formic and acetic acids in degradation products of plant ... · of plant volatiles elicit olfactory...
TRANSCRIPT

Published by Oxford University Press on behalf of US Government 2016. 325
Chemical Senses, 2016, Vol 41, 325–338doi:10.1093/chemse/bjw005
Original ArticleAdvance Access publication February 8, 2016
Original Article
Formic and Acetic Acids in Degradation Products of Plant Volatiles Elicit Olfactory and Behavioral Responses from an Insect VectorJustin George1, Paul S. Robbins1, Rocco T. Alessandro1, Lukasz L. Stelinski2 and Stephen L. Lapointe1
1Subtropical Insects and Horticultural Research Unit, United States Horticultural Research Laboratory, USDA-ARS, 2001 South Rock Road, Fort Pierce, FL 34945, USA and 2Entomology and Nematology Department, Citrus Research and Education Center, University of Florida, 700 Experiment Station Road, Lake Alfred, FL, 33850, USA
Correspondence to be sent to: Stephen L. Lapointe, Subtropical Insects and Horticultural Research Unit, United States Horticul-tural Research Laboratory, USDA-ARS, 2001 South Rock Road, Fort Pierce, FL 34945, USA. e-mail: [email protected]
Accepted 11 January 2016.
Abstract
Volatile phytochemicals play a role in orientation by phytophagous insects. We studied antennal and behavioral responses of the Asian citrus psyllid, Diaphorina citri, vector of the citrus greening disease pathogen. Little or no response to citrus leaf volatiles was detected by electroantennography. Glass cartridges prepared with β-ocimene or citral produced no response initially but became stimulatory after several days. Both compounds degraded completely in air to a number of smaller molecules. Two peaks elicited large antennal responses and were identified as acetic and formic acids. Probing by D. citri of a wax substrate containing odorants was significantly increased by a blend of formic and acetic acids compared with either compound separately or blends containing β-ocimene and/or citral. Response surface modeling based on a 4-component mixture design and a 2-component mixture-amount design predicted an optimal probing response on wax substrate containing a blend of formic and acetic acids. Our study suggests that formic and acetic acids play a role in host selection by D. citri and perhaps by phytophagous insects in general even when parent compounds from which they are derived are not active. These results have implications for the investigation of arthropod olfaction and may lead to elaboration of attract-and-kill formulations to reduce nontarget effects of chemical control in agriculture.
Key words: Asian citrus psyllid, electroantennography, insect feeding behavior, insect–plant interactions
Introduction
Plants produce volatile organic compounds (VOCs) such as monoter-penes, sesquiterpenes, terpenoid alcohols, acetates, and aldehydes by multiple metabolic pathways. They contribute to plant odor plumes involved in attraction of insects (Beyaert and Hilker 2014). Plant catabolic processes can break down and release these compounds as lower chain aldehydes, ketones and carboxylic acids by oxidative pathways (Oikawa and Lerdau 2013). Catabolism and degradation of VOCs can result from plant stress, environmental stress, herbivore
attack, plant–atmosphere interactions, etc., and can have important implications for plant-plant signaling and plant–insect interactions. For example, plants produce methanol as a byproduct of cell wall metabolism that is emitted at high levels during plant growth (Fall and Benson 1996; Hüve et al. 2007). Enzyme-mediated oxidation of methanol in plants produces formaldehyde that can be further oxi-dized to formic acid and CO2 (Cossins 1964). Ethanol, acetaldehyde and acetic acid are also produced during primary metabolism. Under stress conditions, ethanol can be transported from roots to leaves
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

and undergo enzyme-mediated oxidation to acetaldehyde and acetic acid (Kreuzwieser et al. 1999, 2001). The emission levels of these catabolic byproducts can be regulated by ozone (O3) levels, stress conditions, pollution and freezing events (Kimmerer and Kozlowski 1982) as well as pests or diseases. Measurements of net ecosystem exchange (flux) of ethanol, acetaldehyde and acetic acid above an orange grove (Citrus sinensis) found higher net fluxes of acetalde-hyde and acetic acid compared with ethanol. The ratio of catabo-lite (acetaldehyde, acetic acid) was found to be 676% and 311% greater, respectively, than the precursor (ethanol) (Park et al. 2013). Increased flux rates of catabolic or degradation byproducts such as acetic acid and formic acid may act as significant olfactory cues for insect herbivores including pathogen vectors.
Host plant finding is of paramount importance for phytophagous insects to fulfill their nutritional requirements and to find suitable oviposition sites (Bruce et al. 2005). Orientation of Diaphorina citri Kuwayama to host plants appears to be multimodal and includes visual, auditory and as yet unidentified chemical cues (Wenninger et al. 2009a, 2009b; Rohde et al. 2013). Olfactory cues based on VOCs and their catabolic byproducts or abiotic degradation prod-ucts may play an important role in host finding and feeding behavior of insects (Visser and Yan 1995; Raguso and Light 1998; Hansson et al. 1999; Tol and Visser 2002). Peripheral olfactory systems of insects consist of antennae and maxillary or labial palps that house olfactory receptor neurons (ORNs) important in the detection of host volatiles. Electrophysiological responses from insect antennae are processed and integrated within neural glomeruli in the insect brain resulting in behavior (Bruce et al. 2005). Onagbola et al. (2008) characterized eleven morphologically unique sensillae on the antennae of male and female D. citri. Some of these were identified as porous sensilla trichoidea and rhinarial plates with a likely olfac-tory function (Onagbola et al. 2008). The olfactory funtion of these was subsequently proven electrophysiologically with single sensil-lum recordings (Coutinho-Abreu et al. 2014a).
Diaphorina citri is a vector of the causal pathogen of the dev-astating citrus greening disease (aka huanglongbing in Chinese or HLB). The disease is caused by fastidious, phloem-restricted bacte-rium (Candidatus Liberibacter spp.) whose symptoms include chlo-rotic leaf mottling, shoot dieback, small, misshapen and bitter fruit and eventual tree death (Gottwald et al. 2007). The vector was first reported in Florida in 1998 (Halbert 1998) followed by the confir-mation of HLB in Florida in 2005 (Halbert 2005). Adult D. citri feed on various leafy parts of rutaceous plants but oviposition and nymphal development require tender young leaves and shoots. Host selection and feeding behavior are influenced by host plant vola-tiles (Halbert and Manjunath 2004; Bove 2006). Constitutive host plant volatiles and their role in host plant finding have been studied extensively for members of the Liviidae (Moran and Brown 1973; Nehlin et al. 1994; Soroker et al. 2004; Cen et al. 2005; Nissinen et al. 2008; Rouseff et al. 2008; Patt and Sétamou 2010; Onagbola et al. 2011; Patt et al. 2011; Poerwanto et al. 2012; Robbins et al. 2012).
Volatile stimuli emitted by flushing shoots may play an impor-tant role in the detection, location, and evaluation of potential host plants by D. citri (Patt and Sétamou 2010; Sule et al. 2012). Mann et al. (2012) reported that citrus plants infected with HLB were more attractive to D. citri than noninfected plants, possibly due to higher amounts of methyl salicylate released from infected compared with healthy plants. Induced release of methyl salicylate by psyllid feeding may act as a conspecific cue that could lead to increased transmis-sion of the HLB pathogen (Mann et al. 2012).
Very little is known about the orientation of D. citri to host plant volatiles or of insects in general to the degradation products of host plant volatiles. In the literature of insect–plant interactions, degrada-tion products of headspace volatiles have not been considered as insect kairomones. Instead, the primary focus has been on phyto-chemicals as they are released by the plant (Metcalf and Metcalf 1992). The current study reports a convenient method for recording electroantennograms (EAGs) from whole insect preparations of D. citri. We screened common host plant volatiles previously identified from headspace collected over young leaves of Rutaceae species of varying susceptibility to D. citri (Patt et al. 2011; Poerwanto et al. 2012). We report for the first time antennal response of D. citri to formic and acetic acids formed as degradation products of constitu-tive terpenoids. Oxidative byproducts such as formic and acetic acids produced from citrus VOCs may act as olfactory cues for D. citri to locate stressed plants that are more susceptible to insect attack and infection by the HLB pathogen. Antennal responses of D. citri to plant volatiles and their degradation products are discussed. Our results suggest that formic and acetic acids, possibly arising as deg-radation products from host plant volatiles or directly emitted by plants, may serve as important orientation cues for phytophagous insects and disease vectors.
Materials and methods
Asian citrus psyllidsA healthy (HLB-free) colony of D. citri was maintained at the US Horticultural Research Laboratory, Fort Pierce, FL. Insects were reared on seedlings of Citrus macrophylla Wester and maintained at 28 °C, 14:10 L: D. All insects used for recordings were adults between 8 and 10 days old.
ElectroantennographyIndividual D. citri adults were placed in the large end of 10 µL plastic pipette tips (USA Scientific) from which the tip had been cut so the resultant opening provided room for the head and antennae of the psyllid to project (Figure 1). Psyllids were placed in the large opening of the pipette tip and allowed to move upward toward the smaller opening. Using a small piece of cotton wool, psyllids were gently pushed partially through the small end of the plastic pipette tip until the head and antennae were exposed. The pipette tip was threaded to a custom stage (Figure 1) that held the psyllid in place prior to placement of electrodes. A filtered and humidified airstream passed over the antenna at 30 cm/s. Glass electrodes were created by pull-ing glass capillary cartridges (World Precision Instruments, TW150-4, 1.5 mm OD, 1.12 mm ID) in a capillary puller (World Precision Instruments) and then filling them with 0.5 M KCl. Lengths of sil-ver wire (0.5 mm diameter, Alfa Aesar) were inserted through the glass electrode holder and into the KCl solution. Using microman-ipulators, the indifferent electrode was inserted into the back of the insect’s head; the recording electrode encircled the distal end of the antenna as described by Wenninger et al. (2009b). A Kombi-Probe amplifier (Syntech) was used to acquire the signal from the antenna. The signal from the amplifier was conditioned using a Syntech IDAC-4 interface (Syntech). Signals acquired from the IDAC-4 were displayed and stored on a computer running Syntech software.
Odorant preparation and EAG recordingsAn initial survey of volatile plant odors produced by young emergent leaves of C. macrophylla was conducted. A sample of leaves from pot-ted plants was ground to a powder using liquid nitrogen. A portion
326 Chemical Senses, 2016, Vol. 41, No. 4
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

(2 g) was extracted with 100 mL of methanol:water (3:1). The aqueous methanolic extract was back-extracted 3 times with 50 mL aliquots of 1:1 ether:hexane. The combined extracts were dried over anhydrous sodium sulfate, concentrated and analyzed by GC-MS (Figure 2).
To test for direct release of acetic and/or formic acids by citrus plants, potted plants of three citrus genotypes were placed in sepa-rate glass collection chambers (0.2 m × 0.8 m, Analytical Research Systems, Inc.): C. macrophylla and two varieties of sweet orange, C. sinensis (Valencia and Madam Vinous). Headspace volatiles were collected using SPME (50 mm Carboxen/PDMS/DVB fiber) exposed for 1 h through a sampling port 1 h after introduction of the plants into the chambers and daily thereafter up to day 7. Condensate that appeared on the glass walls of the collection chamber was col-lected on filter paper, placed in a 20 mL sampling vial and sampled
by SPME for 1 h. Controls consisted of a chamber containing an identical pot with soil but no plant and a chamber into which 10 µL each of acetic and formic acid were introduced and allowed to vola-tilize. Samples were analyzed by coupled gas chromatograph-mass spectroscopy (GC-MS) using a ThermoElectron Trace GC coupled to a DSQII quadrupole MS. The GC-MS was equipped with a PTV injector operated in CT-splitless mode at 250 °C. After 2 min, the split and purge vents were pened with the split vent flow at 20 mL/min and purge vent flow at 3.5 mL/min. The oven program started at 10 °C for 4 min then increased at 4 °C/min to 280 °C with a 10 min hold. The MS was scanned for the first 5.5 min from m/z 10 to 200, then from 5.5 to 40 min from m/z40 to 400, and 40 min from m/z50 to 550. The MS source was maintained at 220 °C and the GC transfer line at 300 °C.
Figure 1. Custom stage for positioning whole insect preparations of Diaphorina citri for electroantennogram recording. Inset shows an adult psyllid constrained in a plastic pipette tip cut to allow protrusion of the insect’s head.
Figure 2. Chromatogram showing major odorant peaks from injection of a concentrated 1:1 ether hexane back extraction of aqueous methanolic extracts of young leaves of Citrus macrophylla, a preferred host of Diaphorina citri. Neral and geranial (double bond isomers together known as citral) and (Z) and (E)-β-ocimene were used in odorant degradation studies as they consistently showed degradation in glass cartridges under laboratory conditions. 1, α-pinene; 2, β-myrcene; 3, limonene; 4, (Z)-β-ocimene; 5, γ-terpinene; 6, linalool; 7, β-citronellal; 8, neral; 9, geranial; 10, citral; 11, δ-elemene; 12, isocaryophyllene; 13, α-humulene; 14, murolene; 15, dibutylhydroxytoluene; 16, germacrene B.
Chemical Senses, 2016, Vol. 41, No. 4 327
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

To determine antennal responses to individual odorants, log doses of purchased odorants were prepared in methylene chloride. Stimulus cartridges were prepared for electroantennography by pipetting 10 µL of each of the various odorant doses onto 0.75 cm × 3 cm strips of filter paper (Whatman No. 1) before insertion into glass Pasteur pipettes (14.5 cm). Pipettes were capped at both ends with 1000 µL disposable plastic pipette tips briefly exposed to flame to close the distal end. Stimulus cartridges were prepared fresh for each recording or allowed to age as described under EAG response to degradation products of citral and β-ocimene. The volatile con-tents of stimulus cartridges were wafted over the insect preparation by removing the pipette tip caps and connecting the large end of the Pasteur pipette to tubing connected to a Syntech Stimulus Controller (Syntech) that, upon actuation, delivered a 1 mL puff of filtered and humidified air through the stimulus cartridge and into the airstream 2 cm upwind from the insect.
For each recording, a blank stimulus cartridge containing filter paper was puffed over the antennae to control for nonspecific signals from mechanoreceptors reacting to pressure changes associated with the stimulus puff. The nonspecific response from the corresponding run was subtracted from antennal response values. One mg of formic acid (10 µL of 100 µg/µL) pipetted on to filter paper was puffed at the end of each recording as a positive control, given its known activity. In the initial experiment, 24 chemical odorants known to be present in citrus leaf extracts or volatiles, or that had elicited behavioral response from psyllids were tested (Table 1). All compounds were tested against one antenna of each of 3 male and 3 female D. citri.
EAG experiments were also performed using log doses (100 ng, 1 µg, 10 µg, 100 µg, 1 mg, 10 mg) of citral, β-ocimene, geranyl ace-tate, linalyl acetate, neryl acetate, β-caryophyllene, acetic acid, and formic acid (Sigma–Aldrich). Citral is any blend of its geometric isomers (Z, E) referred as neral and geranial, respectively. For odor-ant preparations and detection by EAG of citral and β-ocimene, the commercially available isomeric blends were used. Detection by gas chromatogram-coupled electroantennography (GC-EAD) allowed for antennal discrimination of the individual isomers. The individual odorants for EAG were prepared by serial dilutions in methylene chloride. Ten microliters of each dilution were placed on filter paper strips and allowed to dry in a fume hood before insertion into glass Pasteur pipettes. One antenna from each of 4 male and 4 female D. citri were tested against log doses of each odorant. A new set of log dose pipettes was used for each insect. Log doses were tested beginning with a methylene chloride control, followed by 100 ng, 1 µg, 10 µg, 100 µg, 1 mg, and 10 mg of odorants, and finally fol-lowed by 1 mg of formic acid as a positive control.
EAG response to degradation products of citral and β-ocimeneStimulus cartridges were prepared with citral or β-ocimene [(Z-) and (E-) isomers] tested at 0, 3, 6, and 9 days after preparation. Both compounds were tested at 100 µg, 10 mg, and neat by applying 10 µL of each dilution to filter paper strips. For the neat dose, 10 µL of pure odorant (99.7% or 95%) (Sigma–Aldrich) was applied directly to the filter paper strips. The aged cartridges were puffed over one antenna of each of 4 male and 4 female D. citri. A new set of pipettes was used for each individual. A negative control (meth-ylene chloride) was puffed at the beginning of each run. One mil-ligram of formic acid was puffed at the end of each run as a positive control.
GC-EAD analysisThe GC-EAD system consisted of an Agilent 7890A GC equipped with a SOLGEL-WAX capillary column 30 m × 0.25 mm (ID) × 0.25 μm film (SGE Incorporated) and a post-column glass Y-tube splitter (Supelco) dividing the column effluent in a 1:1 ratio between the flame ionization detector (FID) and a heated (200 °C) EAD trans-fer line (Syntech). The heated transfer line emptied into a charcoal-filtered, humidified air stream (30 cm/s) that carried the effluent over the antennal preparation. Lengths of deactivated column (0.32 mm ID) were used to carry the column effluent from the Y-tube splitter to the FID and into the humidified air stream. EAG and FID sig-nals acquired from the IDAC-4 interface were displayed and stored on a computer running Syntech GC-EAD software. At the start of each GC-EAD run, the GC oven temperature was held at 35 °C for 3 min, increased to 260 °C at 15 °C/min with a 10 min final hold. Injector and FID temperatures were 220 °C and 300 °C, respectively. Splitless injection was used with helium as the carrier gas at 2 mL/min. Additional makeup helium gas was added before the effluent splitter at 10 mL/min.
Antennally active peaks from the aged citral or β-ocimene were identified via GC-EAD by extracting filter paper strips from stimulus cartridges that had been aged for 10 days. Filter paper strips from 10 aged cartridges of each compound were soaked in 4 mL of meth-ylene chloride solvent for 24 h. Filter paper strips were removed and the remaining extract was concentrated to 1 mL under nitrogen. For each GC-EAD analysis, 1 μL of extract was injected. One antenna from each of 2 male and 2 female D. citri were tested against each extract.
Table 1. Mean ± SEM (n = 3) electroantennogram responses from male and female Diaphorina citri to odorants present in headspace volatile collection from Citrus macrophylla leaf flushes
Citrus odorants (100 µg) Male response (mV) Female response (mV)
β-Pinene 0.00 ± 0.00 0.03 ± 0.01Sabinene 0.00 ± 0.00 0.16 ± 0.27D-Limonene 0.18 ± 0.28 0.10 ± 0.21(Z)&(E)-β-Ocimene 0.10 ± 0.23 0.04 ± 0.12γ-Terpinene 0.00 ± 0.00 0.07 ± 0.11α-Humulene 0.00 ± 0.00 0.04 ± 0.08β-Caryophyllene 0.00 ± 0.00 0.00 ± 0.00(Z)-2-Hexenol 0.04 ± 0.18 0.06 ± 0.02(Z)-3-Hexenol 0.00 ± 0.00 0.23 ± 0.33(R)-(−)-Linalool 0.05 ± 0.19 0.00 ± 0.00β-Citronellol 0.03 ± 0.19 0.04 ± 0.08Geraniol 0.00 ± 0.00 0.04 ± 0.01(E)-2-Hexenal 0.00 ± 0.00 0.01 ± 0.02Nonanal 0.00 ± 0.00 0.08 ± 0.02Citral 0.30 ± 0.31 0.02 ± 0.13Methyl salicylate 0.05 ± 0.23 0.09 ± 0.20Linalyl acetate 0.11 ± 0.03 0.24 ± 0.11Geranyl acetate 0.21 ± 0.15 0.16 ± 0.24Neryl acetate 0.04 ± 0.09 0.12 ± 0.14Lauric acid 0.14 ± 0.23 0.02 ± 0.01Myristic acid 0.00 ± 0.00 0.00 ± 0.00Pentadecanoic acid 0.00 ± 0.00 0.11 ± 0.12Palmitic acid 0.58 ± 0.57 0.02 ± 0.00Stearic acid 0.00 ± 0.00 0.09 ± 0.13Formic acid (1 mg) 3.10 ± 0.57 3.00 ± 0.36
Responses were adjusted by subtracting responses to clean air during the corresponding run from odorant responses. 100 µg of synthetic odorant was applied to filter paper in glass stimulus cartridges and puffed into a filtered and humidified airflow upstream from the antenna. Formic acid (1 mg) was puffed at the end of each odorant run as a positive control.
328 Chemical Senses, 2016, Vol. 41, No. 4
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

GC-MS analysisChemical analysis of degradation products was accomplished by GC-MS using a ThermoElectron Trace GC coupled to a DSQII quadrupole MS controlled by Xcalibur software. The GC-MS was equipped with a PTV injector, operated in the CTSPLIT mode and a 30 m × 0.25 mm × 0.25 µm film DB5MS fused silica open tubular column (Agilent) using helium carrier gas at 1 mL/min. The injector temperature was maintained at 220 °C with a split flow of 10 mL/min and a constant septum purge. The oven temperature program started at 30 °C with a 1 min hold and then increased at 4 °C/min to 240 °C. The transfer line from the GC to the MS was maintained at 250 °C. The MS source was maintained at 200 °C and the MS was scanned from m/z 10 to 200 for the first 5 min of the run then scanned from m/z 40 to 400 for the remainder of the run. Compounds were identified by retention index (RI) of the eluted peaks and the background-subtracted mass spectrum averaged across the total ion chromatogram (TIC) peak or across a characteristic extracted ion current peak when co-elution of multiple compounds was detected. The RI database used for identification was generated at USHRL using the same column and GC-MS system. Mass spectra were com-pared with those recorded in the NIST Library. For identification purposes, RIs had to match within 5 RI units of the average database value and the reverse search of the sample spectrum had to achieve a match of >500 of 1000.
Degradation of citral and β-ocimene to oxidative byproducts was studied over a period of 9 days by collecting headspace vola-tiles from the Pasteur pipettes stored under laboratory conditions. Toluene was used as a solvent to wash the inside of glass pipettes when needed. Headspace volatiles in Pasteur pipettes loaded with citral or β-ocimene were analyzed by GC-MS at 0, 3, 6, and 9 days after loading. A 1 μL injection in the splitless mode was made using a gas-tight syringe to sample cartridges through a natural rubber septum capping the large end of the Pasteur pipette. Identification of antennally active peaks detected by GC-EAD was done using GC-MS by injecting 1 μL of each of the previously described filter paper extracts of aged citral or β-ocimene.
Statistical analyses of antennal responses to headspace over aged stimulus tubes containing citral and ß-ocimene, and to log doses of 8 citrus volatiles were performed using Statistix software (Statistix 9). Tukey’s HSD test was used for post hoc means comparison (α = 0.05) after a significant ANOVA using GLM procedures.
Psyllid probing behaviorA choice assay was designed to study the probing behavior of D. citri in response to odorants. This assay measured both insect orientation from several cm to the source of volatiles (olfaction) and subsequent probing behavior that may result from a combination of olfaction and gustation upon contact with a wax substrate containing odor-ants. Test compounds were incorporated in a slow-release wax matrix for volatiles (SPLAT, ISCA Technologies, Inc.) and offered to caged D. citri adults (Patt et al. 2013). Release matrix blends were prepared by adding 6 µL of green food coloring to 10 mL of white SPLAT (resulting in a yellow-green mixture), which was mixed in a vortex rotor for 5 min and combined with odorant compounds. Treatments (SPLAT plus odorants) were applied as narrow strips or beads (2.0 cm × 0.5 cm × 0.1 cm) to glass cover slips (22 mm × 22 mm, Fisherbrand Microscope Cover Glass 12-542-B). Each cover slip received approximately 0.17 mL of SPLAT and 0.7 µL of the odorant(s). The beads were air-dried for 2 h and placed in a com-pletely randomized pattern with replications on the floor of a cubi-cal cage (60 cm × 60 cm × 60 cm, BioQuip). Cohorts of 200 5- to
8-day-old D. citri adults were starved for 6 h and then released into each cage and allowed to probe for 21 h. Cages were held in a tem-perature and humidity controlled incubator at 26 °C, 75% RH, 24 h light conditions L:D. To visualize stylet sheaths produced by feeding attempts on the SPLAT beads, the cover slips were removed from the cages and beads were stained with Coomassie blue dye, washed in water, and allowed to air-dry. The number of salivary sheaths in each bead was counted under a stereomicroscope at 4× magnification.
Two experiments were performed using response surface meth-ods to identify primary drivers in a 4-component mixture and then in a 2-component mixture-amount design to identify an optimal blend of the primary drivers for maximum probing by D. citri. Experiment designs were generated and analyzed with Design-Expert software (v9, Stat-Ease, Inc.). For the 4-component mixture, the total amount of odorant was constant at 40 µL/10 mL SPLAT. Odorants were formic acid, acetic acid, β-ocimene, and citral. A modified D-optimal mixture design sufficient to satisfy a Scheffé cubic polynomial response surface model (Cornell 2002) consisting of 24 odorant combinations was used (Table 2). The entire experi-ment was replicated 6 times for a total of 144 individual cover slips in 6 blocks (cages). The same design was used in each cage but with unique randomizations of treatments within each cage. All cages were kept in the same environmental chamber at 26 °C and popu-lated simultaneously with cohorts of D. citri. In addition to those needed to satisfy model terms, points (odorant combinations) were added to estimate lack of fit and several points were duplicated to attain sufficient degrees of freedoms to estimate pure error across the design space, provide estimates of block effects, and to attain a near uniform leverage for all points (Weisberg 1985). The resulting quadratic design had 5 block, 9 model, 87 lack of fit, and 42 pure error degrees of freedom (Myers and Montgomery 2002). Based
Table 2. Proportions of 4 odorant components varied in a 3-dimen-sional mixture design to study probing response of Diaphorina citri
Blend Formic acid Acetic acid (Z)-β-Ocimene Citral
1 0.00 0.00 0.00 1.002 0.00 0.00 0.00 1.003 0.00 0.00 0.50 0.504 0.00 0.00 1.00 0.005 0.00 0.00 1.00 0.006 0.00 0.33 0.33 0.337 0.00 0.50 0.00 0.508 0.00 0.50 0.50 0.009 0.00 1.00 0.00 0.0010 0.00 1.00 0.00 0.0011 0.13 0.13 0.13 0.6312 0.13 0.13 0.63 0.1313 0.13 0.63 0.13 0.1314 0.25 0.25 0.25 0.2515 0.25 0.25 0.25 0.2516 0.38 0.13 0.13 0.3817 0.50 0.00 0.00 0.5018 0.50 0.00 0.50 0.0019 0.50 0.00 0.50 0.0020 0.50 0.50 0.00 0.0021 0.50 0.50 0.00 0.0022 0.63 0.13 0.13 0.1323 1.00 0.00 0.00 0.0024 1.00 0.00 0.00 0.00
Total amount of odorant was constant at 0.7 µL incorporated into each bead of wax substrate.
Chemical Senses, 2016, Vol. 41, No. 4 329
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

on the results from the 4-component blend experiment, we built a 2-component mixture-amount design to identify the optimal blend and concentration of formic and acetic acids to encourage probing of the wax substrate by D. citri. An I-optimal design sufficient to satisfy a Scheffé cubic × cubic polynomial consisting of 40 runs was used (Table 3) with 4 replications of the compete design in separate cages. The total amount of odorant varied between 10 and 40 µL. The design had 8 model, 12 lack of fit, and 19 pure error degrees of freedom. Blend proportions selected by the design were produced and assayed for probing response.
Statistical analyses of response surfacesLapointe et al. (2008) provided a detailed description of the statisti-cal methods used to evaluate similar blend designs. For the probing assays, models were calculated with Design-Expert. Model selec-tion was then based on a lack of aliased terms, low residuals, a low P value, nonsignificant lack of fit, a low SD, high R2 and a low PRESS (prediction error sum of squares) value. The selected models were then evaluated by adequacy tests described by Anderson and Whitcomb (2005). A Box–Cox plot (Box and Cox 1964) was used to select the square-root transformation of count data. The optimal blend of formic and acetic acids was calculated with Design-Expert using a simplex hill-climbing algorithm (Anderson and Whitcomb 2005) in a multi-dimensional pattern search that converges at a sta-tionary point. The parameter for optimization was maximum num-ber of probes within the range of odorant proportions tested. The control (no odorant) was, by definition, not included in the 4-com-ponent mixture design space because there is no possible intercept in a mixture. To test the hypothesis that the best blend resulted in a greater number of probes than the unscented control, the number of probes on the control was compared by ANOVA with the number of probes observed on the actual blend design point that fell closest to the predicted optimum blend. This prediction was then validated
in the 2-component mixture-amount experiment and the predicted optimum was compared with the control treatments to determine if increased probing resulted from inclusion of odorants in the wax matrix.
Results
Methanolic extracts of citrus leaf flushes contained a variety of volatile compounds including α-pinene, β-myrcene, limonene, (Z)-β-ocimene, linalool, citral (neral + geranial), β-caryophyllene and others (Figure 2). These and other odorants were also identi-fied from the headspace volatiles collected by SPME over 3 citrus varieties (Supplementary Figure S1). β-Ocimene, neral and geranial were detected in most plant headspace samples. Formic and acetic acids were not detected in the headspace samples, nor were they detected in the samples of the moisture condensate from the sides of the sampling jars. Acetic and formic acids were readily detected after spiking the collection chamber with the acids (Supplementary Figure S1).
Odorants associated with citrus leaf volatile components did not elicit significant EAG responses from D. citri antennae (Table 1). Antennae from male or female D. citri did not respond to stimulus cartridges loaded with citral or β-ocimene (<3 h-old). However, the same sealed stimulus cartridges elicited large responses from antennae of both male and female D. citri after 3 to 6 days at ambient laboratory conditions (23 °C) suggesting that degradation products were responsible for the electrophysiologi-cal response.
GC-MS analysis revealed a large and rapid decline in both (Z)- and (E)-β-ocimene (Figure 3A) and citral (Figure 3C) in stimulus cartridges after 3 days and concomitant appearance of degradation products. Significantly lower amounts of (Z)- and (E)-β-ocimene (P = 0.0007 and P = 0.0005, respectively) were present in the stimu-lus cartridges on day 3 and successive days compared with fresh stimulus cartridges prepared on day 0. The stimulus cartridges ini-tially loaded with β-ocimene contained higher amounts of acetic acid than formic acid and these degradation products were great-est on day 6 (Figure 3B). The peak areas were similar for formic and acetic acids in the stimulus cartridges initially loaded with citral (Figure 3D). Peak areas of acetaldehyde, acetone, α,α-dimethyl allyl alcohol, acetic acid and formic acid increased over the duration of the experiment (Figure 4).
EAG response to degradation products of citral and β-ocimeneEAG recordings of male and female D. citri showed responses to citral and β-ocimene degradation products on days 3, 6, and 9. Antennae of both sexes were not responsive to fresh citral or β-ocimene (day 0) at 100 µg, 10 mg, or neat doses (Figure 5). The degradation of citral elicited significantly higher antennal responses on day 9 (F2,11 = 7.64; P = 0.01) from males compared with females, but no significant antennal responses were observed on day 0, 3, and 6 [(F2,11 = 1.08; P = 0.38) (F2,11 = 3.50; P = 0.07) and (F2,11 = 4.16; P = 0.05), respectively] to the odorant doses tested (Figure 5A). The overall male antennal responses showed a dose effect (F2,47 = 15.08; P = 0.000) and no effects of day (F3,47 = 1.43; P = 0.25) or dose × day interaction (F6,47 = 0.94; P = 0.48) were observed. However, the dif-ferent citral odorant doses did not elicit significant female antennal responses on days 0, 3, 6, and 9 [(F2,11 = 0.38; P = 0.69) (F2,11 = 0.50; P = 0.62) (F2,11 = 0.47; P = 0.63) and (F2,11 = 4.11; P = 0.054), respec-tively] (Figure 5B).
Table 3. Proportion and amounts of formic and acetic acids var-ied in a 2-component mixture-amount design to study probing re-sponse of Diaphorina citri
Blend Proportion Amount (µL) Replicates
Formic acid Acetic acid
1 1.00 0.00 10 32 1.00 0.00 25 23 1.00 0.00 40 34 0.75 0.25 18 15 0.75 0.25 30 16 0.75 0.25 36 17 0.50 0.50 10 28 0.50 0.50 25 49 0.50 0.50 32 110 0.50 0.50 40 211 0.38 0.63 10 212 0.38 0.63 20 213 0.38 0.63 30 214 0.38 063 40 215 0.25 0.75 14 116 0.25 0.75 36 117 0.13 0.88 20 118 0.00 1.00 10 319 0.00 1.00 25 220 0.00 1.00 32 121 0.00 1.00 40 3
330 Chemical Senses, 2016, Vol. 41, No. 4
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

Male and female psyllids showed higher antennal responses on day 3 onwards with log doses of citral and β-ocimene degrad-ing to new byproducts. Degradation of β-ocimene over 3, 6, and 9 days produced significantly higher antennal responses com-pared with day 0 (α = 0.05). There were significant differences in antennal responses between the different odorant doses from males (Figure 5C) [(F2,11 = 7.29; P = 0.01) (F2,11 = 15.35; P = 0.001)
(F2,11 = 10.33; P = 0.004), respectively] and females (Figure 5D) [(F2,11 = 12.42; P = 0.002) (F2,11 = 9.19; P = 0.006) (F2,11 = 52.00; P = 0.0001), respectively] on day 3, 6, and 9. Male antennal responses to log doses of β-ocimene were significant (F2,35 = 29.32; P < 0.0001; n = 4), and no effects of day (F2,35 = 0.75; P = 0.48) or day × dose interaction (F2,35 = 1.01; P = 0.42) were observed. Female responses to log doses of β-ocimene treatment were also
Figure 3. Mean ± SEM (n = 6 for β-ocimene, n = 2 for citral) relative amount by TIC of neat β-ocimene (A, n = 6) and citral (C, n = 2) in glass stimulus cartridges over 9 days and concurrent formation of formic and acetic acids in the same stimulus cartridges initially containing β-ocimene (B, n = 6) and citral (D, n = 2).
Figure 4. Chromatogram showing degradation products in the headspace of glass cartridges initially containing neat (Z) and (E)-ß-ocimene after 6 days. Inset shows the TIC (top) and, below are extracted ion current profiles used for identification of formic acid (mol.wt = 46) and acetic acid (mol.wt = 60) of toluene washes of glass cartridges.
Chemical Senses, 2016, Vol. 41, No. 4 331
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from
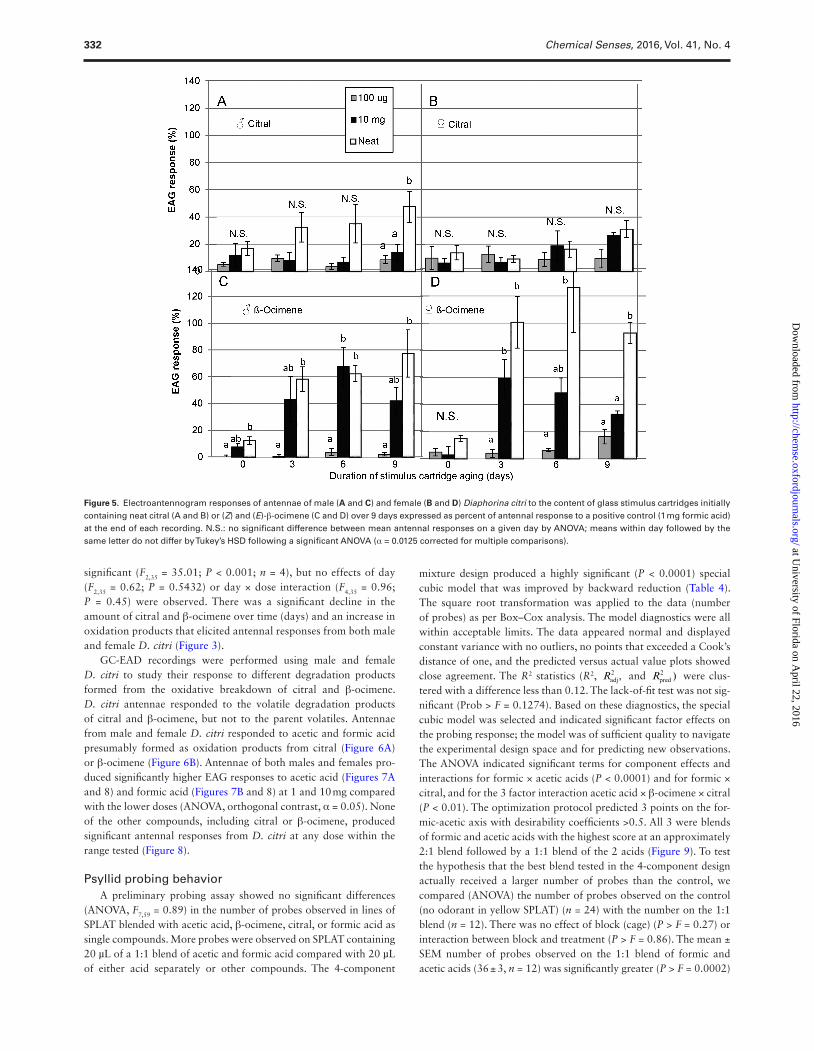
significant (F2,35 = 35.01; P < 0.001; n = 4), but no effects of day (F2,35 = 0.62; P = 0.5432) or day × dose interaction (F4,35 = 0.96; P = 0.45) were observed. There was a significant decline in the amount of citral and β-ocimene over time (days) and an increase in oxidation products that elicited antennal responses from both male and female D. citri (Figure 3).
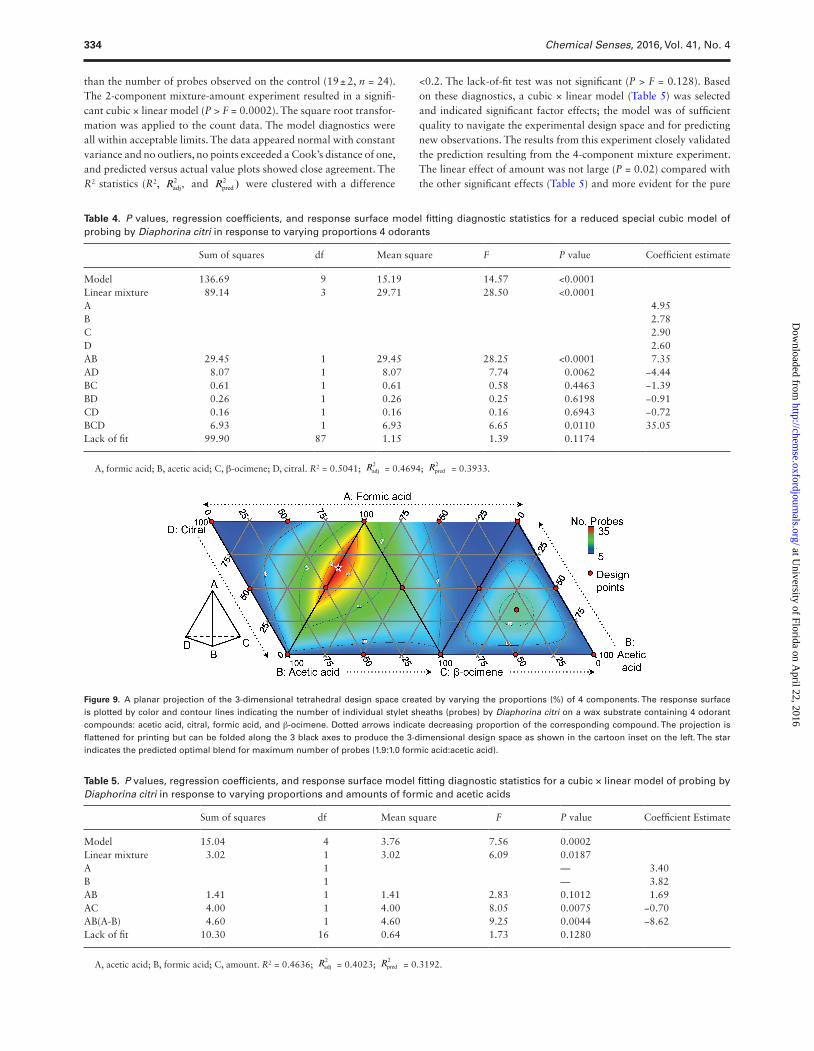
GC-EAD recordings were performed using male and female D. citri to study their response to different degradation products formed from the oxidative breakdown of citral and β-ocimene. D. citri antennae responded to the volatile degradation products of citral and β-ocimene, but not to the parent volatiles. Antennae from male and female D. citri responded to acetic and formic acid presumably formed as oxidation products from citral (Figure 6A) or β-ocimene (Figure 6B). Antennae of both males and females pro-duced significantly higher EAG responses to acetic acid (Figures 7A and 8) and formic acid (Figures 7B and 8) at 1 and 10 mg compared with the lower doses (ANOVA, orthogonal contrast, α = 0.05). None of the other compounds, including citral or β-ocimene, produced significant antennal responses from D. citri at any dose within the range tested (Figure 8).
Psyllid probing behaviorA preliminary probing assay showed no significant differences
(ANOVA, F7,59 = 0.89) in the number of probes observed in lines of SPLAT blended with acetic acid, β-ocimene, citral, or formic acid as single compounds. More probes were observed on SPLAT containing 20 µL of a 1:1 blend of acetic and formic acid compared with 20 µL of either acid separately or other compounds. The 4-component
mixture design produced a highly significant (P < 0.0001) special cubic model that was improved by backward reduction (Table 4). The square root transformation was applied to the data (number of probes) as per Box–Cox analysis. The model diagnostics were all within acceptable limits. The data appeared normal and displayed constant variance with no outliers, no points that exceeded a Cook’s distance of one, and the predicted versus actual value plots showed close agreement. The R2 statistics (R2, Radj
2 , and Rpred2 ) were clus-
tered with a difference less than 0.12. The lack-of-fit test was not sig-nificant (Prob > F = 0.1274). Based on these diagnostics, the special cubic model was selected and indicated significant factor effects on the probing response; the model was of sufficient quality to navigate the experimental design space and for predicting new observations. The ANOVA indicated significant terms for component effects and interactions for formic × acetic acids (P < 0.0001) and for formic × citral, and for the 3 factor interaction acetic acid × β-ocimene × citral (P < 0.01). The optimization protocol predicted 3 points on the for-mic-acetic axis with desirability coefficients >0.5. All 3 were blends of formic and acetic acids with the highest score at an approximately 2:1 blend followed by a 1:1 blend of the 2 acids (Figure 9). To test the hypothesis that the best blend tested in the 4-component design actually received a larger number of probes than the control, we compared (ANOVA) the number of probes observed on the control (no odorant in yellow SPLAT) (n = 24) with the number on the 1:1 blend (n = 12). There was no effect of block (cage) (P > F = 0.27) or interaction between block and treatment (P > F = 0.86). The mean ± SEM number of probes observed on the 1:1 blend of formic and acetic acids (36 ± 3, n = 12) was significantly greater (P > F = 0.0002)
Figure 5. Electroantennogram responses of antennae of male (A and C) and female (B and D) Diaphorina citri to the content of glass stimulus cartridges initially containing neat citral (A and B) or (Z) and (E)-β-ocimene (C and D) over 9 days expressed as percent of antennal response to a positive control (1 mg formic acid) at the end of each recording. N.S.: no significant difference between mean antennal responses on a given day by ANOVA; means within day followed by the same letter do not differ by Tukey’s HSD following a significant ANOVA (α = 0.0125 corrected for multiple comparisons).
332 Chemical Senses, 2016, Vol. 41, No. 4
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from
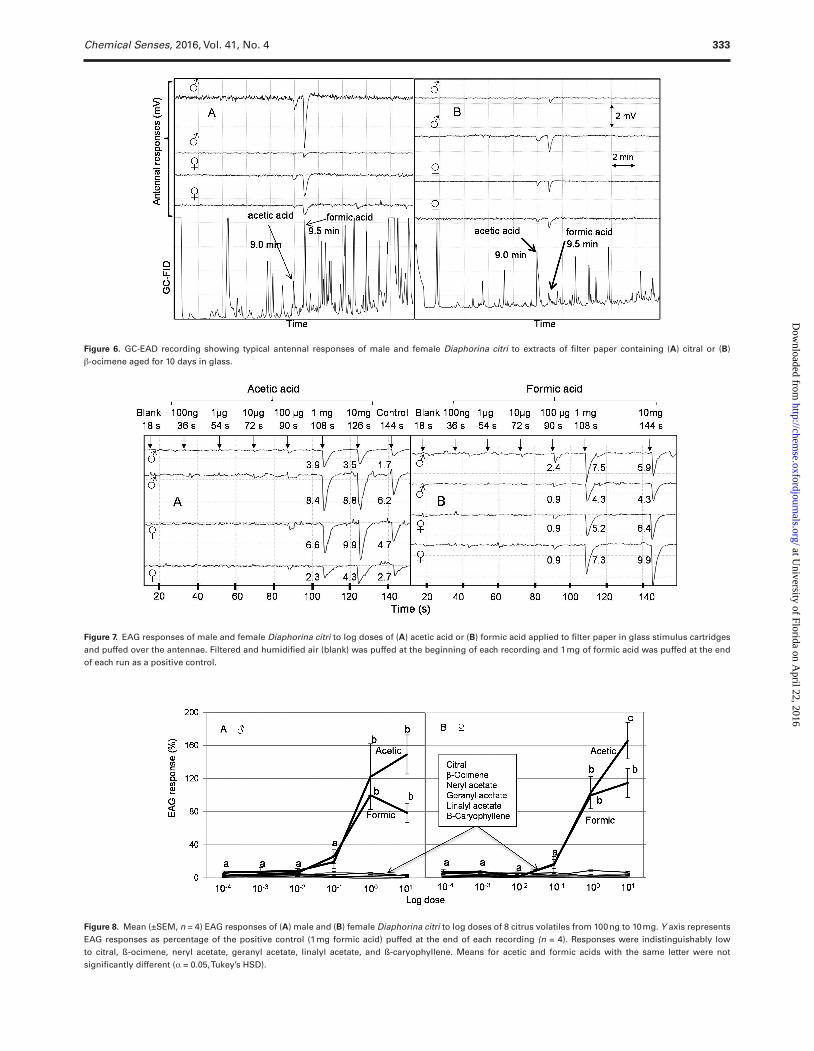
Figure 6. GC-EAD recording showing typical antennal responses of male and female Diaphorina citri to extracts of filter paper containing (A) citral or (B) β-ocimene aged for 10 days in glass.
Figure 7. EAG responses of male and female Diaphorina citri to log doses of (A) acetic acid or (B) formic acid applied to filter paper in glass stimulus cartridges and puffed over the antennae. Filtered and humidified air (blank) was puffed at the beginning of each recording and 1 mg of formic acid was puffed at the end of each run as a positive control.
Figure 8. Mean (±SEM, n = 4) EAG responses of (A) male and (B) female Diaphorina citri to log doses of 8 citrus volatiles from 100 ng to 10 mg. Y axis represents EAG responses as percentage of the positive control (1 mg formic acid) puffed at the end of each recording (n = 4). Responses were indistinguishably low to citral, ß-ocimene, neryl acetate, geranyl acetate, linalyl acetate, and ß-caryophyllene. Means for acetic and formic acids with the same letter were not significantly different (α = 0.05, Tukey’s HSD).
Chemical Senses, 2016, Vol. 41, No. 4 333
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from
than the number of probes observed on the control (19 ± 2, n = 24). The 2-component mixture-amount experiment resulted in a signifi-cant cubic × linear model (P > F = 0.0002). The square root transfor-mation was applied to the count data. The model diagnostics were all within acceptable limits. The data appeared normal with constant variance and no outliers, no points exceeded a Cook’s distance of one, and predicted versus actual value plots showed close agreement. The R2 statistics (R2, Radj
2 , and Rpred2 ) were clustered with a difference
<0.2. The lack-of-fit test was not significant (P > F = 0.128). Based on these diagnostics, a cubic × linear model (Table 5) was selected and indicated significant factor effects; the model was of sufficient quality to navigate the experimental design space and for predicting new observations. The results from this experiment closely validated the prediction resulting from the 4-component mixture experiment. The linear effect of amount was not large (P = 0.02) compared with the other significant effects (Table 5) and more evident for the pure
Table 4. P values, regression coefficients, and response surface model fitting diagnostic statistics for a reduced special cubic model of probing by Diaphorina citri in response to varying proportions 4 odorants
Sum of squares df Mean square F P value Coefficient estimate
Model 136.69 9 15.19 14.57 <0.0001Linear mixture 89.14 3 29.71 28.50 <0.0001A 4.95B 2.78C 2.90D 2.60AB 29.45 1 29.45 28.25 <0.0001 7.35AD 8.07 1 8.07 7.74 0.0062 −4.44BC 0.61 1 0.61 0.58 0.4463 −1.39BD 0.26 1 0.26 0.25 0.6198 −0.91CD 0.16 1 0.16 0.16 0.6943 −0.72BCD 6.93 1 6.93 6.65 0.0110 35.05Lack of fit 99.90 87 1.15 1.39 0.1174
A, formic acid; B, acetic acid; C, β-ocimene; D, citral. R2 = 0.5041; Radj2
= 0.4694; Rpred2
= 0.3933.
Table 5. P values, regression coefficients, and response surface model fitting diagnostic statistics for a cubic × linear model of probing by Diaphorina citri in response to varying proportions and amounts of formic and acetic acids
Sum of squares df Mean square F P value Coefficient Estimate
Model 15.04 4 3.76 7.56 0.0002Linear mixture 3.02 1 3.02 6.09 0.0187A 1 — 3.40B 1 — 3.82AB 1.41 1 1.41 2.83 0.1012 1.69AC 4.00 1 4.00 8.05 0.0075 −0.70AB(A-B) 4.60 1 4.60 9.25 0.0044 −8.62Lack of fit 10.30 16 0.64 1.73 0.1280
A, acetic acid; B, formic acid; C, amount. R2 = 0.4636; Radj2
= 0.4023; Rpred2
= 0.3192.
Figure 9. A planar projection of the 3-dimensional tetrahedral design space created by varying the proportions (%) of 4 components. The response surface is plotted by color and contour lines indicating the number of individual stylet sheaths (probes) by Diaphorina citri on a wax substrate containing 4 odorant compounds: acetic acid, citral, formic acid, and β-ocimene. Dotted arrows indicate decreasing proportion of the corresponding compound. The projection is flattened for printing but can be folded along the 3 black axes to produce the 3-dimensional design space as shown in the cartoon inset on the left. The star indicates the predicted optimal blend for maximum number of probes (1.9:1.0 formic acid:acetic acid).
334 Chemical Senses, 2016, Vol. 41, No. 4
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

acetic acid blend (Figure 10). The optimization algorithm predicted a narrow range of points clustered around a 2:1 blend of formic:acetic acids (Figure 10).
Discussion
Insect vectors and the plant pathogens they transmit limit food production worldwide (Purcell and Almeida 2005). Citrus greening disease, transmitted by D. citri, is a major threat to the economic sustainability of the citrus industry worldwide. Currently, citrus pro-duction in the United States and elsewhere is dependent on intensive insecticide applications to minimize vector density and movement (Tiwari et al. 2011) and nutritional supplementation to ameliorate disease symptoms and extend productive tree life (Schumann et al. 2011). Identifying semiochemicals from host plants and conspecifics for use in traps or other control applications would help to control the vector and the disease, potentially reducing the use of pesticides. Most of the volatile compounds previously identified from head-space of young leaves of susceptible citrus genotypes (Robbins et al. 2012) did not produce a measureable antennal response from male or female D. citri under the conditions of our study by GC-EAD analysis or EAG response to individual neat compounds. This may be related to the relative paucity of chemoreceptors on D. citri antennae (Onagbola et al. 2008) and the possibility that olfaction plays a subordinate or secondary role in mating and host plant ori-entation in this species. Attraction to visual cues and orientation to sex-specific stridulation (Wenninger et al. 2009a; Rohde et al. 2013) may condition or supersede volatile stimuli.
Several citrus volatiles have been suggested as possible attract-ants or repellents for D. citri (Wenninger et al. 2009b; Patt and Sétamou 2010; Mann et al. 2012; Robbins et al. 2012). We tested many citrus volatile odorants by EAG and only a few elicited anten-nal responses (Table 1). During our attempts to study these by EAG and GC-EAD, we discovered that antennal responses to the contents of glass cartridges containing filter paper loaded with small amounts of 2 common citrus compounds, β-ocimene [(Z) and (E)-β-ocimene] and citral (neral and geranial), increased dramatically over time
when the cartridges were stored on a laboratory bench. We also observed an identical effect when filter paper was not used and the compounds were placed into sealed glass cartridges. GC-MS analysis by SPME and extraction of aged filter paper revealed that β-ocimene and citral began to disappear from the cartridges within 3 days (Figure 3). GC-EAD of extracts of aged filter paper allowed us to identify formic and acetic acids as the stimulatory compounds that accumulated in the cartridges, presumably the result of oxidative degradation of β-ocimene and citral. This is the first observation that degradation products of constitutive plant volatiles are stimulatory to insect antennae and the first instance of GC-EAD used to identify chemical compounds stimulatory to the antenna of a psyllid species.
The existence of plant airborne signals has been known for cen-turies and has been well documented for ethylene (Lin et al. 2009), methyl jasmonate, linalool, and β-ocimene (Holopainen and Blande 2012). Monoterpenes such as limonene, α-pinene, β-myrcene, lin-alool, neral, geranial, and β-ocimene (Figure 2) are common citrus volatiles found in the atmosphere surrounding citrus orchards (Park et al. 2013). Some monoterpenes such as isomers of β-ocimene are known to be unstable in air (Fahlbusch et al. 2003). Recent stud-ies (Oikawa and Lerdau 2013; Park et al. 2013) have shown that degradation of VOCs is a common phenomenon and that oxidation of acetaldehyde and formaldehyde can produce acetic and formic acids that are released into the atmosphere. The emission rates of these catabolites (formic or acetic acid) are 200–300% higher than the precursor compounds (ethanol, methanol, acetaldehyde, formal-dehyde, etc.). Park et al. (2013) showed that monoterpenes com-prised approximately 24% of VOC compounds above an orange grove (C. sinensis). They also measured significant amounts of ace-tic acid (8.4%) either from photochemical degradation of parent compounds or direct plant emission. Blande et al. (2014) suggested that emitted plant signals are modified by the environment and that biotic responses are collectively determined by emitted signals and their degradation products.
Understanding the mechanisms involved in insect sensory percep-tion, integration, and behavior is complex and has advanced along a number of paths in recent years. Integration of the knowledge base
Figure 10. Predicted 3-dimensional surface response plot generated by a 2-component mixture-amount design to validate the predicted optimal blend of formic acid:acetic acid to maximize the number of probes on a wax substrate by Diaphorina citri. Circles are data for design points above (red) or below (pink) the predicted value.
Chemical Senses, 2016, Vol. 41, No. 4 335
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

is a challenge, but recent efforts to do so (Renou 2014) emphasize the importance of understanding interactions inherent in mixtures between odorant signals and between odorant receptors (Baker 2009; Su et al. 2012). Our finding suggests that an additional level of complexity may exist in the volatile environment surrounding insect habitats. Although it has been generally accepted that many phy-tophagous insects orient to their host plants based on plant-emitted odors (Bruce et al. 2005; Renou 2014), the peripheral sensory system of D. citri appears to be sensitive to compounds that arise through degradation of common plant volatiles that, by themselves, do not elicit antennal or behavioral responses. As often occurs, serendip-ity contributed to our discovery that common plant volatiles appear to degrade into other distinct small molecules in the environment external to the plant and act as primary physiological and behav-ioral cues for an arthropod vector. This phenomenon may occur more widely than we currently appreciate. The host range of D. citri is restricted to genera of the family Rutaceae. Despite this rather restricted host range, there has been no report of taxonomically spe-cific compound(s) that play a role in the orientation of this species to its host. Antennae of D. citri bear a small number of olfactory sensilla that could play a role in mate or host location (Onagbola et al. 2008). However, evidence has not been found to document orientation to host or conspecific odors from a distance. Coutinho-Abreu et al. (2014b) performed extensive single-cell recording from antennal sensilla of D. citri. They identified a 3-component blend that modestly increased trap catch on sticky yellow cards in the field, but such traps remain highly inefficient. It would appear that sen-sitive olfactory discrimination of suitable host plant tissue from a distance requires a higher degree of olfactory sense sophistication than the insect’s morphology suggests. It is more likely that color is the stronger attractant for flighted D. citri to young, emergent and/or diseased foliage (Mann et al. 2012). Host location by adult D. citri may resemble the “degenerate” mechanism proposed by Kennedy and Stroyan (1959) to explain host plant discovery by species of aphids: random dispersal, visual orientation to yellow wavelengths, and gustatory perception of an arrestant stimulus after landing. This may explain our results wherein formic and acetic acids are highly stimulatory to antennal receptors but do not elicit orientation from a distance. Rather, they may condition a behavioral response (prob-ing) only after alightment on a potential host. This report is the first to identify compounds that elicit antennal responses when presented to the antenna individually by GC-EAD or EAG. The strength and concomitant weakness of these methods is that the odorants are presented to the antennae individually. The perception of a mixture results from the interaction between its individual components giv-ing rise to a neural representation that acquires new characteristics relevant to the odorant mixture (Renou 2014). Diaphorina citri may respond to odorant blends in the context of a particular olfactory background as demonstrated for Manduca sexta (Patt et al. 2013; Riffel et al. 2014). Acetic and formic acids have been shown to be present in the air above citrus orchards (Park et al. 2013). These acids are the only elicitors of antennal response we have been able to consistently demonstrate for D. citri and we hypothesize a role for these compounds in host location or selection. They may play a pri-mary role as attractants or a secondary role as additive or synergistic agents of other, as yet unidentified, odorants. Our inability to dem-onstrate antennal responses to an array of citrus volatiles suggests that acetic and formic acids may act as primary olfactory cues for D. citri and may have practical use for management of this pathogen vector. A great deal more effort will be required to determine how this finding is relevant to movement of the adult psyllids in the field
and how acetic and formic acids interact with other odorants at the level of perception and behavior.
No pheromone has been identified for D. citri. Formic acid, known as an allomone in ants, is also a sex pheromone for 6 ant spe-cies (El-Sayed 2014). The list of semiochemical uses for acetic acid is extensive (El-Sayed 2014). It is designated as an allomone for one spe-cies of Phasmida, one species of Uropygi, 5 species of Coleoptera, one species of Hymenoptera and 10species of Heteroptera. Additionally, it is an attractant for 136 species including Diptera, Heteroptera, Hymenoptera, Lepidoptera, and Neuroptera. It is listed as a sex pheromone in 9 species of Heteroptera, 1 species of Hymenoptera, 4 species of Lepidoptera, and 1 species of Orthoptera (El-Sayed 2014).
Male and female psyllids responded consistently to formic and acetic acids by EAG and GC-EAD. Incorporation of those com-pounds into the feeding assay slow-release matrix allowed us to examine behavioral response to odorant blends. Probing by D. citri of the feeding substrate increased with the addition of a blend of formic and acetic acids compared with the unscented control. We are now pursuing the hypothesis that additional odorant compounds, not stimulatory by themselves, may augment the probing response when combined with formic and acetic acids. The 4-component blend experiment suggested that β-ocimene and citral did not con-tribute to increased probing. The model predicted that an optimal response (greatest number of probes) would occur in response to a 2-component blend along the formic-acetic acid axis even though that particular combination was not included as a design point (Figure 9). Validation of the predicted optimum was obtained by the 2-component mixture-amount experiment (Figure 10) that con-firmed a 2:1 formic:acetic blend as optimal for stimulating the prob-ing response. Current work concentrates on introducing additional odorant compounds into the design space to identify mixtures capa-ble of eliciting greater substrate probing than we have obtained with the 2:1 formic:acetic acid blend. The most evident application of that knowledge could be an attract-and-kill formulation that would greatly increase the specificity of pesticide applications and thereby reduce nontarget effects of insecticides currently used to control D. citri in commercial citrus. These methods could be applied to other phytopathogen vectors for practical management.
We were unable to detect acetic and formic acids in the headspace over healthy potted citrus under the conditions of our study. However, we have not demonstrated that D. citri in nature responds to acetic and formic acids arising as degradation products or simply as natu-rally produced secondary metabolites directly emitted by citrus trees as may occur under field conditions. Nonetheless, the role of degradation products of parent volatile compounds in the orientation of organisms to their hosts may be an area worthy of increased investigation.
Supplementary material
Supplementary material can be found at http://www.chemse.oxford-journals.org/
Funding
Partial funding was provided by the Citrus Research and Development Foundation [853].
AcknowledgmentsWe thank Larry Markle, Jiemin Liu, and Evan Koester (USDA-ARS, Ft. Pierce, FL) for technical assistance and Anna Sara Hill (USDA-ARS, Ft. Pierce, FL) for
336 Chemical Senses, 2016, Vol. 41, No. 4
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

insect rearing. USDA is an equal opportunity provider and employer. Mention of a trademark or proprietary product does not constitute a guarantee or war-ranty of the product by the US Department of Agriculture and does not imply its approval to the exclusion of other products that may also be suitable.
ReferencesAnderson MJ, Whitcomb PJ. 2005. RSM simplified: optimizing processes
using response surface methods for design of experiments. New York: Pro-ductivity Press. p. 292.
Baker TC. 2009. Nearest neural neighbors: moth sex pheromone receptors HR11 and HR13. Chem Senses. 34(6):465–468.
Beyaert I, Hilker M. 2014. Plant odour plumes as mediators of plant-insect interactions. Biol Rev. 89:68–81.
Blande JD, Holopainen JK, Niinemets U. 2014. Plant volatiles in polluted atmospheres: stress responses and signal degradation. Plant Cell Environ. 37(8):1892–1904.
Bove JM. 2006. Huanglongbing: a destructive, newly-emerging, century-old disease of citrus. J Plant Pathol. 88:7–37.
Box GEP, Cox DR. 1964. An analysis of transformations (with discussion). J Roy Stat Soc Series B. 26:211–246.
Bruce TJ, Wadhams LJ, Woodcock CM. 2005. Insect host location: a volatile situation. Trends Plant Sci. 10(6):269–274.
Cen YJ, Ye JM, Xu CB, Feng AW. 2005. The taxis of Diaphorina citri to the vol-atile oils extracted from non-host plants. J S China Agric Univ. 26:41–44.
Cornell JA. 2002. Experiments with Mixtures. 3rd ed. New York: Wiley & Sons.
Cossins EA. 1964. The utilization of carbon-1 compounds from plants. Can J Biochem. 42:1793–1802.
Coutinho-Abreu IV, Forster L, Guda T, Ray A. 2014a. Odorants for surveil-lance and control of the Asian Citrus Psyllid (Diaphorina citri). PLoS One. 9(10):e109236.
Coutinho-Abreu IV, McInally S, Forster L, Luck R, Ray A. 2014b. Odor cod-ing in a disease-transmitting herbivorous insect, the Asian citrus psyllid. Chem Senses. 39(6):539–549.
El-Sayed AM. 2014. Pherobase: database of pheromones and semiochemicals. Available from: http://www.pherobase.com
Fahlbusch KG, Hammerschmidt FJ, Panten J, Pickenhagen W, Schatkowski D, Bauer K, Garbe D, Surburg H. 2003. Flavors and fragrances. Ullmann’s Encyclopedia of industrial chemistry. Weinheim: John Wiley & Sons.
Fall R, Benson AA. 1996. Leaf methanol—the simplest natural product from plants. Trends Plant Sci. 9:296–301.
Gottwald TR, Graca JV, Bassanezi RB. 2007. Citrus huanglongbing: the path-ogen, its epidemiology and impact. Plant Health Prog. 9: 1–36.
Halbert SE. 1998. Entomology section. Tri-ology. 37:6–7.Halbert SE. 2005. The discovery of huanglongbing in Florida. Florida Citrus
Mutual, Orlando, FL, 7–11 Nov. p. 50.Halbert SE, Manjunath KL. 2004. Asian citrus psyllids (Sternorrhyncha: Psyl-
lidae) and greening disease of citrus: a literature review and assessment of risk in Florida. Fla Entomol. 87:330–353.
Hansson BS, Larsson MC, Leal WS. 1999. Green leaf volatile-detecting olfac-tory receptor neurons display very high sensitivity and specificity in a scarab beetle. Physiol Entomol. 24:121–126.
Holopainen JK, Blande JD. 2012. Molecular plant volatile communication. Adv Exp Med Biol. 739:17–31.
Hüve K, Christ MM, Kleist E, Uerlings R, Niinemets U, Walter A, Wildt J. 2007. Simultaneous growth and emission measurements demonstrate an interactive control of methanol release by leaf expansion and stomata. J Exp Bot. 58(7):1783–1793.
Kennedy JS, Stroyan, HLG. 1959. Biology of aphids. Nature. 235:108–109.Kimmerer TW, Kozlowski TT. 1982. Ethylene, ethane, acetaldehyde and etha-
nol production under stress. Plant Physiol. 69:840–847.Kreuzwieser J, Harren FJM, Laarhoven LJJ, Boamfa I, Lintel-Hekkert ST,
Scheerer U, Huglin C, Rennenberg H. 2001. Acetaldehyde emission by the leaves of trees—correlation with physiological and environmental param-eters. Physiol Plantarum. 113:41–49.
Kreuzwieser J, Scheerer U, Rennenbergg H. 1999. Metabolic origin of acet-aldehyde emitted by poplar (Populus tremula × P. alba) trees. J Exp Bot. 50:757–765.
Lapointe SL, Evens TJ, Niedz RP. 2008. Insect diets as mixtures: optimization for a polyphagous weevil. J Insect Physiol. 54(7):1157–1167.
Lin Z, Zhong S, Grierson D. 2009. Recent advances in ethylene research. J Exp Bot. 60(12):3311–3336.
Mann RS, Ali JG, Hermann SL, Tiwari S, Pelz-Stelinski KS, Alborn HT, Ste-linski LL. 2012. Induced release of a plant-defense volatile ‘deceptively’ attracts insect vectors to plants infected with a bacterial pathogen. PLoS Pathog. 8:e1002610. doi:10.1371/journal.ppat.1002610.
Metcalf RL, Metcalf ER. 1992. Plant Kairomones in insect ecology and con-trol. Contemporary topics in entomology 1. New York: Chapman and Hall. p. 168.
Moran VC, Brown RP. 1973. The antennae, host plant chemoreception and probing activity of the citrus psylla, Trioza erytreae (Del Guerico) (Hom-optera: Psyllidae). J Entomol Soc Southern Africa. 36:191–202.
Myers RH, Montgomery DC. 2002. Response surface methodology: process and product optimization using designed experiments. 2nd ed. New York: Wiley & Sons.
Nehlin G, Valterová I, Borg-Karlson AK. 1994. Use of conifer volatiles to reduce injury caused by carrot psyllid, Trioza apicalis, Förster (Homop-tera, Psylloidea). J Chem Ecol. 20(3):771–783.
Nissinen A, Kristoffersen L, Anderbrant O. 2008. Physiological state of female and light intensity affect the host-plant selection of carrrot psyllid, Trioza apicalis (Hemiptera: Triozidae). Euro J Entomol. 105:227–232.
NIST/NIH/EPA (2005) NIST/NIH/EPA. Mass Spectral Library, Standard Ref-erence Database 1, NIST 05. Gaithersburg (MD): Standard ReferenceData Program, National Institute of Standards and Technology.
Oikawa PY, Lerdau MT. 2013. Catabolism of volatile organic compounds influences plant survival. Trends Plant Sci. 18:12.
Onagbola EO, Meyer WL, Boina DR, Stelinski LL. 2008. Morphological char-acterization of the antennal sensilla of the Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), with reference to their probable functions. Micron. 39(8):1184–1191.
Onagbola EO, Rouseff RL, Smoot JM, Stelinski LL. 2011. Guava leaf volatiles and dimethyl disulphide inhibit response of Diaphorina citri Kuwayama to host plant volatiles. J Appl Entomol. 135:404–414.
Park JH, Goldstein AH, Timkovsky J, Fares S, Weber R, Karlik J, Holzinger R. 2013. Active atmosphere-ecosystem exchange of the vast majority of detected volatile organic compounds. Science. 341(6146):643–647.
Patt JM, Meikle WG, Mafra-Neto A, Sétamou M, Mangan R, Yang C, Malik N, Adamczyk JJ. 2011. Multimodal cues drive host-plant assessment in Asian citrus psyllid (Diaphorina citri). Environ Entomol. 40(6):1494–1502.
Patt JM, Sétamou M. 2010. Responses of the Asian citrus psyllid to volatiles emitted by the flushing shoots of its rutaceous host plants. Environ Ento-mol. 39(2):618–624.
Patt JM, Woods D, Dimitratos S, Meikle W, Stockton D, Lapointe SL, Mafra-Neto A. 2013. Novel synthetic ligands enhance the behavioral responses of Asian citrus psyllid to naturally occurring host-plant volatiles. In: Beck J, et al., editors. Pest management with natural products. Washington (DC): ACS Symposium Series. p.111–124, Chap. 8.
Poerwanto ME, Trisyono YA, Beattie GAC, Subandiyah S, Martono E, Holford P. 2012. Olfactory responses of the Asiatic citrus psyllid (Diaphorina citri) to mineral oil-treated mandarin leaves. Amer J Agric Biol Sci. 7(1):50–55.
Purcell HA, Almeida RPP. 2005. Insects as vectors of disease agents. In: Good-man RM, editors. Encyclopedia of plant and crop science. London (UK): Taylor & Francis Group. p. 1–5.
Raguso RA, Light DM. 1998. Electroantennogram responses of male Sphinx perelegans hawkmoths to floral and green-leaf volatiles. Entomol Exp Appl. 86:287–293.
Renou M. 2014. Pheromones and general odor perception in insects. In: Mucignat-Caretta C, editors. Neurobiology of chemical communication. Boca Raton: CRC Press.
Riffel JA, Shilzerman E, Sanders E, Abrell L, Medina B, Hinterwirth AJ, Kutz JN. 2014. Flower discrimination by pollinators in a dynamic chemical environment. Science. 344:1515–1518.
Chemical Senses, 2016, Vol. 41, No. 4 337
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from

Robbins PS, Alessandro RT, Stelinski LL, Lapointe SL. 2012. Volatile profiles of young leaves of Rutaceae spp. varying in susceptibility to the Asian citrus psyllid (Hemiptera: Psyllidae). Fla Entomol. 95:774–776.
Rohde B, Paris TM, Heatherington EM, Hall DG, Mankin RW. 2013. Responses of Diaphorina citri (Hemiptera: Psyllidae) to conspecific vibrational signals and synthetic mimics. Ann Entomol Soc Am. 106:392–393.
Rouseff RL, Onagbola EO, Smoot JM, Stelinski LL. 2008. Sulfur volatiles in guava (Psidium guajava L.) leaves: possible defense mechanism. J Agric Food Chem. 56(19):8905–8910.
Schumann A, Spann T, Rouse R. 2011. Maintaining productivity in the pres-ence of HLB. Citrus Industry, Nov: 6–9. Ocala (FL): AgNet Media, Inc.
Soroker V, Talebaev S, Hararai AR, Wesley SD. 2004. The role of chemical cues in host and mate location in the pear psylla Cacopsylla bidens (Homop-tera: Psyllidae). J Insect Behav. 17:613–626.
Su CY, Menuz K, Reisert J, Carlson JR. 2012. Non-synaptic inhibition between grouped neurons in an olfactory circuit. Nature. 492(7427):66–71.
Sule H, Rita M, Omar D, Kah-Wei AH. 2012. Response of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) to volatiles emitted from leaves of two rutaceous plants. J Agric Sci. 4:152–159.
Tiwari S, Mann RS, Rogers ME, Stelinski LL. 2011. Insecticide resistance in field populations of Asian citrus psyllid in Florida. Pest Management Sci. 67:1258.
Tol VR, Visser JH. 2002. Olfactory antennal responses of the vine weevil Otio-rhynchus sulcatus to plant volatiles. Entomol Exp Appl. 102:49–64.
Visser JH, Yan FS. 1995. Electroantennogram responses of the grain aphids Sitobion avenae (F) and Metopolophium dirhodum (Walk) (Hom, Aphidi-dae) to plant odor components. J Appl Entomol. 119:539–542.
Weisberg S. 1985. Applied linear regression. 2nd ed. New York: Wiley & Sons.Wenninger EJ, Hall DG, Mankin RW. 2009a. Vibrational communication
between the sexes in Diaphorina citri (Hemiptera: Psyllidae). Ann Ento-mol Soc Am. 102:547–555.
Wenninger EJ, Stelinski LL, Hall DG. 2009b. Roles of olfactory cues, visual cues, and mating status in orientation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) to four different host plants. Environ Entomol. 38(1):225–234.
338 Chemical Senses, 2016, Vol. 41, No. 4
at University of Florida on A
pril 22, 2016http://chem
se.oxfordjournals.org/D
ownloaded from