gene regulation in two dimensions: achaete s.cute are
TRANSCRIPT

Gene regulation in two dimensions: the proneural achaete and s.cute genes are controlled by combmanons ot axis-patterning genes through a common intergenic control region James B. Skeath, Grace Panganiban, Jane Selegue, and Sean B. CarrolP
Howard Hughes Medical Institute, Laboratory of Molecular Biology, University of Wisconsin-Madison, Madison, Wisconsin 53706 USA
The mechanisms that generate precise patterns of discrete cell types within developing fields are not well understood. One model for analyzing how cells interpret positional information in two dimensions is the regulation of proneural cluster formation within insect segments. Two adjacent proneural regulatory genes, achaete and scute, are expressed coincidently in cell clusters at reproducible anteroposterior (AP) and dorsoventral (DV) coordinates within the Drosophila embryo from which single neuroblasts later arise. Here, we show that the AP and DV position of these clusters is regulated through a common cis-acting region between the genes under the initial control of the products of the pair-rule and DV polarity genes and is later maintained by selected segment polarity genes. The combination of proneural gene activation/repression in AP stripes and repression within specific DV domains positions each cluster of achaete/scute expressing cells within segments; interactions between these cells then determine individual cell fates.
[Key Words: achaete; scute; proneural gene expression; axis-patterning genes]
Received July 27, 1992; revised version accepted October 2, 1992.
During embryonic development in Drosophila, cell fates are determined according to their position in the em- bryo. Although a great deal has been learned about the genetic regulatory system governing axis formation and global positional information in the embryo, it is less well understood how individual or groups of cells decode this information and follow distinct fates. A combina- tion of classical and molecular genetics has shown that the early zygotic patterning genes act progressively in a regulatory cascade to subdivide the embryo into its seg- mental units (for review, see Ingham 1988; Nfisslein- Volhard 1991). The gap genes interpret maternal antero- posterior (AP) gradients to specify the positional fate of large multisegment embryonic regions. The pair-rule and segment polarity genes refine this information and specify positional information to alternating segment- sized regions and to specific regions within each seg- ment, respectively. Although the analysis of the regula- tory interactions between the different classes of zygotic segmentation genes has yielded a formal understanding of how the basic segmental organization of the Droso- phila embryo is laid down, very little is understood about how these genes specifically contribute to the pat- tern of differentiated cell types or tissues within a segment.
aCorresponding author.
One of the first signs of intrasegmental pattem in the Drosophila embryo is the appearance of proneural gene expression of the achaete-scute complex loci (AS-C), achaete (ac), scute (sc), and lethal of scute (l'sc), in small clusters of cells during gastrulation (Cabrera et al. 1987; Martin-Bermudo et al. 1991; Skeath and Carroll 1992). The earliest phase of proneural gene expression is com- posed of a precise AP and dorsoventral (DV) pattern of cell clusters that is repeated within every segment. Ge- netic and cell ablation experiments suggest that the pro- neural clusters are groups of equipotential cells, all of which have a potential neural fate (Stem 1954; Taghert et al. 1984; Doe and Goodman 1985; Simpson 1990). This potential appears to be conferred to these cells by their expression of the proneural genes (Cabrera et al. 1987; Romani et al. 1989; Jimenez and Campos-Ortega 1990; Cubas et al. 1991; Martin-Bermudo et al. 1991; Skeath and Carroll 1991, 1992) and is eventually re- stricted to a single cell that becomes a neural precursor [a neuroblast (NB) or sensory mother cell]. The dynamics of proneural gene expression directly reflects this cell fate restriction, as proneural gene expression also becomes restricted from all cells of a cluster to just one, the future NB (Cabrera 1990; Cubas et al. 1991; Martin-Bermudo et al. 1991; Skeath and Carroll 1992). All of the first NBs to segregate into the embryonic central nervous system
2606 GENES & DEVELOPMENT 6:2606-2619 �9 1992 by Cold Spring Harbor Laboratory ISSN 0890-9369/92 $3.00
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Intrasegmental pattern formation
(CNS) arise from clusters of cells expressing one or more of the proneural genes of the AS-C (Martin-Bermudo et al. 1991; Skeath and Carroll 1992). Thus, the initial pat- tern of proneural clusters and proneural gene expression within the Drosophila embryo precisely foreshadows the NB pattern in the embryonic CNS. Therefore, the exam- ination of how proneural gene expression is regulated is a direct path to understanding how potential cell fates are assigned at reproducible positions within a segment.
We have concentrated our efforts on unraveling the genetic regulatory mechanisms that establish the initial expression patterns of the proneural genes. These pat- terns are relatively simple; for example, within the pre- sumptive CNS, the ac (Skeath and Carroll 1992) and sc (J.B. Skeath, G. Panganiban, and S.B. Carroll, unpubl.) genes are first expressed coincidently in a pattern of four distinct cell clusters located at reproducible positions within each hemisegment. Through the analysis of DNA rearrangements within the AS-C and l acZ reporter con- structs, we have identified common control regions lo- cated between the ac and sc genes that are required for the initial spatial regulation of both ac and sc. In addi- tion, we show that specific combinations of pair-rule genes act in trans to establish the periodicity of ac ex- pression along the AP axis, whereas the dorsal-ventral (DV) genes restrict ac gene expression to specific do- mains along the DV axis. Surprisingly, the segment po- larity genes play no role in the initiation of the ac ex- pression pattern; however, the products of two loci, gooseberry {gsb) and n a k e d (nkd}, act subsequent to the pair-rule genes to maintain the initial AP boundaries of ac expression.
Results
Registration of proneural gene expression
By early gastrulation, the ac, sc, and l 'sc proteins are expressed in stripes of cells within the neuroectodermal region that gives rise to the CNS (Cabrera 1990; Martin- Bermudo et al. 1991; Skeath and Carroll 1992). During early germ-band extension (stages 7-8, as defined by Campos-Ortega and Hartenstein 1985), these stripes of cells appear to rearrange and form 10 adjoining proneural cell clusters per hemisegment {Fig. la, b,c; Martin-Ber- mudo et al. 1991; Skeath and Carroll 1992). During stage 9, one cell from the central region of each cluster, the future NB, retains proneural protein expression and delaminates into the embryo, whereas the other cells of the cluster slowly lose proneural expression. Before the NBs divide, proneural protein expression in them is ex- tinguished (late stage 9/early stage 10). While ac and sc are not expressed in the neuroectoderm again until late stage 10, a second and third phase of l 'sc expression is superimposed on the initial patterns of ac, sc, and l'sc during stages 9 and 10 (Martin-Bermudo et al. 1991). By stage 11, all three proteins are expressed in the peripheral nervous system (PNS) and CNS in rapidly changing and spatially intricate patterns (Martin-Bermudo et al. 1991; J. Skeath, G. Panganiban, and S.B. Carroll, unpubl.). In
this report we focus entirely on the initial clustered pat- terns of ac, sc, and l 'sc expression (late stage 8/early stage 9) and on the genetic regulatory systems that estab- lish the initial spatial limits of proneural gene expression.
The registration of proneural gene expression in the neuroectoderm is most easily discussed using a Carte- sian coordinate system to refer to the positions of differ- ent proneural clusters within each hemisegment (sum- marized in Fig. l e). Along the AP axis, each hemiseg- ment is divided into four rows or quadrants--A, B, C, D---which successively divide each hemisegment, from the anterior to the posterior, into fourths. Along the DV axis, each hemisegment is divided into three columns: medial (m); intermediate (i); and lateral {1}. Each he- misegment is then composed of 12 squares, each one uniquely identified by its AP and DV coordinates. Each square represents roughly four to six cells and outlines the boundary of a different proneural cluster.
The relative positions of proneural clusters have been precisely determined in single- and double-label experi- ments using antibodies specific for the ac (Fig. la, c), l 'sc (Fig. lb, c), and engrailed (en)(data not shown) proteins, as well as RNA in situ experiments using a probe specific for the sc transcript. During late stage 8, ac (and sc; data not shown) is expressed in four clusters per hemiseg- ment--the m and 1 clusters of rows B and D (Fig. la, c,e; Skeath and Carroll 1992)--whereas l 'sc is expressed in a circular pattern of eight adjoining proneural cell clus- t e r s - the m, i, and 1 clusters of rows A and C, and the m and 1 clusters of row D (Fig. lb, c,e; Cabrera 1990; Martin- Bermudo et al. 1991). By mid-stage 9, one NB has delam- inated from each proneural cluster (summarized in Fig. le; Martin-Bermudo et al. 1991; Skeath and Carroll 1992). Thus, the clustered pattern of proneural gene ex- pression precisely prefigures the pattern of the first 10 NBs to segregate into the embryonic CNS (Fig. 1 e). Along the AP axis, three of the four rows express different com- binations of these genes and no two contiguous rows express the same combination: l 'sc is expressed in rows A and C; ac and sc are expressed in row B; and all three are expressed in row D (Fig. lc,e). At this stage, no pro- neural gene expression is found within the i clusters of rows B and D. During stages 9 and 10, a second and third phase of l'sc expression occurs and these regions, as well as many other regions of each hemisegment, accumulate l 'sc protein (Cabrera 1990; Martin-Bermudo 1991). Be- cause of the relative simplicity of the ac/sc expression pattern and the complications of interpretation that arise from the superimposition of the second and third phases of l 'sc expression on the initial patterns of ac, sc, and l 'sc expression, we focused on elucidating the genetic regu- latory mechanisms that act within the early embryo to establish the initial limits of ac gene expression.
Other AS-C gene products affect the level but not the pattern of ac expression
During the development of the adult nervous system, both ac and sc are again expressed coincidently (Romani et al. 1989; Cubas et al. 1991; Skeath and Carroll 1991).
GENES & DEVELOPMENT 2607
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Skeath et al.
Figure 1. The coincident expression pat- terns of ac and sc overlap partially with that of l'sc. Ventrolateral views of late stage 8/early stage 9 wild-type embryos la- beled for the ac protein (a), l 'sc protein (b), and doubly labeled for the ac (brown) and l 'sc (black)proteins (c). High magnifica- tion ventrolateral view of the abdominal segments of a Df(I)sc 4LgR embryo labeled for the ac protein (d). (e) Schematic repre- sentation of the registration of ac, sc, and l 'sc expression in two consecutive wild- type hemisegments at stage 8 (left) when the proneural genes are expressed in cell clusters and at stage 9 (right) after proneu- ral gene expression has become restricted from clusters of cells to single cells, the future NBs. Each cluster of six cells repre- sents a separate proneural cluster from which a single neuroblast arises. Cells ex- press l 'sc (black); ac and sc (brown); and ac, sc, and l 'sc (purple). Note that proneu- ral gene expression initially divides each hemisegment into a grid consisting of four latitudinal rows [A, B, C, and D (rows B and D are marked in a and b; all rows are marked in c and e)] and three longitudinal columns, medial (m), intermediate (i), and lateral {1) (a,b,c,e). ac protein and sc tran- scripts (not shown) are detected in cell clusters within both the m and 1 columns of rows B and D (a,c,e), whereas the l 'sc protein is detected in cell clusters within the m and 1 columns of row D and within the m, i, and 1 columns of rows A and C (b,c,e). The expression patterns of ac/sc and l'sc coincide in the m and 1 columns of row D (brownish-grey cells in c). In a Df(1)sc aLgR mutant embryo, the removal of l'sc gene function has no effect on ac expression within row B (d, arrows) but causes a reduction in the level of ac expression in row D (d, arrowheads). To compare the expression of ac in a wild-type embyro with that in a Df(1)sc 4LgR mutant embryo compare ac expression (brown and grey cells) in c to ac expression in d. (a,b) Bar, 50 ~m; (c,d) bar, 30 lain.
It has been shown that during adult development the identical nature of these two patterns arises from cross- regulation between the ac and sc genes (Martinez and Modolell 1991; Skeath and Carroll 1991). To determine whether cross-regulation between the products of the proneural genes of the AS-C accounts for the generation of any aspect of the initial embryonic pattern of ac ex- pression, we examined ac protein distribution in em- bryos hemizygous mutan t for l ' s c or sc. As simple point muta t ions have not been generated in ac, sc, or l 'sc, we were forced to use synthetic deficiencies that removed l ' s c or sc as well as - 2 0 kb of surrounding D N A (see Fig. 3a, below; Dambly-Chaudiere et al. 1988; Lindsley and Z i m m 1992). The effects of these deletions on the ac
expression pattern could arise either from the loss of l ' s c
or sc function or from the removal of ac regulatory se- quences. Our conclusions, the evidence for which is de- tailed below, is that l ' s c affects the level but not the spatial pat tern of ac and that disruptions in the ac or sc
patterns in certain synthetic deficiencies in the a c / s c
region result from disruptions of a common c is -regula-
tory region between the genes.
In Df (1 ) sc 4LgR ( l ' s c - ; this deletion removes D N A be- tween +24.9 and +5.1; see Fig. 3a, below; Lindsley and Z i m m 1992) embryos, the pattern of ac expression is not altered (Fig. ld). However, in row D where ac and l ' s c are normally coexpressed, the level of ac expression is fre- quently reduced (Fig. ld, arrowheads), and very rarely ac
expression is absent from the m cluster of row D (not shownl. The NB that arises from this cluster is the NB that is most frequently absent in Df(1)SC 4LgR embryos (Martin-Bermudo et al. 1991). In addition, the t ime at which ac protein is detected within row D is delayed from late stage 7 in wild-type embryos until late stage 8/early stage 9 in Df(1 ) sc 4L9R embryos. Thus, l ' s c ap- pears to enhance the level of ac expression but does not control the spatial pattern of ac expression.
A s h a r e d r e g u l a t o r y reg ion b e t w e e n t h e ac a n d sc g e n e s
Critical insights into the c i s - a c t i n g elements required for the spatial regulation of ac and sc come from the analysis of gene expression in embryos containing inversion breakpoints or deletions between ac and sc. These mu-
2608 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Intrasegmental pattern formation
tants reveal that common intergenic control e lements regulate a c and s c expression. For example, in embryos that lack the s c region of the AS-C [ D f ( 1 ) s c 8L4R (Lindsley and Z i m m 1992)], a striking effect on the a c pattern is observed where a c is expressed only in row B (Fig. 2a, arrows) but almost completely absent from row D (Fig. 2a, arrowheads). The s c 8LaR deletion extends proximally for 22 kb from a point - 1 0 kb downstream of the ac
transcription uni t (Fig. 3a; coordinates + 47.4 to + 24.9; Lindsley and Z i m m 1992). Curiously, there is a recipro- cal effect on s c expression in D f ( 1 ) y 3 P L s c 8R ( a c - ) em- bryos: s c is expressed in row D (Fig. 2b, arrowheads) but not in row B (Fig. 2b, arrows). The y3PLscSR deletion re- moves the a c transcription unit and - 1 7 kb of 5' and 10 kb of 3' DNA [Fig. 3a, coordinates + 73.0 to + 47.4 (Lind- sley and Z i m m 1992)]. Two different interpretations can explain the reciprocal pattern alterations to ac and sc in these mutan t embryos: (1) ac expression could be re- quired to activate s c in row B and s c expression could be required to activate a c in row D; or (2) the D f ( 1 ) s c 8L4R
and D f ( 1 ) y 3 P L s c 8R deletions could remove control ele- ments required to activate ac in row D or s c in row B, respectively. Experiments described below establish that these deletions are removing control elements.
To dist inguish between the two possibilities, we ex- amined both a c and s c expression in I n v e r s i o n ( I ) s c 8 em- bryos (Fig. 2c, d). The distal breakpoint of I n ( 1 ) s c 8 lies
between the a c and s c genes at + 47.4, - 1 2 kb upstream of sc and 10 kb downstream of a c (Fig. 3a; Lindsley and Z i m m 1992), coincident wi th the distal breakpoint of D f ( 1 ) s c 8L4R and the proximal breakpoint of the D f ( 1 ) y 3 P L s c sR deletions (Fig. 3a). I n ( 1 ) s c 8, then, precisely separates the c i s - a c t i n g sequences deleted in Df(1)SC 8L4R away from the a c transcription uni t and the c i s - a c t i n g
sequences deleted in D f ( 1 ) y 3 P L s c 8R away from the s c
transcription unit but preserves the coding regions of both genes. We reasoned that if ac activated s c in row B a n d s c activated ac in row D, then should a c be expressed in row B in I n ( 1 ) s c s embryos, s c expression would also be found there, and should sc be expressed in row D in these same embryos, a c would also be expressed there. On the other hand, if the control e lements required to activate ac in row D are located, for the most part, proximal to the s c s breakpoint (the region deleted in D f ( 1 ) s c 8LaR em- bryos) and the control e lements necessary to activate s c
in row B are located distal to the s c 8 breakpoint (the region deleted in D f ( I ) y 3 P L s c 8R embryos), then in I n ( 1 ) s c 8 embryos, one would expect a c not to be ex- pressed in row D and sc not to be expressed in row B. The results are consistent with the second model: a c is ex- pressed only in row B (Fig. 2c, arrows}, and s c is expressed only in row D of I n ( 1 ) s c 8 embryos (Fig. 2d, arrowheads). This suggests that the init ial embryonic patterns of a c
and sc are not determined by cross-regulation but that
Figure 2. Dual control of ac and sc by a common intergenic regulatory region. (a) Ventrolateral view of an early stage 9 Df(1)sc 8L4R {sc- ) embryo labeled for the ac (arrows) and en (arrowheads) proteins; en
expression marks row D. {b) Ventrolateral view of a stage 8 D f ( 1 ) ~ P L s c 8R (ac - ) em- bryo probed for both sc (arrowheads) and wg RNA (wg expression marks row C). (c) High magnification ventrolateral view of an early stage 9 In(1)sc 8 embryo labeled for ac (arrows) and en (arrowheads). (d) Lateral view of a late stage 8 In(1)sc 8 embryo probed for sc RNA. (e) Lateral view of a stage 8 In(1)sc v2 embryo labeled for ac pro- tein. (f) lateral view of a stage 8 and (h) ventral view of a stage 9 In(1)sc ve embryo both probed for sc RNA. (g) Ventral view of an early stage 9 In (1)ac 3 embryo labeled for the ac protein. In a -h , arrows mark the position of row B; arrowheads mark the position of row D. Note that the expres- sion pattem of ac in Df(1)sc 8LaR and In(1)sc 8 embryos is identical: ac is ex- pressed in row B (a,r arrows) but not in row D (a,c, arrowheads). The reciprocal pattern alteration occurs to sc in both Df(1)y3PLsc 8R and In(1)sc 8 embryos: sc is expressed in row D (b,d, arrowheads) but not in row B (b,d, arrows). In comparison to In(1)sc 8 embryos, ac expression in In(1)sc v2 embryos is reduced in row B (e, arrows), whereas sr expression is en-
hanced in row B (f,h, arrows). Finally, although the level of ac expression is greatly diminished in In (1)ac 3 embryos (g), its pattern is essentially normal. (a ,b ,d- f ,h ) Bar, 50 ~tm; (c,g) bar, 30 ~tm.
GENES & DEVELOPMENT 2609
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from
Skeath et al.
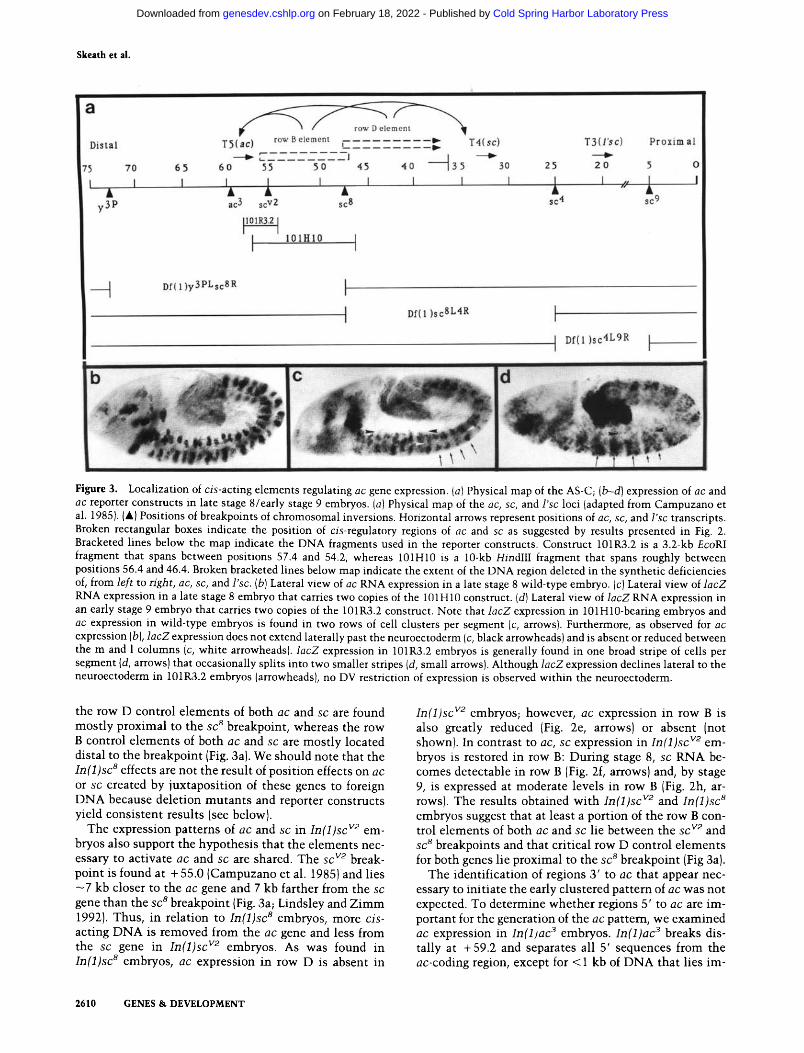
Figure 3. Localization of c is -ac t ing elements regulating ac gene expression. (a) Physical map of the AS-C; (b--d) expression of ac and ac reporter constructs in late stage 8/early stage 9 embryos. (a) Physical map of the ac, sc, and l 'sc loci (adapted from Campuzano et al. 1985). (A) Positions of breakpoints of chromosomal inversions. Horizontal arrows represent positions of ac, sc, and l ' sc transcripts. Broken rectangular boxes indicate the position of cis-regulatory regions of ac and sc as suggested by results presented in Fig. 2. Bracketed lines below the map indicate the DNA fragments used in the reporter constructs. Construct 101R3.2 is a 3.2-kb EcoRI
fragment that spans between positions 57.4 and 54.2, whereas 101H10 is a 10-kb HindII I fragment that spans roughly between positions 56.4 and 46.4. Broken bracketed lines below map indicate the extent of the DNA region deleted in the synthetic deficiencies of, from l e f t to right , ac, sc, and l 'sc. (b) Lateral view of ac RNA expression in a late stage 8 wild-type embryo. (c) Lateral view of l a c Z
RNA expression in a late stage 8 embryo that carries two copies of the 101HIO construct. Id) Lateral view of l a c Z RNA expression in an early stage 9 embryo that carries two copies of the 101R3.2 construct. Note that l a c Z expression in 101H10-bearing embryos and ac expression in wild-type embryos is found in two rows of cell clusters per segment (c, arrows). Furthermore, as observed for ac expression (b), l a c Z expression does not extend laterally past the neuroectoderm (c, black arrowheads) and is absent or reduced between the m and 1 columns (c, white arrowheads), l a c Z expression in 101R3.2 embryos is generally found in one broad stripe of cells per segment (d, arrows) that occasionally splits into two smaller stripes (d, small arrows). Although l a c Z expression declines lateral to the neuroectoderm in 101R3.2 embryos (arrowheads), no DV restriction of expression is observed within the neuroectoderm.
the row D control elements of both ac and sc are found most ly proximal to the s c 8 breakpoint, whereas the row B control elements of both a c a n d s c are mostly located distal to the breakpoint (Fig. 3a). We should note that the I n ( 1 ) s c 8 effects are not the result of position effects on a c
or s c created by juxtaposition of these genes to foreign D N A because deletion mutan t s and reporter constructs yield consistent results (see below).
The expression patterns of a c a n d s c in I n ( 1 ) s c v2 em- bryos also support the hypothesis that the elements nec- essary to activate a c and s c are shared. The sc v2 break- point is found at + 55.0 (Campuzano et al. 1985) and lies - 7 kb closer to the a c gene and 7 kb farther from the sc
gene than the s c 8 breakpoint (Fig. 3a; Lindsley and Z i m m 1992). Thus, in relation to I n ( 1 ) s c 8 embryos, more c is -
acting D N A is removed from the a c gene and less from the s c gene in I n ( 1 ) s c v2 embryos. As was found in I n ( 1 ) s c 8 embryos, ac expression in row D is absent in
I n ( 1 ) s c v2 embryos; however, a c expression in row B is also greatly reduced (Fig. 2e, arrows) or absent (not shown). In contrast to ac, s c expression in I n ( 1 ) s c v2 em- bryos is restored in row B: During stage 8, s c RNA be- comes detectable in row B (Fig. 2f, arrows) and, by stage 9, is expressed at moderate levels in row B (Fig. 2h, ar- rows). The results obtained with I n ( 1 ) s c v2 a n d I n ( 1 ) s c s
embryos suggest that at least a portion of the row B con- trol elements of both ac and s c lie between the s c v2 and s c 8 breakpoints and that critical row D control elements for both genes lie proximal to the sc 8 breakpoint (Fig 3a).
The identification of regions 3' to a c that appear nec- essary to initiate the early clustered pa t tem of a c was not expected. To determine whether regions 5' to a c are im- portant for the generation of the a c pattern, we examined a c expression in I n ( 1 ) a c 3 embryos. I n ( 1 ) a c 3 breaks dis- tally at + 59.2 and separates all 5' sequences from the ac-coding region, except for < 1 kb of D N A that lies im-
2610 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Intrasegmental pattern formation
mediately 5' to ac (Fig. 3a; Campuzano et al. 1985). The level of ac expression is reduced markedly in these em- bryos, but the initial clustered pattern of ac expression is normal (Fig. 2g}. This result, in conjunction with the observed patterns of ac and sc in In(1)sc 8 and In(1)sc v2 embryos, argues that the regulatory regions necessary to initiate the early spatial pattern of ac expression are lo- cated predominantly 3' to ac between the ac and sc genes.
Further support for the role of the intergenic region in the spatial control of ac and sc expression comes from reporter gene constructs. Different, but partially overlap- ping, fragments from the intergenic region are sufficient to drive weak l acZ expression within the neuroectoderm in periodic patterns that resemble the endogenous pat- tern of ac expression (Fig. 3, cf. c and d with b). As is observed for ac (Fig. 3b) and sc (not shown), l acZ expres- sion is detected in two rows of cell clusters per segment (Fig. 3c, arrows) when a 10-kb fragment of intergenic DNA between coordinates 56.5 and 46.5 (Fig. 3a, 101H10) is inserted upstream of the l acZ gene in a re- porter construct. In 101H10, the DV registration of gene expression also closely parallels the endogenous pattern of ac/sc expression, as no l a c Z expression is observed dorsal to the neuroectoderm (Fig. 3c, black arrowheads); and gene expression is repressed, albeit incompletely, be- tween the m clusters (not shown) and the m and 1 clus- ters (Fig. 3c, white arrowheads). A 3.2-kb fragment (101R3.2) that spans from coordinate 57.4 (just 3' to the ac transcription unit) to 54.2 and partially overlaps with 101H10 (Fig. 3a) also drives l acZ expression in a pattern that highlights some aspects of the ac/sc pattern (Fig. 3d). Along the AP axis, expression is observed in one broad stripe of cells that occupies roughly three-fourths of each segment (Fig. 3d, long arrows). Occasionally, the cells found in the middle of these stripes do not express lacZ or express it at very low levels yielding two stripes per segment (Fig 3d, short arrows). Although reporter gene expression is lower in more 1 regions (Fig 3d, black arrowheads), it is more derepressed along the DV axis of the neuroectoderm than in thel01H10 construct (Fig. 3, cf. d with c). Thus, whereas the 101H10 construct yields a fairly faithful reproduction of the endogenous spatial pattern of ac/sc expression, the 101R3.2 construct gen- erates a pattern that is only partially correct in the AP and DV aspects. Clearly, these results strongly support the hypothesis that a complex cis-acting regulatory re- gion exists between ac and sc and that it is required to initiate expression of both genes in their proper spatial patterns. The fact that even large constructs drive only weak l acZ expression in the above-mentioned patterns, however, suggests that the proper level of gene expres- sion may require the additional presence of 5' regulatory elements (see Discussion).
The ini t ial A P posi t ions of proneural clusters are set up by pair -ru le genes and are independen t of s egmen t polar i ty genes
Having established that the initial limits of ac expres- sion are set up largely independently of the other mem-
bers of the AS-C and having identified cis-acting regions required to initiate the clustered expression patterns of ac and sc, we then determined which trans-acting factors might act directly on these regulatory elements to gen- erate the precise expression patterns of ac and sc. The AP and DV registration of these clusters and the time at which ac expression becomes detectable (early gastrula- tion) suggest that the segment polarity, the pair-rule, and the zygotic DV genes are the prime candidate genes to regulate ac expression directly. It has been proposed that the intersection of the expression patterns of these genes forms a grid within which the proneural genes are activated at specific coordinates (Ghysen and Dambly- Chaudiere I989; Jan and Jan 1990). With this model in mind, we sought to identify the genes that control ac expression along each axis.
The similarity between ac and segment polarity gene expression is striking (Fig. 4a). Although ac is expressed in two transverse rows of cell clusters per segment and many segment polarity genes are expressed in one stripe of cells around the embryo per segment, both are ex- pressed in similar and sometimes identical domains along the AP axis. For example, the A and P borders of ac expression within row D coincide perfectly with the A and P borders of en expression, whereas ac expression within row B is found midway between stripes of en- expressing cells (Fig. 4a; Skeath and Carroll 1992). ac and segment polarity gene expression, then, appear to respect similar boundaries. Because of this correspondence, it seemed likely that the establishment of the initial AP limits of ac expression would involve the segment polarity genes. We were surprised to find that this was not the case. In embryos singly mutant for all of the zygotic segment polarity genes [en, armadil lo , wing- less (wgl, hedgehog, patched, gsb, nkd , and Cubi tus interruptus D~ and in embryos doubly mutant for en and invected, the initial embryonic pattern of ac was essentially identical to that observed in wild-type em- bryos (data not shown). This suggested that the genes that establish the boundaries of segment polarity gene expression, the pair-rule genes, also regulate directly the initial AP boundaries of ac gene expression within seg- ments.
In embryos homozygous for null mutations for each pair-rule gene examined [fushi tarazu (ftz), odd-sk ipped (odd), paired (prd), even-sk ipped (eve), odd-paired (opa), sloppy-paired (slp), runt (data not shown), and hairy (data not shown)], the early clustered pattern of ac expression is altered (Figs. 4 and 5). The effect of removing any one of these genes on ac expression, with the exception of runt, is to cause specific pattem alterations at a double segment periodicity. On the basis of their effect on ac expression, most pair-rule genes can be grouped broadly into two categories: those required to set up ac expres- sion within row D (Fig. 4; ftz, odd, prd, eve); and those necessary to establish ac expression within row B (Fig. 5; opa, slp, prd). It should be noted that in all pair-rule mutant embryos where it has been examined (ftz, prd, and odd), sc expression is altered in a manner identical to ac (not shown). Therefore, the sc expression pattern ap-
GENES & DEVELOPMENT 2611
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Skeath et al.
Figure 4. Pair-rule genes positioning ac gene expression within row D. Ventral or lateral views of late stage 8 wild-type (a) and pair-rule mutant (b-h) embryos either singly labeled for the ac protein (b,c,e,g) or doubly labeled for the ac (brown) and en proteins (a,d,f,h, black/purple). The do- main of en-expressing cells marks row D of each segment. (a) Lateral view of a wild- type embryo. At this stage, every segment contains two transverse rows of ac clus- ters: one found midway between stripes of en-expressing cells; the other coincident with the en stripes. In embryos mutant for particular pair-rule mutants, this reiter- ated pattern is disturbed (b--h). In eve 12z embryos (b), all ac clusters normally found in row D are deleted. In ftz embryos (c,d), every fourth transverse row of ac clusters disappears, corresponding to row D of ev- ery even-numbered segment (c,d, arrows). In odd embryos (e,f), the same row of ac clusters expands posteriorly and is three times its normal width (e,f, arrows). Every fourth transverse row of ac clusters is lost in prd embryos (g,h), and these clusters correspond to those found in row D of odd- numbered segments (g,h, arrows). In addi- tion, the row B clusters found in the ante- rior half of each even-numbered segment expands posteriorly and fuses with the ac clusters in row D (g,h, arrowheads). (a,b, c,e,g) Bar, 50 ~m; {d,f,h)bar, 30 ~m.
pears to be regulated in trans in the same way as ac,
which is also consistent wi th ac and sc being regulated by common cis-act ing elements.
T h e c o m b i n a t o r i a l ac t ion of four p a i r - r u l e genes e s tab l i shes ac express ion w i t h i n r o w D
In embryos mu tan t for f tz , every fourth row of ac clus- ters is removed (Fig. 4c, d, arrows). The deleted clusters are those normally found in row D in every even-num- bered segment. The same row of clusters expands poste- riorly to roughly three t imes its normal width in em- bryos mu tan t for o d d (Fig. 4e,f, arrows). Within this re- gion in wild-type embryos, the anterior edges of ac and f t z gene expression coincide precisely, whereas the pos- terior border of ac and the anterior border of o d d expres- sion abut [the registration of ac expression to pair-rule gene expression was deduced from the known expression patterns of ac and the indicated pair-rule genes (Carroll et al. 1988; Coulter et al. 1990)]. Thus, the borders of ac expression defining row D of the even-numbered seg- ments may result from the combinatorial effect of f t z activation and o d d repression.
A similar type of combinatorial mechanism involving different pair-rule genes can explain the borders of ac
expression within row D of the odd-numbered segments. In prd mutan t embryos, every fourth transverse row of ac clusters is deleted; these are the normal row D clusters of odd-numbered segments {Fig. 4g, h, arrows). This same row of ac clusters is removed in embryos mu tan t for eve (Fig. 4b). Because, in wild-type embryos, the anterior bor- ders of ac and eve expression and the posterior borders of ac and prd expression coincide in row D of odd-num- bered segments {Frasch and Levine 1987; Carroll et al. 1988; Baumgartner and Noll 1990}, activation of ac ex- pression within row D of odd-numbered segments may result from the overlapping domains of prd and eve gene expression. We note that prd and eve have additional effects beyond the odd-numbered D rows: p r d - embryos also exhibit fusion of the B and D rows of even-numbered segments (see below}, whereas in e v e - embryos, the D rows of even-numbered segments are deleted, probably because of an indirect effect of eve on [tz expression (Car- roll and Scott 1986; Frasch and Levine 1987).
In contrast to row D, we cannot propose a simple model to explain the establ ishment of ac expression in row B. In every segment, the loss of opa function re- moves ac expression from row B (Fig. 5a, b, arrows) and decreases the cluster size and level of ac expression in row D. In embryos singly mutan t for either sip (Fig. 5c) or
2612 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Intrasegmental pattern formation
Figure 5. Pair-rule genes positioning ac expression within row B. Ven- tral view of a stage 9 opa embryo (a}; lateral view of a late stage 8/early stage 9 slp embryo (c) both labeled for ac protein; and high magnification ventral view of a stage 9 opa embryo labeled for ac (brown) and en (blue/ black) expression (b). In embryos mutant for opa, every transverse row of ac clusters within row B is deleted (a,b, arrows). In slp embryos, the row of ac clusters found in row B of odd-numbered segments fuses with the ac clusters of the same (c, arrows).
prd (Fig. 4g, h), the B and D rows of ac expression fuse in the odd- and even-numbered segments, respectively. When the ac pattern is established, opa is expressed uni- formly throughout the segmental primordia (M. Benedyk and S. DiNardo, pets. comm.), whereas the anterior edge of prd expression abuts the posterior edge of ac expres- sion in row B in even-numbered segments (Baumgartner et al. 1991) and the posterior edge of slp expression abuts the anterior edge of ac expression in row D in odd-num- bered segments (Grossniklaus et al. 1992). The expres- sion domains of ac and prd suggest that prd represses ac expression at the posterior border of row B in even-num- bered segments, whereas the relative expression patterns of slp and ac suggest that slp could repress ac at the anterior border of row D in odd-numbered segments. It is also possible that slp acts alone, or in conjunction with other genes, to repress ac expression at the posterior bor- der of row B in odd-numbered parasegments. The recent cloning and characterization of the slp locus may help to unravel the role slp plays in establishing the ac expres- sion pattern (Grossniklaus et al. 1992). No simple model can be made for the effect of opa on ac. It is possible that opa acts in row B as a general activator and in row D as an enhancer of ac expression, whereas other as yet un- identified genes would define the anterior borders of ac expression in row B. Regardless of the exact mechanism that establishes ac expression within row B, these results support the idea that the combinatorial action of distinct sets of pair-rule genes defines the periodicity of ac ex- pression in the B and D rows of odd- and even-numbered segments.
gsb a n d nkd s e g m e n t po lar i t y genes m a i n t a i n the A P borders of ac e c t o d e r m a l cell c lus ters
Although the initial pattern of ac expression (stage 8) in embryos singly mutant for all of the zygotic segment
polarity genes is essentially identical to that observed in wild-type embryos, adjacent rows of ac clusters fuse in older embryos (mid-late stage 9) mutant for either gsb or n k d (Fig. 6). In embryos mutant for gsb, ac expression in row B expands posteriorly into row C and eventually merges with ac expression in row D (Fig. 6c, e; arrows mark row C), whereas expression in row D appears to dissipate prematurely (Fig. 6, cf. a with c and el. The directional expansion of ac expression in gsb mutant em- bryos is demonstrated by double-labeling with ac and en antibodies [Fig. 6e; en expression (purple) marks row D). In wild-type embryos, the anterior limit of gsb expres- sion abuts the posterior edge of ac expression in row B (J.B. Skeath and S.B. Carroll, data not shown; Baumgart- ner et al. 1987; A. Ungar and R. Holmgren, in prep.), gsb activity appears necessary to maintain the posterior bor- der of ac expression in row B by maintaining ac gene repression in row C. In embryos mutant for n k d , ac ex- pression in rows B and D appears to expand: that of row B anteriorly into row A and that of row D posteriorly into row A (Fig. 6d, f; arrows mark row A). The oriented ex- pansion of ac expression into row A is most obvious in n k d mutant embryos double-labeled for ac and en (Fig. 6f}. Thus, along the AP axis, the concerted action of gsb and n k d appears to preserve the initial pair-rule-defined borders of ac expression by maintaining the repression of ac in rows C and A, respectively (Fig. 6b). Interestingly, the removal of either of these genes has apparently no effect on l ' sc expression (Martin-Bermudo et al. 1991).
It should be noted that although only n k d and gsb control the initial pattern of ac gene expresssion, all zy- gotic segment polarity genes, with the possible excep- tion of gsb, govern the spatial patterning of the later phases of ac gene expression in the PNS and CNS (J.B. Skeath and S.B. Carroll, unpubl.). This raises the inter- esting possibility that two different sets of genes estab- lish the AP pattern of ac gene expression at different times during embryogenesis.
GENES & DEVELOPMENT 2613
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Skeath et al.
Figure 6. The gsb and nkd segment polarity genes maintain the AP registration of ac expression. Ventral views of stage 9 wild-type (a), gsb (c), and nkd (d) mutant embryos labeled for the ac protein; high magnification ventral views of stage 9 gsb [e) and nkd (f) mutant embryos labeled for the ac (brown) and en (black/purple) proteins; en expression marks row D. In embryos mutant for either gsb or nkd, adjacent rows of ac clusters fuse (c-f). In embryos mutant for gsb, ac expression in row B expands posteriorly into row C {c,e; arrows mark row C). In nkd embryos, ac expression within row B expands anteriorly into row A and that of row D expands posteriorly into row A (d,f; arrows mark row A). (b} Schematic diagram of the roles gsb and nkd play in maintaining the integrity of the imtial registration of ac ectodermal cell clusters. Cells express ac (brown); cells express ac and en (purple). (a,b,d) Bar, 50 ~m; (e,f) bar, 30 ~m.
The zygot ic D V genes repress ac expression wi th in specific D V doma ins
In a survey of representative zygotic (but by no means all) DV pattern mutants , we found that the ac domain frequently expanded, indicating that DV regulation of ac expression appears to occur largely through repression of ac expression wi th in specific domains along the DV axis. These domains correspond to the region between the m clusters [regulated by tw i s t (twi) and snail (sna)], the re- gion dorsal to the 1 clusters [regulated by decapentaple- gic (dpp) and tol loid (tld)], and the region between the m and 1 clusters (regulators unknown). In embryos mutan t for tw/, the expression of which is largely restricted to the region between the two columns of m clusters (Thisse et al. 1988; Kosman et al. 1991), a partial fusion of these clusters occurs (Fig. 7b, arrowheads). The dorso- lateral extent of ac expression appears to be regulated by dpp and tld; the ventral l imi t of expression of each of these two genes extends roughly to the dorsal extent of ac expression wi th in the 1 clusters (St. Johnston and Gelbart 1987; Shimell et al. 1991}. Embryos mutan t for each gene exhibit essentially identical alterations to the
ac patterns where the 1 clusters expand dorsally (Fig. 7, cf. c and e with the wild-type embryo in a). The severity of the phenotype increases wi th age, as in stage 9 dpp (Fig. 7e) or t ld (not shown) mutan t embryos; the 1 extent of these clusters has expanded further dorsally than in stage 8 mutant embryos. In embryos mutan t for zerknf i l l t (zen), the 1 clusters may expand dorsally to a small degree (Fig. 7d); however, this effect might also be attributable to the inabil i ty of cells to rearrange and con- dense during germ-band extension. Curiously, we found no mutants in which the m and 1 clusters fuse. These results argue that these DV genes directly or indirectly repress ac expression wi th in specific domains of the DV axis.
D i s c u s s i o n
The role of global pat terning genes in regulating m trasegm en tal pat terns
The analysis of the regulation of the early zygotic genes involved in embryonic pattern formation has yielded a
2614 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Intrasegmental pattern formation
a
C
O
b
C Figure 7. DV genes restrict a c expression to specific regions of the DV axis. Ven- trolateral views of a stage 8 wild-type (a), a stage 8 tld (c), a stage 8 zen (d), and an early stage 9 dpp (e) mutant embryo and ventral view of a late stage 8 tw/mutant embryo, which has been splayed open to visualize better the ventral region of the embryo (b). Mutations in tw/{b) and sna (not shown) result in the partial fusions of the m clus-
ters, whereas mutations in tld (c) or dpp (e) cause the expansion of the lateral ac clusters dorsally. In a stage 9 dpp (e) or tld {not shown) embryo, the clusters have expanded farther dorsally than observed in stage 8 dpp (not shown) or tld (c) embryos. Mutations in zen (d) have a very slight effect on ac expression laterally, which may arise from an inability of cells to rearrange and condense during gastrulation. In a-e, the m and 1 clusters are labeled; arrowheads in b mark the ventral midline. (a-e) Bar, 50 ~m.
formal genetic understanding of how the basic segmental organization of the Drosophila embryo is created (for re- view, see Ingham 1988; Niisslein-Volhard 1991; Ingham and Martinez-Arias 1992); however, little is understood about the mechanisms that generate the precise patterns of discrete cell types within segments. For example, seg- ment polarity genes, the expression of which are the most restricted of the early pattern-forming genes, are expressed in one stripe per segment that consists by stage 8 of -100 cells. Although segment polarity genes specify positional cell identity (i.e., anterior or posterior position) within segments, they do not assign cells dis- crete fates: Cells found within the expression domain of any one such gene differentiate into a number of discrete cell types (e.g., NB, myoblast), which may be part of much larger tissue systems or structures (e.g., the ner- vous system, musculature, imaginal discs). Even though it has been widely assumed for some time that zygotic segmentation and DV genes play a fundamental role in the generation of complex spatial patterns of distinct cell types and structures, the mechanisms through which this is accomplished are only just now being deciphered.
Our results here on the establishment of proneural clusters and recent studies on the specification of the limb primordia (Cohen et al., 1992) and of the salivary glands (Panzer et al. 1992) in Drosophila suggest that spatial arrays of distinct cell types or structures are ini- tially specified by regulatory genes acting along both em- bryonic axes. It appears that the overlap of the expression patterns of particular early zygotic patterning genes de- termine the initial patterns of cell- or tissue-specific reg- ulatory genes within groups of cells located at specific and reproducible coordinates within the embryo. For ex- ample, the spatially overlapping combinations of pair-
rule and DV genes appear to carve out domains of ac expression as proneural clusters. Within the embryonic thoracic segments, the points of intersection between the expression domains of a segment polarity gene, wg, and a DV gene, dpp, appear to activate Brista/Distal-Iess in small groups of cells, as they are set aside as the pre- cursors of the larval leg imaginal discs (Cohen et al., 1992 ). The position of the salivary gland precursors is specified through activation by the homeotic gene Sex combs reduced along the AP axis and by repression by dpp and dorsal along the DV axis (Panzer et al. 1992).
The levels of proneural gene regulation reflect a stepwise refinement of cell fate
The genetic regulation of proneural gene expression, spe- cifically that of ac, is a model for the progressive refine- ment of positional identity that leads to organized arrays of specific cell types (NBs). The establishment of the segmentally repetitive pattem of ac-expressing proneu- ral clusters is the first step in this process. We have shown that along the AP axis the combinatorial action of specific pair-rule genes generates the periodic pattern of ac expression (Fig. 8). Within the D register of even-num- bered segments, the f tz gene, through activation, and the odd gene, through repression, appear to define the pre- cise anterior and posterior borders of ac expression, re- spectively. Within the D register of odd-numbered seg- ments the coexpression of prd and eve proteins appar- ently activates ac expression. It is important to note that within row D, the prd, eve, ftz, and odd genes control en gene expression (which is limited to the D register of every segment; DiNardo and O'Farrell 1987) in the same way that they control ac expression. Furthermore,
GENES & DEVELOPMENT 2615
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Skeath et al.
achaete D elements scute
O�9 O�9 OC: O�9 O I OQ
odd
A B
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
O 0 O 0 O 0 O 0 O 0 O 0
Figure 8. Two-dimensional control of gene expression. A sche- matic indicating the expression domains of ftz and odd pair- rule genes and a theoretical DV regulator within a ventrolateral region of an even-numbered parasegment (bracketed boxes in- dicate the domain of each protein). The specific position of pro- neural gene expression in a cluster of four (to six) cells within the D register (see text) results from a combination of activation by ftz (in the D, A, and B registers), and repression by odd lin the A, B, and C register) and the DV regulators (in the dorsolateral region). The action of each protein appears to be mediated by at least one cis-acting region between the ac and sc loci that in- fluences the expression of both genes.
whereas the pair-rule genes also appear to be the prime determinants of the AP periodicity of l 'sc (Martin-Ber- mudo et al. 1991), l 'sc is expressed largely out of phase (in the A and C register) with respect to ac/sc (Fig. 1). Clearly, a more detailed knowledge of the cis-acting el- ements involved in en, ac, and l 'sc regulation will be necessary to sort out how pair-rule proteins position the expression of each of these target genes.
The fact that the pair-rule, and not the segment po- larity genes, define the precise AP borders of proneural gene expression within segments was surprising on two accounts. First, it has been debated whether or not the graded distribution of pair-rule proteins is sufficient to draw precise boundaries or, rather, only to direct a crude subdivision of the segment that is later refined by seg- ment polarity gene expression (Ingham and Martinez- Arias 1992). Our results demonstrate that most pair-rule genes can contribute directly to intrasegmental patterns and raise the possibility that pair-rule genes are involved in regulating other sets of genes in addition to segment polarity, proneural, and other pair-rule genes.
Second, this observation is also challenging in light of the recent suggestion that pair-rule genes may not func- tion at all in the subdivision of the ectoderm of other insects. Patel et al. (1992)have shown that in the grass- hopper, the eve pair-rule gene is expressed in a highly
conserved {with respect to Drosophila) pattern in the CNS but not at all during segmentation. It is possible that the crucial role that pair-rule genes play in the first wave of NB segregation in Drosophila is not widely con- served in insects and that the later regulators of the pro- neural genes in Drosophila, the segment polarity, and homeotic genes (J.B. Skeath and S.B. Carroll, unpubl.) play a central role in CNS patterning in other insects. Thus, the control of the later phases of proneural gene expression in Drosophila may reflect a second level of AP control that is more widely conserved in insects.
Superimposed on the AP periodicity of ac expression is what we believe to be a pattern of repression imposed by domain-specific DV genes acting directly or indirectly (Fig. 8). tw/and sna are required to repress ac expression between the m clusters, whereas dpp and t ld are required to maintain the dorsal border of the 1 ac clusters. No DV genes have been identified that repress ac expression be- tween the m and 1 clusters, although we expect that a repressing activity exists between these clusters. The su- perimposition of the activities of the pair-rule and DV genes on the ac gene appears then to carve out clusters of ac expression at reproducible coordinates within each segment. It is possible that the regulatory elements (en- hancers) involved in mediating AS-C gene regulation are composed of target sites for both AP and DV regulators. For instance, we can imagine that a f tz- and odd-respon- sive element also contains sites for DV control such that the band of cells defined by f t z activation and odd re- pression that would potentially encircle the embryo is restricted to ventrolateral sites by a dorsolateral repres- sor (Fig. 8). In this model we need not postulate any ventral activators of ac/sc in the neurogenic region (ac- tivation is provided by pair-rule genes), just repressors in the non-neurogenic region.
Dist inct cis-actmg regulatory e lements are invo lved in the control of the pattern and level of ac and sc expression
Our initial dissection of the cis-acting regions of the ac gene suggests that the elements required to initiate the correct spatial expression patterns of ac and sc lie be- tween the two genes and are roughly separable into at least two distinct regions: one critical for the initiation of expression in row B, located 3' to the ac gene and 5' to the sc 8 breakpoint, and one critical for the initiation of expression in row D, located 3' to the sc 8 breakpoint and 5' to the sc gene {Figs. 2 and 3). Furthermore, our initial results from reporter constructs containing large regions of DNA from between the two genes show that this re- gion can drive weak l acZ expression in a pattern that resembles the endogenous ac (and sc) pattern. It is pos- sible that these regions may be further subdivided into enhancers operating in either even- or odd-numbered parasegments responsive to specific combinations of pair-rule genes. It is also likely that each register of pro- neural gene expression is driven by redundant elements spread over the large intergenic regulatory region.
2616 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Intrasegmental pattern formation
Although the intergenic region may be sufficient to initate the proper spatial pattern of ac (sc) gene expres- sion, the proper level of ac expression may require addi- tional control elements located 5' to ac responsive to the input of the ac, sc, and l ' s c proteins. We have shown that the level of ac gene expression (in row D) is reduced in l ' sc mutant embryos, and Van Doren et al. (1991) have identified a number of consensus-binding sites for the ac,
sc, and I ' sc proteins that lie within 1 kb 5' of the ac
promoter. Taken together, these observations suggest that 3' regulatory regions would largely control the spa- tial patterning of ac gene expression, whereas 5' ele- ments would augment its level of expression. Further analysis of these 3' spatial control elements will require higher levels of reporter gene expression than that cre- ated here. It may be necessary to surround the reporter gene with both 5' and 3' regulatory regions of ac in fu- ture constructs to achieve the levels of reporter gene ex- pression necessary for a more detailed dissection of these spatial control elements.
Once the global pattern of ac-positive proneural clus- ters is generated, local interactions between cells deter- mine individual cell fates. Initially, wi thin each cluster, a competi t ive process occurs, involving interactions be- tween the proneural and neurogenic genes, which singles out one cell from the cluster as the future NB (de Celis et al. 1991a, b; Heitzler and Simpson 1991). An increased level of proneural protein expression in this cell relative to the remaining cells of the cluster reflects this decision (Cubas et al. 1991; Martin-Bermudo et al. 1991; Skeath and Carroll 1991, 1992). The future NB, then, starts a process termed lateral inhibition in which the neuro- genie genes function in a cell communica t ion path- way to remove proneural gene expression and thus the neural potential of the remaining cells of a cluster (Leh- mann et al. 1983; de la Concha et al. 1988; Brand and Campos-Ortega 1989; Cabrera 1990; Skeath and Carroll 1992; for review, see Simpson 1990; Campos-Ortega 1991). This repression of proneural gene expression al- lows for the subsequent epidermal development of these cells.
Gene t i c control of t w o - d i m e n s i o n a l pa t t erns
The body of evidence presented here strongly supports the idea that the two-dimensional pattern of NBs arises from the sequential action of two genetic regulatory sys- tems. Initially, the two axis-determining genetic regula- tory systems generate the precise spatial array of proneu- ral clusters. Then, wi th in each cluster, regional and cell- specific genetic regulatory systems (e.g., the proneural and neurogenic genes} assign cells discrete fates. In the future it will be very useful to dissect the complex com- posite enhancers of ac and sc that appear to be regulated by different batteries of spatial regulators. It will be a challenge to identify min imal elements that direct gene expression in different cell clusters and to determine how regulatory proteins act on these elements to gener- ate a two-dimensional pattern of gene expression.
Materials and methods
Fly strains
Fly stocks were obtained from the laboratories of Walter Geh- ring, Michael Hoffmann, Steve DiNardo, Eric Wieschaus, and Alain Ghysen and from the Bowling Green, Indiana, and T/ib- ingen stock centers. Fly stocks harboring the following DNA rearrangements within the AS-C were used: Df(1)yaPLsc 8R, Df(1)sc 8taR, Df(1)sc aLgR, In(1)sc 8, In(1)ac 3, and In(I)sc v2. Fly stocks carrying the following mutations to segment polarity genes were used: en cxl, armadillo xK22, wg acaa, hedgehog lacaa, patched 1Nl~ gsb IIx62, nkd 7Esg, Cubitus interruptus D~ (Ci~ and en a (en and invected double mutant). Fly strains car- rying the following pair-rule mutations were used: ftz wa~ od- d II1D36, prd nBaa, eve 1a7, opa nC71, slp aaaB, runt [Df(1)B57], and hairy 7Hoa. Fly stocks harboring the following DV mutations were used: dpp 61, sna u6~ tld Ba, twi u~ and zen wa6. The iden- tity of the In(1)sc s, In(1)ac 3, In(1)sc vr Df(1)yaPLsc eR, Df(1)sc 8LaR, and Ci ~ fly stocks was verified by the cuticular phenotypes of adult flies. The identity of all pair-rule mutant stocks and the armadillo x~aa, wg ~caa, hedgehog l acaa, patched INt~ and nkd 7E89 fly stocks was confirmed by deter- mining the expression pattern of en in each stock. In all stocks the expected alteration to en expression occurred in ~25% of the properly staged embryos. The identity of the en B, gsb nxez, and Df(1)sc 4LvR fly strains was determined by staining embryos from each stock for en, gsb, or l'sc protein, respectively, and observing that ~25% of the appropriately staged embryos of each stock were devoid of protein expression. The identity of the en cx~ stock was verified by the cuticular phenotype of em- bryos from this stock.
ac reporter constructs
Putative regulatory regions of ac were cloned into phsplacCa- SpeR (H. Nelson and A. Laughon, in prep.), which has a disabled heat shock promoter fused to the lacZ gene. Transformed lines were established by standard methods using the white gene as a selectable marker. Reporter constructs were coinjected with a helper P-element, pPi 25.7wc {Karess and Rubin 1984), into pre- cellular embryos from the y w line (Lindsley and Zimm 1992). The 3.2-kb ac construct shown in Figure 3 (101R3.2) was con- strutted by religation of the 1.0- and 2.2-kb EcoRI fragments (between 57.4 and 54.2) from ~, scl01 (Campuzano et al. 1985] into pBluescnpt(KS) (Stratagene) to generate p101R3.2. Con- struct 101H10 was subcloned as a HindIII fragment from k scl01 (a kind gift from Juan Modolell, Centro de Biologia Mo- lecular, Madrid, Spain) into pBluescript(KS) (Stratagene) to gen- erate p 101H 10. Both fragments were then isolated as KpnI-NotI fragments and were cloned into the unique KpnI- and NotI- cloning sites of phsplac CaSpeR in the sense orientation with respect to lac.
Four lines of the 101H10 construct and three lines of the 101R3.2 construct were obtained. All lines assayed resulted from independent integration events. Three of the four 101H10 lines exhibited the identical expression pattern (shown in Fig. 3), with the other line showing no detectable lacZ expression. All three of the 101R3.2 lines exhibited the expression pattern shown in Figure 3, although two lines expressed lacZ at lower levels.
Immunohistochemistry and in situ hybridization
Embryos were fixed in 4 ml of fix buffer (100 mM PIPES at pH 6.9; 2 mM EGTA; 1 rnM MgSO4), 1 ml of 37% formaldehyde {Sigma), and 5 ml of heptane, following standard protocols. Sin-
GENES & DEVELOPMENT 2617
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

Skeath et al.
gle- and double-antibody labeling experiments were carried out in Eppendorf tubes, as described in Skeath and Carroll (1992) except as noted below.
For staining embryos using peroxidase, Vector's ABC elite kit was used and the following changes were made to the peroxi- dase staining protocol (Skeath and Carroll 1992): After 30 min of washes in 100 mM Tris-HC1 (pH 6.8) and before the develop- ment of the peroxidase reaction (see Skeath and Carroll 1992), embryos were preincubated for 2 rain either in a solution con- taining 100 mM Tris-HC1 (pH 6.8) and 0.5 mg/ml of diamino- benzidine (DAB; for a brown reaction product) or in a solution containing 100 mM Tris-HC1 (pH 6.8), 0.5 mg/ml DAB, and 0.03% (wt/vot) Co 2+ and Ni 2+ ions (for a black reaction prod- uct). After 2 min of preincubation, the solution was removed and the stain was developed by the addition of 100 mM Tris-HC1 (pH 6.8), 0.5 mg/ml of DAB, and 0.002% H20 2. The reaction was monitored visually by placing -20 Izl of embryos on a mi- croscope slide and observing the reaction through a dissecting scope. When the stain had developed to the desired intensity, the reaction was stopped by the addition of 5 ~1 of 20% sodium azide. All other steps were performed as described previously (Skeath and Carroll 1992).
For detection of ac protein, monoclonal antibody (mAb) 990ESF1 was used at a 1 : 3 dilution; for detection of l'sc pro- tein, a rat anti-l'sc antibody (generously provided by Fernando Jimenez, Centro de Biologia Molecular, Madrid, Spain) was used at a 1 : 500 dilution; for detection of en protein, mAbINV4D9 (kindly provided by Nipam Patel, Carnegie Institute of Embry- ology, Baltimore, MD), which recognizes both en and invected, was used at a 1 : 1 dilution; and for detection of gsb-distal protein, rat mAbl6F12 and mAblOE10 (generously provided by Anne Ungar and Bob Holmgren, Northwestern University, Evanston, ILl were used together each at a 1 : 5 dilution. For sequentially double-labeling embryos for ac protein and either l'sc, en, or gsb protein, embryos were first stained with the ac antibody and then stained with one of the other three antibod- ies. Double-labeling embryos for ac and l'sc was done by use of two sequential peroxidase staining reactions, whereas double- labeling embryos for ac and either en or gsb was done by use of a peroxidase staining reaction for ac followed by an alkaline- phosphatase staining reaction for en or gsb (see Skeath and Car- roll 1992). To identify unambiguously early gsb and nkd em- bryos, we balanced each stock over balancer chromosomes that express lacZ under the control of the ftz stripe element. Em- bryos were then doubly or triply labeled for ac and lacZ or for ac, lacZ, and en. Mutant embryos were identified as those that did not express lacZ. Df(1)sc atgR embryos were identified un- ambiguously by double-labeling embryos for ac and l'sc. Mutant embryos were identified as those that expressed no l'sc protein.
In situ hybridization was carried out as described in Tautz and Pfeifle (1989). For detection of sc RNA expression, embryos were hybridized in a total volume of 100 vtl of hybridization solution containing 17 ng of a digoxygenin-labeled DNA probe from the 1.5-kb EcoRI fragment of a sc eDNA (kindly provided by the late Carlos Cabrera). For the simultaneous detection of both sc and wg RNA expression, embryos were hybridized (100 }xl total volume) with the sc probe (17 ng) and with 17 ng of a digoxygenin-labeled DNA probe from the 1-kb EcoRV-HindIII fragment of a wg cDNA (Rijsewijk et al. 1987). Detection of lacZ RNA was carried out as described in Tautz and Pfeifle (I989), as modified by Jiang et al. {1991).
A c k n o w l e d g m e n t s
We thank Juan Modolell, Sonsoles Campuzano, and the late Carlos Cabrera for providing AS-C genomic and cDNA clones.
We are grateful to Ueli Grossniklaus and Walter Gehring and to Mark Benedyk and Steve DiNardo for communication of results before publication and for sending the slp a34B and en B fly strains, respectively. We are also grateful to Steve Cohen for communication of results before publication. We thank Anne Ungar and Bob Holmgren for providing monoclonal antibodies to gsb-distal before publication and Femando limenez for send- ing the 1'sc antiserum. We also thank Heidi Nelson and Allen Laughon for providing us with the phsplacCaSpeR vector. We thank Allen Laughon, Lisa Nagy, Teresa Orenic, and Jim Williams for their comments on the paper; Kathy Vorwerk for technical assistance; Leanne Olds for help with the figures and diagrams; and Jamie Wilson for help with the preparation of the manuscript. This work was supported by a National Institutes of Health (NIH) predoctoral traineeship (GM-07215) to J.B.S., a NIH postdoctoral fellowship (GM-1441) to G.P., a National Sci- ence Foundation Presidential Young Investigators Award, the Shaw Scholar's Program of the Milwaukee Foundation, and the Howard Hughes Medical Institute.
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 USC section 1734 solely to indicate this fact.
R e f e r e n c e s
Baumgartner, S., D. Bopp, M. Burri, and M. Noll. 1987. Structure of two genes at the gooseberry locus related to the paired gene and their spatial expression during Drosophila neuro- genesis. Genes & Dev. 1: 1247-1267.
Baumgartner, S. and Noll, M. 1990. Network of interactions among pair-rule genes regulating paired expression during primordial segmentation of Drosophila. Mech. Dev. 1: 1-18.
Brand, M. and J.A. Campos-Ortega. 1989. Two groups of inter- related genes regulate neurogenesis in Drosophila melano- gaster. Wilhelm Roux's Arch. Dev. Biol. 197: 457-470.
Cabrera, C.V. 1990. Lateral inhibition and cell fate during neu- rogenesis in Drosophila: The interactions between scute, Notch, and Delta. Development 109: 733-742.
Cabrera, C.V., A. Martinez-Arias, and M. Bate. 1987. The ex- pression of three members of the achaete-scute complex cor- relates with neuroblast segregation in Drosophila. Cell 50: 425-433.
Campos-Ortega, J. and V. Hartenstein. 1985. The embryonic development of Drosophila melanogaster. Springer-Verlag, Berlin, Germany.
Campos-Ortega, J.A. 1991. Genetic mechanisms of early neuro- genesis in Drosophila melanogaster. Int. Rev. Cytol. 124: 1- 41.
Campuzano, S., L. Carramolino, C.V. Cabrera, and M. Ruiz- Gomez. 1985. Molecular genetics of the achaete-scute gene complex of D. melanogaster. Cell 40: 327-338.
Carroll, S.B. and M.P. Scott. 1986. Zygotically-active genes that affect the spatial expression of the fushi tarazu segmenta- tion gene during early Drosophila embryogenesis. Cell 45:113-126.
Carroll, S.B., A. Laughon, and B.S. Thalley. 1988. Expression, function, and regulation of the hairy segmentation protein in the Drosophila embryo. Genes & Dev. 2: 883-890.
Cohen, B., A.A. Simcox, and S.M. Cohen. 1992. Allocation of the thoracic imaginal primordia in the Drosophila embryo. Development (in press).
Coulter, D.E., E.A. Swaykus, M.A. Beran-Koehn, D. Goldberg, I. Wieschaus, and P. Schedl. 1990. Molecular analysis of odd-
2618 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

lntrasegmental pattern formation
skipped, a zinc finger encoding segmentation gene with a novel pair-rule expression pattern. EMBO J. 8: 3795--3804.
Cubas, P., J.-F. Celis, S. Campuzano, and J. Modolell. 1991. Pro- neural clusters of achaete-scute expression and the genera- tion of sensory organs in the Drosophila imaginal wing disc. Genes & Dev. 5: 996-1008.
Dambly-Chaudiere, C., A. Ghysen, L.Y. Jan, and Y.N. Jan. 1988. Independent subpatterns of sense organs require indepen- dent genes of the achaete-scute complex in Drosophila lar- vae. Genes & Dev. 1: 297-306.
de Celis, J. F., M. Mari-Beffa, and A. Garcia-Bellido. 1991a. Func- tion of trans-acting genes of the achaete-scute complex in sensory organ patterning in the mesonatum of Drosophila. Wilhelm Roux's Arch. Dev. Biol. 200: 64-76.
- - . 1991b. Cell-autonomous role of Notch, an epidermal growth factor homologue, in sensory organ differentiation in Drosophila. Proc. Natl. Acad. Sci. 88: 632--636.
de la Concha, A., A. Dietrich, D. Weigel, and J.A. Campos- Ortega�9 1988. Functional interactions of neurogenic genes of Drosophila melanogaster. Genetics 118: 499-508.
DiNardo, S. and P. O'Farrell. 1987. Establishment and refine- ment of segmental pattern in the Drosophila embryo: Spa- tial control of engrailed expression by pair-rule genes. Genes & Dev. 1: 1212-1225.
Doe, C.Q. and C.S. Goodman. 1985. Early events in insect new rogenesis. I. Development and segmental differences in the pattern of neuronal precursor ceils�9 Dev. Biol. 111: 193-205.
Frasch, M. and M. Levine. 1987. Complementary patterns of even-skipped and fushi tarazu expression involve their dif- ferential regulation by a common set of segmentation genes in Drosophila�9 Genes & Dev. 1: 981-995.
Ghysen, A. and C. Dambly-Chaudiere. 1989. Genesis of the Drosophila peripheral nervous system. Trends Genet. 5: 251-255.
Grossniklaus, U., R.K. Pearson, and W.J. Gehring. 1992. The Drosophila sloppy paired locus encodes two proteins that are homologous to mammalian transcription factors. Genes & Dev. 6: 1030-1051.
Heitzler, L. and P. Simpson. 1991. The choice of cell fate in the epidermis of Drosophila. Cell 64: 1083-1092.
Hiromi, Y. and W. Gehring. 1987. Regulation and function of the Drosophila segmentation gene fushi tarazu. Cell 50: 963-974.
Ingham, P.W. 1988. The molecular genetics of embryonic pat- tern formation in Drosophila. Nature 335: 25-34.
Ingham, P.W. and A. Martinez-Arias. 1992. Boundaries and fields in embryos. Cell 68: 221-236.
Jan, Y.N. and L.Y. Jan. 1990. Genes required for specifying cell fates in the Drosophila embryonic nervous system. Trends Neurosci. 13: 493-498.
Jiang, l., D. Kosman, Y.T. Ip, and M. Levine. 1991. The dorsal morphogen gradient regulates the mesoderm determinant twist in early Drosophila embryos. Genes & Dev. 5: 1881- 1891.
Jimenez, F. and J.A. Campos-Ortega. 1990. Defective neuroblast commitment in mutants of the achaete-scute complex and adjacent genes of D. melanogaster. Neuron 5: 81-89.
Karess, R. and G. Rubin. 1984. Analysis of P transposable ele- ment functions in Drosophila�9 Cell 38: 135-146.
Kosman, D., Y.T. Ip, M. Levine, and K. Arora. 1991. Establish- ment of the mesoderm-neuroectoderm boundary in the Drosophila embryo. Science 254:118-122.
Lehmann, R., F. Jimenez, U. Dietrich, and J.A. Campos-Ortega. 1983. On the phenotype and development of mutants of early neurogenesis in Drosophila melanogaster. Wilhelm Roux's Arch. Dev. Biol. 192: 62-74.
Lindsley, D.L. and G.G. Zimm. 1992�9 The genome of Drosophila melanogaster. Academic Press, San Diego, CA.
Martin-Bermudo, M.D., C. Martinez, A. Rodriguez, and F. Jimenez. 1991. Distribution and function of the lethal of scute gene during early neurogenesis in Drosophila. Devel- opment 113: 445--454.
Martinez, C. and J. Modolell. 1991. Cross-regulatory interac- tions between the proneural achaete and scute genes of Drosophila. Science 251: 1485-1487.
Nfisslein-Volhard, C. 1991. Determination of the embryonic axes of Drosophila. Development (Suppl.) 1: 1-10.
Panzer, S., D. Weigel, and S.K. Beckendorf. 1992. Organogenesis in Drosophila metanogaster: Embryonic salivary gland de- termination is controlled by homeotic and dorsoventral pat- terning genes. Development 114: 49-57.
Patel, N.H., E.E. Ball, and C.S. Goodman. 1992. Changing role of even-skipped during the evolution of insect pattern forma- tion. Nature 357: 339-342.
Rijsewijk, F.M., E. Schuerrnann, P. Wagenaar, D. Parren, D. Weigel, and R. Nusse. 1987. The Drosophila homolog of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless. Cell 50: 649--657.
Romani, S., S. Campuzano, E. Macagno, and J. Modolell. 1989. Expression of achaete and scute genes in Drosophila imagi- nal discs and their function in sensory organ development. Genes & Dev. 3: 997-1007.
Shimell, M.J., E.L. Ferguson, S.R. Childs, and M.B. O'Connor. 1991. The Drosophila dorsal-ventral patterning gene tolloid is related to human bone morphogenetic protein 1. Cell 67:469--481.
Simpson, P. 1990. Lateral inhibition and the development of the sensory bristles of the adult peripheral nervous system of Drosophila. Development 109: 509-519.
Skeath, J.B. and S.B. Carroll. 1991. Regulation of achaete-scute gene expression and sensory organ pattern formation in the Drosophila wing. Genes & Dev. 5: 984-995.
�9 1992. Regulation of proneural gene expression and cell fate during neuroblast segregation in the Drosophila em- bryo. Development 114: 939-946�9
St. Johnston, R.D. and W.M. Gelbart. 1987. Decapentaplegic transcripts are localized along the dorsal-ventral axis of the Drosophila embryo. EMBO ]. 6: 2785-2791.
Stem, C. 1954. Two or three bristles�9 Am. Sci. 42: 213-247. Taghert, P.H., C.Q. Doe, and C.S. Goodman. 1984. Cell deter-
mination and regulation during development of neuroblasts and neurones in grasshopper embryos. Nature 307: 163-165.
Tautz, D. and C. Pfeifle. 1989. A non-radioactive in situ hybrid- ization method for the localization of specific RNAs in Drosophila embryos reveals translational control of the seg- mentation gene hunchback. Chromosoma 98: 81-85.
Thisse, B., C. Stoetzel, C. Gorostiza-Thisse, and F. Perrin- Schmitt. 1988. Sequence of the twist gene and nuclear local- ization of its protein in endodermal cells of early Drosophila embryos. EMBO J. 7: 2175-2183.
Van Doren, M., H.M. Ellis, and J.W. Posakony. 1991. The Droso- phila extramacrochaetae protein antagonizes sequence-spe- cific DNA binding by daughterless/achaete-scute protein complexes. Development 113: 245-255.
GENES & DEVELOPMENT 2619
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from

10.1101/gad.6.12b.2606Access the most recent version at doi: 6:1992, Genes Dev.
J B Skeath, G Panganiban, J Selegue, et al. through a common intergenic control region.genes are controlled by combinations of axis-patterning genes Gene regulation in two dimensions: the proneural achaete and scute
References
http://genesdev.cshlp.org/content/6/12b/2606.full.html#ref-list-1
This article cites 47 articles, 18 of which can be accessed free at:
License
ServiceEmail Alerting
click here.right corner of the article or
Receive free email alerts when new articles cite this article - sign up in the box at the top
Copyright © Cold Spring Harbor Laboratory Press
Cold Spring Harbor Laboratory Press on February 18, 2022 - Published by genesdev.cshlp.orgDownloaded from