foot-shock stress accelerates non-striatal dopamine synthesis without activating tyrosine...
TRANSCRIPT

Naunyn-Schmiedeberg 's Arch. Pharmacol. 308, 1 5 5 - 1 5 7 (1979) Naunyn-Schmiedeberg's
Archives of Pharmacology �9 by Springer-Verlag 1979
Foot-Shock Stress Accelerates Non-Striatal Dopamine Synthesis Without Activating Tyrosine Hydroxylase
A. H. Tissari *, A. Argiolas, F. Fadda **, G. Serra, and G. L. Gessa
Institutes of Pharmacology and Physiology **, Universitfi di Cagliari, Via Porcell 4, 1-09100 Cagliari, Italy
Summary. Electric foot-shock stress (20 min) increases DOPAC content in the frontal cortex (by about 80 %) and in the nucleus accumbens (by 35 %) but not in the striatum. However, foot shock stress failed to modify the kinetic properties of tyrosine hydroxylase (Vmax, K~ for DMPH 4 cofactor) in any of the above areas. Similar results were obtained in rats in which noradrenergic terminals in the n. accumbens and in the frontal cortex had been eliminated by injection of 6-OH-dopamine into the ascending dorsal noradrenergic bundle. The results support the hypothesis that limbic and cortical DA is involved in emotional states and indicate that DA synthesis may be regulated independently from changes in the kinetics properties of tyrosine hydroxylase.
Key words: Foot-shock stress - DOPAC - Dopamine - Tyrosine hydroxylase.
Introduction
Different kinds of stress increase brain dopamine (DA) synthesis in rats and mice (Thierry et al., 1968; Bliss and Ailion, 1971). Recently, Thierry et al. (1976) have shown that foot shock stress selectively augments DA synthesis in the mesolimbic and, more effectively, in the mesocortical dopaminergic system, but that is in- effective in other brain areas. In different experimental conditions increase in DA synthesis is associated with, and is considered to be dependent on, changes in the kinetic properties of tyrosine hydroxylase (TH). Thus, neuroleptic administration or electrical stimulation of the nigro-striatal dopaminergic pathways results both
Send offprint requests to G. L. Gessa at the above address
* Permanent address: Institute of Pharmacology, University of Helsinki, Finland
in increased DA synthesis, and in an activation of TH, characterized by an increased affinity for its cofactor (Zivkovic et al., 1974). However, we have recently shown that neuroleptics still increase DA synthesis but lose their ability to activate TH after destruction of postsynaptic receptors with kainic acid, suggesting the existence of two mechanisms regulating DA synthesis in vivo: one operating by means of postsynaptic DA- receptors and regulating DA synthesis through the activation of TH, the other acting via presynaptic DA- receptors controlling DA synthesis independently from TH activation (Di Chiara et al., 1978). In order to clarify whether the two mechanisms might evidentiated under non-pharmacological conditions, we compared the effect of foot-shock stress on TH activity and on 3,4-dihydroxyphenylacetic acid (DOPAC) level, the primary DA metabolite. When DA levels are un- changed, changes in DOPAC concentrations often reflect parallel changes in the synthesis of DA (Roth et al., 1976; Roffler-Tarlov et al., 1971).
The present results show that electrical foot-shock selectively increases DOPAC levels in the frontal cortex and in the nucleus accumbens but fails to activate TH in these areas.
Material and Methods
Male Sprague Dawley rats, weighing 180 - 210 g were housed at 24 ~ C with lights on from 6.00 to 18.00 and had water and standard laboratory food ad libitum. Stress consisted of a series of electrical foot shocks delivered in individual cages, with floors made of brass rods. Shocks were provided by a st imulator which delivered a shock of 2 m A every 320ms with 160ms duration for a total period of 20nain. Animals were killed immediately after foot shock. Brains were rapidly removed and frontal cortex, nucleus accumbens and str iatum were dissected on ice as described by Westerink and Kor f (1976). TH activity was measured according to the method of Waymire et al. (1971) modified by Zivkovic et al. (1974). Norepinephrine (NE), DA and DOPAC were assayed in an aliquot of the same homogenate used for TH assay, according to a slight modification of the method of Argiolas and Fadda (1978).
0028-1298/79/0308/0155/$01.00
156 Naunyn-Schmiedeberg's Arch. Pharmacol. 308 (1979)
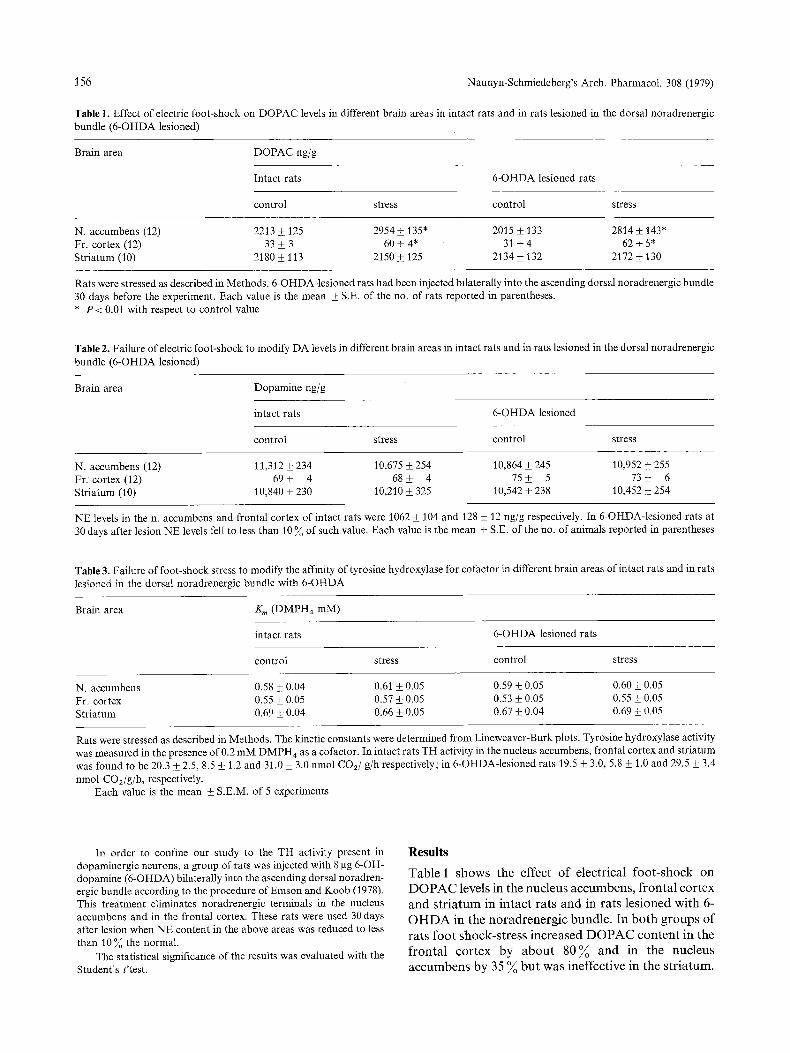
Table 1. Effect of electric foot-shock on DOPAC levels in different brain areas in intact rats and in rats lesioned in the dorsal noradrenergic bundle (6-OHDA lesioned)
Brain area DOPAC ng/g
Intact rats 6-OHDA lesioned rats
control stress control stress
N. accumbens (12) 2213 • 125 2954 • 135" 2015 _+ 133 2814 • 143" Fr. cortex (12) 33 • 3 60 + 4* 31 +_ 4 62 • 5* Striatum (10) 2180 • 113 2150 • 125 2134 • 132 2172 • 130
Rats were stressed as described in Methods. 6-OHDA-lesioned rats had been injected bilaterally into the ascending dorsal noradrenergic bundle 30 days before the experiment. Each value is the mean • S.E. of the no. of rats reported in parentheses. * P < 0.01 with respect to control value
Table 2. Failure of electric foot-shock to modify DA levels in different brain areas in intact rats and in rats lesioned in the dorsal noradrenergic bundle (6-OHDA lesioned)
Brain area Dopamine ng/g
intact rats 6-OHDA lesioned
control stress control stress
N. accumbens (12) 11,312 _+ 234 10,675 + 254 10,864 • 245 10,952 • 255 Fr. cortex(12) 69• 4 68_+ 4 75_+ 5 73_+ 6 Striatum (10) 10,840 _+ 230 10,210 • 325 10,542 _+ 238 10,452 • 254
NE levels in the n. accumbens and frontal cortex of intact rats were 1062 • 104 and 128 _+ 12 ng/g respectively. In 6-OHDA-lesioned rats at 30 days after lesion NE levels fell to less than 10 ~o of such value. Each value is the mean • S.E. of the no. of animals reported in parentheses
Table 3. Failure of foot-shock stress to modify the affinity of tyrosine hydroxylase for cofactor in different brain areas of intact rats and in rats lesioned in the dorsal noradrenergic bundle with 6-OHDA
Brain area K m (DMPH 4 mM)
intact rats 6-OHDA lesioned rats
control stress control stress
N. accumbens 0.58 • 0.04 0.61 • 0.05 0.59 • 0.05 0.60 +_ 0.05 Fr. cortex 0.55 • 0.05 0.57 _+ 0.05 0.53 • 0.05 0.55 • 0.05 Striatum 0.69 _+ 0.04 0.66 +_ 0.05 0.67 • 0.04 0.69 • 0.05
Rats were stressed as described in Methods. The kinetic constants were determined from Lineweaver-Burk plots. Tyrosine hydroxylase activity was measured in the presence of 0.2 mM DMPH 4 as a cofactor. In intact rats TH activity in the nucleus accumbens, frontal cortex and striatum was found to be 20.3 • 2.5, 8.5 _+ 1.2 and 31.0 • 3.0 nmol CO2/g/h respectively; in 6-OHDA-lesioned rats 19.5 _+ 3.0, 5.8 + 1.0 and 29.5 _+ 3.4 nmol CO2/g/h, respectively.
Each value is the mean • S.E.M. of 5 experiments
In order to confine our study to the TH activity present in dopaminergic neurons, a group of rats was injected with 8 gg 6-OH- dopamine (6-OHDA) bilaterally into the ascending dorsal noradren- ergic bundle according to the procedure of Emson and Koob (1978). This treatment eliminates noradrenergic terminals in the nucleus accumbens and in the frontal cortex. These rats were used 30 days after lesion when NE content in the above areas was reduced to less than 10 ~ the normal.
The statistical significance of the results was evaluated with the Student's t'test.
Results
T a b l e 1 s h o w s the e f fec t o f e lec t r ica l f o o t - s h o c k on
D O P A C levels in the n u c l eu s a c c u m b e n s , f r o n t a l c o r t e x
a n d s t r i a t u m in i n t a c t r a t s a n d in ra t s l e s i o n ed w i t h 6-
O H D A in the n o r a d r e n e r g i c b u n d l e . In b o t h g r o u p s o f
ra t s f o o t s h o c k - s t r e s s i n c r e a s e d D O P A C c o n t e n t in the
f r o n t a l c o r t e x by a b o u t 8 0 % a n d in the n uc l eus
a c c u m b e n s by 35 % b u t was inef fec t ive in t he s t r i a t u m .

A. H. Tissari et al. : Foot Shock Stress and DA Metabolism 157
DA levels were not modified in any of the above areas (Table 2).
Table 3 shows the effect of 20 min foot-shock stress on the kinetic properties of TH in intact rats and in 6- OHDA-lesioned rats. It appears that foot-shock altered neither the Km of TH for DMPH4 cofactor nor the apparent Vmax of the enzyme in any of the brain areas studied. This applies both to intact and to 6-OHDA- lesioned rats. As reported by Emson and Koob (1978) removal of noradrenergic afferents resulted in a modest reduction in the Vm, x of the enzyme in the frontal cortex, but failed to change it in the nucleus accumbens, indicating that TH activity measured in these areas is mainly, or solely, the one to be found in dopaminergic terminals.
Discussion
This study indicates that electrical foot-shock stress increases DOPAC levels in the nucleus accumbens and, to a larger extent, in the frontal cortex. These results are in accordance with those of Thierry et al. (1976), who showed that foot-shock stress selectively increases DA synthesis in the above areas, as measured by the rate of decline of DA after synthesis inhibition; they support the contention that mesocortical and mesolimbic DA systems have a critical role in the regulation of emo- tional states.
Moreover this study also shows that foot-shock stress fails to modify the kinetic properties of TH, such as the K,, for its pteridine cofactor and the apparent Vma x, in the same areas where it stimulates DA syn- thesis. The possibility that changes in the properties of TH in DA neurons might be masked by opposite changes in the enzyme present in noradrenergic ter- minals can be ruled out since similar results were obtained in rats in which noradrenergic terminals had been eliminated. The results indicate that changes in DA synthesis can be dissociated from changes in the kinetic properties of TH. It is possible that the latter take place only when the firing rate of dopaminergic neurons exceeds physiological limits, and that, vice versa, under physiological conditions, overstimulation is prevented by DA released by dendrites or nerve terminals onto "autoreceptors" (Carlsson, 1975).
On the other hand, changes in the K m have been observed after electrical stimulation of the substantia nigra (Roth et al., 1975), as well as after administration ofneuroleptics; in the first condition, autoinhibition by DA cannot operate because the firing rate is imposed and, in the second, because autoreceptors are blocked (Walters and Roth, 1976).
Indeed recent evidence from our laboratory sup- ports the contention that changes in DA synthesis can be differentiated from changes in the kinetic properties
of TH: we showed that after destruction of striatal post-synaptic DA receptors with kainic acid, halope- ridol no longer activates TH but fully maintains its capability to increase DA synthesis (Di Chiara et al., 1978).
In conclusion the present results suggest that the rate of synthesis of DA, at least in mesolimbic and mesocortical DA systems, is regulated, under phy- siological conditions, independently from changes in the kinetic properties of TH.
References
Argiolas, A., Fadda, F.: A radioenzymatic method to measure picogram amounts of dopamine and 3,4-dihydroxyphenylacetic acid (DOPAC) in small samples of brain tissue. Experientia 34, 739-741 (1978)
Bliss, E. L., ANon, J.: Relationship of stress and activity to brain dopamine and homovanillic acid. Life Sci. 10, 1161 - 1169 (1971)
Carlsson, A. : Receptor-mediated control of dopamine metabolism. In: Pre- and postsynaptic receptors (E. UsdJn and W. E. Bunney, Jr., eds.), pp. 49-64 . New York: Marcel Dekker, Inc. 1975
Di Chiara, G., Onali, P. L., Tissari, A. H., Porceddu, M. L., Morelli, M., Gessa, G. L.: Destruction of postsynaptic dopamine re- ceptors prevents neuroleptic-induced activation of striatal ty- rosine hydroxylase but not dopamine synthesis stimulation. Life Sci. 23, 691-696 (1978)
Emson, P. C., Koob, G. F. : The origin and distribution of dopamine containing afferents to the rat frontal cortex. Brain Res. 142, 249-267 (1978)
Roffier-Tarlov, S., Sharman, D. F., Tegerdine, P. :3,4-dihydroxyphe- nylacetic acid and 4-hydroxy-3-methoxyphenylacetic acid in the mouse striatum: a reflection of intra- and extra-neuronal metabolism of dopamine? Br. J. Pharmacol. 42, 342 - 351 (197I)
Roth, R. H., Waiters, J. R., Murrin, L. C., Morgenroth, V. H.: Dopamine neurons: role of impulse flow and presynaptic receptors in the regulation of tyrosine hydroxylase. In: Pre- and postsynaptic receptors (E. Usdin and W. E. Bunney, Jr., eds.), pp. 5 -46 . New York: Marcel Dekker, Inc. 1975
Roth, R. H., Murrin, L. C., Walters, J. R.: Central dopaminergic neurons: effects of alteration in impulse flow on the accumu- lation of dihydroxyphenylacetic acid. Eur. J. Pharmacol. 36, 163-171 (1976)
Thierry, A. M., Javoy, F., Glowinski, J., Kety, S. S. : Effect of stress on the metabolism of norepinephrine, dopamine and serotonine in the central nervous system of the rat. J. Pharmacol. Exp. Ther. 163, 163-171 (1976)
Thierry, A. M., Tassin, J. P., Blanc, G., Glowinski, J.: Selective activation of the mesocortical DA system by stress. Nature 263, 242-244 (1976)
Waymire, V. L., Bjur, R., Weiner, N. : Assay of tyrosine hydroxylase by couple decarboxylation of DOPA formed from lt4C-L- tyrosine. Anal. Biochem. 43, 588-600 (1971)
Walters, J. R., Roth, R. H.: Dopaminergic neurons: An in vivo system for measuring drug interactions with presynaptic re- ceptors. Naunyn-Schmiedeberg's Arch. Pharmacol. 296, 5 - 14 (1976)
Westerink, B. H. C., Korf, J.: Acidic dopamine metabolites in cortical areas of the brain: localization and effects of drugs. Brain Res. 113, 429--434 (1976)
Zivkovic, B., Guidotti, A., Costa, E.: Effects of neuroleptics on striatal tyrosine hydroxylase : changes in affinity for the pteridine cofactor. Mol. Pharmacol. 10, 727-735 (1974)
Received October 2, 1978~Accepted April 9, 1979