germinaÇÃo e propagaÇÃo in vitro de myracrodruon …§ões-teses/teses/2016... · Às minhas...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E ZOOTECNIA
Programa de Pós-Graduação em Agricultura Tropical
GERMINAÇÃO E PROPAGAÇÃO in vitro DE Myracrodruon
urundeuva Allemão (AROEIRA-DO-SERTÃO)
DAYANE ÁVILA FERNANDES
C U I A B Á - MT 2016
0

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E ZOOTECNIA
Programa de Pós-Graduação em Agricultura Tropical
GERMINAÇÃO E PROPAGAÇÃO in vitro DE Myracrodruon
urundeuva Allemão (AROEIRA-DO-SERTÃO)
DAYANE ÁVILA FERNANDES
Bióloga
Orientador: Prof. Dr. SEBASTIÃO CARNEIRO GUIMARÃES
Co-orientadora: Profª. Dra. PATRICIA HELENA DE AZEVEDO
Tese apresentada à Faculdade de Agronomia, Medicina Veterinária e Zootecnia da Universidade Federal de Mato Grosso, para obtenção do título de Doutora em Agricultura Tropical.
C U I A B Á - MT
2016
0

0


Aos meus pais, Vera e Osmar,
os melhores anjos que Deus me deu
a quem devo tudo que sou.
DEDICO.
Ao meu irmão Osmar Junior,
a melhor parte de mim,
meu orgulho e meu parceiro.
OFEREÇO.

AGRADECIMENTOS
Primeiramente a Deus criador do céu e da terra, de todas as coisas visíveis e
invisíveis, que permitiu que tudo isso pudesse ser realizado.
À Universidade Federal de Mato Grosso e ao Programa de Pós- Graduação em
Agricultura Tropical pela oportunidade da realização do doutorado, ao dispor da
infra-estrutura física e acadêmica, fundamentais a continuidade da formação de
seus alunos.
Aos professores do Programa de Pós-Graduação em Agricultura Tropical, que
repassaram seus conhecimentos contribuindo tanto para o crescimento pessoal,
quanto para o desenvolvimento profissional dos alunos.
À CAPES pela concessão da bolsa de estudos.
Ao meu orientador Prof. Dr. Sebastião Carneiro Guimarães, pelos conhecimentos
transmitidos, sugestões e críticas enriquecedoras ao trabalho. Pelo respeito,
profissionalismo e confiança na realização do doutorado.
À minha co-orientadora Prof.ª Dr.ª Patricia Helena de Azevedo e também à Prof.ª
Dr.ª Virginia Helena de Azevedo, pelo apoio e sábios ensinamentos, pelas
contribuições valiosas ao nosso trabalho, por toda paciência na correção e ótimas
ideias para melhoria do texto. Além de tudo por todo incentivo ao longo do curso e
por acreditarem em mim.
Ao Prof. Dr. Gustavo Alves Pereira por todo apoio na realização dos experimentos e
pelas importantes contribuições neste trabalho. Gostaria que soubesse que você é
um exemplo de dedicação ao ensino superior e à pesquisa a ser seguido.
À Prof.ª Dr.ª Andrea Carla Dalmolin por todas as sugestões para melhoria deste
trabalho e, acima de tudo pela amizade, por todas as palavras de força, carinho e
cuidado de sempre. Agradeço ainda pela oportunidade de um dia ter sido chamada
por você para dar uma palestra num evento do Univag Centro Universitário, o local
que hoje construo diariamente minha carreira profissional.
Ao Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da
Empresa Mato-grossense de Pesquisa, Assistência e Extensão Rural (EMPAER-
MT) em especial, por disponibilizar o espaço do Laboratório de Cultura de Tecidos
Vegetais, indispensável para a realização do experimento. Inclusive aos meus
queridos Lefayette Michele, Elder Cassemiro, Marcílio Santaella, Islayne Flenga e
Humberto pela companhia, amizade e por todo apoio. Não posso esquecer também
de agradecer pelos lanches e risadas! Vocês são muito especiais para mim.

Ao Univag Centro Universitário, instituição que fiz minha graduação pela
oportunidade profissional. Em especial aos meus coordenadores Rogério Castro,
Marcia Nassarden e Eliana Santini pela confiança e compreensão. Sou
profundamente grata pelo aprendizado que tenho diariamente nas disciplinas que
vocês me confiaram em seus cursos.
Ao meu querido Fabio pela paciência, amor, carinho, companheirismo, por estar ao
meu lado dando toda atenção e amparo. E também a minha madrinha Edileuza
Machado e sua família por fazerem parte da minha vida, pessoas que amo
incondicionalmente. E ainda a Maia, minha companhia de quatro patas, que está
sempre comigo inclusive nos incontáveis dias vendo esse trabalho ser construído e
crescer.
Aos colegas e amigos da pós-graduação, turma de mestrado e doutorado de 2012,
pela agradável companhia nos bons momentos, viagens, palestras e intervalos das
aulas e estudos, assim como nos sufocos e desesperos!!! Em especial a Debora
Jardini, Andressa Ricci e Ana Carla Stieven, e também às quatis Danielle Muller,
Glauce Portela e Josilaine Gonçalves, sou muito feliz por ter conhecido vocês e por
todas as coisas boas que passamos juntas, que possamos manter esta amizade
para sempre.
Às minhas amigas Ana Carolina Borges, Erika Segóvia, Juliane Dias e Silvia
Pimenta, vocês são as melhores amigas que a vida me permitiu escolher, obrigada
pelas conversas, conselhos e carinho, por estarem ao meu lado mesmo na minha
ausência nos fins de semana intermináveis de estudo para conclusão desta etapa.
Às Rossitas Isadora Pacheco, Francieli Peruzzo e Larissa Turini minhas amigas e
vizinhas, pela presença quase diária em minha vida e por toda força e incentivo.
Aos meus alunos de graduação, em especial aos meus orientados Francieli
Peruzzo e Josemar Potencio, e também ao Diego Costa por toda ajuda em campo
e no laboratório, carregando caixas de sementes no sol ou lavando vidraria sem
fim, mas sempre com sorriso no rosto e com boa vontade no coração.
Por fim, a todos que direta ou indiretamente contribuíram para a realização deste
trabalho, muito obrigada de todo coração.

SUMÁRIO 1 INTRODUÇÃO .......................................................................................... 13
1.1 Referências Bibliográficas...................................................................... 15 2 REVISÃO DE LITERATURA .................................................................... 17 2.1 A espécie Myracrodruon urundeuva Allemão ........................................ 17
2.2 Propagação ............................................................................................ 19 2.3 Fatores que influenciam na micropropagação ....................................... 22
2.3.1 Desinfestação do material vegetal ...................................................... 22 2.3.2 Estabelecimento in vitro a partir da germinação de sementes ............ 23
2.3.3 Multiplicação de brotações .................................................................. 24 2.3.4 Aclimatização e enraizamento ex vitro ................................................ 26
2.4 Referências Bibliográficas...................................................................... 28 3 HIPOCLORITO DE SÓDIO NA DESINFESTAÇÃO E GERMINAÇÃO in vitro DE SEMENTES DE Myracrodruon urundeuva Allemão (AROEIRA-DO-SERTÃO) ..................................................................................................... 37
RESUMO ..................................................................................................... 37
ABSTRACT .................................................................................................. 38 3.1 Introdução .............................................................................................. 39 3.2 Material e Métodos ................................................................................ 40
3.3 Resultados e Discussão ........................................................................ 43 3.4 Conclusão .............................................................................................. 48
3.5 Referências Bibliográficas...................................................................... 48 4 FUNGICIDAS NA DESINFESTAÇÃO DE SEMENTES DE Myracrodruon urundeuva Allemão (AROEIRA-DO-SERTÃO) ............................................ 51
RESUMO ..................................................................................................... 51 ABSTRACT .................................................................................................. 52
4.1 Introdução .............................................................................................. 53 4.2 Material e Métodos ................................................................................ 54
4.3 Resultados e Discussão ........................................................................ 58 4.4 Conclusão .............................................................................................. 62
4.5 Referências Bibliográficas...................................................................... 62 5 CONCENTRAÇÕES DE SAIS DO MEIO DE CULTURA MS NO ESTABELECIMENTO in vitro DE Myracrodruon urundeuva Allemão ......... 66
RESUMO ..................................................................................................... 66 ABSTRACT .................................................................................................. 67 5.1 Introdução .............................................................................................. 68 5.2 Material e Métodos ................................................................................ 69
5.3 Resultados e Discussão ........................................................................ 73 5.4Conclusão ............................................................................................... 78
5.5 Referências Bibliográficas...................................................................... 78 6 MULTIPLICAÇÃO in vitro DE Myracrodruon urundeuva Allemão EM CONCENTRAÇÕES DE BAP (6-BENZILAMINOPURINA) ........................ 81 RESUMO ..................................................................................................... 81 ABSTRACT .................................................................................................. 82
6.1 Introdução .............................................................................................. 83 6.2 Material e Métodos ................................................................................ 84 6.3 Resultados e Discussão ........................................................................ 85

6.4 Conclusão .............................................................................................. 91 6.5 Referências Bibliográficas...................................................................... 91 7 ÁCIDO INDOLBUTÍRICO NO ENRAIZAMENTO ex vitro DE Myracrodruon urundeuva Allemão (AROEIRA-DO-SERTÃO) ............................................ 95
7.1 Introdução .............................................................................................. 97 7.2 Material e Métodos ................................................................................ 98 7.3 Resultados e Discussão ...................................................................... 101 7.4 Conclusão ............................................................................................ 105
7.5 Referências Bibliográficas.................................................................... 105 8 CONCLUSÃO GERAL ........................................................................... 111

GERMINAÇÃO E PROPAGAÇÃO in vitro DE Myracrodruon urundeuva
Allemão (AROEIRA-DO-SERTÃO)
RESUMO - A aroeira-do-sertão (Myracrodruon urundeuva Allemão) é uma Anacardiaceae com propriedades medicinais e visibilidade econômica na indústria madeireira. Os objetivos neste trabalho foram: 1) avaliar tempos de exposição e doses de hipoclorito de sódio visando a desinfestação e germinação de sementes; 2) analisar a eficiência de fungicidas na assepsia; 3) comparar diferentes concentrações de sais do meio de cultura MS no desenvolvimento inicial in vitro; 4) verificar o efeito de diferentes concentrações de 6-benzilaminopurina (BAP) na indução de brotações; 5) testar diferentes concentrações de ácido indolbutírico (AIB) e substratos no enraizamento ex vitro. Foram realizados cinco experimentos compreendendo as etapas da micropropagação: assepsia, estabelecimento, multiplicação e aclimatação. Os experimentos relativos ao cultivo in vitro foram conduzidos no Laboratório de Cultura de Tecidos do Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da EMPAER-MT. A aclimatação (ex vitro) foi realizada no viveiro do Univag - Centro Universitário, sendo ambos localizados no município de Várzea Grande – MT. Primeiramente foi definida a assepsia pelo teste de tempos de exposição (10, 15 e 20 minutos) e concentrações de hipoclorito de sódio (0,0; 0,5; 1,0; 1,5 e 2,0%) em três matrizes de aroeira-do-sertão (M1, M2 e M3). Posteriormente foram avaliados oito tratamentos de desinfestação química através do contato de fungicidas nas sementes. No estabelecimento foi verificada a concentração adequada do meio de cultura MS [Murashige e Skoog, 1962] (0; 25; 50; 75 e 100%) na germinação in vitro. Com relação à multiplicação foram avaliadas concentrações de BAP (0,0; 3,0; 6,0; 9,0 e 12,0 ml L-1) em duas matrizes de aroeira (M2 e M3). Na última etapa, a aclimatização e o enraizamento ex vitro foram testados com brotações da matriz M3 em dois substratos e concentrações de AIB (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1). Na assepsia, a dose de 1,5% de hipoclorito de sódio em contato por 15 minutos e 0,5 ml do fungicida Vitavax-thiram® (1% m/v carboxina + 1% m/v tiram), apresentou eficiência na desinfestação de aroeira-do-sertão nas matrizes testadas. No estabelecimento, 25% da concentração de sais do meio MS favoreceu a baixa contaminação, maior germinação e maior comprimento de parte aérea em plântulas de aroeira-do-sertão. A concentração de 3,0 mg L-1 de BAP apresentou efeito satisfatório no desenvolvimento de brotações para a multiplicação in vitro. E para finalizar a elevada taxa de mortalidade (97,9%) pode ser associada às mudanças drásticas nas condições ambientais sofridas pelas plântulas, sugerindo o desenvolvimento de protocolos mais eficientes nessa etapa.
Palavras-chave: micropropagação, assepsia, estabelecimento, aclimatação.
5
5

GERMINATION AND SPREAD in vitro OF Myracrodruon urundeuva
Allemão
ABSTRACT – The Myracrodruon urundeuva Allemão is an Anacardiaceae with medicinal properties and economic visibility in the timber industry. The objectives of this study were: 1) to assess exposure times and sodium hypochlorite doses aimed at disinfestation and seed germination; 2) analyze the fungicide efficiency in aseptic; 3) compare different concentrations of MS medium salts in the early development in vitro; 4) verify the effect of different concentrations of BAP (6-benzylaminopurine) in shoot induction; 5) test different concentrations of indolbutyric acid (IBA) and substrates in the ex vitro rooting. Five experiments were carried out comprising the steps of micropropagation: aseptically establishment, multiplication and acclimatization. The experiments for the in vitro cultivation were conducted in Tissue Culture Laboratory of the Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) of EMPAER-MT. Acclimatization (ex vitro) was held at the nursery UNIVAG – Centro Universitário, both located in the municipality of Várzea Grande - MT. It was first defined by aseptic test exposure times (10, 15 and 20 minutes) and sodium hypochlorite concentrations (0,0; 0,5; 1,0; 1,5 and 2,0%) in three M. urundeuva All. (M1, M2 and M3). Subsequently they were assessed eight treatments of chemical disinfection through contact fungicide in the seed. The establishment was verified the proper concentration of MS medium [Murashige e Skoog, 1962] (0, 25, 50, 75 and 100%) in vitro germination. Regarding the multiplication were evaluated concentrations of BAP (0,0; 3,0; 6,0; 9,0 and 12,0 ml L-1) in two matrices of M. urundeuva All. (M2 and M3). In the last step, the acclimatization and rooting ex vitro were tested with shoots of M3 matrix in two substrates and concentrations of IBA (0,0; 0,5; 1,0; 1,5 and 2,0 mg L-1) . In aseptically dose of 1,5% sodium hypochlorite in contact for 15 minutes and 0.5ml of the fungicide Vitavax-thiram® (1% w / v carboxin + 1% w / v drawing) showed efficient disinfestation in matrices of M. urundeuva All. tested. The establishment, 25% of the concentration of MS medium salts favored the low contamination, higher germination and higher shoot length in seedlings. The concentration of 3,0 mg L-1 BAP presented a satisfactory effect on the development of shoots for the in vitro multiplication. In order to accomplish the high mortality rate (97,9%) can be associated to drastic changes in environmental conditions experienced by plantlets, suggesting the development of more efficient protocols at this stage.
Keywords: micropropagation, aseptically establishment acclimatization.

13
1 INTRODUÇÃO
A aroeira-do-sertão (Myracrodruon urundeuva Allemao [ex- Astronium
juglandifolium Griseb., Astronium urundeuva (Fr. All.) Engl.], é uma Anacardiaceae
não endêmica do Brasil (Silva-Luz e Pirani, 2014) de ocorrência natural desde o
Ceará até a Argentina e o Paraguai, sendo encontrada em formações vegetais de
caatinga, cerrado e floresta pluvial (Andrade, 2000).
Destaca-se por ser praticamente imputrescível, com múltiplos usos, que vão
desde a indústria madeireira e construção civil (Paes et al., 2009); o terapêutico cujo
extrato apresentou potencial atividade antimicrobiana, antifúngica (Alves et al.,
2009), antiulcerogênico (Carlini et al. 2010), antioxidante (Araújo Sá, 2008); inclusive
com estudos que evidenciaram potente atividade larvicida contra o Aedes aegypti
(Souza et al., 2012).
O desenvolvimento de espécies nativas ocorre principalmente por via seminal,
que interfere diretamente na longevidade e no vigor das plantas, tanto do ponto de
vista vegetativo como produtivo (Gentil e Minami, 2005). Um grande desafio do setor
florestal é justamente atender aos anseios do mercado e ainda, manter áreas de
conservação com qualidade para as florestas nativas. Portanto, é necessária a
adaptação de outras técnicas de produção de mudas que possibilitem avanços no
mercado comercial para essas espécies.
A propagação assexuada é uma alternativa para a rápida multiplicação de
mudas em larga escala, com produtividade homogênea, precocidade e tolerância às

14
pragas e doenças, produzidas a partir de plantas matrizes selecionadas (Lira Júnior
et al., 2007).
As etapas que compõem a micropropagação apresentam dificuldades que
vão desde a seleção da planta matriz até a aclimatação das mudas produzidas in
vitro. Na escolha das árvores matrizes, além de se considerar as várias
características (bom crescimento, porte, forma do tronco, sanidade, vigor, etc) das
mesmas, é necessário estar atento para a finalidade do plantio. Em espécies
destinadas ao reflorestamento, as sementes devem ter a maior variabilidade
possível, ou seja, deve ser evitada a coleta de sementes de árvores muito próximas
(Dantas et al., 2012). A coleta do explante exige cuidado para não ocorrer mistura de
material entre as matrizes diferentes e ainda para reduzir o contato com agentes
patogênicos encontrados no ambiente natural.
O nível de contaminação tende a ser maior quando as plantas matrizes são
provenientes do campo e a assepsia visa eliminar os agentes contaminantes do
material que será introduzido em laboratório. Assim, compostos químicos
(detergente, hipoclorito de sódio, fungicidas, bactericidas etc.) são utilizados sendo
necessário o ajuste da concentração e tempo de exposição para evitar injúrias no
tecido vegetal.
A fonte de nutrição do explante está no meio de cultura no qual ocorre a
inoculação e as exigências nutricionais variam entre espécies e até dentro da
mesma espécie, sendo indispensável o ajuste na concentração de sais
(macronutrientes, micronutrientes e vitaminas). O suprimento inadequado, carência
ou excesso de determinado componente pode influenciar negativamente no
desenvolvimento do tecido vegetal.
Sucessivas repicagens proporcionam o aumento exponencial no número de
explantes micropropagados e para isso o ajuste na disponibilidade de reguladores
de crescimento no meio nutritivo se mostra fundamental. As citocininas que
favorecem a citocinese são então utilizadas para estimular o aumento no número de
brotações. As condições de cultivo (temperatura, luminosidade, tipo de frasco etc.) e
a sinergia entre os hormônios endógenos e as substâncias exógenas podem afetar o
metabolismo modificando as respostas ou inativando a ação dos fitorreguladores.
A eliminação da etapa de enraizamento in vitro consiste numa estratégia para
redução de custos na produção, além do estímulo ao desenvolvimento do sistema

15
radicular mais completo e funcional. No transplantio de mudas enraizadas podem
ainda ocorrer danos nas raízes dificultando na absorção dos nutrientes do substrato.
Uma das etapas mais críticas é a aclimatação, pois os explantes são
transferidos de uma condição ambiental controlada e com alta disponibilidade de
nutrientes para o ambiente natural ficando suscetível a diversos estresses. Deste
modo é essencial o cuidado com o ambiente principalmente em relação a
luminosidade, temperatura, irrigação e manutenção da umidade. Além disso, é
preciso definir um substrato uniforme, com boa capacidade de retenção de água,
aerado, livre de patógenos e economicamente viável, que permita a iniciação e a
formação do sistema radicular.
Para estimular o desenvolvimento de raízes que são indispensáveis para a
sobrevivência da planta pode ser feita ainda a aplicação de auxina exógena,
especialmente em espécies lenhosas que apresentam dificuldade de enraizamento.
Com a aplicação da auxina sintética, ocorre um aumento imediato no nível
endógeno e consequentemente dando início da formação de raízes primordiais.
Diversos obstáculos são observados no processo de produção de mudas pela
micropropagação, porém observa-se que existem protocolos bem estabelecidos
para espécies exóticas e do mesmo modo os avanços devem ser aplicados
contemplando as espécies nativas. Desta maneira, essa pesquisa teve como
objetivo a propagação in vitro de aroeira-do-sertão (Myracrodruon urundeuva
Allemão) a partir da germinação sementes.
1.1 Referências Bibliográficas ALVES, P.M.; QUEIROZ, L.M.G.; PEREIRA, J.V.; PEREIRA, M.S.V. Atividade antimicrobiana, antiaderente e antifúngica in vitro de plantas medicinais brasileiras sobre microrganismos do biofilme dental e cepas do gênero Candida. Revista da Sociedade Brasileira de Medicina Tropical, Uberada, v.42, n.2, p.222-224, 2009. ANDRADE, M.W.; LUZ, J.M.Q.; LACERDA, A.S.; MELO, P.R.A. Micropropagação da aroeira (Myracrodruon urundeuva Fr. All.). Ciência e Agrotecnologia, Lavras, v.24, n.1, p.174-180, 2000. ARAÚJO SÁ, R. Constituintes químicos da madeira-de-lei Myracrodruon urundeuva com propriedades antioxidantes e ação contra fungos, bactérias e insetos. 2008. 171 f. Tese (Doutorado em Química) – Universidade Federal de Pernambuco, Recife –PE, 2008.

16
CARLINI, E.A.; DUARTE-ALMEIDA, J.M.; RODRIGUES, E.; TABACH, R. Antiulcer effect of the pepper trees Schinus terebinthifolius Raddi (aroeira-da-praia) and Myracrodruon urundeuva Allemão, Anacardiaceae (aroeira-do-sertão). Revista Brasileira de Farmacognosia, Curitiba, v.20, n.2, p.140-146, 2010. DANTAS, B.F.; SILVA, F.F.S.; RIBEIRO-REIS, R.C. Coleta de Sementes Florestais na Caatinga. Instruções Técnicas da Embrapa Semiárido 104. Petrolina, 2012. GENTIL, D.F.O.; MINAMI, K. Uvaieira, pitangueira e jabuticabeira: cultivo e utilização. Piracicaba: FEALQ, 2005. GRATTAPAGLIA, D.; MACHADO, M.A. Micropropagação. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa – SPI / Embrapa – CNPH, 1998, v.1, p.183-260. LIRA JÚNIOR, J. S.; BESERRA, J. E. F.; LEDERMAN, I. E.; SILVA JÚNIOR, J. F. Pitangueira. Recife: Empresa Pernambucana de Pesquisa Agropecuária – IPA, 2007. 87 p. PAES, J.B.; MORAIS, V.M.; LIMA, C.R.; SANTO, G.J.C. Resistência natural de nove madeiras do semiárido brasileiro a fungos xilófagos em simulares de campo. Revista Árvore, Viçosa, v.33, n.3, p.511-520, 2009. SILVA-LUZ, C.L.; PIRANI, J.R. Anacardiaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. 2014. Disponível em: <http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB4394>. Acesso em: 01 Jan. 2016. SOUZA, T.M.; CUNHA, A.P.; FARIAS, D.F.; MACHADO, L.K.; MORAIS, S.M.; RICARDO, N.M.; CARVALHO, A.F. Insecticidal activity against Aedes aegypti of m-pentadecadienyl-phenol isolated from Myracrodruon urundeuva seeds. Pest Management Science, v.68, p.1380–1384, 2012.

17
2 REVISÃO DE LITERATURA
2.1 A espécie Myracrodruon urundeuva Allemão
A aroeira (Myracrodruon urundeuva Allemão) é uma espécie nativa da família
Anacardiaceae, não endêmica do Brasil, com distribuição geográfica nas regiões
Norte (Tocantins), Nordeste (Alagoas, Bahia, Ceará, Maranhão, Paraíba,
Pernambuco, Piauí, Rio Grande do Norte, Sergipe), Centro-oeste (Distrito Federal,
Goiás, Mato Grosso do Sul, Mato Grosso), Sudeste (Minas Gerais, São Paulo) e Sul
(Paraná, Rio Grande do Sul, Santa Catarina), sendo encontrada em formações
vegetais de domínio fitogeográfico da Caatinga, Cerrado e Mata Atlântica (Silva-Luz
e Pirani, 2014).
O porte varia de acordo com a região onde é encontrada, chegando a cerca
de 25 m de altura e com um tronco que pode atingir 1 m de diâmetro (Andrade,
2000). Os baixos índices de umidade estimulam a abcisão foliar, coincidindo com a
época de floração, geralmente, entre julho e setembro, ocorrendo a maturação dos
frutos de setembro a outubro (Nunes et al., 2008). Clima propício à dispersão de
seus diásporos anemocóricos e antes da estação chuvosa, facilitando uma posterior
germinação das sementes e crescimento das plântulas no início da estação úmida
(Nunes et al., 2005).
As sementes da aroeira-do-sertão (Figura 1) não apresentam dormência física
(Dorneles et al., 2005; Nunes et al., 2008) e a germinação é determinada
principalmente pela absorção de água, ressaltando-se que a velocidade desse
processo depende da permeabilidade do tegumento, temperatura e composição
química da semente (Souza-Silva et al., 2001).

18
FIGURA 1. A. Diásporos anemocóricos. B. Sementes sem a estrutura alada. (Fonte: Fernandes, 2015)
Sua madeira é pardo-avermelhada e destaca-se por ser praticamente
imputrescível quando em contato com a água, possui substâncias capazes de
resistir ao ataque de fungos e cupins. Sendo empregada na fabricação de postes,
dormente para ferrovia, armação de pontes, mourões, estacas de cercas e moendas
de engenhos (Paes et al., 2009).
São atribuídas ainda atividades medicinais a essa espécie, da casca interna
de M. urundeuva isolou-se as chalconas diméricas: urundeuvina A, B, C e matosinha
(Bandeira et al., 2003), cujo extrato apresentou potencial atividade antimicrobiana no
biofilme dental e antifúngica sobre cepas de Candida isoladas da cavidade oral
(Alves et al., 2009); frações de chalconas isoladas do tronco e da casca apresentam
ação neuroprotetora (Nobre Junior et al., 2009); efeito antiulcerogênico e protetor da
mucosa gástrica (Carlini et al. 2010); além da detecção de compostos com atividade
antioxidante (Araújo Sá, 2008), entre outros. Estudos apontaram ainda que os
compostos lectina (MuHL) isolada do cerne (Araújo Sá, 2008) e m-pentadecadienyl-
fenol isolado a partir de sementes podem ter potente atividade larvicida contra o
Aedes aegypti (Souza et al., 2012).
Deve-se salientar a exploração predatória a qual a M. urundeuva e outras
espécies florestais nativas vêm sendo submetidas sem nenhuma reposição, as
populações naturais vêm diminuindo drasticamente nas últimas décadas (Kiill e
Lima, 2011). Sendo, portanto, considerada ameaçada de extinção com alto risco de
desaparecimento de acordo com Instrução Normativa publicada em setembro de

19
2008 (Ministério do Meio Ambiente, 2008). Neste sentido é importante buscar
métodos de propagação que contribuam com a perpetuação da espécie.
Assim, a biotecnologia tem sido um instrumento de resgate e preservação da
biodiversidade da floresta tropical (Daquinta et al., 2001) e a micropropagação tem
sido aplicada com sucesso para a conservação de um grande número de espécies
de plantas ameaçadas, que têm limitada capacidade reprodutiva e/ou existem em
habitats ameaçados.
2.2 Propagação
O período reprodutivo é uma fase fundamental para o entendimento da
dinâmica populacional, sendo a identificação de padrões fenológicos de floração e
frutificação importante para planos de manejo e conservação de espécies vegetais
(Cesário e Gaglione, 2008). O desenvolvimento de espécies nativas ocorre
principalmente por via seminal, que interfere diretamente na longevidade e no vigor
das plantas, tanto do ponto de vista vegetativo como produtivo (Gentil e Minami,
2005).
De acordo com descrição morfológica e anatômica da semente madura de M.
urundeuva feita por Carmello-Guerreiro e Paoli (1999), destacam-se: possui uma
semente por fruto; o endosperma é ausente (exalbuminosa); composta por dois
cotilédones compostos por reserva proteica principalmente; tegumento delgado com
duas colorações (uma clara-tegumentar e outra escura-paquicalazal). Embora
possua estruturas comuns aos representantes da família Anacardiaceae que
apresentam comportamento recalcitrante, a aroeira-do-sertão é comprovadamente
ortodoxa segundo Medeiros e Cavallari (1992), e também com estudos relacionados
ao armazenamento (Medeiros et al., 2000; Teófilo et al., 2004; Souza et al., 2007;
Caldeira e Perez, 2008)
Pesquisas referentes ao comportamento fisiolóco revelaram que apresenta
uma germinabilidade muito variada, com médias abaixo de 70% frequentes
(Medeiros et al., 2000, Silva et al., 2002, Teófilo et al. 2004; Dorneles et al., 2005;
Caldeira e Perez, 2008), contudo registrando em alguns estudos valores entre 80%
a 90% (Teófilo et al., 2004; Pacheco et al., 2006; Nunes et al., 2008). O potencial
germinativo máximo das sementes ocorre na faixa de temperatura entre 20 e
30°C (Silva et al., 2002; Pacheco et al., 2006; Guedes et al., 2011; Virgens et al.,

20
2012). O tempo médio de germinação registrado para os diásporos da espécie está
entre dois e sete dias, sendo a maior velocidade registrada (3,5 e 2,0) em maiores
temperaturas (30 e 35°C, respectivamente) (Silva et al. 2002).
Com relação ao regime hídrico, Figuerôa et al. (2004) perceberam que o
déficit hídrico (25% cc) reduziu a altura das plantas, área foliar e massa seca, porém
com maior alocação de biomassa para as raízes. Mostrando-se sensível a variação
da quantidade de água no solo, porém mesmo em situação de estresse apresenta
potencial hídrico menos negativo em comparação a outras espécies, devido a
adaptações morfo-fisiológicas (Trovão et al., 2004).
Sobre a influência da luz na germinação, Silva et al. (2002) observaram maior
germinação (acima de 60%) na ausência de luz, porém tanto na presença como na
ausência houve germinação, sendo portanto considerada fotoblástica negativa
preferencial. Resultado semelhante obtido por Miranda et al. (2007) ao analisar
sementes coletadas em dois fragmentos de Floresta Estacional Decidual em Minas
Gerais, verificaram germinação elevada na presença e na ausência de luz (médias
de 77 e 74%). Fato este comum em espécies secundárias e clímax, que possuem
capacidade de germinar a sombra do dossel, sem luz solar direta (Kageyama e
Viana, 1991). Complementando, ao verificar aspectos no desenvolvimento inicial de
plântulas de aroeira, Scalon et al. (2012) analisaram mudas com 200 dias de idade
constatando maior crescimento e desenvolvimento a pleno sol.
No que se refere ao tipo de substrato, Pacheco et al. (2006) verificaram que
as temperaturas de 25 e 27ºC combinado com os substratos vermiculita e pó de
coco permitiram bom desempenho germinativo (cerca de 80%), não sendo
necessária a reposição diária de água durante a condução dos testes. Deve-se
evitar o reumedecimento dos substratos após a semeadura, uma vez que pode
causar variações adicionais aos resultados (Brasil, 2009). Conforme Scalon et al.
(2012) a semeadura no Plantmax® proporcionou 34,77% de germinação para as
sementes de aroeira, sendo maior em comparação a terra+areia nas proporções 1:1
e 1:2 (14,04 e 17,73%). Ainda segundo estes autores, resultado favorecido
provavelmente pelo fato do Plantmax® possuir maior quantidade de matéria
orgânica, disponibilidade de água e de oxigênio necessária para a realização da
respiração nesta fase.

21
Ao avaliar a interação entre adubos (Osmocote®, esterco bovino, composto
orgânico e lodo de esgoto seco) e diferentes composições (25; 50 e 75%), Kratka e
Correia (2015) constataram morte com nos tratamentos com uso de Osmocote® e de
lodo de esgoto, provavelmente causada pela dose utilizada que acarretou em
sintomas evidentes de fitotoxicidade (clorose e necrose foliar), mesmo sendo
recomendada para espécies nativas de acordo com o fabricante. Os tratamentos a
base de esterco bovino e composto orgânico (25%) apresentaram resultados
superiores em diâmetro do coleto (3,80 e 3,02 mm respectivamente); número de
folhas (15,40 e 14,33); matéria fresca da parte aérea (1,37 e 0,95) e apesar da
análise química apontar diferença da quantidade de matéria orgânica entre eles
(70,8 e 27,4 g/dm³), ainda assim foram estatisticamente iguais na maioria das
variáveis analisadas.
A amplitude de resultados encontrados dos diversos estudos sobre
germinação de M. urundeuva pode estar relacionada à diferença no vigor dos
diásporos, que provavelmente é decorrente da característica genética das matrizes
selecionadas, condições edafo-climáticas onde se encontra estabelecida, além de
especificidades entre os laboratórios e técnicas utilizadas (Dorneles et al., 2005).
Destacaram ainda que a quantidade final de diásporos germinados tem sido
informação prioritária, pois do ponto de vista prático, esta informação é importante
na produção de mudas em larga escala.
Assim, a propagação assexuada é uma alternativa para a rápida multiplicação
de mudas em larga escala, com produtividade homogênea, precocidade e tolerância
às pragas e doenças, além da antecipação do início da produção comercial com
redução da fase juvenil da planta, produzidas a partir de plantas matrizes
selecionadas (Lira Júnior et al., 2007). Conforme a Lei nº 10.711, de 5 de agosto de
2003, do Ministério da Agricultura, Pecuária e Abastecimento, (BRASIL, 2003),
plantas matrizes são conceituadas como plantas fornecedoras de material de
propagação que mantêm as características da planta básica da qual seja
proveniente.
A micropropagação é um método de propagação vegetativa amplamente
estudada em diversas espécies vegetais, sendo a modalidade dentro da cultura de
tecidos que mais se tem difundido e encontrado aplicações práticas comprovadas
(Carvalho et al., 2006). Pode ser definida como a cultura de células, tecidos e órgãos

22
vegetais em condições assépticas (Grattapaglia e Machado, 1998; Serafini et al.
2001), por períodos indefinidos num meio de cultura apropriado (Andrade, 2002).
Os explantes provenientes de sementes e partes juvenis de plantas adultas
são preferencialmente mais utilizados, pois tecidos meristemáticos se desenvolvem
mais rapidamente e são mais estáveis geneticamente (Souza et al., 2006). Em
espécies florestais a variabilidade dos propágulos, de origem seminal na maioria dos
trabalhos, constitui em uma barreira ao cultivo in vitro, exigindo ajustes específicos
nas etapas da micropropagação dos genótipos (Oliveira et al., 2013).
2.3 Fatores que influenciam na micropropagação
Vários elementos influenciam no comportamento da cultura in vitro, tais como,
fatores biológicos (tipo e condição fisiológica do explante, características genéticas),
químicos (composição do meio de cultura, pH, reguladores de crescimento) e físicos
(temperatura, luminosidade, fotoperíodo), entre outros (Andrade, 1998). Conforme
Oliveira et al. (2013), após o estabelecimento in vitro, os explantes são multiplicados,
alongados, enraizados in vitro ou ex vitro, e por último, aclimatizados em ambiente
ex vitro.
A variabilidade genética entre espécies e dentro da mesma espécie
pressupõe o ajuste da metodologia para cada material genético, o que torna o
processo demorado, e em certas ocasiões, oneroso (Wendling et al., 2006)
2.3.1 Desinfestação do material vegetal
A primeira etapa da micropropagação é o estabelecimento in vitro do material
a ser multiplicado e, para tanto, deve-se determinar a metodologia mais adequada
para desinfestação dos explantes a serem inoculados (Ferreira et al., 2009). Em
consequência da contaminação que muitos trabalhos de cultura de tecidos são
iniciados partindo de plântulas assépticas de origem seminal (Andrade et al., 2000).
Porém para que a plântula formada a partir da germinação in vitro possa ser
fonte de explante, os métodos de desinfestação devem ser eficazes, proporcionando
total ausência de agentes patológicos (Nascimento et al., 2007). Em sementes de
espécies florestais tais como aroeira-vermelha (Schinus terebinthifolius) (Strapasson
et al., 2002), ipê-amarelo (Tabebuia serratifolia) e ipê-roxo (T. impetiginosa) (Botelho
et al., 2008), mangueira (Mangifera indica) (Andrade et al., 2008), , paineira (Ceiba

23
speciosa) (Lazarotto et al., 2010), dentre os gêneros de fungos mais comumente
relatados estão: Aspergillus, Curvularia, Fusarium, Mucor, Penicillium , Pestalotia
sp., Rhizopus e Trichoderma (Santos et al., 2000).
A alta incidência de contaminação fúngica e/ou bacteriana, proveniente do
explante ou do ambiente é uma das grandes limitações da técnica, pois podem
trazer simultaneamente danos ou até mesmo causar a morte dos tecidos vegetais.
Os microrganismos contaminantes competem pelos nutrientes do meio de cultura e
provocam danos diretos e indiretos pela colonização de seus tecidos, podendo
eliminar no meio, metabólitos tóxicos às plantas (Montarroyos, 2000).
Na assepsia, os produtos químicos mais utilizados são os compostos à base
de cloro, como o hipoclorito de sódio (NaHClO) em várias concentrações de cloro
ativo (1 a 2,5%) obtido com o uso de alvejante comercial de fácil acesso
(Grattapaglia e Machado, 1998; Camolesi et al., 2007; Pereira et al., 2011). Ao
estudar a regeneração in vitro de mogno (Swietenia macrophylla King) a partir de
partes de plântulas obtidas da germinação de sementes, Couto et al., (2004)
observaram resultados superiores com a embebição das sementes em 2,5 e 5% de
hipoclorito de sódio por 30 e 20 minutos, a maior germinação (48%) e baixa
contaminação (15 e 10%, respectivamente).
Para desinfestação de sementes de louro pardo (Cordia trichotoma (Vell.)
Arráb. Ex Steud.), Fick et al. (2007) testaram a imersão em hipoclorito de sódio
(NaOCl) (2% e 5%) por 0, 5, 10, 15 ou 20 minutos, porém não encontraram
diferença para nenhum dos fatores na desinfestação e germinação, com médias que
variaram de 90 a 100% e 85 a 100% respectivamente.
Em sementes de guabijuzeiro (Myrcianthes pungens (Berg) Legr.), Souza et
al. (2011) fizeram a desinfestação com imersão em soluções com diferentes
concentrações de NaOCl (0, 2, 4, 6 e 8%). Verificaram 100% de contaminação na
dose 0% de hipoclorito e nas demais houve baixa contaminação (2 a 14%) sem
diferença estatística entre si. A germinação não foi afetada, apresentando valores de
98, 92 e 91% nos tratamentos 4, 6 e 8% respectivamente.
2.3.2 Estabelecimento in vitro a partir da germinação de sementes
Após a descontaminação, o isolamento é sempre realizado em câmara de
fluxo laminar exigindo perícia e cuidado com o explante. Uma rápida excisão,

24
inoculação no meio de cultura e imediata vedação do frasco são fundamentais para
evitar a desidratação (Ferreira e Pasqual, 2008), podendo determinar a
sobrevivência do material. O meio de cultura é composto por uma mistura
balanceada de macronutrientes, micronutrientes, fontes orgânicas de nitrogênio,
vitaminas e hormônios vegetais (Dorris, 2010).
O meio MS formulado por Murashige e Skoog em 1962, foi desenvolvido
inicialmente para o cultivo de tecido medular de Nicotiana tabacum (Caldas et al.,
1990) e suas modificações e diluições, tem sido utilizadas com sucesso para
diversas espécies (Grattapaglia e Machado, 1998). Todavia adequações nas
concentrações são feitas e se baseiam nas exigências nutricionais de cada espécie,
pois controlam o padrão de desenvolvimento in vitro (Caldas et al., 1990; Santos-
Serejo et al., 2006). Meios de cultura não adequados podem causar sintomas de
deficiência mineral e até a morte dos propágulos (Monteiro et al., 2000).
Na germinação de sementes de erva-cidreira (Melissa officinalis – Lamiaceae)
feita por Reis et al. (2008) foram analisadas a concentração de sais (25, 50 e 100%)
do meio MS (Murashige e Skoog, 1962) e observaram no meio 25% a média de
64,75% de germinação. Em cerejeira (Amburana acreana - Fabaceae), maiores
percentuais de germinação (86,7%) ocorreram nos meios de cultura com redução de
50% nas concentrações salinas do meio MS (Fermino Junior e Scherwinski-Pereira
2012). Em pesquisa desenvolvida por Schwalbert et al. (2014) com plântulas de
pega-pega (Desmodium incanum – Fabaceae), o meio MS 50% foi superior nas
variáveis: altura das brotações (média 1,1 cm) e número de folhas (média 6,8),
situação que pode estar ligada às exigências nutricionais e condição natural de
desenvolvimento da espécie.
A maior concentração de sais no meio de cultura interferiu no potencial
osmótico e, consequentemente, na disponibilidade de água para o processo de
embebição da semente na germinação.
2.3.3 Multiplicação de brotações
Assim que estão estabelecidos in vitro, os explantes passam para a etapa de
multiplicação, que é a fase mais longa da micropropagação e visa a produzir o maior
número possível de plantas uniformes e de alta qualidade (Grattapaglia e Machado,
1998). A combinação entre citocininas e auxinas promove a diferenciação celular e

25
também a organogênese, esta necessidade se refere ao tipo, concentração,
genótipo das plantas matrizes e o conteúdo endógeno de hormônios (Lima et al.,
2008). A definição do tipo e da concentração ótima de citocinina para a multiplicação
in vitro constitui um passo importante e dependerá do genótipo da planta; o seu uso
estimula uma maior produção de brotos e alta taxa de multiplicação (Lemos et al.,
2001).
Para indução de brotações de tomate-de-capucho (Physalis peruviana –
Solanaceae), Chaves et al. (2005) avaliaram os meios de cultura MS e MS¾
(reduzido em 25% dos sais) e concentrações de BAP (0,0; 0,1; 0,2 ou 0,3 mg L-1).
Verificaram que o número de brotações utilizando meio de cultura MS¾ apresentou
comportamento linear crescente nas concentrações de BAP utilizadas, sendo que o
número médio de brotações passou de 1 brotação por explante no tratamento
testemunha, para 1,75 brotações na concentração de 0,3 mg L -1 de BAP. Com
relação ao comprimento da parte aérea, o ponto de máxima foi atingido sem adição
de BAP (0,0 mg L-1) com 6,06 cm e a menor média de 1,73 cm na concentração 0,3
mg L-1. Para esta espécie, observaram que concentrações elevadas de BAP
aumentaram a taxa de proliferação das brotações, no entanto, o tamanho tende a
ser pequeno.
A multiplicação in vitro de erva-cidreira (Melissa officinalis – Fabaceae) foi
avaliada por Reis et al. (2008) em meio MS acrescido ou não com a citocinina BAP
(6-benzilaminopurina) na concentração de 4,44 µmol/L (aproximadamente 0,49 mg
L-1). No primeiro subcultivo não houve diferença no número de brotos no meio MS e
MS+BAP com médias de 1,91 e 1,98 respectivamente. No segundo e terceiro
subcultivos o meio MS+BAP se sobressaiu com médias de 3,25 e 3,7, enquanto o
meio MS sem BAP apresentou 1,59 e 2. Porém para o número de entrenós
(importante na multiplicação dos propágulos) o MS sem BAP obteve média superior
de 3,8, bem como para o comprimento das brotações (4,16 cm), com relação aos
três subcultivos. De acordo com Grattapaglia e Machado (1998) a concentração da
citocinina é um fator que influencia o resultado visto que o excesso pode ser tóxico e
comprometer o desenvolvimento das culturas.
Em eucalipto (Eucalyptus benthamii x Eucalyptus dunnii), Brondani et al.
(2009) utilizaram doses de BAP (0; 0,25; 0,50; 0,75; e 1,0 mg L-1) na multiplicação de
segmentos nodais provenientes de minicepas previamente estabelecidas in vitro.

26
Aos 30 dias, uma produção de 6,9 gemas por explante foi observado na
concentração de BAP em torno de 0,31 mg L-1 (de acordo com o ajuste da equação
de regressão). Comportamento semelhante aos 60 dias, com elevado acréscimo na
taxa de multiplicação, com estimativa de 20,2 gemas por explante no ponto de
máxima com BAP de 0,30 mg L-1. De acordo com os autores, baseado nas
equações ajustadas, pode-se inferir que a faixa situada entre 0,25 e 0,50 mg L-1 de
BAP resultou nos melhores valores quanto ao número de gemas emitidas do
material híbrido avaliado. Importante ressaltar que concentrações de BAP acima de
0,75 mg L-1 não resultaram em efeitos positivos quanto à proliferação de gemas, e
na concentração de 1 mg L-1 , além de sintomas de hiperhidricidade houve ainda
oxidação dos explantes.
À medida que as pesquisas são desenvolvidas percebe-se que as respostas
das plantas às técnicas de micropropagação são variáveis, dependem da espécie,
variedade e/ou cultivar, época de coleta, tipo de explante utilizado e condições de
cultivo (Wendling et al., 2006).
2.3.4 Aclimatização e enraizamento ex vitro
A fase de aclimatização das plantas cultivadas in vitro é apontada como uma
das mais críticas do processo de regeneração, em que pode ocorrer a maioria das
perdas (Rocha et al., 2008). Os explantes são expostos à redução de umidade do ar
e temperatura instável, a fim de que possam suportar a transferência para o novo
substrato e, posteriormente, sobreviver e se desenvolver sem complicações em
condições naturais de campo (Silva et al., 1995).
Um dos fatores importantes na produção de mudas, além do controle de
umidade e condições de cultivo, é o tipo de substrato (Paiva Sobrinho et al., 2010),
pois as propriedades do substrato podem facilitar ou impedir o desenvolvimento das
plantas (Calvete et al., 2000; Rocha et al., 2008). Conforme Fernandes et al. (2006)
vários materiais podem ser utilizados como substratos para plantas, dentre eles a
vermiculita, turfa, casca de arroz, casca de pinus, fibra da casca de coco, entre
outros. A vermiculita é um mineral inerte, de baixa densidade e estrutura variável,
constituído de lâminas ou camadas justapostas em tetraedros de sílica e octaedros
de ferro (Fe) e magnésio (Mg) (Gomes e Paiva 2006; Caldeira et al., 2013).

27
A turfa do gênero Sphagnum tem como principais características a leveza, pH
ácido, alta capacidade de retenção de água, baixa drenagem, fácil manuseio e ser
estéril (Ristow et al, 2012). A casca de arroz possui boa aeração permitindo a troca
de ar na base das raízes, grande capacidade de drenagem, tem forma floculada,
leve, de fácil manuseio, tem volume constante seja seca ou úmida, é livre de
patógenos e não necessita de tratamento químico para esterilização, devido ao
processo de carbonização (Souza, 1993).
A casca de pinus constitui um dos resíduos da atividade madeireira, é
utilizado na cobertura do solo, pois apresenta lenta degradação e altos índices de
cálcio e magnésio; este subproduto também é utilizado na produção de substrato
comercial (Nóbrega et al., 2007). O aproveitamento da casca de coco é viável por
serem suas fibras quase inertes e terem alta porosidade, apresentando ótima
aeração, aliada a uma boa capacidade de retenção de água (Wendling e Gatto,
2002). A facilidade de produção, baixo custo e alta disponibilidade são outras
vantagens adicionais apresentadas por este tipo de substrato (Carrijo et al., 2002).
O enraizamento ex vitro diretamente no substrato apresenta como vantagens
a diminuição das dificuldades associadas à sobrevivência na aclimatização (Augusto
et al., 2006). É desejável do ponto de vista econômico (Yan et al., 2010),
possibilitando uma redução nos custos de obtenção da muda micropropagada
(Pelizza et al., 2013). A indução das raízes pode ser obtida por reguladores de
crescimento análogos de auxina, no entanto o modo de ação e o transporte de cada
molécula diferem dentro dos tecidos da planta (Vieten et al., 2007).
Ao analisar as pesquisas em diferentes espécies observou-se que Rocha et
al. (2008) testaram AIB (0; 0,25; 1,0; 2,0 e 3,0 mg L-1) em dois genótipos de
jenipapeiro (Genipa americana) e pela regressão observaram que a concentração de
1,1 mg L-1 foi mais eficiente com percentual acima de 70%, para o genótipo JRB69, e
43,3% para o genótipo JRB59. Em pesquisa com teca (Tectona grandis), Fermino
Junior et al. (2011) verificaram o efeito de concentrações de AIB (0, 100, 1000, 2000,
4000 mg L-1) e obtiveram enraizamento em todos os tratamentos, inclusive sem
adição da auxina, com valores de 100% a partir de 1000 mg L-1. A mesma tendência
foi observada por Fernandes et al. (2015) ao testarem as concentrações de AIB (0,0;
0,5; 1,0; 1,5 e 2,0 mg L-1) no enraizamento ex vitro de teca; e Xavier et al. (2003) ao
aplicar doses de AIB entre 0 (zero) e 4000 mg L-1 em miniestacas de cedro-rosa

28
(Cedrela fissilis). Utilizando AIA (1000, 2000, 3000 e 4000 mg L-1) em pistóporo-
japonês (Pittosporum tobira), Meneguzzi et al. (2015) obtiveram maior porcentagem
de enraizamento (60%) no tratamento com 2.000 mg L-1.
Vários fatores podem influenciar o enraizamento tanto os intrínsecos,
relacionados à própria planta, como os extrínsecos, relacionados as condições
ambientais (Norberto et al., 2001). É notório que a concentração depende da
espécie, condições de cultivo e que principalmente, deve-se considerar o balanço
hormonal para obter resultados satisfatórios.
2.4 Referências Bibliográficas
ALBUQUERQUE, R.J.M.; RODRIGUES, L.V.; VIANA, G.S.B. Análise clínica e morfológica da conjuntivite alérgica induzida por ovalbumina e tratada com chalcona em cobaias. Acta Cirúrgica Brasileira, São Paulo, v.19, n.1, p.43-68, 2004. ALVES, P.M.; QUEIROZ, L.M.G.; PEREIRA, J.V.; PEREIRA, M.S.V. Atividade antimicrobiana, antiaderente e antifúngica in vitro de plantas medicinais brasileiras sobre microrganismos do biofilme dental e cepas do gênero Candida. Revista da Sociedade Brasileira de Medicina Tropical, Uberada, v.42, n.2, p.222-224, 2009. ANDRADE, L.B. Efeito do meio de cultura, tipos de explante e períodos de escuro sobre a micropropagação da batata (Solanum tuberosum L.), cv. Cristal.1998. 60 f. Dissertação (Mestrado em Agronomia) - Universidade Federal de Pelotas, Pelotas, 1998. ANDRADE, M.W.; LUZ, J.M.Q.; LACERDA, A.S.; MELO, P.R.A. Micropropagação da aroeira (Myracrodruon urundeuva Fr. All.). Ciência e Agrotecnologia, Lavras, v.24, n.1, p.174-180, 2000. ANDRADE, S.R.M. Princípios da cultura de tecidos vegetais. Embrapa Cerrados, Planaltina, 2002, 16p. ANDRADE, S.R.M.; OLIVEIRA, W.C.; REIS JUNIOR, F.B. et al. Controle do crescimento e identificação de microrganismos contaminantes visando à micropropagação de gemas laterais de mangueira. Embrapa Cerrados, Planaltina, 27 p., 2008. (Boletim de pesquisa e desenvolvimento 217 / Embrapa Cerrados). ARAÚJO SÁ, R. Constituintes químicos da madeira-de-lei Myracrodruon urundeuva com propriedades antioxidantes e ação contra fungos, bactérias e insetos. 2008. 171 f. Tese (Doutorado em Química) – Universidade Federal de Pernambuco, Recife –PE, 2008.

29
AUGUSTO, C.S.S.; BIASI, L.A.; TELLES, C.A. Enraizamento e aclimatização de plantas micropropagadas de amoreira-preta cv. Brazos. Revista Brasileira de Fruticultura, v.28, n.3, p.473-476, 2006. BANDEIRA, M.A.M., MATOS, F.J.A., BRAZ-FILHO, R. Structural elucidation and total assignment of the H and C NMR spectra of new chalcone dimers. Magnetic Resonance in Chemistry, v.41, p.1009-1014, 2003. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Lei nº 10.711, de 5 de agosto de 2003. Dispõe sobre o Sistema Nacional de Sementes e Mudas e dá outras providências. [Diário Oficial da República Federativa do Brasil], Brasília, DF, 2003. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa Agropecuária. Brasília, DF: Mapa/ACS, 2009. 395p. BRONDANI, G.E.; DUTRA, L.F.; GROSSI, F.; WEDLING, I.; HORNING, J. Estabelecimento, multiplicação e alongamento in vitro de Eucalyptus benthamii Maiden & Cambage x Eucalyptus dunnii Maiden. Revista Árvore, Viçosa, v.33, n.1, p.11-19, 2009. CALDAS, L.S.; HARIDASAN, P.; FERREIRA, M.E. Meios nutritivos. In: TORRES, A. C., CALDAS, L.S. Técnicas e aplicações da cultura de tecidos de plantas. Brasília: ABCTP, EMBRAPA-CNPH, 1990. p.37-70. CALDEIRA, M.V.; DELARMELINA, W.M.; PERONI, L.; GONÇALVES, E.O.; SILVA, A.G. Lodo de esgoto e vermiculita na produção de mudas de eucalipto. Pesquisa Agropecuária Tropical, Goiânia, v. 43, n. 2, p. 155-163, 2013. CALDEIRA, S.F.; PEREZ, S.C.J.G.A. Qualidade de diásporos de Myracrodruon urundeuva Fr. All. armazenados sob diferentes condições. Revista Brasileira de Sementes, Londrina, v.30, n.3, p.185-194, 2008. CALVETE, E.O.; KÄMPF, A.N.; DAUDT, R. Efeito do substrato na aclimatização ex vitro de morangueiro cv Campinas, Fragaria x ananassa Duch. In: KÄMPF, A.N.; FERMINO, M.H. (Ed.). Substrato para plantas: a base da produção vegetal em recipientes. Porto Alegre: Genesis, 2000. p.257-264. CAMOLESI, M.R.; KAIHARA, E.S.; SACONI, C.G.; FARIA, R.T.; NEVES, C.S.V.J. Redução da oxidação na propagação in vitro da bananeira maçã. Ciência Agrotécnica, Lavras, v. 31, n. 4, p. 1237-1241, 2007. CARLINI, E.A.; DUARTE-ALMEIDA, J.M.; RODRIGUES, E.; TABACH, R. Antiulcer effect of the pepper trees Schinus terebinthifolius Raddi (aroeira-da-praia) and Myracrodruon urundeuva Allemão, Anacardiaceae (aroeira-do-sertão). Revista Brasileira de Farmacognosia, Curitiba, v.20, n.2, p.140-146, 2010. CARMELLO-GUERREIRO, S.M.; PAOLI, A.A.S. Aspectos morfológicos e anatômicos da semente de aroeira (Myracrodruon urundeuva Fr.Allem. –

30
Anacardiaceae), com notas sobre paquicalaza. Revista Brasileira de Sementes, Londrina, v.21, n.1, p.222-228, 1999. CARRIJO, O.A.; LIZ, R.S.; MAKISHIMA, N. Fibra da casca do coco verde como substrato agrícola. Horticultura Brasileira, Brasília, v.20, n.4, p.533-540, 2002. CARVALHO, J.M.F.C.; SILVA, M.M.A.; MEDEIROS, M.J.L. Fatores inerentes à micropropagação. Campina Grande: Embrapa Algodão, 2006. 28p. (Embrapa Algodão. Documentos, 148). CESÁRIO, L.F.; GAGLIANONE, M.C. Biologia floral e fenologia reprodutiva de Schinus terebinthifolius Raddi (Anacardiaceae) em Restinga do Norte Fluminense. Acta Botânica Brasilica, Feira de Santana, v.22, n.3, p.828-833, 2008. CHAVES, A.C.; SCHUCH, M.W.; ERIG, A.C. Estabelecimento e multiplicação in vitro de Physalis peruviana L. Ciência e Agrotecnologia, Lavras, v.29, n.6, p.1281-1287, 2005. COUTO, J.M.F.; OTONI, W.C.; PINHEIRO, A.L.; FONSECA, E.P. Desinfestação e germinação in vitro de sementes de mogno (Swietenia macrophylla King). Revista Árvore, Viçosa, v.28, n.5, p.633-642, 2004. DAQUINTA, M.; RAMOS, L; CAPOTE, I. et al. Micropropagación de la teca (Tectona grandis L.f.). Revista Florestal Centroamericana, Comunicación técnica, n.35, p. 25-28, 2001. DORNELES, M.C.; RANAL, M.A.; SANTANA, D.G. Germinação de diásporos recém-colhidos de Myracrodruon urundeuva Allemão (Anacardiaceae) ocorrente no cerrado do Brasil Central. Brazilian Journal of Botany, São Paulo, v.28, n.2, p.399-408, 2005. DORRIS, M. What is plant tissue culture? Journal of the Bromeliad Society, v.60, n.4, p.172-178, 2010. FERMINO JUNIOR, P.C.P.; RAPOSO, A.; SCHERWINSKI-PEREIRA, J.E. Enraizamento ex vitro e aclimatização de plantas micropropagadas de Tectona grandis. Floresta, Curitiba, v.41, n.1, p.79-86, 2011. FERMINO JUNIOR, P.C.P.; SCHERWINSKI-PEREIRA, J.E. Germinação e propagação in vitro de cerejeira (Amburana acreana (Ducke) A.C. Smith - Fabaceae). Ciência Florestal, Santa Maria, v.22, n.1, p. 1-9, 2012. FERNANDES, C.; CORÁ, J.E.; BRAZ, L.T. Alterações nas propriedades físicas de substratos para cultivo de tomate cereja, em função de sua reutilização. Horticultura Brasileira, Campinas, v. 24, n. 1, p. 94-98, 2006. FERNANDES, D. A.; MARTINEZ, D.T.; COSTA, R. B. Sacarose e ácido indolbutírico no enraizamento de Tectona grandis L.f. Revista de Agricultura, Piracicaba, v. 90, n.1, p. 87-99, 2015.

31
FERREIRA, E.A.; PASQUAL, M. Otimização de protocolo para micropropagação da figueira “Roxo de Valinhos”. Ciência Rural, Santa Maria, v.38, n.4, p.1149-1153, 2008. FERREIRA, M.G.R.; SANTOS, M.R.A.; BRAGADO, A.C.R. Propagação in vitro d cupuazeiro: desinfestação de explantes florais. Saber Científico, Porto Velho, v.2, n.2, p.37-44, 2009. FICK, T.A.; BISOGNIN, D.A.; QUADROS, K.M.; HORBACH, M.; REINIGER, L.R.S. Estabelecimento e crescimento in vitro de plântulas de louro pardo. Ciência Florestal, Santa Maria, v.17, n.4, p.343-349, 2007. FIGUEIRÔA, J.M.; BARBOSA, D.C.A.; SIMABUKURO, E.A. Crescimento de plantas jovens de Myracrodruon urundeuva Allemão (Anacardiaceae) sob diferentes regimes hídricos. Acta Botânica Brasílica, São Paulo, v.18, n.3, p.573-580, 2004. GENTIL, D.F.O.; MINAMI, K. Uvaieira, pitangueira e jabuticabeira: cultivo e utilização. Piracicaba: FEALQ, 2005. GOMES, J.M.; PAIVA, H.N. Viveiros florestais (propagação assexuada). 3. ed. Viçosa: UFV, 2006. GUEDES, R.S.; ALVES, E.U.; GONÇALVES, E.P.; COLARES, P.N.Q.; MEDEIROS, M.S.; VIANA, J.S. Germinação e vigor de sementes de Myracrodruon urundeuva Allemão em diferentes substratos e temperaturas. Revista Árvore, Viçosa, v.35, n.5, p.975-982, 2011. GRATTAPAGLIA, D.; MACHADO, M.A. Micropropagação. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa – SPI / Embrapa – CNPH, 1998, v.1, p.183-260. KAGEYAMA, P.Y.; VIANA,V.M. Tecnologia de sementes e grupos ecológicos de espécies arbóreas tropicais. In: SIMPÓSIO BRASILEIRO SOBRE TECNOLOGIA DE SEMENTES FLORESTAIS, 2., 1989, Atibaia. Anais... São Paulo: Instituto Florestal, 1991. p.197-215. KIILL, L.H.P.; LIMA, P.C.F. Plano de manejo para espécies da Caatinga ameaçadas de extinção na Reserva Legal do Projeto Salitre. Petrolina: Embrapa Semiárido, 2011, 55p. (Embrapa Semiárido. Documentos, 243). KRATKA, P.C.; CORREIA, C.R.M.A. Crescimento inicial de aroeira do sertão (Myracrodruon urundeuva allemão) em diferentes substratos. Revista Árvore, Viçosa, v.39, n.3, p.551-559, 2015. LAZAROTTO, M.; MUNIZ, M.F.B.; SANTOS, A.F. Detecção, transmissão, patogenicidade e controle químico de fungos em sementes de paineira (Ceiba speciosa). Summa Phytopathologica, Botucatu, v.36, n.2, p.134-139, 2010. LEMOS, E.E.P.; FERREIRA, M.S.; ALENCAR. L.M.C.; OLIVEIRA, J.G.L.; MAGALHÃES, V.S. Micropropagação de clones de banana cv. terra em biorreator de

32
imersão temporária. Revista Brasileira de Fruticultura, Jaboticabal, v.23, n.3, p.482-487, 2001. LIMA, E.C.; PAIVA, R.; NOGUEIRA, R.C. Callus induction in leaf segments of Croton urucurana Baill. Ciência e Agrotecnologia, Lavras, v.32, n.1, p.17-22, 2008. LIRA JÚNIOR, J. S.; BESERRA, J. E. F.; LEDERMAN, I. E.; SILVA JÚNIOR, J. F. Pitangueira. Recife: Empresa Pernambucana de Pesquisa Agropecuária – IPA, 2007. 87 p. LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Nova Odessa: Ed. Plantarium, 1992. 352p. MEDEIROS, A.C.S.; CAVALLARI, D.A.N. Conservação d germoplasma de aroeira (Astronium urundeuva (Fr. All.) Engl. I. Germinação de sementes após imersão em nitrogênio líquido (-196ºC). Revista Brasileira de Sementes, v.14, n.1, p.73-75, 1992. MEDEIROS, A.C.S.; SMITH, R.; PROBERT, R.; SADER, R. Comportamento fisiológico de sementes de aroeira (Myracrodruon urundeuva Fr. All.) em condições de armazenamento. Boletim de Pesquisa Florestal, Colombo, n. 40, p. 85-98, 2000. MENEGUZZI, A.; NAVROSKI, M.C.; LOVATEL, Q.C.; MARCO, F.T.; PEREIRA, M.O.; TONETT, E.L. Ácido indolacético influencia no enraizamento de estacas de Pittosporum tobira. Revista de Ciências Agroveterinárias, Lages, v.14, n.1, p.24-28, 2015. MINISTÉRIO DO MEIO AMBIENTE. Lista oficial das espécies brasileiras ameaçadas de extinção. 2008. Disponível em: http://www.mma.gov.br/estruturas/ascom_boletins/_arquivos/83_19092008034949.pdf MIRANDA, W.O.; BRANDÃO,D.O ; NUNES, Y.R.F.; VELOSO, M.D.M.; ARRUDA, D. M. Influência da luz na germinação de sementes de aroeira (Myracrodruon urundeuva) ocorrentes em dois fragmentos de Floresta Estacional Decidual no norte de Minas Gerais. In: VIII Congresso de Ecologia do Brasil, 2007, Caxambú. Anais... VIII Congresso de Ecologia do Brasil, 2007. MONTARROYOS, A.V.V. Contaminação in vitro. ABCTP Notícias, Brasília, n.36 e 37, p.5-10, 2000. MONTEIRO, A.C.B.A.; HIGASHI, E.N.; GONÇALVES, A.N.; RODRIGUEZ, A.P.M.A novel approach for the definition of the inorganic medium components for micropropagation of yellow passion fruit (Passiflora edulis Sims f. flavicarpa Deg.). In vitro Cellular e Developmental Biology-Plant, Columbia, v.36, n.6, p.527-531, 2000.

33
NASCIMENTO, P.K.V.; FRANCO, E.T.H; FRASSETTO, E.G. Desinfestação e germinação in vitro de sementes de Parapiptadenia rígida Bentham (Brenam). Revista Brasileira de Biociências, Porto Alegre, v. 5, supl. 2, p. 141-143, 2007. NOBRE-JR, H.V., OLIVEIRA, R.A., MAIA, F.D., NOGUEIRA, M.A.S., DE MORAES, M.O., BANDEIRA, M.A.M., ANDRADE, G.M., VIANA, G.S.B. Neuroprotective effects of chalcones from Myracrodruon urundeuva on 6-Hydroxydopamineinduced cytotoxicity in rat mesencephalic cells. Neurochemical Research, v.34, p.1066-1075, 2009. NÓBREGA, R.S.A.; BOAS, R.C.V.; NÓBREGA, J.C.A.; PAULA, A.M.; MOREIRA, F.M.S. Utilização de biossólido no crescimento inicial de mudas de aroeira (Schinus terebinthifolius Raddi). Revista Árvore, Viçosa, v.31, n.2, p. 239-246, 2007. NORBERTO, P.M.; CHALFUN, N.N.J.; PASQUAL, M. et al. Efeito da época de estaquia e do AIB no enraizamento de estacas de figueira (Ficus carica L.). Ciência e Agrotecnologia, Lavras, v.25, n.3, p.533-541, 2001. NUNES, Y.R.F.; FAGUNDES, M.; SANTOS, R.M. et al. Atividades fenológicas em Guazuma ulmifolia Lam. (Malvaceae) em uma floresta estacional decidual no norte de Minas Gerais. Lundiana, Belo Horizonte, v.6, n.2, p.99-105, 2005. NUNES, Y.R.F.; FAGUNDES, M.; ALMEIDA, H.S. et al. Aspectos ecológicos da aroeira (Myracrodruon urundeuva Allemão – Anacardiaceae): fenologia e germinação de sementes. Revista Árvore, Viçosa, v.32, n.2, p.233-243, 2008. OLIVEIRA, L.S.; DIAS, P.C.; BRONDANI, G.E. Micropropagação de espécies florestais brasileiras. Pesquisa Florestal Brasileira, Colombo, v.33, n.76, p.439-453, 2013. PACHECO, M.V.; MATOS, V.P.; FERREIRA, R.L.C.; FELICIANO, A.L.P.; PINTO, K.M.S. Efeito de temperaturas e substratos na germinação de sementes de Myracrodruon urundeuva Fr. All. (Anacardiaceae). Revista Árvore, Viçosa, v.30, n.3, p.359-367, 2006. PAIVA SOBRINHO, S.; LUZ, P.B.; SILVEIRA, T.L.S.; RAMOS, D.T.; NEVES, L.G.; BARELLI, M.A.A. Substratos na produção de mudas de três espécies arbóreas do cerrado. Revista Brasileira de Ciências Agrárias, Recife, v.5, n.2, p.238-243, 2010. PAES, J.B.; MORAIS, V.M.; LIMA, C.R.; SANTO, G.J.C. Resistência natural de nove madeiras do semiárido brasileiro a fungos xilófagos em simulares de campo. Revista Árvore, Viçosa, v.33, n.3, p.511-520, 2009. PEREIRA, G.A.; CORREA, L.S.; BOLIANI, A.C. Desinfestação e estabelecimento in vitro de explantes de bananeira „Grande naine‟ em diferentes concentrações de hipoclorito de sódio. Revista Brasileira de Fruticultura, Jaboticabal, volume especial, p.222-226, 2011.

34
REIS, E.S.; PINTO, J.E.B.P.; ROSADO, L.D.S.; CORRÊA, R.M. Influência do meio de cultura na germinação de sementes in vitro e taxa de multiplicação de Melissa officinalis L. Revista Ceres, Viçosa, v.55, n.3, p.160-167, 2008. RISTOW, N.C.; ANTUNES, L.E.C; CARPENEDO, S. Substratos para o enraizamento de microestacas de mirtileiro cultivar Georgiagem. Revista Brasileira de Fruticultura, Jaboticabal - SP, v. 34, n. 1, p. 262-268, 2012. ROCHA, M.A.C.; COSTA, M.A.P.C.; SILVA, S.A.; LEDO, C.A.S.; MOREIRA, M.J.S.; BASTOS, L.P. Enraizamento in vitro e aclimatização de genótipos de jenipapeiro (Genipa americana L.). Revista Brasileira de Fruticultura, Jaboticabal - SP, v. 30, n. 3, p. 769-774, 2008. SANTOS, A.F.; GRIGOLETTI JUNIOR, A.; AUER, S.G. Transmissão de fungos por sementes de espécies florestais. Revista Floresta, Curitiba, v.30, n.12, p.119-128, 2000. SANTOS-SEREJO, J.A.; JUNGHANS, T.G.; SOARES, T.L.; SILVA, K.M. Meios nutritivos para micropropagação de plantas. In: SOUSA, A.S.; JUNGHANS, T.G. Introdução à micropropagação de plantas, Cruz das Almas: Embrapa Mandioca e Fruticultura Tropical, 2006. p.79-98. SCALON, S.P.Q.; SCALON FILHO, H.; MASETTO, T.E. Aspectos da germinação e desenvolvimento inicial de plântulas de aroeira. Cerne, Lavras, v.18, n.4, p.533-539, 2012. SCHWALBERT, R.; MALDANER, J.; AITA, M.F.; AMARAL, G.A.; TAROUCO, A.K. Concentrações de sais do meio MS no cultivo in vitro de Desmodium incanum. Enciclopédia Biosfera, Goiânia, v.10, n.18, p.1009-1015, 2014. SERAFINI, L.A.; BARROS, N.M.; AZEVEDO, J.L. Biotecnologia: princípios e aplicações. Guaíba: Agropecuária, p. 25–74, 2001. SILVA, A.T.; PASQUAL, M.; ISHIDA, J.S.; ANTUNES, L.E.C. Aclimatação de plantas provenientes da cultura in vitro. Pesquisa Agropecuária Brasileira, Brasília, v.30, n.1, p.49-53, 1995. SILVA, L.M.M.; RODRIGUES, T.J.D.; AGUIAR, I.B. Efeito da luz e da temperatura na germinação de sementes de aroeira (Myracroduon urundeuva Allemão). Revista Árvore, Viçosa, v.26, p.691-697, 2002. SILVA-LUZ, C.L.; PIRANI, J.R. Anacardiaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. 2014. Disponível em: <http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB4394>. Acesso em: 01 Jan. 2016. SOUZA, F.V.D.; JUNGHANS, T.G.; SOUZA, A.S.; SANTOS-SEREJO, J.A.; COSTA, M.A.P.C. Micropropagação. In: SOUZA, A.S.; JUNGHANS, T.G. Introdução à micropropagação de plantas. Cruz das Almas, BA: Embrapa Mandioca e Fruticultura Tropical, 2006. p. 38-52.

35
SOUZA, F.X. Casca de arroz carbonizada: um substrato para a propagação de plantas. Lavoura Arrozeira, Porto Alegre, v.46, n.406, p. 11, 1993. SOUZA, L.S.; FIOR, C.S.; SOUZA, P.V.D.; SCHWARZ, S.F. Desinfestação de sementes e multiplicação in vitro de guabijuzeiro a partir de segmentos apicais juvenis (Myrcianthes pungens O.Berg) D. Legrand. Revista Brasileira de Fruticultura, Jaboticabal, v.33, n.3, p.691-697, 2011. SOUZA, S.C.A.; BORGES, G.R.A.; BRANDÃO, D.O.; MATOS, A.M.M.; VELOSO, M.D.M.; NUNES, Y.R.F. Conservação de Sementes de Myracodruon urundeuva Freire Allemão (Anacardiaceae) em Diferentes Condições de Armazenamento. Revista Brasileira de Biociências, Porto Alegre, v.5, supl.2, p.1140-1142, 2007. SOUZA-SILVA, M.; FONTENELLE, J.C.R.; MARTINS, R.P. Seasonal abundance and species composition of flower-visiting flies. Neotropical Entomology, v.30, n.3, p.351-359, 2001. SOUZA, T.M.; CUNHA, A.P.; FARIAS, D.F.; MACHADO, L.K.; MORAIS, S.M.; RICARDO, N.M.; CARVALHO, A.F. Insecticidal activity against Aedes aegypti of m-pentadecadienyl-phenol isolated from Myracrodruon urundeuva seeds. Pest Management Science, v.68, p.1380–1384, 2012. STRAPASSON, M.; SANTOS, A.F.; MEDEIROS, A.C.S. Fungos associados às sementes de aroeira-vermelha (Schinus terebinthifolius). Boletim Pesquisa Florestal, Colombo, n.45, 131-135, 2002. TEÓFILO, E.M.; SILVA, S.O.; BEZERRA, A.M.E.; MEDEIROS FILHO, S.; SILVA, F.D.B. Qualidade fisiológica de sementes de aroeira (Myracrodruon urundeuva Allemão) em função do tipo de embalagem, ambiente e tempo de armazenamento. Revista Ciência Agronômica, Ceará, v.35, n.2 p.371-376, 2004. TROVÃO, D.M.B.M.; FERNANDEZ, P.D.; ANDRADE, L.A.; NETO, J.D.; QUEIROZ, J.A. Avaliação do potencial hídrico de espécies de Caatinga sob diferentes níveis de umidade no solo. Revista de Biologia e Ciências da Terra, v.4, n.2, 2004. VIETEN, A.; SAUER, M.; BREWER, P.B.; FRIML, J. Molecular and cellular aspects of auxin-transport-mediated development. Trends in Plant Science, v.12, p.160–168, 2007. VIRGENS, I.O.; CASTRO, R.D.; FERNANDEZ, L.G.; PELACANI, C.R. Comportamento fisiológico de sementes de Myracrodruon urundeuva Fr. All. (Anacardiaceae) submetidas a fatores abióticos. Ciência Florestal, Santa Maria, v.22, n.4, p.681-692, 2012. WENDLING, I.; DUTRA, L.F.; GROSSI, F. Produção de mudas de espécies lenhosas. Colombo: Embrapa Florestas. 2006. 54 p. (Embrapa Florestas. Documentos, 130).

36
WENDLING, I.; GATTO, A. Substratos, adubação e irrigação na produção de mudas. Viçosa: Aprenda Fácil, 2002. XAVIER, A.; SANTOS, G.A.; WENDLING, I.; OLIVEIRA, M.L. Propagação vegetativa de Cedro-rosa por miniestaquia. Revista Árvore, Viçosa, v.27, n.2, p.139-143, 2003.

37
3 HIPOCLORITO DE SÓDIO NA DESINFESTAÇÃO E GERMINAÇÃO in vitro DE
SEMENTES DE Myracrodruon urundeuva Allemão (AROEIRA-DO-SERTÃO)
RESUMO – A desinfestação dos explantes para o estabelecimento in vitro é fundamental para evitar o desenvolvimento de microrganismos, visto que a contaminação é um fator limitante. Objetivou-se avaliar tempos de exposição e doses de hipoclorito de sódio na desinfestação e germinação de sementes de matrizes de aroeira-do-sertão (Myracrodruon urundeuva All.). O experimento foi conduzido no Laboratório de Cultura de Tecidos do Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da EMPAER-MT, em Várzea Grande – MT de maio a junho de 2015. O delineamento foi inteiramente casualizado em esquema fatorial 3x5, considerando-se três tempos de contato (10, 15 e 20 minutos) e cinco concentrações de hipoclorito (0,0; 0,5; 1,0; 1,5 e 2,0%), constituindo 15 tratamentos com três repetições em três matrizes (M1, M2 e M3). Primeiramente foi realizada assepsia das sementes com Tween 20® (3,0 ml L-1) e álcool 70% (v/v), posteriormente em hipoclorito de sódio nos tempos e concentrações e para finalizar foram misturadas por fricção em 0,5 ml de fungicida (1% m/v carboxina + 1% m/v tiram). Foram então inoculadas em meio de cultura MS e levadas para sala de crescimento. Após 20 dias avaliou-se a contaminação por bactérias (%) e a germinação (%). As matrizes M2 e M3 apresentaram 0% de contaminação nos tratamentos 1,0 e 2,0% por 10 e 20 minutos e 1,5% por 10 minutos, respectivamente. Na M1 a germinação máxima de 45% foi obtida com 2,0% de hipoclorito. A utilização de 1,5% de hipoclorito por 15 minutos é eficiente na assepsia e germinação de sementes de M. urundeuvaI All.
Palavras-chave: assepsia, cloro ativo, bactericida.

38
SODIUM HYPOCHLORITE IN disinfection and germination in vitro OF SEEDS
Myracrodruon urundeuva Allemão
ABSTRACT - The disinfection of the explants for in vitro establishment is essential to prevent the growth of microorganisms since the infection is a limiting factor. This study aimed to evaluate exposure times and sodium hypochlorite disinfection doses and germination in matrices of Myracrodruon urundeuva All. The experiment was conducted at Tissue Culture Laboratory of the Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) of EMPAER-MT in Várzea Grande – MT, to May-June 2015. The design was completely randomized in a factorial 3x5, considering If three contact times (10, 15 and 20 minutes) and five hypochlorite concentrations (0,0; 0,5; 1,0; 1,5 and 2,0%), constituting 15 treatments with three replicates in three matrices (M1, M2 and M3). First carried out a sterilization of seeds with Tween 20® (3,0 ml L-1) and ethanol 70% (v/v), then sodium hypochlorite concentrations in time and to finish and were mixed by friction in 0,5 ml fungicidal (1% m/v carboxin + 1% m/v thiram). They were then inoculated in MS medium and taken to a growth room. After 20 days we were evaluated contamination by bacteria (%) and germination (%). The matrices M2 and M3 showed 0% contamination in treatments 1,0 and 2,0% for 10 and 20 minutes and 1,5% for 10 minutes, respectively. In M1 maximum germination of 45% was obtained with 2,0% hypochlorite. The use of 1,5% hypochlorite for 15 minutes is effective in sterilization and germination of M. urundeuvaI All. seeds.
Keywords: aseptic, active chlorine, bactericidal.

39
3.1 Introdução
Uma das grandes limitações para a micropropagação de várias espécies é a
alta incidência de contaminação fúngica e/ou bacteriana, proveniente do explante ou
do meio ambiente (Carneiro et al., 2000). Depende ainda da procedência do material
vegetal utilizado, que pode ser da casa de vegetação ou do campo, o nível de
contaminação tende a ser maior quando as plantas matrizes são provenientes do
campo (Ferreira et al., 2009). Esses microrganismos competem pelos nutrientes do
meio de cultura e provocam danos diretos e indiretos pela colonização de seus
tecidos, podendo eliminar no meio, metabólitos tóxicos às plantas (Montarroyos,
2000).
Conforme Camara et al. (2010), a principal meta da micropropagação é
produzir plantas em condições adequadas e com qualidade fitossanitária, uma vez
que microrganismos silenciosos podem se manifestar em estágios tardios do cultivo
in vitro. As espécies lenhosas apresentam dificuldades para o estabelecimento in
vitro, pois podem apresentar elevado grau de contaminação e presença de
microrganismos associados aos explantes (Couto et al., 2004; Golle et al., 2013). As
bactérias constituem o mais comum e problemático tipo de infecção em cultura de
tecidos, pois podem ser sistêmicas, dificultando assim sua detecção (Costa et al.,
2007).
A frequência da descontaminação não é total, pois os tecidos vegetais podem
interferir de alguma forma no controle, sejam destoxificando essas substâncias, ou
servindo como habitat para os microrganismos que se translocam para os tecidos
(Scherwinski-Pereira e Costa, 2010). O controle da assepsia deve ser feito desde a
esterilização dos instrumentos e recipientes utilizados na manipulação do explante,
que ocorre durante todo o processo. Os instrumentos (pinças, bisturis, placas de
petri, etc) devem ser submergidos em etanol 96%; flambados e colocados sobre um
suporte limpo, dentro da câmara de fluxo laminar (Carvalho, 2006).
A desinfestação do material vegetal apresenta-se como uma importante etapa
no processo de estabelecimento da espécie em laboratório. É iniciada com o uso de
substâncias que possuem ação germicida, dentre as quais, compostos à base de
cloro, como: o hipoclorito de sódio (NaHClO) em várias concentrações de cloro ativo
(1 a 2,5%) (Grattapaglia e Machado, 1998; Camolesi et al., 2007; Pereira et al.,
2011) e o hipoclorito de cálcio [Ca(OCl)2]. Podem ser utilizados ainda o etanol 70%

40
ou absoluto por 1 a 3 minutos, devido sua toxicidade (Gerra et al., 2001) e o
formaldeído (Pardo et al., 2006).
A concentração da solução desinfestante e o tempo de exposição podem
variar muito os índices de descontaminação, sendo necessária adequação de
acordo com a espécie (Moraes et al., 2007; Ferreira et al., 2009). Altas
concentrações e tempo elevado de imersão podem desinfestar de maneira
satisfatória, porém, a viabilidade do tecido vegetal pode reduzir, provocando perdas.
Os métodos de desinfestação devem ser eficazes, para que a plântula sirva de fonte
de explante livre de fungos e bactérias (Couto et al., 2004).
A partir do exposto, o objetivo neste trabalho foi avaliar tempos de exposição
e doses de hipoclorito de sódio na desinfestação e germinação de sementes de
matrizes de aroeira-do-sertão (Myracrodruon urundeuva All.).
3.2 Material e Métodos
O experimento foi realizado no Laboratório de Cultura de Tecidos do Centro
Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da Empresa Mato-
grossense de Pesquisa, Assistência e Extensão Rural (EMPAER-MT) localizado no
município de Várzea Grande – MT, no período de maio a junho de 2015. As
sementes utilizadas estavam armazenadas há oito meses.
O delineamento experimental foi inteiramente casualizado em esquema
fatorial 3x5, considerando-se três tempos de contato (10, 15 e 20 minutos) e cinco
concentrações de hipoclorito (0,0; 0,5; 1,0; 1,5 e 2,0%), constituindo 15 tratamentos
com três repetições de 24 sementes para cada matriz (M1, M2 e M3).
As sementes foram coletadas em uma área de reserva pertencente a uma
empresa comercial situada na Estrada da Guia, Rodovia MT 010, KM 27 - Cuiabá-
MT, em Setembro de 2014. Ocorreu precipitação na região dez dias antes da coleta,
no dia o tempo estava ensolarado com temperatura média registrada de 36°C. As
matrizes selecionadas de aroeira-do-sertão estavam a cerca de 30 metros de
distância entre si.
Para a coleta, embaixo de cada matriz selecionada foi colocada uma lona de
10x10 m2, em seguida os galhos foram agitados para que as sementes caíssem em
cima evitando assim o contato direto com o solo. Posteriormente foi feita a limpeza
separando as sementes de gravetos e demais impurezas, sendo então

41
armazenadas em câmara fria no Laboratório de Sementes da Universidade Federal
de Mato Grosso, na temperatura de 18ºC ±2 e umidade de 68%.
Os recipientes utilizados para inocular as sementes foram frascos cilíndricos
de vidro com capacidade de 250 ml, diâmetro externo de 68 mm e altura de 100 mm.
Em cada frasco foi adicionado 20 ml de meio de cultura MS, formulado por
Murashige e Skoog (1962), suplementado com 30 g L-1 de sacarose e solidificado
com 2,5 g L-1 de Phytagel® e pH ajustado em 5,8 ±0,2. Após o preparo, os frascos
foram levados para a esterilização em autoclave, em temperatura de 120°C e 1 kgf
cm-2 de pressão, por 20 minutos. Na sequência foram retirados da autoclave,
armazenados em estante na sala de repicagem para resfriar em temperatura
ambiente e solidificar o meio de cultura.
Após o preparo do meio de cultura, as sementes de cada matriz armazenadas
foram homogeneizadas e 20 g sementes foram pesadas e separadas em frascos de
vidro. Foram separados 15 frascos de vidro para cada matriz (M1, M2 e M3),
contendo as sementes que posteriormente foram utilizadas para compor as
repetições. Estas sementes foram lavadas com detergente Tween 20® (3 ml L-1)
diluído em água destilada, mexendo por três minutos e enxaguadas três vezes com
água destilada. Na sequência foram adicionados 20 ml de álcool etílico 70% (v/v) em
cada frasco com as sementes que foram agitadas por três minutos, seguido por três
enxágues em água destilada. Em seguida, as sementes foram embebidas pelos
tempos estabelecidos no experimento (10, 15 e 20 minutos), em soluções contendo
as doses de hipoclorito de sódio 0,0; 0,5; 1,0; 1,5 e 2,0% (v/v) de cloro ativo (Start®
12%), com subsequentes três enxagues em água destilada e autoclavada.
Para finalizar a assepsia foram então colocadas em sacos plásticos (15x27
cm) contendo 0,5 ml do fungicida Vitavax-thiram® (1% m/v carboxina + 1% m/v tiram)
e homogeneizadas por fricção até a cobertura total das sementes pelo produto. A
determinação da dose foi feita de acordo com a quantidade adequada para
cobertura das sementes e não foi realizado mais nenhum enxágue. Os tratamentos
com 0,0% de hipoclorito das três matrizes passaram apenas pela limpeza com
detergente Tween 20®, álcool etílico 70% (v/v) e contato com o fungicida.
Após a desinfestação as sementes foram então inoculadas nos frascos
contendo meio de cultura previamente preparado em câmara de fluxo laminar
previamente homogeneizada com álcool 70% e as pinças utilizadas para colocar as

42
sementes em contato com o meio de cultura dentro dos frascos foram esterilizadas
em autoclave. Para cada frasco inoculado, as pinças passaram por flambagem com
álcool 95% e as foram utilizadas frias para evitar injúrias ao tecido vegetal.
Para cada tratamento foram semeados três frascos (subamostras) por
repetição com oito sementes cada. No momento da inoculação, frascos contendo
meio de cultura ficaram abertos para verificar a incidência de possíveis
contaminantes dentro da câmara e/ou resultantes do manuseio do operador. Os
frascos vazios foram acondicionados juntamente com os tratamentos na sala de
crescimento e não foi observado nenhum tipo de contaminação durante e até
mesmo após a avaliação do experimento.
Os frascos foram então identificados de acordo com os tratamentos, vedados
com plástico filme para evitar possíveis contaminações, e levados para uma sala de
crescimento, onde foram dispostos aleatoriamente em uma prateleira de metal. A
temperatura média desta sala foi de 25 ±2°C e fotoperíodo de 16 horas, sendo a
intensidade luminosa de 25 mol m-2 s-1 (medida com luxímetro) fornecida por
lâmpadas do tipo fluorescente branca fria.
Aos 20 dias após a semeadura ocorreu à avaliação por se tratar do período
onde se observou a finalização da germinação e consistiu na observação e
contagem individual das sementes contaminadas por bactérias (%) e a germinação
(%). Foram consideradas germinadas às sementes que apresentaram emissão de
raiz (cerca de 2 mm). Na contaminação por bactérias a porcentagem foi obtida pela
expressão: X(%) = N*100 / 24, em que N = número de sementes contaminadas. A
germinação foi calculada a partir da expressão: G (%) = N/A * 100, em que: N =
número de sementes germinadas; A= número total de sementes colocadas para
germinar.
Os dados coletados foram submetidos à análise da variância. Para
germinação (%) foram transformados em arco seno (Santana e Ranal, 2000). As
médias dos tratamentos foram comparadas por regressão polinomial. Foi utilizado o
programa estatístico Assistat versão 7.7 Beta (Silva, 2011).

43
3.3 Resultados e Discussão
Na análise de variância para a variável contaminação por bactérias observou-
se interação significativa (p < 0,01) entre os tempos de exposição e as
concentrações de hipoclorito nas três matrizes M1, M2 e M3.
As estimativas para a contaminação por bactérias na matriz M1 apresentaram
as menores médias de contaminação nos tratamentos com 10 (2,77%), 15 (4,16%) e
20 (4,16%) minutos foram observadas nas concentrações de 1,5; 2,0 e 2,0% de
hipoclorito de sódio respectivamente (Figura 1).
FIGURA 1. Contaminação por bactérias (%) em sementes da matriz M1 de
aroeira-do-sertão (M. urundeuva All.) tratadas com concentrações de
hipoclorito de sódio em diferentes tempos de exposição (CV10=
34,64%; CV15= 37,50%; CV20= 15,21%, Cuiabá-MT, 2015).
Na matriz M2 as concentrações de 1,0 e 2,0% de hipoclorito de sódio por 10 e
20 minutos respectivamente, não apresentaram contaminação. No tempo de 15
minutos observou-se menor contaminação nos tratamentos 0,5; 1,0 e 2,0% com
4,16% em cada um (Figura 2).
y = -3,614x + 10,554 R² = 0,7042
y = -13,909x2 + 27,001x + 4,9537 R² = 0,8351
y = 7,56x2 - 23,724x + 22,368 R² = 0,7471
0
5
10
15
20
25
30
0 0,5 1 1,5 2Co
nta
min
ação
por
bacté
rias (
%)
Ma
triz
1
Concentrações de hipoclorito (%)
10 minutos 15 minutos 20 minutos
10 minutos 15 minutos 20 minutos
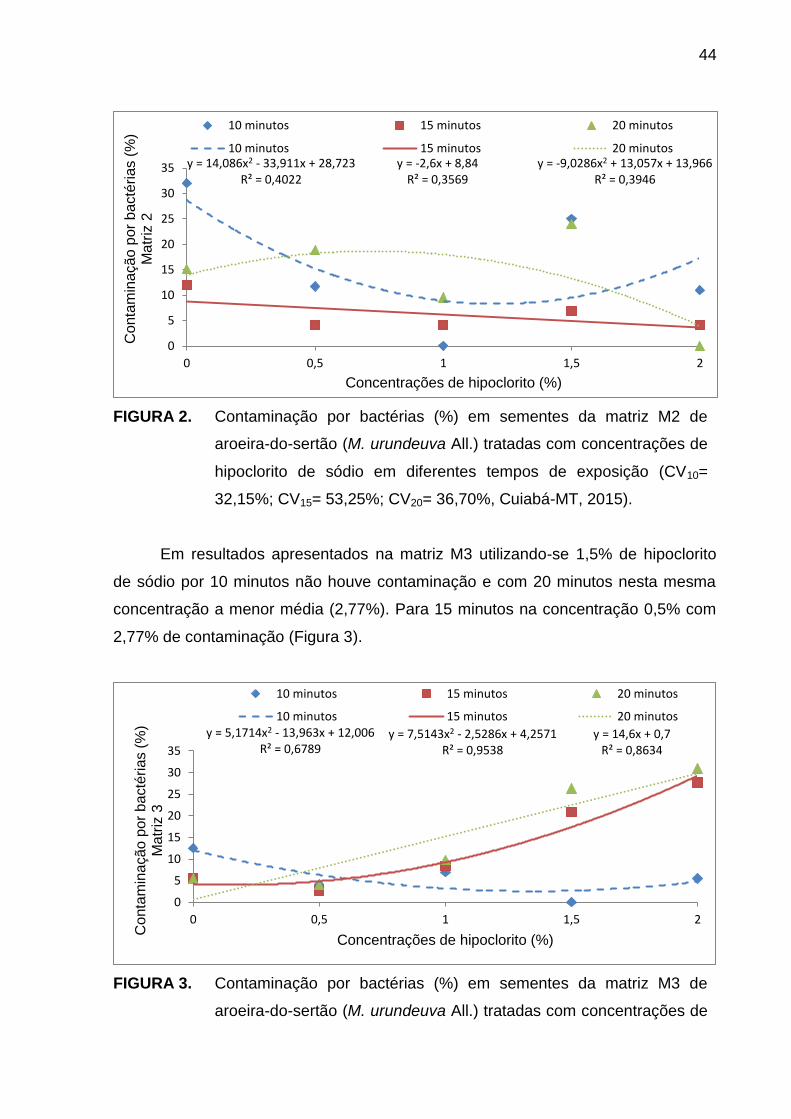
44
FIGURA 2. Contaminação por bactérias (%) em sementes da matriz M2 de
aroeira-do-sertão (M. urundeuva All.) tratadas com concentrações de
hipoclorito de sódio em diferentes tempos de exposição (CV10=
32,15%; CV15= 53,25%; CV20= 36,70%, Cuiabá-MT, 2015).
Em resultados apresentados na matriz M3 utilizando-se 1,5% de hipoclorito
de sódio por 10 minutos não houve contaminação e com 20 minutos nesta mesma
concentração a menor média (2,77%). Para 15 minutos na concentração 0,5% com
2,77% de contaminação (Figura 3).
FIGURA 3. Contaminação por bactérias (%) em sementes da matriz M3 de
aroeira-do-sertão (M. urundeuva All.) tratadas com concentrações de
y = 14,086x2 - 33,911x + 28,723 R² = 0,4022
y = -2,6x + 8,84 R² = 0,3569
y = -9,0286x2 + 13,057x + 13,966 R² = 0,3946
0
5
10
15
20
25
30
35
0 0,5 1 1,5 2
Co
nta
min
ação
por
bacté
rias (
%)
Ma
triz
2
Concentrações de hipoclorito (%)
10 minutos 15 minutos 20 minutos
10 minutos 15 minutos 20 minutos
y = 5,1714x2 - 13,963x + 12,006 R² = 0,6789
y = 7,5143x2 - 2,5286x + 4,2571 R² = 0,9538
y = 14,6x + 0,7 R² = 0,8634
0
5
10
15
20
25
30
35
0 0,5 1 1,5 2
Co
nta
min
ação
por
bacté
rias (
%)
Ma
triz
3
Concentrações de hipoclorito (%)
10 minutos 15 minutos 20 minutos
10 minutos 15 minutos 20 minutos

45
hipoclorito de sódio em diferentes tempos de exposição (CV10=
26,08%; CV15= 30,15%; CV20= 30,69%, Cuiabá-MT, 2015).
A determinação exata da fonte de contaminação muitas vezes é dificultada e
os microrganismos podem ser introduzidos em vários pontos do processo de
micropropagação (Pasqual et al., 2010). Em princípio, existem quatro fontes de
contaminação: a fonte de explante, o meio nutritivo, o ambiente e o operador (Souza
et al., 2007), assim sendo deve-se haver um controle em todos os possíveis modos
de contaminação.
A variação na porcentagem de contaminação bacteriana pode estar
associada à infinidade de microrganismos encontrados no ambiente natural que
podem infiltrar no tecido vegetal. Diferentemente dos fungos, as bactérias são
capazes de penetrar num vegetal por aberturas naturais ou causadas por ferimentos
(Scherwinski-Pereira e Alterthum, 2010). Em condições controladas de luz,
temperatura e nutrientes conforme verificado na micropropagação podem encontrar
o ambiente favorável para se desenvolver.
A persistente contaminação dos tecidos por bactérias é um problema sério e o
uso sucessivo de soluções de hipoclorito pode ser mais prejudicial ao explante do
que à bactéria (Pasqual et al., 2010). A contaminação depende da procedência do
material vegetal utilizado, e tendem a ser maior quando as plantas matrizes usadas
como fonte de explantes são provenientes do campo (Ferreira et al., 2009). Os
plantios com espécies nativas ocupam reduzidas áreas de floresta e o controle
fitossanitário da planta doadora de propágulos torna-se difícil (Oliveira et al., 2013).
A assepsia superficial visa evitar o crescimento de microrganismos, pois a
disputa por nutrientes no meio de cultura podem inviabilizar a técnica levando a
perda do material vegetal. A maior dificuldade nesta etapa está em obter o tecido
descontaminado sem conduzi-lo à morte quando isolado (Grattapaglia e Machado,
1998). As bactérias constituem um tipo de contaminação comum e problemática na
cultura de tecidos, pois podem ser sistêmicas e sua detecção muitas vezes é difícil
(Pasqual et al., 2010).
Além da substância desinfestante utilizada e a concentração adequada, o
tempo de exposição deve ser ajustado de acordo com o tipo e tamanho do explante
selecionado devido à sensibilidade do tecido. Algumas substâncias com ação

46
germicida são recomendadas para realizar a limpeza, sendo os mais comuns o
etanol e compostos à base de cloro, tais como hipoclorito de cálcio e sódio
(Grattapaglia e Machado, 1998; Pasqual et al., 2010). Pressupõe-se que há uma
combinação entre o cloro com a proteína da membrana dos microrganismos que
formam compostos tóxicos, levando à inibição de enzimas essenciais (Donini et al.,
2005).
No resumo da análise de variância das matrizes M1, M2 e M3 para a
germinação (%), observou-se interação significativa (0,01 =< p < 0,05) entre os
tratamentos (tempo e concentrações) apenas na matriz M1. Na matriz M2 não houve
diferença em nenhum dos fatores testados, sendo a média de germinação
observada foi de 50,8%. Na matriz M3 houve diferença significativa (p < 0,01)
apenas para as concentrações de hipoclorito de sódio.
No desdobramento das concentrações em cada tempo testado verificou-se
que a germinação da matriz M1 foi significativa apenas no tempo de 10 minutos para
as concentrações testadas e maior porcentagem de germinação (45,81%) na
concentração 2,0% de hipoclorito de sódio (Figura 4).
FIGURA 4. Germinação (%) em sementes da matriz M1 de aroeira-do-sertão (M.
urundeuva All.) tratadas com concentrações de hipoclorito de sódio
por 10 minutos de exposição (CV10= 14,86%, Cuiabá-MT, 2015).
Ao fazer uma relação entre as variáveis observou-se que para as sementes
da matriz M1 tratadas com as concentrações de 1,5 e 2,0% de hipoclorito nos
y = 8,752x + 25,992 R² = 0,8717
0
10
20
30
40
50
0 0,5 1 1,5 2
Ge
rmin
açã
o (
%)
Ma
triz
1
Concentrações de hipoclorito (%)
10 minutos
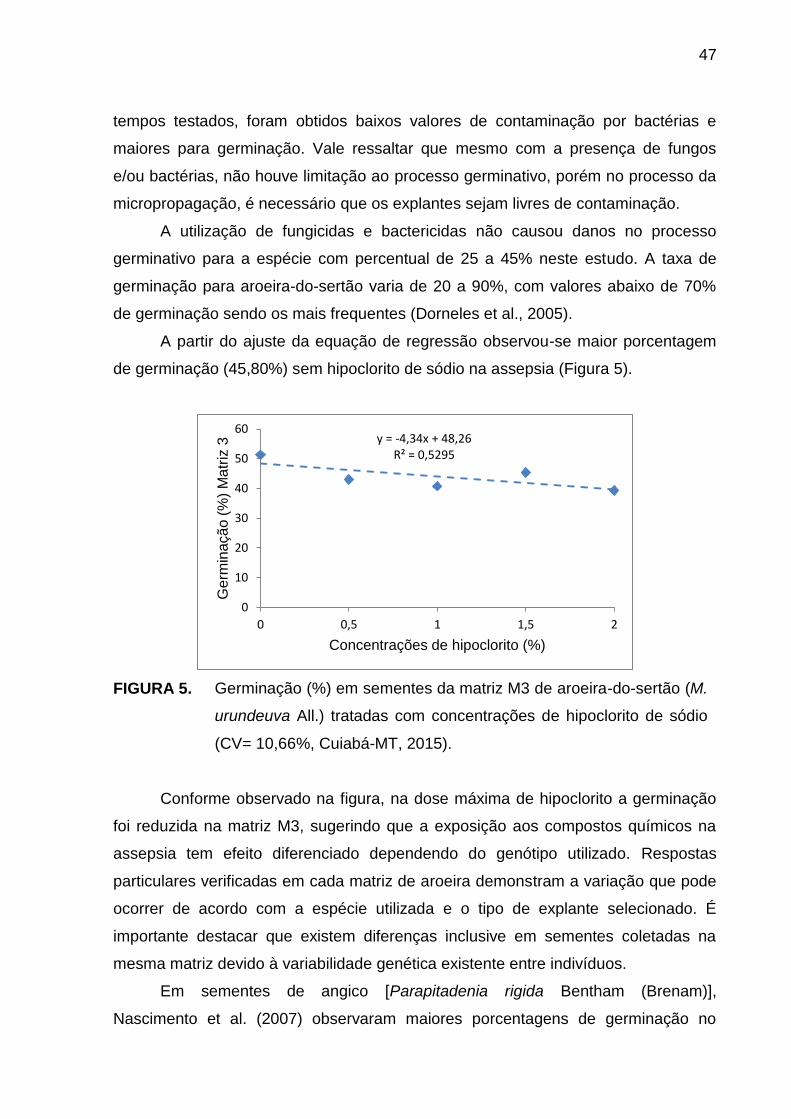
47
tempos testados, foram obtidos baixos valores de contaminação por bactérias e
maiores para germinação. Vale ressaltar que mesmo com a presença de fungos
e/ou bactérias, não houve limitação ao processo germinativo, porém no processo da
micropropagação, é necessário que os explantes sejam livres de contaminação.
A utilização de fungicidas e bactericidas não causou danos no processo
germinativo para a espécie com percentual de 25 a 45% neste estudo. A taxa de
germinação para aroeira-do-sertão varia de 20 a 90%, com valores abaixo de 70%
de germinação sendo os mais frequentes (Dorneles et al., 2005).
A partir do ajuste da equação de regressão observou-se maior porcentagem
de germinação (45,80%) sem hipoclorito de sódio na assepsia (Figura 5).
FIGURA 5. Germinação (%) em sementes da matriz M3 de aroeira-do-sertão (M.
urundeuva All.) tratadas com concentrações de hipoclorito de sódio
(CV= 10,66%, Cuiabá-MT, 2015).
Conforme observado na figura, na dose máxima de hipoclorito a germinação
foi reduzida na matriz M3, sugerindo que a exposição aos compostos químicos na
assepsia tem efeito diferenciado dependendo do genótipo utilizado. Respostas
particulares verificadas em cada matriz de aroeira demonstram a variação que pode
ocorrer de acordo com a espécie utilizada e o tipo de explante selecionado. É
importante destacar que existem diferenças inclusive em sementes coletadas na
mesma matriz devido à variabilidade genética existente entre indivíduos.
Em sementes de angico [Parapitadenia rigida Bentham (Brenam)],
Nascimento et al. (2007) observaram maiores porcentagens de germinação no
y = -4,34x + 48,26 R² = 0,5295
0
10
20
30
40
50
60
0 0,5 1 1,5 2
Ge
rmin
açã
o (
%)
Ma
triz
3
Concentrações de hipoclorito (%)

48
tratamento com 0% de hipoclorito, porém apesar deste bom resultado constataram
que este tratamento não se mostrou eficiente em relação ao parâmetro
desinfestação das sementes.
A utilização de substâncias com ação germicida tais como, etanol e
compostos a base de cloro (hipoclorito de sódio 0,5 a 2,0%), são comumente
empregadas na desinfestação superficial de explantes (Pasqual et al., 2010). Por
isso a importância em adaptar a dose adequada e tempo de exposição visando à
manutenção da viabilidade dos tecidos. A variabilidade genética entre espécies e
dentro da mesma espécie sugere o ajuste da metodologia para cada material
genético, tornando o processo demorado, e em certas ocasiões, oneroso (Wendling
et al., 2006).
3.4 Conclusão
A utilização da solução de hipoclorito de sódio a 1,5% por 15 minutos é
eficiente para a assepsia e germinação de sementes de aroeira-do-sertão (M.
rurundeuva All.).
3.5 Referências Bibliográficas
CAMARA, T.R.; WILLADINO, L.; ALBUQUERQUE, C.C. Microrganismos assintomáticos do cultivo in vitro: natureza e riscos para o cultivo de plantas. In: SCHERWINSKI-PEREIRA, J.E. (editor técnico). Contaminações microbianas na cultura de células, tecidos e órgãos de plantas. Brasília-DF: Embrapa Informação Tecnológica, 2010, p.221-260. CAMOLESI, M.R.; KAIHARA, E.S.; SACONI, C.G.; FARIA, R.T.; NEVES, C.S.V.J. Redução da oxidação na propagação in vitro da bananeira maçã. Ciência Agrotécnica, Lavras, v. 31, n. 4, p. 1237-1241, 2007. CARNEIRO, M.F.; SILVA, G.D.; XIMENES, P.A.; CARNEIRO, I.F.; BORGES, J.D. Avaliação de produtos na descontaminação de explantes de banana (Musa AAB cv. Maçã). Pesquisa Agropecuária Tropical, Goiania, v.30, n.1, p.29-35, 2000. CARVALHO, J.M.F.C.; SILVA, M.M.A.; MEDEIROS, M.J.L. Fatores inerentes à micropropagação. Campina Grande, Embrapa Algodão. Documentos, 148. 2006. 28p. COSTA, A.S.; ARRIGONI-BLANCK, M.F.; BLANCK, A.F.; MENDONÇA, A.B.; AMANCIO, V.F.; LEDO, A.S. Estabelecimento de alecrim-pimenta in vitro. Horticultura Brasileira, Brasília, v.25, n.1, p.68-72, 2007.

49
COUTO, J.M.F.; OTONI, W.C.; PINHEIRO, A.L.; FONSECA, E.P. Desinfestação e germinação in vitro de sementes de mogno (Swietenia macrophylla King). Revista Árvore, Viçosa, v.28, n.5, p.633-642, 2004. DONINI, L.P.; FERREIRA-MOURA, I.; GUISSO, A.P.; SOUZA, J.A.; VIÉGAS,J. Preparo de lâminas foliares de aráceas ornamentais: desinfestação com diferentes concentrações de hipoclorito de sódio. Arquivo do Instituto de Biologia, São Paulo, v. 72, n. 4, p. 517-522, 2005. DORNELES, M. C.; RANAL, M. A.; SANTANA, D.G. Germinação de diásporos recém-colhidos de Myracrodruon urundeuva Allemão (Anacardiaceae) ocorrente no cerrado do Brasil Central. Revista Brasileira de Botânica, São Paulo, v.28, n.2, p.399-408, 2005. FERREIRA, M.G.R.; SANTOS, M.R.A.; BRAGADO, A.C.R. Propagação in vitro de cupuazeiro: desinfestação de explantes florais. Saber Científico, Porto Velho, v.2, n.2, p.37-44, 2009. GERRA, M.P.; VESCO, L.D.; DUCROQUET, J.P.; NODARI, R.O.; REIS, M.S. Somatic embryogenesis is in goiabeira serrana: genotype response, auxinic shock and synthetic seeds. Revista Brasileira de Fisiologia Vegetal, Campinas, v.13, n.2, p.117-128, 2001. GOLLE, P.D.; REINIGER, L.R.S.; BELLÉ, R.A.; CURTI, A.R. Desinfestação superficial de explantes isolados de ramos semilenhosos e herbáceos de Eugenia involucrata DC. (Myrtaceae). Cerne, Lavras, v.19, n.1, p. 77-82, 2013. GRATTAPAGLIA, D.; MACHADO, M.A. Micropropagação. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa – SPI / Embrapa – CNPH, 1998, v.1, p.183-260. MONTARROYOS, A.V.V. Contaminação in vitro. ABCTP Notícias, Brasília, n.36 e 37, p.5-10, 2000. MORAES, A.M.; ALMEIDA; F.A.C.; CAZÉ FILHO, J. Desinfestação e estabelecimento in vitro de gemas axilares de abacaxizeiro. Tecnologia e Ciência Agropecuária, João Pessoa, v.1, n.2, p. 39-44, 2007. MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiologia Plantarum, Copenhagen, v.15, p.473-97, 1962. NASCIMENTO, P.K.V.; FRANCO, E.T.H.; FRASSETTO, E.G. Desinfestação e germinação in vitro de sementes de Parapitadenia rigida Bentham (Brenam). Revista Brasileira de Biociências, Porto Alegre, v.5, n.2, p.141-143, 2007. OLIVEIRA, L.S.; DIAS, P.C.; BRONDANI, G.E. Micropropagação de espécies florestais brasileiras. Pesquisa Florestal Brasileira, Colombo, v. 33, n. 76, p. 439-453, 2013.

50
PARDO, V.M.A.; FERREIRA, A.G.; NUNES, V.F. Seed disinfestation methods for in vitro cultivation of epiphyte orchids from southern Brazil. Horticultura Brasileira, Campinas, v.24, n.2, p.217-220, 2006. PASQUAL, M.; DUTRA, L.F.; ARAUJO, A.G.; PEREIRA, A.R. Prevenção de contaminações microbianas na cultura de células, tecidos e órgãos de plantas. In: SCHERWINSKI-PEREIRA, J.E. (editor técnico). Contaminações microbianas na cultura de células, tecidos e órgãos de plantas. Brasília-DF: Embrapa Informação Tecnológica, 2010, p.61-161. PEREIRA, G.A.; CORREA, L.S.; BOLIANI, A.C. Desinfestação e estabelecimento in vitro de explantes de bananeira „Grande naine‟ em diferentes concentrações de hipoclorito de sódio. Revista Brasileira de Fruticultura, Jaboticabal, volume especial, p.222-226, 2011. SANTANA, D.G.; RANAL, M.A. Análise estatística na germinação. Revista Brasileira de Fisiologia Vegetal, Campinas, v.12 (Edição especial), p.205-237, 2000. SCHERWINSKI-PEREIRA, J.E.; ALTERTHUM, F. Detecção, isolamento e preservação de microrganismos contaminantes da cultura de células, tecidos e órgãos de plantas. In: SCHERWINSKI-PEREIRA, J.E. (editor técnico). Contaminações microbianas na cultura de células, tecidos e órgãos de plantas. Brasília-DF: Embrapa Informação Tecnológica, 2010, p.163-185. SILVA, F.A.S. ASSISTAT versão 7.7 beta. Campina Grande-PB: Assistência Estatística, Departamento de Engenharia Agrícola do CTRN - Universidade Federal de Campina Grande, Campus de Campina, 2011. SOUZA, G.C.; CLEMENTE, P.L.; ISAAC, V.L.R.; FARIA, S.P.; CAMPOS, M.R.C. Contaminação Microbiana na Propagação in vitro de Cattleya walkeriana e Schomburgkia crispa. Revista Brasileira de Biociências, Porto Alegre, v. 5, supl. 1, p. 405-407, 2007. WENDLING, I.; DUTRA, L.F.; GROSSI, F. Produção de mudas de espécies lenhosas. Colombo: Embrapa Florestas. 2006. 54 p. (Embrapa Florestas. Documentos, 130).

51
4 FUNGICIDAS NA DESINFESTAÇÃO DE SEMENTES DE Myracrodruon urundeuva Allemão (AROEIRA-DO-SERTÃO)
RESUMO - Os fungicidas são ocasionalmente utilizados visando desinfestar e evitar a proliferação in vitro de fungos prejudiciais ao desenvolvimento dos explantes. Objetivou-se avaliar diferentes fungicidas na desinfestação e germinação in vitro de sementes de aroeira-do-sertão (Myracrodruon urundeuva Allemão). O experimento foi conduzido no Laboratório de Cultura de Tecidos do Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da EMPAER-MT, localizado no município de Várzea Grande – MT, no período de abril a maio de 2015. O delineamento foi inteiramente casualizado em esquema fatorial 3x8, sendo três matrizes de aroeira (M1, M2 e M3) e oito tratamentos de desinfestação química (carboxina + tiram; carbendazim + tiram; oxicloreto de cobre + mancozebe; piraclostrobina; combinação entre cada um dos três primeiros ingredientes com o piraclostrobina; testemunha) com cinco repetições de 24 sementes cada. As sementes passaram por assepsia inicial com Tween 20® (3,0 ml L-1), álcool 70% (v/v) e hipoclorito de sódio a 1,5% (v/v), posteriormente foram misturadas nos fungicidas por fricção. Aos 20 dias após a semeadura em meio de cultura MS ocorreu à avaliação da contaminação por fungos (%) e da germinação (%). A menor contaminação fúngica ocorreu utilizando-se carboxina + tiram com cerca de 95% de controle. A germinação ocorreu por volta do sexto dia com médias entre 20 e 46%. O fungicida contendo carboxina + tiram (Vitavax-thiram®) foi eficiente na desinfestação, reduzindo a contaminação em sementes de aroeira-do-sertão (M. urundeuva All.) germinadas in vitro.
Palavras-chave: carboxina, carbendazim, contaminação, tiram.

52
FUNGICIDES IN DISINFECTION OF SEEDS Myracrodruon urundeuva Allemão
ABSTRACT - Fungicides are occasionally used in order to disinfect and prevent the proliferation of harmful fungi in vitro development of explants. This study aimed to evaluate different fungicides in disinfection and in vitro germination in seeds of Myracrodruon urundeuva Allemão. The experiment was conducted at the Regional Center Tissue Culture Laboratory of Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) of EMPAER-MT, in the municipality of Várzea Grande - MT, from April to May 2015. The design was completely randomized in a factorial scheme 3x8, three matrices (M1, M2 and M3), and eight chemical disinfestation treatments (carboxin+thiram, carbendazim+thiram, copper oxychloride+mancozeb; pyraclostrobin; combination of each of the first three ingredients with the pyraclostrobin, witness) with five replicates of 24 seeds each. Seed aseptically passed through initial and Tween 20® (3,0 ml L-1), 70% ethanol (v/v) sodium hypochlorite and 1,5% (v/v) were subsequently mixed in the friction fungicides . At 20 days after sowing in MS medium was the evaluation of fungal contamination (%) and germination (%). The smallest fungal contamination occurred using carboxin+thiram about 95% of control. Germination occurred around the sixth day with averages between 20 and 46%. The fungicide containing carboxin + thiram (Vitavax-thiram®) was effective in disinfection, reducing contamination in seed of M. urundeuva All. germinated in vitro.
Keywords: carboxin, carbendazim, contamination, thiram.

53
4.1 Introdução
A micropropagação das espécies florestais brasileiras tem sido realizada
basicamente utilizando-se propágulos provenientes de sementes germinadas in
vitro, dada a dificuldade em se obter material de plantas adultas, livre de
microrganismos (Oliveira et al., 2013). Diversos fatores podem afetar o potencial
germinativo das sementes, dentre eles, a presença de microrganismos, como fungos
(Effegem et al., 2014).
Um dos maiores problemas na produção de mudas in vitro, é a contaminação
por fungos e bactérias, principalmente na superfície dos explantes durante as etapas
de micropropagação, além da contaminação endógena (Oda et al., 2003; Londe et
al., 2007). O índice de contaminação pode ser resultante das próprias matrizes ou
do manuseio em laboratório (Debergh; Zimmerman, 1991).
Para que a plântula formada a partir da germinação in vitro possa ser fonte de
explante, os métodos de desinfestação devem ser eficazes, proporcionando total
ausência de agentes patológicos (Nascimento et al., 2007). Assim, as concentrações
das soluções desinfestantes, bem como as combinações dos ingredientes ativos e
os tempos de exposição, podem variar consideravelmente (Montarroyos, 2000). Este
controle está destinado a proteger as plantas de duas maneiras, seja pela destruição
do inóculo antes que se espalhe e forme micélio ou bloqueando a doença quando já
instalada na planta, seja local ou sistêmica (Cid e Zimmermann, 2006).
O uso de fungicidas pode ser recomendado tanto para a desinfestação
superficial dos tecidos, quanto, para ser adicionado ao meio de cultura (Grattapaglia
e Machado 1998; Torres et al., 1998), quando há contaminação microbiana
proveniente de infecções sistêmicas das plantas matrizes (Wendling et al., 2006).
A escolha do fungicida deve ser baseada no seu amplo espectro de ação, que
seja pouco tóxico às culturas nas concentrações necessárias para controle de
patógenos (Pasqual et al., 2010). A eficácia do tratamento de sementes pode ser
comprometida pelo uso de um único ingrediente ativo, sendo a mistura de fungicidas
com diferentes princípios ativos uma prática recomendável (Coutinho et al., 2012).
Existem diferentes grupos de ingredientes ativos na formulação dos
fungicidas, assim o mecanismo de ação no metabolismo dos fungos é muito variado.
Conforme Cid e Zimmermann (2006) o cobre (presente em alguns fungicidas) é
absorvido pelas células do fungo ligando-se a proteínas e causando um transtorno

54
geral no metabolismo das células do fungo. Para o composto thiram, que tem
afinidade com metais pesados, ocorre a ligação em metais presentes nas enzimas.
No grupo das estrobilurinas (pyraclostrobin) ocorre inibição da germinação de
esporos, desenvolvimento de tubos germinativos e a formação de apressórios
(Zauza et al., 2008).
Sendo assim, torna-se necessário o uso de produtos que promovam a
desinfecção dos tecidos sem comprometer a viabilidade dos mesmos. Portanto o
objetivo no presente trabalho foi analisar a eficiência de fungicidas na desinfestação
e germinação in vitro de sementes de aroeira-do-sertão (Myracrodruon urundeuva
Allemão).
4.2 Material e Métodos
O experimento foi realizado no Laboratório de Cultura de Tecidos do Centro
Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da Empresa Mato-
grossense de Pesquisa, Assistência e Extensão Rural (EMPAER-MT) localizado no
município de Várzea Grande – MT, no período de abril a maio de 2015.
O delineamento experimental foi inteiramente casualizado em esquema
fatorial 3x8, sendo três matrizes (M1, M2 e M3) de aroeira-do-sertão e oito
tratamentos de desinfestação química (Tabela 1), totalizando 24 tratamentos com
cinco repetições de 24 sementes cada.
TABELA 1. Tratamentos com os fungicidas utilizados.
Tratamento Produto comercial Ingrediente ativo
T Testemunha (sem fungicida)
V Vitavax-thiram carboxina + tiram
D Derosal Plus carbendazim + tiram
C Cuprozeb oxicloreto de cobre + mancozebe
P Pyraclostrobin Piraclostrobina
V+P Vitavax-thiram + Pyraclostrobin carboxina + tiram + piraclostrobina
D+P Derosakl Plus + Pyraclostrobin carbendazim + tiram + piraclostrobina
C+P Cuprozeb + Pyraclostrobin oxicloreto de cobre + mancozebe + piraclostrobina

55
As sementes foram coletadas em matrizes posicionadas a uma distância de
cerca de 30 metros entre si, em uma área de reserva pertencente a uma empresa
comercial situada na Estrada da Guia, Rodovia MT 010, KM 27 - Cuiabá-MT, na
segunda quinzena de setembro de 2014. Ocorreu precipitação na região dez dias
antes da coleta, no dia o tempo estava ensolarado com temperatura média
registrada de 36°C.
Foram feitos testes para identificar as espécies de fungos presentes nos
tratamentos testados, pois essa informação poderia contribuir no ajuste da assepsia
com a utilização de produtos direcionados às espécies. Observaram-se em
microscópio óptico comum lâminas com partes de fungos isolados em placas de
Petri contendo meio de cultura próprio, pois a identificação é baseada na morfologia.
Os fungos foram repicados e cultivados em novas placas e passaram por estresse,
porém não foi possível a identificação pela ausência de estruturas reprodutivas
necessárias para o reconhecimento da espécie (Figura 1).
FIGURA 1. Fungos obtidos a partir da germinação de sementes de aroeira-do-
sertão (M. urundeuva All.). A e B= observação macroscópica de
fungos isolados. D e E= manipulação dos fungos para causar
estresse. E e F= observação microscópica de fungos isolados
(Cuiabá-MT, 2015).

56
Os recipientes utilizados para inocular as sementes foram frascos cilíndricos
de vidro com capacidade de 250 ml, diâmetro externo de 68 mm e altura de 100 mm.
Em cada frasco foi adicionado 20 ml de meio de cultura MS, formulado por
Murashige e Skoog (1962) suplementado com 30 g L-1 de sacarose, solidificado com
2,5 g L-1 de Phytagel® e pH ajustado em 5,8± 0,2. Após o preparo, os frascos foram
esterilizados em autoclave, em temperatura de 120°C e 1 kgf cm-2 de pressão, por
20 minutos. Na sequência, foram retirados da autoclave e armazenados em
temperatura ambiente para solidificação do meio de cultura.
Na organização do experimento, primeiramente foram separados 24 frascos
de vidro representando os tratamentos, em seguida as sementes armazenadas
foram homogeneizadas e 20 g de sementes, armazenadas há seis meses, foram
pesadas e separadas em cada um dos frascos, que posteriormente foram utilizadas
para compor as repetições.
As sementes passaram então por uma assepsia inicial antes da aplicação dos
fungicidas. Após a pesagem foram lavadas com detergente Tween 20® (3,0 ml L-1),
mexendo por três minutos e enxaguadas três vezes com água destilada. Na
sequência foram adicionados 20 ml de álcool 70% (v/v) em cada frasco com as
sementes que foram agitadas por três minutos, seguido por três enxágues em água
destilada. A próxima etapa foi acrescentar a solução de hipoclorito de sódio (Start®
cloro ativo 10% v/v) diluído a 1,5% (v/v) por 15 minutos com subsequentes três
enxágues em água destilada e autoclavada.
Para o contato com os fungicidas, as 20 g de sementes de cada tratamento
foram colocadas em sacos plásticos (15x27 cm) contendo 0,5 ml para cada produto
químico. A determinação da dose foi feita de acordo com a quantidade adequada
para cobertura das sementes, após esse procedimento não foi realizado nenhum
enxágue. Foram então homogeneizadas por fricção até a cobertura total das
sementes pelo produto (Figura 2). Para o tratamento testemunha as sementes
passaram apenas pela assepsia inicial e foram inoculadas sem o contato com o
fungicida.

57
FIGURA 2. Sementes de aroeira-do-sertão (M. urundeuva All.) em contato com
fungicida. A= saco plástico com o fungicida. B= sementes
homogeneizadas e cobertas pelo fungicida (Cuiabá-MT, 2015).
Após a assepsia, as sementes foram levadas à câmara de fluxo laminar para
semeadura em meio de cultura. A câmara foi limpa com álcool 70% e as pinças
utilizadas para colocar as sementes em contato com o meio de cultura dentro dos
frascos foram esterilizadas em autoclave. Para cada frasco inoculado, as pinças
passaram por flambagem com álcool 95% e as foram utilizadas frias para evitar
injúrias ao tecido vegetal. Para cada tratamento foram semeados três frascos
(subamostras) por repetição com oito sementes cada, totalizando 24 sementes.
Ao final da semeadura e identificação de acordo com os tratamentos, os
frascos foram vedados com plástico filme para evitar possíveis contaminações, e
levados para uma sala de crescimento com temperatura média de 25 ±2°C e
fotoperíodo de 16 horas, sendo a intensidade luminosa de 25 mol m-2 s-1 (medida em
luxímetro) fornecida por lâmpadas do tipo fluorescente branca fria.
Aos 20 dias após a semeadura ocorreu à avaliação por se tratar do período
onde se observou a finalização da germinação das sementes. Consistiu na
observação e contagem das sementes contaminadas por fungos (%) e a germinação
(%) de cada semente. Foram consideradas germinadas às sementes que
apresentaram emissão de raiz (cerca de 2 mm). Na contaminação por fungos, a
porcentagem foi obtida por: X(%) = N*100 / 24, em que N = número de sementes
contaminadas. A germinação foi calculada a partir da expressão: G (%) = N/A * 100,
em que: N = número de sementes germinadas; A= número total de sementes
colocadas para germinar.

58
Os dados coletados foram submetidos à análise da variância e para
germinação (%) foram transformados em arco seno (Santana e Ranal, 2000). As
médias foram então comparadas pelo teste de Tukey a 5% de probabilidade. Foi
utilizado o programa estatístico Assistat versão 7.7 Beta (Silva, 2011).
4.3 Resultados e Discussão
A análise estatística revelou interação significativa (p < 0,01) entre os fatores
testados [matrizes (M1, M2, M3) e tratamentos com fungicidas (Testemunha, V, D,
C, P, VP, DP, CP)], para as três variáveis analisadas.
A presença de bactérias no experimento ocorreu de maneira esporádica entre
os tratamentos testados, não sendo considerada limitante ao desenvolvimento dos
explantes. O uso do hipoclorito na assepsia inicial pode estar relacionado a este
fato, pois a ação bactericida do hipoclorito se deve ao ácido hipocloroso (HOCl) que
gera uma reação oxidativa que interfere no metabolismo celular bacteriano (Pasqual
et al., 2010).
Nas médias para a variável contaminação por fungos (%), houve interação
entre os fatores testados, verificando-se melhores resultados nas matrizes M2 e M3
utilizando o fungicida V (carboxina + tiram), com 0,83% e 5,83% respectivamente
(Tabela 2).
TABELA 2. Médias para variável contaminação por fungos (%) em sementes de
diferentes matrizes de aroeira-do-sertão (M. urundeuva All.),
submetidas ao tratamento com fungicidas (Cuiabá-MT, 2015).
Fungicidas
Matrizes M1 M2 M3
Testemunha 74,1 Aa 100 Aa 100 Aa V 61,6 ABa 0,8 Db 5,8 Db D 9,1 Cc 39,1 BCb 100 Aa C 29,1 BCb 27,5 CDb 65,8 ABCa P 18,3 Cb 44,1 BCab 68,3 ABa
VP 12,5 Cc 40,0 BCb 69,1 ABa DP 10,8 Cb 65,8 ABa 54,1 BCa CP 6,6 Ca 12,5 CDa 32,5 CDa
Médias seguidas pela mesma letra não diferem estatisticamente entre si de acordo com o Teste de Tukey ao nível de 5% de probabilidade. Letras maiúsculas se referem às colunas e letras minúsculas às linhas (CV= 40,18%).

59
Ao analisar as médias para esta variável observou-se que as matrizes
apresentaram comportamentos diferentes para cada produto aplicado. Na matriz M1
os fungicidas D e CP apresentaram um controle em torno de 90% em comparação à
testemunha com apenas 26%. Nas matrizes M2 e M3 o controle com o uso do
fungicida V foi por volta de 99 e 95% respectivamente em comparação a
testemunha. O que sugere a necessidade do uso de fungicidas no processo de
desinfestação para a espécie.
A variabilidade genética é encontrada naturalmente dentro das populações e
entre os indivíduos, portanto deve-se considerar que sementes retiradas da mesma
matriz possuem características únicas. E mesmo com a uniformização das amostras
para realização do experimento foram observadas diferença dentro de cada matriz
analisada.
É importante destacar que mesmo com a utilização de agentes químicos
ainda assim pode ocorrer contaminação. A aplicação se justifica pela necessidade
de controle, minimizando as perdas de explantes no cultivo in vitro. A melhor
estratégia para controlar a contaminação é estabelecer culturas assépticas, pois a
liberação de plantas contaminadas no ambiente traz consigo o risco de doença em
larga escala e/ou mau desempenho das plantas no campo (Cassells, 2001).
As espécies arbóreas apresentam dificuldades para o estabelecimento in vitro
principalmente se for utilizado material vegetal de indivíduos adultos, que podem
apresentar infestação interna ou externa por microrganismos (Couto et al., 2004). Há
relatos da presença de microrganismos endofíticos em tecidos vegetais em um
grande número de espécies de plantas (Pasqual et al., 2010).
Na coleta das sementes utilizadas neste experimento houve cuidado em não
misturar material genético entre as matrizes, além da utilização de lona para evitar
contato dos diásporos com o solo que aumentaria a possibilidade de contaminação
dos tecidos. Em cultura de tecidos o uso de explantes provenientes do campo
(raízes, sementes, gemas ou folhas), apresenta grandes problemas de
contaminação, mesmo ao realizar uma assepsia previa a inoculação (Cid e
Zimmermann, 2006).
O surgimento da contaminação ocorreu em diferentes dias após a semeadura
e implantação do experimento, fato que não limitou a germinação e estabelecimento
das plântulas obtidas. A contaminação latente reduz a confiabilidade do sistema de

60
produção de mudas in vitro, pois pequenas mudanças nas condições ambientais
(temperatura, pH e composição do meio de cultura, aclimatização) podem permitir a
rápida proliferação do contaminante (Costa et al., 2010).
Ao adicionar o fungicida na assepsia pode-se provocar uma situação de
estresse na cultura. Condições de estresse podem estimular o desenvolvimento de
fungos endofíticos, que geralmente se encontram em estado de latência na semente
e que passam a competir com o embrião por nutrientes minerais e orgânicos,
presentes no endosperma dessa semente (Almeida et al., 2005).
A eficácia na assepsia depende de alguns fatores tais como o produto
utilizado, tempo de exposição e dose adequada, além disso, características da
semente também devem ser consideradas. O tegumento da semente de aroeira-do-
sertão não é totalmente liso, apresentando rugosidade na superfície. Essa estrutura
da semente pode favorecer o transporte de microrganismos e dificultar a assepsia,
pois as rugosidades se tornam barreiras para a penetração e ação dos agentes
desinfestantes (Figura 3).
FIGURA 3. Aspecto do tegumento das sementes de aroeira-do-sertão (M.
urundeuva All.) (Cuiabá-MT, 2015).
Nos dados da variável germinação (%) foi possível observar que as sementes
das três matrizes não tiveram a germinação prejudicada pela utilização dos
fungicidas. Porém a matriz M2 se sobressaiu às demais por obter resultados
superiores em todos os tratamentos testados (Tabela 3).

61
TABELA 3. Médias para a variável germinação (%) em sementes de diferentes
matrizes de aroeira-do-sertão (M. urundeuva All.), submetidas ao
tratamento com fungicidas (Cuiabá-MT, 2015).
Fungicidas
Matrizes M1 M2 M3
Testemunha 20,3 Bb 34,6 Aa 25,6 BCab V 35,9 Aa 31,1 Aa 22,3 Cb
D 33,6 ABa 36,0 Aa 2,3 Db C 34,6 ABa 35,0 Aa 38,9 ABa P 29,2 ABb 40,6 Aa 41,6 Aa
VP 30,4 ABb 45,9 Aa 39,9 ABab DP 32,6 ABa 36,2 Aa 42,6 Aa
CP 21,8 ABb 44,0 Aa 43,5 Aa Médias seguidas pela mesma letra não diferem estatisticamente entre si de acordo com o Teste de Tukey ao nível de 5% de probabilidade. Letras maiúsculas se referem às colunas e letras minúsculas às linhas (CV= 22,43%).
A germinação iniciou por volta do sexto dia após a semeadura em sementes
tratadas com fungicidas e na testemunha, por volta do terceiro dia. Verificou-se que
o uso dos fungicidas retardou a germinação das sementes, porém, não inibiu e nem
reduziu a porcentagem em nenhuma das matrizes testadas.
A média de germinação observada foi entre 20 e 46% nos genótipos
utilizados neste estudo. Vale ressaltar que as sementes de aroeira não apresentam
dormência física (Nunes et al., 2008) e a taxa de germinação varia de 20 a 90%,
com valores abaixo de 70% de germinação sendo os mais frequentes (Dorneles et
al., 2005).
De acordo com a observação visual no momento da avaliação do
experimento, não foi verificado nenhum dano nas plântulas formadas em nenhum
dos tratamentos testados. Não foram observadas injúrias cloróticas e necróticas nas
folhas, nem presença de superbrotações ou outros sintomas de desenvolvimento
anormal nas plântulas produzidas, sugerindo ausência de fitotoxicidade.
Ao comparar os dados da variável contaminação por fungos com a
porcentagem de germinação das sementes foi constatado que a presença desses
microrganismos não afetou o processo. Com exceção do tratamento D para a matriz
M3 que apresentou a menor média de germinação, provavelmente afetada pela
grande incidência observada na variável contaminação por fungos.

62
Infecções fúngicas podem interferir no crescimento in vitro e geralmente
resultam na morte das plantas (Pasqual et al., 2010). Portanto, apesar de ocorrer
germinação e formação de plântula, para a cultura de tecidos é importante que o
material propagado seja livre de agentes contaminantes. No experimento foi possível
observar que em alguns frascos ocorreu contaminação em algumas sementes, mas
outros foram totalmente contaminados pelos microrganismos, dessa forma a perda
do material foi inevitável (Figura 4).
FIGURA 4. Aspecto visual das plântulas obtidas a partir da germinação de
sementes de aroeira-do-sertão (M. urundeuva All.) (Cuiabá-MT,
2015).
De modo geral, as matrizes apresentaram diferentes respostas quando
colocadas em contato com determinado tipo de fungicida neste estudo. As espécies
florestais respondem de maneiras diversas, uma vez que os indivíduos, dentro de
uma população de uma área de coleta de sementes, podem apresentar grande
variabilidade, principalmente em se tratando de espécies alógamas (Mantell et al.,
1994; Ferreira et al., 2005).
4.4 Conclusão
O tratamento utilizando o fungicida a base de 1% m/v carboxina + 1% m/v
tiram na dose de 0,5 ml em 20 g de sementes foi eficiente na desinfestação,
reduzindo a contaminação em sementes de aroeira-do-sertão (M. urundeuva All.),
germinadas in vitro.
4.5 Referências Bibliográficas

63
ALMEIDA, C.V.; YARA, R.; ALMEIDA, M. Fungos endofíticos isolados de ápices caulinares de pupunheira cultivada in vivo e in vitro. Pesquisa Agropecuária Brasileira, Brasília, v.40, p.467-470, 2005. CASSELLS, A.C. Contamination and its impact in tissue culture. Acta Horticulturae, The Hague, v.560, p.353-359, 2001. CID, L.P.B.; ZIMMERMANN, M.J. A contaminação in vitro de plantas. Embrapa Recursos Genéticos e Biotecnologia, 2006. 20.p. (Boletim de Pesquisa e Desenvolvimento n.122). COSTA, M.G.C.; SCHERWINSKI-PEREIRA, J.E.; OTONI, W.C. Importância das contaminações e dos microrganismos endêmicos na cultura de células, tecidos e órgãos de plantas. In: SCHERWINSKI-PEREIRA, J.E. (editor técnico). Contaminações microbianas na cultura de células, tecidos e órgãos de plantas. Brasília-DF: Embrapa Informação Tecnológica, 2010, p.17-59. COUTINHO, W.M.; ALMEIDA, R.P.; DANTAS, F.V.; SOARES, D.J.; ARAÚJO, A.E.; MILANI, M. Eficácia de misturas de fungicidas químicos na micobiota e na qualidade fisiológica de sementes de mamoneira. Embrapa Algodão, 2012. 22p. (Boletim de pesquisa e desenvolvimento n.91) COUTO, J.M.F.; OTONI, W.C.; PINHEIRO, A.L.; FONSECA, E.P. Desinfestação e germinação in vitro de sementes de mogno (Swietenia macrophylla King). Revista Árvore, Viçosa, v.28, n.5, p.633-642, 2004. DEBERGH, P.C.; ZIMMERMAN, R.H.; Micropropagation: Technology and Application. 1. ed. London: Kluwer Academic Publishers, 1991. 48p. DORNELES, M. C.; RANAL, M. A.; SANTANA, D.G. Germinação de diásporos recém-colhidos de Myracrodruon urundeuva Allemão (Anacardiaceae) ocorrente no cerrado do Brasil Central. Revista Brasileira de Botânica, São Paulo, v.28, n.2, p.399-408, 2005. EFFEGEM, C.; GONTIJO, A.B.P.L.; CAMPANHARO, A.; GONTIJO, I. Desinfestação e germinação in vitro de sementes de pimenta-do-reino (Piper nigrum L.). Enciclopédia Biosfera, Goiânia, v.10, n.18, p. 1221-1228. FERREIRA, R.A.; DAVIDE, A.C.; ALCÂNTARA, E.N. de; MOTTA, M.S. Efeito de herbicidas de pré-emergência sobre o desenvolvimento inicial de espécies arbóreas. Revista Brasileira de Herbicidas, Londrina, v.4, p.133-145, 2005. GRATTAPAGLIA, D.; MACHADO, M.A. Micropropagação. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa – SPI / Embrapa – CNPH, 1998, v.1, p.183-260. LONDE, L.N.; SOUZA, C.S.; VIEIRA, C.U.; BONETTI, A.M.; KERR, W.E. Efeito do benomyl e identificação de fitopatógenos em meio MS para controle da contaminação na micropropagação de Anacardium humile (Anacardiaceae). Bioscience Journal, Uberlândia, v.23, n.3, p. 94-100, 2007.

64
MANTELL, S.H.; MATTHEWS, J.A.; McKEE, R.A. Princípios de biotecnologia em plantas. Ribeirão Preto: Sociedade Brasileira de Genética, 1994. 344p. MONTARROYOS, A.V.V. Contaminação in vitro. ABCTP Notícias, Brasília, n. 36/37, p. 5-10, 2000. MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiologia Plantarum, Copenhagen, v.15, p.473-97, 1962. NASCIMENTO, P.K.V.; FRANCO, E.T.H; FRASSETTO, E.G. Desinfestação e germinação in vitro de sementes de Parapiptadenia rígida Bentham (Brenam). Revista Brasileira de Biociências, Porto Alegre, v. 5, supl. 2, p. 141-143, 2007. NUNES, Y.R.F.; FAGUNDES, M.; ALMEIDA, H.S.;VELOSO, M.D.M.. Aspectos ecológicos da aroeira (Myracrodruon urundeuva Allemão- Anacardiaceae): fenologia e germinação de sementes. Revista Árvore, Viçosa, v. 32, n. 2, p. 233-243, 2008. ODA, M.L.; FARIA, R.T.; FONSECA, I.C.B.; SILVA, G.L. Avaliação da fitotoxicidade de fungicidas e germicida na propagação in vitro de Oncidium varicosum Lindl. (Orchidaceae) para o controle de microorganismos. Semina: Ciências Agrárias, Londrina, v. 24, n. 2, p. 273-276, 2003. OLIVEIRA, L.S.; DIAS, P.C.; BRONDANI, G.E. Micropropagação de espécies florestais brasileiras. Pesquisa Florestal Brasileira, Colombo, v.33, n.76, p.439-453, 2013. PASQUAL, M.; DUTRA, L.F.; ARAUJO, A.G.; PEREIRA, A.R. Prevenção de contaminações microbianas na cultura de células, tecidos e órgãos de plantas. In: SCHERWINSKI-PEREIRA, J.E. (editor técnico). Contaminações microbianas na cultura de células, tecidos e órgãos de plantas. Brasília-DF: Embrapa Informação Tecnológica, 2010, p.61-161. SANTANA, D.G.; RANAL, M.A. Análise estatística na germinação. Revista Brasileira de Fisiologia Vegetal, Campinas, v.12 (Edição especial), p.205-237, 2000. SILVA, F.A.S. ASSISTAT versão 7.7 beta. Campina Grande-PB: Assistência Estatística, Departamento de Engenharia Agrícola do CTRN - Universidade Federal de Campina Grande, Campus de Campina, 2011. TORRES, A. C.; CALDAS, L.S.; FERREIRA, A.T. Retrospectiva da cultura de tecidos de plantas. In: In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa – SPI / Embrapa – CNPH, 1998, v.1, p.11-20. WENDLING, I.; DUTRA, L.F.; GROSSI, F. Produção de mudas de espécies lenhosas. Colombo: Embrapa Florestas. 2006. 54 p. (Embrapa Florestas. Documentos, 130).

65
ZAUZA, E.A.V.; COUTO, M.M.F.; MAFFIA, L.A.; ALFENAS, A.C. Eficiência de fungicidas sistêmicos no controle da ferrugem do Eucalyptus. Revista Árvore, Viçosa, v. 32, n. 5, p. 829-835, 2008.

66
5 CONCENTRAÇÕES DE SAIS DO MEIO DE CULTURA MS NO
ESTABELECIMENTO in vitro DE Myracrodruon urundeuva Allemão
RESUMO - As soluções que compõem os meios de cultura não exercem efeito puramente nutritivo, mas também influenciam no crescimento celular e morfogênese, assim as modificações no teor de macro e micronutrientes visam o melhor desempenho dos tecidos. O objetivo foi comparar diferentes concentrações de sais do meio de cultura MS na germinação e desenvolvimento inicial in vitro de Myracrodruon urundeuva All. O experimento foi conduzido no Laboratório de Cultura de Tecidos do Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da EMPAER-MT, em Várzea Grande – MT, de junho a julho de 2015. O delineamento foi inteiramente casualizado em esquema fatorial 3x5, sendo três matrizes de aroeira (M1, M2 e M3) e cinco concentrações de sais (0; 25; 50; 75 e 100%), compondo 15 tratamentos com quatro repetições. As sementes passaram por assepsia inicial e em câmara de fluxo laminar foram inoculadas em meio de cultura MS nas concentrações testadas. A avaliação ocorreu aos 25 dias após a semeadura. Em relação às matrizes, M1 apresentou 78,5% de germinação e 0,81 cm de média do comprimento da parte aérea. A concentração de 25% de sais do meio MS proporcionou resultados superiores para germinação (68,7%) e comprimento da parte aérea (0,64 cm). Houve interação significativa apenas para contaminação. A matriz M3 não apresentou contaminação, em M1 a menor média foi 4,1% e em M2, 23,1% no tratamento com 50% dos sais. O estabelecimento in vitro de M. urundeuva pode ser realizado utilizando-se 25% da concentração de sais do meio de cultura MS.
Palavras-chave: germinação, micropropagação, nutrição.

67
SALT CONCENTRATIONS IN THE CULTURE MEDIUM in vitro ESTABLISHMENT
OF Myracrodruon urundeuva Allemão
ABSTRACT - The solutions comprise the culture media did not exert a purely nutritional effect, but also influence cellular growth and morphogenesis, as changes in the level of macro and micronutrients are aimed at improved performance of fabrics. The objective was to compare different concentrations of MS medium salts on germination and early development in vitro of Myracrodruon urundeuva All. The experiment was conducted at Tissue Culture Laboratory of the Regional Center for Research and Technology Transfer (CRPTT) of EMPAER- MT in Varzea Grande - MT, June-July 2015. The design was completely randomized in a factorial 3x5, three matrices (M1, M2 and M3) and five salt concentrations (0, 25, 50, 75 and 100%), making up 15 treatments with four replications. The seeds have undergone aseptic initial and laminar flow chamber were inoculated in MS medium at the concentrations tested. The evaluation occurred 25 days after sowing. Regarding the matrices, M1 showed 78,5% germination and 0,81 cm of shoot length average. The concentration of 25% of the MS culture medium provided superior results for germination (68,7%) and shoot length (0,64 cm). There was a significant interaction only for contamination. The M3 matrix showed no contamination in M1 the lowest average was 4,1% and M2, 23,1% in the treatment with 50% of salts. The in vitro establishment of M. urundeuva can be performed using 25% concentration of MS culture medium salts.
Keywords: germination, micropropagation, nutrition.

68
5.1 Introdução
A biotecnologia moderna, que inclui metodologias avançadas nas áreas de
genética, biologia molecular, cultura de tecidos, engenharia genética, clonagem de
espécies e variedades biológicas, tem sido apontada como um instrumento de
resgate da biodiversidade da floresta tropical (Daquinta et al., 2001; Carvalho et al.,
2006).
A micropropagação visando a multiplicação e preservação de espécies deve
assegurar a manutenção da variabilidade genética (Bellintani et al., 2007), desse
modo, a germinação in vitro garante a manutenção de características oriundas de
diferentes populações. Constitui um modo de se manter disponíveis explantes livres
de contaminação para aplicação de técnicas de regeneração por cultura de tecidos e
transformação genética, além de ser altamente conveniente para manutenção de
coleções de plantas de genótipos diferentes (Cabral et al., 2003).
O meio de cultura é formulado considerando-se as exigências nutricionais
sendo ajustados às necessidades de cada espécie e etapas in vitro (Santos-Serejo
et al., 2006), fornecendo as substâncias essenciais para o crescimento e
controlando o padrão de desenvolvimento in vitro (Caldas et al., 1998). Assim sendo,
o suprimento inapropriado de um elemento essencial, em excesso ou deficiência,
pode acarretar em prejuízos (Schmildt et al., 2015), tais como, sintomas de
deficiência mineral e até a morte dos propágulos (Monteiro et al., 2000).
No cultivo in vitro, as soluções que compõem os meios de cultura não
exercem efeito puramente nutritivo, mas também influenciam o crescimento celular e
a morfogênese por meio de propriedades osmóticas (George, 1996). Alguns foram
especificamente desenvolvidos para fornecer os requisitos particulares à espécie
trabalhada, como o meio básico de cultura de Murashige e Skoog, desenvolvido
inicialmente para o tecido de Nicotiana tabacum em 1962. O meio MS é um dos
meios nutritivos mais utilizados na micropropagação de diversas espécies vegetais,
desde arbóreas (Mantovani et al., 2001; Rocha et al., 2007; Brondani et al., 2010;
Fernandes et al., 2013), ornamentais (Oliveira et al., 1996; Ribeiro et al., 2008;
Carvalho et al., 2009), frutíferas (Pasqual e Ando, 1989; Oliveira e Oliveira, 1997;
Barboza et al., 2004; Braga et al., 2009; Pereira et al., 2011;), entre outras.
A possibilidade de reduzir a concentração de sais do meio MS visa um
desenvolvimento adequado do tecido vegetal e com a vantagem da redução de

69
custos. Trabalhos relacionados a esse tema foram desenvolvidos com crisântemo,
no qual Oliveira et al. (1996) obtiveram resultados eficientes ao cultivar brotos de
crisântemo (Dendranthema grandifolia Tzvelev) em meio com 75% de sais MS. Em
pesquisa com a germinação de sementes de limoeiro „cravo‟ (Citruslimonia Osbeck),
Schmildt et al. (2015) mostraram bons resultados no tratamento com 33,3% de sais
MS. Além de outras espécies, tais como, manjericão (Ocimum basilicum L.) com MS
na concentração total de sais (Ribeiro et al., 2007); erva-cidreira (Melissa officinalis)
com 25% (Reis et al., 2008) mostrando a importância de ajustes da concentração do
MS de acordo com as necessidades do tecido micropropagado.
A partir do exposto, o objetivo neste trabalho foi comparar concentrações de
sais no meio de cultura MS para a germinação e desenvolvimento inicial in vitro de
aroeira-do-sertão (Myracrodruon urundeuva Allemão).
5.2 Material e Métodos
O experimento foi realizado no Laboratório de Cultura de Tecidos do Centro
Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da Empresa Mato-
grossense de Pesquisa, Assistência e Extensão Rural (EMPAER-MT) localizado no
município de Várzea Grande – MT, no período de junho a julho de 2015.
O delineamento experimental foi inteiramente casualizado em esquema
fatorial 3x5, sendo três matrizes de aroeira-do-sertão (M1, M2 e M3) com nove de
meses de armazenamento, cinco concentrações das soluções de sais do meio de
cultura MS (0; 25; 50; 75 e 100%), compondo 15 tratamentos com quatro repetições
de 24 sementes cada, totalizando 360 sementes. A testemunha foi composta pela
concentração 0% de sais e foi preparada apenas com água e ágar (8 g L-1).
As sementes foram coletadas em matrizes posicionadas a uma distância de
cerca de 30 metros entre si, em uma área de reserva pertencente a uma empresa
comercial situada na Estrada da Guia, Rodovia MT 010, KM 27 - Cuiabá-MT, na
segunda quinzena de setembro de 2014. Ocorreu precipitação na região dez dias
antes da coleta, no dia o tempo estava ensolarado com temperatura média
registrada de 36°C.
Os indivíduos foram selecionados de forma aleatória considerando-se
visualmente características semelhantes como a copa, altura, produção de frutos e
sementes, bem como pela facilidade de acesso. E todos eles tinham sementes

70
abaixo de sua copa, o que dá indícios de que as sementes já tinham atingido a
maturidade fisiológica.
Posteriormente foram levadas para o Laboratório de Sementes da
Universidade Federal de Mato Grosso, no qual foi feita a limpeza separando as
sementes de gravetos e demais impurezas, sendo então armazenadas em câmara
fria na temperatura de 18ºC ±2 e umidade de 68%.
O meio de cultura MS, formulado por Murashige e Skoog (1962) foi preparado
de acordo com as concentrações de sais propostas 0, 25, 50, 75 e 100% (Tabela 1),
suplementado com 30 g L-1 de sacarose, solidificado com 2,5 g L-1 de Phytagel® e
pH ajustado para 5,8 ±0,2.
TABELA 1. Composição do meio de cultura com as concentrações de sais
utilizadas no experimento.
Componente (100%) (75%) (50%) (25%)
----------------------------------gL-1---------------------------------
Ma
cro
nu
trie
nte
s
NH4NO3 82,5 61,875 41,25 20,625
KNO3 95,0 71,25 47,5 23,75
CaCl2.H2O 44 33 22 11
MgSO4.7H2O 37 27,75 18,5 9,25
KH2PO4 17 12,75 8,5 4,25
NA2EDTA 3,73 2,7975 1,865 0,9325
FeSO4.7H2O 2,78 2,085 1,39 0,695
Mic
ron
utr
ien
tes
K 0,083 0,06225 0,0415 0,02075
H3BO3 0,63 0,4725 0,315 0,1575
MnSO4.H2O 1,69 1,2675 0,845 0,4225
ZnSO4.7H2O 0,86 0,645 0,43 0,215
Na2MoO4.2H2O 0,025 0,01875 0,0125 0,00625
CuSO4.5H2O 0,0025 0,001875 0,00125 0,000625
CoCl2.H2O 0,0026 0,00195 0,0013 0,00065
Vita
min
a
s
Tiamina 0,01 0,0075 0,005 0,0025
Piridoxina 0,05 0,0375 0,025 0,0125

71
Ác. Nicotínico 0,05 0,0375 0,025 0,0125
Inositol 10 7,5 5 2,5
Glicina 0,2 0,15 0,1 0,05
Em cada frasco cilíndrico de vidro (capacidade de 250 ml, diâmetro externo
de 68 mm e altura de 100 mm) foi adicionado 20 ml de meio de cultura. E Após o
preparo, foram levados para a esterilização em autoclave, em temperatura de 120°C
e 1 kgf cm-2 de pressão, por 20 minutos. Na sequência foram retirados da autoclave,
armazenados em estante na sala de repicagem para resfriar em temperatura
ambiente e solidificar o meio de cultura.
Para o experimento, primeiramente as sementes armazenadas foram
homogeneizadas e 20 g sementes foram retiradas, pesadas e colocadas em cada
um dos cinco frascos cilíndricos de vidro para cada matriz, totalizando 15 frascos.
Em seguida as sementes foram lavadas com detergente Tween 20® (3 ml L-1) diluído
em água destilada, mexendo por três minutos e enxaguadas três vezes com água
destilada. Posteriormente foram adicionados 20 ml de álcool 70% (v/v) em cada
frasco com as sementes que foram agitadas por três minutos, seguido por três
enxágues em água destilada.
A próxima etapa foi acrescentar a solução de hipoclorito de sódio (cloro ativo
10%) diluído a 1,5% (v/v) por 15 minutos com subsequentes três enxágues em água
destilada e autoclavada. Para finalizar a assepsia, foram então colocadas em sacos
plásticos (15x27 cm) contendo 0,5 ml do fungicida Vitavax-thiram® (carboxina +
tiram) e homogeneizadas por fricção. A determinação da dose foi feita de acordo
com a quantidade adequada para cobertura completa das sementes. Não houve
enxágue após o contato com o fungicida.
Prontamente as sementes foram levadas à câmara de fluxo laminar para
semeadura em meio de cultura. A câmara foi higienizada com álcool 70% e as
pinças utilizadas para colocar as sementes em contato com o meio de cultura dentro
dos frascos foram esterilizadas em autoclave. Para cada frasco inoculado, as pinças
passaram por flambagem utilizando-se álcool 95% e foram utilizadas frias para evitar
injúrias ao tecido vegetal. Para cada repetição de cada tratamento foram utilizados
três frascos (subamostras) com oito sementes cada.

72
Ao final da semeadura e identificação de acordo com os tratamentos, os
frascos foram vedados com plástico filme para evitar possíveis contaminações, e
levados para uma sala de crescimento, onde foram dispostos aleatoriamente em
uma prateleira de metal. A temperatura média desta sala foi de 25 ±2°C e
fotoperíodo de 16 horas, sendo a intensidade luminosa de 25 mol m-2 s-1 (medida em
luxímetro) fornecida por lâmpadas do tipo fluorescente branca fria.
A avaliação foi realizada aos 25 dias após a semeadura, na qual foi feita a
contagem individual das sementes germinadas e/ou plântulas contaminadas por
microrganismos, e a determinação da altura média de parte aérea considerada a
partir da base da brotação até o ápice, medida com régua milimetrada através do
frasco de vidro para não destruir o explante (Figura 1).
FIGURA 1. Medição da altura de plântulas (parte aérea) de aroeira-do-sertão (M.
urundeuva All.) (Cuiabá-MT, 2015).
Foram avaliadas as variáveis: germinação (%), altura da parte aérea (cm) das
plântulas e contaminação por microrganismos (%). A germinação (%G) foi calculada
pela expressão: G (%) = N/A * 100, em que: N = número de sementes germinadas;
A= número total de sementes colocadas para germinar. Na contaminação por
microrganismos, a porcentagem foi obtida por: X(%) = N*100 / 24, em que N =
número de sementes contaminadas.
Os dados coletados foram submetidos à análise da variância, as médias dos
tratamentos qualitativos foram comparadas pelo teste de Tukey a 5% de

73
probabilidade e as médias dos tratamentos quantitativos pela regressão. Foi
utilizado o programa estatístico Assistat versão 7.7 Beta (Silva, 2011).
5.3 Resultados e Discussão
Na análise de variância das matrizes testadas em diferentes concentrações
do meio MS, observou-se diferença estatística em todas as variáveis analisadas
para os dois fatores (matrizes e concentração de sais do meio MS). Para as
variáveis germinação (%) e altura da parte aérea (cm) ocorreu efeito significativo (p
< 0,01) das matrizes e concentrações testadas de forma independente. Houve
interação significativa (p < 0,01) apenas na contaminação por microrganismos (%).
O início da germinação das sementes de aroeira-do-sertão foi verificado cerca
de seis dias após a inoculação das mesmas nos diferentes meios de cultura. Na
comparação entre matrizes, a M1 apresentou médias superiores para germinação
(78,54%) e altura da parte aérea (0,81 cm) (Tabela 2).
TABELA 2. Médias para variável germinação (%) e altura média de parte aérea
(cm) em matrizes de aroeira-do-sertão (M. urundeuva All.) (Cuiabá-
MT, 2015).
Matrizes Germinação1 (%) Altura de parte aérea2 (cm)
M1 78,5 a 0,81 a
M2 56,8 b 0,37 b
M3 59,3 b 0,38 b
Médias seguidas pela mesma letra não diferem estatisticamente entre si. Foi aplicado o Teste de Tukey ao nível de 5% de probabilidade (CV1= 14,12%; CV2= 39,42%).
O desenvolvimento diferenciado das plântulas entre as matrizes
possivelmente foi influenciado pelo próprio genótipo da semente, que interfere na
quantidade de reservas de nutrientes na semente que são disponibilizados para o
crescimento inicial de plântulas. Considera-se que a capacidade de regeneração e
crescimento in vitro podem estar associadas não apenas ao genótipo, mas também
a planta-matriz, sob o controle de diversos fatores endógenos (Gattapaglia e
Machado, 1998; Bevitori, 2013).

74
Conforme Diniz et al. (2015) em espécies florestais podem existir variações
individuais entre árvores devido às influências ambientais durante o
desenvolvimento das sementes e à variabilidade genética.
Na regressão polinomial o ajuste foi quadrático para ambas as variáveis, com
ponto de máxima de 29% para germinação (%) (Figura 2) e 42% para altura média
de parte aérea (cm) (Figura 3).
FIGURA 2. Germinação (%) de aroeira-do-sertão (M. urundeuva All.), inoculadas
em concentrações de sais do meio MS (%) (CV= 14,12%; Cuiabá-MT,
2015).
FIGURA 3. Altura média de parte aérea (cm) em plântulas de aroeira-do-sertão
(M. urundeuva All.), em concentrações de sais do meio MS (%) (CV=
39,42%; Cuiabá-MT, 2015).
y = -0,0024x2 + 0,1408x + 66,971 R² = 0,9959
0
10
20
30
40
50
60
70
80
0 25 50 75 100
Germ
inação (
%)
Concentração de sais do meio MS (%)
y = -9E-05x2 + 0,0076x + 0,4925 R² = 0,9717
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 25 50 75 100
Altu
ra m
édia
de p
art
e
aére
a (
cm
)
Concentração de sais do meio MS (%)

75
As maiores concentrações de sais podem ter influenciado na absorção de
água necessária para iniciar o processo de germinação das sementes. A presença
de maior concentração de sais no meio de cultura pode interferir no potencial
osmótico, afetando por consequência na disponibilidade de água para o processo de
embebição da semente na germinação (Reis et al., 2008). Para que ocorra o
crescimento, a célula deve absorver água através da membrana plasmática, o que é
impulsionado pelo gradiente de potencial hídrico (o potencial hídrico no interior da
célula é menor que no meio externo ou no apoplasto). A entrada de água na célula
produz aumento no potencial de turgescência (Ψp) que ao superar a pressão limite,
atua na parede celular que se distende e a célula cresce (Taiz e Zeiger, 2009).
A quantidade elevada dos sais somada a reserva de nutrientes provenientes
da semente na formação da plântula podem ter influenciado esse resultado, visto
que houve aumento na altura da parte aérea até 50% da concentração de sais
seguida de decréscimo em concentrações maiores (75 e 100%). O tecido utilizado
como explante possui reservas de metabólitos essenciais e substâncias acumuladas
que podem mascarar os resultados de crescimento (Caldas et al., 1998)
dependendo da concentração do meio de cultura.
O meio de cultura não fornece apenas macro e micronutrientes, mas também
pode ser fonte de carboidratos, geralmente devido a adição de sacarose. Além
disso, visando maior crescimento, incluem-se vitaminas, aminoácidos e reguladores
de crescimento, em concentrações que irão depender das respostas dos diversos
materiais genéticos e condições ambientais (Brondani et al., 2010).
Com relação à contaminação por microrganismos houve somente a incidência
de fungos, não sendo observada a presença de bactérias em nenhum dos
tratamentos testados. O problema neste tipo de contaminação é que o material não
pode ser utilizado e precisará passar por autoclavagem evitando a disseminação de
esporos pelo ambiente.
Nos dados para a variável contaminação por microrganismos em que houve
interação entre os fatores testados, não houve ocorrência desses contaminantes na
matriz M3 em nenhuma das concentrações do meio MS utilizadas. Na análise de
regressão não houve ajuste da equação para M1, assim, no teste de médias a
menor contaminação (4,16%) foi observada com 75% da concentração do MS

76
(Tabela 3). Na M2 o ajuste da equação foi quadrático com menor contaminação
(23,18%) no tratamento com 50% (Figura 4).
TABELA 3. Médias para contaminação por fungos (%) em sementes e plântulas
da matriz M1 de aroeira-do-sertão (M. urundeuva All.), inoculada em
concentrações de sais do meio MS (%) (Cuiabá-MT, 2015).
Concentração de sais do
meio MS (%)
Contaminação por microrganismos (%)
0 8,3 bc
25 14,5 ab
50 21,8 a
75 4,1 c
100 17,7 ab
Médias seguidas pela mesma letra não diferem estatisticamente entre si de acordo com o Teste de Tukey ao nível de 5% de probabilidade (CV= 34,70%).
FIGURA 4. Contaminação por microrganismos (%) em sementes e plântulas de
M2 de aroeira-do-sertão (M. urundeuva All.), em concentrações de
sais do meio MS (%) (CV= 27,87%; Cuiabá-MT, 2015).
A diferença entre a matriz M3 que não apresentou contaminação para as
matrizes M1 e M2 que apresentaram incidência de contaminantes, pode estar
y = 0,0093x2 - 0,8119x + 40,561 R² = 1
0
10
20
30
40
50
60
0 25 50 75 100Co
nta
min
ação
por
mic
rorg
anis
mos
(%)
Ma
triz
M2
Concentração de sais do meio MS (%)

77
relacionada a amostra de sementes retirada ao acaso no momento do preparo do
experimento. Embora não tenha sido possível identificar os patógenos, a assepsia
não atingiu na erradicação de todos os contaminantes. Observou-se ainda, que nas
matrizes nas quais ocorreu o desenvolvimento de fungos, dentro do mesmo frasco
haviam indivíduos contaminados e sem contaminação (Figura 5).
FIGURA 5. Aspecto da contaminação por fungos em sementes e plântulas de
aroeira-do-sertão (M. urundeuva All.) (Cuiabá-MT, 2015).
Quando em contato com o meio de cultura, os fungos podem crescer como
saprófitas e então o crescimento de hifas pode aparecer no meio, no explante ou em
ambos (Pasqual et al., 2010). A contaminação é um dos pontos críticos no processo
de inoculação do material vegetal in vitro. Grattapaglia e Machado (1998)
destacaram que a qualidade da planta fornecedora do material vegetal; incluindo seu
manejo cultural e ambiental, têm elevada influência no posterior comportamento da
cultura. Fato este que pode estar associado à diferença na contaminação entre as
matrizes testadas neste estudo.
Os fungos são organismos heterotróficos e necessitam dos compostos
orgânicos que são utilizados como fonte de carbono e energia (Pasqual et al., 2010),
encontrando esse componentes no meio de cultura. Os microrganismos
contaminantes se estabelecem no meio de cultura competindo pelos nutrientes,
produzindo substâncias tóxicas e inibindo o desenvolvimento do explante,
ocasionando sua perda (Souza et al., 2007).

78
5.4 Conclusão
A germinação e o desenvolvimento inicial de plântulas de aroeira-do-sertão
podem ser realizados utilizando-se 25% da concentração de sais do meio MS,
reduzindo custos de produção sem comprometer a sobrevivência do material
vegetal.
5.5 Referências Bibliográficas
BARBOZA, S.B.S.C; CALDAS, L.S.; SOUZA, L.A.C. Micropropagação do híbrido PExSC-52 e da cultivar Smooth Cayenne de abacaxizeiro. Pesquisa Agropecuária Brasileira, Brasília, v.39, n.8, p.725-733, 2004. BELLINTANI, M.C.; LIMA, C.C.; BRITO, A.L.; SANTANA, J.R.F.; DORNELLES, A.L.C. Estabelecimento in vitro de Orthophytum mucugense e Neoregelia mucugensis, bromélias endêmicas da Chapada Diamantina, Bahia - Brasil. Revista Brasileira de Biociências, Porto Alegre, v.5, supl.2, p. 1101-1103, 2007. BEVITORI, R. Cultivo in vitro do arroz (Oryza sativa L.): conceitos básicos e protocolo. Embrapa Arroz e Feijão, 2013. 68 p. - (Documentos / Embrapa Arroz e Feijão, 285). BRAGA, F.T.; NUNES, C.F.; FAVERO, A.C.; PASQUAL, M.; CARVALHO, J.G.; CASTRO, E.M. Características anatômicas de mudas de morangueiro micropropagadas com diferentes fontes de silício. Pesquisa Agropecuária Brasileira, Brasília, v.44, n.2, p.128-132, 2009. BRONDANI. G.E.; HANSEL, F.A.; DUTRA, L.F.; WENDLING, I. Desinfestação e meio de cultura para o estabelecimento in vitro de segmentos nodais de Liquidambar styraciflua. Floresta, Curitiba, v.40, n.3, p.541-554, 2010. CABRAL, G.B.; PIRES, M.V.V.; LACERDA, A.L.; CARNEIRO, V.T.C. Introdução in vitro, micropropagação e conservação de plantas de Brachiaria sp. Brasília: Embrapa Recursos Genéticos e Biotecnologia, 2003. 4p. (Embrapa Recursos Genéticos e Biotecnologia. Comunicado Técnico, 101). CALDAS, L.S.; HARIDASAN, P.; FERREIRA, M.E. Meios nutritivos. In: TORRES, A. C., CALDAS, L. S. Técnicas e aplicações da cultura de tecidos de plantas. Brasília: ABCTP, EMBRAPA-CNPH, 1998. p.37-70. CARVALHO, A.C.P.P.; PINHEIRO, M.V.M.; DIAS, G.M.G.; MORAIS, J.P.S. Multiplicação in vitro de abacaxi ornamental por estiolamento e regeneração de brotações. Horticultura Brasileira, Brasília, v.27, n.1, p.103-108, 2009. CARVALHO, J.M.F.C.; SILVA, M.M.A.; MEDEIROS, M.J.L. Fatores inerentes à micropropagação. Campina Grande, Embrapa Algodão. Documentos, 148. 2006. 28p.

79
DAQUINTA, M.; RAMOS, L; CAPOTE, I.; LEZCANO, Y.; RODRÍGUEZ, R.; TRINA, D.; ESCALONA, M. Micropropagación de la teca (Tectona grandis L.f.). Revista Florestal Centroamericana, Turrialba, n.35, p.25-28, 2001. DINIZ, R.Q.; DINIZ, B.L.M.T.; AZEREDO, G.A.; SOUZA, V.C.; PEREIRA, E.M. Potencial germinativo de sementes de Aroeira Myracrodruon urundeuva Fr. coletadas de população no cariri paraibano. Revista Verde, Pombal, v.10, n.1, p. 154 - 159, 2015. FERNANDES, D.A.; AZEVEDO, P.H.; COSTA, R.B.; BRONDANI, G.E. Tipos de vedação e concentrações de sacarose no cultivo in vitro de Tectona grandis L.f. Revista de Agricultura, Piracicaba, v.88, n.3, p.218 – 228, 2013. GEORGE, E. F. Plant propagation by tissue culture: part 1, the technology. Edington: Exegetics, 1996. 1574 p. GRATTAPAGLIA, D.; MACHADO, M.A. Micropropagação. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa – SPI / Embrapa – CNPH, 1998, v.1, p.183-260. LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Nova Odessa: Ed. Plantarium, 1992. 352p. MANTOVANI, N.C.; FRANCO, E.T.H.; VESTENA, S. Regeneração in vitro de louro-pardo (Cordia trichotoma (Vellozo) Arrabida ex Steudel). Ciência Florestal, v.11, n.2, p. 93-101, 2001. MONTEIRO, A.C.B.A.; HIGASHI, E.N.; GONÇALVES, A.N.; RODRIGUEZ, A.P.M. A novel approach for the definition of the inorganic medium components for micropropagation of yellow passion fruit (Passiflora edulis Sims f. flavicarpa Deg.). In vitro Cellular e Developmental Biology-Plant, New York, v.36, n.6, p.527-531, 2000. MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiologia Plantarum, Copenhagen, v.15, p.473-97, 1962. OLIVEIRA, P.D.; PASQUAL, M.; PAIVA, R. Efeito de diferentes concentrações do meio MS, nitrogênio e sacarose na micropropagação de crisântemo „Orange Reagen‟. Bragantia, Campinas, v. 55, n.1, p.9-18, 1996. OLIVEIRA, R.P.; OLIVEIRA, S. Avaliação da micropropagação comercial em bananeira. Pesquisa Agropecuária Brasileira, Brasília, v.32, n.4, p.415-420, 1997. PASQUAL, M.; ANDO, A. Micropropagação de „Trifoliata‟ através da cultura de gemas axilares in vitro. Pesquisa Agropecuária Brasileira, Brasília, v.24, n.2, p.217-220, 1989. PEREIRA, G.A.; CORREA, L.S.; BOLIANI, A.C. Desinfestação e estabelecimento in vitro de explantes de bananeira „Grande naine‟ em diferentes concentrações de

80
hipoclorito de sódio. Revista Brasileira de Fruticultura, Jaboticabal, volume especial, p.222-226, 2011. REIS, E.S.; PINTO, J.E.B.P.; ROSADO, L.D.S.; CORREA, R.M. Influência do meio de cultura na germinação de sementes in vitro e taxa de multiplicação de Melissa officinalis L. Revista Ceres, Viçosa, v.55, n.3, p.160-167, 2008. RIBEIRO, M.F.; DONINI, L.P.; SOUZA, J.A.; GUISSO, A.P.; FERREIRA-MOURA, I.; BOBROWSKI, V.L.; VIÉGAS, J. Influência de diferentes concentrações de sais de MS e açúcares no cultivo in vitro de manjericão roxo (Ocimum basilicum L.). Revista Brasileira de Biociências, Porto Alegre, v. 5, supl. 2, p.57-59, 2007. RIBEIRO, M.N.O.; PASQUAL, M.; SILVA, A.B.; RODRIGUES, V.A. Diferentes concentrações de sais do meio MS e de sacarose na multiplicação in vitro de Zantedeschia aethiopica L. Spreng. (copo-de-leite). Revista Ciência Agronômica, Fortaleza, v.39, n.1, p.101-106, 2008. ROCHA, S.C.; QUORIM, M.; RIBAS, L.L.F.; KOEHLER, H.S. Micropropagação de Cabralea canjerana. Revista Árvore, Viçosa, v.31, n.1, p.43-50, 2007. SANTANA, D.G.; RANAL, M.A. Análise estatística na germinação. Revista Brasileira de Fisiologia Vegetal, Campinas, v.12 (Edição especial), p.205-237, 2000. SANTOS-SEREJO, J.A.; JUNGHANS, T.G.; SOARES, T.L.; SILVA, K.M. Meios nutritivos para micropropagação de plantas. In: SOUSA, A.S.; JUNGHANS, T.G. Introdução à micropropagação de plantas, Cruz das Almas: Embrapa Mandioca e Fruticultura Tropical, 2006. p.79-98. SCHMILDT, O.; SCHMILDT, E.R.; OLIVEIRA, M.J.V. Sais e sacarose na germinação in vitro de limoeiro „cravo‟. Revista Nucleus, Ituverava, v.12, n.1, p.215-222, 2015. SILVA, F.A.S. ASSISTAT versão 7.7 beta. Campina Grande-PB: Assistência Estatística, Departamento de Engenharia Agrícola do CTRN - Universidade Federal de Campina Grande, Campus de Campina, 2011. TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 4.ed. Porto Alegre: Artmed, 2009. 819p.

81
6 MULTIPLICAÇÃO in vitro DE Myracrodruon urundeuva Allemão EM
CONCENTRAÇÕES DE BAP (6-BENZILAMINOPURINA)
RESUMO - As citocininas são fundamentais pelo efeito na quebra da dominância apical, na proliferação de gemas axilares e influenciam no sucesso da multiplicação in vitro. O objetivo foi verificar o efeito de diferentes concentrações de BAP (6-benzilaminopurina) na indução de brotações in vitro de matrizes de aroeira-do-sertão (Myracrodruon urundeuva Allemão). O experimento foi conduzido no Laboratório de Cultura de Tecidos do Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da EMPAER-MT, localizado no município de Várzea Grande – MT, de outubro de 2015 a janeiro de 2016. O delineamento experimental foi inteiramente casualizado com cinco tratamentos compostos pelas concentrações de BAP (0,0; 3,0; 6,0; 9,0 e 12,0 mg L-1) em duas matrizes de aroeira (M2 e M3), com cinco repetições de cinco explantes cada. Foram realizados dois ciclos de repicagem com intervalo de 30 dias cada e as avaliações ocorreram ao final de cada ciclo. Foram feitas duas avaliações (av1 e av2): av1= número de brotações e altura da parte aérea (cm) e av2= número de brotações, altura da parte aérea (cm) peso da matéria fresca total (mg). Na matriz M2 foi feita apenas a avaliação 1, a maior média de brotações (3) e altura (1,38 cm) foram obtidas com 6,0 e 0 mg L-1 de BAP respectivamente. Na M3 foram feitas as duas avaliações, com média de brotações de 8,6 e 13,6 em 3,0 e 6,0 mg L-1 de BAP respectivamente, na primeira e segunda avaliações. Para altura da parte aérea as maiores médias de 1,85 (avaliação 1) e 1,56 cm (avaliação 2) foram obtidas sem o uso do BAP e o peso da matéria fresca foi superior na concentração de 3,0 mg L-1. A concentração de 3,0 mg L-1 de BAP favoreceu o desenvolvimento das brotações em matrizes de aroeira-do-sertão (M. urundeuva All.).
Palavras-chave: Anacardiaceae, citocinina, regulador vegetal.

82
MULTIPLICATION in vitro OF Myracrodruon urundeuva Allemão IN BAP (6-
BENZYLAMINOPURINE) CONCENTRATION
ABSTRACT - Cytokinins are essential for the effect in breaking the apical dominance, the proliferation of axillary buds and influence the success of in vitro multiplication. The objective was to verify the effect of different concentrations of BAP (6-benzylaminopurine) in the induction of shoots in vitro of matrices Myracrodruon urundeuva Allemão. The experiment was conducted at the Regional Center of the Tissue Culture Laboratory of Research and Technology Transfer (CRPTT) of EMPAER-MT, in the municipality of Várzea Grande - MT, from October 2015 to January 2016. The experimental design it was completely randomized with five treatments comprised of BAP (0,0; 3,0; 6,0; 9,0 and 12,0 mg L-1) in two matrices of M. urundeuva All. (M2 and M3), with five repetitions every five explants. The nodal segments used as explants were obtained from in vitro germination. Pricking two cycles were performed with 30 days apart each and evaluations occurred at the end of each cycle. Two evaluations were made of the variables: number of shoots and shoot height (cm) in the evaluation 1 and number of shoots, shoot height (cm) weight of the total fresh weight (mg) in the evaluation 2. In the M2 matrice was made only the evaluation 1, the highest average shoot (3) and height (1,38 cm) were obtained with 6,0 and 0 mg L-1 BAP respectively. In the M3 in the two evaluations were made, averaging shoots 8,6 and 13,6 at 3,0 and 6,0 mg L-1 BAP respectively, in the first and second evaluations. To shoot height the highest average of 1,85 (evaluation 1) and 1,56 cm (review 2) were obtained without the use of BAP and the fresh weight was higher in the concentration of 3,0 mg L-1. The concentration of 3,0 mg L-1 BAP favored the development of shoots in matrices of M. urundeuva All.
Keywords: Anacardiaceae, cytokinin, phytoregulator.

83
6.1 Introdução
A micropropagação é um processo, no qual pequenos fragmentos de tecido
vegetal são isolados dos organismos ou obtidos a partir destes, sendo cultivados
assepticamente em meio de cultura apropriado em condições controladas
(Grattapaglia e Machado, 1998; Amaral e Silva, 2003). Na fase de multiplicação, o
principal objetivo é produzir o maior número de plantas em menor espaço de tempo.
Partindo-se do pressuposto de que o sucesso do cultivo in vitro depende de fatores
associados à indução da morfogênese e da organogênese (regeneração de brotos e
raízes), se faz necessário que o meio nutritivo forneça substâncias essenciais para o
crescimento da planta in vitro (Cordeiro et al., 2004; Morais et al., 2014).
Para Gaspar et al. (1996) o fator chave nesta fase é a presença dos
reguladores de crescimento vegetal, especialmente aqueles dos grupos das
citocininas e auxinas. Segundo Staden et al. (2008), a interação entre os
fitorreguladores pode ocorrer de maneira sinérgica ou antagonicamente, a depender
de inúmeras circunstâncias. Afirmaram ainda que a presença de um determinado
regulador pode afetar a biossíntese ou metabolismo de outro, alterando os níveis
das substâncias endógenas, ou ainda os fatores ambientais inerentes ao processo
podem modificar as respostas dos reguladores ou, até mesmo, inativá-los.
As citocininas são fundamentais na fase de multiplicação da
micropropagação, promovendo nas plantas divisão (citocinese), alongamento e
diferenciação celular (Brum et al., 2002; Pasqual, 2004). Além disso, controlam a
expressão de genes envolvidos na senescência de tecidos e órgãos vegetais,
inibindo ou retardando os efeitos fisiológicos degenerativos, resultando no aumento
da longevidade celular (Morgante e Lombardi, 2004).
O tipo de citocinina e sua concentração influenciam no sucesso da
multiplicação in vitro, pois a necessidade dessas substâncias varia de acordo com a
espécie a ser trabalhada e de seus níveis hormonais endógenos (Grattapaglia e
Machado, 1998). A 6-benzilaminopurina (BAP) dentro dessa classe, é o
fitorregulador sintético que tem se mostrado mais eficaz na multiplicação de diversas
espécies lenhosas, além disso, ainda proporciona o menor custo de aquisição (Brum
et al., 2002; Cordeiro et al., 2004; Fráguas et al., 2008; Aragão et al., 2011). Na
maioria das plantas superiores, a dominância apical promovida pela ação da auxina
estimula o crescimento da gema apical inibindo o crescimento das gemas laterais.

84
As citocininas executam um papel importante favorecendo a quebra da dominância
apical, a expansão foliar e a indução da proliferação de brotações adventícias
(Macêdo et al., 2003; Pozo et al., 2005; Nascimento et al., 2008).
Diante do exposto, objetivou-se verificar o efeito de diferentes concentrações
de BAP (6-benzilaminopurina) na indução de brotações in vitro de matrizes de
aroeira-do-sertão (Myracrodruon urundeuva Allemão).
6.2 Material e Métodos
O experimento foi realizado no Laboratório de Cultura de Tecidos do Centro
Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da Empresa Mato-
grossense de Pesquisa, Assistência e Extensão Rural (EMPAER-MT) localizado no
município de Várzea Grande – MT, no período de outubro de 2015 a janeiro de
2016.
O delineamento experimental foi inteiramente casualizado com cinco
tratamentos compostos pelas concentrações de BAP (0,0; 3,0; 6,0; 9,0 e 12,0 mg L-
1) no meio de cultura MS em duas matrizes de aroeira (M2 e M3), com cinco
repetições de cinco explantes cada.
O experimento iniciou-se a partir da germinação de sementes in vitro de duas
matrizes de aroeira-do-sertão para obtenção de plântulas que serviram como fonte
de explante. As sementes foram coletadas em matrizes posicionadas a uma
distância de cerca de 30 metros entre si, em uma área de reserva pertencente a
uma empresa comercial situada na Estrada da Guia, Rodovia MT 010, KM 27 -
Cuiabá-MT, na segunda quinzena de setembro de 2014. Ocorreu precipitação na
região dez dias antes da coleta, no dia o tempo estava ensolarado com temperatura
média registrada de 36°C.
Primeiramente foi feita a assepsia nas sementes para inoculação em meio de
cultura MS (Murashige e Skoog, 1962) suplementado com 30 g L-1 de sacarose,
solidificado com 2,5 g L-1 de Phytagel®, pH ajustado para 5,8± 0,2 contendo as
concentrações de BAP testadas. Os frascos foram então vedados com plástico filme
para evitar possíveis contaminações, e levados para sala de crescimento,
posicionados aleatoriamente em uma prateleira de metal. A temperatura média desta
sala foi de 25 ±2°C e fotoperíodo de 16 horas, sendo a intensidade luminosa de 25

85
mol m-2 s-1 (medida em luxímetro) fornecida por lâmpadas do tipo fluorescente
branca fria.
Aos 30 dias após a semeadura, os frascos foram levados para a câmara de
fluxo laminar, as plântulas foram retiradas de dentro dos frascos com o auxílio de
pinças e repicadas sob uma superfície de papel de 15x15 cm utilizando-se bisturis,
sendo todo material empregado nesse procedimento previamente esterilizado.
No processo de repicagem as plântulas foram manipuladas como
microestacas, sendo a raiz eliminada e as plântulas cortadas em porções localizadas
nos entrenós contendo gemas axilares, denominadas segmentos nodais. Em
seguida os explantes, com cerca de 1,0 cm, foram inoculados, posicionados
próximos à borda do frasco para facilitar na posterior avaliação, em meio de cultura
MS suplementado com as doses descritas de BAP, compondo cinco frascos para
cada dose contendo cinco explantes cada, para cada matriz. Os frascos foram então
vedados com plástico filme, levados para sala de crescimento a 25 ±2°C e
fotoperíodo de 16 horas.
Foram realizados dois ciclos de repicagem com intervalo de 30 dias. Aos 30
dias após a primeira repicagem (2015) avaliaram-se as seguintes variáveis: número
de brotações e altura da parte aérea (cm). A altura da parte aérea foi considerada a
partir da base da brotação até o ápice, medida com régua milimetrada através do
frasco de vidro para não destruir o explante. Em seguida foi realizado novamente
todo o procedimento de repicagem descrito anteriormente.
Aos 30 dias após a segunda repicagem (2016) foram avaliados número de
brotações, altura da parte aérea (cm) e massa da matéria fresca (mg). Os dados
foram submetidos à análise de variância e as médias comparadas pela regressão.
Foi utilizado o programa estatístico Assistat versão 7.7 Beta (Silva, 2011).
6.3 Resultados e Discussão
Foi feita apenas a primeira avaliação na matriz M2, pois os explantes não se
desenvolveram após a segunda repicagem, foi observada alta incidência bacteriana
que prejudicou o crescimento nesta etapa. Na M3 foram realizadas as duas
avalições conforme proposto na metodologia. Isto mostra a importância de se
estabelecer as condições de cultivo não só para diferentes espécies ou gêneros,
mas também para diferentes indivíduos dentro de uma espécie.

86
De acordo com a análise de variância para número de brotações e altura da
parte aérea (cm) houve diferença significativa (p < 0,01) na primeira avaliação para a
matriz M1 e em M2 nas duas avaliações para ambas as variáveis.
Para o número de brotações na matriz M1 o ajuste quadrático da curva de
regressão apontou valores crescentes até o tratamento com 6,0 mg L-1 de BAP com
média de 3 brotações, seguido de decréscimo em concentrações maiores (Figura 1).
FIGURA 1. Número de brotações de explantes da matriz M2 de aroeira-do-sertão
(M. urundeuva All.), inoculados em meio de cultura com diferentes
concentrações de BAP (mg L-1) (CV= 36,76%; Cuiabá-MT, 2015).
Na matriz M3 houve aumento no número de brotações em relação às duas
avaliações realizadas, sendo a primeira com média de 8,6 e a segunda com 13,6
brotações, respectivamente para as concentrações de 3,0 e 6,0 mg L-1 de BAP
(Figura 2). Vale ressaltar que em todas as concentrações utilizadas de BAP,
inclusive na sua ausência, observou-se formação de brotações, sugerindo ação de
reservas endógenas.
y = - 0,0079x2 + 0,0619x + 2,7771 R² = 0,8242
0
0,5
1
1,5
2
2,5
3
3,5
0 3 6 9 12Núm
ero
de
bro
taçõ
es
Ma
triz
2
Concentrações de BAP (mg L-1)
Avaliação 1

87
FIGURA 2. Número de brotações de explantes da matriz M3 de aroeira-do-sertão
(M. urundeuva All.), inoculados em meio de cultura com diferentes
concentrações de BAP (mg L-1) (CVav1= 21,51% CVav2= 13,77%;
Cuiabá-MT, 2015 e 2016).
Muitos fatores podem influenciar o processo de indução organogênica, como
a concentração e os tipos de reguladores vegetais, a idade do explante, o genótipo,
as condições físicas do cultivo, a composição e consistência do meio de cultura
(Dobránszki e Silva, 2010).
Diferentemente do resultado deste estudo, Andrade et al. (1998) verificaram
que independente da concentração de BAP, ocorreu desenvolvimento de M.
urundeuva All. em brotação única, sendo 97 e 99% de explantes a partir dos
segmentos nodal e apical, respectivamente. Em pesquisa com cerejeira (Amburana
acreana) Fermino Junior e Scherwinski-Pereira (2012) observaram aumento no
número de brotos até a concentração de 4,0 mg L-1 de BAP e, acima desta
concentração, ocorreu efeito inibitório. Similarmente em sansão-do-campo (Mimosa
caesalpiniifolia), a adição de BAP ao meio de cultura em concentrações de até 4,0
mg L-1 ampliou o número de brotações por explante, seguido por decréscimo
mediante o incremento na concentração do regulador vegetal (Bezerra et al., 2014).
A definição do tipo e da concentração ótima de citocinina para a multiplicação
in vitro constitui um passo importante e dependerá do genótipo da planta; o seu uso
tende a estimular uma maior produção de brotos e alta taxa de multiplicação (Lemos
y = -0,0698x2 + 0,9048x + 5,4229 R² = 0,752
y = -0,1698x2 + 1,8448x + 8,4229 R² = 0,9963
0
2
4
6
8
10
12
14
16
0 3 6 9 12
Nú
me
ro d
e b
rota
çõe
s M
atr
iz 3
Concentrações de BAP (mg L-1)
Avaliação 1 Avaliação 2
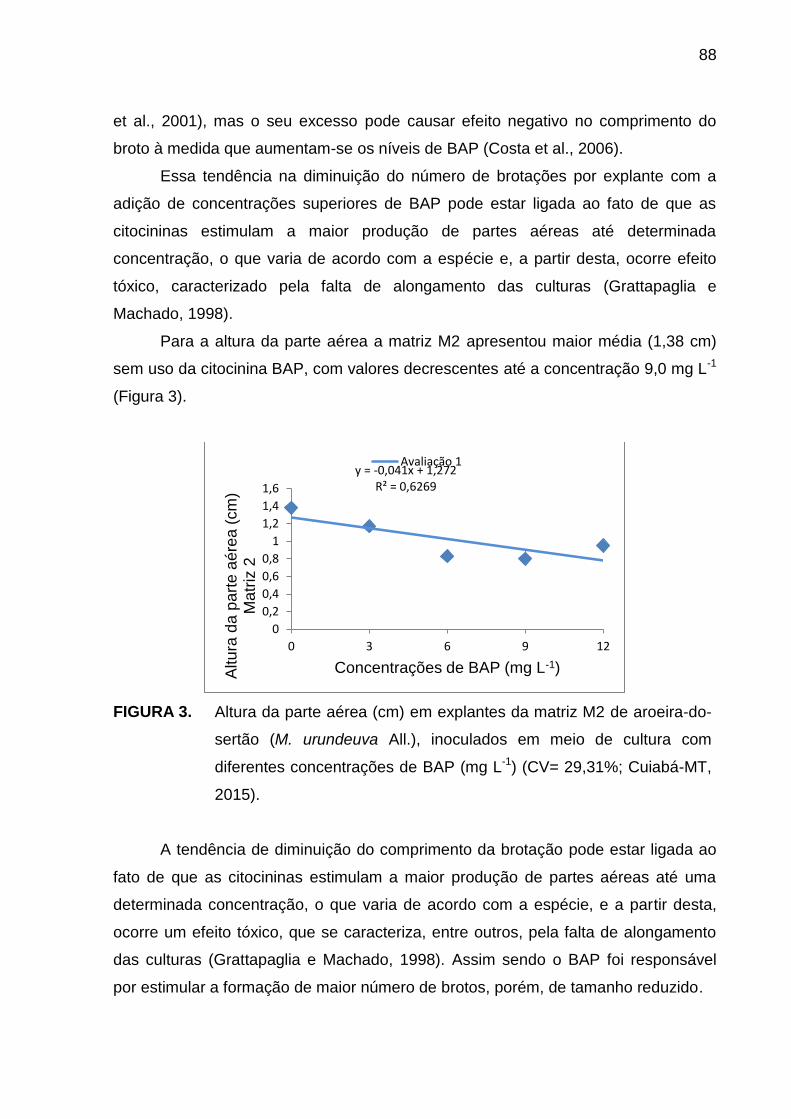
88
et al., 2001), mas o seu excesso pode causar efeito negativo no comprimento do
broto à medida que aumentam-se os níveis de BAP (Costa et al., 2006).
Essa tendência na diminuição do número de brotações por explante com a
adição de concentrações superiores de BAP pode estar ligada ao fato de que as
citocininas estimulam a maior produção de partes aéreas até determinada
concentração, o que varia de acordo com a espécie e, a partir desta, ocorre efeito
tóxico, caracterizado pela falta de alongamento das culturas (Grattapaglia e
Machado, 1998).
Para a altura da parte aérea a matriz M2 apresentou maior média (1,38 cm)
sem uso da citocinina BAP, com valores decrescentes até a concentração 9,0 mg L-1
(Figura 3).
FIGURA 3. Altura da parte aérea (cm) em explantes da matriz M2 de aroeira-do-
sertão (M. urundeuva All.), inoculados em meio de cultura com
diferentes concentrações de BAP (mg L-1) (CV= 29,31%; Cuiabá-MT,
2015).
A tendência de diminuição do comprimento da brotação pode estar ligada ao
fato de que as citocininas estimulam a maior produção de partes aéreas até uma
determinada concentração, o que varia de acordo com a espécie, e a partir desta,
ocorre um efeito tóxico, que se caracteriza, entre outros, pela falta de alongamento
das culturas (Grattapaglia e Machado, 1998). Assim sendo o BAP foi responsável
por estimular a formação de maior número de brotos, porém, de tamanho reduzido.
y = -0,041x + 1,272 R² = 0,6269
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
0 3 6 9 12
Altu
ra d
a p
art
e a
ére
a (
cm
) M
atr
iz 2
Concentrações de BAP (mg L-1)
Avaliação 1

89
Para a matriz M3 o comprimento da parte aérea também foi maior sem o uso
do BAP com médias de 1,85 e 1,56 cm, respectivamente para as avaliações 1 e 2,
apresentando redução a medida de aumentou-se a concentração de BAP (Figura 4).
FIGURA 4. Altura da parte aérea em explantes da matriz M3 de aroeira-do-sertão
(Myracrodruon urundeuva Allemão), inoculados em meio de cultura
com diferentes concentrações de BAP (mg L-1) (CVav1= 20,10%
CVav2= 23,55%; Cuiabá-MT, 2015 e 2016).
Assim como observado neste estudo, em resultados obtidos por Ayub et al.
(2010), o meio de cultura na ausência de BAP proporcionou maiores brotações (1,7
cm) de videira cv. Bordô, afirmaram ainda que a citocinina presente nos demais
tratamentos foi prejudicial ao desenvolvimento da parte aérea. Com alecrim [Lippia
alba (Mill.) N.E. Brown] foram observados maiores brotos (18,86 cm) na ausência de
BAP, ocorrendo redução significativa a medida que aumentou-se a concentração.
Estes resultados podem estar relacionados ao fato deste regulador de crescimento
não ser responsável em promover o alongamento de brotações (Asmar et al., 2012).
Ao estudar o efeito de BAP em porta-enxerto de videira (Vitis spp. L.) „P1103‟,
Villa et al. (2007) observaram que o comprimento da parte aérea atingiu o valor
máximo (3,02 g) utilizando 2,0 mg L-1 de BAP e a partir dessa dose ocorreu
decréscimo no comprimento. Do mesmo modo, Fermino Junior e Scherwinski-
Pereira (2012) obtiveram a maior altura dos brotos de cerejeira (Amburana acreana)
nos tratamentos 0,0 ou com 0,25 mg L-1 de BAP, e observaram que maiores
y = -0,0653x + 1,908 R² = 0,9534
y = -0,0637x + 1,388 R² = 0,8123
0
0,5
1
1,5
2
2,5
0 3 6 9 12
Altu
ra d
a p
art
e a
ére
a (
cm
) M
atr
iz 3
Concentrações de BAP (mg L-1)
Avaliação 1 Avaliação 2

90
concentrações aplicadas exogenamente tendem a desenvolver brotos com menor
altura.
Na multiplicação in vitro de pau-pombo (Tapirira guianensis) Gutiérrez et al.
(2013) verificaram maiores médias para comprimento do maior broto de 10,22 e 9,5
mm em meio de cultura acrescido de 0,92 e 1,15 mg L-1 de BAP (valor estimado
pelas equações quadráticas). Notaram ainda a formação de brotos mesmo na
ausência de reguladores vegetais e perceberam que incrementos de BAP, em
concentrações superiores desfavoreceram a formação das brotações. Vale ressaltar
que elevadas concentrações de citocininas podem reduzir o tamanho das brotações
no cultivo in vitro e estimular a ocorrência de hiperidricidade e formação de folhas
anormais (Pasqual, 2001; Machado et al., 2006).
Como não houve desenvolvimento da matriz M2 após a segunda repicagem,
a massa da matéria fresca foi realizado apenas na M3 o maior peso (1,121 mg) foi
observado com 3,0 mg L-1 de BAP, decrescendo nas demais concentrações. O
ponto de máxima observado foi de 5,69 mg L-1 (Figura 5).
FIGURA 5. Massa da matéria fresca de brotações da matriz M3 de aroeira-do-
sertão (M. urundeuva All.), inoculados em meio de cultura com
diferentes concentrações de BAP (mg L-1) (CV= 21,51%; Cuiabá-MT,
2016).
Esse efeito pode ter ocorrido provavelmente devido às plantas conseguirem
absorver mais água para seus tecidos nesta concentração de regulador resultando
em incremento na massa fresca. A redução do peso da matéria fresca em
y = -0,0129x2 + 0,1469x + 0,4655 R² = 0,528
0
0,2
0,4
0,6
0,8
1
1,2
0 3 6 9 12
Ma
ssa
da
ma
téria
fre
sca
(m
g)
Ma
triz
3
Concentrações de BAP (mg L-1)

91
concentrações maiores da citocinina foi verificada também por Asmar et al. (2012)
em pesquisa com alecrim [Lippia alba (Mill.) N.E.Brown] com maiores valores da
massa fresca de parte aérea (1,07 g) utilizando-se 0,5 mg L-1 de BAP, decrescendo
até a concentração máxima de 1,5 mg L-1.
Em estudo com oliveira (Olea europaea L.) Dutra et al. (2004) obtiveram a
maior média para matéria fresca (0,14 g) da parte aérea na ausência de BAP, com
subsequente redução à medida que se aumentou a concentração (1, 2 e 4 mg L-1)
desse regulador de crescimento. Para os autores essa resposta pode ter ocorrido
devido a um desbalanço na relação auxina/citocinina. Em físalis (Physalis peruviana
L.) foi observada maior massa fresca (0,43g) na ausência de BAP juntamente com a
adição de 75% dos sais do meio MS, neste caso foi sugerido que o aumento na
massa obtida deve-se ao maior comprimento médio dos brotos para esse mesmo
tratamento (Rodrigues et al., 2013).
Com Araújo et al. (2004) não foi observado efeito significativo nas
concentrações de BAP testadas (0, 1, 2 e 4 mg L-1) em gloxínia (Sinningia speciosa
L.H.) para o peso da matéria fresca. Assim como com Villa et al. (2010) ao avaliar o
efeito de concentrações 6-benzilaminopurina (0; 0,5; 1,0; 2,0 e 4,0 mg dm-3)
adicionados à diversos meios de cultura na multiplicação in vitro de amoreira-preta
(Rubus sp.) cultivares Tupy e Brazos.
De modo geral, nas variáveis analisadas concentrações elevadas de BAP
promoveram redução no desenvolvimento em M. urundeuva All. Essa citocinina
apesar de ter um efeito mais expressivo na multiplicação in vitro de diversas
espécies quando comparado às outras, segundo alguns autores pode apresentar
fitotoxicidade quando utilizado em concentrações não adequadas, acarretando em
efeito negativo (Grattapaglia e Machado, 1998; Pasqual, 2001).
6.4 Conclusão
A concentração de 3,0 mg L-1 de BAP favoreceu o desenvolvimento de
brotações em matrizes de aroeira-do-sertão (M. urundeuva All.).
6.5 Referências Bibliográficas ANDRADE, M.W.; LUZ, J.M.Q.; LACERDA, A.S.; MELO, P.R.A. Micropropagação da aroeira (Myracrodruon urundeuva Fr. All.). Ciência e Agrotecnologia, Lavras, v.24, n.1, p.174-180, 2000.

92
AMARAL, C.L.F.; SILVA, A.B. Melhoramento biotecnológico de plantas medicinais: produção de alcalóides e óleos essenciais. Revista Biotecnologia Ciência e Desenvolvimento, Rio de Janeiro, n.30, p.55-59, 2003. ARAGAO, A.K.O.; ALOUFA, M.A.I.; COSTA, I.A. O efeito do bap (6-benzilaminopurina) sobre a indução de brotos em explantes de pau-brasil. Cerne, Lavras, v.17, n.3, p.339-345, 2011. ARAÚJO, A.G.; FIORINI, C.V.A.; PASQUAL, M.; SILVA, A.B.; VILLA, F. Multiplicação in vitro de gloxínia (Sinningia speciosa LOOD. HIERN.). Revista Ceres, Viçosa, v.51, n.253, p.117-127, 2004. ASMAR, S.A.; RESENDE, R.F.; ARARUNA, E.C.; MORAIS, T.P.; LUZ, J.M.Q. Concentrações de BAP sobre a proliferação in vitro de brotos de Lippia alba [(Mill.) N.E.Brown]. Revista Brasileira de Plantas Medicinais, Botucatu, v.14, n.esp., p.149-153, 2012. AYUB, R.A.; SPINARDI, B.; BASSO, M.F.; BIASI, L.A. Indução de multibrotação in vitro em videira cv. Bordô. Revista Brasileira de Fruticultura, Jaboticabal, v.32, n.3, p.675-681, 2010. BEZERRA, R.M.F.; ALOUFA, M.A.I.; FREIRE, F.A.M.; SANTOS, D.D. Efeito de 6-benzilamnopurina sobre a propagação in vitro de Mimosa caesalpiniifolia Benth. (Fabaceae). Revista Árvore, Viçosa, v.38, n.5, p.771-778, 2014. BRUM, G.R.; SILVA, A.B.; PASQUAL, M. Efeito de diferentes concentrações de BAP e ANA na propagação in vitro de figueira (Ficus carica L.). Ciência e agrotecnologia, Lavras. Edição Especial, p.1403-1409, 2002. CALDAS, I.S.; HARIDASON, P.; FERREIRA, M.E. Meios Nutritivos. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa – SPI / Embrapa – CNPH, 1998, v.1, p.87-132. CORDEIRO, I.M.C.C.; LAMEIRA, O.L.; OHASHI, S.T.; ROSAL, L.F. Efeito de BAP sobre a proliferação de brotos in vitro de Schizolobium amazonicum Huber ex Ducke (paricá). Cerne, Lavras v.10, n.1, p.118-124, 2004. COSTA, F.H.S.; PEREIRA, J.E.S.; PEREIRA, M.A.A.; OLIVEIRA, J.P. Efeito da interação entre carvão ativado e N6-benzilaminopurina na propagação in vitro de bananeira, cv. Grand Naine (AAA). Revista Brasileira de Fruticultura, Jaboticabal, v.28, n.2, p.280-283, 2006. DOBRÁNSZKI J.; SILVA J.A.T. Micropropagation of apple - A review. Biotechnology Advances, v.28, n.4, p.462-488, 2010. DUTRA, L.F.; OLIVEIRA, A.F.; FRÁGUAS, C.B.; PASQUAL, M. Multiplicação in vitro de oliveira (Olea europaea L.). Ciência e Agrotecnologia, Lavras, v.28, n.1, p.220-223, 2004.

93
FERMINO JUNIOR, P.C.P.; SCHERWINSKI-PEREIRA, J.E. Germinação e propagação in vitro de cerejeira (Amburana acreana (Ducke) A.C. Smith – Fabaceae). Ciência Florestal, Santa Maria, v.22, n.1, p.1-9, 2012. FRÁGUAS, C.B.; DORNELLES, C.M.V.; LIMA, G.P.P. Benzilaminopurina e Ácido naftaleno acético na indução e multiplicação in vitro de gemas de abacaxizeiro da cultivar „IAC Gomo-de-mel‟. Ciência Rural, Santa Maria, v.39, n.6, p.1682-1687, 2008. GASPAR, T.; KEVERS, C.; PENEL, C.; GREPIN, H.; REID, D. M.; THORPE, T. A. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell Development Biology – Plant, Columbia, v. 32, p. 272-289, Oct./Dec. 1996. GRATTAPAGLIA, D.; MACHADO, M.A. Micropropagação. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa – SPI / Embrapa – CNPH, 1998, v.1, p.183-260. GUTIERREZ, I.E.M.; NEPOMUCENO, C.F.; SILVA, T.S.; FONSECA, P.T. et al. Multiplicação in vitro de Tapirira guianensis Aubl. (Anacardiaceae). Ceres, v.60, n.2, p.143-151, 2013. LEMOS, E.E.P.; FERREIRA, M.S.; ALENCAR. L.M.C.; OLIVEIRA, J.G.L.; MAGALHÃES, V.S. Micropropagação de clones de banana cv. terra em biorreator de imersão temporária. Revista Brasileira de Fruticultura, Jaboticabal, v.23, n.3, p.482-487, 2001. LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Nova Odessa-SP. Instituto Plantarium, 1992, 352p. MACÊDO, C.E.C.; SILVA, G.M.; NÓBREGA, F.S.; MARTINS, C.P. Concentrações de ANA e BAP na micropropagação de abacaxizeiro L. Merrill (Ananas comosus) e no cultivo hidropônico das plântulas obtidas in vitro. Revista Brasileira de Fruticultura, Jaboticabal, v.25, n.3, p.501-504, 2003. MACHADO, M.P.; BIASI, L.A.; RITTER, M.; RIBAS, L.L.F.; KOEHLER, H.S. Multiplicação in vitro do porta-enxerto de videira VR 043-43 (Vitis vinifera x Vitis rotundifolia). Ciência e Agrotecnologia, Lavras, v. 30, n. 4, p. 648-655, 2006. MORAIS, T.P.; ASMAR, S.A.; LUZ, J.M.Q. Reguladores de crescimento vegetal no cultivo in vitro de Mentha x Piperita L. Revista Brasileira de Plantas Medicinais, Campinas, v.16, n.2, p.350-355, 2014. MORGANTE, C.V.; LOMBARDI, S.P. Hormônios vegetais e biotecnologia. Piracicaba: Esalq, 2004, 13p. MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiologia Plantarum, Copenhagen, v.15, p.473-97, 1962.

94
NASCIMENTO, A.C.; PAIVA, R.; ABBADE, L.C.; VARGAS, D.P.; SOARES, F.P. Micropropagação de uvaieira (Eugenia pyriformis Cambess): efeitos do BAP e AIB. Revista Verde, Mossoró, v.3, n.2, p.20-26, 2008. PASQUAL, M. Propagação de plantas ornamentais. Lavras: UFLA/FAEPE, 2004. 106p. PASQUAL, M. Cultura de tecidos. Lavras: FAEPE/UFLA, 2001. POZO, J. C. D.; LOPEZ-MATAS, M. A.; RAMIREZ-PARRA, E.; GUTIERREZ, C. Hormonal control of the plant cell cycle. Biologia Plantarum, Czech Republic, v. 123, p. 173-183, 2005. RODRIGUES, F.A.; PENONI, E.S.; SOARES, J.D.R.; PASQUAL, M. Diferentes concentrações de sais do meio MS e BAP na multiplicação in vitro de Physalis peruviana L. Biosciene Journal, Uberlândia, v.29, n.1, p.77-82, 2013. SILVA, F.A.S. ASSISTAT versão 7.7 beta. Campina Grande-PB: Assistência Estatística, Departamento de Engenharia Agrícola do CTRN - Universidade Federal de Campina Grande, Campus de Campina, 2011. STADEN J., ZAZIMALOVA E.; GEORGE E.F. Plant growth regulators II: Cytokinins, their analogues and antagonists. In: GEORGE E.F., HALL M.A; DE KLERK G.J. (Eds.) Plant propagation by tissue culture. 3ª ed. Dordrecht, Springer. p.205-226, 2008. VILLA, F.; PASQUAL, M.; PIO, L.A.S.; ASSIS, F.A.; TEODORO, G.S. Influência do carvão ativado e BAP na multiplicação in vitro de duas frutíferas de clima temperado. Ceres, Lavras, v.54, n.352, p.118-124, 2007. VILLA, F.; PASQUAL, M.; SOUZA, A.G; VILELA, X.M.S. Meios de cultura e reguladores de crescimento na multiplicação in vitro de amoreira-preta. Scientia Agraria, Curitiba, v.11, n.2, p.109-117, 2010.

95
7 ÁCIDO INDOLBUTÍRICO NO ENRAIZAMENTO ex vitro DE Myracrodruon
urundeuva Allemão (AROEIRA-DO-SERTÃO)
RESUMO – No enraizamento ex vitro, as plantas micropropagadas são estimuladas a formar raízes rapidamente para absorver os nutrientes necessários à sua sobrevivência na fase de aclimatação. Objetivou-se testar diferentes substratos e concentrações de ácido indolbutírico no enraizamento ex vitro de aroeira-do-sertão (M. urundeuva All.). O experimento foi realizado no viveiro do curso de Agronomia do Univag Centro Universitário, no município de Várzea Grande – MT, de janeiro a fevereiro de 2016. O delineamento experimental foi inteiramente casualizado em esquema fatorial 2x5, sendo dois tipos de substrato (Tecnomax® e Vivatto®) e cinco concentrações de ácido indolbutírico [AIB] (0; 500; 1000; 1500 e 2000 mg L-1), com três repetições de três explantes cada. As miniestacas foram produzidas in vitro e passaram por dois ciclos de repicagem. Posteriormente foram retiradas dos frascos e levadas ao viveiro onde tiveram suas bases imersas nas soluções contendo o fitorregulador, foram transplantadas em bandejas de poliestireno expandido. As bandejas possuiam 128 células, com volume de 34,6 cm3 e medindo 3,5x3,5x5,0 cm, contendo os substratos testados. Aos 20 foram avaliadas a sobrevivência (%), comprimento da parte aérea (cm) e da raiz (cm), diâmetro do colo (cm) e peso da matéria fresca (mg). Apenas o tratamento utilizando o substrato Vivatto® e 1,5 g L-1 de AIB teve a sobrevivência de dois indivíduos. Verificou-se que as microplantas que morreram não haviam desenvolvido raízes, impossibilitando dar sequência às análises. A taxa de mortalidade (97,9%) pode ser atribuída a ausência de desenvolvimento radicular e ainda às mudanças drásticas nas condições ambientais sofridas pelas plântulas.
Palavras-chave: aclimatização, auxina, fitorregulador, rizogênese.

96
INDOLBUTYRIC ACID IN ROOTING ex vitro OF Myracrodruon urundeuva Allemão
ABSTRACT - In the ex vitro rooting, as micropropagated plants are encouraged to Form paragraph roots quickly absorb the nutrients necessary for their survival in the acclimatization phase. This study aimed to test different substrates and indolbutyric acid concentrations on rooting ex vitro of Myracrodruon urundeuva All. The experiment was held in Agronomy course nursery of UNIVAG Centro Universitário in the city of Várzea Grande - MT, from January to February 2016. The experimental design was completely randomized in a factorial 2x5 scheme, being two types of the substrate (Tecnomax® and Vivatto®) and five concentrations of indolbutyric acid [AIB] (0, 500, 1000, 1500 and 2000 mg L-1), with three repetitions three explants each. As minicuttings were produced in vitro and passed by two pricking cycles. He was subsequently removed from bottles and taken the nursery Where were immersed in solutions bases containing the plant regulator, Were transplanted into expanded polystyrene trays. As trays had 128 cells, with volume 34,6 cm3 and measuring 3,5x3,5x5,0 cm, containing the substrates tested. At 20 were evaluated survival (%), shoot length (cm) and root (cm), stem diameter (cm) and weight of fresh matter (mg). Only the treatment using the Vivatto® substrate and 1,5 g L-1 IBA had a survival two individuals. It was as microplants que what not died had developed roots, making it impossible to follow up the analyzes. The mortality rate (97,9%) it can be attributed to lack of root development and the drastic changes in environmental conditions experienced by seedlings.
Keywords: acclimatization, auxin, plant regulator, root formation.

97
7.1 Introdução
A fase de aclimatização das plantas oriundas da micropropagação é apontada
como uma das mais críticas do processo de regeneração, em que pode ocorrer a
maioria das perdas (Grattapaglia e Machado, 1998). Um dos fatores importantes na
produção de mudas, é o tipo de substrato (Paiva Sobrinho et al., 2010), pois suas
propriedades podem facilitar ou impedir o desenvolvimento das plantas (Calvete et
al., 2000; Rocha et al., 2008).
O substrato tem o papel fundamental de fornecer às mudas condições
químicas, físicas e biológicas, para um crescimento saudável, oferecendo assim
condições de transformar seu potencial genético em produtividade (Kämpf et al.,
2000). Possibilitar, com a quantidade adequada de espaço poroso, o suprimento de
oxigênio, permitindo a iniciação e a formação do sistema radicular (Smiderle et al.,
2001; Ferreira et al., 2015), bem como o desenvolvimento da parte aérea.
Por consequência, busca-se encontrar uniformidade em sua composição, ser
rico em nutrientes, com elevada capacidade de retenção de água e troca catiônica,
isento de pragas e patógenos, e viável economicamente (Melo et al., 2003; Pozza et
al., 2007). Conforme Fernandes et al. (2006) vários materiais podem ser utilizados
como substratos para plantas, dentre eles a vermiculita, turfa, casca de arroz, casca
de pinus, fibra da casca de coco, entre outros.
A transferência para o substrato de plantas enraizadas in vitro pode dificultar
a técnica, pois estas raízes são frágeis podem se quebrar, resultando em má
qualidade no transplantio e até a morte das plântulas. Portanto, o enraizamento ex
vitro é desejável do ponto de vista econômico (Yan et al., 2010), com redução nos
custos de obtenção da muda micropropagada (Pelizza et al., 2013). Além disso,
induzir o enraizamento na aclimatização beneficia na qualidade do sistema de
raízes, visto que, nesta fase este tende a ser mais completo e funcional
(Grattapaglia e Machado, 1998).
O processo de formação de raízes adventícias ocorre de uma a três semanas
e pode ser dividido em três fases: indução, iniciação e elongação. As fases de
indução e iniciação, geralmente são dependentes de auxina, mas o crescimento
(elongação) pode ser inibido pela presença desse hormônio (Grattapaglia e
Machado, 1998). A dificuldade encontrada em espécies lenhosas com relação à
capacidade de enraizamento, devido a maturidade da planta, pode ser em parte

98
resolvida no processo da micropropagação pelas contínuas subculturas que
promovem o rejuvenescimento dos tecidos (Pasqual, 2001).
Dentre as formas sintéticas comuns mais aplicadas na micropropagação,
estão o ácido 3-indolbutírico (AIB), 2,4-diclorofenoxiacético (2,4-D) e ácido
nafataleno acético (ANA) (Lopes et al., 2001), cujas concentrações variam de acordo
com a espécie. O AIB é a auxina mais utilizada para estimular o enraizamento dos
propágulos (Brondani et al., 2010; Dias et al., 2012; Machado et al, 2013; Tronco et
al., 2015), devido à baixa toxidez para a maioria das espécies, em uma larga faixa
de concentração (Iacona e Muleo, 2010). No entanto, a dose utilizada deve ser
adequada para cada espécie e tipo de propágulo (Titon et al., 2003). Com a
aplicação da auxina sintética, ocorre um aumento imediato no nível endógeno de
auxina natural e consequentemente dando início da formação de raízes primordiais
(Assis e Teixeira, 1998).
Diversos autores testaram o efeito dessa auxina no enraizamento em diversas
espécies arbóreas, tais como, canjarana [Cabralea canjerana (Vell.) Mart.] (Rocha et
al., 2007); cedro-australiano [Toona ciliata var. australis (F. Muell.) Bahadur] (Pereira
et al., 2015); Eucaliptus (Titon et al., 2003; Wendling; Xavier, 2005; Almeida et al.,
2007; Lana et al., 2008; Brondani et al., 2010; Brondani et al., 2014); louro-pardo
[Cordia trichotoma (Vell.) Arrab. ex Steud] (Kielse et al., 2013; Faganello et al.,
2015); teca [Tectona grandis L.f.] (Fermino Junior et al., 2011; Georgin et al., 2014;
Fernandes et al., 2015), entre outras.
Dessa forma, o objetivo neste trabalho foi testar diferentes substratos e
concentrações de ácido indolbutírico no enraizamento ex vitro de aroeira-do-sertão
(Myracrodruon urundeuva Allemão).
7.2 Material e Métodos
O experimento foi conduzido na casa de vegetação do curso de Agronomia do
Univag Centro Universitário, situado no município de Várzea Grande - Mato Grosso,
no período de janeiro a fevereiro de 2016.
O delineamento experimental foi inteiramente casualizado em esquema
fatorial 2x5, sendo dois tipos de substrato (Tecnomax® e Vivatto®) e cinco
concentrações de AIB (0; 500; 1000; 1500 e 2000 mg L-1), compondo 10 tratamentos

99
com três repetições de três explantes. Os substratos utilizados foram Bioplant® e
Tecnomax® cujas composições estão descritas na Tabela 1.
TABELA 1. Composição dos substratos comerciais utilizados no experimento
(Cuiabá-MT, 2016).
A produção de mudas in vitro foi realizada no Laboratório de Cultura de
Tecidos do Centro Regional de Pesquisa e Transferência de Tecnologia (CRPTT) da
Empresa Mato-grossense de Pesquisa, Assistência e Extensão Rural (EMPAER-
MT), localizado no município de Várzea Grande – MT. As plântulas utilizadas como
fonte de explante foram obtidas a partir da germinação in vitro de sementes que
foram coletadas em matrizes localizadas em uma área de reserva pertencente a
uma empresa comercial situada na Estrada da Guia, Rodovia MT 010, KM 27 -
Cuiabá-MT, na segunda quinzena de setembro de 2014. Ocorreu precipitação na
região dez dias antes da coleta, no dia o tempo estava ensolarado com temperatura
média registrada de 36°C.
Após a germinação, os explantes da matriz M3, com cerca de 1,0 cm, foram
subcultivados por dois ciclos de repicagem de 30 dias cada em meio de cultura MS
(Murashige e Skoog, 1962), suplementado com 30 g L-1 de sacarose, 3,0 mg L-1 de
BAP, solidificado com 2,5 g L-1 de Phytagel® e pH ajustado em 5,8 ±0,2. Aos 60 dias
de cultivo, os explantes com cerca de 2,5 cm foram retirados dos frascos com auxílio
de pinça, removeu-se o meio de cultura da base com água destilada e em seguida
foram acondicionados em caixa de isopor forrada com papel toalha umedecido.
Prontamente foram encaminhados para a casa de vegetação e manipulados como
microestacas.
Para a instalação do experimento na casa de vegetação, foram utilizadas
bandejas de poliestireno expandido com 128 células medindo 3,5 x 3,5 x 5,0 cm e
Substrato Composição
Bioplant® Casca de pinus, casca de arroz, pó de coco, vermiculita, macro e micronutrientes
Tecnomax® Vermiculita expandida, casca de pinus, cinza, fibra de coco e
carvão vegetal.

100
volume de 34,6 cm3 (por célula). Primeiramente foi efetuada a assepsia nas
bandejas com hipoclorito de sódio a 10% (cloro ativo 10% v/v) e álcool 70% (v/v).
Para melhor aderência os substratos foram umedecidos de acordo com a
capacidade máxima de retenção com água de poço artesiano e em seguida
colocados nas células. A determinação da capacidade de retenção foi feita de
acordo com as Regras para Análise de Sementes (Brasil, 2009), onde a quantidade
de água a ser adicionada ao substrato foi obtida pesando-se 500 g do material seco,
que foi colocado em filtro de papel, tipo coador de café comercial, em seguida,
adicionou-se 1000 ml de água. Após todo o excesso de água ser drenado, o volume
foi determinado por diferença da quantidade de água que ficou retida no substrato.
No preparo da solução hidroalcóolica de AIB, foram pesadas em balança
analítica as concentrações descritas anteriormente, em seguida dissolvidas em 50
ml de álcool etílico 95% e por último completou-se o volume para 100 ml com água
destilada (Yamamoto et al., 2010; Machado et al., 2013; Tronco et al., 2015). Os
explantes produzidos in vitro tiveram então sua base imersa nas soluções de AIB por
cinco segundos e, em seguida, foram transplantados nas bandejas de poliestireno
contendo os substratos. No tratamento testemunha (0,0 mg L-1 de AIB) os explantes
foram colocados em contato com água destilada apenas. Todos os tratamentos e
repetições foram devidamente identificados com etiquetas plásticas resistentes à
umidade.
As bandejas foram então levadas à casa de vegetação protegida com
sombrite a 50% de sombreamento (conforme fabricante), na qual permaneceram
suspensas em cavaletes a distância de 1,0 m do solo. A temperatura média foi de
cerca de 35ºC. A irrigação foi realizada manualmente com uso de proveta graduada
duas vezes ao dia, às 07 hrs e às 17 hrs. A quantidade de água fornecida variou
conforme a necessidade, entre 10 e 20 ml por dia por célula.
Aos 20 dias após a implantação do experimento, foram avaliadas as variáveis:
taxa de sobrevivência (%), comprimento da parte aérea (cm), comprimento da raiz
(cm), diâmetro do colo (cm) e massa fresca total [parte aérea + raiz] (mg). A taxa de
sobrevivência (%) foi determinada pela contagem de plantas vivas.
O comprimento da parte aérea (cm) foi determinado realizando-se a medição
a partir da base onde está inserida a raiz até o ápice da planta. Para o comprimento
da raiz (cm) foi considerada a maior raiz, medida a partir da base de inserção. O

101
diâmetro do colo (cm) foi demarcado na base da planta, acima do local de inserção
da raiz. Todas as medições foram realizadas com paquímetro digital. Por último, a
massa fresca total (mg) foi realizada com as três plantas por repetição pesadas
juntas em balança semi-analítica.
Os dados foram submetidos à análise de variância, as médias dos
tratamentos qualitativos foram comparadas pelo teste de Tukey a 5% de
probabilidade e as médias dos tratamentos quantitativos pela regressão. Foi
utilizado o programa estatístico Assistat versão 7.7 Beta (Silva, 2011).
7.3 Resultados e Discussão
As plantas da aroeira-do-sertão (M. urundeuva All.) apresentaram elevada
mortalidade (97,9%) nas mudas transplantadas aos 20 dias de cultivo, apenas o
tratamento utilizando o substrato Vivatto® e 1,5 g L-1 de AIB teve a sobrevivência
(Figura 1) de dois indivíduos. Foi verificado que as microplantas que morreram não
haviam desenvolvido sistema radicular, impossibilitando dar sequência às análises
estabelecidas na metodologia.
FIGURA 1. Aspecto das plantas sobreviventes aos vinte dias após a
transferência para o substrato (Cuiabá-MT, 2016).
É importante ressaltar que alguns dos explantes (seis) de aroeira-do-sertão
ainda no cultivo in vitro apresentaram raízes fibrosas e quebradiças que se
danificaram ao serem removidos dos frascos. Esses indivíduos foram colocados nos

102
substratos testados separadamente aos demais tratamentos, mas também não
houve sobrevivência. Segundo Sutter (1988), quando transferidas para a
aclimatização, as plantas enraizadas in vitro são submetidas a uma condição de alta
transpiração que, associada à alta condutividade hídrica, provoca baixa
funcionalidade ou ausência de controle sobre o fechamento dos estômatos. Durante
a aclimatação, além do estresse hídrico há outras adaptações importantes como a
passagem de um estado heterotrófico para um autotrófico, que podem levar muitas
mudas ao óbito (Torres et al., 1998).
A iniciação de raízes adventícias é estimulada por altos níveis de auxina que
estimulam a divisão celular até formar meristema apical da raiz (Taiz e Zeiger, 2009).
Conforme Damiani et al. (2009) é reconhecido que a capacidade de enraizamento é
variável com a maturidade do material a ser propagado, diferindo entre espécies e
entre genótipos. Fatores genéticos estão ligados direta ou indiretamente à
capacidade de determinadas espécies formar ou não raízes adventícias (Assis e
Teixeira,1998). Assim, a mortalidade pode ainda ser atribuída à baixa quantidade de
reserva para o crescimento ex vitro (Costa et al., 2008).
Em comparação ao aspecto das microplantas no dia da transferência
(21/01/2016) e aos sete dias após a transferência (Figura 2), a coloração foi se
diferenciando e a mortalidade foi intensificada a partir desse período, tanto com os
indivíduos que não tiveram contato com a auxina exógena como os demais, e
principalmente aqueles que estavam no substrato Tecnomax®. Foi observada a
sobrevivência geral de cerca de 70%, consideravelmente elevada até os sete dias.
FIGURA 2. Aspecto das plantas após a transferência para o substrato. A= dia da
transferência; B= sete dias após a transferência (Cuiabá-MT, 2016).

103
Dentre as alterações morfológicas quando as plantas são transferidas para a
casa de vegetação, destaca-se a perda da camada de cera epicuticular, que
minimiza a taxa de transpiração, consequentemente ocorre a rápida desidratação
dos tecidos (Grattapaglia e Machado, 1998).
Santos et al. (2009) avaliariam o processo morfofisiológico de mudas de
orquídeas (Cattleya labiata e Dendrobium anosmum) na aclimatização, as mesmas
foram transplantadas em vasos com capacidade de 120 ml, contendo substrato
(casca de Pinnus e carvão), com 2 pulverizações semanais de ANA durante 60 dias
e armazenadas sob telado com sombreamento de 70%. Observaram mortalidade
em todas as mudas de Cattleya labiata transplantadas aos 15 dias de cultivo. Em D.
anosmum o índice de mortalidade foi mais baixo que a C. labiata, mas as mudas só
sobreviveram até 45 dias após o transplantio.
Andrade et al. (2000) verificaram na aclimatação de aroeria-do-sertão (M.
urundeura All.) enraizada in vitro com ANA, apenas 30% de sobrevivência, cujo
plantio foi feito em copos plásticos (200 ml) contendo substrato Plantmax®, fechados
com saco de polietileno transparente (por 15 dias) e colocados em sombrite (50%) à
temperatura ambiente, até o plantio em vaso. Conforme os autores, a alta perda de
plantas provavelmente teve como causa o elevado índice de podridão do colo, que
indicou que o substrato não estava devidamente esterilizado e aliado à alta umidade
no ambiente do saco plástico, culminou na morte das plantas. Em um segundo
momento outras 15 plantas tiveram 80% de sobrevivência, justificada pela isenção
de patógenos no substrato e retirada do saco plástico aos 10 dias.
Em orquídea (Oncidium baueri) Sorace et al. (2007) fizeram a imersão das
plântulas em auxina e pulverizações semanais no substrato, o transplantio foi
realizado em bandejas de isopor contendo esfagno como substrato. A irrigação feita
manualmente duas vezes ao dia no verão, e uma vez ao dia no inverno.
Identificaram assim maior enraizamento e desenvolvimento vegetativo em plântulas
submetidas a pulverização com ANA (200 mg L-1), enquanto que a imersão não
promoveu resultados satisfatórios.
Silva et al (2007) na aclimatização de mudas de antúrios (Anthurium
andreanum) em diferentes substratos, fizeram a irrigação uma vez ao dia por
submersão das bandejas em água, e a cada 3 dias utilizando a solução nutritiva do
MS. Neste caso não foi avaliada sobrevivência, porém os substratos pó de coco

104
seco e vermiculita apresentaram resultado superior aos demais para as variáveis
analisadas. Os autores relacionaram esse resultado as características físicas e
químicas dos substratos, considerados adequados para o crescimento e
desenvolvimento do antúrio, uma vez que todos os tratamentos receberam a mesma
solução nutritiva.
Plantas de barbatimão (Stryphnodendron adstringens) em Plantmax
apresentaram 50% de sobrevivência e em vermiculita + Plantmax® 41% após a
aclimatização de 30 dias em sala de crescimento, com diminuição gradual do
sombrite e abertura parcial dos sacos plásticos que envolviam os tubetes. A
reduzida sobrevivência foi verificada após a retirada total do saco plástico que
envolveu os tubetes, aos 21 dias após o início da aclimatização (Nicioli et al., 2008).
Souza et al. (2011) fizeram a aclimatação de inhame (Dioscorea multiflora)
em areia com 50 plantas enraizadas in vitro e 50 não-enraizadas, cobertas com
frascos de vidro por 40 dias e observaram que as plântulas formaram raízes ex vitro
aos 90 dias após o transplantio, com índice de sobrevivência (75%) idêntico ao das
plântulas enraizadas in vitro. Conforme os autores esse resultado foi relevante no
contexto da otimização do protocolo de micropropagação, uma vez que a etapa de
enraizamento ex vitro tem sido apontada como vantajosa do ponto de vista de
redução de custos e do tempo para a obtenção de mudas.
Fermino Junior e Scherwinski-Pereira (2012) transferiram plantas enraizadas
in vitro com AIB de cerejeira (Amburana acreana) para casa de vegetação, em
câmaras plásticas transparentes contendo vermiculita e perfuradas na tampa
durante 30 dias. Nessas condições observaram sobrevivência de 54%, considerada
baixa pelos autores em comparação a outras espécies arbóreas, indicando a
necessidade de estudos mais aprofundados.
Em orquídea dama-da-noite (Brassavola tuberculata), Souza et al. (2015)
obtiveram maior sobrevivência (67,7%) em substrato esfagno com a imersão das
raízes produzidas in vitro em ANA (1,8 mg L-1), concentrações diferentes dessa
auxina (até 4,5 mg L-1) propiciaram decréscimos na porcentagem de sobrevivência
das plantas. Neste caso a aclimatização da da espécie foi feita em copos plásticos
mantidos por 30 dias em sala de crescimento, após foram acondicionadas em viveiro
com sobreposição de três telas de sombreamento (50% cada), com duas irrigações
diárias por difusores acionados automaticamente por temporizador digital e válvula

105
solenóide. Apesar desse cuidado, observaram senescência e posterior queda das
folhas no início da aclimatização.
A fase de transferência para aclimatação e rustificação das plantas
estabelecidas in vitro, constitui-se em uma importante etapa na formação de mudas
de qualidade, principalmente porque o tecido vegetal passa de uma condição
heterotrófica para a autotrófica, sofrendo estresses fisiológicos (Bandeira et al.,
2007). Diante do exposto, a mortalidade das plântulas no processo de aclimatação
pode ter sido a mudança drástica de condições ambientais, segundo Torres et al.
(1998) a transferência do ambiente in vitro para o ex vitro exige que se adapte
rapidamente a estas novas condições ambientais para sobreviver.
7.4 Conclusão
A elevada taxa de mortalidade (97,9%) pode ser associada a diversos fatores,
dentre eles a concentração endógena de hormônios e o balanço adequado para
ocorrer o estímulo ao desenvolvimento radicular. Além disso, pode ainda estar
relacionado às mudanças drásticas nas condições ambientais sofridas pelas
plântulas, sugerindo o desenvolvimento de protocolos mais eficientes nessa etapa.
7.5 Referências Bibliográficas
ALMEIDA, F.D.; XAVIER, A.; DIAS, J.M.M.; PAIVA, H.N. Eficiência das auxinas (AIB e ANA) no enraizamento de miniestacas de clones de Eucalyptus cloeziana F. Muell. Revista Árvore, Viçosa, v.31, n.3, p.455-463, 2007. ANDRADE, M.W.; LUZ, J.M.Q.; LACERDA, A.S.; MELO, P.R.A. Micropropagação da aroeira (Myracrodruon urundeuva Fr. All.). Ciência e Agrotecnologia, Lavras, v.24, n.1, p.174-180, 2000. ASSIS, T.F.; TEIXEIRA, S.L. Enraizamento de plantas lenhosas. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. (Eds.). Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa-SPI / Embrapa-CNPH, 1998. v.1, p.261-96. BANDEIRA, F. S.; XAVIER, A.; OTONI, W. C.; LANI, E. R. G. Aclimatação ex vitro de plantas propagadas pela enxertia in vitro de Eucalyptus urophylla x E. grandis. Revista Árvore, Viçosa, v.3,n.5, p.773 - 781, 2007. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Regras para Análise de Sementes. Secretaria de Defesa Agropecuária. Brasília: MAPA/ACS5, 2009. 395p.

106
BRONDANI, G.E.; BACCARIN, F.J.B.; BERGONCI, T.; GONÇALVES, A.N.; ALMEIDA, M. Miniestaquia de Eucalyptus benthamii: efeito do genótipo, AIB, zinco, boro e coletas de brotações. Cerne, Lavras, v.20, n.1, p.147-156 , 2014. BRONDANI, G.E.; GROSSI, F.; WENDLING, I.; DUTRA, L.F.; ARAUJO, M.A. Aplicação de IBA para o enraizamento de miniestacas de Eucalyptus benthamii Maiden & Cambage x Eucalyptus dunnii Maiden. Acta Scientiarum.Agronomy, Maringá, v.32, n.4, p.667 - 674, 2010. CALDEIRA, M.V.; DELARMELINA, W.M.; PERONI, L.; GONÇALVES, E.O.; SILVA, A.G. Lodo de esgoto e vermiculita na produção de mudas de eucalipto. Pesquisa Agropecuária Tropical, Goiânia, v. 43, n. 2, p. 155-163, 2013. CALVETE, E.O.; KÄMPF, A.N.; DAUDT, R. Efeito do substrato na aclimatização ex vitro de morangueiro cv Campinas, Fragaria x ananassa Duch. In: KÄMPF, A.N.; FERMINO, M.H. (Ed.). Substrato para plantas: a base da produção vegetal em recipientes. Porto Alegre: Genesis, 2000. p.257-264. CALVETE, E.O.; KÄMPF, A.N.; SUZIN, M. Concentração de sacarose no enraizamento in vitro de morangueiro. Horticultura Brasileira, Brasília, v. 20, n.2, p. 186-191, 2002. COSTA, F.H.S.; PASQUAL, M.; Scherwinski Pereira, J.E.; RODRIGUES, F.A.; MIYATA, L.Y. Relação entre o tempo de enraizamento in vitro e o crescimento de plantas de bananeira na aclimatização. Revista Brasileira de Fruticultura, Jaboticabal, v.30, n.1, p.31-37, 2008. DAMIANI, C.R.; PELIZZA, T.R.; SCHUCH, M.W.; RUFATO, A.R. Luminosidade e IBA no enraizamento de microestacas de mirtileiro dos grupos rabbiteye e southern highbush. Revista Brasileira de Fruticultura, Jaboticabal, v.31, n.3, p.650-655, 2009. DIAS, P.C.; OLIVEIRA, L.S.; XAVIER, A.; WENDLING, I. Estaquia e miniestaquia de espécies florestais lenhosas do Brasil. Pesquisa Florestal Brasileira, Colombo, v.32, n.72, p.453 - 462, 2012. FAGANELLO, L.R.; DRANSKI, J.A.L.; MALAVASI, U.C.; MALAVASI, M.M. Efeito dos ácidos indolbutírico e naftalenoacético no enraizamento de estacas semilenhosas de Cordia trichotoma (Vell.) Arrab. ex Steud. Ciência Florestal, Santa Maria, v. 25, n. 4, p. 863-871, 2015. FERMINO JUNIOR, P.C.P.; RAPOSO, A.; SCHERWINSKI-PEREIRA, J.E. Enraizamento ex vitro e aclimatização de plantas micropropagadas de Tectona grandis. Floresta, Curitiba, v.41, n.1, p.79-86, 2011. FERMINO JUNIOR, P.C.P; SCHERWINSKI-PEREIRA, J.E. Germinação e propagação in vitro de cerejeira (Amburana acreana (Ducke) A.C. Smith – Fabaceae). Ciência Florestal, Santa Maria, v.22, n.1, p.1-9, 2012.

107
FERNANDES, C.; CORÁ, J.E.; BRAZ, L.T. Alterações nas propriedades físicas de substratos para cultivo de tomate cereja, em função de sua reutilização. Horticultura Brasileira, Campinas, v. 24, n. 1, p. 94-98, 2006. FERNANDES, D.A.; MARTINEZ, D.T.; COSTA, R.B. Sacarose e ácido indolbutírico no enraizamento de Tectona grandis L.f. Revista de Agricultura, Piracicaba, v.90, n.1, p. 87-99, 2015. FERREIRA, M.C.; COSTA, S.M.L.; PASIN, L.A.A. Uso de resíduos da agroindústria de bananas na composição de substratos para produção de mudas de pau pereira. Nativa, Sinop, v. 03, n. 02, p. 120-124, 2015. GEORGIN, J.; BAZZOTI, R.; PERRANDO, E. Indução ao enraizamento de estacas de teca (Tectona grandis l.f). REGET - Revista Eletrônica em Gestão, Educação e Tecnologia Ambiental, Santa Maria, v.18, n.3, p.1246-1256, 2014. GRATTAPAGLIA, D.; MACHADO, M.A. Micropropagação. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa – SPI / Embrapa – CNPH, 1998, v.1, p.183-260. IACONA, C.; MULEO, R. Light quality affects in vitro adventitious rooting and ex vitro performance of cherry rootstock colt. Scientia Horticulturae, v.125, p.630-636, 2010. KÄMPF, A. N. et al. Substratos para plantas: a base da produção vegetal em recipientes, Porto Alegre: Gênesis, 2000. 312 p. KIELSE, P.; BISOGNIN, D.A.; HEBERLE, M.; FLEIG, F.D.; XAVIER, A.; RAUBER, M.A. Propagação vegetativa de Cordia trichotoma (Vell.) Arrab. ex Steudel por estaquia radicular. Revista Árvore, Viçosa, v.37, n1., p.59-66, 2013. LANA, R.M.Q.; LANA, A.M.Q.; BARREIRA, S.; MORAIS, T.R.; FARIA, M.V. Doses de ácido indolbutírico no enraizamento e crescimento de estacas de eucalipto (Eucalyptus urophylla). Bioscience Journal, Uberlândia, v.24, n.3, p.13-18, 2008. LEITZKE, L.N.; DAMIANI, C.R.; SCHUCH, M.W. Meio de cultura, concentração de AIB e tempo de cultivo no enraizamento in vitro de amoreira-preta e framboeseira. Revista Brasileira de Fruticultura, Jaboticabal - SP, v. 31, n. 2, p. 582-587, 2009. LOPES, S.C.; LAMEIRA, O.A.; FORTES, G.R.L.; NOGUEIRA, R.C.; PINTO J.E.B.P. Enraizamento in vitro de mogno (Swietenia macrophylla King). Cerne, Lavras, v.7, n.1, p.124-128, 2001. MACHADO, M.P.; DESCHAMPS, C.; BIASI, L.A. Application of IBA on in vitro and ex vitro rooting microcutting of Lavandula angustifolia Miller. Journal of Biotechnology and Biodiversity, Gurupi, v.4, n.2, p.153-161, 2013. MEDEIROS, A.C.S.; SMITH, R.; PROBERT, R.; SADER, R. Comportamento fisiológico de sementes de aroeira (Myracrodruon urundeuva Fr. All.) em condições de armazenamento. Boletim de Pesquisa Florestal, Colombo, n.40, p. 85-98, 2000.

108
MELO, B.; MENDES, A.N.G.; GUIMARÃES, P.T.G. Tipos de fertilizações e diferentes substratos na produção de mudas de cafeeiro (Coffea arabica L.) em tubetes. Bioscience Journal, Uberlândia, v.19, n.1, p. 33-42, 2003. MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiologia Plantarum, Copenhagen, v.15, p.473-97, 1962. NICIOLI, P.M.; PAIVA, R.; NOGUEIRA, R.C.; SANTANA, J.R.F. et al. Ajuste do processo de micropropagação de barbatimão. Ciência Rural, Santa Maria, v.38, n.3, p.685-689, 2008. NORBERTO, P.M.; CHALFUN, N.N. J.; PASQUAL, M.; VEIGA, R.D.; PEREIRA, G.E.; MOTA, J.H. Efeito da época de estaquia e do AIB no enraizamento de estacas de figueira (Fícus carica L.). Ciência e Agrotecnologia, Lavras, v. 25, n. 3, p.533-541, 2001. PAIVA SOBRINHO, S.; LUZ, P.B.; SILVEIRA, T.L.S.; RAMOS, D.T.; NEVES, L.G.; BARELLI, M.A.A. Substratos na produção de mudas de três espécies arbóreas do cerrado. Revista Brasileira de Ciências Agrárias, Recife, v.5, n.2, p.238-243, 2010. PASQUAL, M. Cultura de tecidos. Lavras: UFLA/FAEPE, 2001. 165p. PELIZZA, T.R.; MUNIZ, J.; CAMARGO, P.; KRETZSCHMAR, A.A.; RUFATO, L. Enraizamento ex vitro e aclimatização de plântulas micropropagadas de amoreira-preta „xavante‟. Revista Brasileira de Fruticultura, Jaboticabal, v.35, n.1, p.329-332, 2013. PEREIRA, M.O.; WENDLING,I.; NOGUEIRA, A.C.; KALIL FILHO, A.N.; NAVROSKI, M.C. Resgate vegetativo e propagação de cedro-australiano por estaquia. Pesquisa Agropecuária Brasileira, Brasília, v.50, n.4, p.282-289, 2015. POZZA, A.A.A.; GUIMARÃES, P.T.G.; POZZA, E.A.; CARVALHO, J.G.; MONTARINI, M.; SOUZA, R.F. Efeito do tipo de substrato e da presença de adubação suplementar sobre o crescimento vegetativo, nutrição mineral, custo de produção e intensidade de cercosporiose em mudas de cafeeiro formadas em tubetes. Ciência e Agrotecnologia, Lavras, v. 31, n. 3, p. 685-692, 2007. ROCHA, M.A.C.; COSTA, M.A.P.C.; SILVA, S.A.; LEDO, C.A.S.; MOREIRA, M.J.S.; BASTOS, L.P. Enraizamento in vitro e aclimatização de genótipos de jenipapeiro (Genipa americana L.). Revista Brasileira de Fruticultura, Jaboticabal - SP, v. 30, n. 3, p. 769-774, 2008. ROCHA, S.C.; QUORIM, M.; RIBAS, L.L.F.; KOEHLER, H.S. Micropropagação de Cabralea canjerana. Revista Árvore, Viçosa, v.31, n.1, p.43-50, 2007. SANTOS, L.A.R.; SIQUEIRA, I.T.D.; PEREIRA, J.A.F.; ULISSES, C. Avaliação dos processos morfofisiológicos em mudas de orquídeas utilizando auxina na fase de

109
aclimatização. 2009. Disponível em: http://www.eventosufrpe.com.br/jepeX2009/cd/resumos/R0307-1.pdf Acesso em: 12 jan. 2016. SILVA, E.A.; OLIVEIRA, A.C.; MENDONÇA, V.; SOARES, F.M. Substratos na produção de mudas de mangabeira em tubetes. Pesquisa Agropecuária Tropical, Goiânia, v. 41, n. 2, p. 279-285, 2011. SILVA, F.A.S. ASSISTAT versão 7.7 beta. Campina Grande-PB: Assistência Estatística, Departamento de Engenharia Agrícola do CTRN - Universidade Federal de Campina Grande, Campus de Campina, 2011. SILVA, J.W.; HERNANDÉZ, F.F.F.; BEZERRA, F.C.; DINIZ, J.D.N. Aclimatização “ex vitro” de mudas de antúrio em diferentes substratos. Revista Ciência Agronômica, Fortaleza, v.38, n.2, p.188-191, 2007. SMIDERLE, O.J.; Produção de mudas de alface, pepino e pimentão em substrato combinando areia, solo e Plantmax®. Horticultura Brasileira, Brasília, v. 19, n. 2, p. 253-257, 2001. SORACE, M.; FARIA, R.T.; YAMAMOTO, L.Y.; SCHNITZER, J.A.; TAKAHASHI, L.S.A. Influência de auxina na aclimatização de Oncidium baueri (Orchidaceae). Semina: Ciências Agrárias, Londrina, v.28, n.2, p.195-200, 2007. SOUZA, A.V.; BERTONI, B.W.; FRANÇA, S.C.; PEREIRA, A.M.S. Micropropagação de Dioscorea multiflora Grised. Ciência e Agrotecnologia, Lavras, v.35, n.1, p.92-98, 2011. SOUSA G.G.; ROSA Y.B.C.J.; MACEDO M.C.; SOARES J.S. Aclimatização de Brassavola tuberculata com a utilização de ANA em diferentes substratos. Horticultura Brasileira, Brasília, v.33, n.2, p.208-215, 2015. STUEPP, C.A.; WENDLING, I.; KOEHLER, H.S.; ZUFFELLATO-RIBAS, K.C. Estaquia de árvores adultas de Paulownia fortunei var. mikado a partir de brotações epicórmicas de decepa. Ciência Florestal, Santa Maria, v. 25, n. 3, p. 667-677, 2015. SUTTER, E. Stomatal and cuticular water loss from apple, cherry and sweetgum plants after removal from in vitro culture. Journal of American Society for Horticultural Science, Washington, v.113, n.2, p.234-238, 1988. TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 4 ed. Porto Alegre: Artmed, 2009. 848 p. TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: EMBRAPA-SP/EMBRAPA-CNPH, 1998. 509p. TRONCO, K.M.Q.; BISOGNIN, D.A.; FLEIG, F.D.; HORBACH, M.A. Enraizamento ex vitro e aclimatização de microestacas de Ilex paraguariensis A. St Hil. Cerne, Lavras, v.21, n.3, p.371-378, 2015.

110
WENDLING, I.; XAVIER, A. Influência do ácido indolbutírico e da miniestaquia seriada no enraizamento e vigor de miniestacas de clones de Eucalyptus grandis. Revista Árvore, Viçosa, v.29, n.6, p.921 - 930, 2005. YAMAMOTO, L.Y.; BORGES, R.S.; SORACE, M.; RACHID, B.F.; RUAS, J.M.F.; SATO, O.; ASSIS, A.M.; ROBERTO, S.R. Enraizamento de estacas de Psidium guajava L. „Século XXI‟ tratadas com ácido indolbutírico veiculado em talco e álcool. Ciência Rural, Santa Maria, v.40, n.5, p.1037-1042, 2010. YAN, H.; LIANG, C.; YANG, L.; LI, Y. In vitro and ex vitro rooting of Siratia grosvenorii, a traditional medicinal plant. Acta Physiologiae Plantarum, Poland, v.32, n.1, p.115-120, 2010.

111
8 CONCLUSÃO GERAL
As etapas de micropropagação alcançadas nesse estudo indicam que a
assepsia, estabelecimento através da germinação in vitro e multiplicação de brotos
de M. urundeuva All. podem ser obtidos com sucesso. Entretanto, a aclimatização
com o enraizamento ex vitro necessita de estudos mais aprofundados para não
comprometer a produção em larga escala.