freeze-fracture study of phagocytosis in...
TRANSCRIPT

J. Cell Sci. 51, 63-84 (1981) 63Printed in Great Britain © Company of Biologists Limited 1081
FREEZE-FRACTURE STUDY OFPHAGOCYTOSIS IN DICTYOSTELIUMDISCOIDEUM
COLETTE FAVARD-SE"R£NO, MARIE-ALINE LUDOSKYAND ANTOINETTE RYTER
Laboratoire de Microscopie Electronique, Institut de Recherche en BiologieMoliculaire du CNRS, UniversiU Paris VII, 2, place Jussieu, 75221 Paris Cedex 05,and Department de Biologie Moliculaire, Institut Pasteur, 75724 Paris Cedex 15
SUMMARY
The plasma membrane and its derivative, the phagosome membrane, were studied duringand after ingestion of yeast or latex beads in Dictyostelium discoideum. Freeze-fracture electronmicroscopy, which provides information on the internal architecture of the membranes, andobservation of thin sections of cells treated by cytochemical methods were used in parallel.For visualization of membrane sterols in the replicas, the cells were fixed in the presence ofdigitonin or the antibiotic filipin.
No lateral phase separation occurred during yeast engulfment: the intramembranous par-ticles (IMPs), phospholipids and sterols remained distributed at random in the formingphagosome membrane. In contrast architectural modifications of the membrane were observedupon phagosome internalization. Compared to the plasma membrane, the phagosome membranedisplayed 2-3 times more IMPs, a shift in the IMP size distribution and a higher sterol content.These changes were completed soon after phagosome closure; they were not related eitherto the nature of the ingested particles (yeast, latex beads) or to the pH in the membrane environ-ment. The membrane changes took place when the phagosomes began to fuse with pre-existingdigestive or autophagic vacuoles and lysosomes. Some of the experimental evidence suggests thatthe restructuring of the membrane may be related to the presence of hydrolases.
INTRODUCTION
In many cell types the endocytic process leading to the internalization of the plasmamembrane seems to be accompanied or followed by various changes in the compositionor the organization of the internalized plasma membrane. The biochemical dataobtained during recent years indicate that in varied cell types, the plasma membrane(PM) that is internalized during phagocytosis or pinocytosis, does not contain all ofthe cell-surface constituents. Some proteins, enzymes, glycoconjugates or lipids,characteristic of the PM, are either not found or at least not found in the sameamount in the phagosome or pinosome membranes (Nachman, Ferris & Hirsch,1971; Ukena & Berlin, 1972; Parish & Pelli, 1974; Smolen & Shohet, 1974; vanVliet et al. 1976; Vincenzi & Larsen, 1980). This suggests either that endocytosistakes place in specialized areas of the PM, or that some segregation phenomenonoccurs during the endocytic process; an alternative interpretation being that thephagosome membrane (PhM) is modified after its internalization.
Rearrangements of proteins in the membrane architecture were also detected

64 C. Favard-Sereno, M.-A. Ludosky and A. Ryter
either during or soon after the end of the phagocytic process. Thorne, Oliver &Glauert (1980) observed that 2 proteins of the eosinophil plasma membrane becomeexposed to the cell surface only during the endocytic process. Furthermore, freeze-fracturing revealed changes in intra-membrane particle (IMP) density and size.In Acanihamoeba castellanii (Bowers, 1980) and Tetrahymena (Kitajima & Thompson,1977), the number of IMPs increased soon after phagosome formation. In Paramecium(Allen, 1976) some changes were observed during the complex circulation of foodvacuoles.
Moreover, morphological evidence of the disappearance of glucidic membranecomponents was obtained during pinocytosis and phagocytosis, de Bruyn, Michelson& Becker (1978) observed that sialic acid residues were not detectable in pinocyticcups of an endothelial cell type, and one of us (Ryter & Hellio, 1980) observed thedisappearance of wheat germ agglutinin receptors from the phagocytic membraneduring yeast engulfment, in D. discoideum amoeba.
To obtain a better insight into the evolution of membrane constituents during andafter phagocytosis, we undertook an electron microscopic study of yeast and latex-bead phagocytosis in D. discoideum amoebae using mainly freeze-fracturing methods.With this technique the different membrane components can be visualized: intrinsicproteins are visible as intramembranous particles (IMPs) (for a review, see Singer,1971), phospholipids (PL) appear as smooth surfaces and sterols, when complexedto the antibiotic filipin, are viewed as 25 nm protrusions (Tillack & Kinsky, 1973;Verkleij et al. 1973) in the fractured membrane replicas.
We observed that the internal architecture of the PM was not modified during theengulfment process. In contrast, structural alterations were found in fully formedphagosomes: increased IMP density and shift in the IMP size distribution, whichtook place soon after the phagosome closure and release into the cytoplasm. Thesechanges coincided with fusions between phagosome and lysosome membranes andthey seemed to be accompanied by sterol enrichment of the phagosome membrane.
MATERIALS AND METHODS
Culture conditions and phagocytosis
D. discoideum AX-2 strain was grown in HL-5 axenic medium as described elsewhere(Ryter & Hellio, 1980). The conditions of phagocytosis were also identical to those given inthis paper. In phagocytosis experiments with concanavalin A (Con-A)-coated yeast (10* per ml),previously fixed with 2 % glutaraldehyde and dehydrated with acetone, was treated with 100 figCon A for 15 min at 20 °C. The yeast Was washed twice before being added to starved cells.
Electron microscopy
Five ml of cell suspension were added to the same volume of 2-5 % glutaraldehyde ino-i M-cacodylate buffer (pH 6-8) containing o-i M-sucrose. After 15 min, the cells were spundown and resuspended in the same fixative. After 1 h at 20 °C they were prepared either forthin sectioning as described previously (Ryter & de Chastellier, 1977) or for freeze-fracturing.The cytochemical demonstration of acid phosphatase was performed as described by Ryter &de Chastellier (1977).

Phagocytosis in D. discoideum 65
Preparation of cells for freeze-fracturing
Following fixation, the cells were washed several times in cacodylate buffer and then im-pregnated with increasing concentrations of glycerol (5, 10 and 20%, v/v) in distilled waterfor a total of 2 h. After centrifugation, small samples of the cell pellet were placed on gold alloyspecimen carriers and rapidly frozen in the liquid phase of partially solidified Freon 22 cooledby liquid nitrogen.
The samples were freeze-fractured at —140 °C in a Balzers BA300 apparatus, according tothe technique of Moor & Miihlethaler (1963). The freeze-fracture replicas were cleaned insodium hypochlorite, washed in distilled water and finally mounted on copper grids andexamined in a Philips EM300 electron microscope.
Incubation with digitonin
The cells were prefixed in the culture medium for 15 min as usual, then centrifuged andresuspended in the fixative containing 0-2 % digitonin. They were incubated in this mixturefor 1-3 h before being processed either for embedding, or for freeze-fracturing as reportedabove.
Incubation with filipin
The filipin solutions in dimethylformamide (DMF) were prepared just prior to use. Inorder to prevent membrane disruption during sterol-filipin complex formation, the cells wereprefixed with glutaraldehyde in the culture medium for 30 min, centrifuged and resuspendedin the fixative containing the antibiotic at a final concentration of i8o/*g/ml (DMF finalconcn, 1 %). The cells were incubated in this medium for 17 h at room temperature, washed andprocessed for freeze-fracturing as mentioned above. Control cells were treated in the sameconditions with 1 % DMF in the absence of filipin.
Ouantitation of IMP size and concentration
The negatives were printed at a final magnification of x 200000 for measurement of particlesize and of x 100000 for counting particle numbers. The diameters of the IMP were measuredas the width of the shadow perpendicular to the direction of shadowing. To perform thequantitative evaluation, the membrane fracture faces were carefully selected for flatness.
Quantitation of sterol-filipin complexes
The sterol-filipin complexes appeared as protrusions in the freeze-fractured membranes andwere counted on micrographs at a final magnification of x 100000.
RESULTS
Yeast phagocytosis
Phagocytosis experiments were all performed with cells starved for 2 h in phosphatebuffer, because the speed and level of yeast uptake are then twice as high as in vegeta-tive cells growing in axenic medium (de Chastellier & Ryter, 1977). After this starva-tion (2 h) the digestive vacuoles start to be replaced by autophagic vacuoles containingessentially membrane fragments (de Chastellier & Ryter, 1977). Cells fixed 10 minafter yeast addition already contained several phagosomes. In most of them, the yeastsurface was not in tight apposition to the phagosome membrane, but instead wasseparated by a space containing many patches of spongy material or membranefragments (Figs. 1, 3). The presence of these 2 kinds of material, quite similar to that

66 C. Favard-Sereno, M.-A. Ludosky and A. Ryter
found inside digestive (Ryter & de Chastellier, 1977) or autophagic vacuoles (Fig. 3),suggested that phagosomes had fused with those 2 types of vacuoles. The cyto-chemical demonstration of acid phosphatase (AcPase) showed, moreover, that theappearance of this hydrolase was concomitant with these fusions, although thespongy material itself was seldom covered with lead deposit (Ryter & de Chastellier,1977). These fusions occurred very soon after yeast ingestion, since 5 min after yeastaddition almost 50% of the phagosomes contained some spongy material or membranefragments (Fig. 5). These fusions continued to increase for 15 min and then seemedto be much less frequent, because the number of phagosomes containing this materialremained almost constant. The space between the yeast and the phagosome membranethus seems to result from an increase in area of the phagosome membrane producedby these fusions. We must mention that such fusions also occurred very rapidlywhen undigestible particles such as latex beads were ingested (Fig. 2).
When studied by freeze-fracture electron microscopy, the plasma membranes ofstarved cells displayed a symmetrical distribution of IMPs (Favard-Ser^no & Livrozet,1979). The mean number of particles per /im2 was then similar on the protoplasmic(PF) and exoplasmic (EF) fracture faces (Fig. 6). When the PM invaginated to formthe phagocytic vacuole, the IMP density in the forming phagosome membrane wasidentical to that of the plasma membrane from which it originated. Moreover, theparticles remained distributed at random, even in the region where the yeast was inclose contact with the PM. The only exception to this random distribution was visibleon the E face in PM regions between the forming phagosomes, where clusters oflarge particles were observed (Fig. 7). Such clusters were never found on the P face(Fig. 8).
We did not observe the closure of the phagocytic vacuole, an event that doesrequire membrane fusion and seems to be very rapidly performed in Dictyosteliumamoebae. Among the numerous membrane-bound vesicles found in the cytoplasm offreeze-fractured cells, the internalized phagosomes were easily identified by theirlarge size and, sometimes, by the configuration of the bud-scar region of the yeastthey contained. The newly formed phagosomes were surrounded by vesicles some of
Fig. 1. Thin section of a yeast-containing phagosome showing many patches ofspongy material (arrowheads) in the space between the yeast and the phagosomemembrane. Most are localized in regions where the phagosome membrane is curvedinwards. This picture suggests that the phagosome has fused with many lysosomesor digestive vacuoles. Bar, 1 fim.
Fig. 2. Latex-bead phagosomes in which the phagosome membrane is not tightlyapposed to the bead. Spongy material is visible in this space (arrowheads) suggestingthat fusions with lysosomes have occurred, dv, digestive vacuoles. Bar, 1 fim.Fig. 3. Part of a yeast phagosome showing membranous material in the space betweenyeast and membrane (arrowheads). This membranous material is quite similar tothat visible inside the surrounding autophagic vacuoles (an). Bar, 1 fim.Fig. 4. High magnification of a portion of phagosome containing Con A-coated yeast(y). The phagosome membrane (ph m) is tightly apposed to the yeast surface and nospongy or membranous material is visible between them. Bar, 0-2 fim.

Phagocytosis in D. discoideum
_ri

68 C. Favard-Sereno, M.-A. Ludosky and A. Ryter
which most probably corresponded to lysosomes (Figs, n , 12). In cross-fractures acontinuity was sometimes observed between the phagosome membrane and themembranes of vesicles of various sizes, suggesting that fusions had occurred. Alongthe ordinarily intact membrane fracture faces, the fusion sites with vesicles appearedas small round-shaped holes. Moreover, the phagosome membrane displayed manyIMP-free areas of high intrinsic curvature (Figs. 11, 12), which represent regionswhere lysosome and phagosome membranes are apposed prior to fusion. Fusionsbetween phagosomes and digestive, or autophagic vacuoles are completed by the
100-
7 5 -
5 0 -
2 5 -
5 10 15 30Time (min)
Fig. 5. Frequency of yeast phagosomes containing spongy or membranous materialbetween the yeast and their membrane, in cells fixed at different times after yeastaddition.
same process. The contents of autophagic vacuoles were sometimes visible in thephagosome cavity in the form of stacked pieces of smooth bilayer always deprived ofIMPs. In neither case could we detect rosette-forming arrays of IMPs at the siteswhere the phagosome membrane was interacting with the limiting membranes ofother vesicles, or vacuoles.
By the time the phagosome transformed into a phagolysosome, we observed anenhancement in the number of IMPs/^m2 on both fracture faces of its membrane(Figs. 9, 10). This increase in IMP density was gradual and reached a maximal valuein the membrane of 15-min-old phagosomes (Fig. 6). Both faces of the phagosomemembrane then displayed a 2- to 3-fold increase in IMP density as compared to the

Phagocytosis in D. discoideum 69
PM (Fig. 6). As for the size distribution: in the PF IMPs were considerably largerthan in the PM, whereas in the EF the enrichment was represented mainly by small-sized particles (Fig. 13). Some of the particles of the phagolysosome membranepresented an indented shadow and thus seemed to be made up of linear arrays of2-4 individual particles (Figs. 9, 10); the values of the diameters of these formationswere not plotted. When the amoebae were given latex beads instead of yeast particles,IMP enrichment and changes in IMP size distribution were observed as well. In thiscase also the changes were subsequent to fusions of the phagosomes with lysosomes.
4000-
E 3000-a.
XL£
. 2000-o
1000 -
Plasmamembrane
2 mm 5 min 15 mm
PhagosomesFig. 6. Number of IMPs per /*m' on the EF and PF of the plasma membrane, and ofphagosome membranes in cells fixed 2, 5 or 15 min after yeast addition. The barsrepresent the standard deviation.
Ryter & de Chastellier (1977) have demonstrated that the membranes of the diges-tive system of D. discoideum are recycled back to the plasma membrane when thedebris of digested yeast are released into the culture medium. Nevertheless, neitherphagosome-plasma membrane fusions nor PM areas with a high content of large IMPswere visualized, even after long incubations (more than 2 h) in the presence of yeast.
Ingestion of Con A-coatedyeast
The fact that the increase in number of IMPs seemed to coincide with vacuolefusion suggested that these 2 events were tightly correlated. We have therefore tried.

7° C. Favard-Sereno, M.-A. Ludosky and A. Ryter
to find conditions in which these fusions did not occur. Because Edelson & Cohn(1974) observed that Con A-containing vacuoles did not fuse with lysosomes orother pinosomes in macrophages, we coated the yeast with this lectin before givingthem to amoebae.
Observations in thin sections. Con A-coated yeast were added to cells starved for 2 has for untreated yeast. Light microscopic observations showed that they very quicklyadhered to the cell surface but their uptake was slower and less than with untreatedyeast. In thin sections, Con A-coated yeast phagosomes contained no space betweenyeast and membrane (Fig. 14), and no spongy or membranous material was foundinside the phagosomes. The tight and continuous contact between yeast and membrane(Fig. 4) persisted for about 45 min. A quantitative comparison was made betweenphagosomes containing untreated yeast and those containing Con A-coated yeast inthe cells fixed 30 min after yeast addition. Of the 35 phagosomes counted in eachsample, 33 phagosomes containing untreated yeast had a large space between mem-brane and yeast and many patches of spongy or membranous material. An inverseratio was found for Con A-coated yeast: 2 contained some material between theyeast and the membrane, and 33 contained no space and no material. After 45 min,the space started to appear and the Con A layer became progressively detached fromthe yeast surface and the phagosome membrane. These observations thus indicatedthat the yeast-Con A layer strongly bound to the phagosome membrane and preventedits fusion with digestive or autophagic vacuoles. They also suggested that phagosomesdid not receive lysosomal enzymes. To determine whether this latter assumption wascorrect or not, acid phosphatase was revealed cytochemically in cells fixed 15 or30 min after addition of untreated or Con A-coated yeast. In fact, both samplescontained the same number of AcPase positive phagosomes in light or electronmicroscopy, but the localization of lead deposit was different (Figs. 15, 16). Inuntreated yeast, lead was distributed in the yeast cytoplasm whereas it remainedconfined to the surface of Con A-coated yeast. AcPase is probably immobilized by thelectin because its glucidic moiety contains Con A receptors.
These results thus show that, although Con A-coated yeast phagosomes showed novisible increase of membrane area due to fusion with digestive vacuoles, they haveapparently received a normal amount of AcPase. This suggests that this hydrolasecomes from tiny primary lysosomes in which it is highly concentrated.
Observation of freeze-fractured cells. In cross-fractures of cells fixed 15 min afteraddition of Con A-coated yeast to the culture medium, the membrane limiting thephagosomes appeared tightly apposed to the yeast envelopes. As compared to thoseof the PM, the fracture faces of the yeast-Con A phagosome membrane displayed the
Figs. 7, 8. Plasma membrane E face (EF) (Fig. 7) and P face (PF) (Fig. 8) of a cellfixed during yeast phagocytosis. On the EF some IMP clusters are visible. Bar, o-1 fim.Figs. 9, 10. Phagosome membrane EF (Fig. 9) and PF (Fig. 10) of a cell fixed 15 minafter yeast addition. On both faces, the number of IMPs is much higher than in Figs. 7and 8. Bar, o-i fim.

Phagocytosis in D. discoideum

C. Favard-Sereno, M.-A. Ludosky and A. Ryter
tigs, I I , 12. Detail ot a yeast-containing phagosome showing contacts with smallvesicles or vacuoles (black arrowheads). Small areas devoid of IMPs probably corres-pond to regions of lysosome fusion (white arrows). Bar, i fim.
same progressive increase in IMP density and the same alterations of IMP size dis-tribution as the membrane of untreated yeast phagosomes. Fusions with lysosomesdid not seem to be delayed by the presence of Con A-coated yeast. In contrast,fusions with autophagic and digestive vacuoles, and subsequent increase in surfacearea of the phagosome membrane, were observed only 30-40 min after ingestion ofCon A-coated yeast. Because the change in the number and size of IMPs seemed to berelated to the presence of hydrolases inside phagosomes we studied the influence ofpH on the plasma membrane.

Phagocytosis in D. discoideum 73
Effect of low pH on the plasma membrane
Cells starved for 2 h were incubated for 15 min in phosphate buffer at pH 5. Theywere fixed for 15 min in this buffer with 2 5 % glutaraldehyde and then transferred tothe usual fixative at pH 6-8 and fixed for 1 h. Observation of cryo-fractures did notreveal any changes in the number or size of IMPs in the plasma membrane or intra-cellular membranes but showed frequent IMP clusters on the EF similar to thoseobserved after Con A labelling (Favard-Se're'no & Livrozet, 1979).
PF
50-
4 0 -
- 30-
2 0 -
1 0 -
EF
8 12 16 20 4 8 12 16Size (nm) Size (nm)
Fig. 13. IMP size distribution in PF and EF of PM (open bars) and Ph M (cross-hatched bars) in cells fixed 15 min after yeast addition.
Effect of chloroquine
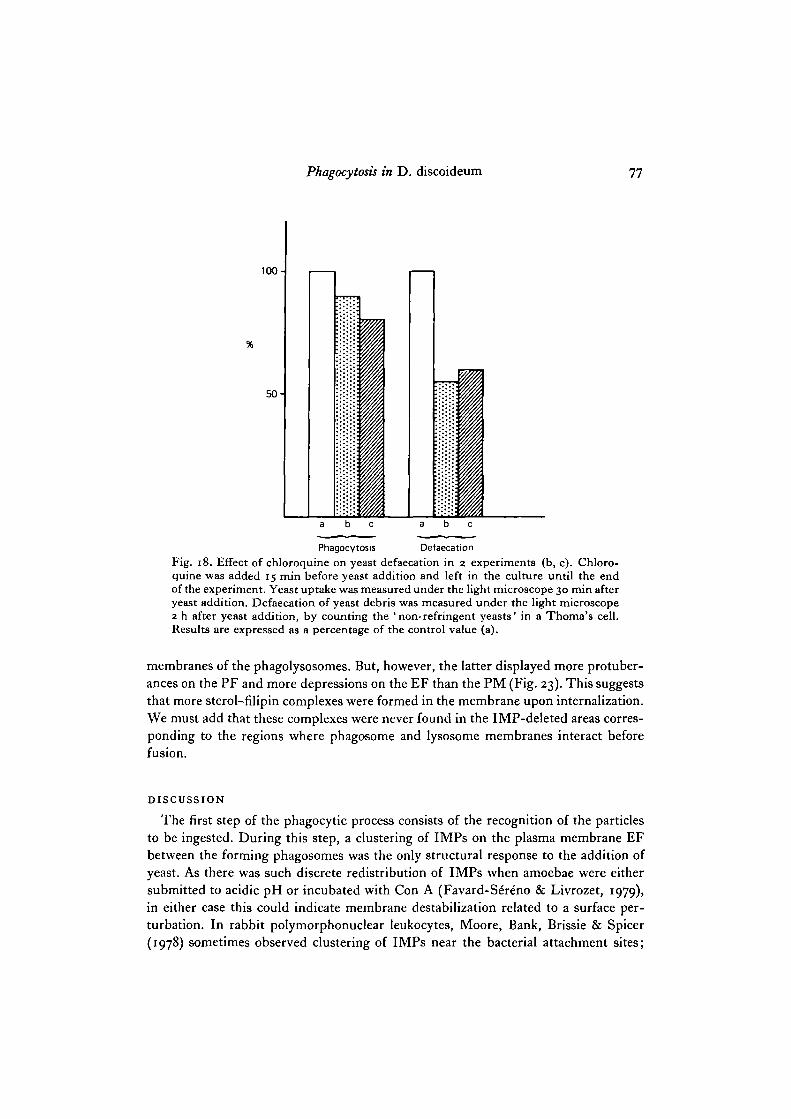
It has been shown (Wibo & Poole, 1974; Gonzales-Noriega, Grubb, Talkad &Sly, 1980; Tietze, Schlesinger & Stahl, 1980) that the addition of chloroquine to theculture medium of many animal cell types inhibits the digestion of endocytizedmaterial by maintaining a high pH in secondary lysosomes (Ohkuma & Poole, 1978).Chloroquine (o-i mM) was added at different times before yeast addition, and yeastuptake was determined under the light microscope 30 min after addition of theseparticles. Chloroquine inhibited yeast phagocytosis and this inhibition increasedwith the length of contact before yeast addition (Fig. 17). A 15 min preincubation waschosen to follow the effect on yeast defaecation. Yeast digestion was determined bycounting with a Thoma's cell counter the yeast debris defaecated into the culturemedium after 2 h (Ryter & de Chastellier, 1977). These debris correspond to indiges-tible yeast cell wall and appear under the light microscope as ' dark yeast' in contrastto the bright intact yeast. In these conditions, yeast phagocytosis decreased by 15-20% and yeast defaecation by 50 -55% (Fig. 18). It can be concluded that in Dicty-ostelium chloroquine has an inhibitory effect on both phagocytosis and digestion, asin mammalian cells (Wibo & Poole, 1974; Gonzales-Noriega et al. 1980; Tietze et al.

74 C. Favard-Sereno, M.-A. Ludosky and A. Ryter
1980). This inhibition of digestion had no effect, however, on the cellular morphology.In thin sections phagosomes were seen fused with many vacuoles and freeze-fracturepreparations also presented an IMP increase.
Membrane sterol visualization
Phagocytosis is accompanied by deformation and polarization of the PM and thusmight give rise to alterations in the composition and microviscosity of the membrane(Kitajima, Sekiya & Nozawa, 1976; Kitajima & Thompson, 1977). As the presenceof sterols within the bilayer matrix causes modulations in the lipid fluidity, we havemade 2 attempts to estimate possible variations in the sterol content of the PM duringthe internalization process and the completion of yeast digestion.
Some authors (Elias, Goerke & Friend, 1978) observed that addition of digitoninduring glutaraldehyde fixation leads to characteristic membrane deformations, visibleon thin sections and in cryo-fracture. They showed that, according to the sterolcontent of the cellular membrane, significant differences could be seen in the intensityof these deformations. This method, applied to Dictyostelium during or after phago-cytosis, showed in thin sections that the phagosome membrane was much moreindented than usual. No difference could be found between phagocytic cups andyoung or old phagosomes. These indentations were not so striking on the plasmamembrane because, even in the absence of digitonin, starved cells present a highlyfolded plasma membrane (de Chastellier & Ryter, 1977). Autophagic vacuoles anddigestive vacuoles showed the same deformations as phagosomes, whereas endoplasmicreticulum, nuclear membrane and mitochondrial membrane were not affected.
Upon fracturing, PM as well as phagosomal and lysosomal membranes of digitonin-treated cells appeared heavily convoluted and displayed, on both fracture faces,convexities and concavities that correspond to sterol-digitonin complexes. Despitethe fact that digitonin is a powerful detergent, no membrane disruption was observed,possibly because the cells were fixed with glutaraldehyde before being treated withdigitonin. In between the digitonin-induced corrugations the IMPs were randomlydistributed, no particle clusters being observed. The nuclear envelope, endoplasmicreticulum and mitochondrial membranes were not perturbed by treatment withdigitonin, whilst phagosomal and lysosomal membranes in their vicinity were modified.This clearly indicates sterol deficiencies in the membranes of these organelles.Although the reacting membranes looked uniformly plicated, more convexities wereobserved in the EF and more concavities on the PF face. This could suggest anasymmetric distribution of sterols between the 2 membrane halves, but the large size
Fig. 14. Thin section of Con A-coated yeast-containing phagosomes in a cell fixed30 min after yeast addition. No space and no spongy or membranous material can beseen between the yeast and the phagosome membrane. Bar, 1 fim.Figs. 15, 16. Cytochemical demonstration of acid phosphatase in cells fixed 10 minafter yeast addition. The phagosome of Fig. 15 contains an untreated yeast; leaddeposit is randomly distributed. The phagosome of Fig. 16 contains a Con A-coatedyeast; lead deposit remains exclusively located along the yeast surface. Bar, 1 fun.

Phagocytosis in D. discoideum
14

76 C. Favard-Sereno, M.-A. Ludosky and A. Ryter
of the digitonin molecule and the complexes it forms with sterols in the membranesdo not allow us to make any statement on this point.
In an attempt to obtain more quantitative information concerning distribution ofsterols in the membranes we examined cells treated with filipin, a polyene antibioticthat forms complexes of reduced size with sterol molecules. These complexes are notvisualized in thin sections, but produce membrane alterations that are easily viewedin freeze-fracture replicas (Tillack & Kinsky, 1973; Verkleij et al. 1973). The cellswere incubated for 7 min in the presence of yeast before prefixation and subsequenttreatment with filipin. Both the PM and the phagosome membranes strongly inter-acted with the antibiotic as shown by the numerous deformations visible on thefracture faces (Figs. 19, 20, 21, 22). The sterol-filipin complexes appeared as bumps
100 -
50-
25 -
45 60C 0 15 30Time (min)
Fig. 17. Effect of chloroquine on yeast phagocytosis. Cells were treated with o-i mM-chloroquine for different periods before yeast addition (o to 60 min), and during yeastuptake. The number of ingested yeast per cell was determined by light microscopy60 min after yeast addition. The results are expressed as a percentage of the controlvalue (C) observed without chloroquine.
protruding from the rest of the fracture-plane of the membrane. These protruberanceswere ellipsoid and the width of their platinum shadow was between 15 and 25 nm.An asymmetry obviously exists in the distribution of the complexes between the Eand P fracture faces of the plasma membrane. Three to four-fold more protuberanceswere encountered in 1 /tm2 of the EF than in the same area of the PF (Fig. 23).Between the bumps depressions about 15 nm in diameter were observed that presentan inverted shadow as compared to the protuberances (Figs. 19, 20, 21, 22). Therewere twice as many craters in the PF as in the EF of the PM (Fig. 23). No redistribu-tion of the IMPs of plasma membrane was induced by the antibiotic and the 2 typesof alteration produced by the formation of the complexes were seen at random in themembrane plane. No special pattern of the alterations described above was observed,either in the regions of the PM that contain the forming phagosomes, or in the
Phagocytosis in D. discoideum 77
100-
50 -
Phagocytosis DefaecationFig. 18. Effect of chloroquine on yeast defaecation in 2 experiments (b, c). Chloro-quine was added 15 min before yeast addition and left in the culture until the endof the experiment. Yeast uptake was measured under the light microscope 30 min afteryeast addition. Defaecation of yeast debris was measured under the light microscope2 h after yeast addition, by counting the 'non-refringent yeasts' in a Thoma's cell.Results are expressed as a percentage of the control value (a).
membranes of the phagolysosomes. But, however, the latter displayed more protuber-ances on the PF and more depressions on the EF than the PM (Fig. 23). This suggeststhat more sterol-filipin complexes were formed in the membrane upon internalization.We must add that these complexes were never found in the IMP-deleted areas corres-ponding to the regions where phagosome and lysosome membranes interact beforefusion.
DISCUSSION
The first step of the phagocytic process consists of the recognition of the particlesto be ingested. During this step, a clustering of IMPs on the plasma membrane EFbetween the forming phagosomes was the only structural response to the addition ofyeast. As there was such discrete redistribution of IMPs when amoebae were eithersubmitted to acidic pH or incubated with Con A (Favard-SeY^no & Livrozet, 1979),in either case this could indicate membrane destabilization related to a surface per-turbation. In rabbit polymorphonuclear leukocytes, Moore, Bank, Brissie & Spicer(1978) sometimes observed clustering of IMPs near the bacterial attachment sites;

C. Favard-Sereno, M.-A. Ludosky and A. Ryter
. 20
A »
^ - ^̂ ^
2 - 1 '•••" 22'

Phagocytosis in D. discoideum 79
this was never found with yeast ingested by Dictyostelium. Furthermore, the formingphagosomes presented IMPs with size range, distribution and density similar to thoseof the PM. This means that the membrane curvature and the presence of a thick layerof contractile proteins under this area of the membrane do not have any influence onthe distribution of IMPs. It can also be concluded that the disappearance of wheatgerm agglutinin (WGA) receptors from the membrane of phagocytic cups (Ryter &Hellio, 1980) cannot be related to a change in IMPs, either because this disappearanceonly concerns the glucidic moiety of glycoproteins, or because WGA receptors arenot transmembrane proteins or transmembrane proteins detectable as IMPs.
Phagocytic vacuole closure is a very rapid process (Bowers, 1980; Segal, Dorling &Coade, 1980). In Dictyostelium also it seems to be completed within seconds and wasnever detected by freeze-fracturing methods or in thin sections. In contrast, fusionsbetween phagosome membranes and lysosomes (appearing as IMP-free areas) wereobserved very soon after phagosome formation, as in other cell types (Batz & Wunder-lich, 1976; Pryzwansky, MacRae, Spitznagel & Cooney, 1979; Bowers, 1980). Thus inDictyostelium, as in many other cell types, clearing of IMPs seems to be one of theintermediate stages in the fusion between regions of exposed phospholipids (Loyter &Lalazar, 1980). In some Protozoa, unusual arrays of IMPs, the rosettes, mark thesites of interaction of granules with the PM; this is thought to precede and triggermembrane fusion (Satir, Schooley & Satir, 1973; Plattner, 1974). Rosettes are neverobserved during the digestive process in Dictyostelium or Acanthamoeba (Bowers1980), nor during the functioning of the contractile vacuole of Leptomonas (Linder &Staehelin, 1978), and it seems most likely that these are related specifically to exocy-tosis rather than to the general membrane fusion process.
The most prominent feature of the PM upon internalization was a marked increasein density of IMPs accompanied by changes in the IMP size distribution on bothfracture faces of the phagosome membrane. These changes are thus rather similar tothose found in Acanthamoeba (Bowers, 1976, 1980) and Tetrahymena (Kitajima &Thompson, 1977). These modifications were observed soon after yeast engulfmentbut they were not linked specifically to the presence of yeast since they also occurredin the membrane of phagosomes containing latex beads and in the digestive vacuolesof cells growing in axenic medium. These changes seem to coincide in time with thenumerous fusions occurring between phagosomes and digestive or autophagic vacu-oles. The observations made with Con A-coated yeast showed, in addition, that thepresence of the lectin on the yeast surface prevented fusion with digestive or auto-phagic vacuoles, since no space and no foreign material could be seen between theyeast and the phagosome membrane. The presence of acid phosphatase proved,however, that fusions had occurred, probably with tiny primary lysosomes, the small
Figs. 19-22. Filipin-sterol complex formation in the PM (Figs. 19, 20) and Ph M(Figs. 21, 22). These complexes appear as large protrusions (black arrowheads),particularly abundant in the EF (Figs. 19, 21). In the PF (Figs. 20 and 22) they areless numerous but many depressions of the same size are visible (white arrows).Bar, o-i /*m.

80 C. Favard-Sereno, M.-A. Ludosky and A. Ryter
size and high membrane curvature of which allowed discrete contacts with thephagosome membrane. Moreover, the same increase in size and density of IMPs wasobserved as that found in untreated yeast phagosomes. This suggests that the changesin IMPs are not related to fusion with digestive or autophagic vacuoles but only withprimary lysosomes. It also leads us to think that a correlation may exist between thesechanges and the presence of hydrolases.
The real cause of this membrane reorganization is, however, difficult to understand.Although no accurate measurements of lysosomal IMPs could be made, due to thesmall size and high curvature of lysosomes, it seems very unlikely that the lysosomalmembrane could provide the bulk of the extra IMPs visualized in the phagosomemembrane.
50 -
E 40 -
8 30-X5.
3 20 -
10-
iPM Ph
50 -
E 40a.
30 -
20-
10 -
Ph
PM
EF PF PF EF
Fig. 23. Filipin—sterol complex formation. A. Mean number per ^m1 of filipin-sterolcomplexes visible in the form of protrusions in the 2 fracture faces of PM and Ph M(Ph).B. Mean number per /1m1 of depressions corresponding to the complexes of thecomplementary half in PM and Ph M (Ph). The bars represent the standard deviation.
Bowers (1980) suggested that the increase in density of IMPs could reflect a changein the protein/lipid ratio and she, in effect, found a lower lipid content in the phago-some membrane (personal communication). She proposed that selective budding ofpure phospholipid vesicle leads to an increase in protein concentration. It is possiblethat the same phenomenon occurs in Dictyostelium but we were unable to detect it.
To our minds, the enhanced density of IMPs could result essentially from theassembling of pre-existing membrane proteins. This reorganization of proteins mightbe correlated with changes in membrane permeability. It is indeed well known thatthe plasma membrane of amoeba is very impermeable as well as that of Dictyostelium(personal observation). It is probable that the products of digestion of the ingestedmaterial can diffuse into the cytoplasm only if the phagosome membrane becomesmore permeable than the plasma membrane. It is interesting to point out that an

Phagocytosis in D. discoideum 81
increase in density of IMPs was also observed in heat-activated spores of Dictyoste-lium (Holh, Buhlmann & Wehrli, 1978), a phenomenon that also requires an increasein membrane permeability.
As sterols are membrane constituents that modulate diffusion and viscosity of lipidwithin the bilayer and could regulate membrane permeability (Papahadjopoulos,Cowden & Kimelberg, 1973), we were interested in testing possible variations of thecontent and distribution of sterols in Dictyostelium membranes during yeast ingestionand digestion, by using digitonin and filipin. Filipin gave more accurate indicationsthan digitonin on sterol concentration and allowed us to conclude that sterols areuniformly distributed in both forming and fully formed phagosome membranes.There is one exception: the IMP-free areas, which correspond to regions wherephagosome and lysosome membranes are apposed, are sterol-depleted as they nevershow any deformations. This micro-heterogeneity in the distribution of sterols inthe plane of the phagosome membrane seems to be related to the temporary membraneremodelling that precedes membrane fusions, as observed in the phospholipid-richmembrane areas of the sperm head that are destined to fuse (Elias et al. 1978; Friend,1980).
Filipin-sterol interaction being equimolar (De Kruijff & Demel, 1974), an appraisalof the relative sterol contents of different membranes can be inferred from thecomparison between the total number of complexes present in a given unit of area ofthese fractured membranes. In this respect, as more protrusions were observed inthe phagosome membrane (EF + PF) than in the PM (EF + PF) per /tm2, we canconclude that the latter is enriched with sterol upon internalization. This seems ingood agreement with the biochemical results of van Vliet et al. (1976) in Entamoeba.The same conclusion was also reported by Charalampous (1979) for neuroblastomacells. This means that if the permeability of the phagosome membrane is higher thanthat of the plasma membrane, this change is not produced by a decrease in sterolcontent.
The higher number of protuberances found on the EF of both PM and phagosomemembrane could indicate a 2-3 times higher level of sterols in the external half ofthese membranes as compared to the inner half; such an asymmetry was also demon-strated for the erythrocyte PM (Zwaal, Roelafsen & Colley, 1973; Fisher, 1976). Twokinds of deformation, i.e. protrusions and depressions, were observed on the fracturefaces of the membrane upon treatment with filipin. The question is, do these deforma-tions arise independently, or are they causally related, the depressions being com-plementary aspects of the protrusions situated in the complementary half-membrane?The relative diameters of protrusions (15-25 nm) and depressions (15 run at most)suggest that the 2 types of deformation represent complementary aspects of thecomplexes, situated in register in the 2 membrane leaflets. The data obtained from thestudy of cholesterol in the erythrocyte PM also favour this hypothesis. In filipin-treated erythrocytes, the EF of the PM displays numerous protrusions and the PF, incontrast, mostly depressions (Verkleij et al. 1973; Kitajima et al. 1976). This couldindicate that the sterol-filipin complexes are not lying within the hydrophobic coreparallel to the membrane plane (according to one of the models proposed by De

82 C. Favard-Sereno, M.-A. Ludosky and A. Ryter
Kruijff & Demel, 1974), but rather are oriented perpendicular to the plane of thebilayer (as the complexes formed with other polyene antibiotics; Finkelstein & Holz,X973)-
Another puzzling aspect of the filipin-treated membranes is the disparity betweenthe density of protrusions (number//im2) observed on the EF and the density ofdepressions visible in the PF, on one hand; and on the other hand, the similaritybetween protrusion density on the PF and depression density on the EF. One temptinginterpretation refers to transbilayer asymmetry of phospholipid and/or fatty acid, ashas been found in many biological membranes (see Malhotra, 1980, for a review). Inthese terms our observations could indicate that the external, as compared to theinternal, membrane leaflet of the PM and phagosome membrane in Dictyosteliumcontains lipid species that confer on it more plasticity.
Many thanks to R. Daty for her excellent technical assistance. This work was supported bythe Centre National de la Recherche Scientifique (laboratoire associi 269) and the D616gationGe'ne'rale a la Recherche Scientifique et Technique (contract 80.7.0363).
REFERENCESALLEN, R. D. (1976). Freeze fracture evidence for intramembrane changes accompanying
membrane recycling in Paramecium. Cytobiologie 12, 254-273.BATZ, W. & WUNDERLICH, F. (1976). Structural transformation of the phagosomal membrane
in Tetrahymena cells endocytosing latex beads. Arch. Microbiol. 109, 215-220.BOWERS, B. (1976). Changes in plasma membrane structure after internalization in phagosomes.
J. Cell Biol. 70, 326.BOWERS, B. (1980). A morphological study of plasma and phagosome membranes during
endocvtosU in Acanthamoeba. J. Cell Biol. 84, 246-260.CHARALAMPOUS, F. C. (1979). Levels and distributions of phospholipids and cholesterol in
the plasma membrane of neuroblastoma cells. Biochim. biophys. Acta 556, 38-51.DE BRUYN, P. P. H., MICHELSON, S. & BECKER, R. P. (1978). Non random distribution of
sialic acid over the cell surface of bristle-coated endocytic vesicles of the sinusoidal endo-thelium. J. Cell Biol. 78, 379-389.
DE CHASTELLIER, C. & RYTER, A. (1977). Changes of the cell surface and of the digestiveapparatus of Dictyostelium discoideum during the starvation period triggering aggregation.J. Cell Biol. 75, 218-236.
DE RRUIJFF, B. & DEMEL, R. A. (1974). Polyene antibiotic-sterol interactions in membranes ofAcholeplasma laidlawii cells and lecithin liposomes. Biochim. biophys. Acta 339, 57-70.
EDELSON, P. J. & COHN, Z. A. (1974). Effects of concanavalin A on mouse peritoneal macro-phages. J. exp. Med. 140, 1364-1368.
ELIAS, P. M., GOERKE, J. & FRIEND, D. S. (1978). Freeze-fracture identification of sterol-digitonin complexes in cell and liposome membranes. J. Cell Biol. 78, 577-596.
FAVARD-SERENO, C. & LIVROZET, M. (1979). Plasma membrane structural changes correlatedwith the acquisition of aggregation competence in Dictyostelium discoideum. Biologie cell. 35,45-54-
FINKELSTEIN, A. & HOLZ, R. (1973). Polyene antiobiotics. In Membranes: Lipid Bilayers andAntibiotics, vol. 2 (ed. G. Eisenman), pp. 377-408. New York: M. Dekker.
FISHER, K. A. (1976). Analysis of membrane halves: cholesterol. Proc. natn. Acad. Sci. U.S.A.73. 173-177-
FRIEND, D. S. (1980). Freeze-fracture alterations in Guinea pig sperm membranes precedinggamete fusion. In Membrane-Membrane Interactions (ed. N. B. Gilula), pp. 153-165. NewYork: Raven Press.

Phagocytosis in D. discoideum 83
GONZALES-NORIEGA, A., GRUBB, J. H., TALKAD, V. & SLY, W. S. (1980). Chloroquine inhibitslysosomal enzyme pinocytosis and enhances lysosomal enzyme secretion by impairingreceptor recycling. J. Cell Biol. 85, 839-852.
HOHL, H. R., BULHMANN, M. & WEHRLI, E. (1978). Plasma membrane alterations as a resultof heat activation in Dictyostelium spores. Arch. Microbiol. 116, 239-244.
KITAJIMA, Y., SEKIYA, T. & NOZAWA, Y. (1976). Freeze-fracture ultrastructural alterationsinduced by filipin, pimaricin, nystatin and amphotericin B in the plasma membranes ofEpidermophyton, Saccharomyces and red blood cells. A proposal of models for polyene-ergosterol complex-induced membrane lesions. Biochem. biophys. Acta 455, 452-465.
KITAJIMA, Y. & THOMPSON, G. A. Jr (1977). Differentiation of food vacuolar membranesduring endocytosis in Tetrahymena. J. Cell Biol. 75, 436-445.
LINDEB, J. C. & STAEHELIN, L. A. (1978). Membrane fusion and specialized membranes ofthe contractile vacuole apparatus. J. Cell Biol. 79, 231a.
LOYTER, A. & LALAZAR, A. (1980). Induction of membrane fusion in human erythrocyteghosts: involvement of spectrin in the fusion process. In Membrane-Membrane Interactions(ed. N. B. Gilula), pp. 11-26. New York: Raven Press.
MALHOTRA, S. K. (1980). Organization, composition and biogenesis of animal cell membranes.In Membrane Structure and Function, vol. 1 (ed. E. E. Bittar), pp. 1-72. London, New York:Wiley Interscience.
MOOR, H. & MUHLETHALER, K. (1963). Fine structure in frozen-etched yeast cells. J. Cell Biol.17, 609-628.
MOORE, P. L., BANK, H. L., BRISSIE, N. T. & SPICER, S. S. (1978). Phagocytosis of bacteriaby polymorphonuclear leukocytes. J. Cell Biol. 76, 158-174.
NACHMAN, R. L., FERRIS, B. & HrascH, J. G. (1971). Macrophage plasma membrane. J. exp.Med. 133, 807-820.
OHKUMA, S. & POOLE, B. (1978). Fluorescence probe measurement of the intralysosomal pHin living cells and the perturbation of pH by various agents. Proc. natn. Acad. Sci. U.S.A. 75,3327-333I-
PAPAHADJOPOULOS, D., COWDEN, M. & KIMELBERG, H. (1973). Role of cholesterol in membranes.Effects of phospholipid-protein interactions, membrane permeability and enzymatic activity.Biochim. biophys. Acta 330, 8-26.
PARISH, W. & PELLI, C. (1974). Alkaline phosphatase of Dictyostelium discoideum: cell surfacelocation and colchicine effects on internalization during phagocytosis. FEBS Lett. 48,293-296.
PLATTNER, H. (1974). Intramembranous changes on cationophore-triggered exocytosis inParamecium. Nature, Lond. 252, 722-724.
PRYZWANSKY, K. B., MACRAE, E. K., SPITZNAGEL, J. K. & COONEY, M. H. (1979). Early de-granulation of human neutrophils: immunocytochemical studies of surface and intracellularphagocytic events. Cell 15, 1025-1033.
RYTER, A. & DE CHASTELLIEH, C. (1977). Morphometric and cytochemical studies of Dictyost-elium discoideum in vegetative phase. Digestive system and membrane turnover. J. Cell Biol.75, 200-217.
RYTER, A. & HELLIO, R. (1980). Electron-microscope study of Dictyostelium discoideum plasmamembrane and its modifications during and after phagocytosis. J. Cell Sci. 41, 75-88.
SATIR, B., SCHOOLEY, C. & SATIR, P. (1973). Membrane fusion in a model system. Mucocystsecretion in Tetrahymena. J. Cell Biol. 56, 153-176.
SEGAL, A. W., DORLING, J. & COADE, S. (1980). Kinetics of fusion of the cytoplasmicgranules with phagocytic vacuoles in human polymorphonuclear leukocytes. J. Cell Biol. 85,42-59-
SINGER, S. J. (1971). The molecular organization of biological membranes. In MembraneStructure and Function (ed. L. I. Rothfield), pp. 146-222. New York, London: AcademicPress.
SMOLEN, J. E. & SHOHET, S. B. (1974). Remodeling of granulocyte membrane fatty acids duringphagocytosis. J. din. Invest. 53, 726.
THORNE, K. J. I., OLIVER, R. C. & GLAUERT, A. M. (1980). Eosinophil interaction with anti-body-coated, non-phagocytosable surfaces: changes in cell surface proteins. J. Cell Sci. 42,367-378.

84 C. Favard-Sereno, M.-A. Ludosky and A. Ryter
TIETZE, C, SCHLESINGER, P. & STAHL, P. (1980). Chloroquine and ammonium ion inhibitreceptor-mediated endocytosis of mannose-glycoconjugates by macrophages: apparentinhibition of receptor recycling. Biochem. biophys. Res. Comrmtn. 93, 1-8.
TILLACK, T. W. & KINSKY, S. C. (1973). A freeze-etch study of the effects of filipin on lipo-somes and human erythrocyte membranes. Biochim. biophys. Acta. 323, 43-54.
UKENA, T. E. & BERLIN, R. D. (1972). Effect of colchicine and vinblastine on the topographicalseparation of membrane functions. J. exp. Med. 136, 1-7.
VAN VLIET, H. H. D. M., SPIES, F., LINNEMANS, W. A. M., KLEPKE, A. OP DEN KAMP, J. A. F.& VAN DEENEN, L. L.M. (1976). Isolation and characterization of subcellular membranes ofEntamoeba invadens. J. Cell Biol. 71, 357-369.
VERKLEIJ, A. J., DE KRUIJFF, B., GERRITSEN, W. F., DEMBL, R. A., VAN DEENEN, L. L. M. &VERVERGAERT, P. H. J. (1973). Freeze-etch electron microscopy of erythrocytes, Achole-plasvta laidlatcii cells and liposomal membranes after the action of filipin and amphotericin B.Biochim. Biophys. Acta 291, 577-581.
VINCENZI, F. F. & LARSEN, F. L. (1980). The plasma membrane calcium pump: regulation bya soluble Ca1+ binding protein. Fed. Proc. 39, 2427-2431.
WIBO, M. & POOLE, B. (1974). Protein degradation in cultured cells. J. Cell Biol. 63, 430-440.ZWAAL, R. F. A., ROELAFSEN, B. & COLLEY, C. M. (1973). Localization of red cell membrane
constituents. Biochim. biophys. Acta 300, 159-182.
{Received 6 January 1981)