genetics of complement c4. two homoduplication haplotypes c4s c4s and c4f c4f in a family
TRANSCRIPT

Hum Genet (1982) 61 : 36-38
© Springer-Verlag 1982
Genetics of Complement C4. Two Homoduplication Haplotypes C4S C4S and C4F C4F in a Family
G. Bruun-Petersen 1, Lars U. Lamm 2, Bodil K. Jacobsen ~, and Tom Kristensen 3
1 Institute of Human Genetics and ZTissue Typing Laboratory, University of Aarhus, DK-8000 Aarhus C, Denmark 3 Tissue Typing Laboratory, State University Hospital, DK-2200 Copenhagen N, Denmark
Summary. A family in which two homoduplicated C4 haplo- types (or supergenes) segregate is described. One haplotype C4F'3 C4F'2.2 is composed of two C4F alleles and the other C4S'5.1 C4S*1 of two C4S alleles. The C4F duplication haplo- type is a partial inhibitor of the Rodgers antigen, and judged from our family and population material, it seems to be rather frequent and associated with HLAB*35, B f 'F, and HLAD/DR* 1. The C4S duplication haplotype is Rg(a-) and is not identified in individuals without another S, Ch(a+) variant.
Introduction
A C4 polymorphism in man was demonstrated by Teisberg et al. (1976, 1977) using agarose gel electrophoresis. The observation of O'Neill et al. (1978 a, b) that persons lacking the fast moving C4 bands are negative for the Rodgers antigen, Rg(a-), and that those lacking the slow moving bands are negative for the Chido antigen, Ch(a-), was a keystone in our understanding of the genetics of the C4 system. They concluded that the Chido antigen is a component of the C4S molecule and that the Rodgers antigen is a component of the C4F molecule and proposed that C4 is determined by two loci.
Olaisen et al. (1979) used the IgG fraction of an exceptionally strong anti-Chido antiserum to precipitate specifically the C4S fraction from persons with different C4 phenotypes and showed that some haplotypes contained both functioning C4F and C4S loci.
The introduction by Awdeh and Alper (1980) of pretreat- ment of plasma with neuraminidase before electrophoresis was a great technical advance. This technique gives easily recognizable C4 gene products with a three-band pattern, of which the most anodic band has the highest concentration.
By using this technique and by defining a C4F gene product as having hemolytically inactive or weak bands and being Rg(a+), Ch(a-), and a C4S gene product as having hemolytically strong bands and being Rg(a-), Ch(a+), Bruun-Petersen et al. (1981) were able to distinguish at least 12 different structural C4 gene products. These gene products gave rise to at least 12 different C4 haplotypes, of which 7 were duplications, each composed of an F and an S gene.
Nordhagen et al. (1981) described a double haplotype C4FI coding for the two fast C4 proteins F and I and carrying a weak Rg activity.
This work was supported by grant No 12-1727 from the Danish Medical Research Council Offprint requests to: G. Bruun-Petersen
We present a Danish family in which two homoduplication haplotypes segregate, one haplotype with two C4S genes and the other haplotype with two C4F genes.
Materials and Methods
The family with one grandparent and four children was found through an investigation of C4 in families with crossovers between the HLA system and GLO. The methods used for C4 typing and typing of other markers were as described by Bruun- Petersen et al. (1981).
Results
The C4 phenotypes as seen after immunofixation are shown in Fig. 1 and after hemolytical development, in Fig. 2 for the seven members of the family. In the following, gene products are referred to by the main or most anodic band. The mother's mother (MM) has two fast, hemolytically weak gene products and one slow, hemolytically strong product (F3F2.2S1). The mother (M) has four gene products of which the two fastest are hemolytically weak and the two slow are hemolytically strong (F3F2.2S5.1S1). The father (P) has one fast, hemolytically weak and two slow, hemolytically strong gene products (F3S2S 1). The first two children have identical C4 phenotypes composed of two hemolytically strong gene products ($5.1S1). Child 3 has one fast, hemolytically weak and three slow, hemolytically strong gene products (F3S5.1S2S1) and child 4 has two fast and weak gene products and one slow, hemolytically strong product (F3F2.2S 1).
Figure 3 is a graphic representation of the five gene products in the family. The products F3, F2.2, $2, and S1 have been described previously (Bruun-Petersen et al. 1981), while the product assigned $5.1 is positioned between the two most cathodic bands of F3. In Fig. 3 the results of the determination of the Rodgers and Chido antigens have also been given. When segregating together, the gene products F3 and F2.2 are partial inhibitors of Rodgers. The $5.1 product is Rg(a-) and is not identified in individuals without another S, Ch(a+) variant.
The genetic interpretation of the family is given in Fig. 4, in which the order of the loci is HLAA, HLAC, HLAB, Bf C4, D/DR, GLO. Child 1 shows a maternal recombinant between the C4 and GLO loci, but since the mother is a D/DR homozygote, the crossover cannot be localized with respect to that locus. Apart from this, no crossover is seen in the family. The central observation is that child 3 has three C4S gene products and there-
0340-6717/82/0061/0036/$ 01.00
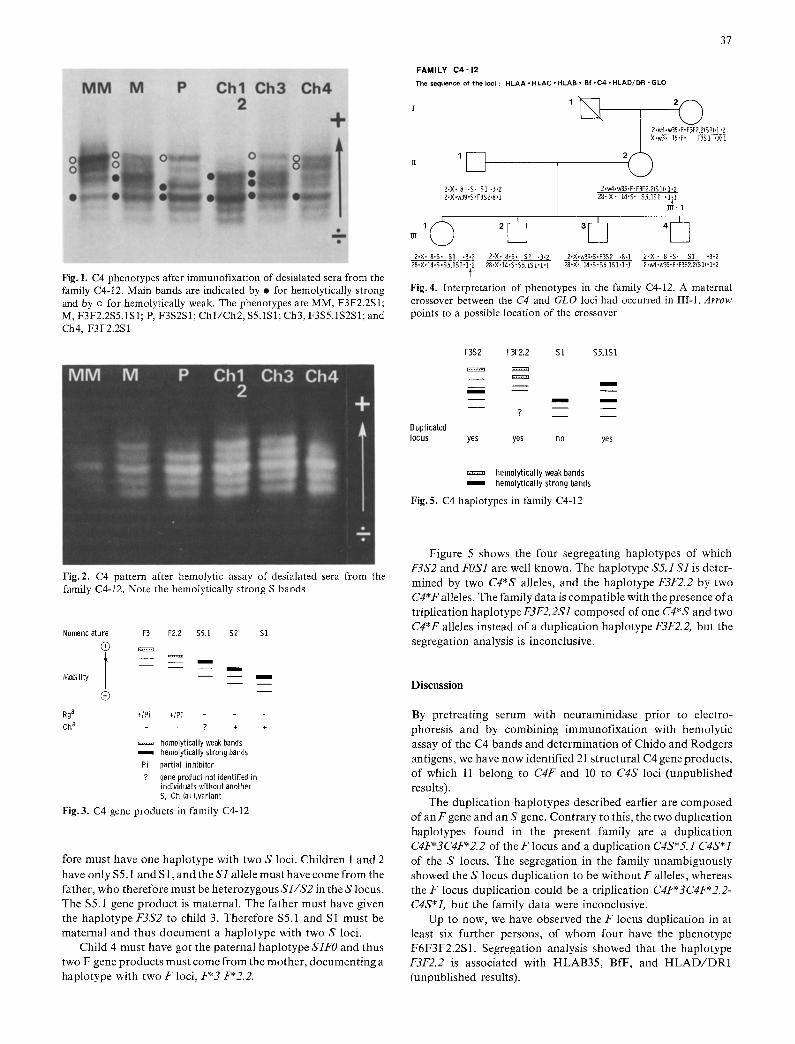
Fig. 1. C4 phenotypes after immunofixation of desialated sera from the family C4-12. Main bands are indicated by • for hemolytically strong and by o for hemolytically weak. The phenotypes are MM, F3F2.2S1; M, F3F2.2S5.1S 1; P, F3S2S1; Chl/Ch2, S5.1SI; Ch3, F3S5.1S2S1; and Ch4, F3F2.2S 1
37
II1
2.X- 8-S- S1 .3.2 2-X. 8.S. S1 .3.2 2.X.w39.S-F3S2 -8-1 2.X • 8 -S' S1 -3-2
FAMILY C 4 - 12
The sequence o f the loci : HLAA oHLAC ,HLAB • Bf ,C4 , HLAD/DR ,GLO
2,X. 8 -S- S1.3.2 2.w4.w35-F-F3F2.2(S1).I.2 2.X.w39"S-F3S2.8'l 28. X. 14"S" 55.1S1 '1~1
11I-1
28-X.14.S.SS.lSl']i2 28.X'lLL'S' $5.1S 1 • 1.I 28'X" 14"S'$3.1S1'1-1 2.w4.w33'F'F3F2.2(S 1)'1-2
Fig.4. Interpretation of phenotypes in the family C4-12. A maternal crossover between the C4 and GLO loci had occurred in III-1. Arrow points to a possible location of the crossover
F3S2 F3F2.2 SI SS.ISI
?
D up licated locus yes yes no yes
hemolytically weak bands hemolytically strong bands
Fig. 5. C4 haplotypes in family C4-12
Fig.2. C4 pattern after hemolytic assay of desialated sera from the family C4-12. Note the hemolytically strong S bands
Nomenclature ®
Mobility l
®
Rg a
Ch a
F3 F2.2 $5.1 $2 SI
m h l i
+/Pi +/Pi ? +
hemolytically weak bands hemolytically strong bands
Pi partial inhibitor
? gene product not identified in i nd iv idua ls w i thou t another S, Ch (a+),variant
Fig. 3. C4 gene products in family C4-12
fore must have one haplotype with two S loci. Children 1 and 2 have only $5.1 and S 1, and the $1 allele must have come from the father, who therefore must be heterozygous S1 /$2 in the S locus. The $5.1 gene product is maternal. The father must have given the haplotype F3S2 to child 3. Therefore $5.1 and S1 must be maternal and thus document a haplotype with two S loci.
Child 4 must have got the paternal haplotype S1FO and thus two F gene products must come from the mother, documenting a haplotype with two F loci, F*3 F'2.2.
Figure 5 shows the four segregating haplotypes of which F3S2 and FOS1 are well known. The haplotype $5.1 $1 is deter- mined by two C4"S alleles, and the haplotype F3F2.2 by two C4*F alleles. The family data is compatible with the presence of a triplication haplotype F3F2.2S1 composed of one C4"S and two C4*F alleles instead of a duplication haplotype F3F2.2, but the segregation analysis is inconclusive.
Discussion
By pretreating serum with neuraminidase prior to electro- phoresis and by combining immunofixat ion with hemolytic assay of the C4 bands and determination of Chido and Rodgers antigens, we have now identified 21 structural C4 gene products, of which 11 belong to C4F and 10 to C4S loci (unpublished results).
The duplication haplotypes described earlier are composed of an F gene and an S gene. Contrary to this, the two duplication haplotypes found in the present family are a duplication C4["3C4F'2.2 of the F locus and a duplication C4S'5.1 C4S*1
of the S locus. The segregation in the family unambiguously showed the S locus duplication to be without F alleles, whereas the F locus duplication could be a triplication C4[~3C4F'2.2 -
C4S*1, but the family data were inconclusive. Up to now, we have observed the F locus duplication in at
least six further persons, of whom four have the phenotype F6F3F2.2S1. Segregation analysis showed that the haplotype F3F2.2 is associated with HLAB35, BfF, and H L A D / D R 1 (unpublished results).

38
Ins tead of assuming that the mo the r has two rare haplotypes, it would, of course, be more reasonable to assume that she has two c o m m o n haplotypes, bo th with an F and an S locus. However, this is not consistent with the family data wi thout the occurrence of several double crossovers between HLAB and GLO and in addi t ion to that , unequal crossovers in the C4 region. Not even illegitimacy could offer a simpler explanat ion of the family wi thout these assumptions.
The de te rmina t ion of the Rodgers and Chido antigens showed the F dupl icat ion to be a part ial inhibi tor of the Rodgers and thus confirms the observat ion made by Nordhagen et al. (1981). The S locus dupl icat ion gene p roduc t was not identified in individuals wi thout ano the r S, Ch(a+) var iant and therefore was inconclusive with respect to normal Chido activity.
Acknowledgements. Our thanks to Mrs. Dorte Laursen for skillful secretarial assistance and to Ms. Kirsten Milgaard and Mrs. Karen Tobler for excellent photographic work.
References
Bruun-Petersen G, Lamm LU, Sorensen I J, Buskjaer L, Mortensen JP (1981) Family studies of complement C4 and HLA in man. Hum Genet 58 : 260-267
Nordhagen R, Olaisen B, Teisberg P, Gedde-Dahl T Jr, Thorsby E (198 I) C4 haplotype products and partial inhibition of anti-Rodgers sera. J Immunogenet 8:485-491
Olaisen B, Teisberg P, Nordhagen R, Michaelsen T, Gedde-Dahl T Jr (1979) Human complement C4 locus is duplicated on some chromo- somes. Nature 279:736-737
O'Neill G J, Yang SY, Tegoli J, Berger R, Dupont B (1978a) Chido and Rodgers blood groups are distinct antigenic components of human complement C4. Nature 273:668-670
O'Neill GJ, Yang SY, Dupont B (1978b) Two HLA-linked loci con- trolling the fourth component of human complement. Proc Natl Acad Sci USA 75:5165-5169
Teisberg P, /~kesson I, Olaisen B, Gedde-Dahl T, Thorsby E (1976) Genetic polymorphism of C4 in man and localisation of a structural C4 locus to the HLA gene complex of chromosome 6. Nature 264: 253-254
Teisberg P, Olaisen B, Jonassen R, Gedde-Dahl T, Thorsby E (1977) The genetic polymorphism of the fourth component of human complement: Methodological aspects and a presentation of linkage and association data relevant to its localization in the HLA region. J Exp Med 146:1380-1389
Awdeh ZL, Alper CA (1980) Inherited structural polymorphism of the fourth component of human complement. Proc Natl Acad Sci USA 77:3576-3580 Received April 4, 1982