gradientes de productividad primaria, clorofila … · el estudio de la distribución espacial de...
TRANSCRIPT

65Gradientes de productividad y biomasa del fitoplancton en Aysén
GRADIENTES DE PRODUCTIVIDAD PRIMARIA, CLOROFILA-a Y COMPOSICIÓN POR TAMA-ÑOS DEL FITOPLANCTON EN CANALES OCCIDENTALES DE AYSÉN EN NOVIEMBRE 2002*
PRIMARY PRODUCTIVITY GRADIENTS, CHLOROPHYLL-a AND SIZE COMPOSITION OF PHYTOPLANKTONIN WESTERN CHANNELS OF AYSÉN REGION: NOVEMBER 2002
VIVIAN MONTECINO1
M. ALEJANDRA PAREDES1
MICHELLE MANLEY2
ROSA ASTORECA3
PAULINA URIBE4
GADIEL ALARCÓN5
GEMITA PIZARRO6
CAROLINA VARGAS7
1Universidad de Chile, Facultad de Ciencias, Departamento de Ciencias Ecológicas,Casilla 653, Santiago, Chile.
2 SHOA-Valparaíso,3 Ecologie des Systèmes Aquatiques-Université Libre de Bruxelles,
4 Fundación Ciencia para la Vida,5PROFC-Universidad de Concepción,
6IFOP- Zonal Punta Arenas,7Dirección General de Aguas (Dirección OO. PP.).
RESUMEN
Se analizó la influencia en la composición por tamaños del fitoplancton en la productividad deecosistemas acuáticos, específicamente para los canales occidentales de Aysén (43o - 47o S) durante elcrucero CIMAR 8 en noviembre de 2002. El estudio de la distribución espacial de la clorofila-a, sinfraccionar y fraccionada en tres clases de tamaño, reveló tendencias opuestas entre los distintos cana-les en los gradientes transversales de biomasa y la relación de la biomasa de la clorofila-a con el índicede diversidad de tamaños y los parámetros fotosintéticos y bio-ópticos del fitoplancton. El parámetrofotosintético PB
máx mostró promedios diferentes entre las estaciones del sur (canales Darwin y Pulluche)
y las del norte (canales Moraleda y King). En estas últimas estaciones fue donde se midieron lasmayores biomasas totales. En la mayoría de las estaciones y profundidades, la biomasa de la clorofila-a estuvo dominada por la fracción microfitoplanctónica (>20 micras), y sus valores más altos ocurrieronen aquellos sectores con gradientes parciales de salinidad y temperatura. Con estos resultados, seconfirma la predicción sobre la mayor abundancia de la fracción microfitoplancton asociado a columnasde agua con características eutróficas, turbulentas y parcialmente mezcladas.
Palabras claves: Diversidad de tamaños del fitoplancton, parámetros fotosintéticos y bio-ópticos. Fiordos y cana-les chilenos.
ABSTRACT
The influence phytoplankton size composition on the productivity of aquatic ecosystems is analyzed,specifically for the Aysén western channels (43o - 47o S) during CIMAR 8 cruise in November 2002. Thestudy of the distribution of non-fractionated chlorophyll-a and chlorophyll-a fractionated in three sizeclasses, showed opposite tendencies between channels of the along-shore gradients of biomass and inthe relationship of chlorophyll-a with a size-diversity index and the photosynthetic and bio-opticalparameters. The photosynthetic parameter PB
max showed different mean values between the southern
stations (channels Darwin and Pulluche) and the northern ones (channels Moraleda and King). In theselatter stations the highest total biomass were measured. In most stations and depths, the chlorophyll-a
Cienc. Tecnol. Mar, 29 (2): 65-85, 2006
* Proyecto CONA-C8F -02-09.

66
biomass was dominated by the microphytoplanktonic fraction (>20 microns) and their highest valuesoccurred in those sites with partial salinity and temperature gradients. From these results, the predictionabout higher microphytoplankton abundance associated to eutrophic characteristics of the water column,turbulent and partially mixed conditions is confirmed.
Key words: Phytoplankton size diversity, photosynthetic and bio-optical parameters. Chilean fjords and channels.
INTRODUCCIÓN
Los modelos que dan cuenta de la relaciónentre tamaño corporal y biomasa del fitoplanctonpredicen que en columnas de agua turbulentas,parcialmente estratificadas, eutróficas y sin li-mitación por luz, predomina el microfitoplancton(>20 µm) con elevadas biomasas totales, mien-tras que en columnas de agua estratificadas,oligotróficas y con limitación por luz, predominanel pico (<2 µm) y nanofitoplancton (2-20 µm)asociado a bajas biomasas totales (Kiorboe,1993). El predominio de la clase de mayor tama-ño del fitoplancton y su abundancia relativa tie-ne fuertes implicancias en la productividad delos sistemas (Margalef, 1978). El clima tambiéninfluye directa o indirectamente en la composi-ción y concentración del fitoplancton (Lehman etal., 2000) cuya diversidad específica como re-sultado de la competencia se relaciona con ladisponibilidad de recursos y a los regímenes deperturbaciones abióticas (Reynolds et al., 1993;Huisman et al., 1999).
El sistema de los fiordos y canales de Ay-sén, comprendidos entre los 43o - 47o S, y carac-terizados física y químicamente por Silva et al.(1998a) y Guzmán & Silva (2002), constituye unsistema abierto, altamente heterogéneo y de bajaestabilidad. Aquí, bajo condiciones hidrodinámi-cas longitudinales y transversales inducidas porel viento y frentes de marea (Cáceres et al.,2002), con diferentes situaciones ecológicas ypor lo tanto con diferente composición y abun-dancia del fitoplancton, se dan las condicionespara estudiar el comportamiento de la biomasafitoplanctónica total y su relación con la compo-sición por clases de tamaño. Estas característi-cas se deben principalmente al efecto de unaalta variabilidad local y estacional en los proce-sos de turbulencia, en la concentración de nu-trientes y en la disponibilidad de luz (Pizarro etal., 2005a).
Las asociaciones fitoplanctónicas y algunospatrones de su distribución espacial y temporal envarios fiordos y canales australes, han sido descri-tas por Vera et al. (1996), Muñoz & Avaria (1997) yPizarro et al. (2000, 2005a, 2005b). Sin embargo,en relación con la dinámica y estructura de estos
ecosistemas australes, la relevancia de la estruc-tura de tamaños del fitoplancton o de su diversidadde tamaños es poco conocida.
Para el presente estudio, se analizaron dife-rentes sitios (estaciones oceanográficas) quedistribuidos en gradientes “océano-canales”,presentan diferentes condiciones hidrográficasde estratificación, mezcla parcial o mezcla tur-bulenta y en consecuencia diferente productivi-dad por diferente apor te de nutrientes. Seconsideró como hipótesis, que las áreas másoceánicas con termo-nutriclinas más profundasy con menor fertilización por procesos de mez-cla, presentarían una biomasa fitoplanctónica yuna productividad primaria menor que en loscanales. Este patrón estaría asociado a unamenor proporción de células microfitoplanctóni-cas en la estructura de tamaños de estas co-munidades fitoplanctónicas.
METODOLOGÍA
Durante el crucero CIMAR 8-2 (noviembre,2002) se realizaron mediciones diarias de lapenetración de la radiación solar para determi-nar la dimensión de la zona eufótica y definir lasprofundidades de muestreo. En las estacionestanto diurnas como nocturnas se recolectaronmuestras de agua hasta en 5 profundidades dis-cretas para estimar la biomasa fitoplanctónicatotal y de tres fracciones de tamaño (>20, 11-20y <11 µm), considerando como un indicador desu biomasa la concentración clorofila-a (CL-a).Cada muestra de 250-500 ml se filtró con y sinpre-filtración con mallas de 20 y 11 µm, sobrefiltros de fibra de vidrio de tamaño de poro 0,7µm (nominal). Los filtros se conservaron en ni-trógeno líquido hasta su análisis en laboratorio(Jeffrey et al., 1997), donde se extrajeron lospigmentos con acetona y los extractos fueronleídos en un fluorómetro 10 AU TURNER Designpreviamente calibrado con CL-a estándar (Sigma).
Simultáneamente, se fijaron muestras deagua de diferentes profundidades con lugol, parael posterior análisis cuantitativo del fitoplanctonen microscopio invertido con contraste de fasesy cámaras de 10 y 50 ml (Pizarro et al., 2005a).El fitoplancton cualitativo fue identificado en

67Gradientes de productividad y biomasa del fitoplancton en Aysén
muestras recolectadas mediante lances vertica-les de una red de 40 µm hasta 50 m de profun-didad, fijadas con formalina al 5%, que se anali-zaron en tres alícuotas de cada una (Pizarro etal., op. cit.). Para el estudio de picoplancton porcitometría de flujo, se fijaron 2 mL de agua demar con paraformaldehido 10% y se almacena-ron en nitrógeno líquido (Vaulot et al., 1989). Elanálisis de las poblaciones picoplanctónicas fuerealizado en un citómetro de flujo FACSCaliburcon un láser Ion-Argon de 488 nm de 15 mW(Becton Dickinson, San José, CA). El recuentocelular fue procesado usando los programasCellQuest-Pro (BD) and CytoWin (D. Vaulot, Sta-tion Biologique, Roscoff, Francia). La discrimina-ción entre las diferentes poblaciones fue realiza-da en base a sus propiedades ópticas de fluo-rescencia y tamaño (Marie et al., 1999). Comoel bacterioplancton carece de pigmentos, debeser teñido usando tinciones moleculares, por loque a cada muestra se le agregó 10–4 de la solu-ción stock de SYBR Green I (Molecular Probes)(Marie et al., 1997).
La diversidad de tamaños (H’tam
) se obtuvocon el índice de Shannon & Weaver modificado(Paredes, 2005), H’
tam= –∑ pi ln pSi, donde:
pi = la proporción de CL-a de la clase de tama-ños i de la misma muestra.
S = número total de fracciones de tamaño de lamisma muestra.
En 11 estaciones diurnas y de muestras ob-tenidas a distintas profundidades discretas, sefiltraron entre 200 y 500 ml de agua para deter-minar la absorbancia del fitoplancton in vivo (a
ph).
Posteriormente también se estimó la absorciónespecífica (a*) normalizando a
ph por el corres-
pondiente valor de CL-a sin feopigmentos. Paraestas mediciones se utilizaron filtros GF/F What-man de 0,7 µm (nominal), los que fueron conser-vados en placas histológicas en nitrógeno líqui-do para determinar absorbancia espectral in vivode partículas (a
p) en un espectrofotómetro (Shi-
madzu UV_Visible) en el rango 300-700 nm (Mit-chell et al., 2000). Luego de medir a
ph, los pig-
mentos se extraen con metanol y el filtro se midenuevamente en el mismo intervalo de 300-700nm para determinar la absorbancia por partícu-las no algales (a
PNA). Los resultados de absor-
bancia de cada uno de los espectros se convier-ten en coeficientes de absorción según:
ap/
PNA(λ)= 2,303*ODp/
NAP(λ)/X, donde X es
la razón entre el volumen filtrado y el área delfiltro con material filtrado. Para corregir la dis-
persión de luz, se sustrajo el valor promedio en-tre 700 y 750 nm de a
p y de a
PNA y luego se sus-
trajo de aph
el valor de aPNA
para obtener laabsorbancia espectral de los pigmentos (a
ph). Los
espectros aPNA
(λ) se corrigieron ajustando la fun-ción exponencial:
aPNA
(λ)= aPNA
(λ)* e(-s(λ-λ–1)). Posteriormente losresultados de a
ph y a* se expresan de acuerdo al
valor obtenido a 440 nm (Tabla I).
De las mismas muestras originales se sub-muestrearon 100 ml para medir materia orgánicadisuelta coloreada (a
g). Las estimaciones de a
g
se realizaron a partir de la filtración de estas mues-tras por filtros precalcinados de fibra de vidrio yluego por filtros de policarbonato de 0,2 µm, am-bos filtros pre-lavados con 100 ml de aguaultrapura Milli Q (filtrada en equipo Barnstead).Para calcular a
g se registró la densidad óptica en
una cubeta de 10 cm de paso de luz en unespectrofotómetro Shimadzu UV-Visible utilizandoagua ultrapura Milli Q como referencia. Los valo-res de absorbancia se transformaron en coeficien-te de absorción según a
g(λ)= 2,303*ODa
g(λ)/L
donde L es el largo de la cubeta. La funciónexponencial para ajustar los datos es la mismaque la utilizada para a
PNA (Mitchell et al., 2000,
Pizarro et al., 2005b) y los resultados se expre-san de acuerdo con el valor obtenido a 375 nm(Tabla I).
Para cuantificar la productividad primaria seutilizaron muestras de agua de dos profundida-des super ficiales, 5 y 25 m. Las mediciones defotosíntesis versus irradiancia (P-E) se realiza-ron en un fotosintetrón. Para cada profundidadse incubaron entre 10-12 submuestras de 1 ó2 ml inoculadas con 14C (1µCi/ml) durante 1hora en un bloque termorregulado ajustado a latemperatura promedio superficial. El gradientede intensidades de la luz (E) varió entre 2,5 y777,2 µmoles de fotones m–2 s–2 de radiaciónfotosintéticamente activa en cada serie. Paraobtener los parámetros fotosintéticos Alpha (efi-ciencia en la fijación de carbono con limitaciónde luz) y PB
máx (tasa de fijación máxima de car-
bono normalizado por clorofila-a), se ajustaronlas curvas P-E utilizando el programa Systat (Wil-kinson, 1989) con el siguiente modelo: PB =PBmax tanh (Alpha*I/PBmax), (Montecino &Quiroz, 2000).
Cada día y alrededor del mediodía, se reali-zaron mediciones de los perfiles de radiaciónespectral descendente E
d(λ) en cuatro longitu-
des de onda del espectro visible (411, 442, 489y 555 nm) y tres λ el rango del UV, utilizando un
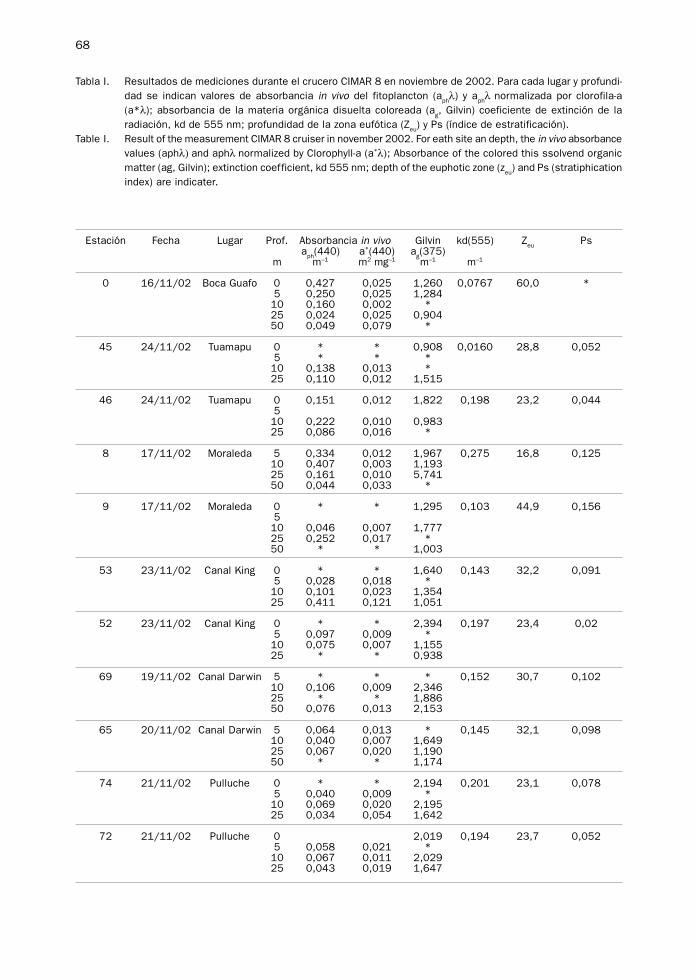
68
Tabla I. Resultados de mediciones durante el crucero CIMAR 8 en noviembre de 2002. Para cada lugar y profundi-dad se indican valores de absorbancia in vivo del fitoplancton (a
phλ) y a
phλ normalizada por clorofila-a
(a*λ); absorbancia de la materia orgánica disuelta coloreada (ag, Gilvin) coeficiente de extinción de la
radiación, kd de 555 nm; profundidad de la zona eufótica (Zeu
) y Ps (índice de estratificación).Table I. Result of the measurement CIMAR 8 cruiser in november 2002. For eath site an depth, the in vivo absorbance
values (aphλ) and aphλ normalized by Clorophyll-a (a*λ); Absorbance of the colored this ssolvend organicmatter (ag, Gilvin); extinction coefficient, kd 555 nm; depth of the euphotic zone (z
eu) and Ps (stratiphication
index) are indicater.
Estación Fecha Lugar Prof. Absorbancia in vivo Gilvin kd(555) Zeu
Psa
ph(440) a*(440) a
g(375)
m m–1 m2 mg–1 m–1 m–1
0 16/11/02 Boca Guafo 0 0,427 0,025 1,260 0,0767 60,0 *5 0,250 0,025 1,28410 0,160 0,002 *25 0,024 0,025 0,90450 0,049 0,079 *
45 24/11/02 Tuamapu 0 * * 0,908 0,0160 28,8 0,0525 * * *10 0,138 0,013 *25 0,110 0,012 1,515
46 24/11/02 Tuamapu 0 0,151 0,012 1,822 0,198 23,2 0,044510 0,222 0,010 0,98325 0,086 0,016 *
8 17/11/02 Moraleda 5 0,334 0,012 1,967 0,275 16,8 0,12510 0,407 0,003 1,19325 0,161 0,010 5,74150 0,044 0,033 *
9 17/11/02 Moraleda 0 * * 1,295 0,103 44,9 0,156510 0,046 0,007 1,77725 0,252 0,017 *50 * * 1,003
53 23/11/02 Canal King 0 * * 1,640 0,143 32,2 0,0915 0,028 0,018 *10 0,101 0,023 1,35425 0,411 0,121 1,051
52 23/11/02 Canal King 0 * * 2,394 0,197 23,4 0,025 0,097 0,009 *10 0,075 0,007 1,15525 * * 0,938
69 19/11/02 Canal Darwin 5 * * * 0,152 30,7 0,10210 0,106 0,009 2,34625 * * 1,88650 0,076 0,013 2,153
65 20/11/02 Canal Darwin 5 0,064 0,013 * 0,145 32,1 0,09810 0,040 0,007 1,64925 0,067 0,020 1,19050 * * 1,174
74 21/11/02 Pulluche 0 * * 2,194 0,201 23,1 0,0785 0,040 0,009 *10 0,069 0,020 2,19525 0,034 0,054 1,642
72 21/11/02 Pulluche 0 2,019 0,194 23,7 0,0525 0,058 0,021 *10 0,067 0,011 2,02925 0,043 0,019 1,647

69Gradientes de productividad y biomasa del fitoplancton en Aysén
espectroradiómetro sumergible Satlantic. Loscoeficientes de extinción espectral K
d(λ) de la
radiación (Kirk, 1994) en la columna de agua a555 nm (longitud de onda más penetrante) secalcularon mediante la ecuación:
Ed(z) = E
d(0)*exp(-K
d(0-z).
Donde Ed(z) es la irradiancia descendente a pro-
fundidad z.
Ed(0) es la irradiancia descendente a profundi-
dad 0.
Kd(0-z) es el coeficiente de extinción de la luz
entre las profundidades 0 y z.
Para analizar la relación entre la concentra-ción de CL-a integrada entre 0 y 25 m, y la di-mensión de los procesos de mezcla en la colum-na de agua, se calculó el parámetro de estratifi-cación (Ps) de acuerdo a Prandle (1985) desdelos 0 m hasta la profundidad máxima de muestreopara cada estación: Ps = dδ/<s>
Donde δS = Diferencia entre salinidad de lasuperficie menos la salinidad del fondo.
<s> = promedio de salinidad transversal.
Las mediciones de temperatura, salinidad ydensidad en la columna de agua de cada esta-ción, se obtuvieron mediante un CTD SBE19, ylos datos fueron procesados en el Centro Nacio-nal de Datos Hidrográficos y Oceanográficos deChile (CENDHOC).
RESULTADOS
Patrones espaciales físico-biológicos
En al menos dos estaciones por cada canal, semuestran en paralelo la distribución de las abundan-cias de microfitoplancton, cianobacterias y picoeuca-riontes y la distribución vertical de la biomasa del fito-plancton medido como CL-a (Fig. 1 derecha) junto conlos registros de la temperatura y la salinidad (Fig. 1izquierda). A profundidades discretas, los valores máxi-mos de CL-a fueron registrados en estaciones con di-ferente desarrollo de estratificación. Por ejemplo, laestación 8 presentó 45,4 µg CL-a l–1 a los 10 m, resul-tado que concuerda con los intervalos máximos de CL-a integrada hasta los 25 m superficiales de la colum-na de agua (>370 µg CL-a m2) señalados en la figura2, para las estaciones ubicadas hacia el norte del ca-nal Moraleda. Este canal presentó sobre los 50 m,gradientes verticales de temperatura (10–11,8 oC) y
salinidad (28,2–32,2 psu) en la columna de agua (Fig.1, Moraleda 8 y 9), diferenciándose del resto de loscanales, que no presentaron gradientes significativosde estas variables, debido a discontinuidad de la es-tratificación (procesos de mezcla parcial de la columnade agua) y a una menor influencia de agua dulce deacuerdo a lo que se observa en los perfiles de tempe-ratura y salinidad (Fig. 1 Tuamapu; King; Darwin). Si seconsideran todas las estaciones, el rango de tempera-tura fluctuó entre 10 oC y 11,8 oC, y la salinidad entre27,1 y 32,6 psu. Las estaciones E65-Darwin, E72 yE74 en el canal Pulluche presentaron un máximo su-perficial de CL-a ≥ 5,0 µg.L–1 (Fig. 1, Darwin; Pulluche)que aunque es un valor alto, es comparativamenteinferior a aquellos del resto de las estaciones.
La distribución espacial de la CL-a integrada, mostróen los canales Tuamapu (44o S) y Pulluche (45o 70’ S) ungradiente positivo este-oeste, lo cual significa que lasconcentraciones de CL-a integrada aumentaron hacialas estaciones más oceánicas en aquellos canales (Fig.2). En cambio, en los canales Ninualac (45o S) y Darwin(45o 40’ S), se observó que las concentraciones deCL-a integrada presentan un gradiente inverso, es de-cir, las concentraciones disminuyen hacia las estacio-nes más oceánicas. Por último en el canal King (44o
56’ S) no se observa un gradiente transversal de la CL-a integrada. Las estaciones meridionales del áreamuestreada, presentaron los valores mínimos deCL-a integrada, con 91,0 mg.m–2 en E67-Darwin y94,6 mg.m–2 en E74-Pulluche (Fig. 2).
La relación entre la CL-a con el comportamientode la columna de agua fue evaluada utilizando el“Parámetro de Estratificación” (Ps), cuyos valoresvan desde Ps <0,15 = Estuario Bien Mezclado has-ta >0,32 = Estuario Estratificado. La fluctuación dePs entre 0,02 y 0,16 (Tabla I), significa que casitoda el área en noviembre 2002 se puede clasificarcomo de “estuario bien mezclado” con lo cual la CL-a integrada en los canales, no se encuentra regu-lada de manera significativa por diferencias en laestratificación de la salinidad o la temperatura.La excepción se dio en el canal Moraleda, dondecoinciden los mayores valores de Ps (0,12-0,16)con los mayores valores integrados de CL-a (224,7-1018,2 mg.m–2) en las estaciones E9 y E7 res-pectivamente.
En relación con la dimensión de la zona eufóticaz
eu, que queda determinada en gran medida por la
biomasa —en sistemas dominados por absorciónde luz por parte del fitoplancton— y la materia or-gánica coloreada disuelta (a
g λ), las estaciones que
presentaron menores zeu
(<23 m) en los canales,tenían altos valores de a
g375 > 1,5 m–1 (Tabla I) y
también los valores máximos de CL-a integrada. Elcoeficiente de extinción de la longitud de onda

70
Fig. 1, a-e: Izquierda: Perfiles de temperatura y salinidad y valores de clorofila-a a profundidades discretas en estacio-nes seleccionadas. En todos los gráficos la escala de clorofila es 0-25 mg.m–3 con excepción de la estaciónØ8 que es hasta 60 mg.m–3.Derecha: Perfiles verticales de las abundancias de cianobacterias, picoeucariontes y bacterias en célulaspor ml y del microfitoplancton en células por L.Las estaciones se ordenan secuencialmente de norte a sur (Fig. 2) igual que en la Tabla I, para canales a)Tuamapu (E45 y E46), b) Moraleda (E8 y E9), c) King (E52 y E53), d) Darwin (E65 y E69) y e) Pulluche (E72y E74). Noviembre 2002.
Fig. 1, a-e: Left: Profiles of temperature and salinity and chlorophyll-a values at specific depths in selected stations. Inall plots the chlorophyll scale is 0-25 mg.m–3. With exception of station Ø8 where it is up to 60 mg.m–3. Right:Vertical profiles of the abundances of cyanobateria, picoeucarionts and bacteria in cells per ml and of themicrophytoplankton in cells per L.Stations are arranged sequentially from north to south (Fig. 2) in the same order as in Table I, for channelsa) Tuamapu (E45 and E46), b) Moraleda (E8 and E9), c) King (E52 and E53), d) Darwin (E65 and E69) ande) Pulluche (E72 y E74). November 2002.
Pr
fund
idad
(m)
bacterias cyanobacteriaspicoeucariontes microfitoplancton
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000 10000
células ml –1x 103 o células L –1 x 103
o
Tuamapu 45
bacterias cyanobacteriaspicoeucariontes microfitoplancton
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000 10000
células ml –1x 103 o células L –1 x 103
Pro
fundid
ad
(m)
Tuamapu 46
0
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fun
did
ad
(m)
Temperatura ( C)o
0 5 10 15 20 25Clorofila (ug/l)
26 28 30 31 33 3527 29 32 34Salinidad
Tuamapu 45
0
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fun
did
ad
(m)
Temperatura ( C)o
0 5 10 15 20 25Clorofila (ug/l)
26 28 30 31 33 3527 29 32 34Salinidad
Tuamapu 46
cianobacterias
cianobacterias
A

71Gradientes de productividad y biomasa del fitoplancton en Aysén
Fig. 1, a-e: Continuación.Fig. 1, a-e: To be continued.
bacterias cyanobacteriaspicoeucariontes microfitoplancton
células ml –1x 103 o células L–1x 10 3
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000 10000
Pro
fundid
ad
(m)
Moraleda 08
0
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fun
did
ad
(m)
Temperatura ( C)o
0 10 20 30 40 60
Clorofila (ug/l)
26 28 30 31 33 3527 29 32 34Salinidad
Moraleda 08
50
células ml –1x 103 o células L –1 x 103
bacterias cyanobacteriaspicoeucariontes microfitoplancton
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000 10000
Pro
fundid
ad
(m)
Moraleda 09
0
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fun
did
ad
(m)
Temperatura ( C)o
0 5 10 15 20 25Clorofila (ug/l)
26 28 30 31 33 3527 29 32 34Salinidad
Moraleda 09
B
cianobacterias
cianobacterias

72
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000 10000
células ml –1x 103 o células L –1 x 10 3
Pro
fundid
ad
(m)
bacterias cyanobacteriaspicoeucariontes microfitoplancton
King 52
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000 10000
células ml -1 x 103 o células L -1 x 103
Pro
fundid
ad
(m)
bacterias cyanobacteriaspicoeucariontes microfitoplancton
King 53
0
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fundid
ad
(m)
Temperatura ( C)o
0 5 10 15 20 25Clorofila (ug/l)
26 28 30 31 33 3527 29 32 34Salinidad
King 52
0
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fundid
ad
(m)
Temperatura ( C)o
26 28 30 31 33 3527 29 32 34Salinidad
King 53
0 5 10 15 20 25Clorofila (ug/l)
Fig. 1, a-e: Continuación.Fig. 1, a-e: To be continued.
C
cianobacterias
cianobacterias

73Gradientes de productividad y biomasa del fitoplancton en Aysén
células ml –1x 103 o células L –1x 103
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000 10000
Pro
fun
did
ad
(m)
bacterias Cianobacteriaspicoeucariontes microfitoplancton
Darwin 69
bacterias Cianobacterias
picoeucariontes microfitoplancton
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000 10000
Pro
fundid
ad
(m)
Darwin 65
células ml –1x 103 o células L –1x 103
0
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fundid
ad
(m)
Temperatura ( C)o
0 5 10 15 20 25Clorofila (ug/l)
26 28 30 31 33 3527 29 32 34Salinidad
Darwin 65
0
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fundid
ad
(m)
Temperatura ( C)o
0 5 10 15 20 25Clorofila (ug/l)
26 28 30 31 33 3527 29 32 34Salinidad
Darwin 69
Fig. 1, a-e: Continuación.Fig. 1, a-e: To be continued.
D

74
células ml –1x 103 o células L –1x 10 3
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000 10000
Pro
fun
did
ad
(m)
bacterias Cianobacteriaspicoeucariontes microfitoplancton
Pulluche 720
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fundid
ad
(m)
Temperatura ( C)o
26 28 30 31 33 3527 29 32 34Salinidad
Pulluche 72
0 5 10 15 20 25Clorofila (ug/l)
Pro
0
5
10
15
20
25
30
35
40
45
50
0,01 0,1 1 10 100 1000
células ml –1x 103o células L–1x 103
fundid
ad
(m)
bacterias Cianobacteriaspicoeucariontes microfitoplancton
Pulluche 74
0
5
10
15
20
25
30
35
40
45
509 9,5 10 10,5 11 11,5 12
Pro
fundid
ad
(m)
Temperatura ( C)o
0 5 10 15 20 25
Clorofila (ug/l)
26 28 30 31 33 3527 29 32 34
Salinidad
Pulluche 74
Fig. 1, a-e: Continuación.Fig. 1, a-e: To be continued.
E

75Gradientes de productividad y biomasa del fitoplancton en Aysén
47°
46°
45°
44°
43° S
75° 74° 73°
Boca del Guafo
IslaGuafo
76° W
Seno Aysén
LagunaSan Rafael
Islade Chiloé
IslaGuamblín
Canal Ninualac
Canal DarwinCanal Pulluche
Bahía Tictoc
Can
alM
ora
led
a
Canal Puyuguapi
Península de Taitao
Bahía Anna Pink
Océan
oP
acíf
ico
Constricción de Meninea
0
4
7
8
9
10
31
444546
4950515253
6263
64 656667 69
707274777879
60 A
50 to 129.9
130 to 199.9
200 to 279.9
280 to 369.9
370 to 1200
Isla de Pascua
109 20’o
26
27
’o
105 20’o
26
27
’o
Isla San Ambrosio
Isla San Félix
80 05’o
Isla Salas y Gómez
Arch. Juan Fernández
33
37’
o
53
46’
o
78 49’o
60 46’o
I.Alejand
ro
Selkirk I. Robinson Crusoe
20° S80° W 60° W
20°
30°
40°
50°
40°
30°
Territorio Chileno Antártico
90°W 53° W
60° 60°
Canal King
Canal Tuamapu
Fig. 2: Distribución a mesoescala de la clorofila-a integrada entre 0 y 25 metros en mg CL-a m2. Colores indicanlos intervalos de abundancia de la CL-a para cada una de las estaciones muestreadas en canales occi-dentales de Aysén (I-VII), desde boca del Guafo hasta el canal Pulluche entre el 16 y 24 de noviembre2002.
Fig. 2: Mesoscale distribution of integrated chlorophyll-a between 0-25 meters, in mg CL-a m2. Ranges for CL-aabundances are shown in colors for each of the sampled stations in the western channels of Aysén (I-VII),from Guafo passage to Pulluche channel between 16-24 of November 2002.

76
más penetrante (kd555 nm) alcanzó valores muy
similares en sitios muy diferentes y distantes (Ta-bla I). Por ejemplo 0,194 y 0,198 m–1 en la esta-ción interior 72 (canal Pulluche) y la estaciónoceánica 46 (afuera del canal Tuamapu) y valoresmuy diferentes en sitios muy cercanos como en elcanal Moraleda en la estación 8, que fue de 0,275m–1 y disminuyó a 0,103 m–1 en la estación 9.
Producción primaria
Al comparar los resultados de los experi-mentos de P versus E realizados con dos mues-tras subsuper ficiales (Fig. 3), se observa va-riabilidad entre los diferentes lugares presen-tando un gradiente este-oeste y también entrelas profundidades de 5 y 25 m. Los valores dePBmax más altos de las muestras de 5 m (>6,0mgC (mg CL-a) h–1)), se obtuvieron en E65-Darwin y E72-Pulluche y los valores máximospara el parámetro de eficiencia Alpha se obtu-vieron en E65-Darwin (0,029 mgC (mg CL-a)–1
(µmol m–2 s–1)) y E72-Pulluche (0,079 mgC (mgCL-a)–1 (µmol m–2 s–1)). Los valores de PBmax ≤2,0 mgC (mg CL-a) h–1) se midieron en las es-taciones 8, 52 y 53 (Moraleda y King respecti-vamente) y con bajas eficiencias sólo en E53-King y E69-Darwin.
En relación con el parámetro aph
, su valor masbajo se midió en E9-Moraleda (0,002 m–1) y elmás alto en la vecina estación 8 (0,334 m–1)asociado a un orden de magnitud de diferenciaentre la concentración de la CL-a (2,8 y 28,0 mgm–3, respectivamente) aunque con valores simi-lares de H’
tam lo que concuerda con lo esperado,
considerando la presencia de bajos valores deH’
tam cuando hay bajos y altos valores de CL-a.
Agrupando espacialmente los resultados de losparámetros fotosintéticos en dos: nor te(Moraleda y King) y sur (Darwin y Pulluche), seobservan diferencias significativas en PB
max (F:
5,12; p<0,05) (Fig. 4), siendo menor en las es-taciones norte E8, E9, E52 y E53.
En varios lugares los experimentos realizadoscon la muestra de 25 m difieren notablemente,por esta razón y para facilitar el análisis, se orde-naron junto a estas curvas P-E, los de absorciónin vivo de las mismas muestras y también con ladistribución vertical de H’
tam (Fig. 3a, b y c).
Biomasa y estructura de tamaños
La mayoría de los valores de CL-a medidasen noviembre 2002 fueron mayores a 2 mg m–3 yla biomasa del fitoplancton estuvo dominada entodas las localidades y profundidades por la frac-
ción microfitoplanctónica (> 20 µm) (Fig. 5a). Engeneral, debido a la relación directa de la diver-sidad de tamaños con la biomasa encontrada eneste crucero (Fig. 5b) los bajos valores relativosde este índice H’
tam reflejan las variaciones de la
abundancia de la clase de tamaños dominante.
DISCUSIÓN
Los menores valores integrados de clorofi-la-a asociados a salinidades bajas de 27,1; 30,6y 31,7 psu alcanzaron respectivamente 142,7;124,1 y 94,6 mg CL-a m–2 en las estacionesinteriores del canal Pulluche (E70, E72 y E74);en tanto las estaciones más oceánicas (E78 yE79) presentaron concentraciones de CL-a inte-grada mayores de 322,8 y 363,6 mg m–2 lo quepodría estar asociado a una mayor/menor in-fluencia de las aguas de baja salinidad y me-nor/mayor transparencia respectivamente. Sinembargo, comparando con estudios anteriores(Pizarro et al., 2000, 2005a, 2005b), estosvalores de CL-a medidas en noviembre 2002,son los más altos reportados hasta la fechapara la región de Aysén.
Los patrones espaciales físico-biológicos jun-to con los cambios en z
eu (Tabla I) reflejan una
dinámica de la biomasa en parches según lascondiciones particulares de los diferentes sitios.Los incrementos de la biomasa al interior de loscanales ocurren cuando en sitios determinadosse producen condiciones favorables por un pe-ríodo de tiempo compatible con el crecimientode las células fitoplanctónicas y la dispersión delas poblaciones, y están determinados por losintensos flujos horizontales y los procesos decirculación. En sistemas similares, estos inter-cambios están modulados por la fuerte boyan-tés de las aguas, difundiendo los efectos loca-les de la mezcla sobre varios kilómetros en po-cos días (Gargett & Marra, 2002). Como se indicóen los resultados, la condición parcialmente mez-clado o de mezcla fue dominante para la mayoríade los sitios analizados aquí. En la cuenca sur delcanal Moraleda (45o 30’ S), los procesos de mez-cla obedecen a la presencia de ondas internas queson generadas por las corrientes de mareas ya queen estos sectores existen corrientes barotrópicasintensas, donde el tiempo de residencia de lasaguas depende tanto de los aportes de agua dulcecomo de la intensidad del viento (Salinas et al.,2002, Salinas & Hormazábal, 2004). Si se compa-ran lugares donde se encontraron cambios vertica-les de densidad con aquellos donde éstos fueronmenos intensos, la eficiencia fotosintética (Alfa) nose relacionó significativamente con el parámetrode estratificación.

77Gradientes de productividad y biomasa del fitoplancton en Aysén
Fig.
3, a-
c:Pa
ra c
ada
esta
ción
, en
la p
rimer
a fil
a se
mue
stra
n lo
s es
pect
ros
de a
bsor
banc
ia d
el fi
topl
anct
on (a
ph) e
ntre
400-7
00 n
m, p
ara
dist
inta
s pr
ofun
dida
des
(ver
sim
bolo
gía)
. Est
oses
pect
ros
se r
ealiz
aron
con
mue
stra
s de
las
mis
mas
est
acio
nes
y pr
ofun
dida
des
de lo
s ex
perim
ento
s P-
E (fot
osín
tesi
s ve
rsus
irra
dian
cia)
, cu
yos
resu
ltado
s es
tán
disp
ues-
tos
en la
seg
unda
y ter
cera
fila
(5 y
25 m
etro
s re
spec
tivam
ente
). E
n la
cua
rta
fila
se u
bica
n lo
s pe
rfile
s ve
rtic
ales
de
dive
rsid
ad d
e ta
mañ
os (H
’ tam). D
e iz
quie
rda
a de
rech
ael
ord
enam
ient
o de
las
esta
cion
es e
s de
oes
te-e
ste
para
los
dife
rent
es c
anal
es o
rden
ados
de
nort
e a
sur.
Fig.
3, a-
c:Fo
r ea
ch s
tatio
n, t
he f
irst
horiz
onta
l row
sho
ws
the
plot
s of
phy
topl
ankt
on a
bsor
banc
e sp
ectr
a (a
ph)
betw
een
400-7
00 n
m,
for
the
diff
eren
t de
pths
(no
te t
he c
olor
cod
es).
Thes
e sp
ectr
a w
ere
done
with
the
sam
e st
atio
ns a
nd d
epth
s of
the
P-E
exp
erim
ents
(ph
otos
ynth
esis
ver
sus
irra
dian
ce), res
ults
tha
t ar
e si
tuat
ed in
the
sec
ond
and
third
row
(5 a
nd 2
5 m
eter
s re
spec
tivel
y). In
the
fou
rth
row
the
siz
e di
vers
ity (H
tam) ve
rtic
al p
rofil
es a
re lo
cate
d. F
rom
left to
right
the
seq
uenc
e of
sta
tions
is fro
m w
est to
eas
t fo
r th
edi
ffer
ent
chan
nels
tha
t w
ere
arra
nged
fro
m n
orth
to
sout
h.
λλλλλ
λλλλλ
λλλλλ
a)

78
Da
rwin
E6
5
0,0
0,1
0,2
0,3
0,4
40
05
00
60
07
00
Lo
ng
itu
dd
eo
nd
a(n
m)
e6
50
5
e6
51
0
e6
52
5
Da
rwin
E6
55
m
0,0
4,0
8,0
02
00
40
06
00
80
0
�m
ole
sd
efo
ton
es
mgC(mgChl-a)h
Da
rwin
E6
52
5m
0,0
4,0
8,0
02
00
40
06
00
80
0
�m
ole
sd
efo
ton
es
mgC(mgChl-a)h
Da
rwin
E6
9
0,0
0,1
0,2
0,3
0,4
40
05
00
60
07
00
Lo
ng
itu
dd
eo
nd
a(n
m)
e6
91
0
e6
95
0
Da
rwin
E6
95
m
0,0
4,0
8,0
02
00
40
06
00
80
0
�m
ole
sd
efo
ton
es
mgC(mgChl-a)h
Da
rwin
E6
92
5m
0,0
4,0
8,0
02
00
40
06
00
80
0
�m
ole
sd
efo
ton
es
mgC(mgChl-a)h
Mo
rale
da
E9
0,0
0,1
0,2
0,3
0,4
40
05
00
60
07
00
Lo
ng
itu
dd
eo
nd
a(n
m)
e0
91
0
e0
92
5
Mo
rale
da
E9
5m
0,0
4,0
8,0
02
00
40
06
00
80
0
�m
ole
sd
efo
ton
es
mgC(mgChl-a)h
Mo
rale
da
E9
25
m
0,0
4,0
8,0
02
00
40
06
00
80
0
�m
ole
sd
efo
ton
es
mgC(mgChl-a)h
0
10
20
30
40
50
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Darw
inE
65
Profundidad(m)
H´t
am
años
0
10
20
30
40
50
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Darw
inE
69
Profundidad(m)
H´t
am
años
0
10
20
30
40
50
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mora
leda
E9
Profundidad(m)
H´t
am
años
aph()(m–
1)
aph()(m–
1)
aph()(m–
1)–
1
–
1
–
1
–
1
–
1
–
1
Fig.
3, a-
c: C
ontin
uaci
ón.
Fig.
3, a-
c: T
o be
con
tinue
d.
b)
λλλλλ
λλλλλ
λλλλλ

79Gradientes de productividad y biomasa del fitoplancton en Aysén
Pulluche E74
0,0
0,1
0,2
400 500 600 700
Longitud de onda (nm)
aph
()
(m)
e7405
e7410
e7425
Pulluche E74 5m
0,0
4,0
8,0
0 200 400 600 800
�moles de fotones
mg
C(m
gC
hl-a)
h
Pulluche 74 25 m
0,0
4,0
8,0
0 200 400 600 800
�moles de fotones
mg
C(m
gC
hl-a
)h
Pulluche E72
0,0
0,1
0,2
400 500 600 700
Longitud de onda (nm)
aph
()
(m)
e7205
e7210e7225
Pulluche E72 5m
0,0
4,0
8,0
0 200 400 600 800
�moles de fotones
mg
C(m
gC
hl-
a)
h
0,0
4,0
8,0
0 200 400 600 800
�moles de fotones
mg
C(m
gC
hl-
a)h
0
10
20
30
40
50
0.0 0.2 0.4 0.6 0.8 1.0 1.2
Pulluche E74
Pro
fundid
ad
(m)
H´tamaños
0
10
20
30
40
50
0.0 0.2 0.4 0.6 0.8 1.0 1.2
Pulluche E72
Pro
fun
did
ad
(m)
H´tamaños
Pulluche E72 25m
–
1
–
1
–
1
–
1
Fig. 3, a-c: Continuación.Fig. 3, a-c: To be continued.
c)
λλλλ λ λλλλ λ
-1 -1

80
Fig. 4: Promedio de parámetros fotosintéticos PBmax (mgC(mg CL-a)h–1) y Alpha (mgC(mg CL-a)–1 (µmol m–2 s–1) delas muestras de 5 m (cuadrado) y 25 m (triángulo), en las estaciones de los canales de la zona norteidentificadas con símbolos vacíos (Estación 8, 9 (Moraleda) y 52 y 53 (King)) y de la zona sur identificadascon símbolos llenos (Estación 65-69 (Darwin) y 72-74 (Pulluche)).
Fig. 4: Mean value of photosynthetic parameters PBmax (mgC (mg CL-a) h–1) and Alpha (mgC (mg CL-a) –1
(µmol m–2 s–1) of samples from 5 m (square) and 25 m (triangle) from stations in the northern channelsshown with empty symbols (Station 8- 9 (Moraleda) and 52-53 (King)) and from the southern zone shownwith full symbols (Station 65-69 (Darwin) and 72-74 (Pulluche)).
Norte 25 m
Sur 25 m
Norte 5 m
Sur 5 m
12
10
8
6
4
2
0
0,080,060,040,020
Alpha (mgC (mgCl-a) mol m s)–1 –2
(�
PB
ma
x(m
gc
(mg
cl-a
)h
)–1

81Gradientes de productividad y biomasa del fitoplancton en Aysén
Fig. 5: Gráficos de dispersión sobre la relación entre la clorofila-a sin fraccionar (biomasa total de fitoplancton)con A: la clorofila-a de la fracción <20 micras y B: el índice de la diversidad de tamaños (H
tam) del fitoplancton.
Resultados corresponden a la totalidad de las muestras de diferentes profundidades y diferentes estacio-nes en noviembre 2002.
Fig. 5: Scatter plots showing the relationship between chlorophyll-a without fractionation (phytoplankton totalbiomass) with A: Chlorophyll-a from the <20 microns fraction and B: phytoplankton size diversity index(H
tam). Results are for all the samples from different depths and different stations in November 2002.
Y= 0,936x - 0,8373R2 = 0,976
Clorofila-a sin fraccionar (mg m )–3
100
100
Clo
rofila
-afr
accio
nada
(mg
m)
–3
Fra
cció
n>
20
m
10
10
1
1
A
y = -0,2075Ln(x) + 0,8532
R2 = 0,6879
0,0
0,2
0,4
0,6
0,8
1,0
1,2
0,1 1 10 100
Clorofila-a (mg m )-3
H’d
eta
maños
B
.
.
.

82
Aunque los gradientes de H’tam
del fitoplanctonson difíciles de visualizar, se observa que a 5 oa 10 m, H’
tam es <0,4 en las estaciones 8 y 9 del
canal Moraleda y >0,4 hacia las estaciones másoceánicas (E53 10 m), de igual forma como ocu-rre en las estaciones del canal Darwin (E69 ver-sus E65) y del canal Moraleda hacia sur oeste(Fig. 3a, b). Aquí los valores de los parámetrosfotosintéticos y bio-ópticos sugieren que los ba-jos valores de H’
tam reflejan el predominio de una
sola clase de tamaño –la clase pequeña (<11µm)- en baja biomasa y que hacia el océano seincorporarían otras clases de tamaño. El canalMoraleda presenta bajas H’
tam subsuperficiales
(<0,6) que reflejan la predominancia delmicrofitoplancton (>20 µm) en altas biomasas(Figs. 1, 3a y 3b). Ya que a
ph aumenta con la
abundancia de la biomasa total– del fitoplancton(Bricaud et al., 1995), la similitud de los resulta-dos de a
ph y H’
tam indican cuál es la fracción de
tamaños dominante. En áreas oceánicas adya-centes y en varias de estas mismas localidadesen los canales interiores, pero en otros períodosdel año y bajo otras condiciones relativas a laestructura vertical de la columna de agua, sehabía encontrado que el tamaño predominantede la biomasa fitoplanctónica correspondía a lafracción <23 µm (nano y picofitoplancton) (Pizarroet al., 2000, 2005b).
El análisis de la composición por tamañoscorporales del fitoplancton permite conocer laestructura de los ecosistemas, dada la relaciónentre los procesos fisiológicos individuales, eltamaño del cuerpo y los procesos comunitariosy ecosistémicos (Ernest & Brown, 2001, Monte-cino, 2002). Así, los resultados de H’
tam del fito-
plancton, donde la biomasa total estuvo domina-da por la fracción microfitoplanctónica, seríanindicativos de una situación de eutrofía generali-zada con aguas bien mezcladas a mesoescala ycon efectiva aclimatación a la luz de esta frac-ción. En cambio Agawin et al. (2000), indican queel picofitoplancton domina en aguas oligotrófi-cas (menos de 0,3 mg.m–3 de CL-a) de tempera-turas más altas y pocos nutrientes. En Aysén seencontró que la fracción pequeña (< 20 µm) fuemenor del 10% de la biomasa total y sólo au-mentó su proporción cuando la clorofila total (sinfraccionar) disminuyó a menos de 5 mg.m–3. Losresultados sobre abundancia del microfitoplanc-ton (fracción > 20 µm) corroboran estos resulta-dos con concentraciones de 75,8 x 103 cél.L–1,donde la diatomea Stephanopyxis alcanzó un 95%(i.e. E8 del canal Moraleda). Irigoien et al. (2004)al referirse a patrones globales de biodiversidad,encontraron que en situación de incrementos dela biomasa, generalmente domina una sola es-
pecie. En estas condiciones de predominanciade unas pocas especies del microfitoplancton yen consecuencia con baja diversidad específica,podemos inferir que otras especies del fitoplanc-ton han sido desplazadas por efecto de compe-tencia por nutrientes o luz. Relacionando la bio-masa del microfitoplancton con su diversidad es-pecífica se encontró que la H’ específicadisminuyó a medida que la clorofila de la frac-ción >20 aumentó (sobre los 5,0 mg.m–3). Sinembargo, la diversidad específica estimada utili-zando únicamente especies microfitoplanctóni-cas no explica la estructura comunitaria, ya queno considera al pico y nanofitoplancton. Por ello,el uso del índice de diversidad de tamaño, com-prendiendo el amplio espectro de las microalgas,presenta mayor robustez para describir estosconglomerados. La abundancia y distribuciónver tical de cianobacterias y picoeucariontesque a pesar de estar siempre presente en abun-dancias similares (Fig. 1) avalan las diferen-cias encontradas en los resultados de H’
tam aún
cuando el sistema estuviera dominado por al-tas biomasas.
Las altas concentraciones de CL-a encontradasen el canal Moraleda, y las comparativamente ba-jas concentraciones de CL-a en los canales Pulluchey Darwin, son consistentes con un estudio realiza-do en el CIMAR-Fiordo 1 (Octubre 1995) por Silvaet al. (1998b). Estos autores encontraron altosvalores de materia orgánica en los sedimentos(>10%) en el centro del canal Moraleda, mientrasque en los canales Darwin y Pulluche (entre otros),se encontraron bajos porcentajes (<2,5%). Estasdiferencias se atribuyeron al efecto que tienen losglaciares en la zona, los cuales mediante la libera-ción de material inorgánico denominado “glacial silt”producirían una “dilución” del contenido orgánicode los sedimentos por una parte, y atenuarían laproductividad primaria del sector por otra, por limi-tación de luz.
El tamaño celular predominante en elfitoplancton determina las características de loscoeficientes de absorción de la luz (Ciotti et al.,2002). Comparando los resultados de a
ph de
noviembre 2002 (Fig. 3) con los obtenidos ennoviembre 2001 (CIMAR 7), donde los valoresmáximos a 440 nm fueron de 0,07 - 0,11 m–1
(Astoreca et al., 2002), estos fueron más altosen la mayoría de los casos en noviembre 2002,con máximos de 0,427 y 0,407 m–1 en la E0-Boca del Guafo a 0 m y E8-Moraleda a 10 mrespectivamente, acompañado por cambios pocosignificativos en la estructura de tamaños. Deltotal de datos, la fracción menor que 20 µm al-canzó más del 50% solo en un 6% de las mues-

83Gradientes de productividad y biomasa del fitoplancton en Aysén
tras. La estructura de tamaños reflejada por losbajos valores relativos de H’
tam, repercute en
mayores valores de aph
y menores valores en losparámetros Alfa y PBmax comparados con losvalores medidos en octubre y marzo por Pizarroet al. (2005) con promedios de 0,107 mgC (mgCL-a)–1 (µmol m–2 s)) y 9,84 mgC (mg CL-a) h–1
para Alpha y PBmax respectivamente.
Varios autores han centrado su atención en estu-diar la relación entre diversidad de especies y proce-sos de mezcla (i.e. Lindenschmidt & Chorus, 1998) yLi (2002) describió patrones macro ecológicos en elAtlántico Norte, analizando la relación entre la diver-sidad de tamaños y la estabilidad de la columna deagua. Para el área de Aysén, considerando los datoshistóricos junto con los resultados obtenidos en no-viembre 2002, donde los valores de CL-a fueron muyelevados y H’
tam del fitoplancton fue baja, se ha com-
pletado una escala interanual de variabilidad (Pare-des, 2005). Esta amplia variabilidad permitirá esta-blecer junto con nuevos datos, si de acuerdo con laintensidad de las perturbaciones (mezcla, luz, nutrien-tes, entre otros) la diversidad de tamaños presentauna respuesta característica unimodal. La respuestaunimodal esperada, se refiere a que una baja diversi-dad de tamaños, con predominancia de la fracciónmenor o mayor, está asociada a perturbaciones máxi-mas y mínimas, y una diversidad de tamaños alta(todas las fracciones presentes) está asociada a per-turbaciones de tipo intermedio.
El análisis de los resultados del cruceroCIMAR 8 en noviembre de 2002 confirma la pre-dicción de una mayor proporción de la fracciónmicrofitoplanctónica dada por las característicasde aguas eutróficas, turbulentas y parcialmentemezcladas de estos sistemas.
AGRADECIMIENTOS
Se agradece a Claudio Rivas su colaboraciónen cubierta y su trabajo en el laboratorio duranteel crucero. Para la edición agradecemos la cola-boración de Susana Giglio. Al comandante, la tri-pulación y a los ayudantes de cubier ta del AGOR“Vidal Gormaz”, al jefe de crucero y a la organi-zación del CONA del crucero CIMAR 8, nuestroreconocimiento. Este estudio fue posible graciasa los aportes directos del CONA e indirectos dela Universidad de Chile.
REFERENCIAS
ASTORECA, R., G. PIZARRO, M. A. PAREDES, P.URIBE & V. MONTECINO. 2002. Relación en-tre patrones espaciales de productividad y la
abundancia y estructura de tamaños delfitoplancton en sistemas de fiordos y cana-les de la XI región. Resultados Crucero CIMAR-7 Fiordos. Informes Preliminares. ComitéOceanográfico Nacional: 77-88.
BRICAUD, A., BABIN, M., MOREL, A., & H.CLAUSTRE. 1995. Variability in the chlorophyll-specific absorption coef ficients of naturalphytoplankton: analysis and parameterization.Journal of Geophysical Research 100, 13.321–13.332.
CÁCERES, M. A., VALLE-LEVINSON, H. H.SEPÚLVEDA, & K. HOLDERIED. 2002.Transverse variability of flow and density ina Chilean fjord, Continental Shelf Research,22, 1.683-1.698.
CIOTTI, A. M., M. R. LEWIS & J. J. CULLEN. 2002.Assessment of the relationships betweendominant cell size in natural phytoplanktoncommunities and the spectral shape of theabsorption coef ficient. Limnology andOceanography 47: 404-417.
ERNEST, S. K. M. & J. H. BROWN. 2001. Homeos-tasis and compensation: the role of speciesand resources in ecosystem stability. Ecolo-gy 82(8): 2.118-2.132.
GARGETT, A. & J. MARRA. 2002. Effects of upperocean physical processes (turbulence, advectionand air-sea interaction) on oceanic primaryproduction. In The Sea (ed. Alan Robinson, J.Mc Carthy & B. Rotschild. 12: 19-49.
GUZMÁN, D. & N. SILVA. 2002. Caracterizaciónfísica y química y masas de agua en los ca-nales australes de Chile entre boca del Guafoy golfo Elefantes (Crucero CIMAR 4 Fiordos).Cien. Tecnol. Mar, 25(2): 45-76.
HUISMAN, J., P. VAN OOSTVEEN & F. J. WEISSING.1999. Species dynamics in phytoplanktonblooms: Incomplete mixing and competitionfor light. The American Naturalist. 154: 46-68.
IRIGOIEN, X., J. HUISMAN & R. HARRIS. 2004. Glo-bal biodiversity patterns of marine phytoplanktonand zooplankton. Nature 863-867.
JEFFREY, S. W., R. F. C. MANTOURA & S. W.WRIGHT. 1997. Phytoplankton pigments inoceanography: methods. Monographs onOceanographic Methodology. SCOR andUNESCO.

84
KIORBOE, T. 1993. Turbulence, phytoplankton cellsize, and the structure of pelagic food webs.Advances in Marine Biology, 29.
KIRK, J. T. O. 1994. Light and Photosynthesis inAquatic Ecosystems. Second edition.Cambridge University Press, 509 pp.
LEHMAN, P. W. 2000. The influence of climateon phytoplankton community biomass in SanFrancisco Bay Estuar y. Limnology andOceanography, 45: 580-590.
LI, W. K. 2002. Macroecological patterns ofphytoplankton in the nor thwestern Nor thAtlantic Ocean. Nature, 154-157.
LINDENSCHMIDT, K. & I. CHORUS. 1998. The effectof water column mixing on phytoplankton suc-cession, diversity and similarity. Journal ofPlankton Research, 20: 1.927-1.951.
MARGALEF, R. 1978. Life forms of phytoplanktonas survival alternatives in an unstable envi-ronment. Oceanologica Acta 1: 493-509.
MARIE, D., PARTENSKY F. P., JACQUET S., VAULOTD. 1997. Enumeration and cell cycle analysisof natural populations of marine picoplanktonby flow cytometry using the nucleic acid stainSYBR Green I. Applied EnvironmentalMicrobiology, 63: 186-193.
MARIE, D., PARTENSKY F., VAULOT D., & C. BRUS-SAARD. 1999. Enumeration of phytoplankton,bacteria, and viruses in marine samples. In:Robinson J. P., Darzinkiewicz Z., Dean P. N.,Orfao A., Rabinovitch PS, Stewart C.C., TankeH.J., Wheeless L.L., editors. Current proto-cols in cytometry. Supplement 10. Unit 11.11.New York: John Wiley & Sons; 1999. p 1-15.
MITCHELL, G,, A. BRICAUD, K. CARDER, J. CLE-VELAND, G. FERRARI, R. GOULD, M. KAHRU,M. KISHINO, H. MASKE, T. MOISAN, L. MOO-RE, N. NELSON, D. PHINNEY, R. REYNOLDS,H. SOSIK, D. STRAMSKI, S. TASSAN, C.TREES, A. WEIDEMANN, J. WIELAND & A.VODACEK. 2000. Determination of spectralabsorption coefficients of particles, dissol-ved material and phytoplankton for discretewater samples. En: G. S. Fargion & J. L. Mue-ller (eds) Ocean Optics Protocols for satelliteOcean Color Sensor Validation, Revision2.125-153. NASA. Greenbelt, Maryland.
MONTECINO, V. & D. QUIROZ. 2000. Specificprimary production and phytoplankton cell size
structure in an upwelling area off the coastof Chile (30o S). Aquatic Sciences, 62: 364-380.
MONTECINO, V. 2002. Alometría y biodiversidaden fitoplancton en relación con la productivi-dad primaria en ecosistemas pelágicos. En:Sustentabilidad de la biodiversidad. K. Alveal& T. Antezana (eds).
MUÑOZ, P. & S. AVARIA. 1997. Fenómenos demarea roja y otras floraciones algales en Chi-le. Cienc. Tecnol. Mar, 20:175-192.
PAREDES, M. A. 2005. Alteraciones ambienta-les moduladoras de la diversidad alométricadel fitoplancton en fiordos y canales austra-les. Tesis. Magíster en Ciencias Biológicas,mención Ecología y Biología Evolutiva. Univer-sidad de Chile. Chile.
PIZARRO, G., J. L. IRIARTE, V. MONTECINO, J. L.BLANCO & L. GUZMÁN. 2000. Distribuciónde la biomasa fitoplanctónica y productividadprimaria máxima de fiordos y canales austra-les (47o - 50o S) en octubre 1996. Cienc.Tecnol. Mar, 23: 25-48.
PIZARRO, G., V. MONTECINO, L. GUZMÁN, V.MUÑOZ, V. CHACÓN, H. PACHECO, M. FRAN-GÓPULOS, L. RETAMAL & C. ALARCÓN.2005a. Patrones locales recurrentes del fito-plancton en fiordos y canales australes (46o -56o S) en primavera y verano. Cienc. Tecnol.Mar, 28 (2): 63-83.
PIZARRO, G., R. ASTORECA, V. MONTECINO, M.A. PAREDES, G. ALARCÓN, P. URIBE & L.GUZMÁN. 2005b. Patrones espaciales de laabundancia de la clorofila, su relación con laproductividad primaria y la estructura de ta-maños del fitoplancton en julio y noviembrede 2001 en la región de Aysén (43o 46’ S).Cienc. Tecnol. Mar, 28(2): 27-42.
PRANDLE D. 1985. On salinity regimes and thevertical structure of residual flows in narrowtidal estuaries. Estuarine, Coastal and ShelfScience, 20, 615-635.
REYNOLDS, C., I. PADISÁK & U. SOMMER. 1993.Intermediate disturbance in the ecology ofphytoplankton and the maintenance of spe-cies diversity: a synthesis. Hidrobiología. 249:183-188.
SALINAS, S., J. FIERRO, M. CASTILLO & J.LETELIER. 2002. Tiempo de residencia y pro-

85Gradientes de productividad y biomasa del fitoplancton en Aysén
cesos de mezcla en la cuenca sur de la cons-tricción de Meninea. Resultados CruceroCIMAR 7 Fiordos. Informes Preliminares. Co-mité Oceanográfico Nacional: 33-43.
SALINAS, S. & S. HORMAZÁBAL. 2004. Capaci-dad de transpor te de la constricción deMeninea para un flujo de dos capas y el efec-to de la corriente de marea. Cienc. Tecnol.Mar, 27(1): 5-15.
SILVA, N., C. CALVETE & H. SIEVERS. 1998a. Ma-sas de agua y circulación general para algunoscanales australes entre Puerto Montt y lagunaSan Rafael, Chile. (Crucero CIMAR-Fiordo 1).Cienc. Tecnol. Mar, 21: 17-8.
SILVA, N., J. MATURANA, J. SEPÚLVEDA & R.AHUMADA. 1998b. Materia orgánica, C y N,
su distribución y estequiometría, en sedi-mentos super ficiales de la región norte delos fiordos y canales australes de Chile (Cru-cero CIMAR-Fiordo 1). Cienc. Tecnol. Mar,21: 49-74.
VAULOT, D., COURTIES, C., & F. PARTENSKY.1989. A simple method to preserve oceanicphytoplankton for flow cytometric analyses.Cytometry 10, 629-635.
VERA, P., S. AVARIA & P. MUÑOZ. 1996. Composi-ción y distribución del fitoplancton de los fior-dos adyacentes a Campos de Hielo Sur y surelación con algunos parámetros oceanográfi-cos. Cienc. Tecnol. Mar, 19: 73-92.
WILKINSON, L. 1989. SYSTAT: The System forStatistics. Evanston, IL. Systat, Inc., 638 pp.