gelsy torres-oviedo and lena h. ting - georgia …¬c muscle synergies in human balance control are...
TRANSCRIPT

103:3084-3098, 2010. First published Apr 14, 2010; doi:10.1152/jn.00960.2009 J NeurophysiolGelsy Torres-Oviedo and Lena H. Ting
You might find this additional information useful...
74 articles, 46 of which you can access free at: This article cites http://jn.physiology.org/cgi/content/full/103/6/3084#BIBL
including high-resolution figures, can be found at: Updated information and services http://jn.physiology.org/cgi/content/full/103/6/3084
can be found at: Journal of Neurophysiologyabout Additional material and information http://www.the-aps.org/publications/jn
This information is current as of June 14, 2010 .
http://www.the-aps.org/.American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the
publishes original articles on the function of the nervous system. It is published 12 times a yearJournal of Neurophysiology
on June 14, 2010 jn.physiology.org
Dow
nloaded from

Subject-Specific Muscle Synergies in Human Balance Control Are ConsistentAcross Different Biomechanical Contexts
Gelsy Torres-Oviedo and Lena H. TingThe Wallace H. Coulter Department of Biomedical Engineering, at Georgia Tech and Emory University, Atlanta, Georgia
Submitted 2 November 2009; accepted in final form 7 April 2010
Torres-Oviedo G, Ting LH. Subject-specific muscle synergies inhuman balance control are consistent across different biomechanicalcontexts. J Neurophysiol 103: 3084–3098, 2010. First published April14, 2010; doi:10.1152/jn.00960.2009. The musculoskeletal redun-dancy of the body provides multiple solutions for performing motortasks. We have proposed that the nervous system solves this uncon-strained problem through the recruitment of motor modules or func-tional muscle synergies that map motor intention to action. Consistentwith this hypothesis, we showed that trial-by-trial variations in muscleactivation for multidirectional balance control in humans were con-strained by a small set of muscle synergies. However, apparent musclesynergy structures could arise from characteristic patterns of sensoryinput resulting from perturbations or from low-dimensional optimalmotor solutions. Here we studied electromyographic (EMG) re-sponses for balance control across a range of biomechanical contexts,which alter not only the sensory inflow generated by postural pertur-bations, but also the muscle activation patterns used to restore bal-ance. Support-surface translations in 12 directions were delivered tosubjects standing in six different postural configurations: one-leg,narrow, wide, very wide, crouched, and normal stance. Muscle syn-ergies were extracted from each condition using nonnegative matrixfactorization. In addition, muscle synergies from the normal stancecondition were used to reconstruct muscle activation patterns acrossall stance conditions. A consistent set of muscle synergies wererecruited by each subject across conditions. When balance demandswere extremely different from the normal stance (e.g., one-legged orcrouched stance), task-specific muscle synergies were recruited inaddition to the preexisting ones, rather generating de novo musclesynergies. Taken together, our results suggest that muscle synergiesrepresent consistent motor modules that map intention to action,regardless of the biomechanical context of the task.
I N T R O D U C T I O N
The control of balance requires sensorimotor transforma-tions that allow the nervous system to rapidly interpret multiplesensory input signals from all segments of the body to producecontext-dependent muscle activation patterns that stabilize thebody. In humans, we previously observed that the variations inmuscle activation patterns evoked in response to differentdirections of support-surface movements in the horizontalplane could be described by a limited set of muscle synergies(Torres-Oviedo and Ting 2007). We define muscle synergies asinvariant patterns of activation across multiple muscles that arecombined to produce complex muscle activation patterns. Wehypothesize that variations in muscle activity for movement aregenerated through the flexible recruitment of a limited set ofmuscle synergies (for review see Ting and McKay 2007;
Tresch and Jarc 2009). To challenge our hypothesis, we inves-tigated whether muscle synergy structures are consistent acrossconditions in which the biomechanical and sensory context ofthe postural task change and where large variations in muscleactivity are observed. To this end we examined muscle acti-vation patterns in postural responses to perturbations adminis-tered to subjects standing in six different postural configura-tions.
Consistency in muscle synergy structure and function acrossdifferent biomechanical contexts would suggest that musclesynergies represent motor modules that map motor intention toaction. Accordingly, changes in muscle activity would be dueto the modulation of neural commands that recruit musclesynergies, but not to changes in the structure of the musclesynergies themselves. Studies in frogs demonstrate that com-mon muscle synergy structures produce similar functions likeextending the leg across behaviors with different biomechani-cal contexts such as swimming, jumping, and wiping (d’Avellaand Bizzi 2005). Similarly, in cat postural responses, wedemonstrated that changes in mean muscle activity acrossdifferent postural configurations could be explained by modu-lating the recruitment of the same set of muscle synergies(Torres-Oviedo et al. 2006). These muscle synergies appear toproduce the same endpoint force vector relative to the limbacross a range of postural configurations (McKay and Ting2008; Torres-Oviedo et al. 2006). In human pedaling andwalking, consistent functional roles of muscle synergies havebeen demonstrated to allow robust behaviors across locomotorspeeds (Clark et al. 2010; McGowan et al. 2010; Neptune et al.2009; Raasch and Zajac 1999). In human balance control, wedemonstrated that trial-by-trial changes in electromyographic(EMG) patterns across multiple muscles could be accountedfor by the recruitment of a consistent set of muscle synergies torestore balance (Torres-Oviedo and Ting 2007). However, inour prior human study we examined subjects standing only intheir preferred postural configuration. Therefore it is unknownwhether the identified muscle synergies were consistent be-cause they reflected motor modules for mapping intention toaction or because they simply reflected the consistent biome-chanical context of the balance task that we studied.
Although, the consistency of muscle synergies in response toperturbations could be due to the shaping of motor outputs bycharacteristic patterns of somatosensory inflow, evidence sug-gests that this is not the case. During postural perturbations tostanding balance, somatosensory information arising from thejoints and skin is critical to the timing of the initial burst ofpostural muscle activity (Inglis et al. 1994; Stapley et al. 2002).Moreover, the specific pattern of sensory input is determinedby the interactions between the perturbation and the musculo-
Address for reprint requests and other correspondence: L. H. Ting, TheWallace H. Coulter Department of Biomedical Engineering, at Georgia Techand Emory University, 313 Ferst Drive, Atlanta, GA 30322-0535 (E-mail:[email protected]).
J Neurophysiol 103: 3084–3098, 2010.First published April 14, 2010; doi:10.1152/jn.00960.2009.
3084 0022-3077/10 Copyright © 2010 The American Physiological Society www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

skeletal configuration of the limb (Honeycutt et al. 2009).Patterns of sensory input are therefore expected to change withpostural configuration. If muscle synergy structures are shapedby somatosensory patterns, then their structure would be pre-dicted to change in direct relation to sensory input patterns.However, a decoupling of somatosensory patterns and motoroutput patterns has been demonstrated in both human and catpostural responses, whereby perturbations causing oppositejoint motions and loadings give rise to similar muscle activa-tion patterns (Nashner 1977; Ting and Macpherson 2004). Thisdemonstrates that postural muscle activity is not consistentlyrelated to specific patterns of sensory inputs. In contrast, thesame muscle activation patterns across different perturbationparadigms are predicted by the required direction of center ofmass (CoM) motion for balance control, suggesting that themotor output is related to the performance of a task level goal(Gollhofer et al. 1989; Ting and Macpherson 2004). Moreover,in cat postural control we demonstrated that the same musclesynergies could account for EMG responses to perturbationsthat induced opposite changes in joint angle (i.e., differentsensory inflow), but similar (CoM) displacements (Torres-Oviedo et al. 2006). Similarly, muscle synergy structure islargely preserved after deafferentation, in which changes inmotor output patterns are attributable to changes in therecruitment of muscle synergies (Cheung et al. 2005; Kargoet al. 2010). These results further support the idea thatmuscle synergies are motor output modules whose recruit-ment, but not structure, is altered by changing patterns ofsensory inflow. An implication is that changes in patterns oflocal somatosensory feedback associated with different pos-tural configurations would not be expected to alter musclesynergy structure.
It has also been suggested that the identification of musclesynergies in motor tasks reflects the optimal muscle coordina-tion pattern to control the limb in a given configuration, ratherthan modularity of motor outputs (Fagg et al. 2002; Kurtzer etal. 2006). The invariant muscle synergies that we identified inpostural control could be emergent patterns based on optimalcontrol of the musculoskeletal system and not explicitly en-coded in the nervous system. An implication is that the optimalmuscle activation pattern would be expected to change whenthe biomechanical context of the task is altered. However, therehas been surprising robustness in the identification of musclesynergies across different motor tasks, such as swimming andlocomotion in frogs (Cheung et al. 2005; d’Avella and Bizzi2005), different postural configurations in cats (Torres-Oviedoet al. 2006), or when grasping different shaped objects (Over-duin et al. 2008). Musculoskeletal modeling studies have alsodemonstrated that although muscle synergy structures could beoptimized for a particular biomechanical task or objectivefunction, they may be more generally applied to produce awider repertoire of related tasks (Berniker et al. 2009; Raaschand Zajac 1999; Valero-Cuevas et al. 1998) In cat posturalcontrol, we demonstrated that postural forces were predicted torotate, as in experimental observations, only if muscle synergyconstraints were imposed, but not if individual muscle coordi-nation was allowed (McKay and Ting 2008). Thus althoughmuscle synergies might indeed be shaped by optimality prin-ciples, they might still be encoded within the nervous system asa heuristic representation of a particular optimal motor solu-tion. If the identified muscle synergies are emergent from
optimization processes, then we would predict muscle synergystructures to change as the biomechanical context of muscleactions varies across different postural configurations. Alter-nately, if muscle synergies reflect a set of motor modules thatare generally recruited to perform particular motor tasks, thenwe would predict them to be invariant across different posturalconfigurations.
Here we altered the postural configuration of subjects to testthe consistency of muscle synergies across different biome-chanical and sensory contexts. Moderate changes in posturalconfiguration consisted of changes in stance width and havebeen shown to alter the magnitude of EMG responses (Henryet al. 2001). Extreme changes in postural configuration con-sisted of standing in a crouched configuration or on one leg,which have been shown to alter amplitude, latency (crouch;Burtner et al. 1998), and directional tuning (one-leg; Tropp andOdenrick 1988) of EMG responses. We observed that thestructure of muscle synergies previously identified (Torres-Oviedo and Ting 2007) was independent of the musculoskel-etal configuration. The large variations in EMG activity withinand across biomechanical contexts were accounted for by therecruitment of consistent sets of subject-specific muscle syn-ergies. Also, when balance demands were extremely differentfrom the typical stance (e.g., one-legged or crouched stance),task-specific muscle synergies were recruited in addition to thepreexisting ones, rather generating de novo muscle synergies.In sum, these results suggest that muscle synergies simplifybalance control by providing consistent motor outputs that mapintention to action, regardless of the biomechanical context ofthe postural task.
M E T H O D S
Experimental setup
To test the degree to which the biomechanical context of thepostural task influenced the structure and recruitment of musclesynergies used for balance control, we studied human responses tobalance perturbations in multiple stance configurations. Subjects stoodin six postural configurations and experienced unexpected support-surface translations in 12 evenly distributed directions in the horizon-tal plane (Fig. 1), yielding a total of 72 different combinations ofperturbation direction and postural configuration tested in each sub-ject. Data were collected over 2 consecutive days. In each session,both postural configuration and perturbation direction were random-ized. In all, nine healthy subjects (five females and four males; meanage: 21.9 � 3.6 yr; mean height: 67.4 � 2.9 in.; mean weight:140.4 � 17.8 lb; mean body mass index [BMI]: 21.6 � 1.7) partic-ipated in the study. All experimental protocols were approved by theGeorgia Tech and Emory University Institutional Review Boards.
To induce a large degree of variability, EMG responses wereevoked during both “moderate” and “extreme” variations in posturalconfiguration. Additional variability in EMG responses was furtherintroduced within trials of the same stance conditions because we didnot explicitly monitor or prescribe the joint angles or center ofpressure location prior to each trial. For “moderate” variations inpostural configuration subjects were instructed to stand at eithernarrow, normal, or wide stance widths by placing their heels onmarks located 9, 19, and 30 cm apart, respectively. These condi-tions were previously shown to alter the magnitude of EMGresponses in humans (Henry et al. 1998, 2001). For “extreme”variations in postural configuration subjects were instructed tostand at the widest stance width, in one-leg stance, and in acrouched stance. The distance between the feet in the widest stance
3085INVARIANT MUSCLE SYNERGIES ACROSS POSTURAL CONFIGURATIONS
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

was 60 cm, twice that of the wide stance. In the one-leg stance,subjects were instructed to maintain their balance on their right leg(dominant leg for all subjects) throughout the entire platformmotion, without allowing the ankle of the nonstance leg to bebraced against the stance limb. In the crouched stance, subjectsbent their knees (�20° knee flexion) while maintaining their feet incontact with the floor and their torso relatively upright.
On the first day, normal, narrow, wide, and widest stances weretested; normal, one-leg, and crouched stances were tested on thesecond day. The normal stance was collected on each day to act as acontrol for comparisons across days. In each session, five replicates ofeach condition were collected. Thus a total of ten replicates werecollected for normal stance; these data have been previously analyzedand published (Torres-Oviedo and Ting 2007). Ramp-and-hold sup-
AT
DE
MV
0.2
0.6D
EM
G
A Platform perturbation kinematics
B EMG responses to 120° perturbations
4
12
0 0.5 10
0
0.5 1
0 0.5 1 0 0.5 1 0 0.5 1 0 0.5 1 0 0.5 1 0 0.5 1
one-leg narrow normal wide widestcrouched
time (sec)
time (sec) time (sec)
)mc( tne
mecalp sid
)g ( no it are lecca
0
platform kinematics for one-leg-stanceplatform kinematics for all other stances
12 3
BO
XE
OR
EP
TAL
V
narrowone-legcrouched normal wide widest
perturbation direction (deg)
0 180 180 180 180 180 180360 360 360 360 360 3600 0 0 0 0
C EMG tuning curves to all perturbation directions in time bin APR1
FIG. 1. Example of changes in muscle activity patterns across postural configurations during electromyographic (EMG) responses to a leftward-forwardperturbation of the support surface. A: perturbations were induced by a ramp-and-hold motion of the support surface. The same parameters of platform motionwere used for perturbations in all stance conditions except for the one-leg stance, in which a smaller platform motion was used. B: example EMG activity oftibialis anterior (TA), vastus medialis (VMED), and gluteus medius (GMED) in response to the same perturbation direction under different posturalconfigurations. Changes in muscle activity typically occur 100 ms following the onset of platform motion (vertical dashed line). Mean EMG activity in 3 timebins of 75 ms (shaded areas; EMGAPR1, EMGAPR2, EMGAPR3) were used in the analysis. C: representative tuning curves of external oblique (EXOB), vastuslateralis (VLAT), and peroneus (PERO) during EMGAPR1 of a sample subject standing in all stance conditions. Directions of peak muscle activity were conservedin the normal, narrow, and wide configurations, but could change in the one-leg, crouched, and widest stance conditions. Black traces indicate the mean responseand gray dots represent responses in each trial. Intertrial variations in EMG responses were also observed in all stance conditions, as shown by the spread in graydots indicating response level during a single trial.
3086 G. TORRES-OVIEDO AND L. H. TING
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

port-surface translations of 12.4 cm total displacement, 35 cm/s peakvelocity, and 0.5 g (�490 cm/s2) peak acceleration were used for allexperimental conditions except for the one-leg stance, for which asmaller perturbation of 4 cm total displacement, 12 cm/s peak veloc-ity, and 0.2 g (�196 cm/s2) peak acceleration were used (Fig. 1A).
EMG activity was recorded from 16 leg and lower-back muscles ofthe subject’s right side. Electrode positions on the subject’s body weremarked to ensure similar electrode placement in both experimentalsessions. The following muscles were recorded: rectus abdominalis(REAB), tensor fascia lata (TFL), biceps femoris long head (BFLH),tibialis anterior (TA), semitendinosus (SEMT), semimembranosus(SEMB), rectus femoris (RFEM), peroneus (PERO), medial gastroc-nemius (MGAS), lateral gastrocnemius (LGAS), erector spinae(ERSP), external oblique (EXOB), gluteus medius (GMED), vastuslateralis (VLAT), vastus medialis (VMED), and soleus (SOL). RawEMG data were filtered and processed off-line using a set of customMATLAB routines. The entire time course of the raw EMG data washigh-pass filtered at 35 Hz, demeaned, rectified, and low-pass filteredat 40 Hz. EMGs were normalized to their maximum mean valuesmeasured across perturbation directions in the normal stance config-uration. Normalizing the data in this way reduced sensitivity tooutliers.
Data processing
Postural response activity for each trial was characterized by themean EMG activity in each of four time bins: a 280-ms backgroundperiod (BK) that ended 170 ms before the perturbation and each ofthree 75-ms time bins beginning 100 ms (automatic postural response1 [APR1]), 175 ms (APR2), and 250 ms (APR3) after perturbationonset (Fig. 1B). These time bins can characterize the basic temporalphases of EMG responses of individual muscles (Diener et al. 1988;Torres-Oviedo and Ting 2007). Data from each of the 16 muscles ineach postural configuration consisted of a vector of data composed of4 time bins � 12 directions � 5 trials � 240 data points (480 fornormal stance). Therefore across all six postural configurations weanalyzed 26,880 data points.
To ensure the activity in all muscles was equally weighted in themuscle synergy extraction algorithms, muscle data vectors consistingof EMGBK, EMGAPR1, EMGAPR2, and EMGAPR3 across all pertur-bation directions from the normal stance condition were furthernormalized to have unit variance before muscle synergy extraction.Then, muscle data vectors from all the other stance conditions werenormalized with the same normalization factors to maintain consistentunits across conditions.
Data analysis
In all subjects, muscle synergies were extracted for each stancecondition using a nonnegative matrix factorization algorithm de-scribed in detail in the following text. We determined the number ofmuscle synergies that best described each data set by choosing thesmallest number of muscle synergies that could account for 75% ofthe variability of that data set using the same local and global criteriafrom our previous studies (Torres-Oviedo and Ting 2007; Torres-Oviedo et al. 2006). Previously we demonstrated that muscle syner-gies from normal stance were not significantly different when ex-tracted from data collected on 2 consecutive days (Torres-Oviedo andTing 2007). Because these results demonstrate that the differences inelectrode placement did not affect muscle synergy recruitment, wewere able to compare muscle synergies across all conditions.
To overcome the known difficulties when comparing muscle syn-ergies across data sets, we also used an alternate approach of identi-fying shared and specific muscle synergies across conditions as inprevious studies (Cheung et al. 2005, 2009a; Torres-Oviedo et al.2006). We examined whether muscle synergies from normal stancetrials—the control condition—in each subject could account for mus-
cle activation patterns in all of the other postural configurations (testconditions). Task-specific muscle synergies were subsequently ex-tracted when control muscle synergies could not account for importantfeatures in the test conditions. This approach was necessary because1) muscle synergies cannot be reliably identified in data sets wherethey are not recruited (Torres-Oviedo et al. 2006) and 2) musclesynergies that are recruited independently in one condition but covaryin another condition will appear merged and cause an apparentreduction in the number of muscle synergies (Cheung et al. 2005;Clark et al. 2010; Raasch and Zajac 1999; Saltiel et al. 2001; Ting etal. 1999). We then compared the extracted muscle synergies for eachstance to the control and task-specific muscle synergies. Finally tofurther cross-validate our results, we also compared them to musclesynergies extracted from the entire data pool.
Extraction of muscle synergies from a singlepostural configuration
Muscle synergies were extracted using nonnegative matrix factor-ization, a linear decomposition technique previously presented(Cheung et al. 2005; Lee and Seung 2001; Torres-Oviedo and Ting2007; Torres-Oviedo et al. 2006; Tresch et al. 2006). This lineardecomposition technique assumes that each muscle activation patternM, evoked by a perturbation at a given time period and stance configu-ration (e.g., Fig. 8B; Trial 1 in one-leg condition) is composed of a linearcombination of a few (Nsyn) muscle synergies W, each recruited bysynergy scaling coefficient ci. Thus the predicted muscle activationpattern M̂ takes the form
M^ (t) � �i�1
i�Nsyn
ci(t)Wi
ci � 0 Wi � 0(1)
The ith muscle synergy Wi is represented as a vector that specifiesa spatial pattern of muscle activity identified in the control conditiondata set. Each element of Wi represents a muscle whose relativecontribution to the muscle synergy is time invariant and takes a valuebetween 0 and 1. The nonnegative scaling coefficient ci describes therecruitment of the muscle synergy, representing the purported neuralcommand to the muscle synergy that determines the relative contri-bution of Wi to the overall predicted muscle activation pattern M̂. Theset of ci values scaling Wi across all perturbation directions duringquiet stance and during the three automatic postural response (APR)periods is the vector Ci. The components of Ci are tuning curves thatdescribe how the recruitment of Wi changes as a function of pertur-bation direction and time.
In all our subjects we extracted muscle synergies from the controlcondition using an iterative process where Nsyn varied between 1 and16, the number of muscles. Then using the control muscle synergieswe determined the coefficients that would best reconstruct the EMGresponses at each test condition: Cwide, Cwidest, Ccrouched, Cone-legged,and Cnarrow, or Cnormal. We selected the smallest number of musclesynergies that could adequately reconstruct background and APRresponses of each muscle in all the trials of control and test conditions.This was determined by both global and local criteria, which ensuredthat muscle synergies represented actual features in the data set andnot of the noise. We used variability accounted for (VAF), defined as100 � uncentered Pearson correlation coefficient (Torres-Oviedo etal. 2006; Zar 1999), to quantify the goodness of the data reconstruc-tion by the muscle synergies. First, we plotted the overall VAF,indicating the goodness of the fit of the overall data, against thenumber of muscle synergies and looked for the inflection pointbeyond which additional increase in muscle synergies caused onlysmall increments in VAF. Next, we used a local criterion to addmuscle synergies whose recruitment accounted for a relatively smallerpercentage of the overall VAF but provided significant improvement(�75% VAF) in the reconstruction of a particular direction, condition,
3087INVARIANT MUSCLE SYNERGIES ACROSS POSTURAL CONFIGURATIONS
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

or muscle activity pattern. These criteria ensured that each muscletuning curve at all stance configurations would be well reconstructed,so that the critical spatiotemporal features of each muscle activationpattern were well accounted for by the muscle synergies. In general,by satisfying our local criteria, the total VAF in the data set was wellover 90%. If the addition of a muscle synergy contributed evenly tothe VAF of directions, conditions, or muscles, it was not includedbecause it was considered to represent only random variations in thedata and not those due to either direction or condition.
Extraction of task-specific muscle synergies
Task-specific muscle synergies Wtest were extracted in test condi-tions where control muscle synergies extracted from the normal stance(control condition) were not adequate to account for �75% of thevariability in muscle activation patterns. Task-specific muscle syner-gies were relevant only to the test conditions and they were identifiedby applying the same principles used in the reformulation of the NMFalgorithm by Cheung et al. (2005, 2009a). We provided the algorithmwith the test condition data and the control muscle synergies W. Thealgorithm first performs a least-squares fit to determine the nonnega-tive coefficients Ctest that would best reconstruct the test conditiondata using W. Subsequently, the algorithm determines Wtest to recon-struct the test condition data not accounted for by W. Thus similar toa multiple regression, the net muscle activation pattern in the testcondition M̂test was obtained by the projection of the test data onto Wand Wtest extracted from the residual data. Stated formally
M^ test(t) � �i�1
i�Nsyn
ctest i(t)Wi � �i�1
i�N test
citest(t)Wi
test
ctest i, citest � 0 Wi, Wi
test � 0(2)
The ith task-specific muscle synergy Witest is a time-invariant
nonnegative vector that specifies a spatial pattern of muscle activityfeatured in the test data only. The magnitude of its contribution toM̂test is determined by ci
test, representing the neural command to Witest.
Citest is the set of ci
test values scaling Witest across all perturbation
directions and time periods. Thus Ctest describes how the recruitmentof task-specific muscle synergies Wtest change as a function ofperturbation direction and time.
Validation of subject-specific and task-specificmuscle synergies
To validate the number and pattern of muscle synergies per subject,we extracted muscle synergies independently from different portionsof the data set. Muscle synergies were extracted not only from eachstance condition but also from data pooled across all stance condi-tions. For each subject, we calculated correlation coefficients tocompare control and task-specific muscle synergies to those extractedfrom different portions of the entire data set (i.e., from each stancecondition or all stance conditions).
In addition, to test whether intertrial variability had a structurespecific to each subject, we compared the ability to reproduce eachsubject’s EMG patterns in all stance conditions by synergies fromeach subject’s own data versus those from the other subjects. One-wayANOVA on the variability accounted for per muscle synergy set wasperformed. The type of muscle synergies used was set as a factor withFisher’s least significant difference (LSD) post hoc test to determinethe effect of subject specificity and data variability on the reconstruc-tion. We used P � 0.05 as a measure of significance for all statisticalanalyses, which were completed using Statistica (StatSoft, Tulsa, OK)software.
Statistical analysis of synergy recruitment coefficients
To determine the effect of stance configuration on muscle synergyrecruitment we first performed a functional sorting of control and
task-specific muscle synergies across subjects. Muscle synergies ofeach subject were ordered based on muscle composition and synergyrecruitment profiles rather than on percentage of contribution to thetotal data variability (as in other factorization methods such asprincipal component analysis). We performed a functional sortingbecause subjects might use muscle synergies differently, causingcomparable muscle synergies to have large differences in contributionto the total data variability.
To perform the functional sorting we computed averaged synergyrecruitment coefficients across all trials (C� ) for each muscle synergy(W) of each subject. Then an initial sorting was performed bygrouping muscle synergies based on the similarity of W values and/orC� values (r � 0.70) to that of an arbitrary reference subject. From thisinitial sorting, an averaged set of W values and C� values acrosssubjects was computed. Then, using an iterative process, only musclesynergies that were similar to either averaged W values or C� values,or both (r � 0.70), were kept in the group. The averaged set of W andC� vectors across subjects were updated every time a muscle synergywas discriminated from a group. The r values obtained served not onlyas a sorting parameter, but also as a measure to evaluate the generalityof muscle synergies across subjects. Therefore within each group, ther values were used to identify similarities across subjects in both Wand C� , or only W, or only C� .
Subsequent to the functional sorting, we performed a repeated-measures ANOVA on all sample times of the peak activation for eachmuscle synergy. Stance condition and time bin were set as factors withFisher LSD post hoc test to determine the effect of posture and timeperiod on the synergy recruitment coefficients. Bonferroni correctionswere done in all our comparisons and we used P � 0.05 as a measureof significance for all statistical analyses, which were completed usingStatistica software.
R E S U L T S
In all subjects, both moderate and extreme changes inposture introduced variations in balance responses. Althoughmoderate changes to postural configuration induced mainlychanges in EMG amplitude, extreme changes to postural con-figuration induced changes in both directional and temporalpatterning of muscle responses to perturbations. Surprisingly,muscle synergy structures were consistent regardless of thedata set from which they were extracted. Control musclesynergies extracted from the normal stance condition repro-duced most of the variability in balance responses over bothmoderate and extreme changes in postural configuration. Underthe extreme difference in postural configuration induced by one-leg and crouched stance, some muscle synergies were not restruc-tured and only one additional task-specific muscle synergy wasrequired. Similarities in most muscle synergies were found acrosssubjects, but variability in EMG responses across trials and acrossconditions was best accounted for by a subject’s own musclesynergies rather than those obtained from a different subject.
Changes in muscle activity across postural configurations
Similar to previous studies investigating the effect of stancewidth (Henry et al. 2001), moderate changes in postural con-figuration across narrow, normal, and wide stances induceprimarily changes in EMG responses amplitude (Fig. 1). Mus-cle activity decreased as stance width increased, particularly inproximal compared with distal muscles. Both the temporalfeatures (Fig. 1B) and the spatial tuning (Fig. 1C) of individualmuscle activity exhibited similar patterns across moderatechanges in postural response.
3088 G. TORRES-OVIEDO AND L. H. TING
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

Across extreme changes in postural configuration in thecrouched, one-leg, and widest stances, more complex changesin EMG response amplitude, timing, and spatial tuning wereobserved (Fig. 1). Changes in amplitude were nonmonotonic inextreme changes to postural configuration, with some musclesincreasing and others decreasing their amplitude in response tothe same perturbation (Fig. 1B). Changes in onset latencieswere observed primarily in the crouched stance. In this condi-tion onset latencies of proximal muscles, which normallyoccurred during APR2, were activated earlier during APR1,disrupting the distal-to-proximal muscle activation order typi-cally observed in the normal stance condition (Burtner et al.1998; Woollacott and Shumway-Cook 2002). Changes in di-rectional tuning of EMG signals were also observed in extremechanges in postural configuration. For example, the directionsof maximum and minimum EXOB activity were reversed inwidest stance compared with all other conditions (Fig. 1C).
Muscle synergies are conserved across posturalconfigurations
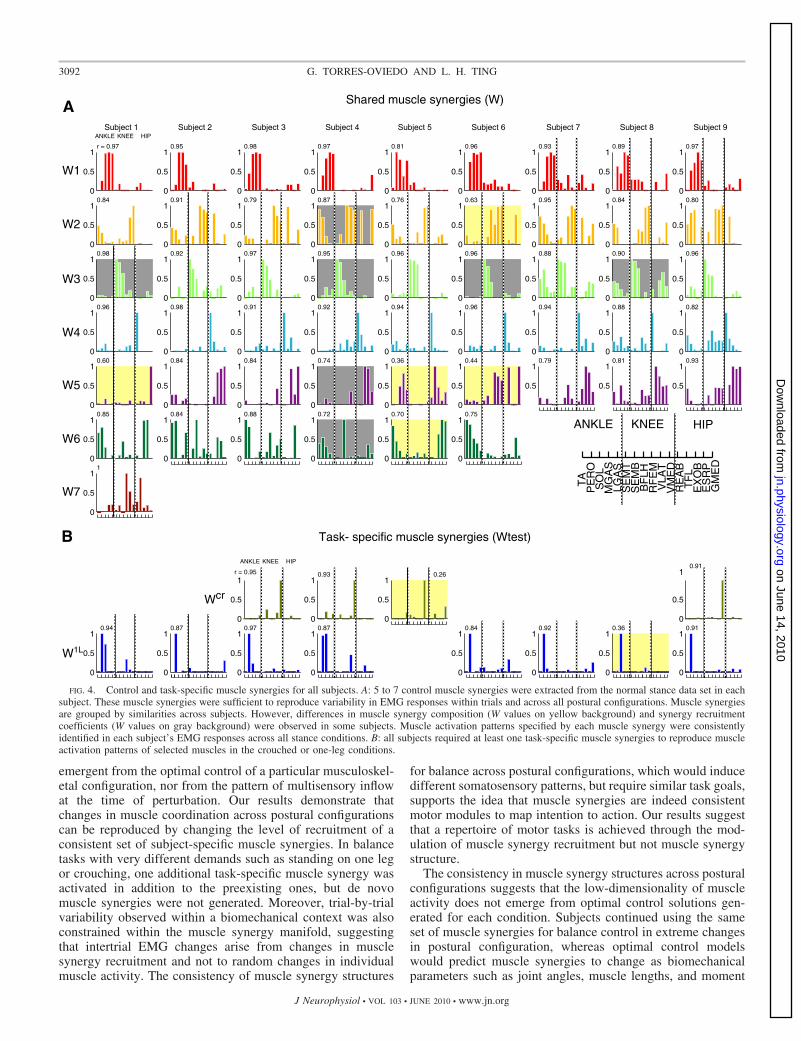
In all subjects, control muscle synergies extracted fromnormal stance were sufficient to account for variations inmuscle activity across moderate changes in postural configu-ration. All subjects had five to seven control muscle synergies(Fig. 4A), accounting for 93 � 1.4, 93 � 0.8, and 93 � 1% ofthe total variability across trials in normal, narrow, and widestances, respectively. In six of nine subjects, the number ofmuscle synergies required to reconstruct muscle activity acrossmoderate changes in postural configuration was the same as thenumber required to reconstruct normal stance only. In theremaining three subjects, we observed the splitting of a musclesynergy extracted from normal stance to account for variationin other stances. In essence, two muscle synergies were con-trolled together in normal stance, but independently on varia-tion in postural configurations (Clark et al. 2010; Saltiel et al.2001; Ting et al. 1999).
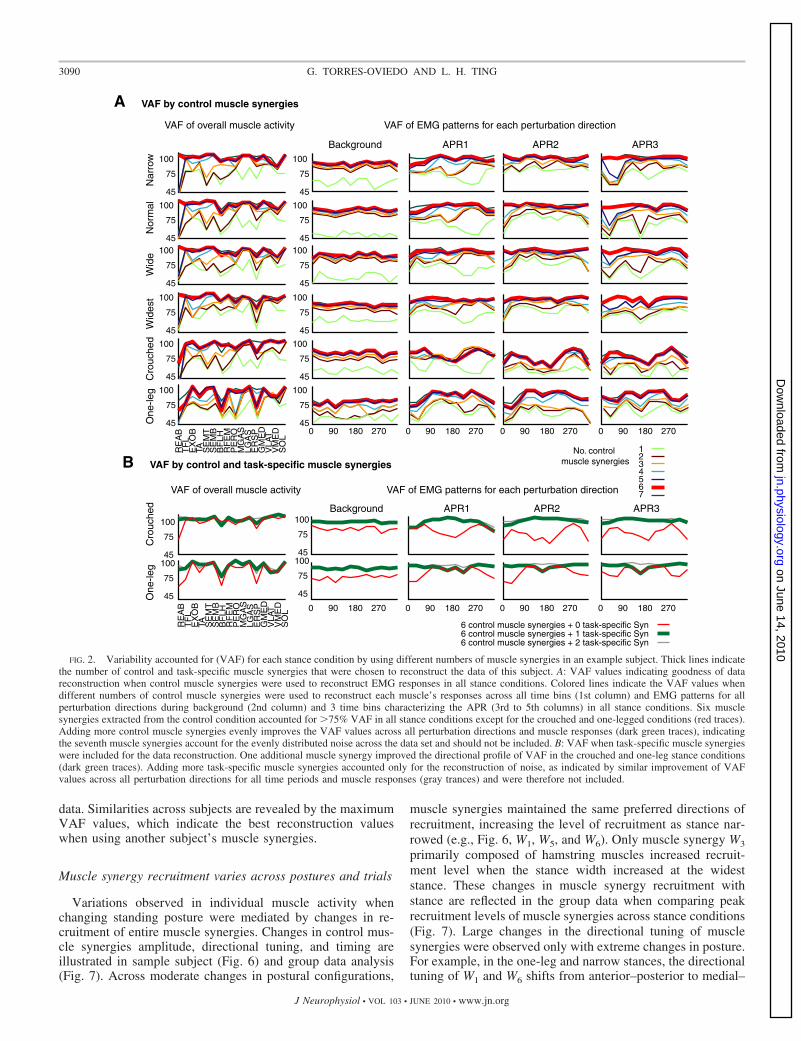
Task-specific muscle synergies were sometimes needed toaccount for the variability in two of the extreme configurations:one-leg and crouched stance. Control muscle synergies ac-counted for 92 � 1% (widest), 84 � 9.9% (crouched), and84 � 4.7% (one-leg) variability across trials in extreme pos-tural configurations. In contrast to moderate changes in pos-tural configuration, the addition of more control synergies didnot improve the VAF values. Task-specific muscle synergieswere necessary to account for the variability in one-leg andcrouched EMG responses of four or fewer muscles of the 16that were recorded. The mean VAF in these conditions in-creased to 91 � 3.6% (crouched) and 92 � 1.3% (one-leg)with the addition of a single task-specific muscle synergy foreach condition. As an example, in subject 3 we observed thatsix control muscle synergies constituted the smallest number ofsynergies that accounted for �75% muscle activity duringbackground and APR periods of all muscles in the narrow,normal, wide, and widest conditions (Fig. 2A, red trace).However, not all muscle activity was well reconstructed in theone-legged and crouched stance, as indicated by the poordirectional profile of VAF for all time periods (Fig. 2A; redtrace on one-legged and crouched panels in the second tofourth columns from right to left) and the low VAF values ofREAB, BFLH, and ERSP (Fig. 2A; on one-legged and
crouched panels in first column on the right). Adding onetask-specific muscle synergy to the reconstruction of these twoconditions dramatically improved the reconstruction of allmuscles and the directional profile of VAF (Fig. 2B). We didnot include more than six control muscle synergies or morethan one task-specific muscle synergy in the crouched andone-legged conditions because the addition of further musclesynergies accounted only for the reconstruction of noise, asindicated by similar improvement of VAF values across allperturbation directions for all time periods and muscles (Fig. 2,A and B, green trace and gray trace, respectively).
Muscle synergies extracted independently from each stancecondition were remarkably similar (e.g., Fig. 3). Although notall the control muscle synergies could be extracted from eachconfiguration, the ones that were identified in the test condi-tions were similar to those extracted in the normal stance.Across subjects, 98, 94, 92, 69, and 67% of the control musclesynergies presented here (e.g., Fig. 3) were also identifiedwhen using the narrow data set (0.81 � 0.08 � r � 0.97 �0.02), the wide data set (0.86 � 0.07 � r � 0.95 � 0.04), thewidest data set (0.81 � 0.08 � r � 0.95 � 0.03), the one-legdata set (0.72 � 0.07 � r � 0.94 � 0.03), and the croucheddata set (0.69 � 0.12 � r � 0.92 � 0.03), respectively. Thefewest numbers of control muscle synergies were extractedfrom the crouched and one-leg stances because of the lowrecruitment of control muscle synergies activating proximalmuscles. This can be explained by the low activity of trunkmuscles in the crouched stance and the low activity of abdom-inal and hamstring muscles in the one-leg stance. Also in allsubjects, control and task-specific muscle synergies presentedhere (Fig. 3) were similar to muscle synergies extracted fromdata pooled across all stances (0.7 � 0.07 � r � 0.97 � 0.04)and task-specific synergies were similar to test muscle synergyextracted from the one-leg and crouched stances alone (r �0.87 � 0.10). SOL, PERO, VMED, and VLAT were typicalexamples of muscles in which their overall activation was repro-duced by control and task-specific muscle synergies (Fig. 4B).
Although the number and muscle synergy structure variedconsiderably across subjects (Fig. 4), the same muscle synergystructures were consistently extracted from different portionsof each subject’s data set. Overall, muscle synergies were similaracross subjects, although we also observed clear subject-specificdifferences in muscle synergy structure (Fig. 4, W values onyellow background) or recruitment (Fig. 4, W values on graybackground).
The total variability accounted for in each subject’s data setwas greatest when using subject-specific muscle synergies(Fig. 5). We observed the variance in the data set containing alltest conditions was roughly twice that of the normal stancealone (Fig. 5A), yet control and task-specific muscle synergieswere sufficient to account for these variations. A range of VAFvalues was obtained when using others’ muscle synergies toreconstruct a subject’s data. Overall, the goodness in thereconstruction decreased when using others’ muscle synergies,as indicated by the significantly lower mean and minimumVAF values (Fig. 5B). The poor variability accounted for byothers’ muscle synergies is also indicated by the low VAFvalues in the reconstruction of individual muscles (Fig. 5C).VAF values in four of nine subjects was �62%, which isapproximately the amount of variability accounted for by usingartificial muscle synergies extracted from randomly generated
3089INVARIANT MUSCLE SYNERGIES ACROSS POSTURAL CONFIGURATIONS
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from
data. Similarities across subjects are revealed by the maximumVAF values, which indicate the best reconstruction valueswhen using another subject’s muscle synergies.
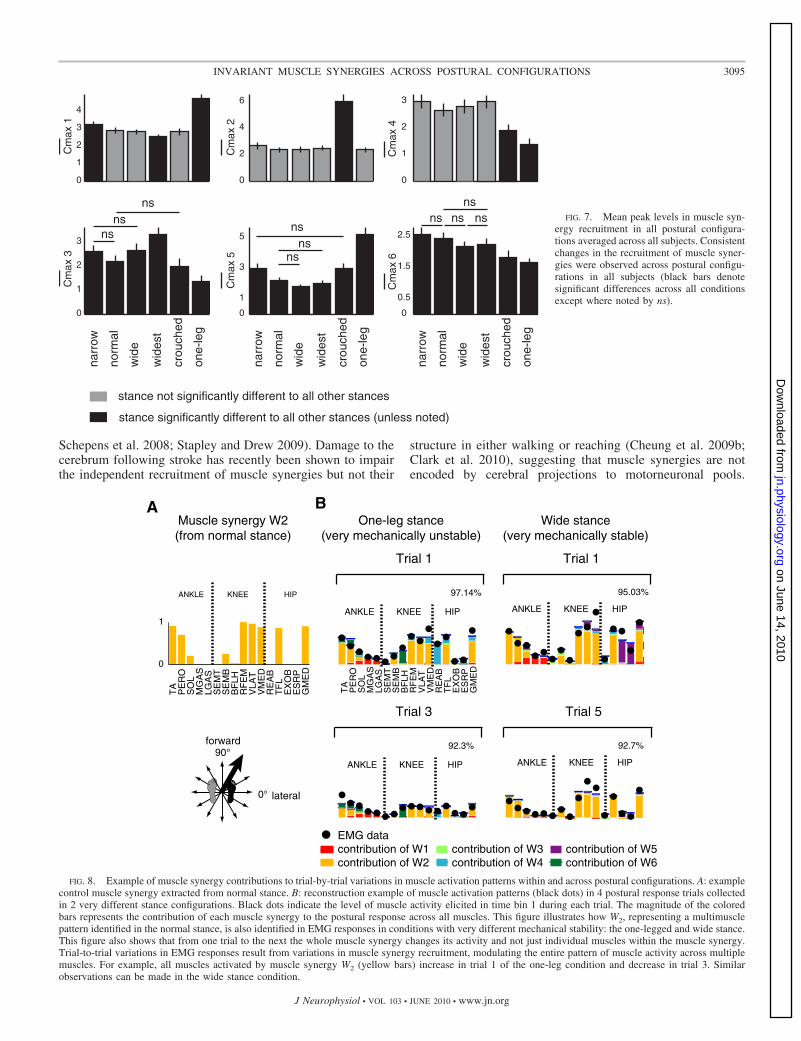
Muscle synergy recruitment varies across postures and trials
Variations observed in individual muscle activity whenchanging standing posture were mediated by changes in re-cruitment of entire muscle synergies. Changes in control mus-cle synergies amplitude, directional tuning, and timing areillustrated in sample subject (Fig. 6) and group data analysis(Fig. 7). Across moderate changes in postural configurations,
muscle synergies maintained the same preferred directions ofrecruitment, increasing the level of recruitment as stance nar-rowed (e.g., Fig. 6, W1, W5, and W6). Only muscle synergy W3
primarily composed of hamstring muscles increased recruit-ment level when the stance width increased at the wideststance. These changes in muscle synergy recruitment withstance are reflected in the group data when comparing peakrecruitment levels of muscle synergies across stance conditions(Fig. 7). Large changes in the directional tuning of musclesynergies were observed only with extreme changes in posture.For example, in the one-leg and narrow stances, the directionaltuning of W1 and W6 shifts from anterior–posterior to medial–
Nar
row
Nor
mal
Wid
eW
ides
t
VAF of overall muscle activity VAF of EMG patterns for each perturbation direction
45
75
100
45
75
100
45
75
100
45
75
100
45
75
100
45
75
100
45
75
100
45
75
100
B
Cro
uche
dO
ne-le
gC
rouc
hed
One
-leg
RE
AB
TF
LE
XO
BTA S
EM
TS
EM
BB
FLH
RF
EM
PE
RO
MG
AS
LGA
SE
RS
PG
ME
DV
LAT
VM
ED
SO
L
VAF by control and task-specific muscle synergies
VAF by control muscle synergiesA
VAF of overall muscle activity VAF of EMG patterns for each perturbation direction
RE
AB
TF
LE
XO
BTA S
EM
TS
EM
BB
FLH
RF
EM
PE
RO
MG
AS
LGA
SE
RS
PG
ME
DV
LAT
VM
ED
SO
L
45
75
100
0 90 180 27045
75
100
45
75
100
45
75
100
0 90 180 270 0 90 180 270 0 90 180 270
45
75
100
0 90 180 270
45
75
10045
75
100
45
75
100
0 90 180 270 0 90 180 270 0 90 180 270
6 control muscle synergies + 0 task-specific Syn6 control muscle synergies + 1 task-specific Syn 6 control muscle synergies + 2 task-specific Syn
1234567
No. control muscle synergies
Background APR1 APR2 APR3
Background APR1 APR2 APR3
FIG. 2. Variability accounted for (VAF) for each stance condition by using different numbers of muscle synergies in an example subject. Thick lines indicatethe number of control and task-specific muscle synergies that were chosen to reconstruct the data of this subject. A: VAF values indicating goodness of datareconstruction when control muscle synergies were used to reconstruct EMG responses in all stance conditions. Colored lines indicate the VAF values whendifferent numbers of control muscle synergies were used to reconstruct each muscle’s responses across all time bins (1st column) and EMG patterns for allperturbation directions during background (2nd column) and 3 time bins characterizing the APR (3rd to 5th columns) in all stance conditions. Six musclesynergies extracted from the control condition accounted for �75% VAF in all stance conditions except for the crouched and one-legged conditions (red traces).Adding more control muscle synergies evenly improves the VAF values across all perturbation directions and muscle responses (dark green traces), indicatingthe seventh muscle synergies account for the evenly distributed noise across the data set and should not be included. B: VAF when task-specific muscle synergieswere included for the data reconstruction. One additional muscle synergy improved the directional profile of VAF in the crouched and one-leg stance conditions(dark green traces). Adding more task-specific muscle synergies accounted only for the reconstruction of noise, as indicated by similar improvement of VAFvalues across all perturbation directions for all time periods and muscle responses (gray trances) and were therefore not included.
3090 G. TORRES-OVIEDO AND L. H. TING
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

lateral directions (Fig. 6, red and dark green). These changesoccur, presumably to compensate for the missing contralateralsupport on medial–lateral perturbations.
Significant temporal differences in the recruitment of musclesynergies was mainly observed in the extreme stance condi-tions (P � 0.001). Temporal patterns of muscle synergiesrecruitment by either starting at an earlier onset (e.g., C2 in thecrouched stance), later onset (e.g., C4 in the widest stance), orincreasing their background activity (e.g., C6 in the one-legstance). For example, the onset of C2, which recruited a musclesynergy that was largely composed by quadriceps muscles,changed to an earlier time bin APR1 in the crouched stance.Changes in background activity of muscle synergies weremainly observed in the extreme stance conditions, such as thelarger recruitment level in CCr and C2 in the crouched stanceand C6, formed by ankle dorsiflexor and evertors and biartic-ular muscle biceps femoris, in the one-leg stance.
In all stance conditions, intertrial variations in EMG activityof individual muscles were not due to noise at the level ofmuscles, but instead corresponded to intertrial variations in therecruitment of entire muscle synergies. Similar trial-to-trialvariations of responses were observed in two trials of stanceswith very different biomechanical stability conditions: theone-leg and wide stances (Fig. 8, black dots). The changes in
activation of individual muscles, however, were accompanied bysimilar changes in the activation of all other muscles belonging toW2 (Fig. 8, yellow bars). Therefore intertrial changes in musclemagnitude were not simply a random variation in individualmuscle activity but corresponded to modulation of an entire musclesynergy from one trial with respect to another.
Although we observed variability in muscle synergy re-cruitment within each stance, the average contribution ofeach muscle synergy to balance responses across stanceswas very consistent (Fig. 9). That is, the relative contribu-tion of muscle synergies indicated by the VAF level permuscle synergy is very similar in the narrow, normal, wide,and widest stances. These results not only indicate that allcontrol muscle synergies were recruited in all stances butthat they were recruited similarly, suggesting there mightalso be constraints on the recruitment of muscle synergies toEMG responses for balance control. These patterns of mus-cle synergy recruitment were shifted in the one-leg andcrouched conditions (Fig. 9).
D I S C U S S I O N
Because muscle synergy structures were consistent acrossdifferent postural configurations, it is unlikely that they were
0
0.5
1
0
0.5
1
0
0.5
1
0
0.5
1
0
0.5
1
0
0.5
1
r = 0.95
0.9
0.94
0.86
0.91
0.96
0.95
0.99
0.94
0.99
0.86
0.93
0.94
0.92
0.93
0.91
0.8
0.93
0.9
0.98
0.9
0.81
0.9
0.82
0.84
0.66
0.79
0.93
Control and Task-specificMuscle Synergies narrow only wide only widest only crouched only one-legged only
ANKLE KNEE HIP ANKLE KNEE HIP ANKLE KNEE HIPANKLE KNEE HIP ANKLE KNEE HIP ANKLE KNEE HIPANKLE KNEE HIP
all stances
W1
W2
W5
W3
W4
W1L
AT
OR
EP
LO
SS
AG
MS
AGL
TM
ES
BM
ES
HLF
BM
EF
RT
A LV
DE
MV
BA
ER
LF
TB
OX
EP
RS
ED
EM
G
ANKLE KNEE HIP
FIG. 3. Muscle synergies extracted from each condition compared with control and task-specific muscle synergies for a sample subject. Five control and onetask-specific (W1L) were sufficient to reproduce variability in EMG responses within trials and across all postural configurations. These muscle synergies werecompared with muscle synergies extracted from each stance condition alone. Muscle synergies mainly composed of proximal muscles could not be identifiedin the crouched and one-legged condition in all subjects, when these muscles are not highly recruited. However, all other muscle synergies extracted from eachstance were similar to those extracted in the normal stance (e.g., in this subject: 0.66 � r � 0.99).
3091INVARIANT MUSCLE SYNERGIES ACROSS POSTURAL CONFIGURATIONS
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from
emergent from the optimal control of a particular musculoskel-etal configuration, nor from the pattern of multisensory inflowat the time of perturbation. Our results demonstrate thatchanges in muscle coordination across postural configurationscan be reproduced by changing the level of recruitment of aconsistent set of subject-specific muscle synergies. In balancetasks with very different demands such as standing on one legor crouching, one additional task-specific muscle synergy wasactivated in addition to the preexisting ones, but de novomuscle synergies were not generated. Moreover, trial-by-trialvariability observed within a biomechanical context was alsoconstrained within the muscle synergy manifold, suggestingthat intertrial EMG changes arise from changes in musclesynergy recruitment and not to random changes in individualmuscle activity. The consistency of muscle synergy structures
for balance across postural configurations, which would inducedifferent somatosensory patterns, but require similar task goals,supports the idea that muscle synergies are indeed consistentmotor modules to map intention to action. Our results suggestthat a repertoire of motor tasks is achieved through the mod-ulation of muscle synergy recruitment but not muscle synergystructure.
The consistency in muscle synergy structures across posturalconfigurations suggests that the low-dimensionality of muscleactivity does not emerge from optimal control solutions gen-erated for each condition. Subjects continued using the sameset of muscle synergies for balance control in extreme changesin postural configuration, whereas optimal control modelswould predict muscle synergies to change as biomechanicalparameters such as joint angles, muscle lengths, and moment
0
0.5
1r = 0.97
Subject 1
0
0.5
10.84
0
0.5
10.98
0
0.5
10.96
0
0.5
10.60
0
0.5
10.85
0
0.5
11
0
0.5
1 0.94
0
0.5
10.95
Subject 2
0
0.5
10.91
0
0.5
10.92
0
0.5
10.98
0
0.5
10.84
0
0.5
10.84
0
0.5
10.87
0
0.5
10.98
Subject 3
0
0.5
10.79
0
0.5
10.97
0
0.5
10.91
0
0.5
10.84
0
0.5
10.88
0
0.5
1r = 0.95
0
0.5
10.97
0
0.5
10.97
Subject 4
0
0.5
10.87
0
0.5
10.95
0
0.5
10.92
0
0.5
10.74
0
0.5
10.72
0
0.5
10.93
0
0.5
10.87
0
0.5
10.81
Subject 5
0
0.5
10.76
0
0.5
10.96
0
0.5
10.94
0
0.5
10.36
0
0.5
10.70
0
0.5
10.26
0
0.5
10.96
Subject 6
0
0.5
10.63
0
0.5
10.96
0
0.5
10.96
0
0.5
10.44
0
0.5
10.75
0
0.5
10.84
0
0.5
10.93
Subject 7
0
0.5
10.95
0
0.5
10.88
0
0.5
10.94
0.5
10.79
0
0.5
10.92
0
0.5
10.89
Subject 8
0
0.5
10.84
0
0.5
10.90
0
0.5
10.88
0.5
10.81
0
0.5
10.36
0
0.5
10.97
Subject 9
0
0.5
10.80
0
0.5
10.96
0
0.5
10.82
0.5
10.93
0
0.5
10.91
0
0.5
10.91
AT
OR
EP
LO
SS
AG
MS
AGL
TM
ES
BM
ES
HLF
BM
EF
RT
ALV
DE
MV
BA
ER
LF
TB
OX
EP
RS
ED
EM
G
W1
W2
W5
W6
W7
W3
W4
Wcr
W1L
Shared muscle synergies (W) A
Task- specific muscle synergies (Wtest) B
ANKLE KNEE HIP
ANKLE KNEE HIP
ANKLE KNEE HIP
FIG. 4. Control and task-specific muscle synergies for all subjects. A: 5 to 7 control muscle synergies were extracted from the normal stance data set in eachsubject. These muscle synergies were sufficient to reproduce variability in EMG responses within trials and across all postural configurations. Muscle synergiesare grouped by similarities across subjects. However, differences in muscle synergy composition (W values on yellow background) and synergy recruitmentcoefficients (W values on gray background) were observed in some subjects. Muscle activation patterns specified by each muscle synergy were consistentlyidentified in each subject’s EMG responses across all stance conditions. B: all subjects required at least one task-specific muscle synergies to reproduce muscleactivation patterns of selected muscles in the crouched or one-leg conditions.
3092 G. TORRES-OVIEDO AND L. H. TING
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

arms changed (Hamilton et al. 2004; Kurtzer et al. 2006).Moreover, when additional biomechanical functions were nec-essary, such as the production of medial forces during theone-leg condition, subjects added an additional muscle syn-ergy, rather than reshaping all muscle synergies. Our findingsare consistent with evidence demonstrating that muscle syner-gies are preserved across biomechanically different loadingconditions, postural configurations, and locomotor tasks(Cheung et al. 2009a; d’Avella and Bizzi 2005; Torres-Oviedoet al. 2006). This suggests that for a given motor task, thenervous system recruits preexisting muscle synergies ratherthan generating a specific optimal solution for each condition(Chhabra and Jacobs 2006; Scott 2004; Todorov and Ghahra-mani 2004a). Muscle synergies may therefore be optimal whenconsidering an entire movement repertoire, rather than beingspecific to a particular movement condition or limb configu-ration (Berniker et al. 2009; McKay and Ting 2008). It remainsto be investigated whether optimization processes determinehow new muscle synergies are developed in the face of newtask demands and whether preexisting muscle synergies rep-
resent the starting point for learning task-specific muscle syn-ergies.
The fact that differences in sensory inflow in differentpostural configuration alter only the recruitment of musclesynergies and not their structure suggests that they reflectmodularity in motor modules and that are not patterned bysensory inflow. However, we show that differences in sensoryinflow may alter the recruitment of muscle synergies. This isconsistent with postural control studies demonstrating thatmuscle stretch is not always predictive of muscle activationsfor balance control (Carpenter et al. 1999; Horak and Nashner1986; Nashner 1977; Ting and Macpherson 2004). Similarly,our previous work in feline postural control demonstrated thatthe same muscle synergies were recruited when opposite jointangle disturbances were induced (Torres-Oviedo et al. 2006).The idea that muscle synergies are motor modules is furthersupported by the observation that deafferentation does not altermuscle synergy structure but only their recruitment (Cheung etal. 2005; Kargo et al. 2010).
The fact that subject-specific muscle synergies were consis-tent across days and across postural configurations suggeststhat they may reflex habitual or heuristic motor solutionsencoded in the nervous system. Although the biomechanics ofthe musculoskeletal system imposes constraints on musclecoordination patterns (Kutch et al. 2008; Valero-Cuevas 2000),individual differences in morphology are unlikely to accountfor all of the subject-specific differences in muscle synergynumber and structure. Because of the musculoskeletal redun-dancy, multiple solutions for any given biomechanical taskexist (Bernstein 1967). Specific to standing balance control, wehave demonstrated that a high degree of flexibility in muscleactivation patterns remains when the biomechanical constraintsof standing are imposed (Bunderson et al. 2008). The similar-ities in muscle synergy tuning curves across subjects provideevidence that similar task constraints are imposed on all sub-jects when the CoM is perturbed in various directions. How-ever, individual differences in the specific patterns of muscleactivity used to achieve those tasks can vary considerably(Torres-Oviedo and Ting 2007; Torres-Oviedo et al. 2006).These differences might be due to the influence of training andexperience, which would affect the range of behaviors that areused to tailor the specific structure of muscle synergies in anindividual. A similar phenomenon has been observed in neu-ronal circuit models, demonstrating not only the necessity ofusing subject-specific parameter sets rather than parametervalues averaged across subjects (Golowasch et al. 2002) butalso the wide range of possible parameter sets that generatesimilar behavioral outcomes (Prinz et al. 2004). Moreover,recent work suggests that motor cortex topological organiza-tion reflects muscle coordination patterns that are most domi-nant in the behavioral repertoire (Aflalo and Graziano 2006,2007; Graziano and Aflalo 2007), which indicates the possi-bility for experience-dependent and subject-specific shaping ofmuscle synergies.
There is no clear consensus of where muscle synergies maybe encoded in the CNS and how they are modulated. Musclesynergies for frog wiping reflexes may be encoded in spinalcenters, as shown by the consistent modular motor activity inthe frog on spinal stimulation (Hart and Giszter 2010; Saltiel etal. 2001). Muscle synergies for locomotion and scratching mayalso be located in spinal networks that are recruited by rhythm-
Nor
mal
stan
ceAl
l oth
erst
ance
s0
1
2
3
A Data variability
Var
0
20
40
60
80
100
0
20
40
60
80
100
*
Others’ synergies
Own synergies from all data
Max
VA
FM
in V
AF
Max
VA
FM
ean
VAF
Min
VA
F
Min
VA
F
Others’ synergiesOwn synergies from all data
B Overall VAF C Individual muscle VAF
ns
Mea
n VA
F
VA
F
VA
F
***
FIG. 5. Variability in muscle activation patterns and percentage VAF bymuscle synergies extracted from 1) each subject’s own trials in normal stanceand 2) other subjects’ EMG responses in normal stance. A: variance in themuscle activation patterns in the pooled test conditions is significantly largerthan that in the normal stance, from which control muscle synergies wereextracted. B: when accounting for total variability across all muscles andconditions, similar reconstruction levels were possible when using musclesynergies from another subject. However, the mean and minimum variabilitiesaccounted for were significantly decreased when using another subject’smuscle synergies. C: variability accounted for in individual muscles is a moresensitive indicator of the goodness of fit and the minimum VAF was signifi-cantly lower when using other subject’s muscle synergies.
3093INVARIANT MUSCLE SYNERGIES ACROSS POSTURAL CONFIGURATIONS
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

generating circuits (McCrea and Rybak 2008). On the otherhand, muscle synergies for posture and balance control may bemediated by brain stem nuclei. Reticulospinal neurons (RSNs)
project to multiple motoneuronal pools and RSNs are activatedduring postural adjustments across different behavioral andbiomechanical contexts (Schepens and Drew 2004, 2006;
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0 90 180 270
0
1
0
1
0
1
0
1
0
1
0
1
0
1
0
1
A
T O
RE
PL
OS
SA
GM
SA
GLT
ME
SB
ME
SHL
FB
ME
FR
TA L
VD
EM
VB
AE
R L
FT
BO
XE
PR
SE
DE
MG
W1
W2
W5
W6
W3
W4
C1
C2
C5
C6
Wcr Ccr
W1L C1L
C3
C4
ANKLE KNEE
Muscle synergies (W)
HIP
Synergy Coefficients (C), APR1 (100-175ms)
NarrowOne-legCrouched Normal Wide Widest
A B
Perturbation direction (deg)
FIG. 6. Mean (�SD) synergy recruitment levels across stance configurations in a representative subject. A: control muscle synergy vectors Wi, extracted fromEMG responses in normal stance, and task-specific muscle synergies for crouched WCr and one-leg W1L stance condition. Each bar represents the relative levelof activation of each muscle within the synergy (see METHODS section for muscle abbreviations). Task-specific muscle synergies are mainly formed by a highlyactivated muscle. B: mean muscle synergy tuning curves during APR1. The mean recruitment level of control muscle synergies Ci and task-specific musclesynergies Ctest vary as a function of perturbation direction. The spatial tuning of muscle synergy recruitment was consistent across moderate changes in posturalconfiguration (narrow, normal, wide stances) but could shift in extreme changes in postural configuration (crouched, one-leg, widest stances).
3094 G. TORRES-OVIEDO AND L. H. TING
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from
Schepens et al. 2008; Stapley and Drew 2009). Damage to thecerebrum following stroke has recently been shown to impairthe independent recruitment of muscle synergies but not their
structure in either walking or reaching (Cheung et al. 2009b;Clark et al. 2010), suggesting that muscle synergies are notencoded by cerebral projections to motorneuronal pools.
worran
lamron
ed iw
tsed iw
deh cuorc
g el-eno
worran
lamron
ed iw
tsed iw
dehcuorc
gel-eno
wor ran
lamron
ed iw
tsed iw
dehcuo rc
g el-eno
0
1
2
3
3 xam
C
ns
ns ns
0
1
2
3
4
1 xam
C
0
2
4
6
2 xam
C
0
1
2
3
4 xam
C
0
1
3
55 xa
mC
0
0.5
1.5
2.5
6 xam
C
ns
ns ns
ns ns ns ns
stance significantly different to all other stances (unless noted)
stance not significantly different to all other stances
FIG. 7. Mean peak levels in muscle syn-ergy recruitment in all postural configura-tions averaged across all subjects. Consistentchanges in the recruitment of muscle syner-gies were observed across postural configu-rations in all subjects (black bars denotesignificant differences across all conditionsexcept where noted by ns).
97.14%
92.7%
A
TO
RE
PL
OS
SA
GM
SA
GLT
ME
SB
ME
SHL
FB
ME
FR
TAL
VD
EM
VB
AE
RL
FT
BO
XE
PR
SE
DE
MG
ANKLE KNEE HIP
ANKLE KNEE HIP
EMG datacontribution of W1contribution of W2
contribution of W3contribution of W4
Trial 1
Trial 5
92.3%
ANKLE KNEE HIP
Trial 3
95.03%
ANKLE KNEE HIP
Trial 1
One-leg stance(very mechanically unstable)
Wide stance(very mechanically stable)
90°
0° lateral
forward
Muscle synergy W2(from normal stance)
0
1
ANKLE KNEE HIP
A
TO
RE
PL
OS
SA
GM
SA
G LT
ME
SB
ME
SH L
FB
ME
FR
TAL
VD
EM
VB
AE
RL
FT
BO
XE
PR
SE
DE
MG
contribution of W5contribution of W6
A B
FIG. 8. Example of muscle synergy contributions to trial-by-trial variations in muscle activation patterns within and across postural configurations. A: examplecontrol muscle synergy extracted from normal stance. B: reconstruction example of muscle activation patterns (black dots) in 4 postural response trials collectedin 2 very different stance configurations. Black dots indicate the level of muscle activity elicited in time bin 1 during each trial. The magnitude of the coloredbars represents the contribution of each muscle synergy to the postural response across all muscles. This figure illustrates how W2, representing a multimusclepattern identified in the normal stance, is also identified in EMG responses in conditions with very different mechanical stability: the one-legged and wide stance.This figure also shows that from one trial to the next the whole muscle synergy changes its activity and not just individual muscles within the muscle synergy.Trial-to-trial variations in EMG responses result from variations in muscle synergy recruitment, modulating the entire pattern of muscle activity across multiplemuscles. For example, all muscles activated by muscle synergy W2 (yellow bars) increase in trial 1 of the one-leg condition and decrease in trial 3. Similarobservations can be made in the wide stance condition.
3095INVARIANT MUSCLE SYNERGIES ACROSS POSTURAL CONFIGURATIONS
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

Rather, cortical projections may modulate muscle synergyrecruitment. Accordingly, projections from motor cortex tomultiple motoneuronal pools for reaching (Holdefer and Miller2002; Schwartz et al. 1988) have been shown to change theirlevel of recruitment as a function of postural configuration(Kakei et al. 1999; Scott and Kalaska 1997). Moreover, stim-ulation of different cortical sites can recruit similar low-dimensional hand movements (Gentner and Classen 2006).
Intertrial variations in muscle synergy recruitment may beexplained by variations in activity of hierarchal neural struc-tures regulating movement. Recent studies demonstrate thatvariability in the firing of premotor structures is correlated totrial-by-trial variability in behavioral features such as pitch inbirdsong (Sober et al. 2008), reaching speed in monkeys(Churchland et al. 2006), and eye movement in monkeys(Medina and Lisberger 2007). Variability in motor output maytherefore be centrally controlled and passed to the behaviorthrough downstream sensorimotor transformations (Soechtingand Flanders 2008; Yanai et al. 2008) of reduced dimension(Flash and Hochner 2005; Lockhart and Ting 2007). Similarly,intertrial variations in muscle synergy activations might arisefrom descending influences such as expectation, habituation, oremotion, which are factors that have been shown to affectEMG responses (Carpenter et al. 2004; Woollacott and Shum-way-Cook 2002). Alternately, the influence of biomechanicalcontext in muscle synergy recruitment that we observe couldexplain the intertrial variations in EMG patterns. It is possiblethat trial-by-trial variations were due to small variations inbiomechanical configuration at the onset of the perturbations,since we did not strictly regulate the initial postural configu-
ration of the subject. Likewise, it is possible that the trial-by-trial variations we observed were due to small differences inthe sensory inflow arising from small variations in biomechani-cal configuration from one trial to the next.
Why are muscle synergies useful?
Muscle synergies may provide a simplifying mechanism formuscle coordination, not because they reduce the number ofvariables controlled by the nervous system, but because theymap high-level task goals into actions. Control of task-levelvariables by muscle synergies has been identified in reachingstudies showing modular control of limb endpoint forces(Georgopoulos et al. 1992) or endpoint motions (Holdefer andMiller 2002; Schwartz et al. 1988; Scott and Kalaska 1997). Inbalance control, this mapping would allow standing balancethrough the control of encoded task variables rather thanindividual muscles. Accordingly, muscle activity in balancecontrol is modulated over time in response to changes in CoMkinematics rather than individual joint motions (Lockhart andTing 2007; Welch and Ting 2008). Moreover, we have foundconsistent relationships between the recruitment of musclesynergies and the modulation of endpoint forces across pos-tural configurations (Torres-Oviedo et al. 2006). Thus thenervous system might use these relationships to reliably coor-dinate the redundant structures of the limb across differentbiomechanical contexts. Muscle synergies controlling taskvariables would thus allow for the rapid reconfiguration ofmuscle coordination patterns in a context-dependent manner.This idea is analogous to the concepts in sparse coding of
1W
2W
3W
4W
5W
6W
7W W
rc
WL1 1
W
2W
3W
4W
5W
6W
7W W
r c
WL1 1
W
2W
3W
4W
5W
6W
7W W
rc
WL1
Variability accounted (VAF) by each synergy per stance
0
10
20
30
40
50 Narrow
0
10
20
30
40
50 Normal
0
10
20
30
40
50 Wide
0
10
20
30
40
50 Widest
0
10
20
30
40
50 Crouched
0
10
20
30
40
50 1L
VAF
VAF
Variability accounted for by red synergy (i.e., W1) in a single subjectMean variability accounted by each muscle synergy
FIG. 9. Muscle synergy contributions to EMG responses at each stance configuration averaged across subjects. Muscle synergies are recruited differentlyacross subjects, as shown by the spread in colored dots indicating the mean contribution of each synergy in a particular subject in each condition. However, similarrecruitment profiles of muscle synergies were observed for the narrow, normal, wide, and widest stance. Although most of the muscle synergies contribute tothe EMG responses in the crouched and one-leg stances, the recruitment profiles of muscle synergies are very different from those observed in the other stances.
3096 G. TORRES-OVIEDO AND L. H. TING
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

sensory inputs (Olshausen and Field 2004), where representa-tions of images are based on a small set of basis functionsselected from a large library. Therefore a few muscle synergiesmay be controlled by the nervous system in any given motorbehavior, but a whole library of muscle synergies may beavailable for the construction of movements (Chiel et al. 2009;McKay et al. 2007). Muscle synergies may therefore provide aconsistent mapping between task-level and execution-levelcontrol of balance. Such a modular organization may beadvantageous in reducing the number of output variables thatare modulated during the rapid adaptation to changing taskdemands (Fiete et al. 2004; McKay et al. 2007), by providingfaster convergence to new solutions during learning (Chhabraand Jacobs 2006; Todorov and Ghahramani 2004b). Usingmotor modules that define task-level actions may therefore beadvantageous for the adaptive neural control of movement.
A C K N O W L E D G M E N T S
We thank J. L. McKay for insightful conversations during the preparationof this manuscript.
Present address of G. Torres-Oviedo: Department of Neurology at KennedyKrieger Institute and Johns Hopkins University, 707 N. Broadway, Baltimore,MD 21201.
G R A N T S
This work was supported by National Institute of Neurological Disordersand Stroke Grant NS-058322.
D I S C L O S U R E S
No conflicts of interest are declared by the authors.
R E F E R E N C E S
Aflalo TN, Graziano MSA. Possible origins of the complex topographicorganization of motor cortex: reduction of a multidimensional space onto atwo-dimensional array. J Neurosci 26: 6288–6297, 2006.
Aflalo TN, Graziano MSA. Relationship between unconstrained arm move-ments and single-neuron firing in the macaque motor cortex. J Neurosci 27:2760–2780, 2007.
Berniker M, Jarc A, Bizzi E, Tresch MC. Simplified and effective motorcontrol based on muscle synergies to exploit musculoskeletal dynamics.Proc Natl Acad Sci USA 106: 7601–7606, 2009.
Bernstein N. The Coordination and Regulation of Movements. New York:Pergamon Press, 1967.
Bunderson NE, Burkholder TJ, Ting LH. Reduction of neuromuscularredundancy for postural force generation using an intrinsic stability crite-rion. J Biomech 41: 1537–1544, 2008.
Burtner PA, Qualls C, Woollacott MH. Muscle activation characteristics ofstance balance control in children with spastic cerebral palsy. Gait Posture8: 163–174, 1998.
Carpenter MG, Allum JHJ, Honegger F. Directional sensitivity of stretchreflexes and balance corrections for normal subjects in the roll and pitchplanes. Exp Brain Res 129: 93–113, 1999.
Carpenter MG, Frank JS, Adkin AL, Paton A, Allum JH. Influence ofpostural anxiety on postural reactions to multi-directional surface rotations.J Neurophysiol 92: 3255–3265, 2004.
Cheung VC, d’Avella A, Bizzi E. Adjustments of motor pattern for loadcompensation via modulated activations of muscle synergies during naturalbehaviors. J Neurophysiol 101: 1235–1257, 2009a.
Cheung VC, Piron L, Agostini M, Silvoni S, Turolla A, Bizzi E. Stability ofmuscle synergies for voluntary actions after cortical stroke in humans. ProcNatl Acad Sci USA 106: 19563–19568, 2009b.
Cheung VCK, d’Avella A, Tresch MC, Bizzi E. Central and sensorycontributions to the activation and organization of muscle synergies duringnatural motor behaviors. J Neurosci 25: 6419–6434, 2005.
Chhabra M, Jacobs RA. Properties of synergies arising from a theory ofoptimal motor behavior. Neural Comput 18: 2320–2342, 2006.
Chiel HJ, Ting LH, Ekeberg O, Hartmann MJ. The brain in its body: motorcontrol and sensing in a biomechanical context. J Neurosci 29: 12807–12814, 2009.
Churchland MM, Santhanam G, Shenoy KV. Preparatory activity in pre-motor and motor cortex reflects the speed of the upcoming reach. JNeurophysiol 96: 3130–3146, 2006.
Clark DJ, Ting LH, Zajac FE, Neptune RR, Kautz SA. Merging of healthymotor modules predicts reduced locomotor performance and muscle coor-dination complexity post-stroke. J Neurophysiol 103: 844–857, 2010.
d’Avella A, Bizzi E. Shared and specific muscle synergies in natural motorbehaviors. Proc Natl Acad Sci USA 102: 3076–3081, 2005.
Diener HC, Horak FB, Nashner LM. Influence of stimulus parameters onhuman postural responses. J Neurophysiol 59: 1888–1905, 1988.
Fagg AH, Shah A, Barto AG. A computational model of muscle recruitmentfor wrist movements. J Neurophysiol 88: 3348–3358, 2002.
Fiete IR, Hahnloser RHR, Fee MS, Seung HS. Temporal sparseness of thepremotor drive is important for rapid learning in a neural network model ofbirdsong. J Neurophysiol 92: 2274–2282, 2004.
Flash T, Hochner B. Motor primitives in vertebrates and invertebrates. CurrOpin Neurobiol 15: 660–666, 2005.
Gentner R, Classen J. Modular organization of finger movements by thehuman central nervous system. Neuron 52: 731–742, 2006.
Georgopoulos AP, Ashe J, Smyrnis N, Taira M. The motor cortex and thecoding of force. Science 256: 1692–1695, 1992.
Gollhofer A, Horstmann GA, Berger W, Dietz V. Compensation of trans-lational and rotational perturbations in human posture: stabilization of thecentre of gravity. Neurosci Lett 105: 73–78, 1989.
Golowasch J, Goldman MS, Abbott LF, Marder E. Failure of averaging inthe construction of a conductance-based neuron model. J Neurophysiol 87:1129–1131, 2002.
Graziano MS, Aflalo TN. Rethinking cortical organization: moving awayfrom discrete areas arranged in hierarchies. Neuroscientist 13: 138–147,2007.
Hamilton AF, Jones KE, Wolpert DM. The scaling of motor noise withmuscle strength and motor unit number in humans. Exp Brain Res 157:417–430, 2004.
Hart CB, Giszter SF. A neural basis for motor primitives in the spinal cord.J Neurosci 30: 1322–1336, 2010.
Henry SM, Fung J, Horak FB. EMG responses to maintain stance duringmultidirectional surface translations. J Neurophysiol 80: 1939–1950, 1998.
Henry SM, Fung J, Horak FB. Effect of stance width on multidirectionalpostural responses. J Neurophysiol 85: 559–570, 2001.
Holdefer RN, Miller LE. Primary motor cortical neurons encode functionalmuscle synergies. Exp Brain Res 146: 233–243, 2002.
Honeycutt CF, Gottschall JS, Nichols TR. Electromyographic responsesfrom the hindlimb muscles of the decerebrate cat to horizontal supportsurface perturbations. J Neurophysiol 101: 2751–2761, 2009.
Horak FB, Nashner LM. Central programming of postural movements:adaptation to altered support-surface configurations. J Neurophysiol 55:1369–1381, 1986.
Inglis JT, Horak FB, Shupert CL, Jones-Rycewicz C. The importance ofsomatosensory information in triggering and scaling automatic posturalresponses in humans. Exp Brain Res 101: 159–164, 1994.
Kakei S, Hoffman DS, Strick PL. Muscle and movement representations inthe primary motor cortex. Science 285: 2136–2139, 1999.
Kargo WJ, Ramakrishnan A, Hart CB, Rome LC, Giszter SF. A simpleexperimentally based model using proprioceptive regulation of motor prim-itives captures adjusted trajectory formation in spinal frogs. J Neurophysiol103: 573–590, 2010.
Kurtzer I, Pruszynski JA, Herter TM, Scott SH. Primate upper limbmuscles exhibit activity patterns that differ from their anatomical actionduring a postural task. J Neurophysiol 95: 493–504, 2006.
Kutch JJ, Kuo AD, Bloch AM, Rymer WZ. Endpoint force fluctuationsreveal flexible rather than synergistic patterns of muscle cooperation. JNeurophysiol 100: 2455–2471, 2008.
Lee DD, Seung HS. Algorithms for non-negative matrix factorization. AdvNeural Info Proc Syst 13: 556–562, 2001.
Lockhart DB, Ting LH. Optimal sensorimotor transformations for balance.Nat Neurosci 10: 1329–1336, 2007.
McCrea DA, Rybak IA. Organization of mammalian locomotor rhythm andpattern generation. Brain Res Rev 57: 134–146, 2008.
McGowan CP, Neptune RR, Clark DJ, Kautz SA. Modular control ofhuman walking: adaptations to altered mechanical demands. J Biomech 43:412–419, 2010.
3097INVARIANT MUSCLE SYNERGIES ACROSS POSTURAL CONFIGURATIONS
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from

McKay JL, Burkholder TJ, Ting LH. Biomechanical capabilities influencepostural control strategies in the cat hindlimb. J Biomech 40: 2254–2260, 2007.
McKay JL, Ting LH. Functional muscle synergies constrain force productionduring postural tasks. J Biomech 41: 299–306, 2008.
Medina JF, Lisberger SG. Variation, signal, and noise in cerebellar sensory-motor processing for smooth-pursuit eye movements. J Neurosci 27: 6832–6842, 2007.
Nashner LM. Fixed patterns of rapid postural responses among leg musclesduring stance. Exp Brain Res 30: 13–24, 1977.
Neptune RR, Clark DJ, Kautz SA. Modular control of human walking: asimulation study. J Biomech 42: 1282–1287, 2009.
Olshausen BA, Field DJ. Sparse coding of sensory inputs. Curr OpinNeurobiol 14: 481–487, 2004.
Overduin SA, d’Avella A, Roh J, Bizzi E. Modulation of muscle synergyrecruitment in primate grasping. J Neurosci 28: 880–892, 2008.
Prinz AA, Bucher D, Marder E. Similar network activity from disparatecircuit parameters. Nat Neurosci 7: 1345–1352, 2004.
Raasch CC, Zajac FE. Locomotor strategy for pedaling: muscle groups andbiomechanical functions. J Neurophysiol 82: 515–525, 1999.
Saltiel P, Wyler-Duda K, D’Avella A, Tresch MC, Bizzi E. Muscle syner-gies encoded within the spinal cord: evidence from focal intraspinal NMDAiontophoresis in the frog. J Neurophysiol 85: 605–619, 2001.
Schepens B, Drew T. Independent and convergent signals from the pontomed-ullary reticular formation contribute to the control of posture and movementduring reaching in the cat. J Neurophysiol 92: 2217–2238, 2004.
Schepens B, Drew T. Descending signals from the pontomedullary reticularformation are bilateral, asymmetric, and gated during reaching movementsin the cat. J Neurophysiol 96: 2229–2252, 2006.
Schepens B, Stapley P, Drew T. Neurons in the pontomedullary reticularformation signal posture and movement both as an integrated behavior andindependently. J Neurophysiol 100: 2235–2253, 2008.
Schwartz AB, Kettner RE, Georgopoulos AP. Primate motor cortex and freearm movements to visual targets in three-dimensional space. I. Relationsbetween single cell discharge and direction of movement. J Neurosci 8:2913–2927, 1988.
Scott SH. Optimal feedback control and the neural basis of volitional motorcontrol. Nat Rev Neurosci 5: 534–546, 2004.
Scott SH, Kalaska JF. Reaching movements with similar hand paths butdifferent arm orientations. I. Activity of individual cells in motor cortex. JNeurophysiol 77: 826–852, 1997.
Sober SJ, Wohlgemuth MJ, Brainard MS. Central contributions to acousticvariation in birdsong. J Neurosci 28: 10370–10379, 2008.
Soechting JF, Flanders M. Sensorimotor control of contact force. Curr OpinNeurobiol 18: 565–572, 2008.
Stapley PJ, Drew T. The pontomedullary reticular formation contributes tothe compensatory postural responses observed following removal of thesupport surface in the standing cat. J Neurophysiol 101: 1334–1350, 2009.
Stapley PJ, Ting LH, Hulliger M, Macpherson JM. Automatic posturalresponses are delayed by pyridoxine-induced somatosensory loss. J Neuro-sci 22: 5803–5807, 2002.
Ting LH, Kautz SA, Brown DA, Zajac FE. Phase reversal of biomechanicalfunctions and muscle activity in backward pedaling. J Neurophysiol 81:544–551, 1999.
Ting LH, Macpherson JM. Ratio of shear to load ground-reaction force mayunderlie the directional tuning of the automatic postural response to rotationand translation. J Neurophysiol 92: 808–823, 2004.
Ting LH, McKay JL. Neuromechanics of muscle synergies for posture andmovement. Curr Opin Neurobiol 17: 622–628, 2007.
Todorov E, Ghahramani Z. Analysis of the synergies underlying complexhand manipulation. Conf Proc IEEE Eng Med Biol Soc 6: 4637–4640,2004a.
Todorov E, Ghahramani Z. Unsupervised learning of sensory-motor primi-tives. In: Proceedings of the 26th Annual International Conference of theIEEE Engineering Medicine and Biology Society. Piscataway, NJ: IEEE,2004b, p. 1750–1753.
Torres-Oviedo G, Macpherson JM, Ting LH. Muscle synergy organizationis robust across a variety of postural perturbations. J Neurophysiol 96:1530–1546, 2006.
Torres-Oviedo G, Ting LH. Muscle synergies characterizing human posturalresponses. J Neurophysiol 98: 2144–2156, 2007.
Tresch MC, Cheung VC, d’Avella A. Matrix factorization algorithms for theidentification of muscle synergies: evaluation on simulated and experimentaldata sets. J Neurophysiol 95: 2199–2212, 2006.
Tresch MC, Jarc A. The case for and against muscle synergies. Curr OpinNeurobiol 19: 601–607, 2009.
Tropp H, Odenrick P. Postural control in single-limb stance. J Orthop Res 6:833–839, 1988.
Valero-Cuevas FJ. Predictive modulation of muscle coordination patternmagnitude scales fingertip force magnitude over the voluntary range. JNeurophysiol 83: 1469–1479, 2000.
Valero-Cuevas FJ, Zajac FE, Burgar CG. Large index-fingertip forces areproduced by subject-independent patterns of muscle excitation. J Biomech31: 693–703, 1998.
Welch TD, Ting LH. A feedback model reproduces muscle activity duringhuman postural responses to support-surface translations. J Neurophysiol99: 1032–1038, 2008.
Woollacott M, Shumway-Cook A. Attention and the control of posture andgait: a review of an emerging area of research. Gait Posture 16: 1–14, 2002.
Yanai Y, Adamit N, Israel Z, Harel R, Prut Y. Coordinate transformation isfirst completed downstream of primary motor cortex. J Neurosci 28: 1728–1732, 2008.
Zar JH. Biostatistical Analysis. Upper Saddle River, NJ: Prentice-Hall, 1999.
3098 G. TORRES-OVIEDO AND L. H. TING
J Neurophysiol • VOL 103 • JUNE 2010 • www.jn.org
on June 14, 2010 jn.physiology.org
Dow
nloaded from