gut lactobacillus acidophilus to cellspent broth culture supernatant or spent broth culture...
TRANSCRIPT

Gut 1994; 35: 483-489
Lactobacillus acidophilus LA 1 binds to culturedhuman intestinal cell lines and inhibits cellattachment and cell invasion by enterovirulentbacteria
M F Bernet, D Brassart, J R Neeser, A L Servin
AbstractFour human Lactobacillus acidophilus strainswere tested for their ability to adhere ontohuman enterocyte like Caco-2 cells in culture.The LA 1 strain exhibited a high calciumindependent adhesive property. This adhesiononto Caco-2 cells required a proteinaceousadhesion promoting factor, which was presentin the spent bacterial broth culture superna-tant. LA 1 strain also strongly bound to themucus secreted by the homogeneous culturedhuman goblet cell line HT29-MTX. The inhib-itory effect of LA 1 organisms against Caco-2cell adhesion and cell invasion by a largevariety of diarrhoeagenic bacteria was investi-gated. As a result, the following dose depen-dent inhibitions were obtained: (a) against thecell association of enterotoxigenic, diffuselyadhering and enteropathogenic Escherichiacoli, and Salmonella typhimurium; (b) againstthe cell invasion by enteropathogenicEschericha coli, Yersinia pseudotuberculosis,and Salmonella typhimurium. Incubations ofL acidophilus LA 1 before and together withenterovirulent E coli were more effective thanincubation after infection byE coli.(Gut 1994; 35: 483-489)
Equipe de PathogenieMicrobienne Cellulaire etMoleculaire Intestinale,Departement deMicrobiologie, UFRSciencesPharmaceutiques, Paris,FranceM F BernetA L Servin
and Nestle ResearchCentre, Lausanne,SwitzerlandD BrassartJ R NeeserCorrespondence to:Dr A L Servin, Departementde Microbiologie etImmunologie, UFR SciencesPharmaceutiques Paris XI,92296 Chatenay-Malabry,France.Accepted for publication26 July 1993
Lactobacilli have been largely introduced intoseveral fermented dairy products. As recentlyunderlined by Klaenhammer,' lactobacillistrains with well defined properties should bescientifically selected and characterised forspecific use in commercial preparations. One ofthese properties should be the ability to adhere tomucosal surfaces, which could confer a competi-tive advantage, important for bacterial main-tenance in the human gastrointestinal tract.Recent reports show that selected strains, such asLactobacillus casei GG,2 Lactobacillus acidophilusBG2FO4,`' and LB6 express adhesive factors,which permit interactions with human entero-cyte like cells. Another desirable effect is apositive role of exogenous lactobacilli in preven-tion and treatment of gastrointestinal disorders.There is numerous evidence showing such nutri-tional and therapeutic benefits of lactobacilli.78To explain this effect, several authors suggestedthat exogenous lactobacilli could exert a 'barriereffect' against pathogens as does the indigenousmicroflora.78The aim of this study was to examine the
adhesion of lactobacilli of human origin tohuman intestinal cells. For this purpose we usedthe enterocyte like Caco-2 cell line,9"' which
displays typical features of enterocytic intestinalcells. " 2Bacterial attachment to the intestinal cellsurface was characterised by electron micro-scopic examination and by quantitative deter-mination. The mechanism through which anadherent L acidophilus strain interacts withCaco-2 cells was investigated. Moreover, it is wellknown that the polarised Caco-2 cell line canmimic the conditions in vivo where infectionoccurs, after cell association of enterotoxigenicbacteria'3'6 and cell invasion by enterovirulentbacteria'7 20 and rotavirus.2' Consequently, weexamined the inhibition of cell association andcell invasion ofpathogens participating in infant22and travellers' diarrhoea23 by adhering Lacidophilus.
Methods
BACTERIALSTRAINS,GROWTHCONDITIONS,ANDRADIOLABELLINGLactobacillus acidophilus LA 1, LA 3, LA 10, andLA 18 strains (Nestec collection, Lausanne,Switzerland) were grown in anaerobic conditions(Gaspack H2+CO2) in De Man, Rogosa, Sharpe(MRS) broth (Biokar, Pantin, France) 2 x24hours at 37°C before adhesion assay.The enterovirulent Escherichia coli strains used
in this study were the enterotoxigenic E coli(ETEC) H10407 expressing the CFA/I adhesivefactor,24 the enteropathogenic E coli (EPEC)JPN15 [pMAR7] (EAF+eae+) (Professor J BKaper, Center for Vaccine Development, Uni-versity ofMaryland, USA),'8 the diffusely adher-ing E coli (DAEC) C-1845, which belongs to theuropathogenic family (Dr S Bilge, WashingtonUniversity, Seattle, USA),2526 Yersinia pseudo-tuberculosis YPIII [pYV-] (inv+) (DrM Simonet,Faculte' Necker-Enfants Malades, Paris,France),27 and Salmonella typhimurium SL 1344(Professor B A D Stocker, Stanford, Cali-fornia). '7
Before adherence assays, ETEC, and DAECstrains were grown on CFA-agar containing 1%casamino acids (Difco Laboratories, Detroit,USA), 0d15% yeast extract, 0-005% magnesiumsulphate, and 0 0005% manganese chloride in2% agar for 18 hours at 37°C. EPEC bacteria werecultured at 37°C for 24 hours in luria broth with200 ,ug/ml ampicillin. S typhimurium wascultured at 37°C for 18 hours in luria broth.Y pseudotuberculosis was cultured 18 hours at28°C in luria broth.For radiolabelling, ETEC, DAEC, and EPEC
bacteria were subcultured twice at 37°C for 24
483
on February 19, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.35.4.483 on 1 A
pril 1994. Dow
nloaded from

Bernet, Brassart, Neeser, Servin
hours in luria broth. They were metabolicallylabelled by the addition of '4C-acetic acid(Amersham, UK (94 mCi/mmol); 100IuCi/1O mltube). Before radiolabelling, S typhimurium wascultured for four hours in luria broth andY pseudotuberculosis was cultured for 18 hours intrypticase soja broth; and for radiolabelling thebacteria were subcultured at 37°C for 45 minutesin methionine medium (Difco) with 35S-meth-ionine (Amersham, 1000 Ci/mmol, 20,uCi/ml).
CELL CULTUREEnterocyte like Caco-2 cells9 were obtained fromDr Jorgen Fogh (Sloan Kettering MemorialCancer Center, Rye, NY, USA). The homo-geneous mucus secreting subpopulation ofHT29-MTX cells28 was obtained from Dr AZweibaum (Unite 178, Institut National de laSante et de la Recherche Medicale, Villejuif,France).
Cells were routinely grown in Dulbecco mod-ified Eagle's minimal essential medium (25 mMglucose) (Eurobio, Paris, France), supplementedwith 10% (HT29-MTX) or 20% (Caco-2) inacti-vated (30 min; 56°C) fetal calf serum(Boehringer, Mannheim, Germany), and 1%non-essential amino acids (Caco-2). For adhesionassay, monolayers of Caco-2 cells and HT29-MTX cells were prepared on glass coverslips,which were placed in six well Corning tissueculture plates (Coming Glass Works, Corning,NY, USA). For inhibition of cell association andcell invasion of pathogens, monolayers of Caco-2cells were prepared in 20 four well tissue cultureplates (Corning Glass Works). Cells were seededat a concentration of 2 x 104 (HT29-MTX) and1-4x 104 (Caco-2) cells/cm2. Maintenance of thecells and all experiments were carried out at 37°Cin a 10% CO190% air atmosphere. The culturemedium was changed daily. Mucus secretingHT29-MTX and Caco-2 cells were used between20-40 and 60-90 cell passages, respectively.Cells were used for adherence assays at late post-confluence - that is, after 15 days (Caco-2) and 20days (HT29-MTX) in culture.
ADHERENCE ASSAYThe adherence ofL acidophilus strains to Caco-2and HT29-MTX cells was examined as describedpreviously for theL acidophilus strain LB and theL acidophilus strain BG2FO4 adhesion assay.56Briefly, the, Caco-2 and HT-29 monolayers,prepared on glass coverslips, which were placedin six well Corning tissue culture plates (CorningGlass Works), were washed twice with phosphatebuffered saline. L acidophilus (1 ml, 4x 108bacteria/ml) in spent culture supernatant, treatedsupernatant or fresh MRS broth were added to 1ml of the cell line culture medium. This suspen-sion (2 ml) was added to each well of the tissueculture plate and the plate incubated at 370C in10% CO]90% air. After one hour of incubation,the monolayers were washed five times withsterile phosphate buffered saline, fixed withmethanol, stained with Gram stain, and exam-ined microscopically. Each adherence assay wasconducted in triplicate over three successivepassages of intestinal cells. For each monolayer
on a glass coverslip, the number of adherentbacteria was evaluated in 20 random microscopicareas. Adhesion were evaluated by two differenttechnicians to eliminate bias.
PHYSICAL AND CHEMICAL TREATMENTS OFBACTERIA AND SPENT CULTURE SUPERNATANTTo characterise the bacterial determinants par-ticipating inL acidophilus adhesion, bacteria withspent broth culture supernatant or spent brothculture supernatant alone were subjected todifferent treatments, as previously described.56All enzymes and chemicals were obtained fromSigma Chemical Co (St Louis, USA). Bacterialcells and spent culture supernatant were sepa-rated by centrifugation (20 000 g, 1 h at 4°C).Bacterial cells and spent broth culture super-natant or spent broth culture supernatant alonewere incubated with trypsin (2'5 mg ml-') for 60minutes at 37°C; trypsin was inactivated byadding inactivated (30 min, 56°C) fetal bovineserum (Boehringer, Mannheim, Germany). Todetermine the influence of calcium on adherenceof L acidophilus,3 the monolayers were washedfive times with the calcium chelating agent[ethylene-bis-(oxyethylenenitrilo)] tetraacetic acid(EGTA 20 mM) in phosphate buffered salineafter the incubation period with bacteria.
SCANNING ELECTRON MICROSCOPYFor scanning electron microscopy, the tissueculture cells were grown on glass coverslips.After the bacterial adhesion assay, cells werefixed with 2-5% glutaraldehyde in 0-1 M phos-phate buffer (pH 7-4) for one hour at roomtemperature. After two washes with phosphatebuffer, cells were postfixed for 30 minutes with2% OsO4 in the same buffer, washed three timeswith phosphate buffer, dehydrated in a gradedseries (30%, 50%, 70%, 80%, 90%, 100%) ofethanol, and passaged through in a graded series(50%, 70%, 90%, 100%) of amyl acetate. Cellswere dried in a critical point dryer (BalzersCPDO30), and coated with gold. The specimenswere then examined with a Jeol JSM 25S scan-ning electron microscope.
ADHESION AND INVASION INHIBITION ASSAYBefore the adhesion and invasion inhibitionassays, the Caco-2 monolayers were washed twicewith phosphate buffered saline.To determine Caco-2 cell associated bacteria
(adhering and invading bacteria) radiolabelledbacteria were used. Bacteria were suspended inthe culture medium and 1 ml (108 CFU cells/ml)of this suspension was added to each well of thetissue culture plates. For ETEC, DAEC, andEPEC, incubations were conducted in the pres-ence of 1% D-mannose that inhibits adhesionmediated by type 1 pili. The plates were incu-bated at 370C in 10% CO2/90% air, 60 minutesfor ETEC, DAEC, and Salmonella, 180 minutesfor EPEC. The monolayers were then washedthree times with sterile phosphate bufferedsaline. Cell associated bacteria and intestinal cellswere dissolved in a 0-2 N NaOH solution. The
484
on February 19, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.35.4.483 on 1 A
pril 1994. Dow
nloaded from

L acidophilus LA I inhibits interactions ofpathogens with intestinal cells
TABLE I Adhesion of Lactobacillus acidophilus strainsonto human intestinal epithelial Caco-2 cells in culture.Determination ofcalcium dependent and independentbindings
Adhesion*
Lactobacilli strains WithoutEGTA With 20 mM EGTA
L acidophilus LA 1 155 (26) 63 (5)L acidophilus LA 3 66 (21) 18 (9)L acidophilus LA 10 18 (3) 4(2)L acidophilus LA 18 23 (7) 5 (3)
*Adhesion of lactobacilli strains onto monolayers of differentiatedCaco-2 cells is expressed as a mean number (SEM) of lactobacilliadhering to the cell monolayer/100 Caco-2 cells. Twentyrandomised microscopic fields/cover slip were counted. Eachadherence assay was conducted in triplicate with three successiveCaco-2 cell passages. EGTA=(Ethylene-bis-(oxyethylenenitrilo))tetraacetic acid.
rate of bacterial adhesion was evaluated by liquidscintillation counting.
Bacterial internalisation was assessed by quan-titative determination of bacteria located withinthe Caco-2 cells using unlabelled bacteria and anaminoglycoside antibiotic. Bacteria were sus-pended in the culture medium and 2 ml (10k
Figure 1: Examination by scanning electron microscopy ofadherence ofLactobacillusacidophilus strain 1 onto the differentiated human intestinal epithelial cells Caco-2. (A) Lowmagnification ofCaco-2 monolayer covered by L acidophilus I bacteria; (B) Highmagnification ofL acidophilus 1 whole cells.
CFU/ml) of this suspension were added to eachwell of the tissue culture plate. The plates wereincubated at 37°C in 10% COI90% air, 180minutes for EPEC, 90 minutes for Yersinia, and60 minutes for Salmonella. For Yersinia, contactwith Caco-2 cells was increased by centrifugationfor 10 minutes at 123 g. After incubation theplates were washed five times with sterile phos-phate buffered saline and then incubated for 120minutes in a medium containing 20-100 ,ug ofgentamicin/ml. As gentamicin does not diffuseacross apical domain ofCaco-2 cells, bacteria thatadhere to the Caco-2 brush border were rapidlykilled, whereas those located within Caco-2 cellswere not. The monolayer was washed withphosphate buffered saline and lysed with ster-ilised water. Appropriate dilutions were plated todetermine the number of viable intracellularbacteria.
Inhibition ofadhesion or invasion ofpathogensby lactobacilli were evaluated as previouslydescribed.' Briefly, 1 ml of radiolabelled patho-genic bacteria (adhesion and invasion) orunlabelled bacteria (invasion) (10k CFU/ml) and1 ml of L acidophilus with spent culture super-natant (109 to 107 CFU/ml as indicated) wereadded together to each well of the tissue cultureplate and incubated as previously described.Each assay was conducted in triplicate with threesuccessive passages of Caco-2 cells.
Results
ADHESION OF L ACIDOPHILUS STRAINS TO HUMANCULTURED INTESTINAL CELL LINES ANDCHARACTERISTICS OF ADHESIONFour human Lactobacillus acidophilus strainswere examined for their ability to adhere tocultured enterocyte like Caco-2 cells (Table I).L acidophilus LA 1 and LA 3 possessed a highcalcium independent capacity of adhesion,whereas low capacities were expressed byL acidophilus LA 10 and LA 18 strains. Thisresult agrees with a previous finding showing thatadhesive properties vary considerably betweenlactobacilli strains.6 As seen by scanning electronmicroscopy, the LA 1 strain showed diffuseadhesion to Caco-2 cells (Fig 1), as do otheradhering L acidophilus strains.56 Because, in thehuman intestine, two main cell phenotypes - thatis, enterocytes and goblets cells - are represented,we examined the binding ofLA 1 strain to humanmucus secreting intestinal cells in culture.L acidophilus LA 1 strongly bound in a diffusepattern to the mucus secreted by the homo-geneous subpopulation ofHT29-MTX cells (Fig2). The rate of adhesion of LA 1 whole cells tomucus secreting cells and to the mucus layerseemed higher than adhesion to enterocyte likeCaco-2 cells.As recently seen, adhering lactobacilli strains
secrete extracellular adhesions to attach tohuman56 and murine3m intestinal cells. In anattempt to identify the components participatingin adhesion of L acidophilus LA 1 to humanintestinal cells in culture, we subjected the LA 1culture to several treatments (Table II). Whenthe spent culture supernatant was discarded andreplaced by a fresh culture medium, a dramatic
485
on February 19, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.35.4.483 on 1 A
pril 1994. Dow
nloaded from

Bernet, Brassart, Neeser, Servin486
0 ilk,' v. '
^ li. -%~ .. .4-i r
,- --x-s_Aic t. J .. 7...W - -.,jwk
Figure 2: Adhesion ofLactobacillus acidophilus strain 1 tomucus secretingHT29-MTX monolayer seen by low andhigh magnification scanning electron microscopy. (A) and (B)Low and high magnification ofHT-29 cell monolayer.Notice that brush border ofthe HT29-MTX cells is notvisible because it is entirely covered by the dense mucus gel;(C) Observation ofhigh level ofadhesion with Lacidophilus strain 1.
loss of adhesion occurred. Trypsin treatment ofthe spent culture supernatant totally abolishedthe adhesive capacity of L acidophilus LA 1.These results show that an extracellular pro-
teinaceous component secreted in the bacterialspent culture supernatant participates in theadhesion ofLA 1 to intestinal cells.
INHIBITION OF PATHOGENS CELL ASSOCIATIONAND CELL INVASION WITH AND WITHIN CACo-2CELLS BY L ACIDOPHILUS LA 1Diarrhoeagenic Escherichia coli 1416 18 31 andSalmonella typhimurium ` interact with the brushborder of differentiated cultured human intes-tinal cell lines. By contrast, Yersinia pseudotuber-culosis interacts only with cell periphery ofundifferentiated Caco-2 cells (A Servin et al,unpublished data), as does Listeria mono-
cytogenes.19We examined the inhibition of cell association
TABLE II Characteristics ofadhesion of Lactobacillusacidophilus LA 1 strain
Adhesion score
Adhesions: With spent culture supernatant 124 (13)With fresh culture medium* 9 (3)Trypsint 0-6
Adhesion is expressed as a mean number (SEM) of lactobacilliadhering to the cell monolayer/100 Caco-2 cells, determined in20 randomised microscopic fields/cover slip. Each adherenceassay was conducted in triplicate with three successive Caco-2 cellpassages. *To determine the presence of adhesive factors secretedby the bacteria in the spent culture supernatant, it was replacedby a fresh culture medium before adhesion assay; tTo furthercharacterise the bacterial determinants participating inL acidophilus LA 1 adhesion, bacteria with spent broth culturesupernatant was subjected to trypsin treatment (2-5 mg/ml) for60 miin at 37C, then trypsin was inhibited by adding inactivated(30 min, 56°C) fetal bovine serum.
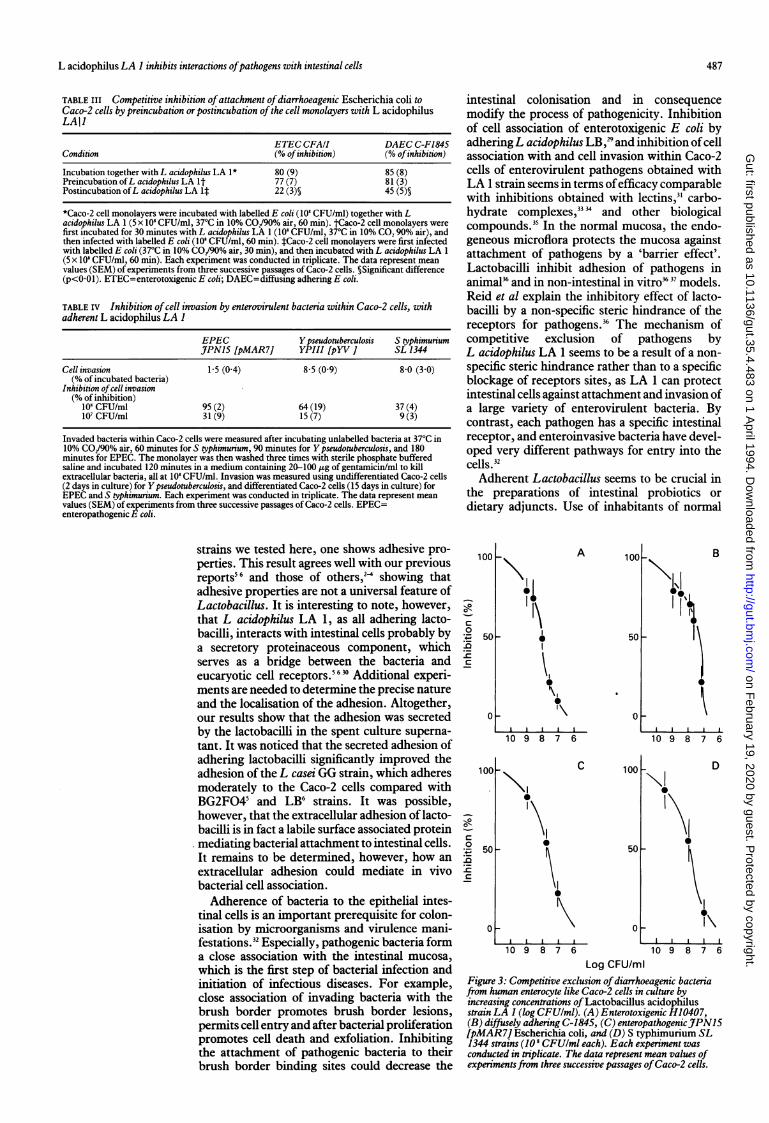
and cell entry of several enterovirulent bacteriabyL acidophilus LA 1 (Tables III and IV, and Fig3). By using radiolabelled bacteria (108 CFU/ml),we evaluated the cell association (adhering andinvading bacteria) of enterotoxigenic (ETECCFA/I), diffusely adhering (DAEC C-1845), andenteropathogenic (EPEC JPN15 [pMAR7])E coli, and S typhimurium (SL 1344) strains withCaco-2 cells. The bacterial entry of these EPEC,S typhimurium, and Y pseudotuberculosis (YPIII[pYV-1) strains into Caco-2 cells was measuredusing unlabelled bacteria (108 CFU/ml) andgentamicine, which selectively kills the non-internalised bacteria.L acidophilus LA 1 strain efficiently inhibited
cell association of these ETEC, DAEC, EPEC,and S typhimurium strains with Caco-2 cells in adose dependent fashion (Fig 3). For all patho-gens, 50% inhibition of cell association wasobtained with 101 lactobacillml. We examineddifferent conditions of bacterial incubation tofurther characterise the inhibitory action ofL acidophilus LA 1 (Table III). When LA 1organisms were incubated with Caco-2 cellsbefore the infection with ETEC or together withETEC, an identical inhibition of E coli cellassociation was seen. By contrast, a significantdecrease of efficacy was seen when the pathogenswere incubated with Caco-2 cells before addingthe L acidophilus LA 1 bacteria (Table III,p<OOl).
Invasion of Caco-2 cells by selectedY pseudotuberculosis, S typhimurium, and EPECstrains was inhibited in a dose dependent way byL acidophilus LA 1 (Table IV). This strain at 108CFU/ml inhibited more efficiently Caco-2 cellsinvasion by EPEC and Y pseudotuberculosis, thanthat by S typhimurium.
DiscussionAs previously reported, the polarised humanintestinal epithelial cells Caco-2 could be used topromote the selection of Lactobacillus strains.2 5 6
This stable and reproducible model permitsevaluation of the comparative adherence capaci-ties of these strains in the human intestine, aproperty that is thought to be essential forintestinal maintenance or colonisation. More-over, this model provides an excellent system tocharacterise the manner by which lactobacilliinteract with a well defined enterocytic brushborder.56 Among the four human L acidophilus
on February 19, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.35.4.483 on 1 A
pril 1994. Dow
nloaded from
L acidophilus LA I inhibits interactions ofpathogens with intestinal cells
TABLE III Competitive inhibition ofattachment ofdiarrhoeagenic Escherichia coli toCaco-2 cells by preincubation or postincubation ofthe cell monolayers with L acidophilusLAI]
ETEC CFAII DAEC C-F1845Condition (% of inhibition) (% ofinhibition)
Incubation together with L acidophilus LA 1* 80 (9) 85 (8)Preincubation ofL acidophilus LA It 77 (7) 81(3)Postincubation ofL acidophilus LA If 22 (3)§ 45 (5)§
*Caco-2 cell monolayers were incubated with labelled E coli (108 CFU/ml) together with Lacidophilus LA 1 (5 x 1O0 CFU/ml, 37'C in 10% CO/90% air, 60 min). tCaco-2 cell monolayers werefirst incubated for 30 minutes with L acidophilus LA 1 (108 CFU/ml, 37'C in 10% CO, 90% air), andthen infected with labelled E coli (108 CFU/ml, 60 min). tCaco-2 cell monolayers were first infectedwith labelled E coli (37'C in 10% CO,/90% air, 30 min), and then incubated with L acidophilus LA 1(5 x 108 CFU/ml, 60 min). Each experiment was conducted in triplicate. The data represent meanvalues (SEM) of experiments from three successive passages of Caco-2 cells. §Significant difference(p<0-01). ETEC=enterotoxigenic E coli; DAEC=diffusing adhering E coli.
TABLE IV Inhibition of cell invasion by enterovirulent bacteria within Caco-2 cells, withadherent L acidophilus LA I
EPEC Ypseudotuberculosis S typhimuriumJPN15 [pMAR7] YPIII [pYV ] SL 1344
Cell invasion 1-5 (0-4) 8-5 (0-9) 8-0 (3 0)(% of incubated bacteria)
Inhibition ofcell invasion(% of inhibition)
lO CFU/ml 95 (2) 64 (19) 37 (4)10' CFU/ml 31 (9) 15 (7) 9 (3)
Invaded bacteria within Caco-2 cells were measured after incubating unlabelled bacteria at 37'C in10% CO}90% air, 60 minutes for S typhimurium, 90 minutes for Y pseudotuberculosis, and 180minutes for EPEC. The monolayer was then washed three times with sterile phosphate bufferedsaline and incubated 120 minutes in a medium containing 20-100 ,ug of gentamicin/ml to killextracellular bacteria, all at lO CFU/mI. Invasion was measured using undifferentiated Caco-2 cells(2 days in culture) for Ypseudotuberculosis, and differentiated Caco-2 cells (15 days in culture) forEPEC and S typhimurium. Each experiment was conducted in triplicate. The data represent meanvalues (SEM) of experiments from three successive passages of Caco-2 cells. EPEC=enteropathogenic E coli.
strains we tested here, one shows adhesive pro-perties. This result agrees well with our previousreports" and those of others,2" showing thatadhesive properties are not a universal feature ofLactobacillus. It is interesting to note, however,that L acidophilus LA 1, as all adhering lacto-bacilli, interacts with intestinal cells probably bya secretory proteinaceous component, whichserves as a bridge between the bacteria andeucaryotic cell receptors."3 Additional experi-ments are needed to determine the precise natureand the localisation of the adhesion. Altogether,our results show that the adhesion was secretedby the lactobacilli in the spent culture superna-tant. It was noticed that the secreted adhesion ofadhering lactobacilli significantly improved theadhesion of the L casei GG strain, which adheresmoderately to the Caco-2 cells compared withBG2F045 and LB6 strains. It was possible,however, that the extracellular adhesion of lacto-bacilli is in fact a labile surface associated proteinmediating bacterial attachment to intestinal cells.It remains to be determined, however, how anextracellular adhesion could mediate in vivobacterial cell association.Adherence of bacteria to the epithelial intes-
tinal cells is an important prerequisite for colon-isation by microorganisms and virulence mani-festations.32 Especially, pathogenic bacteria forma close association with the intestinal mucosa,which is the first step of bacterial infection andinitiation of infectious diseases. For example,close association of invading bacteria with thebrush border promotes brush border lesions,permits cell entry and after bacterial proliferationpromotes cell death and exfoliation. Inhibitingthe attachment of pathogenic bacteria to theirbrush border binding sites could decrease the
intestinal colonisation and in consequencemodify the process of pathogenicity. Inhibitionof cell association of enterotoxigenic E coli byadheringL acidophilus LB,29 and inhibition ofcellassociation with and cell invasion within Caco-2cells of enterovirulent pathogens obtained withLA 1 strain seems in terms ofefficacy comparablewith inhibitions obtained with lectins,3' carbo-hydrate complexes,33 34 and other biologicalcompounds.35 In the normal mucosa, the endo-geneous microflora protects the mucosa againstattachment of pathogens by a 'barrier effect'.Lactobacilli inhibit adhesion of pathogens inanimal36 and in non-intestinal in vitro 37 models.Reid et al explain the inhibitory effect of lacto-bacilli by a non-specific steric hindrance of thereceptors for pathogens. ' The mechanism ofcompetitive exclusion of pathogens byL acidophilus LA 1 seems to be a result of a non-specific steric hindrance rather than to a specificblockage of receptors sites, as LA 1 can protectintestinal cells against attachment and invasion ofa large variety of enterovirulent bacteria. Bycontrast, each pathogen has a specific intestinalreceptor, and enteroinvasive bacteria have devel-oped very different pathways for entry into thecells.32Adherent Lactobacillus seems to be crucial in
the preparations of intestinal probiotics ordietary adjuncts. Use of inhabitants of normal
._
c
0
.0
.C_
.-
-0
c
A 100
50k
0
B
4-I I'
50 F
0I I I
10 9 8 7 6
Cloo00
50k
0
100
50
0I I
10 9 8 7 6
10 9 8 7 6
10N9I 8 7 6
10 9 8 7 6
Log CFU/mlFigure 3: Competitive exclusion ofdiarrhoeagenic bacteriafrom human enterocyte like Caco-2 cells in culture byincreasing concentrations of Lactobacillus acidophilusstrain LA I (log CFUlml). (A) Enterotoxigenic H10407(B) diffusely adhering C-1845, (C) enteropathogenicJ7PNl5[pMAR7] Escherichia coli, and (D) S typhimurium SL1344 strains (10s CFUlml each). Each experiment wasconducted in triplicate. The data represent mean values ofexperimentsfrom three successive passages ofCaco-2 cells.
'I
487
on February 19, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.35.4.483 on 1 A
pril 1994. Dow
nloaded from

488 Bernet, Brassart, Neeser, Servin
human microflora, such as lactobacilli andbifidobacteria, in fermented milk products couldaid in preventing intestinal infections. Althoughcontroversial,78 the protective role of lactobacilliagainst intestinal colonisation by pathogenic bac-teria has recently gained more credibility. Con-sistent findings using well characterisedintestinal cell models2"29 have brought insightson the action oflactobacilli. Indeed, reports showthat adhering lactobacilli survive in the humanstomach38 and that the adherentL caseiGG strain2used in fermented milk products is maintained inthe human intestine after oral administration.39Moreover, recent controlled clinical studies haveproduced convincing evidence concerning thebeneficial effects ofadherent lactobacilli.' In thisreport, we found that in vitro incubation ofL acidophilus before infection with pathogensprovided an important inhibition of pathogenattachment, similar to that obtained by incuba-tion ofpathogens together withL acidophilus. Bycontrast, when the infection of the intestinalCaco-2 cell monolayers by pathogens camebefore the incubation ofL acidophilus, a decreaseof efficacy occurs. This result strongly suggests apreventive role of lactobacilli against intestinalinfections.
In conclusion, the results presented hereclearly show that L acidophilus LA 1 inhibits thecell association and the cell entry of some entero-pathogens present in diarrhoea. The precisemechanism, however, for the inhibitory effect ofthe Lactobacillus has not been fully clarified. Itcould result from (a) the non-specific sterichindrance of the apical enterocytic receptors ofpathogens by the Lactobacillus whole cells, (b)the antimicrobial action of a Lactobacillussecreted molecule with a large spectre of action(bacteriocin like activity), (c) the stimulation ofsecretion by intestinal cells of an antimicrobialsubstance (defensin like activity), after Lac-tobacillus cell binding. Further experiments arerequired to elucidate the nature of the mecha-nism(s) of this event.This work was supported by a grant INSERM-Industrie N°90025. MFB is supported by a doctoral fellowship of Ministere dela Recherche, de la Technologie et de l'Espace. We thank AZweibaum (INSERM U 178, Villejuif, France) for providing uswith Caco-2 and HT29-MTX cell lines. We thank D Guillaumin(Service Microscopie Electronique, CNRS Universite Pierre etMarie Curie Paris VI) for the technical assistance with the electronmicroscopy study.
1 Klaenhammer TR. Microbial considerations in selection andpreparation of Lactobacillus strains for use as dietaryadjuncts. JDairySci 1982; 65: 1339-49.
2 Elo S, Salminen S. Attachment ofLactobacillus casei strainGGto human colon carcinoma cell line Caco-2: comparison withother dairy strains. LettApplMicrobiol 1991; 13: 154-6.
3 Kleeman EG, Klaenhammer TR. Adherence of Lactobacillusspecies to human fetal intestinal cells. J Dairy Sci 1982; 65:2063-9.
4 Hood SK, Zottola EA. An electron microscopic study of theadherence of Lactobacillus acidophilus to human intestinalcells in vitro. Food Microstructure 1989; 8: 91-7.
5 Coconnier MH, Klaenhammer TR, Kerneis S, Bernet MF,Servin AL. Protein-mediated adhesion Lactobacillusacidophilus strain BG2FO4 on human enterocyte andmucus-secreting cell lines in culture. Appl Environ Microbiol1992; 58: 2034-9.
6 Chauviere G, Coconnier MH, Kerneis S, Fourniat J, ServinAL. Adherence of human Lactobacillus acidophilus ontohuman enterocyte-like cells, Caco-2 and HT-29 in culture.J GenMicrobiol 1992;-138:1689-96.
7 Conway P. Lactobacilli: Fact and fiction. In: Grun R, MidvedtT, Normn E, eds. The regulatory and protective role ofthe normalflora. Stockton Press, 1988: 263-81.
8 Reid G, Bruce AW, McGroarty JA, Cheng KJ, Costerton JW.Is there a role for Lactobacilli in prevention of urogenital andintestinal infection. Glin Microbiol Rev 1990; 3: 335-44.
9 Fogh J, Fogh JM, Orfeo T. One hundred and twenty seven
cultured human tumor cell lines producing tumors in nudemice. JNatl Cancer Inst 1977; 59: 221-6.
10 Pinto M, Robine-Leon S, Appay MD, Kedinger M, TriadouN, Dussaulx E, et al. Enterocyte-like differentiation andpolarization of the human colon carcinoma cell line Caco-2 inculture. Biol Cell 1983; 47: 323-30.
11 Zweibaum A, Laburthe M, Grasset E, Louvard D. Use ofcultured cell lines in studies of intestinal cell differentiationand function. In: Schultz SJ, Field M, Frizell RA, eds.Handbook of physiology. The gastrointestinal system vol IV.Bethesda: American Physiological Society, 1991: 223-55.
12 Louvard D, Kedinger M, Hauri HP. The differentiatingintestinal epithelial cell: establishment of functions throughinteractions between cellular structures. Annu Rev Cell Biol1992; 8: 157-95.
13 Aubel D, Darfeuille-Michaud A, Joly B. New adhesive factor(Antigen 8786) on a human enterotoxigenic Escherichia coli01 17:H4 strain isolated in Africa. Infect Immun 1991; 59:1290-9.
14 Darfeuille-Michaud A, Aubel D, Chauviere G, Rich C,Bourges M, Servin A, et al. Adhesion of enterotoxigenicEscherichia coli to the human colon carcinoma cell lineCaco-2 in culture. Infect Immun 1990; 58: 893-902.
15 Kernis S, Bilge S, Fourel V, Chauviere G, Coconnier MH,Servin AL. Use of purified F1845 fimbrial adhesin to studylocalization and expression of receptors for diffusely adher-ing Escherichia coli (DAEC) during enterocytic differentia-tion of human colon carcinoma cell lines HT-29 and Caco-2in culture. Infect Immun 1991; 59: 4013-8.
16 Kerneis S, Chauviere G, Darfeuille-Michaud A, Aubel D,Coconnier MH, Joly B, et al. Expression of receptors ofenterotoxigenic Escherichia coli during enterocytic differen-tiation of human intestinal epithelial cells, Caco-2 andHT-29, in culture. Infect Immun 1992; 60: 2572-80.
17 Finlay BB, Falkow S. Salmonella interactions with polarizedhuman intestinal Caco-2 epithelial cells. J Infect Dis 1990;162: 1096-106.
18 Francis CL, Jerse AK, Kaper JB, Falkow S. Characterizationof interactions of enteropathogenic Escherichia coli0127:H6 with mammalian cells in vitro. J Infect Dis 1991;164: 693-703.
19 Gaillard JL, Berche P, Mounier J, Richard S, Sansonetti JP. Invitro model of penetration and intracellular growth ofListeria monocytogenes in the human enterocyte-like cellline Caco-2. Infect Immun 1987; 55: 2822-9.
20 Mounier J, Vasselon T, Hellio R, Lesourd M, Sansonetti PJ.Shigella flexneri enters human colonic Caco-2 epithelial cellthrough the basolateral pole. Infect Immun 1992; 60: 237-48.
21 Svensson L, Finlay BB, Bass D, VonbonsdorffCH, GreenbergHB. Symmetric infection of rotavirus on polarized humanintestinal epithelial Caco-2 cells. J Virol 1991; 65: 4190-7.
22 Rubino A. Secretory diarrhea in infants and children. In:Lebenthal E, ed. Textbook ofgastroenterology and nutrition ininfancy. New York: Raven Press, 1989: 1159-70.
23 Okhuysen PC, Ericsson CD. Travelers' diarrhea. TravelMedicine 1992; 76: 1357-73.
24 Evans DG, Evans Jr DJ, Clegg S, Pauley JA. Purification andcharacterization of the CFA/I antigen of enterotoxigenicEscherichia coli. Infects Immun 1987; 25: 738-48.
25 Bilge SS, Clausen CR, Lau W, Moseley SL. Molecularcharacterization of a fimbrial adhesin, F1845, mediatingdiffuse adherence of diarrhea-associated Escherichia coli toHEp-2 cells. J Bacteriol 1989; 171: 4281-9.
26 Giron JA, Jones T, Millan-Velasco F, Castro-Munoz E, ZarateL, Fry J, et al. Diffuse-adhering Escherichia coli (DAEC) asa putative cause of diarrhea in Mayan children in Mexico.JIInfectDis 1991; 163: 507-13.
27 Isberg RR, Voorhis DL, Falkow S. Identification of invasin: Aprotein that allows enteric bacteria to penetrate culturedmammalian cells. Cell 1987; 50: 769-78.
28 Lesuffleur T, Barbat A, Dussaulx E, Zweibaum A. Growthadaptation to methotrexate of HT-29 human colon car-cinoma cells is associated with their ability to differentiateinto columnar absorptive and mucus-secreting cells. CancerRes 1990; 50: 6334-43.
29 Chauviere G, Coconnier MH, Kerneis S, Darfeuille-MichaudA, Joly B, Servin AL. Competitive exclusion of diar-rheagenic Escherichia coli (ETEC) from enterocyte-likeCaco-2 cells in culture. FEMS Microbiol Lett 1992; 91:213-8.
30 Conway PL, Kjelleberg S. Protein-mediated adhesion ofLactobacillus frementum strain 737 to mouse stomachsquamous epithelium. J Gen Microbiol 1989; 135: 1175-86.
31 Neeser JR, Chambaz A, Golliard M, Link-Amster H, FryderV, Kolodziejczyk E. Adhesion of colonization antigen factorantigen II-positive enterotoxigenic Escherichia coli strains tohuman enterocyte-like differentiated HT-29 cells: a basis forhost-pathogen interactions in the gut. InfectImmun 1989; 57:3727-34.
32 Falkow S, Isberg RR, Portnoy DA. The interaction of bacteriawith mammalian cells. Annu Rev Cell Biol 1992; 8: 333-63.
33 Neeser JR, Chambaz A, Hoang KY, Link-Amster H. Screen-ing for complex carbohydrates inhibiting hemagglutinationsby CFA/I- and CFA/II-expressing enterotoxingenicEscherichia coli strains. FEMS Microbiol Lett 1988; 49:301-7.
34 Cravioto A, Tello A, Villajan H, Ruiz J, del Vedovo 5, NeeserJR. Inhibition of localized adhesion of enteropathogenicEscherichia coli to HEp-2 cells by immunoglobulin ofoligosaccharide fractions of human colostrum and breastmiLk.JlInfeceDis 1991;-163:1247-55.
35 Reid G, Cook RL, Bruce AWV. Examination of strains oflactobacilli for properties that may influence bacterial inter-ference in the urinary tract. I Urol 1987; 138: 330-5.
on February 19, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.35.4.483 on 1 A
pril 1994. Dow
nloaded from

L acidophilus LA I inhibits interactions ofpathogens with intestinal cells 489
36 Gotteland M, Pochart P, Dabbech M, Bisetti N, Desjeux JF.In vivo effect of yogurt on excretion of enteropathogenEscherichia coli RDEC-1 during acute diarrhea in the just-weaned rabbit.J Pediat Gastroenterol 1992; 14: 264-7.
37 Blomberg L, Henriksson A, Conway PL. Inhibition ofadhesion of Escherichia coli K88 to piglet ileal mucus byLactobacillus spp. Appl Environ Microbiol 1992; 59: 43-9.
38 Conway PL, Gorbach SL, Goldin BR. Survival of lactic acid
bacteria in the human stomach and adhesion to intestinalcells. JDairySci 1987; 70: 1-12.
39 Goldin BR, Sherwood L, Saxelin M, Barakat S, Gualteri L,Salminen S. Survival of Lactobacillus species (Strain GG) inhuman gastrointestinal tract. Dig Dis Sci 1992; 37: 121-8.
40 Isolauri E, Juntunen M, Rautanen T, Sillanaukee P, KoivulaTA. A human Lactobacillus strain (Lactobacillus casei spstrain GG) promotes recovery from acute diarrhea inchildren. Pediatrics 1991; 88: 90-7.
on February 19, 2020 by guest. P
rotected by copyright.http://gut.bm
j.com/
Gut: first published as 10.1136/gut.35.4.483 on 1 A
pril 1994. Dow
nloaded from