gene expression levels of selected factors in hacat...
TRANSCRIPT

Gene expression levels of selected factors in HaCaT
cells upon treatment with plant extracts against hair
loss
H. Aydın1, B. Akyıldırım1, N. Çelik1, H. Sevinç1, M. Kartal2 and M. Türkoğlu1
1Biota Laboratories, Sancaktepe, 34785 Istanbul, Turkey
2Bezmialem Vakıf University, Bezmialem Center for Education, Research and Practice in Phytotherapy
Abstract
Background: Hair loss (alopecia) is a serious problem due to many different reasons. It is of
great interest; therefore, to develop new therapies for treating alopecia and for enhancing hair
growth. Treatments that are utilizing natural products such as plants and their derivatives are
attracting more attention especially studying the mechanism at the cellular level.
Objective: The objective of this study was to examine the hair-growth effect of plant extracts
that have been either traditionally used for treating hair loss or selected through a literature
review. This collection includes the extracts of Myrtus communis L., Urtica dioica L.,
Rosmarinus officinalis L., Raphanus sativus L., Lepidium sativum L., and Ficus carica L..
Our investigation was based on the changes in the gene expression levels of selected factors
such as VEGF, SRD5α (5α-reductase), and IL-1α in HaCaT cells treated with the plants
extracts.
Methods and Results: Plant extracts were prepared with different methods such as Soxhlet
and decoction in different solvents such as water, water-ethanol or oil. Non-cytotoxic
concentrations of the extracts were determined by Cell Proliferation Assay using XTT
reagent. Human keratinocyte (HaCaT) cells were then incubated with the extracts in the
determined concentrations. RNA isolations were carried out for both untreated and treated
cells by using Tri-reagent. Gene expression levels of the factors that have been implicated in
hair growth control were measured by RT-qPCR analysis. Extracts were tested on HaCaT
cells and the effects of these extracts on the VEGF, SRD5α, TNF-α, and IL-1α gene
expression levels were monitored. Among these extracts, Urtica dioica, Ficus carica and
Lepidium sativum led to significant inhibition of SRD5a gene expression (p <0.05). Myrtus
communis, on the other hand, significantly increased the expression level of VEGF.
Discussion and Conclusion: SRD5α gene expression was found to be significantly
downregulated by Urtica dioica, Lepidium sativum and Ficus carica extracts implying the
potential inhibitory effects of the extracts on androgenic activity in HaCaT cells. Upregulation
of VEGF expression by Myrtus communis extract represented the potential hair-growth effect
of the extract. According to these preliminary results, these plant extracts are promising
candidates for enhancing hair growth hence may be used as therapeutic agents against hair
loss. As a result, this study will give an insight about potential hair-growth, anti-inflammatory
and anti-androgenic effects of the selected plants extracts.

Introduction
The presence of hair is characteristic for mammals since it exerts a wide range of functions
including physical protection, thermal insulation, sensory functions and social interactions.
For humans, hair is of greater importance and there are many human diseases associated with
hair loss. Hair loss, also known as alopecia, is a common and distressing problem both for
men and women. The effect of hair disorders on patients’ psychological wellbeing and on
social interactions cannot be ignored. Therefore, treatments for hair loss attract considerable
attention and it is a growing research area which is heavily invested in (Cotsarelis and Millar,
2001). In order to treat hair loss, scientists try to have a molecular understanding of hair loss.
Hair follicle is responsible for the production of hair shaft. The formation of hair follicle takes
place during embryogenesis: hair follicle develops from the embryonic epidermis. Then the
keratinocyte cells of hair follicle differentiate into three enclosed cylindrical parts: the central
most layer forms the hair shaft (fiber), the middle layer forms the inner root sheath (IRS) and
the outermost cylinder forms the outer root sheath (ORS) (Stenn and Paus, 2001; Rogers,
2004). The IRS forms the hair shaft and directs its outward movement whereas the ORS
separates the whole structure from the dermis (Stenn and Paus, 2001). The hair follicle is
remodeled under the control of repetitive cycles of growth (anagen), regression (catagen), rest
(telogen) and shedding (exogen) (Cotsarelis and Millar, 2001). In anagen phase, follicular
keratinocyte cells proliferate rapidly and hair shaft thickens and elongates. Therefore, the
duration of anagen phase determines the length of the hair shaft (Price, 1999). During
catagen, the epithelial cells at the base of the follicle undergo programmed cell death
(apoptosis) reducing the size of the follicle as it enters the resting phase (telogen) (Cotsarelis
and Millar, 2001). At the end of telogen, hair is released and shed in exogen phase and the
next cycle is initiated (Price, 1999).
The cyclic regeneration of hair is thought to be affected and controlled by cyclic cellular
mechanisms. Besides the genetic factors and dietary changes, there are a number of
significant factors which are thought to be responsible for the cellular signaling mechanisms
underlying the hair growth cycle. The hair follicle is highly sensitive to numerous growth
factors, cytokines, neuropeptides and hormones (Schneider et al., 2009). Growth factors such
as vascular endothelial growth factor (VEGF), insulin-like growth factor (IGF-1) and
keratinocyte growth factor (KGF) are thought to be important for anagen maintenance.
VEGF, for example, is best known for its role in angiogenesis which, in turn, promotes cell
proliferation. VEGF is also identified as a major mediator of hair follicle growth and showed
to improve follicle vascularization promoting hair growth (Yano et al., 2001). KGF, as a
member of fibroblast growth factor (FGF) family, was found to be an important endogenous
mediator of normal hair growth and development (Danilenko et al., 1995). On the other hand,
cytokines, such as tumor necrosis factor-α (TNF-α), interleukin-1α/β (IL-1α/β) and interferon-
γ (IFN-γ), have been described to induce catagen either by inducing apoptosis (TNF-α) or by
increasing inflammation (IL-1α/β, IFN-γ) (Schneider et al., 2009; Konur et al., 2005). In
addition to these factors, stereoid 5α-reductase, an enzyme that catalyzes the conversion of
testosterone into 5α-dihydrotestosterone (DHT), was found to play an important role in hair

growth control. The conversion of testosterone into DHT amplifies the androgenic signal in
androgen-related target tissues. Previously, prostate was known to be one of these target
tissues which is rich in 5α-reductase; however, the studies in the 1970s and 1980s have stated
that the skin is also rich in 5α-reductase. Later, it was found that 5α-reductase activity was
associated with the androgen-dependent skin disorders such as acne vulgaris and androgenetic
alopecia (Chen et al., 1998).
Based on this knowledge about the molecular mechanisms of hair growth, treatments for hair
loss basically aim to prolong anagen, convert telogen follicles to anagen, prevent apoptosis of
follicle keratinocytes during catagen and, if possible, generate new follicles. At molecular
level, these therapeutic approaches include regulation of growth factors, cytokines and/or
other signaling molecules that are known to have roles in hair growth. Despite the ever-
increasing demand for treating hair loss, there are only two clinically proven and FDA-
approved treatments for hair loss: finasteride and minoxidil. Finasteride is a highly specific
5α-reductase (type 2) inhibitor whereas minoxidil is a nonspecific drug (Price, 1999).
However, finasteride is approved for the treatment of androgenetic alopecia only in men while
minoxidil is proven to be useful in both men and women with various conditions such as
androgenetic alopecia and alopecia areata (Price, 1999). In order to develop new therapeutic
approaches that overcome the limitations of finasteride and minoxidil current studies require
better understanding of hair biology. Besides the drugs, use of natural products, such as plants
and their derivatives, for hair loss treatment have been gathered ever-increasing attention as in
the case of many other disorders.
Plants are inseparable part of both natural ingredients of drugs and alternative, complementary
medicine which have become popular in the last 10-15 years. Throughout history, plants and
botanical extracts have been used in order to treat diseases within the various kind of
formulation by humans. New innovative hair products are also developed from natural
products, especially from plants. Botanical extracts that support the health, texture and
integrity of hair are widely used in cosmetic formulations.
Turkey is rich in flora and approximately 12.000 varieties of plants are growing naturally
(Davis et al., 1965-85; 1988; Güner et al., 2000; Özhatay and Kültür 2006; Özhatay et al.
2009). Local people are using the plants that are growing naturally in proximity for many
different purposes. “People living in the vicinity of a region to exploit information to meet the
various needs of plants and the effect on plants’ can be used as the definition of the term of
ethnobotany. Recently, the studies performed utilization of the plants for different purposes in
Turkey. Studies stated that nearly 500 plants are used for medicinal purposes.
In the present study we examined the hair-growth effect of plant extracts that have been either
traditionally used for treating hair loss or selected through a literature. This collection
includes the extracts of Myrtus communis, Urtica dioica, Rosmarinus officinalis, Raphanus
sativus, Ficus carica and Lepidium sativum. Although some of the selected plant has proven
effects in different areas, insufficient scientific data on the hair. Our investigation was based
on the changes in the gene expression levels of selected factors (VEGF, SRD5α (5α-
reductase), IL-1α and TNF-α in HaCaT cells treated with the plants extracts.

Materials & Methods Preparation of Plant Extracts
The different parts of plants were collected from Turkey. These plants species listed below in
the Table 1. These plants were identified using the stereomicroscope and Flora of Turkey and
the Aegean Islands.
Latin name of plant The part of used
Raphanus sativus Rhizomes
Lepidium sativum Leaf
Ficus carica Leaf
Urtica dioica Leaf
Myrtus communis Leaf
Rosmarinus officinalis Leaf
Table 1: Plants species and their parts of used
- Raphanus sativus fresh rhizomes juice was prepared.
- Lepidium sativum fresh leaf was fine-cut and extracted with 50% distilled water and
50% ethanol with pressure extractor for 120 minutes at 15°C.
- Ficus carica fresh leaf was fine-cut and extracted with 100% distilled water for 3
hours at 50°C.
- Urtica dioica dried leaf was fine-cut and extracted with 100% distilled water for 3
hours at 50°C.
- Myrtus communis dried leaf was fine-cut and extracted with 100% distilled water for 3
hours at 50°C.
- Rosmarinus officinalis dried leaf was fine-cut and extracted with 30% distilled water
and 70% ethanol with pressure extractor for 120 minutes at 15°C.
The extracts were filtered through a filter paper into a sterile bottle.
Cell Culture
The human keratinocyte cell line (HaCaT) was cultured in Dulbecco’s modified Eagle’s
medium with high glucose supplemented with 10% heat-inactivated fetal bovine serum (FBS)
and 100U/ml gentamicin. Cells were maintained at 37℃ in a humidified atmosphere at 5%
CO2 in Newbrunswick incubator. All supplements and media were purchased from Sigma
Aldrich (Germany).
XTT Cell Proliferation Assay
Roche XTT Cell Proliferation kit was used for cytotoxicity assay. HaCaT cells were seeded
into 96 well plates (1x104 cells/well) and were subjected to different concentrations of plant
extracts. XTT and activator reagents were added to the plates after 72 hour incubation period
according to the manufacturer’s instructions. Then, cells were incubated at 37℃ for 4 hours in
order that XTT reagent was reduced to formazan compound. The optical density of soluble
formazan compound was measured at 495 nm with a BIO-RAD microplate reader (Japan)
(650 nm as reference wavelength).

RNA Isolation
Total RNA was extracted from cells treated with plant extracts and non-treated cells by using
TRI-reagent according to manufacturer’s instructions (Sigma Aldrich, USA). The
concentration and purity of isolated RNA samples were determined by measuring optical
densities at 260 nm and 280 nm using Bio-spect nano (Japan).
Reverse Transcription
Roche Transcriptor First Strand cDNA Synthesis Kit was used for reverse transcription.
cDNA synthesis was performed with 500 ng total RNA, 2 μM final concentration of gene
specific primers of VEGF, IL-1α, 5α- reductase, TNF-α and GAPDH (Integrated DNA
Technologies), 10 U of Transcriptor Reverse Transcriptase, 20 U of Protector RNase
Inhibitor, 1mM each of dNTP mix and Transcriptor Reverse Transcription Buffer (5X)
according to the manufacturer’s instructions (Roche).
Real-Time Quantitative Polymerase Chain Reaction
Real-time PCR (qPCR) reaction was carried out in Light Cycler 96 (Roche). And Fast Start
DNA Green Master Kit (Roche) was used. Briefly, total volume of reaction mix was 20 ul
containing 10 μl SYBR Green Master Mix (2X), 0.5 μM of reverse and forward primers, 2.5
ng cDNA and appropriate amount of nuclease free water. All samples were run as triplicates
in each run including a non-template control and four standards (1:1, 1:10, 1:100, 1:1000).
The PCR parameters were determined separately for each target according to melting and
annealing temperatures of primers. Each parameter includes a pre-incubation step for 10 min
at 95°C and followed by 45 cycles 3-step Amplification and melting step. Melting curve
analysis was performed to verify specificity. For each factor upon treatment with each extract,
five repeats of gene expression analysis was done (n=5). For quantitation of RT-qPCR results,
ΔΔCt method was used (2-ΔΔCt
).
Statistical Analysis
All data are representative of five repeats (n=5) and expressed as mean ±standard error of the
means (SEM). Statistical evaluation was performed by Unpaired t-test using Graph Pad Prism
5 Software (USA) and the results with p value less than 0.05 were accepted as significant.
Results
Cytotoxicity Analysis
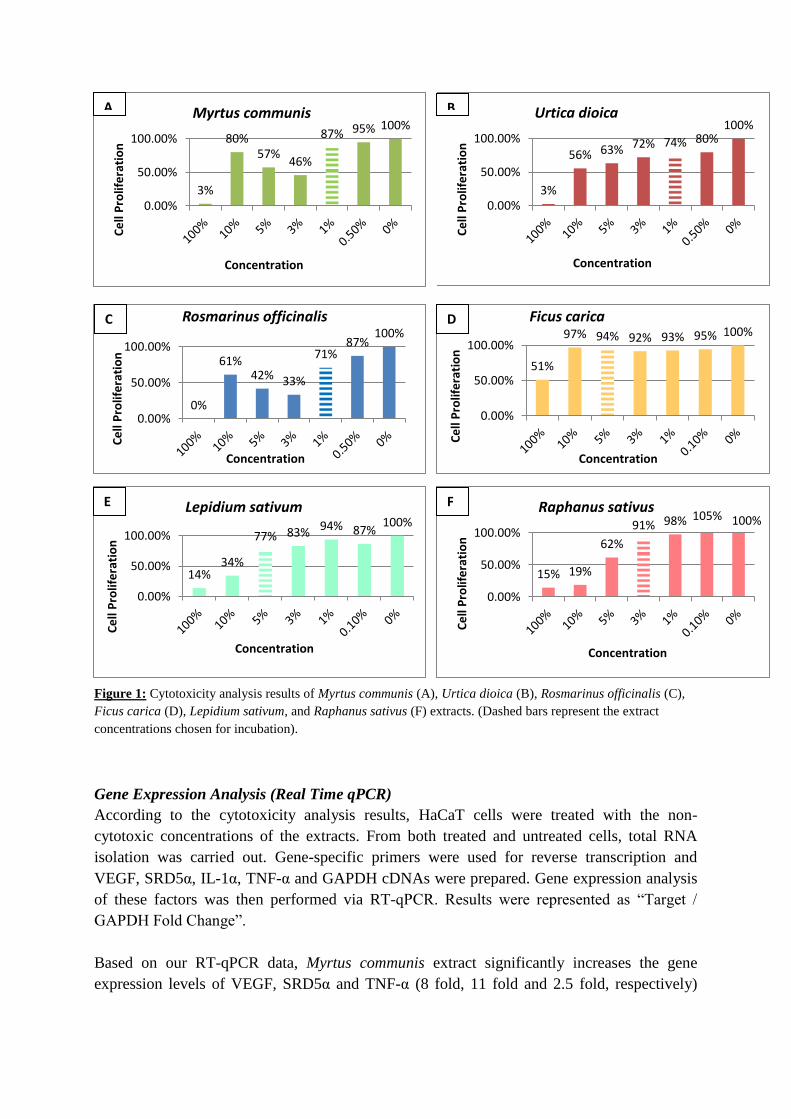
Based on cell proliferation ratios of extract-treated cells with respect to the control cells,
cytotoxicity levels of the extracts were determined. Among all six extracts, Ficus carica
extract was found to have the minimum cytotoxic effect on HaCaT cells. For Myrtus
communis, Urtica dioica and Rosmarinus officinalis extracts, 1% concentration was chosen as
non-cytotoxic dose. 3% concentration was chosen for Raphanus sativus and 5% concentration
was chosen for Lepidium sativum and Ficus carica (Figure 1).
3%
80% 57%
46%
87% 95% 100%
0.00%
50.00%
100.00%
Ce
ll P
rolif
era
tio
n
Concentration
Myrtus communis
0%
61% 42% 33%
71% 87%
100%
0.00%
50.00%
100.00%
Ce
ll P
rolif
era
tio
n
Concentration
Rosmarinus officinalis
51%
97% 94% 92% 93% 95% 100%
0.00%
50.00%
100.00%
Ce
ll P
rolif
era
tio
n
Concentration
Ficus carica
14% 34%
77% 83% 94% 87% 100%
0.00%
50.00%
100.00%
Ce
ll P
rolif
era
tio
n
Concentration
Lepidium sativum
15% 19%
62%
91% 98% 100%
0.00%
50.00%
100.00%
Ce
ll P
rolif
era
tio
n
Concentration
Raphanus sativus
Figure 1: Cytotoxicity analysis results of Myrtus communis (A), Urtica dioica (B), Rosmarinus officinalis (C),
Ficus carica (D), Lepidium sativum, and Raphanus sativus (F) extracts. (Dashed bars represent the extract
concentrations chosen for incubation).
Gene Expression Analysis (Real Time qPCR)
According to the cytotoxicity analysis results, HaCaT cells were treated with the non-
cytotoxic concentrations of the extracts. From both treated and untreated cells, total RNA
isolation was carried out. Gene-specific primers were used for reverse transcription and
VEGF, SRD5α, IL-1α, TNF-α and GAPDH cDNAs were prepared. Gene expression analysis
of these factors was then performed via RT-qPCR. Results were represented as “Target /
GAPDH Fold Change”.
Based on our RT-qPCR data, Myrtus communis extract significantly increases the gene
expression levels of VEGF, SRD5α and TNF-α (8 fold, 11 fold and 2.5 fold, respectively)
3%
56% 63% 72% 74% 80% 100%
0.00%
50.00%
100.00%
Ce
ll P
rolif
era
tio
n
Concentration
Urtica dioica
105%
A B
C D
E F

whereas it causes a significant reduction in IL-1α expression level, compared to untreated
control cells. (Figure 2, Table 2).
Urtica dioica extract, did not significantly affect the expression levels of VEGF, IL-1α and
TNF-α. However, it was found to downregulate 5α-reductase since it reduced gene expression
level of SRD5α almost by half, compared to untreated control cells (Figure 3, Table 2).
Myrtus
communis
VEGF 8,384 ± 1,04 (P=0,0021)
SRD-5α 11,66 ± 0,692 (P=0,0001)
TNF-α 2,623 ± 0,6035 (P=0,0361)
IL-1α 0,4500 ± 0,09891 (P=0,0014)
Urtica dioica
VEGF 0,6102 ± 0,1415 (P=0,0511)
SRD-5α 0,5873 ± 0,05858 (P=0,0021)
TNF-α 492,5 ± 314,2 (P=0,1688)
IL-1α 176,4 ± 86,10 (P=0,0878)
Control Myrtus communis
Control Myrtus communis Control Myrtus communis
Control Myrtus communis
Figure 2: Gene expression levels of VEGF (A), TNF-α (B), SRD5α (C) and
IL-1α (D) in Myrtus communis extract-treated HaCaT cells
Table 2: Fold changes in the
expression levels of VEGF, SRD5α,
TNF-α and IL-1α genes
Control Urtica dioica Control Urtica dioica
Control Urtica dioica Control Urtica dioica
Figure 3: Gene expression levels of VEGF (A), TNF-α (B), SRD5α (C) and
IL-1α (D) in Urtica dioica extract-treated HaCaT cells
Table 3: Fold changes in the
expression levels of VEGF, SRD5α,
TNF-α and IL-1α genes

Cells treated with Rosmarinus officinalis extract showed 31-fold increased expression
of TNF-a gene together with 7.5-fold increase in VEGF gene expression level compared to
untreated cells (Figure 4, Table 4).
Ficus carica extract, which was found to be the least cytotoxic extract of all studied, caused
significant reduction in expression levels of all genes of interest with the most reduction in
SRD5α gene expression (0.07-fold, p<0.0001) (Figure 5, Table 5).
Rosmarinus
officinalis
VEGF 7,450 ± 1,865 (P=0,0258)
SRD-5α 1,292 ± 0,06272 (P=0,0096)
TNF-α 31,01 ± 10,56 (P=0,0295)
IL-1α 0,8000 ± 0,1693 (P=0,2821)
Ficus carica
VEGF 0,2300 ± 0,04041 (p<0.0001)
SRD-5α 0,0700 ± 0,0200
(p<0.0001)
TNF-α 0,2433 ± 0,1033
(p=0.0019)
IL-1α 0,1867 ± 0,03844
(p<0.0001)
Control Control
Control Control
Rosmarinus officinalis
Rosmarinus officinalis
Rosmarinus officinalis
Rosmarinus officinalis
Figure 4: Gene expression levels of VEGF (A), TNF-α (B), SRD5α (C) and
IL-1α (D) in Rosmarinus officinalis extract-treated HaCaT cells
Table 4: Fold changes in the
expression levels of VEGF, SRD5α,
TNF-α and IL-1α genes
Figure 5: Gene expression levels of VEGF (A), TNF-α (B), SRD5α (C) and
IL-1α (D) in Ficus carica extract-treated HaCaT cells
Table 5: Fold changes in the
expression levels of VEGF, SRD5α,
TNF-α and IL-1α genes
Control Ficus carica Control Ficus carica
Control Ficus carica Control Ficus carica
A
C D
B
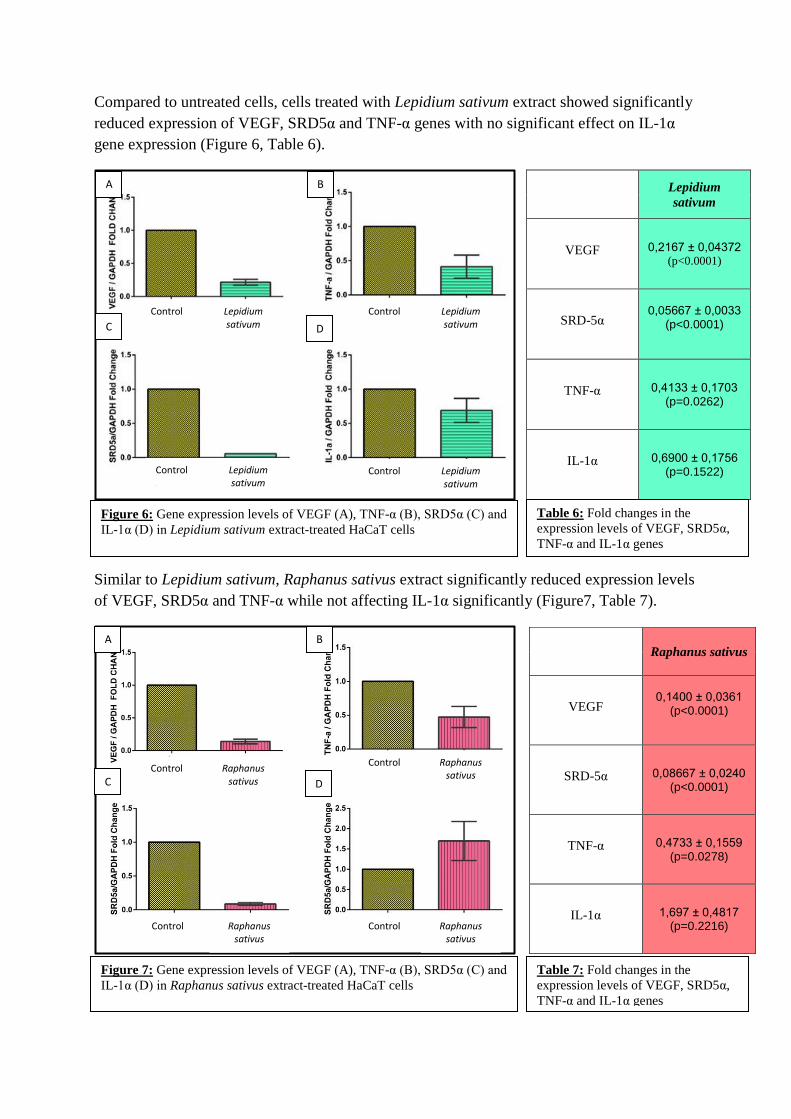
Compared to untreated cells, cells treated with Lepidium sativum extract showed significantly
reduced expression of VEGF, SRD5α and TNF-α genes with no significant effect on IL-1α
gene expression (Figure 6, Table 6).
Similar to Lepidium sativum, Raphanus sativus extract significantly reduced expression levels
of VEGF, SRD5α and TNF-α while not affecting IL-1α significantly (Figure7, Table 7).
Lepidium
sativum
VEGF 0,2167 ± 0,04372 (p<0.0001)
SRD-5α 0,05667 ± 0,0033
(p<0.0001)
TNF-α 0,4133 ± 0,1703 (p=0.0262)
IL-1α 0,6900 ± 0,1756 (p=0.1522)
Raphanus sativus
VEGF 0,1400 ± 0,0361
(p<0.0001)
SRD-5α 0,08667 ± 0,0240 (p<0.0001)
TNF-α 0,4733 ± 0,1559 (p=0.0278)
IL-1α 1,697 ± 0,4817 (p=0.2216)
A
C D
B
Control Lepidium sativum
Control Lepidium sativum
Control Lepidium sativum
Control Lepidium sativum
Figure 6: Gene expression levels of VEGF (A), TNF-α (B), SRD5α (C) and
IL-1α (D) in Lepidium sativum extract-treated HaCaT cells
Table 6: Fold changes in the
expression levels of VEGF, SRD5α,
TNF-α and IL-1α genes
A
C D
B
Control Raphanus sativus
Control Raphanus sativus
Control Raphanus sativus
Control Raphanus sativus
Figure 7: Gene expression levels of VEGF (A), TNF-α (B), SRD5α (C) and
IL-1α (D) in Raphanus sativus extract-treated HaCaT cells
Table 7: Fold changes in the
expression levels of VEGF, SRD5α,
TNF-α and IL-1α genes

Discussion
The demand for treating hair loss with natural products and the botanical wealth of Turkey are
the principal motivations of the present study. Additionally, the lack of studies about the
molecular effects of already-used plants on hair cells paved the way for this study.
There have been many plants commonly used by people for hair care. On the other hand,
there are only two clinically-approved treatments for hair disorders, each having limitations to
some extent. Taking these into account, developing new treatments that utilize plants and
demonstrating their effects on the molecular mechanism of hair growth become a remarkable
research area. In this study, extracts of Myrtus communis, Urtica dioica, Rosmarinus
officinalis, Raphanus sativus, Ficus carica and Lepidium sativum were chosen and tested for
their potential hair-growth effects. In order to demonstrate their effects, the changes in the
expression levels of VEGF, SRD5α, TNF-α, and IL-1α genes upon treatment were monitored.
Myrtus communis extract was found to increase the expression levels of all factors of interest
to some extent. By taking this overall increase into consideration, Myrtus communis extract
may be said to enhance HaCaT cell proliferation hence to upregulate synthesis of all factors.
Especially, the 8-fold increase in VEGF gene expression is remarkable since VEGF is a
growth factor that facilitates hair-growth. Taken together, these results demonstrated potential
hair-growth effect of Myrtus communis extract.
According to our results, Urtica dioica extract affected the gene expression level of only
SRD5α gene and it reduced SRD5α gene expression almost by half. This selective
downregulation of SRD5α gene by Urtica dioica extract represents the potential anti-
androgenic effect of Urtica dioica extract since SRD5α gene encodes for 5α-reductase
enzyme that converts testosterone into dihydrotestosteron which results with the shortening of
anagen phase. Urtica dioica, therefore, is a promising candidate plant to use in hair-loss
treatment, especially in men.
Upon treatment with Rosmarinus officinalis extract, HaCaT cells showed highly increased
levels of both VEGF (7.5-fold) and TNF-α (31-fold) genes. Even if it increases VEGF gene
expression, the 31-fold increase in the expression level of TNF-α gene, which is an apoptosis-
inducer, pointed out the anti-proliferative effect of Rosmarinus officinalis extract on HaCaT

cells. Therefore, Rosmarinus officinalis extract was not included into the list of plants that can
be used for hair-loss treatment.
In contrast to Myrtus communis, Ficus carica extract decreased to gene expression levels of
all factors of interest to some extent. In this regard, it can be said that Ficus carica extract
may have a negative effect on HaCaT cell proliferation hence decreased the synthesis of all
factors. However, the reduction in the expression level of SRD5α gene was the most among
all : its expression was reduced by almost one-fifteenth whereas the expression level of other
factors were reduced by one-fifth. Despite its overall negative effect on HaCaT cell
proliferation, Ficus carica extract, with its significant SRD5α downregulation, can have
potential anti-androgenic effect.
Lepidium sativum extract and Raphanus sativus extract treatments ended up with results
similar to those of Ficus carica just with the difference of no significant effect on IL-1α.
Therefore, Lepidium sativum and Raphanus sativus may also have anti-androgenic effects.
None of all plant extracts tested caused upregulation of VEGF and downregulation of other
factors at the same time. Such extract could be the ideal to use for hair-loss treatment.
Nevertheless, combination of two or more extracts with different but complementary effects
can give results like such an ideal extract can do. For example, Myrtus communis and Urtica
dioica could be a good combination.
This study gave an insight about potential hair-growth, anti-inflammatory and anti-androgenic
effects of the selected plants extracts. Further studies with different factors together with
clinical studies can pave way to use of this plant extracts as therapeutic agents against hair
loss.
Acknowledgements
This study was supported by TUBİTAK in the context of “Industrial R&D Projects Grant
Programme” with the project ID 3130970.

References
1. Cotsarelis, G., & Millar, S. (2001). Towards a molecular understanding of hair loss
and its treatment. Trends in Molecular Medicine, 7: 293-301.
2. Stenn, K. S., and R. Paus (2001). Controls of Hair Follicle Cycling. Physiological
Reviews, 81: 449–494.
3. Rogers, G. (2004). Hair follicle differentiation and regulation. The International
Journal of Developmental Biology Int. J. Dev. Biol., 48: 163-170.
4. Price, V. (1999). Treatment of Hair Loss (A. Wood, Ed.). The New England Journal of
Medicine, 341: 964-973.
5. Schneider, M., Schmidt-Ullrich, R., & Paus, R. (2009). The Hair Follicle as a
Dynamic Miniorgan. Current Biology, 19: 132-142.
6. Yano, K., Brown, L., & Detmar, M. (2001). Control of hair growth and follicle size by
VEGF-mediated angiogenesis. Journal of Clinical Investigation J. Clin. Invest., 107:
409-417.
7. Danilenko, D., Ring, B., Yanagihara, D., Benson, W., Wiemann, B., Starnes, C., &
Pierce, G. (1995). Keratinocyte Growth Factor Is an Important Endogenous Mediator
of Hair Follicle Growth, Development, and Differentiation. American Journal of
Pathology, 147:145-154.
8. Konur, A., Schulz, U., Eissner, G., Andreesen, R., & Holler, E. (2005). Interferon
(IFN)-gamma is a main mediator of keratinocyte (HaCaT) apoptosis and contributes to
autocrine IFN-gamma and tumour necrosis factor-alpha production. Br J Dermatol
British Journal of Dermatology, 152: 1134-1142.
9. Chen, W., Zouboulis, C., Fritsch, M., Blume-Peytavi, U., Kodelja, V., Goerdt, S.,
Luu-The, V., Orfanos, C. (1998). Evidence of Heterogeneity and Quantitative
Differences of the Type 1 5alpha-Reductase Expression in Cultured Human Skin Cells
- Evidence of its Presence in Melanocytes. J Invest Dermatol Journal of Investigative
Dermatology, 110: 84-89.
10. Davis, P.H., Miller, R.R., Tan, K., 1965-1985. Flora of Turkey and the Aegean
Islands. Vol. 1-9, Edinburgh University Press, Edinburgh,
11. Davis, P.H., Miller, R.R., Tan, K., 1988. Flora of Turkey and the Aegean Islands. 10
(Supplement I), Edinburgh University Press, Edinburgh.
12. Güner, A., Özhatay, N., Ekim, T., Başer, K.H.C., 2000. Flora of Turkey and the
Aegean Islands. Vol 11 (Supplement II), Edinburgh University Press, Edinburgh.
13. Özhatay,N. ,Kültür, Ş. 2006 Checklist of additional taxa to the Supplement Flora of
Turkey IIITurk. J. Bot. 30:281-316.
14. Özhatay N, Kültür, Ş.and Aslan S 2009 Checklist of additional taxa to the Supplement
Flora of Turkey IV Turk. J. Bot. 33:191-226.