for celss of diet spirulina potential biomass y · pdf fileabstract spirulina sp. as a...
TRANSCRIPT

NASA Contractor NCC 2-501
Characterization of Spirulina Biomass Y for CELSS Diet Potential / f l& -<Z 1. <WAh/
/ /t/ - g - C r<
Mahasin G. Tadros Alabama AEM University
Normal, AI 35762
(EASA-CE-185319) CEAEACTEfI2193CY GP 1389-2556 1 EPIBULILA B I G B A S S FOB CELSZ IIEP lECfElDTIAE (Alakaaa A E E Uoio . ) 53 p CSCL C6C
Unclas 63/51 0217270
October, 1988
https://ntrs.nasa.gov/search.jsp?R=19890016190 2018-05-22T04:13:42+00:00Z

NASA Contractor NCC 2-501
Title: Characterization of Spirulino Biomass for CELSS Diet Potential
Pr inci pa I I nvestigator Alabama A&M University Normal, AL 35762
Mahasin G o Tadros, Ph-D-
Technical Monitor Robert Do MacElroy, PhoDo NASA/AMES MOffet Field, CA 94035
Period Covered 11/01/87 - 10/31/88
NASA Cooperative Agreement CELSS Program
October 1988

TABLE OF CONTENTS
Page i ACKNOWLEDGMENT AND PERSONNEL ...........................
ABSTRACT ............................................... ii
INTRODUCTION AND BACKGROUND ............................ 1
Objectives ............................................. 3 Significance ........................................... 3
MATERJALS AND METHODS .................................. 4
Culturing .............................................. 4 Growth Conditions ...................................... 5 Analysis ............................................... 7
EXPERIMENTAL DESIGN .................................... 9
Protocol ............................................... 9 Growth Parameters Characterization ..................... 9 Physiological Characterization of Spirulina
in Batch Cultures ................................... 11
RESULTS AND DISCUSSIONS ................................ 13
Temperature and Light 13 Aeration Rate .......................................... 13 Air Enrichment with Carbon Dioxide 17 pH Effect .............................................. 17
physiological Characterization of Spirulina
.................................. .....................
Nutrient Requirement ................................... 20
28 in Batch Cultures ................................... 37 CONCLUSIONS ............................................ 40 FuT[TRE RESEARCH ........................................
REFERENCES ............................................. 42
APPENDIX ............................................... 44

LIST OF FIGURES Page
Figure 1. Flow Diagram for Experimental Design ............. 10
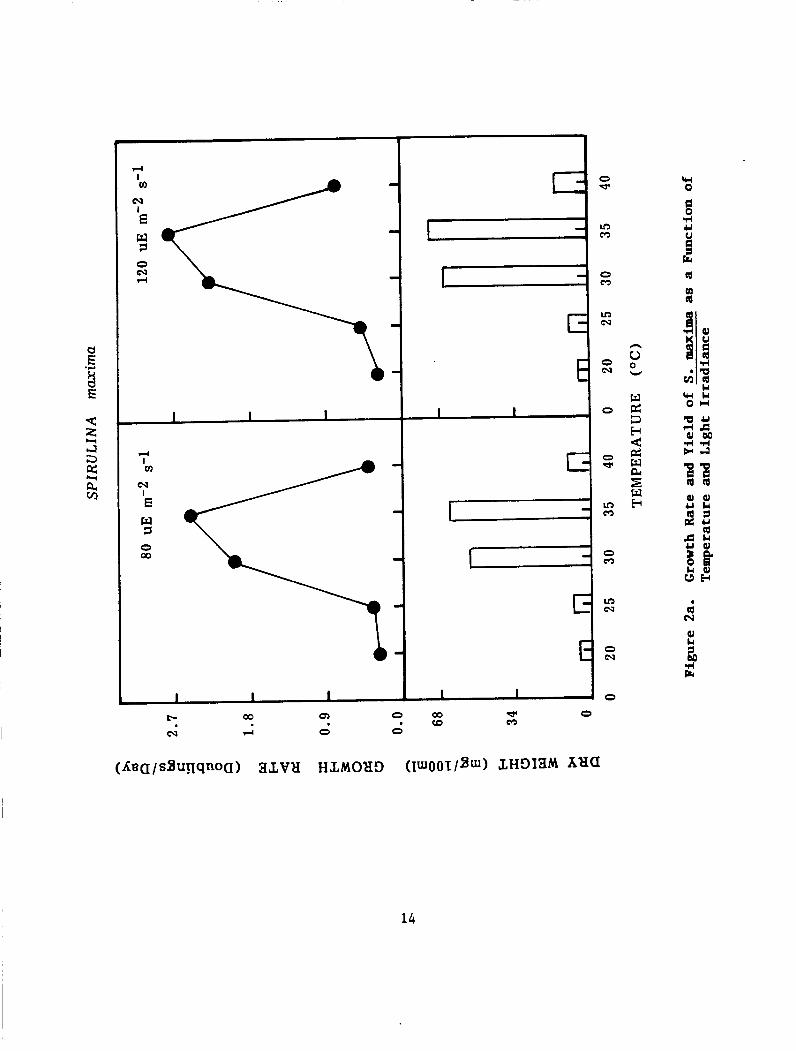
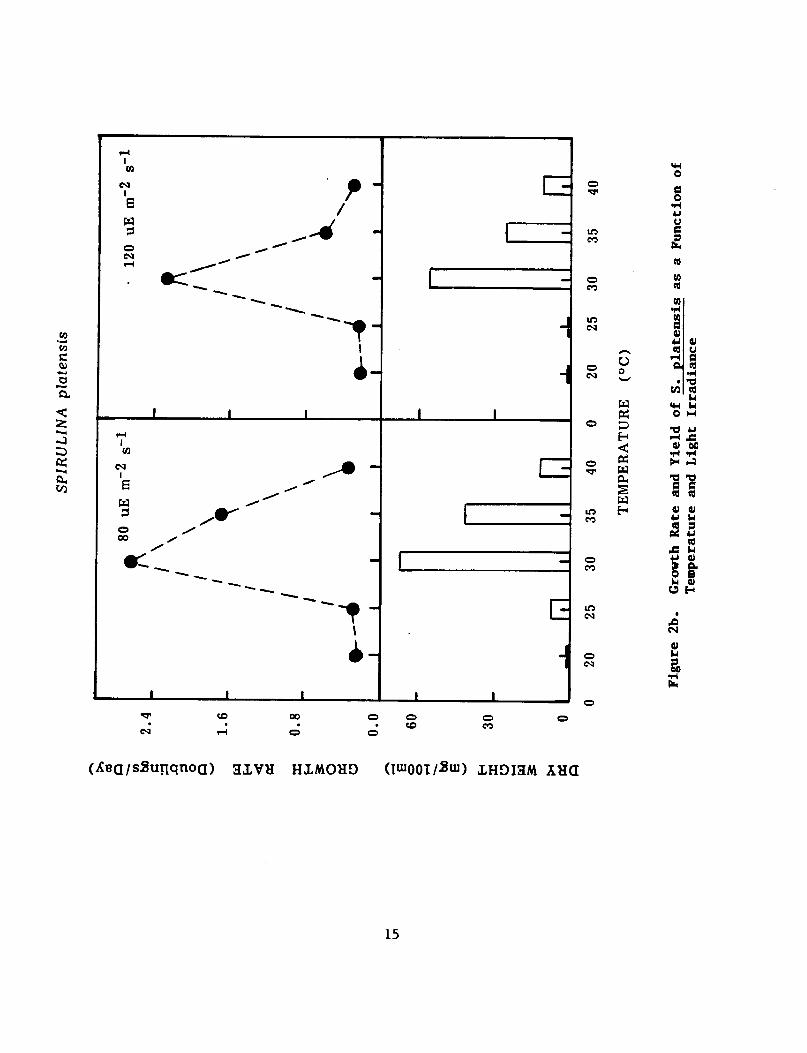
Figure 2. Growth Rate and Yield of S. maxima(a) and S. platensis (b) as a Function of Temperature and Light Irradiance.. ................ 14915
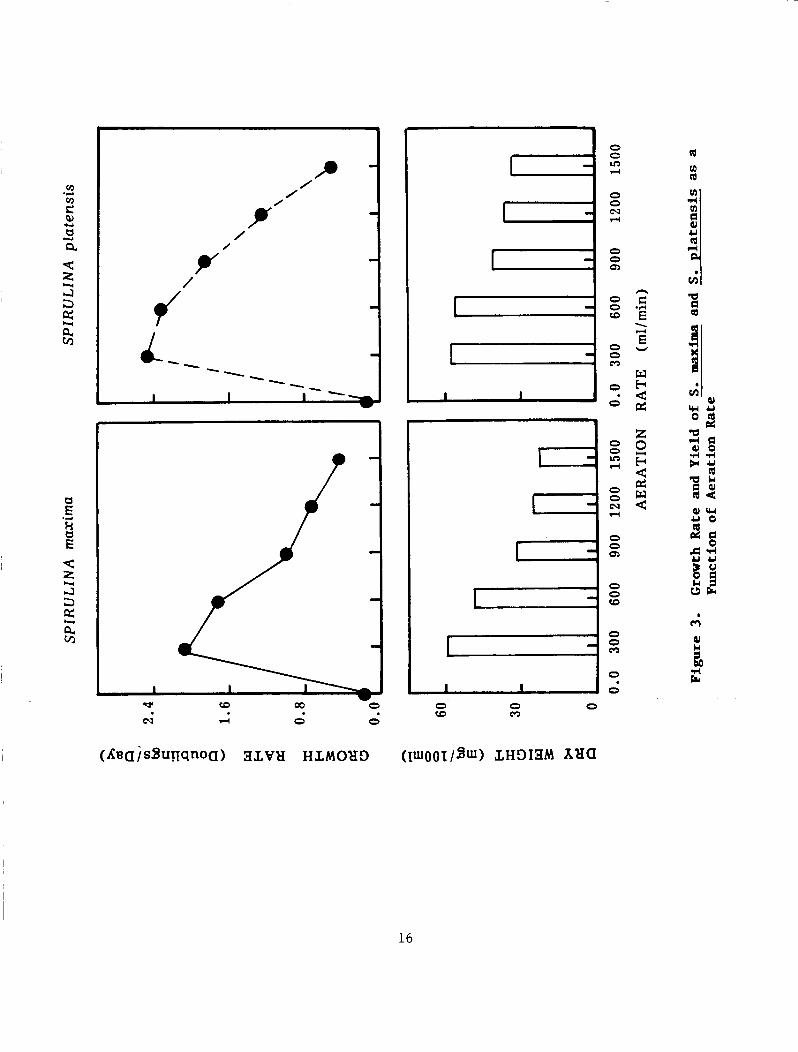
Figure 3. Growth Rate and Yield of S. maxima and S. platensis as a Function of Aeration Rate..................................... 16
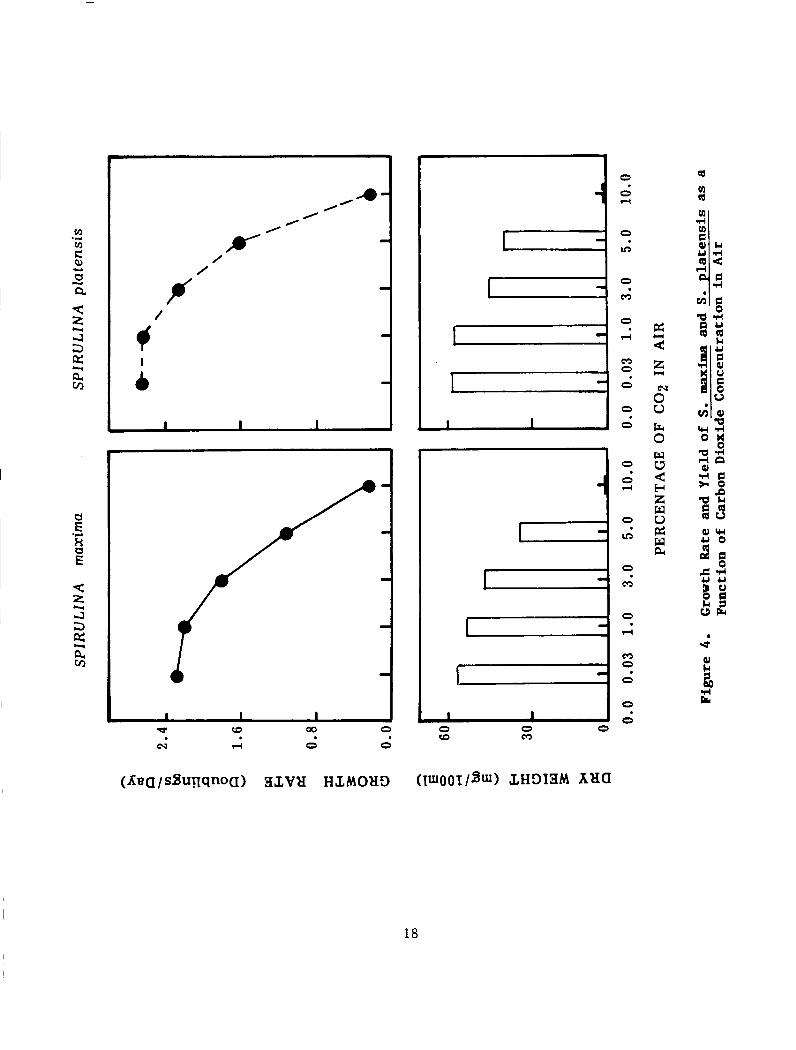
Figure 4. Growth Rate and Yield of S. maxima and S. platensis as a Function of Carbon Dioxide Concentration in Air .............. 18
Figure 5. Growth Rate and Yield of S. maxima and S. platensis as a Function of pH ............................................... 19
Figure 6. Growth Rate and Yield of S. maxima and S. platensis as a Function of
21 Nitrogen Concentration ........................... Figure 7. Growth Rate and Yield of S. maxima
and S. platensis as a Function of Phosphate Concentration .......................... 22
Figure 8. Growth Rate and Yield of S. maxima and S. platensis as a Function of Sodium Chloride Concentration .................... 24
Figure 9. Growth Rate and Yield of S. maxima and S. platensis as a Function of Iron Concentration 25 ...............................
Figure 10. Growth Rate and Yield of S. maxima (a) and S. platensis (b) as a Function of Bicarbonate Concentration ........................ 26,27
Figure 11. Optical Density versus Dry Weight S. maxima (a), S. plantensis (b) ................... 29
Figure 12. Physiological Characteristics, under Optbum Growth Conditions ........................ 31,32
Figure 13. Physiological Characterization of Cultures, under Stress Conditions ................ 35,36
Figure 14. Cells of S. maxima grown under Optimum Conditions (a) and (b) ................... 39

LIST OF TABLES Page
Table 1. Molecular Composition of Spirulina strains ......... 33

ACKNOWLEDGEMENT
Appreciation is extended to N A S A - A M E S for their support of this project.
As the NASA Technical Monitor, Dr . Robert MacEIroy has made invaluable guidance and assistance to the project. him, we are especially grateful.
To
PERSONNEL
The following personnel has been employed on this contract.
Mahasin G . Tadros, Ph.D., P . I . Woodrow Smith, B.Sc., M.Sc. Peter Mabuthi, B.Sc. Beverly Joseph, B . Sc.
i

ABSTRACT
Spirulina sp. as a bioregenerative photosynthetic and an edible alga for space craft crew in a CELSS, was characterized for growth rate and biomass yield in batch cultures, under various environmental conditions. The cell characteristics were identified for two strains of Spirulina: S . maxima and S . platensis. Fast growth rate and high yield of both strains were obtained under the followin conditions: temperature (30°C-350C), light irradiance 60-100 uE m-s s-l, nitrate 30mM, phosphate 2mM, aeration300 ml/min, and pH 9-10. The partitioning of the assimalatory products (proteins, carbohydrates, lipids) were manipulated by varying the environmental growth conditions. Our experiments with Spirulina have demonstrated that under "stress" conditions (i.e. high light nitrogen or phosphate limitation; 0.1 M sodium chloride) carbohydrate increased at the expense of protein. In other experiments, where the growth media were sufficient in nutrients and incubated under optimum growth conditions, the total proteins were increased up to almost 70% of the organic weight.
120 uE m-2 s-l, temperature 38OC,
In other words the nutritional quality of the alga could be manipulated by growth conditions. These results support the feasibility of con- sidering Spirulina as a subsystem in CELSS because of the ease with which its nutrient content can be manipulated.
ii

INTRODUCTION AND BACKGROUND
Certain basic physiological needs must be met in order for human beings to stay alive. On earth, these needs are met by other life forms in conjunction with geochemcial processes that effectively use human waste products in conjunction with energy from the sun to produce fresh supplies of food, oxygen and clean water. In the artificial environment of a spacecraft, these materials must be provided, and human wastes re- moved, without relying on the natural resources of the earth's biosphere.
Pursuit of our national goals in space exploration will eventually require man's long-duration tenancy of celestial vehicles and planetary bases. Requirements for life support could be met through expenditure of stored supplies and by regeneration and reuse of the waste products of human metabolism. The logistics necessary of regeneration for extended space missions are well documented.
The use of biological components Controlled Ecological Life Support System (CELSS) program as subsystems for the revitalization of air, waste processing, and for the production of food has been proposed for the long term - space flight (MacElroy, Bredt, 1985) . Studies of biogenerative life support systems for use in space indicated that they are scientifically feasible. Support of a crew in space, whether in an orbiter or on the surface of a planetary body requires that oxygen, potable water and food be supplied and that waste material be removed. Employment of photosynthetic organisms (higher plants, green algae, cyanobacteria) allows biomass production from relatively simple com- ponents which are readily recycled in a CELSS system, namely carbon dioxide, minerals (NO3-, PO4-3, K+, Na+, etc.) and micronutrients.
The primary source of all man's food and organic raw materials is solar energy. Conventional food sources consist of higher plants and animals. Unconventional food sources for human consumption are photosynthetic algae and bacteria and non-photosynthetic bacteria, yeasts and fungi. Conventional food sources are highly palatable, but require a long time to produce. The photosynthetic energy efficiency of higher plants is less than 3%. Algae, on the other hand, grow rapidly; their metabolism can be controlled; they produce a high ratio of edible to nonedible biomass; and their gas-exchange characteristics are compatible with human requirements.
The semi-microscopic blue-green algae (Cyanophyta; Cyanobacteria) occupy a unique taxonomic position, since they combine an autorophic mode of growth that is common to eukaryotic plant cells with a metabolic system that is generally regarded as bacterial, rather than plant-like.
1

Cyanobacteria single cell protein (SCP) has been used as a food source in various parts of the world (e.g. Mexico, China and Africa) since ancient times; in fact, dried cyanobacteria and cyanobacterial tablets . are now sold in health food stores in Japan, North America and Europe because they are recognized for their nutritional value. The nutri- tional quality of all cyanobacteria which have been tested (See Appendix) appears to be very high. For example, Spirulina, in addition to being the richest known source of vitamin Biz, also contains significant amounts of vitamins B1 and B2. Similarly, one gram of Spirulina contains one-half of the adult daily requirements of vitamin A (B-carotene). The trace elements and iodine found in cyanobacteria are also important when considering the nutritional quality cyanobacteria. The protein of S. maxima and Anabaena cylindrica is easily digestible and approximately 65% of the protein is assimilatible.
Changes in the supply of consumption of metabolites may have con- siderable effects on metabolic patterns. The accumulation of photo- synthetic products in algae can be induced by manipulating the environmental conditions under which the algae are grown (Fogg, 1956). Physiological changes have been indicative of particular changes in nutrient deficiency (Healey, 1975). Studies have shown that limitation of nitrogen, phosphorus, and iron in culture media, affects the growth and physiology of cyanobacteria. Agitationof the culture with air leads to biomass increase. Enrichment with 5% C02 in the bubbling air was an efficient way of obtaining a good productivity (DE la Noue et. al., 1984) . Packer, et. al., (1986), have shown that with proper manipulation of the osmotic environment, macromolecules of carbohydrates can be produced by N2-fixing cyanobacteria.
The most difficult problem in using algae as food is the conversion of algal biomass into products that a space crew could actually eat over a long period of time. If algae are to be considered as a pri- mary food source, it will be necessary to determine that they can be converted into a wide enough range of a palatable complete diet. Therefore, Spirulina, an edible alga with less nucleic acids and no cell walls, offers a gaod prospect for further studies by manipulating growth parameters.
In order to evaluate the potential of Spirulina for a CELSS diet, it is essential to have background information on the environmental tolerances of the species and eventually the responses of physiological characteristics. species in batch and continuous cultures.
This background will be obtained from studying the
The purpose of this project was to evaluate the growth and chemcial composition of two strains of Spirulina under different growth conditions.
2

OBJECTIVES
To characterize batch cultures of Spirulina: Growth, Biomass Yield, and Chemical Compositon under varying:
. Temperature . Light intensity . Aeration rate . Nutrient concentrations SIGNIFICANCE:
. Development of CELSS relies, in part, on the ability to manipulate and control the organisms which are a part of the system. . Biological regeneration of supplies consumed in CELSS. . Direct utilization of algae in space craft crew diet.
This project started in November 1987. The accomplishments in the period November 1987 to October 1988 are described in this report.
3

MATERIALS AND HETEODS
Culturing :
Organism: Two strains of cyanobacteria Spirulina:
. S. Maxima: (UTEXLB 2342) was obtained from Utex Algal Collection
. S. Platensis: was obtained from Dr. Becker W. Germany
. S. Maxima filaments have turns, while S. Plantesis filaments are straight ones.
Growth Medium: Zarrouk medium (1966) was used as follows:
NaHC03 16.0g; K2HP04 0.5g; NaN03 2.5g; K2SO4 1.Og; NaCl 1.Og;
MgS04.7H20 0-2g; CaC12 0.4g; FeS04 0.01g; EDTA 0.08g; Solution
A5 1mL; Solution B6 1mL; in 1L distilled water.
Solution A5 in grams per litre of distilled water: H3BO3, 2.86; MnC12.4H20, 1.81; ZnS04.7H20, 0.222; CuS04.5H20, 0.079; and MoO3, 0.015.
Solution B6, in milligrams per litre of distilled water: NH4V03,
22*96; KCr (s04) 2.12H20, 192.0; NiS04.6H20, 44.8; Na2W04.2H20, 17.94; TiOSOq.H2S04,8H20, 61.1; and Co(N03)2.6H20, 43.98.
The medium was autoclaved without the bicarbonate salts. The bicarbonate solutions were sterilized by filtration through 0.2 mm pore size filters.
The culture medium was modified for nutrient limitation studies:
. For N2 limited cultures, NaN03 was replaced by KC1, and nitrate ammonia, and urea were tested in different concentrations as nitrogen sources.
. For P-limited medium the P was replaced by NaCl and H3P04 was used as P-source, in different concentrations.
. For salinity studies, NaCl was used in different concentrations.
4

. FeSO4 was used in different concentrations.
. pH was maintained in all cases at 9 with 2N NaOH Contamination: The standard plate-count method was used to determine the number of bacteria present in the culture. Aliquots were plated using a bent glass rod on an agar medium, which is prepared from Zarrouk (1966) medium (algae medium mentioned previously) enriched with the following ingredients:
Tryptone glucose yeast agar: Tryptone, 5.0g; Yeast extract, 2.5g; Glucose, 1.Og; Agar, 15.0g; in 1L distilled water, pH 7 .
0.1 ml filtrate was spread on the agar surface and incubated. Colonies were counted, dilution was made if necessary. Plates were incubated at 30°C and counted after 48 hours or longer to detect all organisms.
Purification of Spirulina Culture: The original cultures of Spirulina were contaminated with bacteria. Different procedures were used to purify the cultures. However, the following procedure was the most successful one:
Cells were collected, filtered with 8mm filter (Gelman), washed with basal medium and homogenized. Cells were spread in a plate, exposed to UV 5 min (20 W UV lamp, 30 cm distance) and inoculated in test tube cultures containing the basal medium. One drop size, inoculum per tube. From 100 culture tubes, 10 tubes were bacteria free.
Growth Conditions:
Culture Room: A small room (3m D x 2.45m W2 x 2.lm H), was available for this project. It was provided with shelves, which have been illuminated with cool white fluorescent tubes. Light intensity varied from 80-100 uE m-2 was kept at 25°C.
on the shelves. The room temperature
Light Measurements: Light irradiation measurements were made with a Li-Cor Model Li-185 M (Lambda Instruments) Meter equipped with a spherical quantum sensor.
All experiments were incubated in continuous light.
Culturing Techniques:
a. Culturing bottles: Small bottles (60 ml capacity) containing 30 ml growth medium were inoculated from stock culture in the
5

exponential phase and bubbled with air. These cultures were used for evaluating the growth parameters of the alga.
b. Roux bottles: Experimental amounts of algal cells were grown in roux bottles, containing 800 ml sterile growth medium, by in- oculating them with 50 ml of preadapted rapidly growing culture in a 125 ml erlenmeyer flask. Cultures were illuminated con- tinuously by placing them in front of a bank of two cool white fluorescent lamps (40 W). Light irradiation, measured at the surface of culture bottles was 80 uE m-2 s-l. grown in a water bath kept at 29-30°C by the use of a heater- thermostat combination.
The cultures were
c. Aeration: The cultures were aerated with air (0.03% C02) or air enriched with carbon dioxide. The air was delivered by an oil- less compressor. The air was passed first through concentrated sulfuric acid, a saturated ZnC12 solution and then distilled water. The air was then passed through a cotton filled erlenmeyer followed by a glass wool filled erlenmeyer prefilter. Finally, the air was sterilized by flowing through sterile filter 0.22 um (Gelman). The air flow rate was monitored by a flow meter. The source of carbon dioxide was from a pressurized tank (50 lb) which was provided with a regulator and solenoid valve to shut off the gas automatically through an electric timer. The carbon dioxide flow was monitored by a flow meter and was sterilized by passing through sterile 0.22 um filter (Gelman). Mixtures of air (0.36% C02) and carbon dioxide were obtained by blending gases to a desired mixture in a two-gas proportioner. The flow rate of the mixed gas delivered to the culture was maintained at 300 ml/min.
6

Analysis :
Growth Rate: Growth was measured by monitoring change in absorbance (O.D.) at 560 nm with spectrophotometer (Perkin Elmer Lambda I) and expressed as doublings day'l. The mean daily division rate t, K , is calculated from:
Where, t = days since inoculation
ODt = optical density after t days
OD, = optical density when t = 0.
Harvesting of Cells: Cells were collected by filtration using filter paper 10 mm pore size (Gelman). Cells were washed with buffer solution (pH 8), diluted to known volume and processed for further analysis. Cultures were harvested at O.D. 0.1 units, to avoid light limitation.
Triplicate samples of the algal suspension were taken for each determination. The mean value of these triplicates was recorded. The follwoing determinations were carried out:
Total Chlorophyll: An aliquot from the culture was centrifuged for 2 min at 2000g. The precipitate was suspended in 5 ml methanol for 5 min in a water bath at 7OoC, and thereafter centrifuged. optical density of the supernatant was determined 655 nm.
The
Dry Weight Measurements (DW): A volume from the culture was filtered through a filter 10 um pore size, dried in previously dried, pre- weighed filter paper for 4 h at 80°C, and then weighed after cooling in a desiccator.
Ash-Free Dry Weight (AFDW): After recording the dryweight, the dried cells were ashed at 500°C for 2hrs. Then the ash wt. was recorded. The difference between dry weight and ash weight gave the organic weight of the sample.
Total Carbohydrates: The anthrone sulphuric acid method was followed (Strickland, Pearsons, 1972). The principle of this method is the formation of a blue-green color which is the product of anthrone and the furfural derivatives produced by acid decomposition of the sugar. The anthrone reagent consists of 0.2g anthrone, 8 ml ethyl alcohol, 10 ml distilled water. one ml of algal suspension (containing known weight of alga) and heated in a water bath for seven minutes and cooled.
Then ml of the anthrone reagent was added to
7

The blue-green color was measured by a spectrophotometer at wave length of 620 nm. The value of the reading was calculated as micrograms of glucose from a standard curve for glucose which has been prepared by the same method.
Total lipids: Cellular lipids were solubilized by repeated extraction with methanol and methanol-chloroform ( l : l ) , then phase separated after adjustment of the solvent ratios to 10:10:9 (methanol: chloro- form: water, v/v) (Bligh and Dyer 1959). The chloroform phase was collected, evaporated to dryness under N2, and the weight of the lipid was determined. Lipid content was calculated as the weight of the lipid extract divided by the ash free dry weight of the original sample.
Total Nitrogen and Protein (Kjeldahl): One ml of algal suspension containing a given weight was digested in a Kjeldahl digestion flask containing 0.3, selenium mixture and one ml sulfuric acid. contents became colorless, they were transferred to the Kjeldahl apparatus with 10 ml of 50% sodium hydroxide solution. A strong current of steam was passed for 7 minutes during which the liberated ammonia was received in a 100 ml flask containing 5 ml of 2 % boric acid solution and 4 drops of indicator. The indicator was composed of 0.016g methyl red and 0.83g bromocresol green in 100 ml of alcohol. The distillate in the boric acid solution was back titrated with 0.1 N sulphuric acid, until the color of the indicator turned pale pink. A blank sample was done for each series of nitrogen estimation, using D.W. The value of the readings was calculated in ug N, from a standard curve for nitrogen source as ammonium sulfate, which has been treated by the same method. total N x 6.25.
When the
Total protein was calculated from
8

EXPERIMENTAL DESIGN
Protocol: Flow diagram for the experimental design was followed (Fig. 1).
A. Grovth parameters characterization:
I. Temperature, Light:
The algal growth was evaluated for temperature and light tolerance on a gradient plate. Temperature could be adjusted in range from 10°C to 50°C. Illumination was provided by eight cool white fluorescent tubes (40 W). Different light intensities were obtained by varying the distance between the cultures and light source. The algal species were cultured in small bottles (60 ml capacity) containing 30 ml growth medium. Triplicate cultures were placed on the gradient plate, at temperatures: 20°C, 25"C, 35OC and 40°C. The cultures were exposed to two light intensities and were aerated with air (0.03% C02).
11. pH Effect:
The alga was incubated in small bottles as described in section A(1) at 35°C on a temperature gradient plate and 80 uE m-l s - ~ irradiance. culturing, except the pH used for culturing was varied by using NaOH or HC1. The pH of cultures was adjusted daily to the original pH. The cultures were aerated with air (0.03% C02).
The original medium was used for
111. Aeration Rate, Carbon Dioxide Enrichment, Bicarbonate Con- centration:
The alga was incubated in small bottles described in Section A(1) at 35°C on a temperature gradient plate and 80 uE m-l s - ~ irradiance. Three sets of cultures were treated differently:
a. Cultures were aerated with different flow rates (air 0.03% C02).
b. The flow rate which gave the best growth rate, was selected from 'la''. The cultures were aerated with air enriched with carbon dioxide in different concentrations 1%, 32, 52, 10%.
c. Cultures were treated with different bicarbonate
9

w
rl (d u 4 bl
B
a rl
10

concentrations in which one set was aerated with air (0 .03% C 0 2 ) and other set was aerated with air containing 1% C02.
The pH of all cultures was adjusted twice daily.
IV. Nutrient Requirements:
Cultures were incubated in small bottles under the same conditions as described in Section A ( 1 ) . The original growth medium was modified by changing the concentration of one nutrient. Nitrogen, phosphorus, iron, bicarbonate and sodium chloride were studied in sufficient and limiting concentrations. The bicarbonate effect was studied together with the aeration effect "111".
In all experiments triplicate culture bottles were inoculated from stock cultures in the exponential phase. Growth response was measured as optical density and the growth rate was expressed as doublings per day. The yield of cultures was expressed as the total dry weight after 5 days of growth. The total day weight was determined by harvesting the cells and drying it (see Methods).
B. Physiological Characterization of Spirulina in Batch Cultures:
For this experiment, the alga was grown in batch cultures (Roux bottles) as mentioned in "Methods". The cultures were maintained under optimum growth conditions and monitored in the exponential phase by the absorption measurement (see Methods).
I.
11.
O.D. of Cell Suspension versus D.W. and Chlorophyll: Both species grown intriplicate Roux bottles under the same conditions described before (see Methods). Twenty ml of culture samples were taken daily for measurements of theD.W., and chlorophyll. The experiment was continued for one week, and 20 ml of fresh culture medium were added to the Roux bottles immediately after each sampling in order to maintain the same volume of the culture medium during the cultivation.
Under Optimum Growth Conditions:
Both species were grown in duplicate Roux bottles under the same conditions described before (see Methods). Cultures were analyzed for growth parameters during the eight days.
11

111. Various Stress Conditions:
Light and Temperature:
Batch cultures were incubated under high light irradiation and others at high temperature (38°C) in water bath.
Nutrients:
Both species were grown in Roux bottles under the same conditions described previously. Batch cultures were grown in duplicate until the exponential phase was reached at 0.1 OD, to avoid light limitation. One batch was analyzed and represented the culture sufficient in nutrients. The exponential phase lasted three to five days. Batch cultures were concentrated and diluted to the original batch volume but with a new medium modified in one element. The nitrogen limited batch received 0.05 mM nitrate; the phosphate limited batch received 0.01 mM phosphate; the bicarbonate limited batch received 4g/L bicarbonate; and sodium chloride was added in two concentrations; 0.1 M and 0.5 M. When one element was limited, the others were in sufficient concen- trations. The cultures were incubated under stressed condi- tions for two days and ten harvested for analysis.
12

RESULTS AND DISCUSSION
Temperatue and Light:
Figure 2 (a and b) depict the growth and yield of both strains of Spirulina at two light irradiations and different temperatures ranging from 20°C to 40°C. grow at 25°C at very slow rate. algal fastest growth rate and highest yield of cells. temperature was raised to 40"C, the algal cells turned yellow and
Neither strains grew at 20°C but they started to Temperatures 30 and 35°C enabled the
When the
gave a lower yield. S. platensis, on the other hand gave optimum yield at light irradiation-80 uE m-2 s-l while S. maxima tolerated light irradiance 120 uE m-z s-l.
Aeration Rate:
The effects of air agitation rate on the growth rate and cell yield aredepicted in Figure 3 . The growth rate of both Spirulina strains increased with increasing the flow rate of air in range of 150 ml/min and 500 ml/min. When the flow rate of aeration was increased to 2000 ml/min, the growth rate started to decline and cells turned yellow. On the other hand the cell yield in terms of dry weight was not affected. The pH of all cultures increased to 11. The cell yield of both strains showed parallel fluctuation to the growth rate of the agla.
13
rl I u1
N I E W 3 0 ea d
I I I
rl I rn si
I E w 1 0 00
N
00
rl
Q,
0
0
In m
0 m
Lo N
0 N
0
0 e
Lo e3
0 m
Lo N
0 N
0
0 0 0 e * w m 0
0
W 0
B a
14
d I v1
N I E w 1 0 hl d
I I
\ \
* (D
CJ rl
00
0
t /
4-
---? I
I I I
rl I cn
N I E w s 0 00
0 0
0 w 0 ,I
/ 0
.0@
0 0 * c D 0
0 m 0
0 w
v) m
0 m
v) N
0 CJ
0
0 w
In m
0 c3
In N
0 N
0
15
t‘
/ /
Y’ /-
Y’ /
ea d 0 0
I
0 0 0 W m
16

Air Enrichment with Carbon Dioxide:
Figure 4 shows the effect of air enriched with different concentrations of carbon dioxide on the growth rate and yeild of both Spirulina strains :
. Cultures aerated with 10% C02 - in air, did not grow and turned yellow. The pH dropped rapidly, within 3 hours, from 9.4 to 7 .
. When the C02 - concentration in air was decreased to 5% or 3% the cultures started to grow. The pH of the cultures were maintained at 9.4 by the addition of sodium hydroxide. How- ever, the pH of cultures aerated with 1% C02 - enriched air was maintained stable.
. Cultures aerated with air (0 .3% C02) grew at more or less the same growth rate of those aerated with 1% C02 - enriched air. The yeild of cultures treated with different C02 concentrations, in terms of dry weight, was equivalent to the growth rate.
The results of this experiment are in agreement with those of Faucher and Coupal ( 1 9 7 9 ) . They reported that sparging 1% C02 - air in Spirulina cultures could maintain a constant pH of the culture medium, and at the same time generate HCO3 ions which were used as carbon source for S . maxima. In a similar study with green algae, Goldman and Graham (1981), reported that in batch cultures, maximum growth rates were achieved at the C02 levels present in atmospheric air and at HCO?j concentrations of 3 mM.
pE Effect:
The growth rate of b o t h Spirulina strains is clearly affected by the pH of the growth medium as in shown in Figure 5. Both strains exhibited higher growth rate in media of pH range of 9 to 10. The growth rate decreased with increasing pH above 10 and the cells turned yellow in case of S. maxima while in S . platensis the cells remained in bluish green in color. The cell concentration increased when increasing the pH of the medium from 8 to 10 and then decreased above pH 10 .
1 7
0 0
/ /
I 0
/
W
d
00
0
0
0
4
4 0 0 0 W m
0
0 rl
0
m
0
m
rl
m 0
0
0
d
0
m
0
m
d
m 0
0
18

R
f 2 c-r
f I
\ \
\ \ \
L
v) co m co d
v) b 0
0 0
0 cp
0 e3
0
w 0
19

Nutrient Requirement:
Nitrogen :
Nitrogen sources in the form of nitrate, urea and ammonia were tested in different concentrations in order to determine their effectiveness as N-sources. Ammonia inhibited the growth of both strains and there- fore the data were deleted. The results of nitrate-N and urea-N are represented (Figure 6 ) . The growth rate of both Spirulina strains was inhanced with increasing the concentration of urea-N and nitrate-
- N. The urea-N at 20 mM concentration enhanced the growth rate, while further increase in its concentration limited the growth of both strains. On the other hand, nitrate-N at concentration 30 mM, enabled both strains to reach fast growth rate and high yield in terms of dry weight. the growth of both strains to some extent, the strains bleached and lost their pigments. This experiment demonstrated that the least amount of nitrate-N necessary to maintain the growth of Spirulina in culture was 10 mM. Microscopically, the trichomes became shorter in both strains and with average 6 turns/trichome, in media limited in nitrogen concentration. In agreement with our results, Faucher et. al. (1979) , reported t h a t urea-N in low concentration could support the growth of S. maxima, at high concentration of nitate-N.
Although lower concentration of nitrate-N (10 mM) supported
Phosphate:
Increasing the phoshate-P concentration in the culturing media to 1 mM and 5 mM, enhanced the growth rate of both strains (Figure 7) . But as the concentration increased to 10 mM, the growth rate of both strains declined. The mass yield of both strains showed similar responses coinciding with their growth rate. Microscopically, the trichomes became shorter in media of phosphate-P concentration below 1 mM and with few number of turns in case of S. maxima ( 5 turns/trichome). Generally, cyanobacteria require small concentrations of phosphate-P for growth. They can grow inphosphorus-limited media (Lang and Brown, 1981).
20

w c E I
I \\\ \\\ \\\\ \\
c z 3 n: a v)
3 k
yO< \ \ \
w b
e w a: c p: E I n 5
w N
W
d
00 0
0 0
I 2 3
""1
0 *
0 m
0 el
0 d
0
0 w
0 m
0 hl
0 d
0
rb a rb
n s Q
E 4 W
2 1

2.25
1.5
0.75
(
0 . 0
60
30
0
- - -
- -
2 I I I
S P I R U L I N A m a x i m a
. 0.0 2.0 5.0 10.0
S P I R U L I N A p l a t e n s i s
P, I '
I I
'0 \ \
I I
/ i
\ \ \ b
CONCENTRATION (mM)
PHOSPHATE
Figure 7. Growth Rate and Yield of S. maxima and S. platensis as a Function of Phosphate Con- centration
22

Sodium Chloride:
Both Spirulina strains grew in media lacking sodium chloride (Figure 8). They showed response in growth rate as the sodium chloride concentration increased to 10 mM. Further increase in sodium chloride concentration (100 mM) affected the growth rate of both strains and resulted in lower yield of cells. In addition, microscopic examination of both strains indicated that in media treated with a high concentation of sodium chloride 100 mM, the trichomes were short and with less turns in case of S. maxima (6 turns/trichome). The results of this experi- ment, indicate that Spirulina tolerate increases in sodium chloride concentration up to 100 mM. Spirulina tolerance to salt had been previously reported (Faucher, et. al., 1979).
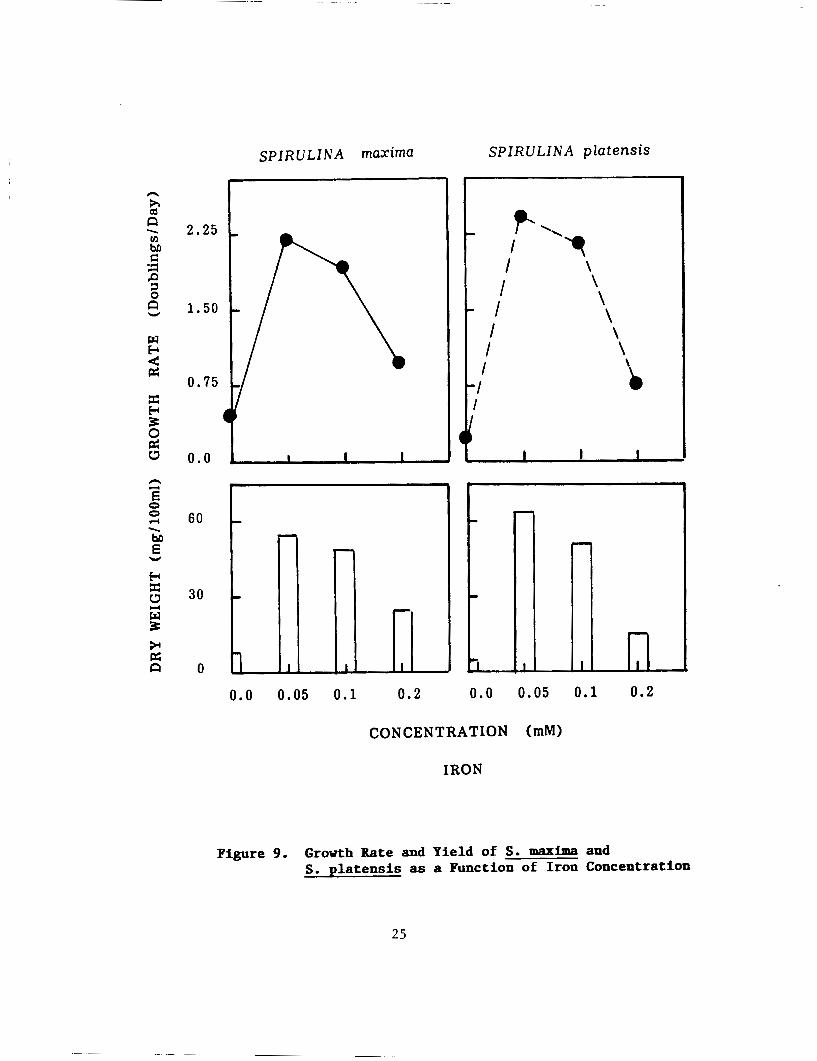
Iron:
Iron concentrations (FeS04) influenced the growth and yield of both strains (Fig. 9). Concentration of 0.05mMwas sufficient for the growth of both strains, although media deficient in iron did not show any growth response. Increasing the concentration of iron beyond 0.1 mM lowered the yield of the alga and cells turned yellow.
Bicarbonate Concentration:
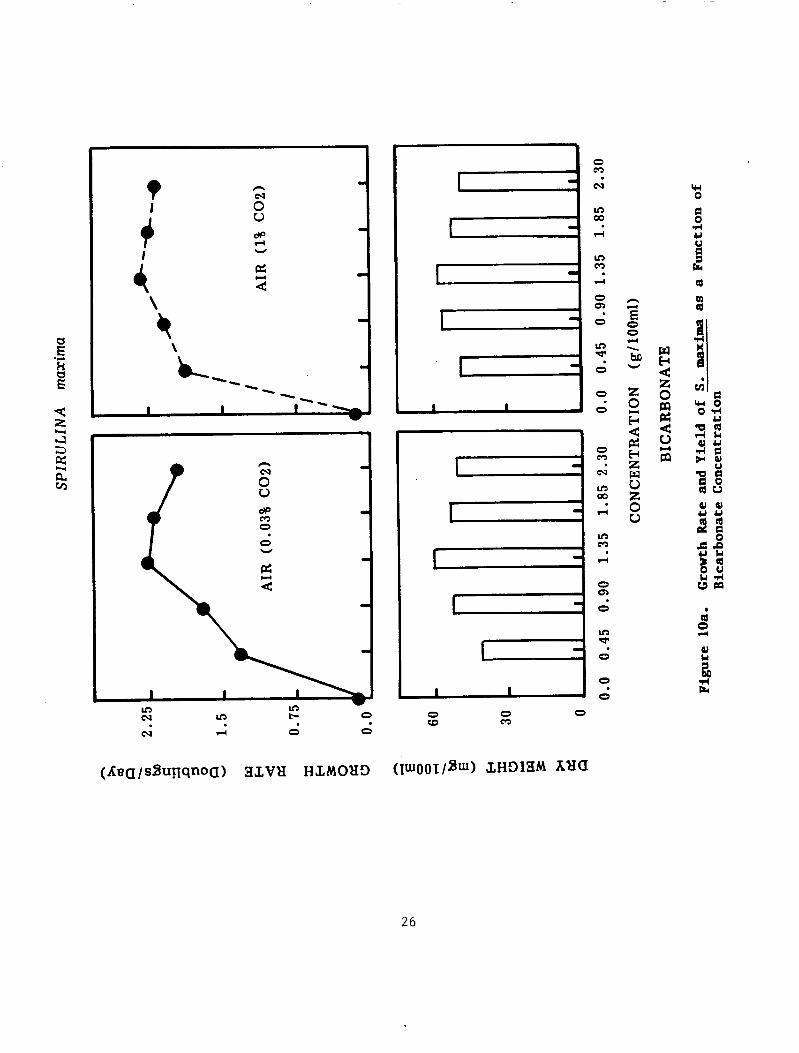
Figure 10 "a" shows that S. maxima grows in the medium even without bicarbonate salt, providing that the culture was aerated with air (0.03% CO2). A s the bicarbonate concentration increased, the growth rate as well as productivity increased. Further increase in bicarbonate concentration above 16g/L (190 mM) did not affect the growth rate. When the carbon dioxide concentration in the air increased from 0.03% to 1%, as shown in Figure 10 "b", the growth rate increased remarkably by decreasing the bicarbonate concentration in the medium as low as 4g/L (48 mM) i.e. one quarter of the concentration in the Zarrouk medium (see Methods). Spirulina platensis, does not show much variation in its response to increasing CO2 concentration in air, when compared to S. maxima (Figure 10 "b"). The cell concentration based on dry/weight measurement was related to the growth rate in both strains, in all treatments. The results of this experiment indicate that both strains can utilize atmospheric carbon dioxide when the media bicarbonate concentration is minimum in the culture medium. was adjusted daily to 9.4.
The pH of all cultures
23

SPIRULINA maxima SPIRU LINA platensis
0 .0 0.01 0.1 0 . 5
I I I
0 . 0 0.01 0.1 0.5
CONCENTRATION ( M )
SODIUM CHLORIDE
Figure 8. Growth Rate and Yield of S. m a x i m a and S. platensis as a Function of Sodium Chloride Concentration
2 4
n
i2 n Q s 70 n
\
c .H
U
w t.c 4 p:
3: w 5 0 p: c3
n
0 0 4
4
E
\
M E
w X c3 w 5 * p:
U
Y
n
2 . 2 5
1.50
0.75
0.0
60
30
0
S P I R U L I N A maxima S P I R U L I N A platensis -
- 7 \
I ?-- \\ \
I I \
- I \ I \ I \
i I -I
1
- -
1 I I I
1 I I I
L 0.0 0.05 0.1 0.2 0.0 0.05 0.1 0.2
CONCENTRATION (mM)
IRON
Figure 9. Growth Rate and Yield of S. maxima and S. platensis as a Function of Iron Concentration
25
?
i i \
\ t
n 0 ea
@a d 0 0
- 0 0 0 CD m
0 m m
In 00
d
In m d
0 aa 0
m e 0
0
0
0 rn ea m 00
d
In m r(
0 Q,
0
In * 0
0
0
26

T
p: 2
i 4
\
t 1
I 1
I t I
i
c3
- 4
0 Q,
- 0
m * 0 0
0
' 0
n
0" u do m 0
0
I
00 00 0
0 m
- m
d 0 0
0 Q,
0
m * a: 0
1 1 4: 0 0 0 W m
n - E 0 0 d \
M
z 0 t-c c p: t-c z W u z 0 u
W
I
27

Physiological Characterization of Spirulina in Batch Cultures:
Batch Cultures:
I. op tical Density (O.D.) of Cell Suspension versus Dry Weight (D.W.) and Chlorophyll:
Growth can be expressed as growth rate or as yield. Yield, as an expression of organic production, is usually given in terms of dry weight of the organic mass produced over a period of time per unit volume. A relationship between optical density, dry weight and chlorophyll was established for both strains of Spirulina.
Results are presented in Figure 11. For all samples within the first three days of cultivation, which contain relatively small concentrations of biomass (400 mg DW/L or less), readings fell within the accurate range of the O.D. scale and they could be read directly from the spectrophotometer without dilution. How- ever, for all samples during the later cultivation periods which contained high concentration of biomass (500 mg DW/L), dilution of the samples with distilled water was necessary prior to OD readings. The graphs show linearity between OD and dry weight. Each OD unit is equivalent to a concentration of 700 mg/L in the case of S. maxima and to 750 mg/L in the case of S. platensis. It is obvious from this experiment that other reliable indicators of estimating algal productivity can be computed from OD measure- ments. Therefore, OD measurements can be translated into biomass yield in terms of dry weight or chlorophyll.
28

1.5
1.0
E c 0 (0 v)
c, a
0.5
0.0 0 250 400 550 700 850
n
0
1.
1.
10
n d + bn E W
0.
0.
DRY WEIGHT (mg/l)
Figure 11. Optical Density versus D r y Weight S. maxima (a) and S. platensis (b)
29
0 p: 0 bJ 3: u
10

11. Physiological Characteristics of Culture, under Optimum Growth Conditions:
Both species were grown in duplicate Roux bottles under the same conditions described before (see Methods). Cultures were assayed for growth parameters during the eight days. (Fig. 12 "a and b"). Increments of carbohydrates, proteins, dry weight and chlorophyll are expressed as ug/ml culture. The results show that increases in the synthesis of chlorophyll, protein and yield of the culture are correlated. Growth parameters of cultures analyzed after 8 days started to level off, due the nutrient exhaustion and light limitation caused by increasing cell concentration.
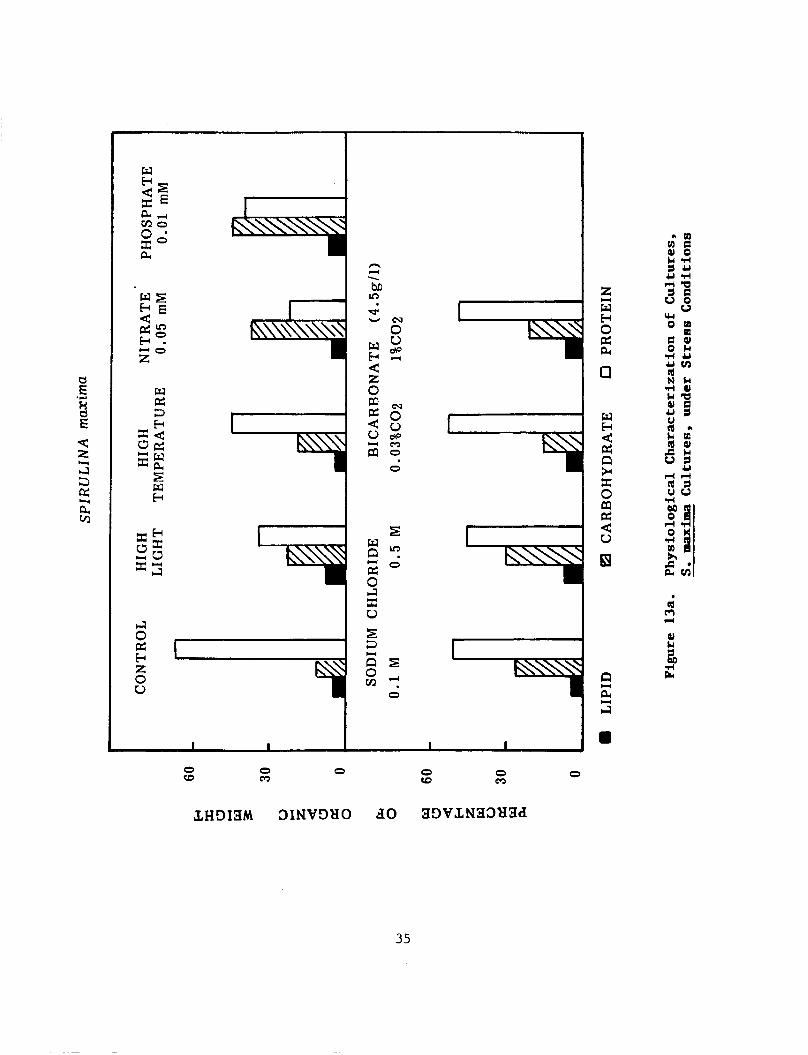
111. Physiological Characterization of Cultures, under Stress Conditions:
The results of analysis were expressed on the basis of organic weight (Ash Free Dry Weight: AFDW) and represented in Table 1 (Figure 13). Results of cultures grown under optimum condi- tions (11) were used as control for all experiments incubated under various growth conditions.
Light Irradiance and Temperature: Increasing the light irradiance to 120 uE m-2 s-l, led to an increase in the total carbohydrate content and a decrease in total protein content: S. maxima 19.58%,29.06%: S. plantensis 15.222, 27.18%. In- creasing the temperature of culture incubation to 38"C, in- fluenced the composition of both strains, in a similar manner to the light irradiation experiment: S. maxima, 18.75%, 45.28%; S. platensis, 13.12%, 35.32%; for protein and carbohydrates, respectively. Both strains produced a low percentage of lipids, when grown in high temperature experiments. Cells turned yellow green in color. Studies with light-limited cyanobacteria showed a high level of polysaccharide formation when they were exposed to high light intensities (Konopka, et. al., 1987). Cohen et. al., (1987) also reported a reduction in fat content of 19 strains of Spirulina by temperature and high light intensity.
Nutrient Limitation: Media limited in nitrate-N and phosphate-P, favored the accumulation of carbohydrate rather than protein. In nitrate and phosphate limited cultures: S. maxima had 37.52%, 35.21% carbohydrate and 21.562, 41.25% protein, while S. platensis had 30.31%, 31.87% carbohydrate, and 32.81%, 34.683 protein. When the cultures were transferred to media limited in nitrogen and phosphate, cultures changed in color from blue to yellow-green. N-limited cultures of Anacystis nidulans (Lehman and Wober, 1978), Anabaena variabilis (Ernst and Boger, 1985) and P-limited cultures
30

SPlRULINA maxima
I B C A R B O H Y D R A T E PROTEIN
8oo t z w c-c c(
0 6 0 0 1 & .I
w 2 c-c- 2 400
* X 0 200 eo p: 4 u
nu
0 1400 c
D R Y WEIGHT M CHLOROPHYLL I n
d 1
bn 1050 E v
* CG n
f 700
350
0
0 48 96 144 192
TIME (hrs)
Figure 12a. Physiological Characteristics of S. maxima under Optimum Growth Conditions
31

800
600
400
200
0 1600
1200
800
400
0
SPIRULINA platensis
C A R B O H Y D R A T E B PROTEIN
n L D R Y WEIGHT
A-A CHLOROPHYLL
0 48 96 144 192
TIME (hrs)
Figure 12b. Physiological Characteristics of S. platensis under Optimum Growth Conditions
15
10
5
0
32
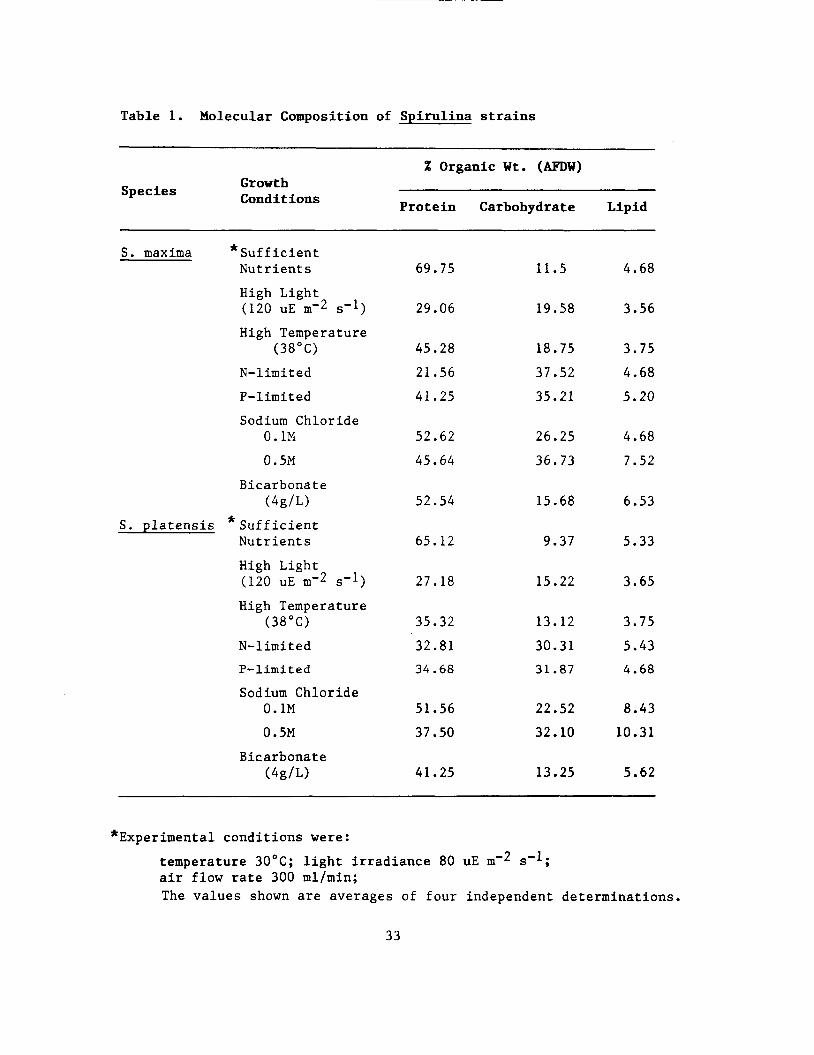
Table 1. Molecular Composition of Spirulina strains
X Organic Ut. (AFDW) Growth Conditions Species
Protein Carbohydrate Lipid
S. maxima *Sufficient Nutrients 69.75 11.5 4.68
High Light (120 UE m-2 S-1)
High Temperature
N-limi t ed
P- I imi t ed Sodium Chloride
(38°C)
0.1M
0.5M
Bicarbonate ( 4 g / L )
29.06 19.58 3.56
45.28
21.56
41.25
18.75
37.52
35.21
3.75
4.68
5.20
52.62
45.64
26.25
36.73
4.68
7.52
52.54 15.68 6.53
S. platensis * Sufficient Nutrients 65.12 9.37 5.33
High Light (120 UE m-2 S-1)
High Temperature (38°C)
27.18 15.22 3.65
35.32 13.12 3.75
N- 1 imi t e d 32.81 30.31 5.43
P-limited 34.68 31.87 4.68
Sodium Chloride 0.1M 51.56 22.52 8.43
0.5M 37.50 32.10 10.31
Bicarbonate (4g/L) 41.25 13.25 5.62
*Experimental conditions were:
temperature 3OoC; light irradiance 80 uE I U - ~ s-l; air flow rate 300 ml/min; The values shown are averages of four independent determinations.
33

of Oscillatori agardhii (Riegman et. al., 1985), showed elevated levels of polysaccharide storage.
Depletion of dissolved phosphate-phosphorus from the culture medium of Anabaena was accompanied by a decline in chlorophyll a , protein, RNA and an increase in carbohydrate per unit dry
weight (Healey, 1 9 7 3 ) . 1) I I
Sodium Chloride, concentration influenced the storage of carbohydrates and proteins of both strains. As media (Zarrouk, see "Methods"), were enriched with 0.1 M and 0 . 5 M NaC1, the carbohydrate content of the cells increased, when compared t o that of the control (Zarrouk: 0.01 M NaCl), to 26.25%, 36.73% in S . maxima and to 22.52%, 32.10% in S . platensis. On the other hand the total protein decreased respectively to: 52.62% 45.64% in S . maxima and 5 1 . 5 6 % , 3 7 . 5 0 2 , in S . platensis. The lipid percentages showed little increase when compared to those of complete media (control). Many cyanobacteria are capable of adapting to a range of salinity in the environment by sythesizing internal osmotic support in the form of carbohydrates (Packer et. al., 1986).
Bicarbonate: When bicarbonate concentration of Zarrouch media was reduced to one quarter (4.g/L), neither strains showed much difference in the chemical composition as compared with the control media except their yield was somewhat below.the control.
34
w I
n H \
hD In
w v
w E c z 0 m d 4 u Y
m
w
d 0 X u E 3
0 cn
n H
Ej
0" u dp d
0" u OQ m 0
0
E In
0
E rl
0
r
z E 0 d a
E;
u
w E c d
3.1 X 0 m d 4 V
Q
n
CJ z z
0 0 0 0 0 0 W c3 W c3
;LHE)IBM 3INVE)UO d O 3 0 V & N 3 3 U 3 d
35

I I I I I I 0 0 0 0 0 0 W m W m
z w t-1 0 p: a
U
a w b c p:
* x 0 m p: c V
61
n
n U
U pc I4
;LHE)I3M 3 I N V D H O do 3 ! 3 V , L N 3 3 H 3 d
36

CONCLUSIONS :
In an attempt to optimize the biomass of cyanobacteria the growth parameters 0.f two strains were characterized in batch cultures to gain basic information to be considered in a continuous culture system.
"Spirulina",
Conclusions of the first year study are summarized as following:
. From the environmental variables studied, optimum growth temperature was clearly species specific. For S. maxima, 35°C and for S. platensis, 3OOC.
. Optimum light irradiation for both strains was 80 uE m-2 s-1.
. Both strains exhibited a tolerance for a wide range of pH, from 8 to 11 with optimum pH in range of 9 to 10.
. When studying the effect of flow rate of aeration and percen- age of C02 present in the air on the growth rate and yield of the alga; it was concluded that the 500 ml/min for aeration rate and 1% C02 gave optimum conditions of growth. It was concluded also that cultures supplied with air (0.03% C02) gave the same responsein terms of productivity as well as those supplied with 1% C02 enriched air, providing that the bicarbonate concentration present in the medium was reduced to 4 g/L instead of 16 g/L (Zarrouk medium).
. Variations in supply of nutrients: Nitrate-N, phosphate-P, sodium chloride, bicarbonate, affected the productivity rate of the alga. They not only influenced the production rates, but also the quality of the produced biomass as measured by the carbohydrate and total protein. In most of the cases the carbohydrate content increased when nutrients were limiting or in excess as sodium chloride concentration of 0.1 M and 0.5 M (see Table 1).
. The lipid percentage, in particular, did not show much increase in different culture treatments. But, increasing the tern erature of culturing to 38°C or the light irradiance t o 120 uE m-$ s-l, reduced the total lipids drastically. However, increasing sodium chloride to 0.1 M in the culturing media, the lipids increased somewhat higher than in the control media.
. The ability of the alga to utilize macroelements and micro- elements, and to convert it into biomass.
37

. Variation of single environmental regulants such as light intensities or temperature, during the present study also revealed their detrimental effect even when the cultures contain sufficient nutrients. Cultures whose growth rates and productivities were reduced by any factor, became pro- gressively more yellow (light, temperature, nutrient limit- ation, pH) and changed in morphology (Fig. 1 4 ) .
. A slight inverse relationship was observed between the protein content and carbohydrates which means that one increased at the expenses of the other. This suggests that quality of biomass may be manipulated for dietary purposes. An edequate supply of nutrients is therefore a pre-requisite for producing a uniform quality of biomass, which in turn could then be used in the formulation of diets. (see Sufficient Nutrients). The possibility of manipulating the quality of the biomass could have potential for the NASA/CELSS Program, when specific diet formuation is needed (e.g. low protein content).
. Overall algal productivity and quality could be manipulated by means of varying nutrient concentrations or temperature and light irradiance.
Further work is needed to characterize the efficiency of the algal cells under such environmental conditions in terms of gas exchange and energy loss or gain in steady state.
“It can be concluded that through manipulating environmental conditions of the algal growth, one can modify the photosynthetic products. Thus, Spirulina can be, through manipulating growth factors, used as palatable diet comparable to higher plants (see Appendix).”
38

Figure 14a. Cells of S. maxima grown under Optimum Conditions. Scale: 1 cm = 25 u
Figure 14b. Cells of S. m a x i m a grown under Stress Conditions. Scale: 1 cm = 40 u
39

FUTURE RESEARCH (1988/1989)
We plan to investigate the effects of: light irradiation, temperature, carbon dioxide on partitioning of macromolecules (protein, carbohydrates, lipids) and on elemental composition (carbon, hydrogen, oxygen) of Spirulina maxima. The manipulation of composition of Spirulina maxima by the environmental factors will be investigated at steady state.
Tasks :
I. Biomass (Dry Weight and Chlorophyll) and Cultivation Time
11. Dilution Rate and Dry Weight
111. Productivity and Dry Weight
IV. Effect of Light Intensity:
This experiment will be performed using the best temperature and aeration obtained from batch culturing. The growth rate as a function of light intensity will be determined at: 30 , 60, 100 uE m-2 s-l and as well as 35°C temperatures in relation to the productivity and efficiency of the alga.
V. Aeration and Carbon Dioxide Concentrations
The following experiments will be performed at steady state:
. Bubbling with air (0.03% C02) . Bubbling with air (0.1% C02) . Bubbling with air (1% C02) . Bubbling with air (5% C02) Experiments IV and V will be analyzed for the following:
. Dry Weight . Chlorophyll Productivity . Light effeciency . Particulate carbon . Particulate nitrogen
, Inorganic nitrogen and carbon . Total phosphorus . Proteins . Carbohydrates . 02 - evolution . C02 - measured in flux in and out of culture
40

EXPECTED OUTCOMES
. Relationhsip between dilution rate (growth rate) and dry weight
. Growth rate as function of light intensity e.g. 30, 60, 100 uE m'2 s-l
. Productivity and chemical analysis in relation to three light intensities will be determined in order to determine efficiency of the alga
. Relationship between C02 mass flux in (mg C. day'l) and algal carbon mass flux out (mg C. day'l) at fixed dilution rate.
. Relationship betwen C02 mass flux (in-out) at different concentrations and algal carbon flux out (mg C. day'l) at fixed dilution rate
. Relationship between C02 mass flux in (mg C. day'l) and algal carbon: nitrogen mass flux out (mg C. day'l)
. Ralationship between C02 mass flux in (mg. C. day'l) and algal com- position (carbohydrate, protein, lipid)
. Relationship between 02 evolved by the culture and C02 flux
. Efficiency evaluation of Spirulina biomass for CELSS, in terms of light, C02 and nutrient parameters.
41

FERERENCES
Cohen, Z., Vonshak, A, Richard, A.: 1987. Fatty acids Composition of Spirulina strains grown under various environmental conditions. Phytochen. 26: 2255-2258.
De la Noue, J., Cloutier-Plantha, L., Walsh, P., Picard, G.: 1984. Influence of agitation and aeration modes on biomass production by Oocystis. Sp. grown on wastewaters. Biomass 4: 43-58.
Ernst, A., Boger, P.: 1985. glycogen accumulation and the induction of nitrogenase activity in the heterocystforming cyanobacterium Anabaena variabilis. J. of Gen. Hicrobio. 131: 3147-3153.
Faucher, O., Coupal, B., Ledny, A.: 1979. Utlization of Seawater - urea as a culture medium for Spirulina maxima. Can. J. Microbiol. 25: 752-759.
Fogg, G. E.: 1956. Phytosynthesis and formation of fats in a diatom. Ann. Bot (Loud.) 20: 265-285.
Goldman, J. C., Graham, S. J.: 1981. Inorganic carbon limitation and chemical composition of tvo freshwater green microalgae. Appl. Environ. Microbiol. 41: 60-70.
Healey, F. P.: 1973. Characteristics of phosphorus deficiency in Anabaena. J. Phycol. 9: 383-394.
Healey, F. P.: 1975. Physiological indicators of nutrient deficiency in algae. Can. Fish. Xar. Serv. Res. Dev. Tech. Rep. 585: 30.
Konopka, A. Kromkamp, J., Mur, L. R.: vesicle content and buoyancy in light or phosphate-limited cultures of Aphanizomenon flos-aquae (Cyanophyceae). J. Phycol. 23: 70-78.
Lang, D. S., Brown, E. S.: 1981. Phosphorus-limited growth of a green alga and a bluejgreen alga.
1987. Regulation of gas
Appl. Environ. Hicrobiol. 42: 1002-1009.
Le-, M., Wober, G.: 1976. Accumulation, mobilization and turn- over of glycogen in the blue-green bacterium Anacystis nidulans. Archiv of Microbiol. 111: 93-97.
IkcElroy, B. D., Bredt, J.: 1985. Current concepts and future directions of CELSS, pp. 1-9, In: Controlled Ecological Life Support System, NASA CP-2378, Xgp COSPAR meeting, Graz, Austria, July 1984 (MacElroy, B. C., Smeroff, D. T., Klein, H. P., eds.).
42

Packer, L., Fry, I., Belkin, S.: 1986. Application of photosynthetic N2-fixing cyanobacteria to the CELSS program p. 339-352, In: Controlled Ecological Life Support Syste: CELSS 1985. Workshop NASA TM 88215.
Riegman, R., Rutgers, M., Mur, L. R.: 1985. Effects of photoperiodicity and light irradiance on phosphate-limited Oscillatoria agardhii in chemostat cultures. I. Photosynthesis and carbohydrrate storage. Archiv. of Microbiol. 142: 66-71.
Strickland, J. D., Parsons, T. R.: 1972, A practical handbook of seawater analysis. Bull. Fish. Res, Bd. CAn. 167.
Zarrouk, C. 1966. Contribution a 1' etude d'une cyanophycee. In- fluence de divers facteurs physiques sur la crossance et la photosynthesis de Spirulina maxima . (France).
Thesis, University of Paris
43

A P P E N D I X
4 4

45

ORfG1FUAL PAGE tS OF POUR QUALtTY
Three billion Year old exoert grants immunity
"I am the immortal descendant of the original life form. Over 3 billion years ago. blue-green algae produced our Dxygen atmosphere so life could :valve. Today, I offer your life imely health benefits.
Your own technology and life- style threaten you. Pollution. toxic :hemicals. radiation. disease, stress. irugs and processed foods anack your immune system. Damage to your body and its cells can cause premature aging and cancer.
Natural Life Preserver Vitamin supplements help out.
but with two limitations. First. synthetic mega-doses are wasted if your body can't absorb them and may even be toxic. But my whole food spirulina vitamins and minerals arc casily absorbed.
Second, because your bod! is not simply a mechanism, it needs more than isolated nuuicnts. Living cells need specific information to remem- ber their function. DNA molecular codes in natural foods contain genet- ic memories of successful life forms for millions of years. I have reju- venated myself since the beginning. My 3 billion years of cell memories can help your body remember iu powers and renew itself. When you need a natural life preserver. put mc IO work inside your body.
Unusually Concentrated Within me is a powerful concen.
tmtion of nutrients, unlike any other single plant. p i n . food or herb. 1 flourished in the nutrients of the original primordial soup. Many of these nutrients arc strongly recommended by scientists to build the immune system. New medical research has focused on my effects 0" choles.ternl reduction. cancer and immune system reaponse.
Protective Nutrients I have more beta carotene than
chlorella algae and ten times more than any other food. Beta carotene is known to lower the incidence of cancer and protect against UV radia- tion. l am the only food that con- tains significant amounts of gamma linolenic acid (GLA). the beneficial numcnt in evening primrose oil. 1 am the best source of vitamin B-12.
Extra iron and trace minerals added to my ponds make Eanhrise Spiru- lina the best iron food.
My living cells are dehydrated in seconds by low temperature spray drying, best preserving valuable nutrients. Then I'm freshly scaled in anti-oxidant containers. Eanhrise is the only company that can guarantee quality from my living ponds to my consumers.
sometimes deficient in vegetarian diets. I am rich in iron, essential for healthy red blood cells and a strong immune system.
I have the highest protein content (60-70'70). all the essential amino acids. and RNA and DNA nucleic acids I'm 1% cleansing chlorophyll ~ ~ b # , , h ~ ~ I
and 15% sfrenethenine Dhvcocvanin. a unique blue &ynen;fouAd only in blue-green algae. Clean Green Energy
Enjoy my energy every day- p a t between meals and before swenuous Naturattv Diaestible
1 am so old ;hat IFvolved betore hard cellulose cell walls. My cell walls are naturally soft proteins. My younger cousin. chlorella. requirch additional factory procehh- ing to break down its hard cell walls. But I'm already perfect - 95% digestible. The most digestible food. Many people say the! feel my energy within minutes.
activity for quick nutrition without feeling too full. I'm also helpful for dieting and cleansing your body.
Once known as il /nod o/ [he /rtrurc. I ani already growing in villages in the developing world. I'm called 'green medicine' food by the children.
So. panake of my immortal body each dav. Eat 3 billion wars of cell memory' and a conccntrihon of pro- tective nutrients. Reneu your own health. reneu your connection with your SISICN and brothers in the third world. and with the ongins of life."
1 S p i ~ l i ~ at healtn tood stores 1
I 1 m m 1 m 1 - 1 1 1
You can iind best selling Earthwe
I or order 11 direct from Earthrise. I Earthrise Quality Standard I Please send me free literature on: I
introduced as a health food supple- I Name I ment in 1979 by Earthrise. Now I'm enjoyed by millions world wide- athletes. vegetarians, dieters, health praciitioners. and people of all ages.
Earthrise is one of my most ad- vanced farms. Here. I'm grown free of pcsticidcs. hcrhicides. additives and prcacrvalivca. and 100% purc. L-----L-~--J San Ralaei CA 94915
Earthrise" Spirulina - nature's most protective food
46