fracture toughness of horns and a reinterpretation of the horning behaviour of bovids
TRANSCRIPT

J. Zool., Lond. (1987) 213,621-639
Fracture toughness of horns and a reinterpretation of the horning behaviour of bovids
ANDREW KITCHENER Biomechanics Group, Department of Pure and Applied Zoology, University of Reading,
Whiteknights, PO Box 228, Reading, Berks RG6 2AJ, UK
(Accepted 9 April 1987)
(With 2 plates and 1 1 figures in the text)
The keratinous horns of bovids are used in intraspecific combat to gain access to females in oestrus. Horn sheath keratin is a composite material consisting of stiffprotein fibres and a pliant protein matrix. Unlike antlers, horns are permanent structures which are likely to accumulate damage during fighting. Therefore, horn sheath keratin should be resistant to fracture (tough) and insensitive to surface defects (scratches and cracks) which may weaken horns by acting as stress concentrators.
The effect of water on the toughness and notch-sensitivity of horn sheath keratin was investigated in three-point bending and tensile tests. Several measures of toughness were made on dry (0% water content), fresh (20%) and wet (40%)) horn keratin, including total work of fracture, Gurney & Hunt work of fracture, critical strain energy release rate and critical stress intensity factor.
The mean total work of fracture offresh horn is about 40 kJ/m2 which is relatively much greater than most biological and synthetic materials. Most of the work of fracture is due to plastic yielding of the matrix (5&75%); the rest is due to crack-tip specific fracture mechanisms such as fibre pull-out and Cook Gordon crack-stopping. Dehydration reduces the total work of fracture of horn keratin by preventing the yielding of the matrix.
The strength of fresh and wet horn is insensitive to notches, but dry horn is very notch-sensitive. Therefore, bovids must avoid dehydration of their horns due to the desiccating effect of the environment. The ‘homing’ behaviour of bovids may be a maintenance activity which ensures that the horn sheath is adequately hydrated to remain tough and notch-insensitive,
Contents
Introduction . . . . . . . . . . . . . . . . Materials and methods . . . . . . . . . . . .
Total work of fracture . . . . . . . . . . . . Notch-sensitivity . . . . . . . . . . . . . . Gurney & Hunt area sector method . . . . . . Critical strain energy release rate Critical stress intensity factor, Kc . . . . . .
Results and discussion . . . . . . . . . . . . Total work of fracture . . . . . . . . . . . . Notch-sensitivity . . . . . . . . . . . . . . Gurney and Hunt area sector method . . . . Critical strain energy release rate and LEFM . . Critical stress intensity factors . . . . . . . . Homing behaviour and the toughness of horns
Summary . . . . . . . . . . . . . . . . . . References . . . . . . . . . . . . . . . . . .
. . . . . .
62 I 0022-5460/87/012621+ 19 $0340
Page . . . . . . . . . . . . . . . . 622 . . . . . . . . . . . . . . . . 623 . . . . . . . . . . . . . . . . 623 . . . . . . . . . . . . . . . . 624 . . . . . . . . . . . . . . . . 625 . . . . . . . . . . . . . . . . 628 . . . . . . . . . . . . . . . . 629 . . . . . . . . . . . . . . . . 630 . . . . . . . . . . . . . . . . 630 . . . . . . . . . . . . . . . . 634 . . . . . . . . . . . . . . . . 635 . . . . . . . . . . . . . . . . 636 . . . . . . . . . . . . . . . . 636 . . . . . . . . . . . . . . . . 637 . . . . . . . . . . . . . . . . 638 . . . . . . . . . . . . . . . . 638
0 1987 The Zoological Society of London

622 A. KITCHENER
Introduction
Male bovids use their horns to fight each other for priority access to females in oestrus (Leuthold, 1977). Unlike antlers, horns are permanent structures so that any damage they may incur due to fighting is accumulated throughout life. The breakage of a horn in fighting is very serious. A new horn cannot be grown so that it would be unlikely that its owner could compete successfully for females in subsequent years, even if it survived. Consequently, reproductive success would be affected severely. Deer cast and regrow their antlers every year so if their antlers break, reproductive success is affected in only one year (Lincoln, 1972; Clutton-Brock, Guinness & Albon, 1982). A bovid may well die from the loss of a horn or be weakened to succumb to predation or secondary infections. Therefore, horns should be made of a material which can withstand the potential weakening effects of accumulated surface defects such as scratches and cracks.
The effects of such defects have been much studied in simpler materials. For instance, the strength of glass is sensitive to surface scratches and cracks (Gordon, 1976). Stress (force per unit area) is concentrated at these defects when a structure is loaded. If the concentration of stress exceeds the local strength of the material, cracks will propagate catastrophically from these defects unless there are mechanisms for preventing this from happening. Consequently, structures appear to fail at much lower loads than they are theoretically designed to withstand. Materials can avoid the propagation of cracks from defects either by mechanisms occurring at the crack tip or by yielding to relieve the stress concentration. Score lines put into glass act as large stress concentrators and weaken the glass locally so that it can be easily and predictably broken. This is the basis of the glazing industry. Glass has no mechanisms for relieving stress concentrations or preventing propagation of cracks and is described as a brittle material. Materials which have mechanisms for relieving stress concentrations due to surface (and other) defects are described as tough materials.
Horns are made up of a sheath of a-keratin and a core of spongy bone. The keratin sheath is the main load-bearing material of the horn and because it is external it is likely to incur scratches and cracks from fighting. Consequently, horn keratin should be a tough material which is insensitive to the presence of surface defects.
Alpha-keratin is a composite material comprising stiff, strong fibres made of protein set in a pliant, weak matrix also made of protein (Fraser & MacRae, 1980). The toughness of, or the ability of fibrous composites to prevent crack propagation depends on its components having different mechanical properties. Any factor which alters the mechanical properties of the components could affect the toughness of horn sheath keratin.
Most environments are desiccating whether they are hot or cold. The external surface of horns would be particularly susceptible to dehydration. As the water content of a-keratin is reduced, the stiffness of the matrix approaches that of the fibres so that the toughening mechanisms of horn sheath could be affected (Fraser & MacRae, 1980). Materials often become more brittle due to dehydration.
Unfortunately, fracture mechanics is only able to investigate the elastic or reversible toughening mechanisms at the tip of a propagating crack. At present, there is no theoretical basis for the investigation of the yielding or plastic component of toughness, which may be very important in the fracture of biological materials.
There are two different approaches to investigating the elastic or reversible component of the toughness of materials. These approaches have been united in Linear Elastic Fracture Mechanics

FRACTURE IN HORNS 623
(LEFM). One approach is to investigate the energy change in a body as a crack develops to derive a parameter, Gc, the critical strain energy release rate, which is independent of the geometry of the crack. The other approach is to consider the stress field in the vicinity of the tip of a crack which is characterized by a parameter, K, the stress intensity factor. Fracture occurs when the stress intensity factor reaches a critical value, Kc. However, these two approaches are interrelated only if the material is elastic linearly up to fracture, undergoes small strains, is isotropic and shows negligible plastic flow at the crack tip (Piggott, 1980; Williams, 1984; Atkins & Mai, 1985). The problems of non-linear elasticity, viscoelasticity, ductility, high strains and anisotropy have hardly been dealt with, which is unfortunate since biological materials usually demonstrate all of these problems. In spite of this, LEFM has been applied successfully to anisotropic, high strain biological materials such as grass, kelp fronds and wood (Williams & Birch, 1976; Vincent 1982; Vincent & Gravell, 1986).
The effect of water on the toughness and notch sensitivity of horn sheath keratin has been investigated using the horns of gemsbok (Oryx g. gazella), mouflon (Ouis rnusimon) and waterbuck (Kobus ellipsipryrnnus) to see if bovids are likely to encounter any problems due to dehydration of their horns in desiccating environments. Linear Elastic Fracture Mechanics was applied to the results in order to assess the relative importance of elastic and plastic fracture mechanisms in the total work of fracture of horn keratin.
Materials and methods
Horns were obtained post-mortem and stored in a domestic freezer until required. Longitudinal strips of horn sheath were cut from the horns. The strips were made roughly square using either rotating silicon carbide discs irrigated with water, or a belt-driven sander. The strips were milled square using an edge or fly cutter until they were devoid of any potential stress-concentrating defects. The effect of water on the notch-sensitivity and fracture toughness of horn sheath was investigated using gemsbok horns. Total work of fracture of the horn sheath was investigated using waterbuck and mouflon horns.
Total work of fracture
The total work of fracture includes all energy used in creating a fracture, including plastic flow which may occur far from the site of the fracture.
Test-pieces were prepared of approximate dimensions 80 mm x 15 mm x 10 mm. The mouflon horn was dry museum material and no special storage conditions were required for it. The waterbuck horn was tested immediately after the test-pieces were prepared so that they did not dehydrate. Horn tested in this way is described as fresh horn and has a water content of about 20% (Kitchener, 1985).
A Tattersall & Tappin notch (TTN) was put into the centre of each test-piece using a hacksaw (Fig. 1) (Tattersall & Tappin, 1966). The notches were sharpened using a razor blade to increase the concentration of stress. The TTN notch conformation was developed specifically for bending tests on brittle materials, but has been found to be suitable for ductile materials such as horn because a crack is driven through the horn keratin before too much plastic flow prevents any crack propagation at all. The notch ensures that a crack is driven through the material in a controlled manner at a particular point far from the end effects of loading (Tattersall & Tappin, 1966).

624 A. KITCHENER
FIG. 1. The conformation of the Tattersall & Tappin notch (TTN) for the determination of total work of fracture. The stippled area ll! indicates the material which is fractured during a test.
All tests were carried out in 3-point bending in an Instron 4202 testing machine using a bending span of 35 mm and a loading speed of 2 mm/min. Although the depth to span ratio was quite low in bending tests, fracture was clearly due to bending rather than shear failure. At greater bending spans, test-pieces continued to bend and yield without crack propagation. The slow speed of loading ensures controlled crack propagation so that all the energy fed into the test-pieces is used in creating fracture surfaces. The Instron was connected to a chart recorder which gave an output of force against deflection.
Work of fracture was measured for test-pieces from along the length of the horns by measuring the area under the force deflection curve and dividing it by the cross-sectional area.
Notch-sensitivity
Test-pieces of approximate dimensions 150 mm x 15 mm x 2 mm were prepared as described above from the horns of gemsbok. Three different water contents of horn keratin were used covering the range from fully hydrated to fully dehydrated. Wet or saturated horn was prepared by soaking in distilled water for at least 3 days. Measurements of the dimensions of the test-pieces twice daily showed that they had swollen fully after this time (Fig. 2). Wet horn has a water content of approximately 40% (Kitchener, 1985). Dry or dehydrated horn was prepared by heating test- pieces in an oven at 1 10 "C for 24 hours. Dry horn has a water content of 0%. Fresh or cut horn was defined as test-pieces which were tested immediately after being cut from the horn. Consequently, no special storage conditions were required for them and they had a water content of approximately 20% (Kitchener, 1985).
Tabs of aluminium of approximate dimensions 40 mm x 15 mm x 2 mm were folded in half and used to sandwich the ends of the test-pieces (Fig. 3). They were glued in place using Araldite epoxy resin adhesive. These tabs ensured that the jaws of the testing machine did not damage the ends of the test-pieces and fed the load evenly into them.

FRACTURE IN HORNS 625
0 1 2 3 4 5 6 Days
F I G . 2. The swelling of test-pieces of horn sheath immersed in water. Swelling is complete after three days.
Notches were put into both edges at the centre of each test-piece using an engraving cutter which ensured that the radius of the notches was constant for different crack lengths. Relative notch depths ranged from 0 to 0.7 for each different water content of horn sheath.
Test-pieces were tested in tension in the Instron 4202 testing machine and were loaded at 2 mm/ min at room temperature until complete failure. Tests were rejected if failure occurred at the aluminium tabs or did not occur at the central notches.
The tensile strength (a') of each test-piece was calculated as follows:
a'= Fmax/(b*d) (1)
where: Fmax is peak load (N), b is breadth of test-piece (m), d is depth of test-piece (m) and b > d. Tensile strength was plotted against the relative notch length of each test-piece. In this case,
relative notch length is the sum of the lengths of the two notches divided by the breadth (b) of the test-piece.
Gurney & Hunt area sector method
One of the best ways to measure the elastic work of fracture of a non-linear elastic material is using the Gurney & Hunt area sector method (Gurney & Hunt, 1967). This method involves the

626 A. KITCHENER
FIG. 3. The double edge notch tensile test-piece. Aluminium tabs are glued to each end to minimize damage to the test- piece in the jaws of the Instron. The arrows (+) indicate the tensile mode of loading.
Deflection (mm)
FIG. 4. Calculation of Gurney & Hunt work of fracture and critical strain energy release rate, Gc. (a) The two force deflection curves are the results for two different notch lengths. The stippled area between the curves is the work needed to extend the crack from one notch length to the other. (b) Lines are drawn parallel to the initial loading curve to intersect with the point at which yielding ends in order to exclude work of yielding. (c) These two lines are superimposed and the stippled area (ABD) is the work needed to extend the crack from one notch length to the other. Alternatively, the areas ABC and ADE represent the work to failure of the test-pieces. The critical strain energy release rate is calculated using an energy calibration factor.

FRACTURE IN HORNS 627
PLATE 1. Electron micrograph of the fracture surface of the horn sheath of the mouflon showing the rough and fibrous surface with a distinct layering and delaminations ( x 100).
repeated loading and unloading of the material as the crack propagates. The work of fracture is calculated as the energy of the loading/unloading curve divided by the area of material fractured in extending the length of the crack. Unfortunately, it was not possible to follow the path of the crack through horn sheath material. This method relies also on the fracture being reversible or elastic (i.e. there is no ductile failure), but horn sheath shows considerable ductile failure. Therefore, a modified Gurney & Hunt method was used to measure directly work of fracture of horn.
The force deflection curves of test-pieces of similar dimensions, but different relative notch lengths, were superimposed. The area between these two curves is the work needed to extend the smaller to the greater notch length (Fig. 4a). Work of fracture (R) was calculated as follows:
(2) R = W/(d . (a2 - u 1))
where: W is the difference in work between two different notch lengths (J), d is depth of test-piece (m), a1 is the smaller notch depth (m), a2 is the larger notch depth (m).
The problem with this method is that because of the variability of the force deflection curves few of them could be superimposed. Also, this method may include some plastic work because the test- pieces yield just before failure. Therefore, a linear curve was drawn parallel to the initial slope of the experimental force deflection curve up to the failure load so that work of yielding could be eliminated (Fig. 4b). These linear force deflection curves for different crack lengths were
628 A. KITCHENER
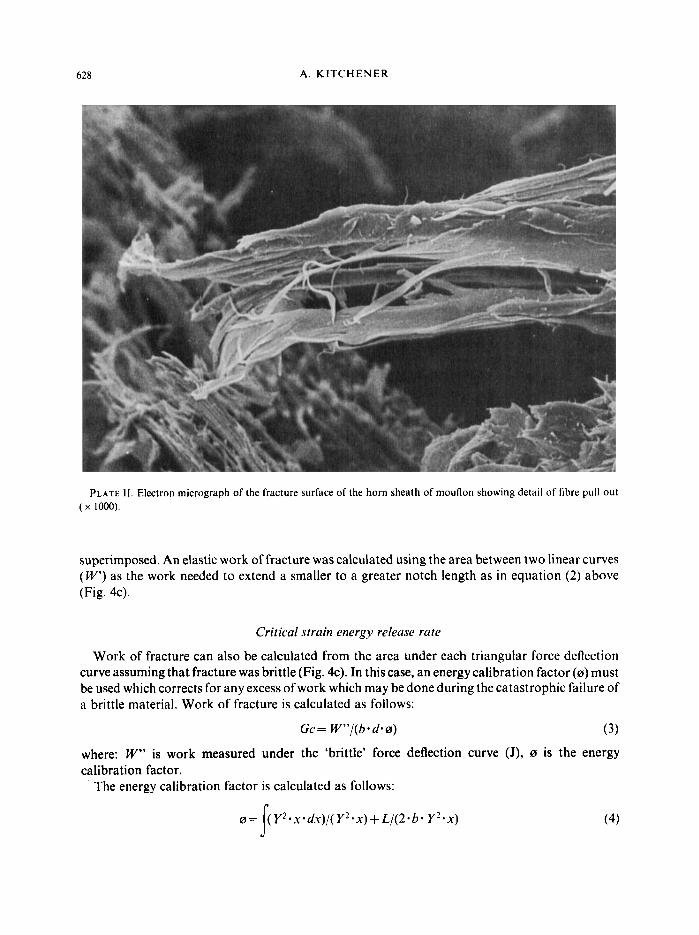
PLATE 11. Electron micrograph of the fracture surface of the horn sheath of mouflon showing detail of fibre pull out ( x 1000).
superimposed. An elastic work of fracture was calculated using the area between two linear curves ( W ’ ) as the work needed to extend a smaller to a greater notch length as in equation (2) above (Fig. 4c).
Critical strain energy release rate
Work of fracture can also be calculated from the area under each triangular force deflection curve assuming that fracture was brittle (Fig. 4c). In this case, an energy calibration factor (0) must be used which corrects for any excess of work which may be done during the catastrophic failure of a brittle material. Work of fracture is calculated as follows:
Gc = W ” / ( b . d.0 ) (3)
where: W” is work measured under the ‘brittle’ force deflection curve (J), 0 is the energy calibration factor.
The energy calibration factor is calculated as follows:
0 = ( Y 2 * x . d x ) / ( Y 2 - x ) + L / ( 2 . b * Y 2 . x ) (4) s

F R A C T U R E IN HORNS 629
. . . . . . . . . , : . . . . I . . . .
. I . .
. . ,
' . . I . . . .
. . . . . . . . . . . . . . . . . . . . (a) . . . . . . . . . . . . . . . . - .
. . . . . . . . . : n
. . . . . . . . . . I
. . . . , . . . . . . , , . , . . . , '
. .
(b) . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . , . . * . I f '
FIG. 5. The Cook-Gordon crack-stopping mechanism in a fibre composite. (a) The crack starts to propagate across the fibres. (b) The stress ahead of the crack tip opens up the weak interface between fibre and matrix. (c) The crack is blunted when it reaches the opened-up interface.
where: x is relative notch depth, L is length of test-piece (m), h is breadth of test-piece (m), Y is the geometry factor.
The geometry factor is calculated as follows (Williams, 1984):
Y = 1.98 + 0 . 3 6 ~ - 2.1 2x2 + 3 . 4 2 ~ ~ ( 5 )
It must be noted that the results calculated in the methods outlined above are supposed to represent the elastic fracture mechanisms occurring only at the crack tip. They assume that horn sheath is linearly elastic to failure and isotropic. The condition of linear elasticity has been enforced to try to see the relative importance of crack tip specific fracture mechanisms to the total work of fracture.
Critical stress intensity factor, Kc
The critical stress intensity factor is calculated from the above data as follows (Williams, 1984):
KC = Y.af.al (6)

630 A. KITCHENER
aa
6C m- E 3 p!
Y - 3 0 c
E 4c c 0
6 - m c :
2(
C I I I I I I 100 200 300 400 500 600
Length from tip (mm)
FIG. 6. The work of fracture of the horn sheath along the length of the horn of a waterbuck. The standard errors of the means for each horn section are indicated by the bars.
where: a is half notch length (m), b' is fracture strength (Pa), Y is the geometry factor (see above). Horn sheath keratin should also be both an isotropic and a linearly elastic material to failure in
order to calculate Kc, particularly if it is to be related to Gc by the following: Kc= E . Gc where E is the stiffness (or Young's modulus) of the material. Therefore, measurements of Kc should be treated with great caution. However, it must be remembered that LEFM has been apparently successfully applied to biological materials (Williams & Birch, 1976; Vincent, 1982; Vincent & Gravell, 1986).
Results and discussion
Total work of fracture
The horn keratin yielded considerably during the three-point bending tests. The fracture surfaces showed delamination of the layers of keratinocytes which make up horn sheath (Plate I) and considerable fibre pull out (Plate 11). Both of these mechanisms contributed to the high total work of fracture of horn sheath keratin.
Sometimes a notch would be deflected along the length of a test-piece and perpendicular to its original direction of propagation. This indicates an effective Cook-Gordon crack-stopping mechanism (Fig. 5) (Gordon, 1976). There is a component of stress ahead of a crack tip which is

60
N-
E 4 Y
5 40-
2
z
0,
V c
c c 0
g 2 20- - I-
0
63 I
-
I I I I I I 0 100 200 300 400 500 600
F I G . 7. The work of fracture of the horn sheath along the length of the horn of a mouflon. The standard errors of the means for each horn section are indicated by the bars.
150
100 -5- a z 5
p! 3
Is, C
50
0
Relative notch length
F I G . 8. The notch-sensitivity of fresh horn (0) and wet horn (0) in tension. T h e y on x regression lines are shown for these two sets of data.
632
800
600
A. KITCHENER
-
-
/
I 3
Deflection (mm)
F I G . 9. Force deflection curves for fresh and dry horn in tension.
150
100
z z 5 m 2 z t
50
0
' 8 0
0 I
'0 \
0 E3-T
I I I I I I I I I 0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8
Relative notch length
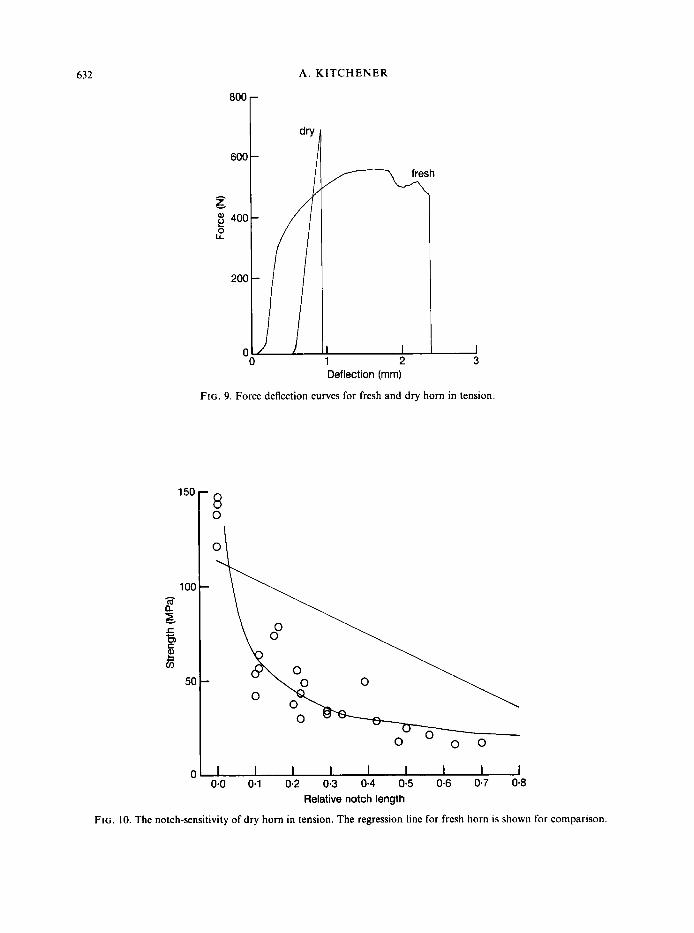
FIG. 10. The notch-sensitivity of dry horn in tension. The regression line for fresh horn is shown for comparison.

FRACTURE IN HORNS 633
1 0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8
Relative notch length
FIG. 1 I . The notch-sensitivity of fresh horn tested in impact. The regression line for fresh horn is shown for comparison.
TABLE I A summary of the mean works of fracture (kJ/m2) of horn sheath
keratin derived from different methods
Dry Fresh Wet
Total work of fracture - 40.0 -
Gurney & Hunt +plastic work - 19.8 13.6 Gurney & Hunt -plastic work - 11.2 Critical strain energy release rate 20.2 8.4
-
-

634 A. KITCHENER
perpendicular to the direction of’the propagating crack so that any weak interfaces in the material will open up ahead of the crack and spread perpendicular to it. Therefore, the crack will be blunted when it reaches the opened-up interface. As a result, the crack must take a very tortuous path through the material if it is to propagate through the material and this absorbs much energy to produce a very high work of fracture. This mechanism is characteristic of composite materials where there is a disparity in mechanical properties between the two components. The Cook- Gordon crack-stopping mechanism could occur either between the fibre and matrix of the keratin or between the layers of keratinocytes. Horn sheath keratin appears to have a variety of toughening mechanisms.
Work of fracture varies along the length of the horns. The total work of fracture of waterbuck horn keratin varies from a mean of about 10 kJ/m2 at the base of the horn to a mean of more than 80 kJ/m2 at the tip of the horn (Fig. 6). The total work of fracture of mouflon horn keratin varies from a mean of about 12 kJ/m2 at the base to a mean of more than 60 kJ/m2 at the tip (Fig. 7).
This variation in work of fracture seems to be due to the presence of annual and minor growth lines which are rich in calcium and which run between layers of keratinocytes along the length of the test-pieces (Kitchener, 1985). There are more growth lines at the base of the horn because horns grow more slowly as they grow older (Schaller, 1977; Nievergelt, 1981; Goss, 1983). These growth lines appear to act as planes of weakness and preferential shear which could reduce bending strength and work of fracture. Calcium is found also bound to phospholipids within keratinocytes (Spearman, 1977). The slower rate of growth at the base of the horn results in more calcium being incorporated into the keratinocytes (Kitchener, 1985) where it may act as filler particles to reduce the yielding of the matrix and which may reduce the strain to failure (Mullins, 1980). This may reduce the total work of fracture at the base of the horns compared to the tip.
Total work of fracture is dependent on the geometry and size of the test-pieces, so that comparisons can only be made between tests carried out under similar conditions. This is because it is not possible to predict the plastic component of the work of fracture using fracture mechanics. Therefore, total work of fracture is not a material property. However, it should reflect the total work of fracture of the horn sheath of the living animal because the thickness of the test-pieces is very close to the thickness of the horn sheath.
The work of fracture of horn keratin is much greater than most biological and synthetic materials due to its yielding. Taking into account specific gravity of different materials, the specific or relative work of fracture of horn keratin (32 kJ/m2) is greater than that of antler (6.6 kJ/m*), bovine femur (1.6 kJ/m2), glass (5 J/m2) and mild steel (25.6+kJ/m2) (Gordon, 1978; Currey, 1979). Only wood (40 kJ/m2) and some synthetic composite materials such as glass fibre epoxy resin (volume fraction of fibres = 0.67) have a higher specific work of fracture (96 kJ/m2) (Gordon, 1978; Hull, 1981).
The work of fracture of horn is very high, as was expected for a permanent structure used repeatedly in fighting. The magnitude of the work of fracture seems to be dependent on the ability of the matrix to yield and a variety of toughening mechanisms at the crack tip.
Notch-sensitivity
Fresh and wet gemsbok horn keratin tested in tension show considerable yielding, which is reversible for wet horn but irreversible for fresh horn. The plasticizing action of water probably allows the large protein chains of the matrix greater mobility in wet horn so that they can recover their former positions after the load is removed. The lower hydration of fresh horn makes this

FRACTURE IN HORNS 635
impossible so that yielding is mostly plastic, although there is some viscoelastic recovery. The notches in the tensile tests were often blunted by the yielding or they were deflected along the test- pieces by the Cook-Gordon crack-stopping mechanism. The mean tensile strength of wet horn (56 MPa, n = 13, S.E. = 4.0) is lower than that for fresh horn (122 MPa, n = 38, S.E. = 2.9) because the shear modulus of the matrix of wet horn is lower due to hydration so that stress is transferred less efficiently to the short fibres (Kitchener & Vincent, 1987).
The tensile strengths of fresh and wet horn are unaffected by the presence of notches, i.e. both are notch-insensitive materials (Fig. 8). For example, a relative notch length of 0.5 reduces the tensile strength of the test-piece to half of the tensile strength of the material.
Dry horn shows no yielding before failure (Fig. 9). It cannot relieve stress concentrations caused by notches so that it is a notch-sensitive material (Fig. 10). A relative notch length of 0.5 reduces the tensile strength of the test-piece to less than half of the tensile strength of the material. The matrix can no longer yield in dry horn because dehydration increases the strength and stiffness of the matrix so that it has similar mechanical properties to the fibres (Fraser & MacRae, 1980). Consequently, toughening mechanisms, such as the Cook-Gordon crack-stopping mechanism, and yielding can no longer occur and total work of fracture is reduced. Dry horn acts like an isotropic, homogeneous material which shows brittle failure. The fracture surface shows no sign of fibre pull out or delamination of keratinocytes.
At high rates of loading, the stiffness of the matrix approaches that of the fibres due to its viscoelastic behaviour (Dorrington, 1980). Test-pieces of gemsbok horn were tested on impact at 3 m/s which revealed that they showed some notch-sensitivity (Fig. 11) (Kitchener, 1985). An analysis of films of fighting bovids shows that bovids clash typically at about 3 m/s (Kitchener, 1985). The degree of notch-sensitivity is slight when compared with dry horn so that the toughness of horn keratin does not seem to be affected by clashing at high speeds.
This experiment shows that i t is important for bovids to keep their horns hydrated adequately to reduce notch-sensitivity and to maintain a high work of fracture by yielding. The mechanism by which bovids regulate the water content of their horns will be discussed below.
Gurney and Hunt area sector method
The force deflection curves for similar-sized test-pieces in tensile tests were very variable so that few of them could be superimposed to work out a Gurney & Hunt work of fracture for gemsbok horn. For example, it was possible only to obtain one result for wet horn and none for dry horn. A conventional Gurney & Hunt method was not possible because it was not possible to follow the path of a crack through the material.
The mean Gurney & Hunt work of fracture, including some plastic work, was 19.8 kJ/m2 (n = 9, S.E. = 2.0) for fresh horn. By partitioning off the plastic work, the true or elastic work of fracture was reduced to I 1.2 kJ/m2 ( n = 6, S.E. = 1.4). A number of fracture mechanisms at the crack tip are responsible for this work of fracture, including fibre debonding and pull out, delamination of layers of keratinocytes and Cook-Gordon crack-stopping (Gordon, 1976; Harris, 1980). All of these mechanisms absorb energy by making the actual fracture surface area much greater than the nominal surface area.
The single Gurney & Hunt work of fracture of wet horn including plastic work was 13.6 kJ/m2, which is slightly lower than that for fresh horn.

636 A. KITCHENER
Critical strain energy release rate and LEFM
The work of fracture of fresh horn derived from the work to failure and an energy calibration factor assuming the conditions for LEFM was 8.4 kJ/m2 (n = 12, S.E. = 0.8) which is very similar to the Gurney & Hunt work of fracture excluding plastic work ( 1 1 *2 kJ/m2). However, the same work of fracture for dry horn was 20.2 kJ/m2 (n = 12, S.E. = 4), which is nearly twice as much as for fresh horn. In view of the small sample sizes, it is difficult to decide whether this difference is real or apparent.
Therefore, both the Gurney & Hunt work of fracture and the critical strain energy release rate indicate the proportion of the total work of fracture which is due to elastic fracture mechanisms at the crack tip. The remainder is due to ductility of the matrix. Only 25-50% of the total work of fracture of fresh horn can be ascribed to fracture mechanisms at the crack tip, which indicates the importance of yielding to the living animal. If the material yields, fracture can be avoided and cracks blunted so that it would be possible to renew or repair material before the next fight. These results are summarized in Table I.
Critical stress intensity factors
The mean critical stress intensity factor, Kc, was 4.5 MPa/mf (n = 36, S.E. =0.98). There was no dependence of Kc on depth or breadth of test-piece. Dry horn (2.2 MPa/mi, n = 21, S.E. = 0.72) and wet horn (2.6 MPa/mi, n= 11, S.E.=0.75) had both lower stress intensity factors, which is consistent with the lower work of fracture expected for a brittle (dry) material, and a weak (wet) material, respectively.
However, it must be remembered that, in the case of fresh and wet horn particularly, the parameter calculated here is not a true stress intensity factor (Kc) because the conditions for LEFM have not been adhered to. Fresh and wet horn are not isotropic and are not elastic to failure.
LEFM has been applied apparently successfully to other biological materials which do not strictly follow its conditions, including wood (Williams & Birch, 1976), grass (Vincent, 1982) and kelp fronds (Vincent & Gravell, 1986). Therefore, it would be interesting to see if this approach could be applied to fresh horn.
The stress intensity factor of an anisotropic material can be converted to a work of fracture using the following equation (Williams & Birch, 1976):
R (1/2 * El * Et); [(Et/El)i + ( ( -2 - G - v * Et + Et * El)/(2 - El - G))]:
Kc2 =
where: R is work of fracture (J/m2), El is longitudinal stiffness (4.5 GPa), Et is transverse stiffness (4.5, 2.25 GPa), G is shear modulus (1 GPa), u is Poisson ratio (0.3).
Makinson (1954) showed that the longitudinal and transverse stiffnesses of sheep horn were similar. This is unlikely to be the case for gemsbok horn, because when it is dried it readily splits longitudinally, which sheep horn does not do. Therefore, it seems to be anisotropic. It was not possible to measure the transverse stiffness of gemsbok horn because of the small dimensions of the horns which made it impossible to manufacture appropriate test-pieces. Therefore, two cases will be considered: an isotropic case and an anisotropic case using an arbitrary transverse stiffness of 2.25 GPa which is half the longitudinal stiffness.

FRACTURE IN H O R N S 637
Warburton (1948) measured a Poisson ratio of 0.23 for sheep horn using an ultrasonic method. Kitchener (1985) measured repeatedly a Poisson ratio of about 0.4 for gemsbok horn using orthogonal strain gauges. In view of the paucity of data concerning the Poisson ratio of horn, an arbitrary value of 0.3 was used.
The Gurney & Hunt work of fracture of 19.8 kJ/m2 gives a critical stress intensity factor of between 8.3 (anisotropic) and 8.6 (isotropic) MPa/mi. This is much greater than the mean Kc of 4.5 MPa/m for fresh horn. The Gurney & Hunt work of fracture excluding plastic work gives an expected Kc of 6.2-6.6 MPa/mf and the critical strain energy release rate, Gc, gives an expected Kc of 54-56 MPa/mf, both of which are closer to the experimental value.
Ductility affects the measured value of Kc, which can be corrected using the following (Knott, 1973):
KC = Y . d * [a * ( 1 + d/(2 *a”))]
where: Y is the geometry factor (see above), a‘ is fracture stress (Pa), a is notch length (m), aY is yield stress of material (122 MPa).
This increases the mean experimental Kc to 4.9 MPa/mj, which is in agreement with the Kc predicted from the Gc. Therefore, it seems that LEFM can be applied to fresh horn.
However, it was not possible to find any similarity between the stress intensity factors and work of fractures for dry and wet horn. Therefore, the results obtained above may be purely coincidental or represent a special case for fresh horn keratin. These results should be treated with caution.
Horning hehaviour and the toughness of horns
The behavioural repertoire of bovids includes a wide variety of agonistic displays which are used in the defence of territories, competition for resources and determination of social rank (Geist, 197 I; Leuthold, 1977; Schaller, 1977; Walther, 1978). Agonistic displays are intended to intimidate and deter rivals because the risks of fighting are very high, but they can never replace the final deterrent of fighting (Geist, 1971; Clutton-Brock, 1982).
Bovids push their horns frequently into the ground (usually wet mud) or thrash them against vegetation. This behaviour is known as horning and seems to be common to all bovids (Leuthold, 1977). In the chamois (Rupicapra rupicupra) and the mountain goat (Oreumnos americanus), it is associated with the scent-marking of the boundaries of territories, using the post-cornual glands at the base of the horns (Schaller, 1977). However, for most bovids it is a poorly understood behaviour and has been variously interpreted as an agonistic display or redirected aggression, or a displacement activity, depending on its context (Walther, 1966; Jacobsen, 1974; Leuthold, 1977; Anderson, 1980; Spinage, 1982; Walther, Mungall & Grau, 1983).
The results above have demonstrated that bovids must be careful to ensure that their horns are adequately hydrated to maintain the toughness and notch-insensitivity of horn keratin against the desiccating environment. If horns were to dry out, scratches and cracks accumulated in fighting could cause the failure of the horns, even when loaded under normal conditions of fighting. The work of fracture would be reduced because the matrix would not be able to yield and crack tip fracture mechanisms could no longer occur. If horns were to break readily, reproductive success would be affected adversely and permanently.
The horning of wet mud by bovids may be a maintenance activity to ensure that the surface of horns do not become too dehydrated and brittle. The thrashing of vegetation may cause resinous substances to be applied to the horns which would prevent water loss as well as plasticizing the

638 A. KITCHENER
horns. This would explain the variety of different circumstances in which horning has been observed. It is easy to see how a maintenance activity could become a ritualized agonistic display.
It is unlikely that horns could absorb too much water when immersed in wet mud and become less tough as a consequence. This is because it would take fresh horn more than two days of constant immersion to become fully hydrated.
Summary
Horn sheath keratin is a very tough composite material which is ideal for its continued use in intraspecific combat. The high work of fracture of horn sheath relies on a variety of toughening mechanisms at the site of crack propagation and the considerable yielding of the matrix phase of the composite. Fresh horn is a notch-insensitive material so that scratches and cracks accumulated in fighting do not affect adversely the strength of horn keratin. However, the desiccating effect of the environment may cause dehydration of horn keratin so that it becomes notch-sensitive and its toughness is reduced. The horning behaviour of bovids may be a maintenance activity to ensure that the surface of horns are sufficiently hydrated against the desiccating effects of the environment.
I would like to thank Richard Cindery of the Zoological Society of London’s Whipsnade Park Zoo and Dr Juliet Clutton-Brock of the British Museum (Natural History) for supplying me with horns. I would like to thank Andrew Jackson and Mike Watkins for carrying out the impact tests. I would also like to thank Prof. A. Atkins for helping me to discover more about fracture mechanics. I am especially grateful to Alan Jenkins and Liz Wheeler for their help with electron microscopy and printing ‘hundreds’ of photographs. Finally, I would like to thank Dr Julian Vincent and Andrew Jackson for their criticism of this manuscript. This work was carried out under the Herbert Knapman Postgraduate Award in Science at the University of Reading.
REFERENCES
Anderson, J. L. (1980). The social organization and aspects of behaviour of the nyala, Trageraphus angasi Gray, 1849.
Atkins, A. G. & Mai, Y.-M. (1985). Elastic andplastic fracture. Chichester: Ellis Horwood. Clutton-Brock, T. H. (1982). The functions of antlers. Behauiour 7 9 108-125. Clutton-Brock, T. H., Guinness, F. E. & Albon, S. A. (1982). Red deer. Behaviour and ecology of two sexes. Edinburgh:
Currey, J. D. (1979). Mechanical properties of bone tissues with greatly differing functions. J . Biomech. 12 313-319. Dorrington, K. L. (1980). The theory of viscoelasticity in biomaterials. Symp. SOC. exp. Biol. No. 34: 289-314. Fraser, R. D. B. & Macrae, T. P. (1980). Molecular structure and mechanical properties of keratins. Symp. SOC. exp. Biol.
Geist, V. (1971). Mountain sheep. A study in behavior and evolufion. Chicago: Chicago University Press. Gordon, J. E. (1976). The new science of strong materials. 2nd edn. London: Penguin. Gordon, I. E. (1978). Structures. London: Penguin. Goss, R. J. (1983). Deer antlers. Regeneration. function and evolution. London: Academic Press. Gurney, C. & Hunt, J. (1967). Quasi-static crack propagation. Proc. R. Soc. ( A ) 299: 508-524. Harris, 9 . (1980). The mechanical behaviour of composite materials. Symp. SOC. exp. Biol. No. 34: 37-74. Hull, D. (1981). An introduction to composite materials. Cambridge: Cambridge University Press. Jacobsen, N. H. G. (1974). Distribution, home range and behaviour patterns of bushbuck in the Lutope and Sengwa
Kitchener, A. C. (1985). The functional design of horns. PhD thesis, University of Reading. Kitchener, A. & Vincent, J. F. V. (1987). Composite theory and the effect of water on the stiffness of horn. J. Mat. Sci. 2 2
2. Saugetierk. 45: 90-123.
Edinburgh University Press.
NO. 34: 21 1-246.
valleys, Rhodesia. J. S. Afr. Wildl. Mgmt Ass. 4 75-93.
138S1389.

FRACTURE I N HORNS 639
Knott, J. F. (1973). Fundamentals offraclure mechanics. London: Butterworth. Leuthold, W. (1977). African ungulates. A comparative review oftheir ethology and behavioralecology. New York: Springer
Lincoln. G. A. (1972). The role of antlers in the behaviour of the red deer. J. exp. Zool. 182: 233-250. Makinson, K. R. (1954). The elastic anisotropy of keratinous solids. I. The dilatational elastic constants. Aust. J. Biol. Sci.
Mullins, L. (1980). Theories of rubber-like elasticity and the behaviour offilled rubbers. Symp. Soc. exp. Biol. No. 34: 273-
Nievergelt, B. (1981). Ibexes in an African environment. New York: Springer Verlag. Piggott, M. R. (1980). Load-bearingjbre composites. Oxford: Pergamon. Schaller, G. B. (1977). Mountain monarchs. Wild sheep and goats of the Himalayas. Chicago: Chicago University Press. Spearman, R. I. C. (1977). Keratins and keratinization. Symp. zool. Soc. Lond. No. 39: 335-352. Spinage, C. A. (1982). A territorial antelope: The Uganda waterbuck. London: Academic Press. Tattersall, H. G. & Tappin, G. (1966). The work of fracture and its measurement in metals, ceramics and other materials.
Vincent, J. F. V. (1982). The mechanical design of grass. J. Mat. Sci. 17: 8 5 6 8 6 0 . Vincent, J. F. V. & Gravel], K. (1986). The mechanical design of kelp, Laminaria digitata. J. Mat. Sci. Lett. 5: 353-354. Walther, F. R. (1966). Mit Horn und Huf. Berlin: Paul Parey. Walther, F. R. (1978). Behavioural observations on oryx antelope (Oryx beisa) invading the Serengeti National Park.
Walther, F. R., Mungall, E. C. & Grau, G. A. (1983). Gazelles and their relatives. A study in territorial behavior. New Jersey:
Warburton, F. L. (1948). Determination of the elastic properties of horn keratin. J. Text. Inst. 3 9 297-308. Williams, J. G. (1984). Fracture mechanics ofpolymers. Chichester: Ellis-Horwood. Williams, J. G. & Birch, M. W. (1976). Mixed mode fracture mechanics in anisotropic media. A.S.T.M. Spec. Publ. 601:
Verlag.
8: 33G347.
288.
J. Mat. Sci. 1: 2 9 6 3 0 I .
J . Mammal. 5 9 243-260.
Noyes Publications.
125-137.