formation mallory
TRANSCRIPT

Proc; Nati. Acad. Sci. USAVol. 76, No. 8, pp. 4112-4116, August 1979Medical Sciences
Formation and involution of Mallory bodies ("alcoholic hyalin") inmurine and human liver revealed by immunofluorescencemicroscopy with antibodies to prekeratin
(alcoholic hepatitis/liver pathology/keratin/intermediate filaments/tonofilaments)
HELMUT DENK*, WERNER W. FRANKEt, ROMANA ECKERSTORFER*, ERIKA SCHMIDt, ANDDONTSCHO KERJASCHKI**Division of Gastroenterologic Pathology and Hepatopathology (Hans Popper Laboratory), Department of Pathology, University of Vienna School of Medicine,Vienna, Austria; and tDivision of Membrane Biology and Biochemistry, Institute of Experimental Pathology, German Cancer Research Center,Heidelberg, West Germany
Communicated by Hans Popper, May 29, 1979
ABSTRACT Antibodies raised against prekeratin intenselyand specifically stain, in immunofluorescence microscopy,Mallory bodies ("alcoholic hyalin") present in livers of humanalcoholics and griseofulvin-treated mice. The high sensitivityof this method allows the identification of small distinct cyto-plasmic structures that are observed during early stages ofMallory body formation, especially frequent in the perinuclearcytoplasm, as well as during stages of Mallory body disinte-gration and disappearance, such as after withdrawal of the drug.In the latter situation, the prekeratin-containing small particlesexhibit a characteristic pattern of arrangement in the hepatocyteperiphery. Electron microscopy illustrates that such smal bodiesare heap-like aggregates of typical Mallory body filaments.Immunofluorescence studies with antibodies to isolated pre-keratin polypeptides from bovine hoof or muzzle epidermisshow that Mallory body filaments, in particular those in humanliver, are immunologically more closely related to prekeratinof tonofilaments from living epidermal cells (stratum spinosum).The data indicate that Mallory bodies contain a pathologic formof prekeratin-like material. They also suggest that disorders ofcytoskeletal structures of the intermediate-sized filament classare associated with specific diseases and can be visualized andcharacterized by immunofluorescence microscopy by usingantibodies to constitutive proteins of such filaments.
Mallory bodies (MBs) ("alcoholic hyalin") are unique cyto-plasmic inclusions, mostly observed in hepatocytes (1). Theirmajor and most characteristic components are randomly oi-ented, unbranched, rod-shaped filaments of 14-20 nm in di-ameter, which often exhibit a dense fimbriated coat (2-5). Theyare characteristic morphologic features of alcoholic hepatitis(1) but have also been observed in diverse liver disorders notrelated to alcoholism (3). Typical MBs can be experimentallyproduced in mouse liver by long-term griseofulvin feeding (6).The major constituents of experimentally induced MB filamentsare a group of polypeptides with apparent Mr values rangingfrom 48,000 to 66,000 (5), and a similar composition has re-cently been reported for MBs from human alcoholics (7). In thisrespect, MBs resemble prekeratins from different sources thatall contain characteristic polypeptides in the 47,000- to68,000-Mr range (see ref. 8 for references). In addition, it hasbeen shown that antibodies to prekeratin from bovine hoofhorny layer, in immunofluorescence microscopy, specificallydecorate murine MBs, which further strengthens the relation-ship between MB filaments and intermediate-sized filamentsof the prekeratin type present in most if not all epithelial cells("cytokeratins") (8). On the basis of these findings it has beenproposed that MB formation represents a pathologic form ofhyperkeratosis (5).
In the present communication the following findings arereported. (i) Antibodies t6 bovine prekeratin react with MBsof human origin, thus extending our original observation inexperimental animals (5) to human disease. (ii) Immunofluo-rescence microscopy using such antibodies as well as antibodiesto MBs of human origin allows the tracing of MB formation andinvolution by detection of early (precursor) stages of MB for-mation as well as MB fragments during involution. (iii) MBs ofhuman liver differ from those in livers of griseofulvin-treatedmice in that they immunologically crossreact with prekeratinfrom desmosome-attached tonofilamepts of bovine muzzle butnot with prekeratin polypeptide components of bovine hoof.(iv) Neither original nor colchicine- or griseofulvin-reinducedmurine MBs are related to the vimentin-type of intermedi-ate-sized filaments (9).
MATERIALS AND METHODSHuman Liver. Liver tissue from patients with alcoholic
hepatitis and cirrhosis was removed at autopsy. For comparison,samples of alcoholic fatty liver and of livers without pathologicalterations were examined.
Animals. Male Swiss albino mice (strain Him:OF 1 SPF;Institute for Laboratory Animal Research, University of ViennaSchool of Medicine, Himberg, Austria) were fed a powderedstandard diet (Altromin, LippeW West Germany) containing2.5% griseofulvin (Qlaxo, Greenford, England) as de-scribed (6). Animals were killed after 1, 2, 3, 4, 6, 8, and 12weeks of continuous griseofulvin feeding, and 1, 2, 3, and 10days and 3, 8, and 12 weeks after withdrawal of the drug inorder to trace development and involution of MBs. In addition,some mice were kept on the following regimen: continuousgriseofulvin treatment for 2.5 months, then 2.5 or 3 months ona drug-free diet, then on the griseofulvin-containing diet againfor 4 days prior to sacrifice. In another group of animals finalgriseofulvin refeeding was replaced by the administration ofcolchicine for 8 days (10) by using a colchicine dosage of 0.5,tg/g of body weight injected subeutaneously twice a day.
Antibodies. Antibodies were (i) guinea pig antisera to totalreconstituted, purified bovine hoof (including stratum cor-neum) prekeratin (5, 8); (ii) IgG from i made monospecific forprekeratin as described (11); (iii) guinea pig anitisera againstelectrophoretically separated individual polypeptides orpolypeptide fractions of bovine hoof prekeratin and desmo-some-associated tonofilaments from bovine muzzle (11-13);(iv) guinea pig IgG from antisera to prekeratin from humanepidermal tissue obtained from autopsy skin material that was
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be hereby marked "ad-vertisemennt" in accordance with 18 U. S. C. §1734 solely to indicatethis fact.
4112
Abbreviations: MBs, Mallory bodies; DT, indicates desmosome-asso-ciated tonofilament prekeratin component; PK, indicates prekeratincomponent from bovine hoof.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 15
, 202
2

Proc. Natl. Acad. Sci. USA 76 (1979) 4113
prepared as described for bovine hoof prekeratin (8); (v) guineapig IgG from antisera to human MBs isolated from autopsy liveraccording to French et al. (14) and further purified as describedby Franke et al. (5); (vi) guinea pig antisera against murinevimentin (9). Controls included preimmune sera and sera ab-sorbed with the specific antigens.
Immunofluorescence Microscopy. Small pieces of liverwere quickly frozen in isopentane cooled with liquid nitrogen.Immunofluorescence microscopy was performed on frozensections as described (5).
Electron Microscopy. Mice were anesthetized with ether,and the livers were perfused through the portal vein first withHanks' balanced salt solution and then with 2.5% glutaral-dehyde in 0.1 M sodium cacodylate buffer, pH 7.4. Small cubesof liver tissue were postfixed for at least 4 hr in the same fixative.They were then osmicated and processed for embedding andelectron microscopy of ultrathin sections according to con-ventional procedures.
RESULTSReaction of Murine and Human MBs with Antibodies to
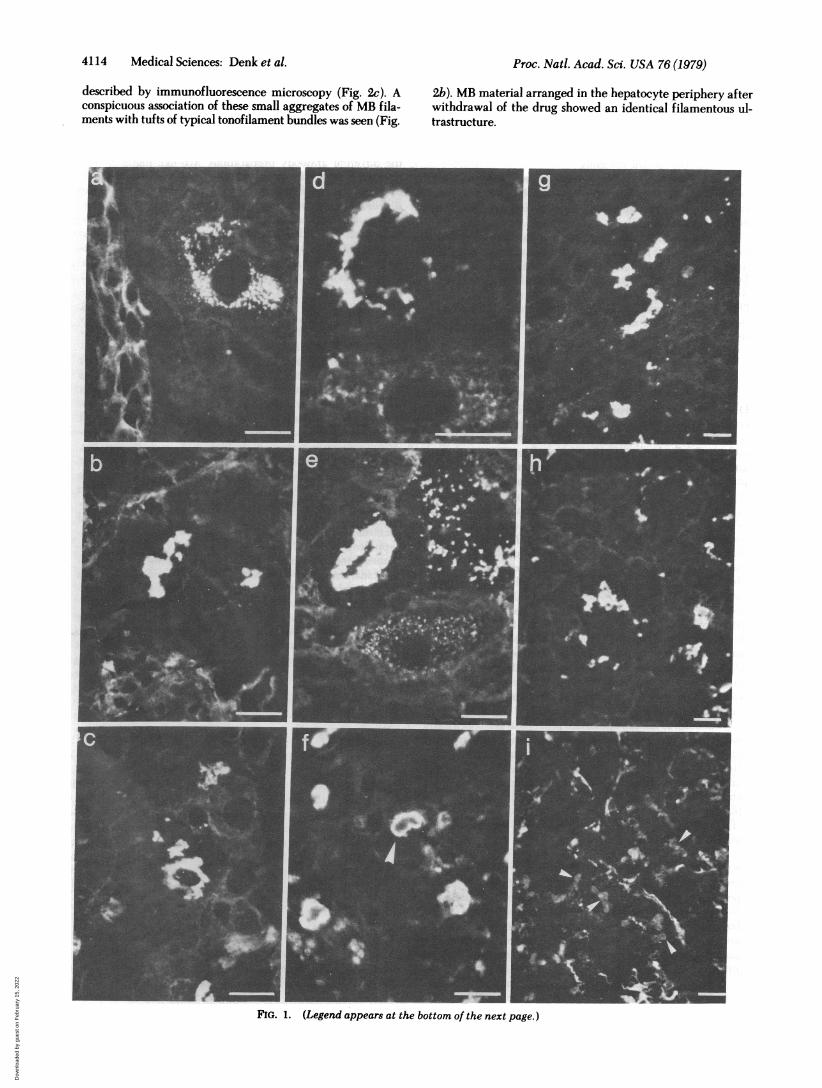
Diverse Prekeratin Preparations and to Human MBs. Anti-bodies to prekeratin from total bovine hoof epidermis and frombovine muzzle desmosome-associated tonofilaments stronglyand specifically decorated MBs in mouse liver (Fig. ig). Whenantisera to individual polypeptide fractions of bovine hoofprekeratin (11) were examined, antibodies to polypeptidefractions PK 1-4-i.e., components with apparent Mr valuesof 54,000-63,000-showed stronger reaction with mouse MBs(Figs. le, 2c, and 3b) than did antisera to components withlower molecular weights (Mr of 48,000-52,000). Antisera to themajor polypeptides electrophoretically separated from des-mosome-attached tonofilaments of bovine muzzle epidermisalso strongly stained MBs, and again antibodies to larger poly-peptides (Mr values: DT 1, 68,000; DT 2,60,000; DT 3,58,000)were more effective (e.g., Fig. la) than antisera to the smallerpolypeptides (Mr values: DT 4, 52,000; DT 5, 51,000) (8, 12).It is noteworthy that some of the antibodies to hoof prekeratinand desmosome-attached tonofilaments (especially PK 1-3 andDT 2, 4, and 5) intensely reacted with tonofilament structuresin bile duct epithelial cells (Fig. la) (15), whereas other pre-
keratin antibodies (PK 4 and 5, and DT 1 and 3) exhibited no
significant reaction.Antibodies to human epidermal prekeratin also specifically
stained MBs in mouse liver (Fig. lb), and so did antisera raisedagainst isolated human MBs (Fig. 1 d, f, and h and Fig 3b).However, our guinea pig antisera to human MBs did not sig-nificantly decorate fibrils of bile duct epithelia of mouse liverand cultured rat kangaroo (PtK2) cells but did decorate cyto-plasmic fibrils present in human (HeLa) and bovine (BMGE;cf. ref. 8) epithelial cells as well as in frozen-sectioned bovinemuzzle epidermis (data not shown).
Antibodies to bovine hoof prekeratin polypeptides stainedhuman MBs only poorly, if at all, in contrast to intensive dec-oration of human MBs with the antibodies to the large poly-peptides of desmosome-associated tonofilament prekeratin (DT1-3; Fig. 1c). Hepatocytes of normal human liver and of alco-holic fatty liver exhibited only very poor, if any, reaction withantibodies to prekeratin, as described in rodent liver (15), mostprobably reflecting the small amounts of tonofilamentspresent.MBs of murine and human origin were not stained to a sig-
nificant extent by antibodies to murine vimentin (Fig. li) andto actin.
Patterns of MB Formation and Involution. Mouse liverdamaged by griseofulvin administration was examined by usingantibodies to bovine prekeratin, to individual prekeratincomponents, and to MBs on consecutive sections of frozen livertissue. Agreement of immunolocalization was observed withthe different antibody preparations. MB-like material wasdetected in some mice as early as 4 weeks after commencementof griseofulvin feeding in the form of small granules that weredistributed either diffusely in the cytoplasm of scattered livercells or, more characteristically, in prominent perinuclear ar-rays. Such perinculear granules seem to be typical for earlystages of MB formation (Fig. 1 a, d, and e and Fig. 2). After 6weeks of continuous griseofulvin administration, sporadichepatocytes in centrolobular position with irregularly ramifiedMB-like structures and aggregates of confluent prekeratin-containing granules were observed. At 8 weeks, MB-containinghepatocytes were already numerous and were arranged ingroups of 4-10 cells, mostly in the center of the lobules sur-rounding the central veins. At this stage, MBs in many hepa-tocytes were large and exhibited irregular, reticular contoursor coiled organization. They were often, but not always, inparanuclear position, sometimes forming thick perinuclear rings(Fig. le). However, despite the presence of typical fully de-veloped MBs in some cells,, adjacent, hepatocytes often stillexhibited small granules of MB material or showed arrange-ments suggestive of stages of confluence of sma'll granules (Fig.1 d and e), indicating continuous development of MBs undersustained griseofulvin administration. Prolonged griseofulvinfeeding resulted in only quantitative changes of the pattern inthat hepatocytes containing MBs in various stages of develop-ment increased in number, and, consequently, the originalcentrolobular predominance of MB-containing liver cells dis-appeared. At such later stages, extracellular MBs were some-times detected by immunofluorescence microscopy in the si-nusoidal lumina or were taken up by Kupffer cells. MBs in cellsof hyperplastic nodules (6) were identical in their immuno-reactivity to those present in hepatocytes.
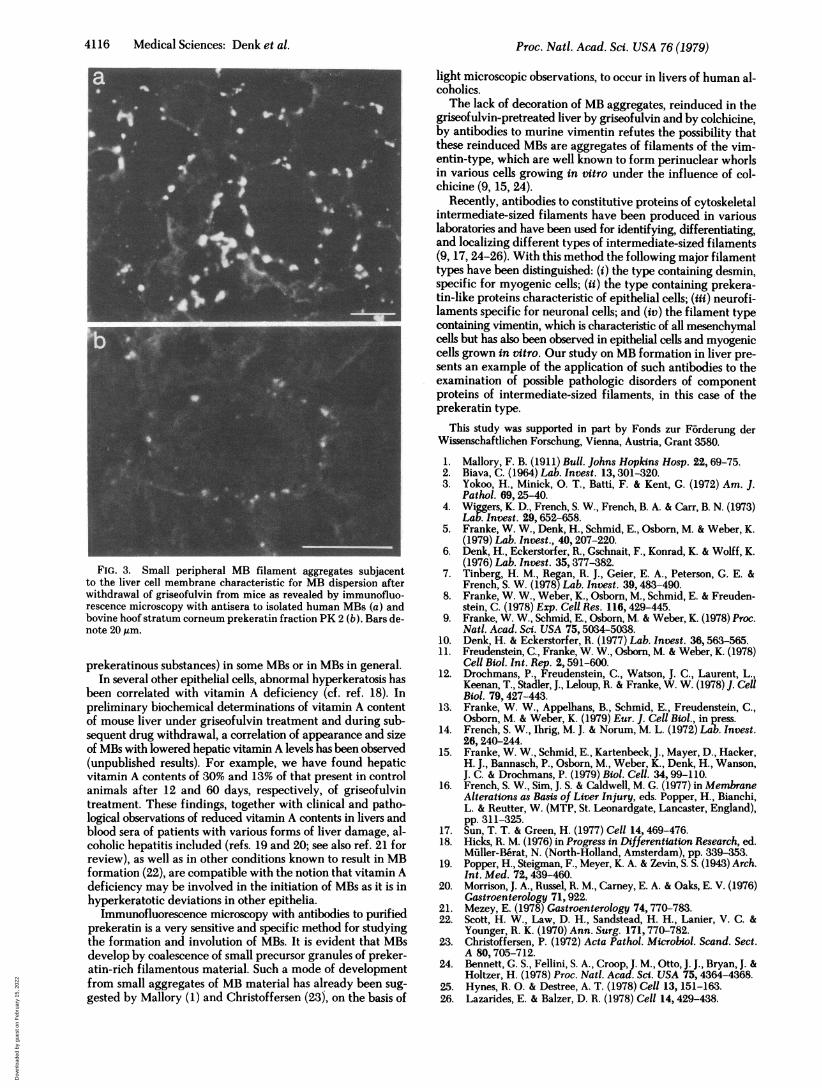
After discontinuation of griseofulvin feeding, the intensityof decoration of MBs with antibodies to prekeratin and to.human MBs progressively decreased, and often a nonfluorescentcore portion was visible (Fig. If). The number of fluorescingMBs gradually decreased. Concomitant with the disappearanceof the typical large MBs, small granules strongly decoratablewith antibodies to human MB-material and to prekeratin ap-peared at the periphery of the hepatocytes (Fig. 3). In somehepatocytes, mostly in centrolobular ones, these granules per-sisted for up to 2-3 months after drug withdrawal.When mice, which originally had been fed a griseofulvin-
containing diet for 3 months, were kept on a griseofulvin-freediet for 1-3 months and then griseofulvin was readministeredfor 4 days, large typical MBs reappeared within 4 days (cf. ref.6). A similar effect of fast reinduction is observed when col-chicine is injected for a total of 8 days (10). Such reinduced MBsreacted with antibodies to prekeratin and to human MBs butnot with antibodies to vimentin (Fig. 1 g-i). Typical MBs canbe reinduced with griseofulvin as well as with colchicine inlivers that still contain hepatocytes with peripheral granules ofMB-like material but also in livers in which no trace of MB-material is detected in the control animals.
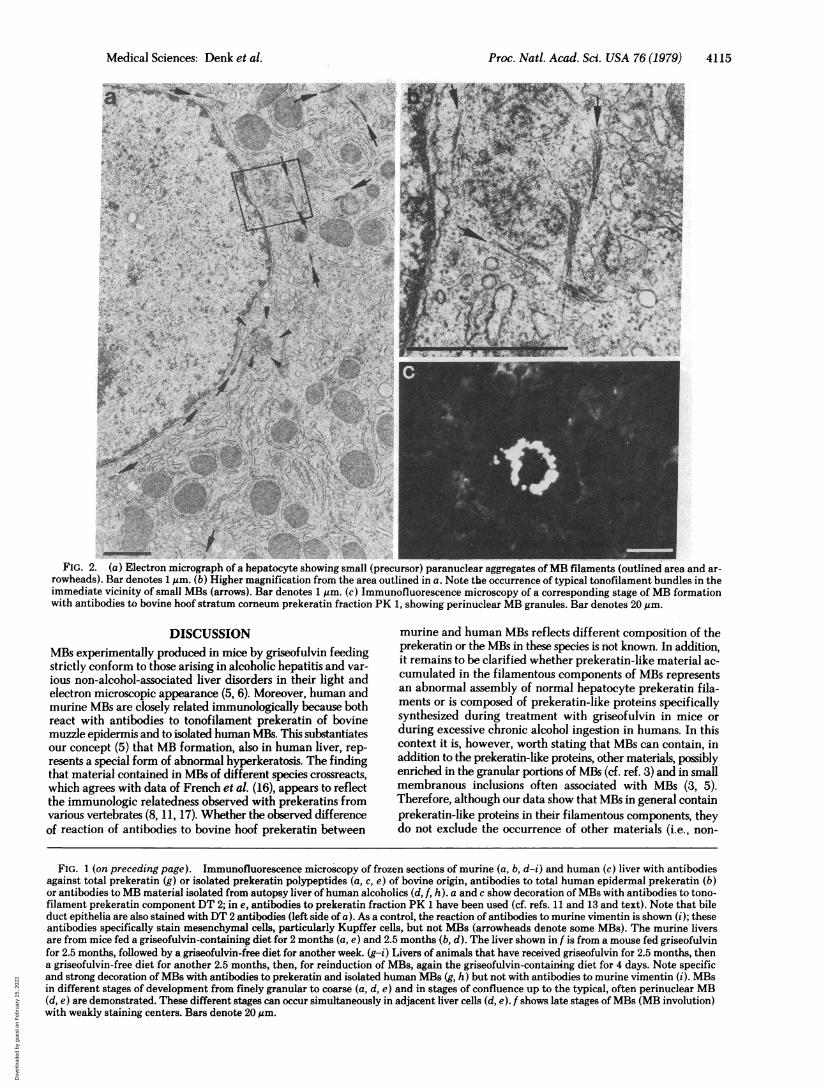
Electron Microscopy. When early stages of MB forma-tion-i.e., when only a few hepatocytes show granules positivelyreacting with antibodies to prekeratin or human MB-mate-rial-were examined by electron microscopy, small heaps(diameters ranging from 0.1 to 1 um) of randomly orientedtypical fimbriated filaments were seen in locations, oftenaround the nucleus (Fig. 2 a and b), corresponding to those
Medical Sciences: Denk et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 15
, 202
2
4114 Medical Sciences: Denk et al.
described by immunofluorescence microscopy (Fig. 2c). Aconspicuous association of these small aggregates of MB fila-ments with tufts of typical tonofilament bundles was seen (Fig.
2b). MB material arranged in the hepatocyte periphery afterwithdrawal of the drug showed an identical filamentous ul-trastructure.
FIG. 1. (Legend appears at the bottom of the next page.)
Proc. Natl. Acad. Sci. USA 76 (1979)
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 15
, 202
2
Proc. Natl. Acad. Sci. USA 76 (1979) 4115
4k%_~~~~~~~~~~~~~~~~~~
Gste
rowheads). 2d
itns r B o (I fmic p s o o
4ithaniodet oin oo trtmconumpekrtn rcto P ,shwngpriularM gauls ardnoe 2 m
ISCUSSION m 4n human reflectsdifferentcompositionofthe
V,, or the i.-~sexpr'mntllyCrducdmmiebglsefullnfe
45 -' '- -.** 1?aatreasobecafdhehrrkmaeal
4~~~
Its; Z 'ci4.1.1....~~~~~~~~c
strletly OlecoWrotoeasginacnlcnpxsAnavrFIG. 2. (a) Electron micrograph of a hepatocyte showing small (precursor) paranuclear aggregates ofMB filaments (outlined area and ar-rowheads). Bar denotes1nm. (b) Higher magnification from the area outlined in a. Note the occurrence of typical tonofilament bundles in theimmediate vicinity of small MBs (arrows). Bar denotes 1 am. (c) Immunofluorescence microscopy of a corresponding stage of MB formationwith antibodies to bovine hoof stratum corneum prekeratin fraction PK 1, showing perinuclear MB granules. Bar denotes 20im.
DISCUSSION murine and human MBs reflects different composition of theour conexperimenthally MBrformatione ainmic y umanlveredg cprekeratin or the MBs in these species is not known. In addition,
Mes ext perimnallpormodueibnormicebygrk iseoulifheefiding aditrmionstobheclrfewhtrprekeratin-likeprtisohe materials ac-ibl
strictly conform to those arising in alcoholic hepatitis and var- cmlrated in the filarmetious compnsofMBsr(f.re ma riprseanious non-alcohol-associated liver disorders in their light and mulated inclamentous assonents of MBs (3,e5).electron microscopic appearance (5, 6). Moreover, human and an abnormal assembly of normal hepatocyte prekeratin fila-murine MBs are closely related immunologically bause both ments or is composed of prekeratin-like proteins specificallyreact with antibodies to tonofilament prekeratin of bovine synthesized during treatment with griseofulvin in mice ormuzzle epidermis and to isolated human MBs. This substantiates during excessive chronic alcohol ingestion in humans. In thisour concept (5) that MB formation, also in human liver, rep- context it is, however, worth stating that MBs can contain, inresents a special form of abnormal hyperkeratosis. The finding addition to the prekeratin-like proteins, other materials, possiblythat material contained in MBs of different species crossreacts, enriched in the granular portions of MBs (cf. ref. 3) and in smallwhich agrees with data of French et al. (16), appears to reflect membranous inclusions often associated with MBs (3, 5).the immunologic relatedness observed with prekeratins from Therefore, although our data show that MBs in general containvarious vertebrates (8, 11, 17). Whether the observed difference prekeratin-like proteins in their filamentous components, theyof reaction of antibodies to bovine hoof prekeratin between do not exclude the occurrence of other materials (i.e., non-
FIG. 1 (on preceding page). Immunofluorescence microscopy of frozen sections of murine (a, b, d-i) and human (c) liver with antibodiesagainst total prekeratin (g) or isolated prekeratin polypeptides (a, c, e) of bovine origin, antibodies to total human epidermal prekeratin (b)or antibodies to MB material isolated from autopsy liver of human alcoholics (d, f, h). a and c show decoration of MBs with antibodies to tono-filament prekeratin component DT 2; in e, antibodies to prekeratin fraction PK 1 have been used (cf. refs. 11 and 13 and text). Note that bileduct epithelia are also stained with DT 2 antibodies (left side of a). As a control, the reaction of antibodies to murine vimentin is shown (i); theseantibodies specifically stain mesenchymal cells, particularly Kupffer cells, but not MBs (arrowheads denote some MMs). The murine liversare from mice fed a griseofulvin-containing diet for 2 months (a, e) and 2.5 months (b, d). The liver shown in f is from a mouse fed griseofulvinfor 2.5 months, followed by a griseofulvin-free diet for another week. (g-i) Livers of animals that have received griseofulvin for 2.5 months, thena griseofulvin-free diet for another 2.5 months, then, for reinduction of MBs, again the griseofulvin-containing diet for 4 days. Note specificand strong decoration of MBs with antibodies to prekeratin and isolated human MBs (g, h) but not with antibodies to murine vimentin (i). MBsin different stages of development from finely granular to coarse (a, d, e) and in stages of confluence up to the typical, often perinuclear MB(d, e) are demonstrated. These different stages can occur simultaneously in adjacent liver cells (d, e). f shows late stages of MBs (MB involution)with weakly staining centers. Bars denote 20 Aim.
Medical Sciences: Denk et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 15
, 202
2
4116 Medical Sciences: Denk et al.
light microscopic observations, to occur in livers of human al-coholics.The lack of decoration of MB aggregates, reinduced in the
griseofulvin-pretreated liver by griseofulvin and by colchicine,by antibodies to murine vimentin refutes the possibility thatthese reinduced MBs are aggregates of filaments of the vim-entin-type, which are well known to form perinuclear whorlsin various cells growing in vitro under the influence of col-chicine (9, 15, 24).
Recently, antibodies to constitutive proteins of cytoskeletalintermediate-sized filaments have been produced in variouslaboratories and have been used for identifying, differentiating,and localizing different types of intermediate-sized filaments(9, 17, 24-26). With this method the following major filamenttypes have been distinguished: (i) the type containing desmin,specific for myogenic cells; (ii) the type containing prekera-tin-like proteins characteristic of epithelial cells; (iii) neurofi-laments specific for neuronal cells; and (iv) the filament typecontaining vimentin, which is characteristic of all mesenchymalcells but has also been observed in epithelial cells and myogeniccells grown in vitro. Our study on MB formation in liver pre-sents an example of the application of such antibodies to theexamination of possible pathologic disorders of componentproteins of intermediate-sized filaments, in this case of theprekeratin type.
FIG. 3. Small peripheral MB filament aggregates subjacentto the liver cell membrane characteristic for MB dispersion afterwithdrawal of griseofulvin from mice as revealed by immunofluo-rescence microscopy with antisera to isolated human MBs (a) andbovine hoof stratum corneum prekeratin fraction PK 2 (b). Bars de-note 20,pm.
prekeratinous substances) in some MBs or in MBs in general.In several other epithelial cells, abnormal hyperkeratosis has
been correlated with vitamin A deficiency (cf. ref. 18). Inpreliminary biochemical determinations of vitamin A contentof mouse liver under griseofulvin treatment and during sub-sequent drug withdrawal, a correlation of appearance and sizeof MBs with lowered hepatic vitamin A levels has been observed(unpublished results). For example, we have found hepaticvitamin A contents of 30% and 13% of that present in controlanimals after 12 and 60 days, respectively, of griseofulvintreatment. These findings, together with clinical and patho-logical observations of reduced vitamin A contents in livers andblood sera of patients with various forms of liver damage, al-coholic hepatitis included (refs. 19 and 20; see also ref. 21 forreview), as well as in other conditions known to result in MBformation (22), are compatible with the notion that vitamin Adeficiency may be involved in the initiation of MBs as it is inhyperkeratotic deviations in other epithelia.
Immunofluorescence microscopy with antibodies to purifiedprekeratin is a very sensitive and specific method for studyingthe formation and involution of MBs. It is evident that MBsdevelop by coalescence of small precursor granules of preker-atin-rich filamentous material. Such a mode of developmentfrom small aggregates of MB material has already been sug-gested by Mallory (1) and Christoffersen (23), on the basis of
This study was supported in part by Fonds zur Forderung derWissenschaftlichen Forschung, Vienna, Austria, Grant 3580.
1. Mallory, F. B. (1911) Bull. Johns Hopkins Hosp. 22, 69-75.2. Biava, C. (1964) Lab. Invest. 13,301-320.3. Yokoo, H., Minick, 0. T., Batti, F. & Kent, G. (1972) Am. J.
Pathol. 69, 25-40.4. Wiggers, K. D., French, S. W., French, B. A. & Carr, B. N. (1973)
Lab. Invest. 29,652-658.5. Franke, W. W., Denk, H., Schmid, E., Osborn, M. & Weber, K.
(1979) Lab. Invest., 40, 207-220.6. Denk, H., Eckerstorfer, R., Gschnait, F., Konrad, K. & Wolff, K.
(1976) Lab. Invest. 35,377-382.7. Tinberg, H. M., Regan, R. J., Geier, E. A., Peterson, G. E. &
French, S. W. (1978) Lab. Invest. 39, 483-490.8. Franke, W. W., Weber, K., Osborn, M., Schmid, E. & Freuden-
stein, C. (1978) Exp. Cell Res. 116,429-445.9. Franke, W. W., Schmid, E., Osborn, M. & Weber, K. (1978) Proc.
Natl. Acad. Sci. USA 75,5034-5038.10. Denk, H. & Eckerstorfer, R. (1977) Lab. Invest. 36,563-565.11. Freudenstein, C., Franke, W. W., Osborn, M. & Weber, K. (1978)
Cell Biol. Int. Rep. 2,591-600.12. Drochmans, P., Freudenstein, C., Watson, J. C., Laurent, L.,
Keenan, T., Stadler, J., Leloup, R. & Franke, W. W. (1978) J. CellBiol. 79, 427-443.
13. Franke, W. W., Appelhans, B., Schmid, E., Freudenstein, C.,Osborn, M. & Weber, K. (1979) Eur. J. Cell Biol., in press.
14. French, S. W., Ihrig, M. J. & Norum, M. L. (1972) Lab. Invest.26,240-244.
15. Franke, W. W., Schmid, E., Kartenbeck, J., Mayer, D., Hacker,H. J., Bannasch, P., Osborn, M., Weber, K., Denk, H., Wanson,J. C. & Drochmans, P. (1979) Biol. Cell. 34, 99-110.
16. French, S. W., Sim, J. S. & Caldwell, M. G. (1977) in MembraneAlterations as Basis of Liver Injury, eds. Popper, H., Bianchi,L. & Reutter, W. (MTP, St. Leonardgate, Lancaster, England),pp. 311-325.
17. Sun, T. T. & Green, H. (1977) Cell 14, 469-476.18. Hicks, R. M. (1976) in Progress in Differentiation Research, ed.
Miiller-B3rat, N. (North-Holland, Amsterdam), pp. 339-353.19. Popper, H., Steigman, F., Meyer, K. A. & Zevin, S. S. (1943) Arch.
Int. Med. 72,439-460.20. Morrison, J. A., Russel, R. M., Carney, E. A. & Oaks, E. V. (1976)
Gastroenterology 71, 922.21. Mezey, E. (1978) Gastroenterology 74,770-783.22. Scott, H. W., Law, D. H., Sandstead, H. H., Lanier, V. C. &
Younger, R. K. (1970) Ann. Surg. 171,770-782.23. Christoffersen, P. (1972) Acta Pathol. Microbiol. Scand. Sect.
A 80, 705-712.24. Bennett, G. S., Fellini, S. A., CroopJ. M., Otto, J. J., Bryan, J. &
Holtzer, H. (1978) Proc. Natl. Acad. SVd. USA 75,4364-4368.25. Hynes, R. 0. & Destree, A. T. (1978) Cell 13, 151-163.26. Lazarides, E. & Balzer, D. R. (1978) Cell 14, 429-438.
Proc. Natl. Acad. Sci. USA 76 (1979)
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 15
, 202
2