frozen tissue sections as an experimental system to reveal specific
TRANSCRIPT

Proc. NatL Acad. Sci. USAVol. 79, pp. 5562-5566, September 1982Cell Biology
Frozen tissue sections as an experimental system to reveal specificbinding sites for the regulatory subunit of type II cAMP-dependent protein kinase in neurons
(immunohistochemistry/microtubule associated protein/dendrites/protein phosphorylation)
P. MILLER*, U. WALTERt, W. E. THEURKAUFt, R. B. VALLEEt, AND P. DE CAMILLI*§*Section of Cell Biology, Yale University, School of Medicine, New Haven, Connecticut 06510; tDepartments of Physiological Chemistry, Wurzburg, FederalRepublic of Germany; and *Cell Biology Group, Worcester Foundation for Experimental Biology, Shrewsbury, Massachusetts 01545
Communicated by George E. Palade, May 28, 1982
ABSTRACT Specific binding sites for the regulatory subunitof type II cAMP-dependent protein Idnase (RI,) were revealed inneurons by an immunohistochemical approach. Fixed frozen sec-tions of several regions of the rat central nervous system were in-cubated in the presence of bovine RI,. Bound bovine RH1 was sub-sequently detected by an immunofluorescence procedure usingantibodies that recognize bovine but not rat RI,. The results in-dicate that RI, binds with high affinity to neurons. Binding is prom-inent in dendrites and almost undetectable in axons and axon ter-minals. The morphological distribution of the R]I binding sites isalmost identical to that of microtubule-associated protein 2 (MAP2) immunoreactivity. Preadsorption ofRI, with a MAP preparationhighly enriched in MAP 2 completely abolished binding of RI, totissue sections, suggesting that the binding is mediated by MAP2. Our results indicate that frozen sections of fixed tissues are asuitable experimental system for study of specific interactions ofcellular macromolecules at a morphological level.
Following the discovery of neurotransmitter-sensitive adenyl-ate cyclases (1), much evidence that cAMP plays an importantmodulatory role in neurons has accumulated (2). cAMP-depen-dent protein kinases, the target enzymes for cAMP, are presentat high concentrations in all regions of the central nervous sys-tem (3, 4). At the subcellular level, however, they seem to beat least partially compartmentalized because such kinase activ-ities have been demonstrated in both soluble and particulatesubfractions (3-9). One of the possible mechanisms of com-partmentalization is a high-affinity interaction of their subunitswith partner molecules localized on structural cellular compo-nents. An association of type II cAMP-dependent protein ki-nases with specific cytoplasmic proteins, mediated by its reg-ulatory subunit, RI,, has recently been reported (10-12).
In the present work we used an immunohistochemical ap-proach to study the existence and distribution of high-affinitybinding sites for RI, in the central nervous system ofthe rat. Forthis purpose we took advantage of the antigenic dissimilaritybetween bovine and rat RI, (13). Antibodies raised against bo-vine RII did not stain any cellular element when used to labelendogenous RI, in rat brain by immunofluorescence. It there-fore has been possible to study binding sites for RI, by usingbovine RI, as a probe followed by the anti-bovine RI, antibodiestolocalize the bound exogenous RI,. The starting premise ofthiswork was that, in spite of the prominent antigenic differencesbetween bovine and rat RI,, physiologically important bindingsites on these two molecules for other cytoplasmic proteinsmight be similar.
Results presented here indicate that RI, binds specifically toneurons and that, in neurons, binding is most prominent indendrites. They also suggest that such binding is probably me-diated by microtubule-associated protein 2 (MAP 2), a highmolecular weight protein that copurifies with brain microtu-bules (14, 15) and is a prominent substrate for cAMP-dependentprotein kinase.
MATERIALS AND METHODSPreparation of Antigens and Antibodies. Bovine heart RI,
was prepared and used to immunize rabbits as described (9).Some of the characteristics of the antiserum thus obtained aredescribed in refs. 9, 13, and 16. Antibodies specific for RI, wereprepared from the serum by affinity-chromatography on bovineheart RI, covalently bound to Sepharose beads (Pharmacia,Uppsala, Sweden). Specific antibodies were eluted from thebeads by pH 2.3 buffer.A MAP fraction highly enriched in MAP 2 and devoid of
MAP 1 (heat-stable MAP fraction) was prepared by exposureoffour times-cycled microtubules (from bovine cerebral cortex)to heat (17-19). MAP 2 represented 75% of the total proteincontent of the fraction (unpublished data). For the preparationof a specific antiserum directed, against MAP 2, peptides con-tained in the crudeMAP fraction were separated by NaDodSO4/polyacrylamide slab gel electrophoresis. The MAP 2-containingregion of the gel was then excised and injected into rabbits.When tested by gel radioimmunolabeling (20) against all thepeptides contained in a homogenate of rat brain, the antiserumrecognized only the MAP 2 doublet (unpublished data).
Gel Electrophoresis and Gel Radioimmunolabeling. Na-DodSO4/polyacrylamide slab gel electrophoresis of homoge-nates of rat and bovine brains was performed as described (20).Gels were radioimmunolabeled as described (20), with the ex-ception that affinity-purified antibodies directed against RI, in-stead of whole serum were used at the primary step of theprocedure.
Immunohistochemical Procedures. Tissues were fixed andfrozen sections were prepared as reported (21). In all cases, tis-sues were frozen by quick immersion in isopentane chilled withliquid nitrogen. Immunohistochemical staining was performedas described (21); whole serum (1: 20-1:50 dilutions) or purifiedIgGs (0.4 mg/ml) were used as the first step and rhodaminated
Abbreviations: MAP, microtubule associated protein; RI and RI,, reg-ulatory subunits of the type I and type II cAMP-dependent proteinkinases; C, catalytic subunit of cAMP-dependent protein Idnases.§ Present address: Consiglio Nazionale delle Ricerche, Center of Cy-topharmacology and Dept. of Pharmacology, UniversitA di Milano,via Vanvitelli 32, 20129 Milano, Italy.
5562
The publication costs ofthis article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertise-ment" in accordance with 18 U. S. C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 79 (1982) 5563
goat anti-rabbit IgGs (Cappel Laboratories, Cochranville, PA)were used as the second step. To study binding of RI, to thetissue, some sections were incubated for 30 min with 0.1 MTris'HCI buffer and then were overlaid with 100 Ald of CEAbuffer [chicken egg albumin (50 mg/ml)/0.45 M NaCl/0.3%Triton X-100/40 mM phosphate buffer] containing 6 ,ug of bo-vine heart RI,. After 3-hr incubation at room temperature, thesections were thoroughly washed with Triton buffer (21) to re-move the unbound RI, and finally immunostained for bovineRI1. In some cases the bound RI, was crosslinked to the tissueby overlaying the section for 15 min with 2% glutaraldehyde/120 mM phosphate before immunostaining. Unreacted glutar-aldehyde was then quenched by an incubation with 1% Na bo-rohydride before the antibody steps. In the experiments carriedout to study the nature of the binding of RI, to the tissue, RI,(6 ,ug) was applied to the sections in 100 ,ud ofCEA buffer con-taining either 350 tug of the heat-stable MAP fraction or 5 mMcAMP. The CEA buffer samples containing the RI,/MAPS andthe RI/cAMP mixtures had been preincubated for 30 min be-fore being overlaid on sections.
All protein-containing solutions were cleared by centrifu-gation in an Eppendorf Microfuge before being applied to tissuesections.
RESULTSImmunological Dissimilarity Between Rat and Bovine RII.
When, affinity-purified antibodies raised against bovine RI,were used to immunolabel gels of bovine brain homogenate,only the doublet of RI, (9) was labeled (Fig. 1). Labeling of RII,however, was practically undetectable when rat brain homog-
_ORIGIN
FIG. 1. Autoradiograph ofpolyacrylamide gel showingspecificity of affinity-purifiedantibodies directed against bo-vine RI,. Peptides contained ina bovine brain homogenate (150,ug) were separated by Na-DodSO4 electrophoresis in an8% gel. The gel was subse-quently radioimmunolabeled byan indirect technique (20) uAingaffinity-purified antibodies di-rectedagainstbovineR1I(15 ,ug/ml) as the first step and 125I-la-
r. beled protein A as the secondstep. The arrow indicates thedirection of protein migration.
A B C
]R1I__m.
FIG. 2. Autoradiograph of an 8% NaDodSO4/polyacrylamide gelshowing the immunological dissimilarity between rat and bovine RH1.Purified bovine heart RI, was applied to lane A (0.2 /Lg), bovine brainhomogenate (150 ,ug) to lane B, and rat brain homogenate (150 u&g) tolane C. After electrophoresis the gel was radioimmunolabeled by af-finity-purified antibodies directed against bovine RH1 as described inFig. 1. The Re-containing region of the autoradiograph is shown. Thearrow indicates the direction of protein migration.
enate was used instead of bovine brain homogenate (Fig. 2).Similar results, indicating poor imunological crossreactivity be-tween bovine and rat R11, have been obtained by immunopre-cipitation (13).
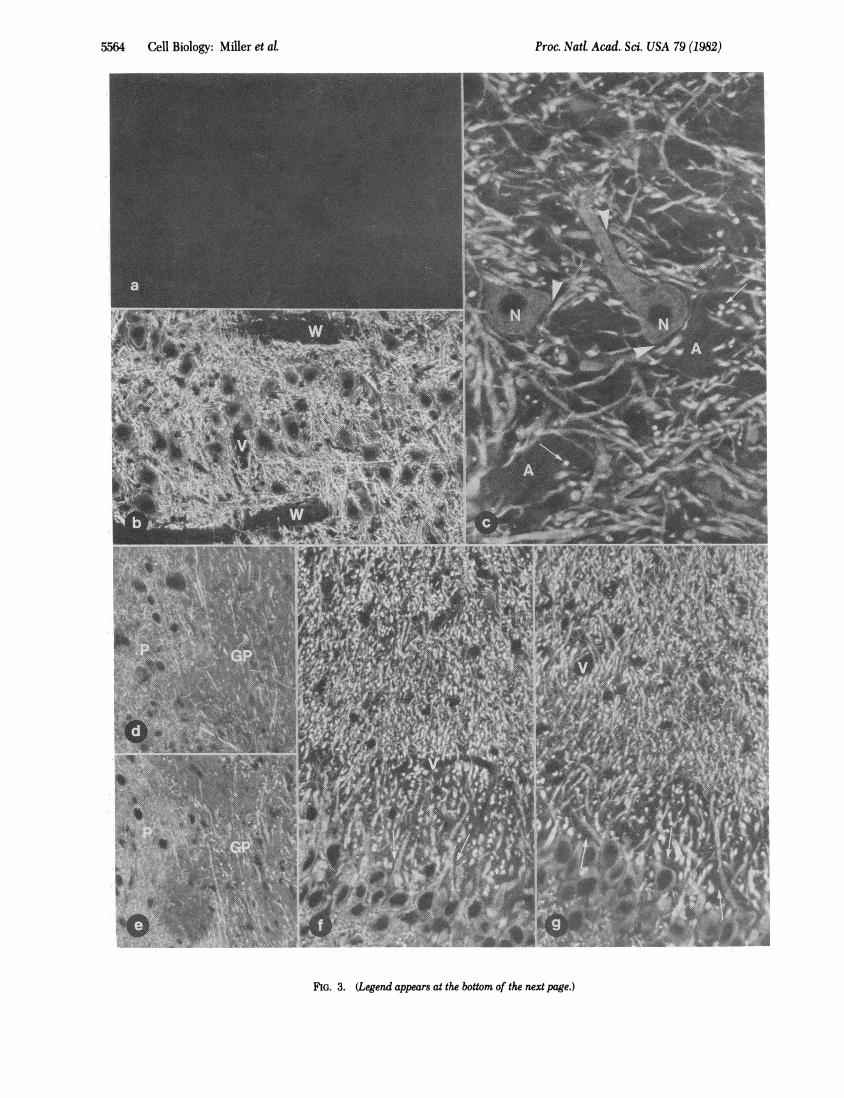
Binding of RI, to Tissue Sections. When the affinity-purifiedantibodies directed against bovine RI, were used to stain frozensections of formaldehyde-fixed rat brain by an indirect immu-norhodamine procedure, no specific staining was detected (Fig.3a). Results were indistinguishable from those obtained whenthe anti-RI, antibodies were replaced by preimmune serum ornonimmune IgGs. This was not surprising in view of the lowcrossreactivity between rat and bovine RI, mentioned above.However, a bright fluorescence was observed in sections im-munostained with affinity-purified antibovine RI, antibodieswhen the immunohistochemical procedure was preceded byincubation of the sections with bovine RI, (Figs. 3 and 4 a andd). This fluorescence had a typical pattern: (a) it was presentin all regions of the gray matter; (b) it was specifically localizedin neurons and outlined them; glial cells were unstained; (c)inside neurons it was unevenly distributed-fluorescence gen-erally was bright in dendrites, present but to a lower and vari-able extent in perikarya, absent from nuclei, and undetectablein axons and axon terminals. Staining intensity was variable indifferent regions of the central nervous system. For instance,staining of dendrites was bright in the gray matter of the brainstem (Figs. 3 b and c and 4d) and the spinal cord but weak inthe cerebellar cortex (not shown).
Because our antibodies directed against RI, did not recognizeendogenous RI, in rat tissue sections, the staining describedabove has to be attributed to the exogenous bound bovine RI,.Such binding of RI, is specific and of high affinity: it occurs ina buffer containing high protein and high salt concentrations andit is resistant to extensive washing of the sections.
Binding of RH1 to the Tissue Is Not Mediated by cAMP orby the Catalytic Subunit of cAMP-Dependent Protein Kinase(C). The morphological results indicate that high-affinity bind-ing sites for heterologous RI, are present in neurons. BecauseRI, is known to bind with high affinity to cAMP and to C (22),it was important to explore whether either one of these mole-cules was responsible for such binding. [It is known that RI, andC of different animal species can assemble to form hybrid holo-enzyme molecules (23). ] We tested therefore whether bindingof RI, could still occur in the presence of a high concentrationofcAMP. If RI, binds to cAMP present in thetissue, then highlevels ofcAMP in the incubation medium should prevent bind-ing. IfRI, binds to tissue C, cAMP should again prevent bindingbecause cAMP promotes the dissociation of RI, from C (22).
Cell Biology: MiUer et aL
Proc. Natl. Acad. Sci. USA 79 (1982)
4
i
I. ..k
i. ..%.'-.:
FIG. 3. (Legend appears at the bottom of the next page.)
5564 Cell Biology:'Miller et al.
F.1 ..f1
I I
4t,Aki i.w'ok
kI
r ,-A:

Proc. Natd Acad. Sci. USA 79 (1982) 5565
FIG. 4. Immunofluorescent staining of fixed frozen sections of rat brain, showing blocking of the RIa binding by aMAP fraction highly enrichedin MAP 2. (a-c) Coronal sections of cerebral cortex. (a) Section incubated in the presence of bovine R11. The exogenous bound bovine R11 was sub-sequently detected by immunofluorescence using anti-bovine RH1 antibodies. Note the bright fluorescence of bundles of apical dendrites (D) of py-ramidal cells, the minor fluorescence of other randomly oriented dendrites, and the pale staining of the cytoplasm of neuronal perikarya (arrows).(b) Section immunostained for MAP 2. The pattern of fluorescence is identical to the pattern in a. (c) Section processed as the one in a except thatRHwas applied to the tissue in the presence of an excess of the MAPfraction highly enriched in MAP 2. No stain is visible. (d and e) Adjacent sectionsof the locus ceruleus processed as described in a and c, respectively. The meshwork of thin dendrites is fluorescent in d but not in e. (a-c, X 160;d and e, x280.)
Specific binding of bovine RI, to rat tissue sections was unaf-fected by the presence of a high concentration ofcAMP in theincubation medium (Fig. 3 d and e).The Morphological Distribution of Binding Sites for RI, Is
Similar to the Morphological Distribution of MAP 2 Immu-noreactivity. It has recently been reported that high molecularweight MAPs are highly compartmentalized in brain, beingmostly concentrated in neuronal dendrites (24). Analogous re-sults have been obtained with an antiserum specific for MAP2 (unpublished data). When the morphological distributions ofRI, binding sites and of MAP 2 immunoreactivity were com-pared, an almost identical pattern could be recognized in all ofthe several regions of the central nervous system we have ex-amined, which include hippocampus (Fig. 3f and g), cerebralcortex (Fig. 4 a and b), thalamus, cerebellum, several brainstemregions, and spinal cord (not shown). In general the outline ofperikarya and dendrites in sections stained for MAP 2 is sharper
due to a higher signal/background ratio, but the relative dis-tributions of staining among different structures are similar.
Preadsorption of RI, with a MAP Fraction Highly Enrichedin MAP 2 Abolishes Binding of RI, to Tissue Sections. The sim-ilarity of the morphological distribution of MAP 2 immuno-reactivity and ofRH1 binding sites is so striking as to suggest thatMAP 2 might be the binding site for RI, in frozen sections ofneuronal tissue. This is consistent with the recent finding thata high proportion (about one-third) of the cytosolic cAMP-de-pendent protein kinase activity in brain is specifically associatedwith MAP 2, such association being mediated by RI, (10, 11).
If MAP 2 is the binding site for exogenous RI, in tissue sec-tions, previous saturation of the MAP 2 binding sites on RI,should prevent binding. To test if this was the case, bovine RI,was applied to tissue sections in the presence of an excess of aMAP preparation (heat-stable MAP fraction) containing MAP2 as the predominant species and not containing MAP 1. The
FIG. 3 (on preceding page). Immunofluorescence staining of fixed frozen sections of rat brain showing the distribution of binding sites for ex-ogenous RH and of endogenous MAP 2 immunoreactivity. Immunoreactive material appears white. (a) Section of a brainstem motor nucleus im-munostained with affinity-purified antibodies directed against bovine RH by an indirect immunorhodamine procedure. No specific stain is visible.(b-f) Sections processed as described for a, after a preincubation in the presence of purified bovine heart RH. (b) Section adjacent to the one shownin a. The network of neuronal dendrites is intensely fluorescent. A pale fluorescence is present in the cytoplasm of perikarya; cell nuclei are com-pletely negative. Also unstained are two tracts of white matter present in the field (W). V, blood vessel. (c) High magnification of a brainstem motornucleus. The perikarya of two motor neurons (N) are visible in the field. Note the bright fluorescence of dendrites and the absence of staining onaxonal bundles (A). Note also the absence of fluorescence at sites that are knowntobe occupiedby the axosomatic and axodendritic terminals whichcover the surface of motor neurons (arrowheads). Arrows point at cross-sectioned motor neuron dendrites. (d) Coronal sections of the basal ganglia.The different dendritic pattern in the putamen (P) and in the globulus pallidus (GP) can be seen. (e) Section of a brain region similar to the oneshown in d. In this case, RI had been applied to the section in the presence of 5mM cAMP. No difference can be observed between the fluorescencepatterns in d and e. (f) Coronal section of the CA4 region of the hippocampus. The perikarya and the apical dendrites of the pyramidal cells canbe seen in the picture. Cross-sectioned dendrites appear as small white dots. Fluorescence is more prominent in dendrites than in the cytoplasmof the perikarya. Note the dark spaces around the proximal portions of the apical dendrites (arrows). These spaces are occupied by the large mossyfiber terminals which are not stained. V, blood vessels. (g) Section adjacent to the one shown in f but immunostained with an antiserum directedagainst MAP 2. Note the remarkable similarity of fluorescent stain in f and g. (a and b, x 192; c x460; d and e, x268; fand g, X358.)
Cell Biology: Miller et d

Proc. Natd Acad. Sci. USA 79 (1982)
molar excess ofMAP 2 over the RI, monomer was approximately8-to 9-fold. When the sections treated with the mixture ofMAPand RI, were subsequently processed for anti-bovine RI, im-munofluorescence to reveal bound RI,, no specific fluorescencewas observed, indicating that no binding of RI, had occurred(Fig. 4 a and c vs. d and e).
DISCUSSIONThe results reported in this paper point to the possibility ofusing a procedure based on immunocytochemistry to study spe-
cific interactions of cellular macromolecules at a morphologicallevel. In spite of the chemical and physical manipulations re-
quired to obtain sections retaining good cell and tissue mor-
phology, at least some proteins of the tissue maintain their abil-ity to interact specifically with partner molecules.
In this study we found specific binding of RI, to componentsofthe nervous system. We also found evidence that this bindingis mediated by MAPs, most likely by MAP 2. This is suggestedby the similar if not identical distributions of MAP 2 immuno-reactivity and of RI, binding sites and by the ability of a prep-
aration of MAPs highly enriched in MAP 2 (and devoid of MAP1) to abolish the binding. Our results are in agreement with thefinding that a major fraction of brain type II cAMP-dependentprotein kinase is associated with MAP 2(10, 11). The latter stud-ies have indicated that the kinase is bound via its regulatorysubunit, RI,, to a fragment of MAP 2 representing a projectionon the microtubule surface. Our results imply that in vivo a largefraction of RI, binding sites on MAP 2 molecules are unoccupiedbecause it is unlikely that, after fixation, bovine RI, might dis-place previously bound rat RI,.MAP 2 is a prominent substrate for cAMP-dependent protein
kinase (14, 15, 19). Its interaction with the regulatory subunitof the enzyme might be a mechanism aimed at achieving a highconcentration of the catalytic subunit in close proximity to itssubstrate.The binding of bovine RI, to rat brain and the observation
that bovine proteins contained in the MAP fraction competewith the rat tissue binding sites indicate that the domain of themolecule involved in the binding is specifically conserved inevolution, as is the case for the cAMP and C binding sites (23),in spite of the marked antigenic dissimilarity of the RI, mole-cule in the two animal species. This further supports the ideaof a physiological significance of the specific binding of RI, totissue sections.
Our results do not indicate that a protein in neuronal den-drites is the only binding site for RI, in the nervous system or
that the distribution of the endogenous RI, overlaps that of thebinding sites revealed by the present study. Other binding sitesmight not withstand fixation, or might require homologous RI,to be detected, or might already be saturated by endogenousRI,, and therefore not be available for the exogenous subunit.Also, binding sites of low affinity, not detectable by our ex-
perimental protocol, might be important physiologically in cellscontaining high concentrations of RI,. Finally, it should bestressed that in our study we used an RI, subunit purified fromheart. It is well known that differences exist between brain andheart RI, (8, 9) and that, when analyzed by NaDodSO4 gel elec-trophoresis, brain RI, appears to consist ofa heterogeneous pop-
ulation of peptides of similar molecular weight (9) (Figs. 1 and
2). It is possible that the ability to bind to neuronal dendritesis peculiar to a subclass of RI, molecules common to brain andheart. Other RI, subclasses, specific to brain, might bind toother structures. This question will be clarified by studying thespecific interaction with brain tissue of RI, purified from brain.
It will be interesting in the future to study whether specificbinding sites exist for RI, the regulatory subunit of type I cAMP-dependent protein kinase, and whether these binding sites aredistinct from those for RI,. In fact, binding sites specific foreither RI or RI, might be the basis for the functional differencebetween the two regulatory subunits. They might mediate theintracellular compartmentalization of the catalytic subunit indistinct supramolecular structures.We thank P. Greengard for support throughout the work. We also
thank N. Borgese and J. Meldolesi for critically reading the manuscriptand S. M. Harris for help in preparing the illustrations. This work wassupported by a Muscular Dystrophy Association grant to P.D.C., byNational Institutes of Health Grant BM 26701 to R.B.V., and by a grantfrom the Deutsche Forschungsgemeinschaft to U.W.1. Kebabian, J. W. & Greengard, P. (1971) Science 174, 1346-1349.2. Greengard, P. (1978) Cyclic Nucleotides, Phosphorylated Pro-
teins and Neuronal Function (Raven, New York).3. Hoffman, F., Bechtel, P. J. & Krebs, E. G. (1977)J. Biol Chem.
252, 1441-1447.4. Walter, U., Kanof, P., Schulman, H. & Greengard, P. (1978) 1.
Biol Chem. 253, 6275-6280.5. Maeno, H., Johnson, E. M. & Greengard, P. (1971) J. Biol.
Chem. 246, 7303-7310.6. Rubin, C. S., Erlichman, J. & Rosen, 0. M. (1972)J. Biol Chemn.
247, 6135-6139.7. Corbin, J. D., Sugden, P. H., Lincoln, T. M. & Keely, S. L.
(1977) J. Biol Chean. 252, 3854-3861.8. Rubin, C. S., Rangel-Aldao, R., Sarkan, D., Erlichman, J. &
Fleischer, N. (1979) J. Biol Chem. 254, 3797-3805.9. Lohmann, S. M., Walter, U. & Greengard, P. (1980) J. Biol.
Chem. 255, 9985-9992.10. Vallee, R. B., Di Bartolomeis, M. J. & Theurkauf, W. E. (1981)
J. Cell Biol 90, 568-576.11. Theurkauf, W. E. & Vallee, R. B. (1982)J. Biol. Chemn., in press.12. Hathaway, D. R., Aldestein, R. S. & Klee, C. B. (1981)J. Biol
Chem. 256, 8183-8189.13. Walter, U., De Camilli, P., Lohmann, S. M., Miller, P. &
Greengard, P. (1981) in Protein Phosphorylation, Cold SpringHarbor Conferences on Cell Proliferation, eds. Rosen, 0. M. &Krebs, E. G. (Cold Spring Harbor Laboratory, Cold Spring Har-bor, NY), Vol. 8, pp. 141-157.
14. Sloboda, R. D., Rudolph, S. A., Rosenbaum, J. L. & Greengard,P. (1975) Proc. Nati Acad. Sci. USA 72, 177-181.
15. Vallee, R. (1980) Proc. Nati Acad. Sci. USA 77, 3206-3210.16. Walter, U., Miller, P., Wilson, F., Menkes, D. & Greengard, P.
(1980) J. Biol Chean. 255, 3757-3762.17. Fellous, A., Francon, J., Lennon, A. M. & Nunez, J. (1977) Eur.
J. Biochemn. 78, 167-174.18. Herzog, W. & Weber, K. (1978) Eur. J. Biochein. 92, 1-8.19. Kim, H., Binder, L. I. & Rosenbaum, J. L. (1979)J. Cell Biol 80,
266-276.20. De Camilli, P., Ueda, T., Bloom, F. E., Battenberg, E. &
Greengard, P. (1979) Proc. Nati. Acad. Sci. USA 76, 5977-5981.21. Lohmann, S. M., Walter, U., Miller, P. E., Greengard, P. & De
Camilli, P. (1981) Proc. Nati Acad. Sci. USA 78, 653-657.22. Krebs, E. G. & Beavo, J. A. (1979) Annu. Rev. Biochein. 48,
923-939.23. Yamamura, H., Kumon, A. & Nishizuka, Y. (1971)J. Biol Chein.
246, 1544-1547.24. Matus, A., Bernhardt, R. & Hugh-Jones, T. (1981) Proc. Nati.
Acad. Sci. USA 78, 3010-3014.
5566 Cell Biology: Miller et aL