hawthorns (crataegus, rosaceae) stomata size in relation · pdf filestomata size in relation...
TRANSCRIPT

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
Stomata Size in Relation to Ploidy Level in North AmericanHawthorns (Crataegus, Rosaceae)Author(s): Brechann V. McGoey Kelvin Chau Timothy A. DickinsonSource: Madroño, 61(2):177-193. 2014.Published By: California Botanical SocietyDOI: http://dx.doi.org/10.3120/0024-9637-61.2.177URL: http://www.bioone.org/doi/full/10.3120/0024-9637-61.2.177
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated contentindicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

STOMATA SIZE IN RELATION TO PLOIDY LEVEL IN NORTH AMERICANHAWTHORNS (CRATAEGUS, ROSACEAE)
BRECHANN V. MCGOEY
Department of Ecology and Evolutionary Biology, University of Toronto, Toronto, ON,Canada M5S 3B2
KELVIN CHAU
Canadian Food Inspection Agency, 1124 Finch Ave. W, Unit 2, Toronto, ON,Canada M3J 2E2
TIMOTHY A. DICKINSON
Green Plant Herbarium (TRT), Department of Natural History, Royal Ontario Museum,100 Queen’s Park, Toronto, ON, Canada M5S 2C6, and Department of Ecology and
Evolutionary Biology, University of Toronto, Toronto, ON, Canada M5S 3B2
ABSTRACT
The impacts of ploidy level changes on plant physiology and ecology present interesting avenues ofresearch, and many questions remain unanswered. Here, we examine the connections betweencytotype, taxon, stomata characteristics, and environmental variables in black-fruited hawthorns ofthe Pacific Northwest (Crataegus ser. Douglasianae; Maleae, Amygdaloideae, Rosaceae). We explorethe extent to which stomatal measurements can be used to predict ploidy level and how differences inploidy level and stomata characteristics relate to geographic distributions. We sampled trees fromacross the geographic ranges of the putative sister taxa Crataegus suksdorfii (Sarg.) Kruschke (diploidsand autotriploids) and C. douglasii Lindl. (tetraploids). We found that stomata differed between thetwo species, with tetraploid C. douglasii having larger average stomata sizes than diploid and triploidC. suksdorfii. We also obtained climatological and elevation data for the sites at which these sampleswere collected, and examined the associations between taxon, ploidy level, stomatal size and density,elevation, and environmental parameters. Our analyses indicate positive associations betweenstomatal size and latitude, and between ploidy level and elevation. Negative associations were foundbetween temperature and precipitation variables and both ploidy level and stomatal size, particularlyfor the fall and winter quarters. There appeared to be no significant association between stomataldensity and any of the environmental variables. Tetraploid C. douglasii occupied a wider range ofenvironmental conditions than did either the diploids or the autotriploids.
Key Words: Agamospermy, climate, Crataegus douglasii, Crataegus gaylussacia, Crataegus suksdorfii,geographical parthenogenesis, polyploidy, stomata.
Determining the causal factors behind speciesdistributions is a fundamental area of study inecology (Krebs 2001). The realized range of anindividual species can be affected by abioticfactors such as climate, biotic interactions suchas competition, or a more complex interplay ofenvironmental factors (Angert and Schemske2005). Species ranges vary dramatically in area,and even closely related species can have verydivergent range sizes (Brown et al. 1996). Forsome close relatives, differences in cytotype maybe the underlying cause of range size differences;polyploidy in one species could lead to an abilityto tolerate a greater or different range ofconditions, and therefore facilitate a wider ornovel distribution, compared to a diploid conge-ner (Levin 1983; Krebs 2001; Parisod et al. 2010;Ramsey 2011). A polyploidization event maypresent opportunities in terms of range expansion,including a possible increase in genetic diversity,and a decrease in barriers to self-fertilization
(Angert and Schemske 2005, Lowry and Lester2006). In addition, polyploids have been found tohave a number of advantages over their diploidrelatives, including larger seed sizes, more vigor-ous seedlings, increased resistance to pathogensand pests, and higher tolerances for poor soils anddrought conditions (Levin 1983; Brown et al.1996). These differences can translate into diver-gent ecological tolerances and geographic distri-butions (Lewis 1980; Lowry and Lester 2006;Ramsey 2011; McIntyre 2012). Although differ-ences between cytotypes in range sizes andenvironmental factors are not universal (Martinand Husband 2009), some rapid divergence afterpolyploidization is theoretically necessary for thenew polyploid population to avoid direct compe-tition with its progenitor, thereby allowing for itspersistence (Coyne and Orr 2004; McIntyre 2012).However, the connections between polyploidyand observed ecological differentiation remainpoorly understood (Li et al. 1996; Ramsey 2011).
MADRONO, Vol. 61, No. 2, pp. 177–193, 2014

Polyploidy is frequently associated with gameto-phytic apomixis, notably in Ranunculus L. species(Horandl 2008) and in many Asteraceae, Poaceae,and Rosaceae (Nogler 1984). In these cases thewider range of apomicts may be related not onlyto polyploidy but also to the ability to set seed inthe absence of other conspecific individuals(Horandl 2006; Lo et al. 2013).
Investigations of the distribution and fre-quency of polyploidy typically depend on theavailability of tissue from which chromosomecounts can be made or, increasingly, fromwhich stainable, intact nuclei can be extractedand passed through a flow cytometer. Neitherof these approaches offers much chance ofsuccess with existing specimens from museumcollections. Taxonomic recognition of majormorphological differences between ploidy levelsmay be sufficient in some cases, as in thecontrast between diploid hawthorns, all ofwhich have 20 stamens per flower, while almostall tetraploid hawthorns have 10 stamens perflower (Talent and Dickinson 2005). Unfortu-nately, this pattern is confounded by intraspe-cific variation in ploidy level (e.g., Crataegussuksdorfii [Sarg.] Kruschke [Dickinson et al.2008]), and morphological variation amongclosely related tetraploids (e.g., C. crus-galli L.sensu lato [Dickinson and Phipps 1986]). Inaddition, flowering traits may not always bevisible on herbarium specimens.
Considering these issues, researchers may seekproxies for direct measurements of chromosomenumber or nuclear DNA content. Cell size is onesuch proxy, since polyploids typically have largercells than diploids (Stebbins 1950; Levin 1983;Otto 2007; Beaulieu et al. 2008). In plants, thesize of pollen grains and of stomatal guard cellpairs may both be accessible with herbariummaterial and work on a variety of taxa, includingCrataegus (Marshall 1978) and other Rosaceae(Joly and Bruneau 2007), has shown how thesetraits can be used to predict ploidy level inherbarium specimens (Buechler 2000; Joachimiakand Grabowska-Joachimiak 2000; Saltonstall etal. 2007; Chen et al. 2009). Relatively simplemethods for light microscopy may have thepotential to reveal the ploidy level of long deadindividuals from which herbarium specimens ofhistorical interest were collected. However, inhawthorns the variation in pollen grain sizewithin ploidy levels is so great that, while overallsize increases with nuclear DNA content mea-sured in leaves, no prediction from pollenmeasurements to ploidy level is possible; onlystainability differentiates the pollen of diploids(highly stainable) from that of polyploids (Dick-inson, unpublished data).
Importantly, features such as stomatal size anddensity have been recognized as varying not onlywith ploidy level, but also with environmental
factors, such as temperature and water availabil-ity (Wang and Clarke 1993). The links betweencytotype, stomata, and environmental variableshave important implications for the realizedranges of plant species, and could help explainthe differences in the breadth and characteristicsof habitats occupied by congeners. Here, weinvestigate the way in which ploidy level,stomatal dimensions, and stomatal density varywith each other, and in relation to geographicdistribution and the environment, as indexed bylatitude, elevation, and climate parameters. Weask the following questions: (1) does size or someother readily measurable characteristic of cellswith a fixed ontogenetic trajectory like stomatavary with ploidy level in such a way that thisparameter can be used to predict ploidy level?, (2)does size or some other characteristic of stomatavary between taxa even when ploidy level is keptconstant?, and finally (3) is cell size or othermorphological variation between ploidy levelsand taxa associated in any way with differences ingeographic distribution?
MATERIALS AND METHODS
Crataegus L. is a genus of approximately 200species of woody plants, part of a mostlyfleshy-fruited clade (tribe Maleae Small) withinan expanded subfamily Amygdaloideae Arn.(Rosaceae) (Potter et al. 2007; McNeill et al.2012). Crataegus is taxonomically complex be-cause, while the subgeneric classification of thegenus into taxonomic sections and series (Phippset al. 1990; Phipps and O’Kennon 2002) isbroadly supported by morphology, not allsubgeneric groups have been shown to bemonophyletic (Lo et al. 2007, 2009). Moreover,species circumscriptions often inadequately re-flect the occurrence of hybridization, gameto-phytic apomixis, and polyploidy. Black-fruitedCrataegus series Douglasianae (Loud.) Rehder isone example of an apparently monophyleticgroup (Lo et al. 2007, 2009). This group iswidespread in the Pacific Northwest of theUnited States and Canada and includes twowell-known species, Crataegus suksdorfii (Sarg.)Kruschke and C. douglasii Lindl. Crataegussuksdorfii is restricted to the mesic conifer forestregion of the Pacific Northwest (Brunsfeld andJohnson 1990), while C. douglasii can grow inmore xeric areas and so has a much widerdistribution, extending from the eastern slopes ofthe Cascades to the Rocky Mountains, withdisjunct populations east of the continentaldivide (Hargrove and Luxmoore 1998; Dickinsonet al. 2008).
Diploid, triploid, tetraploid, and pentaploid C.suksdorfii are now known (Dickinson et al. 2008;Lo et al. 2010a, 2013; Coughlan 2012), althoughonly one of these cytotypes has, as yet, been given
178 MADRONO [Vol. 61

taxonomic recognition. Crataegus gaylussacia A.Heller was described from Sonoma County,California, and has been shown to be triploid(Coughlan 2012, Dickinson et al. unpublisheddata), and to correspond more or less to themorphology of autotriploid individuals foundelsewhere that would otherwise be referred to C.suksdorfii (Lo et al. 2010a). Crataegus gaylussaciahas priority over the name C. suksdorfii if thesenames are applied to all 20-stamen, black-fruitedhawthorns of the Pacific Northwest without regardto ploidy level (Phipps 2012; VASCAN 2013). Inview of the variation in ploidy level documentedfor these plants (Talent and Dickinson 2005; Lo etal. 2013), we will retain the older usage of thesenames here (i.e., as referring to different biologicalentities). A publication clarifying the use of thesenames is currently in preparation (T. A. Dickinson,unpublished manuscript).
In contrast to C. suksdorfii, individuals of C.douglasii are almost exclusively tetraploid (Talentand Dickinson 2005; Lo et al. 2013), anddistinctive morphotypes that have received taxo-nomic recognition (Phipps and O’Kennon 1998,2002) appear to be of hybrid origin (Zarrei et al.2012). Introgression from C. douglasii appears tobe responsible for the allopolyploid cytotypesof C. suksdorfii (Lo et al. 2010a). The widergeographic distribution of tetraploid C. douglasiiis associated with an apparently almost completereliance on asexual seed production (Talent andDickinson 2007; Coughlan 2012; Lo et al. 2013),as is also the case in polyploid C. suksdorfii(Dickinson et al. 1996; Lo et al. 2013). Diploid C.suksdorfii, on the other hand, appears to produceexclusively biparental, sexual seeds (Coughlan2012; Lo et al. 2013).
Sampling
Sampling for this study draws on a largecollection of mainly Pacific Northwest andOntario specimens of Crataegus series Douglasia-nae, assembled and georeferenced for studies ofmorphological, breeding system, and ploidy levelvariation, that are housed for the most part in theGreen Plant Herbarium (TRT) of the RoyalOntario Museum (Table 1). Many, but not all ofthese, are vouchers for studies of DNA sequencevariation in the group (Lo et al. 2007, 2009;Coughlan 2012; Zarrei et al. 2012). To more fullysample the continental United States range of C.douglasii, permission was obtained to remove leafmaterial from two borrowed specimens fromMichigan (Table 1). Specimens were selected thathad sufficient mature leaves to permit removal ofsome for destructive sampling. In the geographicstudy, specimen selection had the further cri-terion that only C. douglasii and C. suksdorfii(both diploids and autotriploids) would becompared, unconfounded by being combined as
in allopolyploid C. suksdorfii (Lo et al. 2009,2010a). Locations of the individuals and popula-tions studied were mapped using SimpleMappr(Shorthouse 2010).
Ploidy level determination. Most specimensstudied are vouchers for flow cytometric ploidylevel determinations (Talent and Dickinson2005). However, because C. douglasii sensu latohas been found to be almost exclusively tetra-ploid (Talent and Dickinson 2005, Talent andDickinson unpublished data), we have includedsome specimens of this taxon for which flowcytometric or chromosome count data areunavailable, and for our purposes have assumedthat they are tetraploid (Table 1).
Stomatal Size – Population Level Comparison
Specimens for this study were selected tocompare population samples of diploid (twopopulations, n 5 7 and n 5 10, Table 1),autotriploid, allotriploid, and allotetraploid (onepopulation each, all n 5 10) C. suksdorfii with twosamples (each n 5 10) of tetraploid C. douglasii,each of the latter sympatric with one of theallopolyploid C. suksdorfii populations Table 1(Dickinson et al. 1996; Lo et al. 2009, 2010a).
Two leaves, randomly chosen from both shortand long shoots were removed from eachspecimen, and two approximately 1 cm 3 1 cmsegments were excised from each leaf. For eachleaf a random number from one to six wasobtained and used to select a sampling position onthe leaf (Fig. 1). Leaf segments were softened for30–40 min in 70% ethanol, rinsed in tap water for15–20 min, and then submerged overnight in100% DranoH (S. C. Johnson & Son, Inc., Racine,WI) to clear the leaf segments (Buechler 2000).The following day, the samples were rinsed inwater for 15–20 min, mounted in water on amicroscope slide, and examined under Nomarskidifferential interference contrast at 8003 magni-fication. Using MorphoSys v. 1.29 (Meacham andDuncan 1991), stomatal outlines were captured inorder to calculate their length, width, area, andperimeter. In each leaf segment, five stomateswere measured in five different quadrants of theleaf fragment: upper left, upper right, bottom left,bottom right, and middle, giving a total of 25stomates measured per segment. To avoid count-ing a stomate more than once, the same countingsequence was used consistently (i.e., after the firststomate, go up, skip one, measure the followingone; next, go left, skip one, count the followingone; then up again and so on).
Stomatal Size and Density – Geographic Study
Specimens for this study were selected tocompare diploid and autotriploid C. suksdorfii
2014] MCGOEY ET AL.: PLOIDY LEVEL AND STOMATA SIZE IN CRATAEGUS 179

TA
BL
E1.
SIT
EA
ND
VO
UC
HE
RD
AT
AF
OR
PO
PU
LA
TIO
NA
ND
GE
OG
RA
PH
ICS
TU
DIE
SO
FC
EL
LS
IZE
VA
RIA
TIO
NIN
RE
LA
TIO
NT
OP
LO
IDY
LE
VE
LIN
CR
AT
AE
GU
SS
ER
IES
DO
UG
LA
SIA
NA
E(A
LL
2004-
CO
LL
EC
TIO
NS
BY
TA
D,
E.
Y.
Y.
LO
,A
ND
S.
NG
UY
EN
;D
1619
AT
MT
02
CO
LL
EC
TE
D2001
BY
TA
D,
F.
GE
RV
AIS
,A
ND
J.S
.D
ICK
INS
ON
).S
ites
are
nu
mb
ered
as
inF
ig.
7,
an
dla
bel
edas
inp
revio
us
stu
die
s.P
loid
yle
vel
det
erm
inati
on
sb
ase
do
nfl
ow
cyto
met
ry(T
ale
nt
an
dD
ick
inso
n2005;
Lo
etal.
2013).
Exce
pt
as
no
ted
,all
vo
uch
ers
are
fro
mth
eco
nti
nen
tal
US
A,
an
dare
ho
use
dat
TR
T.
So
me
plo
idy
level
s,w
her
ek
no
wn
,w
ere
det
erm
ined
by
N.
Tale
nt
fro
mem
bry
o(*
),
rath
erth
an
leaf,
tiss
ue.
Co
llec
tor
isT
AD
un
less
ind
icate
do
ther
wis
e(T
AD
5T
.A
.D
ick
inso
n;
RE
D5
R.
E.
Do
tter
er;
EL
5E
.Y
.Y
.L
o;
RM
L5
R.
M.
Lo
ve)
.
Sta
teT
axo
nC
ou
nty
Sit
eL
at.
N,
Lo
ng.
WE
lev.
(m)
Sit
en
ote
sV
ou
cher
s(p
op
ula
tio
nst
ud
y)
Vo
uch
ers
(geo
gra
ph
icst
ud
y)
Plo
idy
Califo
rnia
C.
dougla
sii
Lin
dl.
sensu
lato
Plu
mas
(1)
CA
R1,
Cre
scen
tM
ills
40.0
94,
120.9
11
1068
wit
hin
120
m,
an
d11
mab
ove,
Ind
ian
Cr.
RE
D-C
AR
036
4x*
Sh
ast
a(2
)C
AR
2,
Dan
a41.1
14,
121.5
67
1017
wit
hin
100–110
mo
f,an
dat
ab
ou
tth
esa
me
level
as,
Dan
aC
r.
2006-1
24x
C.
suk
sdorf
ii(S
arg
.)K
rusc
hk
e
Mari
n(3
)C
A5,
1k
mW
of
To
male
s38.2
47,
122.9
13
87
on
ban
ks
of
seaso
nal
wate
rco
urs
es
2010-5
3x*
Sis
kiy
ou
(4)
CA
R5,
Sco
ttV
all
ey41.3
97,
122.8
38
872
Sco
ttR
.fl
oo
dp
lain
2006-1
52x
2006-2
22x
So
no
ma
(5)
CA
7,
Seb
ast
op
ol
38.3
6,
122.8
180
Cu
nn
ingh
am
Mars
hR
.T
hom
pso
n564
3x
R.
Thom
pso
n565
3x
R.
Thom
pso
n566
3x
Idah
oC
.dougla
sii
Lin
dl.
sensu
lato
Ad
am
s(6
)ID
06,
Last
Ch
an
ceC
am
pgro
un
d
44.9
89,
116.1
91425
wit
hin
10
mo
fG
oo
seC
r.2005-0
5,
2005-0
6,
2005-0
7,
2005-0
8,
2005-1
1,
EL
-166,
EL
-179,
EL
-180,
EL
-181,
EL
-183
4x
(7)
ID06a,
Last
Ch
an
ceR
oad
an
dG
oo
seC
reek
44.9
64,
116.2
06
1293
wit
hin
10
mo
fG
oo
seC
r.2001-8
24x*
Idah
o(8
)W
hit
eB
ird
45.7
58,
116.3
06
476
road
sid
e,ab
ove
Wh
iteb
ird
Cr.
RM
L-9
504
4x*
Lem
hi
(9)
ID16,
Salm
on
Riv
er45.3
67,
113.9
52
1130
wit
hin
70
m,
an
d7
mab
ove,
Salm
on
R.
EL
-190
4x
Nez
Per
ce(1
0)
ID20,
Hig
hw
ay
346.5
22,
116.7
33
291
wit
hin
300
m,
an
d5-
10
mab
ove,
Lit
tle
Po
tlatc
hC
r.
EL
-121
4x
C.
suk
sdorf
ii(S
arg
.)K
rusc
hk
e
Ad
am
s(1
1)
ID06,
Last
Ch
an
ceC
am
pgro
un
d
44.9
989,
116.1
91425
wit
hin
10
mo
fG
oo
seC
r.E
L-1
62,
EL
-163,
EL
-164,
EL
-167,
EL
-168,
EL
-172,
EL
-173,
EL
-179,
EL
-182,
EL
-184
3x
180 MADRONO [Vol. 61
TA
BL
E1.
CO
NT
INU
ED
.
Sta
teT
axo
nC
ou
nty
Sit
eL
at.
N,
Lo
ng.
WE
lev.
(m)
Sit
en
ote
sV
ou
cher
s(p
op
ula
tio
nst
ud
y)
Vo
uch
ers
(geo
gra
ph
icst
ud
y)
Plo
idy
Mic
hig
an
C.
dougla
sii
Lin
dl.
sensu
lato
Ho
ugh
ton
(12)
47.0
33,
88.6
17
200
Ric
hard
s2575
(CM
)-
Kew
een
aw
(13)
47.3
7,
88.3
03
330
Ric
hard
ss.
n.
(DA
O591508)
-
Mo
nta
na
C.
dougla
sii
Lin
dl.
sensu
lato
Lak
e(1
4)
MT
06,
Bea
ver
hea
dD
rive
47.4
86,
114.1
13
923
wit
hin
50
m,
an
dle
ssth
an
1m
ab
ove,
Cro
wC
r.
2001-3
14x*
Po
wel
l(1
5)
MT
02,
Kle
insc
hm
idt
Fla
t
47.0
2,
112.9
46
1319
Co
op
erL
ak
eR
d.,
wit
hin
50
m,
an
dle
ssth
an
1m
ab
ove,
Ro
ckC
r.
D1611,
EL
-24,
EL
-31,
EL
-32,
EL
-34,
EL
-35,
EL
-38,
EL
-39,
EL
-40,
EL
-46
4x
EL
-40
4x
C.
suk
sdorf
ii(S
arg
.)K
rusc
hk
e
Po
wel
l(1
5)
MT
02,
Kle
insc
hm
idt
Fla
t
47.0
2,
112.9
46
1319
Co
op
erL
ak
eR
d.,
wit
hin
50
m,
an
dle
ssth
an
1m
ab
ove,
Ro
ckC
r.
D1619,
EL
-29,
EL
-30,
EL
-33,
EL
-36,
EL
-37,
EL
-41,
EL
-43,
EL
-44,
EL
-45
4x
Ore
go
nC
.dougla
sii
Lin
dl.
sensu
lato
Gra
nt
(16)
So
uth
fork
Rd
.44.3
72,
119.5
47
836
wit
hin
50
m,
and
5m
above,
the
South
Fork
,Jo
hn
Day
R.
RM
LC
2003-2
04x
Mu
ltn
om
ah
(17)
Fro
nta
ge
road
So
fI-
84
nea
rD
od
son
45.6
04,
122.0
429
wit
hin
300m
,an
d20
mab
ove,
the
Co
lum
bia
R.
RM
L-8
521
-
Um
ati
lla
(18)
Blu
eM
ou
nta
ins,
Ssi
de
of
OR
Hw
y204
45.7
95,
118.3
59
896
road
sid
ew
ith
in100
m,
an
d17
mab
ove,
trib
uta
ryo
fP
ine
Cr.
2008-8
14x
C.
suk
sdorf
ii(S
arg
.)K
rusc
hk
e
Co
lum
bia
(19)
OR
11,
Sau
vie
Isla
nd
45.7
34,
122.7
67
4b
each
rid
ge
wit
hin
ap
pro
x.
30
mo
fth
eC
olu
mb
iaR
.
2005-1
6,
EL
-103,
EL
-104,
EL
-105,
EL
-106,
EL
-107,
EL
-109
2x
(19)
bea
chri
dge
wit
hin
ap
pro
x.
30
mo
fth
eC
olu
mb
iaR
.
P.
Zik
a18477
2x
Do
ugla
s(2
0)
OR
04,
Up
per
Elk
Mea
do
ws
Res
earc
hN
atu
ral
Are
a
43.5
27,
122.9
05
1240
op
ense
ep(C
urt
is1986)
RM
LC
2003-3
53x
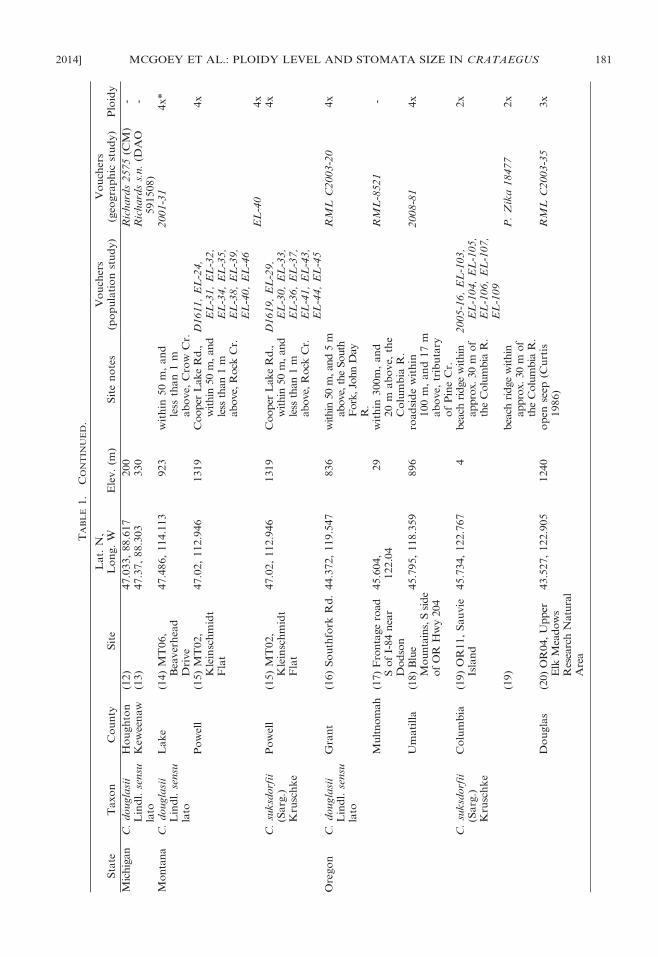
2014] MCGOEY ET AL.: PLOIDY LEVEL AND STOMATA SIZE IN CRATAEGUS 181

TA
BL
E1.
CO
NT
INU
ED
.
Sta
teT
axo
nC
ou
nty
Sit
eL
at.
N,
Lo
ng.
WE
lev.
(m)
Sit
en
ote
sV
ou
cher
s(p
op
ula
tio
nst
ud
y)
Vo
uch
ers
(geo
gra
ph
icst
ud
y)
Plo
idy
Jose
ph
ine
(21)
OR
20,
Dee
rC
reek
Cen
ter
42.2
77,
123.6
47
387
ban
ks
of
Dee
rC
r.2010-1
12x
2010-1
22x
Lan
e(2
2)
OR
06,
Patt
erso
nM
ou
nta
inP
rair
ie
43.7
67,
122.6
19
1307
op
ense
epE
L-4
8,
EL
-49,
EL
-50,
EL
-51,
EL
-52,
EL
-53,
EL
-54,
EL
-56,
EL
-57,
EL
-63
3x
(22)
EL
53
3x
(22)
EL
56
3x
Lin
n(2
3)
OR
01,
Co
gsw
ell-
Fo
ster
Res
erve
44.3
34,
123.1
22
88
ver
nally
mo
ist
Willa
met
teV
alley
pra
irie
,w
ith
in500
man
d2
mab
ove,
Lit
tle
Mu
dd
yC
r.(L
ove
an
dF
eigen
1978)
EL
-68,
EL
-72,
EL
-75
2x
(23)
RM
L-8
712
2x*
Wash
ingto
n(2
4)
Hil
lsb
oro
45.5
57,
122.8
88
72
Ed
ge
of
seco
nd
-gro
wth
wo
od
s,p
ow
erlin
ecl
eari
ng
P.
Zik
a18483
2x
Was
hin
gton
C.
dougla
sii
Lin
dl.
sensu
lato
Kit
tita
s(2
5)
WA
20,
Ro
nald
47.2
4,
121.0
32
725
road
sid
eh
edger
ow
,w
ith
in100
m,
an
d5–10
mab
ove,
trib
uta
ryo
fC
leE
lum
R.
2005-2
14x*
Th
urs
ton
(26)
WA
21,
Mo
un
dP
rair
ie,
Esi
de
of
Sca
tter
Cre
ekR
est
Are
ao
nIn
ters
tate
5
46.8
37,
122.9
84
65
dry
dis
turb
edm
ead
ow
,w
ith
in900
m,
an
d6
mab
ove,
Sca
tter
Cr.
P.
Zik
a18438
4x
C.
suk
sdorf
ii(S
arg
.)K
rusc
hk
e
Cla
rk(2
7)
Rid
gef
ield
45.8
55,
122.7
18
73
thic
ket
sat
edge
of
mea
do
w,
wit
hin
500
m,
an
d70
mab
ove,
Mu
dL
ak
e
P.
Zik
a18486
(sam
ein
div
idu
al
as
P.
Zik
a18918)
2x
P.
Zik
a18918
(sam
ein
div
idu
al
as
P.
Zik
a18486)
2x*
Lew
is(2
8)
Dre
ws
Pra
irie
46.4
67,
122.8
84
90
wet
pra
irie
rem
nan
tP
.Z
ika
18995
2x
182 MADRONO [Vol. 61

with tetraploid C. douglasii across their continen-tal United States ranges (Fig. 2, Dickinson et al.2008). Leaf samples were examined from 15 C.suksdorfii, including eight diploids and sevenautotriploids, and 17 tetraploid C. douglasii trees(Table 1).
The methods were the same as for thepopulation-level study above, with the followingexceptions. Softening the leaves was deemedunnecessary, and leaves were submerged inDranoH for 72 hr. Since the results of theprevious study demonstrated that the samplinglocation on the leaf (Fig. 1) affected stomata sizesignificantly (see Results), we restricted this studyto leaf sections in the middle right of the leaf(section four in Fig. 1). Within each 1 cm2 leafsection we examined an areole in each of fourdifferent, spatially separated regions (areoles areareas bounded by quarternary veins on all sides).We captured images of an areole at different focaldepths using an Infinity 1 digital camera and theInfinity Analyze program supplied with it (Lu-menera Corporation, Ottawa, ON). The imageswere stacked using Combine Z image software(Hadley 2010), and the stacked images were thenimported back to the Infinity Analyze program.This was used to delimit individual areoles,determine their area, and count all the stomatafound within an areole. We also used theprogram to measure the length and width ofeach stomata (Fig. 3). All measurements wereautomatically stored in an M.S. AccessTM data-base (Microsoft, Redmond, WA).
Environmental data. Latitude and longitude foreach collection site (Table 1) were used to obtainclimate data (1971–2000 monthly normals forprecipitation, and maximum and minimum tem-perature) using the online PRISM Data Explorer(PRISM Climate Group 2012). The geographiccoordinates were also used to ascertain the ele-vation at each site using Google Earth (Version7.1.2.2041 [software] available from http://www.google.com/earth/). Google Earth was alsoemployed to estimate the proximity of samplingsites to bodies of water. The analyses describedbelow used seasonal (quarterly) averages of themonthly normal values (Q1, winter, December-February; Q2, spring, March–May; Q3, summer,June–August; and Q4, fall, September–Novem-ber). Some analyses also included the averageminimum and maximum temperatures for theyear, and total normal precipitation. Analyses ofjust the quarterly averages also included siteelevation and latitude.
Statistical analyses. Statistical analyses wereperformed using R (R Development Core Team2004). Normality of the size measurements waschecked using qq plots and the Shapiro-Wilkstest. Measurements found to be lognormallydistributed were log-transformed. When analyz-
TA
BL
E1.
CO
NT
INU
ED
.
Sta
teT
axo
nC
ou
nty
Sit
eL
at.
N,
Lo
ng.
WE
lev.
(m)
Sit
en
ote
sV
ou
cher
s(p
op
ula
tio
nst
ud
y)
Vo
uch
ers
(geo
gra
ph
icst
ud
y)
Plo
idy
Wyo
min
gC
.dougla
sii
Lin
dl.
sensu
lato
Tet
on
(29)
Targ
hee
Nati
on
al
Fo
rest
43.2
77,
110.7
95
1782
Sn
ak
eR
iver
flo
od
pla
inP
hip
ps
7459
-
On
tari
o,
Can
ad
aC
.dougla
sii
Lin
dl.
sensu
lato
Th
un
der
Bay
Dis
tric
t
(30)
ON
19,
Pin
eR
iver
,fi
rst
rap
ids
48.0
54,
89.5
07
211
Sh
rub
by
flo
od
pla
inD
1294
4x*
2014] MCGOEY ET AL.: PLOIDY LEVEL AND STOMATA SIZE IN CRATAEGUS 183

ing the stomata size data, two different methodswere used to avoid the pseudoreplication problemof having many stomata measurements from thesame leaves and trees. First, the mean values foreach tree were calculated, and then compared. Inthe second analysis, we used a linear mixed effectsmodel to determine the effect of ploidy andtaxonomic group while accounting for thereplication within leaves and trees. To analyzethe relationship between stomatal traits, ploidylevel, and climate variables we conducted aMANOVA and principal component analysis.We used the fourth-corner function in R from theade4 package (Legendre et al. 1997, Dray andLegendre 2008) in order to infer relationshipsbetween our morphological data (stomata sizeand density; nuclear DNA content) and theenvironmental features of the sites referred toabove.
RESULTS
While examining our C. douglasii and C.suksdorfii leaves, we made basic observations onthe leaf surface characteristics (Fig. 3). Theoutlines of epidermal cells were wavy and cellssurrounding the stomata appeared anomocytic.This is consistent with Metcalfe and Chalke’s(1979) description of Rosaceae epidermis, but notwith observations of other Crataegus speciesstudied by Ganeva et al. (2009).
FIG. 1. Diagram of a Crataegus leaf divided into sixsections. For the population-level study, the sampledsection was chosen randomly while, for the geographicstudy, the mid-right section (4) was always used. In allcases the abaxial surface of the leaf was examined asshown here.
FIG. 2. Map showing sites where tetraploid C. douglasii (closed squares), diploid C. suksdorfii (open circles), andtriploid C. gaylussacia and C. suksdorfii (open triangles) were collected for the geographic study (Table 1). Sitesused in the population study are circled.
184 MADRONO [Vol. 61

Stomatal Size – Population-Level Study
Variation in stomatal size. The stomata rangedin length from 21 mm to 54 mm and their totalareas ranged from 264 mm2 to 1486 mm2. Asexpected, stomata from tetraploid individualswere the largest (mean length 36 mm, comparedto the overall mean of 34 mm). These results areconsistent with analyses on the other sizemeasures (width, perimeter, and area). Diploids
had slightly larger average sizes than triploids,which were the smallest of the three ploidygroups. This is surprising, since triploids didhave higher DNA content then diploids, which isexpected to be correlated with cell size. We testedfor such a relationship between DNA contentand stomata size within each ploidy level, butthere were no significant correlations (all P .0.05).
The mean values for each leaf were determinedand found to be normally distributed. Averagestomata characteristics were consistently differentbetween ploidy levels (all P values ,0.001, seeFig. 4 for length), but not in a proportionalmanner (see below). Using an ANOVA to assesswhich factors affected stomatal characteristics,we found that both ploidy level and section of theleaf (Fig. 1) were significant (ploidy level andstomata length, P , 0.001; ploidy level and area,P , 0.001; leaf section and stomata length P ,0.001; leaf section and stomata area P , 0.001).
The mixed effects model of all the individualstomatal measurements was consistent with theanalysis using size means. When examining thelength of stomata, we found that ploidy was asignificant factor (P , 0.001), and that the locationmeasured on the leaf was as well (P , 0.001). Inaddition there was a significant interaction betweenploidy level and measured location (P 5 0.0071).The same was true when examining stomata area,with both ploidy level (P , 0.001) and leaf section(P , 0.001) being significant factors, the interactionterm was also significant (P 5 0.0124). Withintetraploids (C. douglasii, 4x C. suksdorfii; Fig. 4;Table 1), species was not a significant factor forstomata size (all P values .0.05).
Stomatal Size – Geographic Study
Variation in stomatal size. Stomata ranged inlength from 15 mm to 57 mm, and the widthsranged from 9.8 mm to 38 mm. As was true in thepopulation-level study, tetraploid C. douglasiihad the largest stomata, and autotriploid C.suksdorfii had the smallest, although they werenot significantly different from their diploidconspecifics (Fig. 5).
Again, two analyses were conducted to avoidpseudoreplication. Tree means were used toexamine the effect of ploidy on stomata lengthand width. Ploidy level was a significant factor forboth length and width. The second analysis, usingthe full data set while accounting for pseudorep-lication within leaves and trees by including themas random factors, confirmed these findings, asthe linear mixed effect models showed that ploidywas a significant factor for both stomata length (P, 0.001) and width (P , 0.001).
Variation in stomatal density. Stomatal densityranged from 88 stomates/mm2 to 323 stomates/
FIG. 3. Panels show Crataegus douglasii stomatalmeasurements: (A) Stomata width (i) and length (ii)were measured as the distances between the midpoints;(B) Crataegus douglasii stomata shown under 803magnification. Stomata density was estimated bycounting the number of stomata within an areole, anddividing by the total area of the areole.
2014] MCGOEY ET AL.: PLOIDY LEVEL AND STOMATA SIZE IN CRATAEGUS 185

mm2 (Fig. 5A). Note that the densities arereported here in stomates/mm2 for ease ofinterpretation, but the areoles were 0.16 mm2 onaverage. The C. douglasii had an average stomatadensity of 166, while the diploid and triploid C.suksdorfii had average densities of 169 and 214respectively. When all data were included in asimple ANOVA model, there was a significantdifference between C. douglasii and C. suksdorfii(P 5 0.0215), but this difference was driven bythe higher density in triploids, so that when treeand leaf were included as random effects, thespecies difference was no longer significant (P 50.21). Likewise, an ANOVA on tree meansshowed that there was not a significant differencein stomata density between ploidy levels (P 50.16) or species (P 5 0.24).
Environmental correlates. Comparisons be-tween sites occupied by tetraploid C. douglasiiand those occupied by C. suksdorfii (bothdiploids and autotriploids) showed the formerto be cooler and more xeric (Figs. 6, 7). It isimportant to note that the environmental datawere atmospheric, and may not be a precisereflection of water relations on the ground at
each site. Nevertheless, except for the twoMichigan sites for which hydrological conditionsare unknown, each of the samples studied hereare on flood plains, adjacent lakeshores, or onslopes down which water moves at or below thesurface in response to local atmospheric condi-tions (Table 1). The results of the MANOVAanalysis demonstrated a significant relationshipbetween three climatological variables (annualprecipitation, minimum and maximum tempera-ture) and the Crataegus species at a site (P 5
0.0058). Fourth-corner analyses demonstrated asignificant relationship between several environ-mental factors and some but not all of thefeatures of the hawthorn stomata (Fig. 8).Stomatal length and width covaried significantlywith latitude, precipitation, and temperature,although the direction of the relationship de-pended on the quarter of the year examined(Fig. 8). Ploidy level was positively associatedwith elevation and negatively associated withprecipitation and temperature (Fig. 8). Therewere no significant relationships between stoma-tal density and any of the environmental factors(Fig. 8).
FIG. 4. Box plot of size (length) of stomata for Crataegus taxa in the population study. The dark middle linesshow the median values, and two ends of the box show the 1st and 3rd quartiles. The whiskers show theinterquartile range multiplied by 1.5. All values outside this range (outliers) are shown as open circles. Both ploidylevel and section of the leaf (Fig. 1) were significant (ploidy level P , 0.0001; leaf section P , 0.0001).
186 MADRONO [Vol. 61

Predicting Ploidy Level fromStomatal Measurements
Data from the population-level study and thegeographical one were pooled in order to depictthe predictive relationship between nuclearDNA content and stomatal size. Plottingnuclear DNA content against stomatal length(Fig. 9) demonstrates the way in which varia-tion at the levels of leaf, individual, andsampling site vitiates any attempt to predictploidy level from stomatal size: the ranges insize for each ploidy level overlap so much thatany given size except the very smallest or the
largest might correspond any one of the threeploidy levels studied here (Fig. 9). Similarrelationships were observed by Marshall (1978)in Manitoba Crataegus chrysocarpa and C.succulenta. Although Marshall was probablyincorrect in his inference that the smaller stomataof C. succulenta meant that this taxon is diploid(Ontario specimens have been shown to betriploid; Talent and Dickinson 2005), their sizerelative to those of C. chrysocarpa (tetraploid;Talent and Dickinson 2005) demonstrates thesame trend we have observed.
FIG. 5. Boxplot of stomata characters of diploid (2x)and autotriploid (3x) C. suksdorfii and tetraploid (4x)C. douglasii in the geographic study: (A) Density; (B)Length (mm); (C) Width (mm). The dark middle linesshow the median values, and two ends of the box showthe 1st and 3rd quartiles. The whiskers show theinterquartile range multiplied by 1.5. All values outsidethis range (outliers) are shown as dark circles.
FIG. 6. Boxplot of climate summaries comparing theC. douglasii and C. suksdorfii sites (diploid andautotriploids pooled) used in the geographic study.Precipitation is the annual total (mm), maximum andminimum temperatures are annual averages. The darkmiddle lines show the median values, and two ends ofthe box show the 1st and 3rd quartiles. The whiskersshow the interquartile range multiplied by 1.5. Allvalues outside this range (outliers) are shown asdark circles.
2014] MCGOEY ET AL.: PLOIDY LEVEL AND STOMATA SIZE IN CRATAEGUS 187

DISCUSSION
The taxonomic complexity of the genus Cra-taegus has long been recognized (Camp 1942), ashas the connection between this complexity andthe occurrence of gametophytic apomixis, hy-bridization, and polyploidy (Dickinson andPhipps 1985, 1986; Lo et al. 2010b). Thisconnection makes knowing the ploidy level andbreeding system critical in order to make cleartaxonomic judgments regarding Crataegus spe-cies. Determining ploidy level from chromosomecounts is difficult because the tissues in whichmeiotic metaphases can be accumulated areavailable for less than a week, annually, at anygiven locality. Flow cytometry is an extremelyvaluable tool when studying Crataegus, but thismethod works mainly on living tissue, making itnearly useless especially with older herbariummaterial (e.g., type specimens). The only excep-tion to this is the ability to obtain flow cytometricdata from seeds (Table 1; Talent and Dickinson2007). This approach has been shown to work ondecades-old seeds, provided they have beenstored in the cold (Talent personal communica-tion). The ability to use a phenotypic trait likestomata size as an indicator of ploidy level wouldgreatly simplify the study of this complicatedgenus. Unfortunately, based on our results,variation in this trait is not consistently relatedto ploidy level.
Although the largest stomata in our study werefrom the tetraploid C. douglasii trees, stomatafrom autotriploid C. suksdorfii individuals weregenerally smaller than those of diploids (Fig. 5).In addition, there was wide variation in sizewithin a ploidy level, and too much overlapbetween ploidy levels to make a comparison ofaverage stomata size useful for assessing ploidy(Fig. 9). The relationship between genome andcell size is weaker in trees than in other plants(Beaulieu et al. 2008). The lack of a differencebetween C. suksdorfii cytotypes in stomata sizemay be a reflection of this. Stomata size is notsolely controlled by genome size; environmentalor physiological factors may select for smaller cellsize, or specifically for smaller stomata. Note thatwe deliberately excluded allopolyploid C. suks-dorfii from the geographic comparison in order tofocus on the contrast between diploid andautotriploid C. suksdorfii and tetraploid C.douglasii (Fig. 5).
There seemed to be more of a taxonomicgrouping with respect to stomata size than adifference between ploidy levels (Fig. 5). Thedifference in stomata size between C. douglasiiand C. suksdorfii could prove useful in determin-ing the identity of previously collected specimens.This might be of use in the case of sterilespecimens, where per flower stamen numbersare unavailable. Other factors affect stomata size,and the relationship between environmental
FIG. 7. Principal component analysis of quarterly climate data of the Crataegus sites used in the geographic study(numbered as in Table 1). The four quarters are winter (Q1), spring (Q2), summer (Q3), and fall (Q4). Left panel:convex hulls enclose sites grouped by taxon and ploidy level present at each one. Right panel: PCA biplot withvectors representing contributions of quarterly climate variables.
188 MADRONO [Vol. 61

characteristics and stomata traits illustrate suchan example. For C. douglasii and C. suksdorfii,spring and summer precipitation averages arepositively correlated with stomata size (Figs. 7,8). There are a limited number of triploid sites,and there is much environmental heterogeneitybetween those in California (presumptively auto-triploid C. gaylussacia) and those in Oregon(autotriploid C. suksdorfii). However, these sitesare representative of the known, and quitelimited, distribution of C. gaylussacia on theone hand, and of autotriploid C. suksdorfii on theother.
Despite large strides in polyploidy research,we still know very little about the ecologicalimplications of polyploidy (Soltis et al. 2010;Manzaneda et al. 2012). Past researchers havehypothesized that genome duplication confersadvantages that lead to an increased or novelrange for the polyploid species. However, littleempirical work has been done to test this idea(Soltis et al. 2010). Stebbins addressed the
uncertainty surrounding the effect of polyploidyon species ranges decades ago and suggested thatclarification could be gained by carefully studyingthe morphological traits and habitats of increas-ing numbers of species groups (Stebbins 1950). Inthe years since, many different taxonomic groupshave been examined. This research has demon-strated that polyploids often tolerate moreextreme conditions than their diploid relatives(Lewis 1980), and one ecological difference thatdoes seem common across many taxa is thatplants with higher ploidy levels are found in morexeric conditions than their diploid relatives(Levin 2002). In our study group, tetraploid C.douglasii is found in more xeric conditions (interms of mean annual precipitation) than diploidand triploid C. suksdorfii (Fig. 6). This isconsistent with studies on other plants, wheretaxa with higher ploidy levels grow in drierconditions than their lower ploidy-level relatives,including in Achillea L. (Ramsey 2011), Chamer-ion (Raf.) Raf. ex Holub (Maherali et al. 2009),
FIG. 8. Associations between Crataegus stomatal characteristics and ploidy level, and the geographic and climaticvariables at the sites from which they were collected, as calculated by a fourth-corner analysis (Legendre et al.1997). Significant positive associations are shown in black, significant negative associations are in grey and non-significant associations are in white. Latitude values were collated from the herbarium specimens used in thegeographic study. Elevation values were gathered from Google Earth. Monthy averages for maximum temperature,minimum temperature, and precipitation were acquired using the PRISM database and used to calculate quarterlyvalues. The four quarters are winter (Q1), spring (Q2), summer (Q3), and fall (Q4).
2014] MCGOEY ET AL.: PLOIDY LEVEL AND STOMATA SIZE IN CRATAEGUS 189

and Brachypodium P. Beauv. (Manzaneda et al.2012). Sites at which C. douglasii occurs are also,on the average, colder than those occupied by C.suksdorfii (Fig. 6). When the climate data arebroken down into quarterly values, we see thatwhile there is some overlap between the environ-ments in which diploid C. suksdorfii and tetra-ploid C. douglasii occur, the tetraploid occupies amuch wider range of environments than does thediploid (Fig. 7). Combined with the variation inbreeding systems, this pattern of apomicticpolyploids distributed over a wider range ofenvironments than closely related sexual diploids(hence exhibiting a larger geographic range;Fig. 2) is what is referred to as geographicalparthenogenesis (Horandl 2006; Lo et al. 2013).
As seen in our study, a higher ploidy level tendsto lead to larger cells and stomata. In many taxathis increase in stomata size is accompanied by asignificant decrease in stomata density. Never-theless, by using the fourth-corner method toinfer the association between nuclear DNAcontent and features of stomata, and quarterlyclimate parameters and elevation, we have shownthat stomata density does not vary significantly inrelation to any of the environmental factors weexamined (Fig. 8). In other groups it does notseem advantageous to have larger stomata inmore xeric conditions, but in many cases, thedecreased density more than compensates, so thatthe total pore space on a leaf is lower in highercytotypes (Levin 2002). Polyploidy may conferother advantages on plants that enable them to
tolerate dry environments. Polyploids tend togrow more slowly than diploid congenerics,which could be an advantage when resourcesare scarce (Grant 1981; Levin 1983; Deng et al.2011). In addition, some polyploids have in-creased leaf water content and transpirationefficiency (Chen and Tang 1945; Tal and Gardi1976). In Crataegus, allopolyploid C. suksdorfiioccurs in drier areas, where no diploid popula-tions occur. Perhaps this is due to the contribu-tion from the C. douglasii genomes in theseindividuals. It is also important to considereffects separate from cytotype; genetic divergencedriven by natural selection will occur betweenlineages after a polyploidization event. Moreresearch on the magnitude and mechanism forincreased tolerance to dry environments inpolyploids is needed.
As noted above, environmental descriptors like‘‘drier’’ or ‘‘wetter,’’ not to mention the precipi-tation data used in our analyses, relate toatmospheric conditions. In fact all of our sitesexcept possibly Hillsboro, Washington, and thetwo Michigan ones, are within a kilometer(usually much less) of at least seasonally runningor standing water, and the topography of many ofthese sites is such that ground water likely flowsthrough the root zone most or all of the year(Table 1; e.g., Curtis 1986). Hawthorns arefrequently found in riparian zones and this factmay help to explain the lack of correlationbetween stomatal size and growing season pre-cipitation (Fig. 8). Figure 8 also documents theway in which diploid C. suksdorfii and autotri-ploid C. gaylussacia are restricted to a region ofMediterranean climate (high winter precipitation,summer drought) where stomatal size is negative-ly correlated with Q2 temperatures, and positivelycorrelated with Q3 precipitation.
CONCLUSIONS
The interplay between cytotype, physiologyand ecology is complex. Our study demonstratesthat neither stomatal density nor stomatal sizecan be used to determine ploidy level in theCrataegus series Douglasianae, However, stomatasize may be useful in differentiating between C.douglasii and C. suksdorfii. The two species occurin distinctive environments, and some climato-logical (precipitation, temperature) and geo-graphic traits (latitude) are significantly relatedto stomata size. Polyploidy is a unique mecha-nism of speciation in its immediacy, and itspropensity to cause speciation in sympatry. Theecological implications of polyploidization con-tinue to be an important avenue of research.
ACKNOWLEDGMENTS
We are greatly indebted to Eugenia Lo and especiallyNadia Talent for sharing flow cytometry data with us;
FIG. 9. Nuclear DNA content and stomatal lengthplotted to show why stomatal size cannot be used as apredictor of ploidy level given the variation betweenleaves, individuals, and sites using data from thecombined population level (leaf region 4 only) andgeographic samples. Tetraploid C. douglasii, filledsquares. Open symbols, C. suksdorfii: circles, diploid;triangles, triploid.
190 MADRONO [Vol. 61

also to Nadia Talent for advice on nomenclature. Wealso thank Rebecca Dotterer, Rhoda Love, and PeterZika for plant collections and collecting site informa-tion, and Annabel Drover, Jenny Bull, Cheying Ng, andthe many Green Plant Herbarium volunteers and 2011summer staff for helping to organize and database thevouchers for this study. The late Steve Brunsfeldsuggested several of our Idaho study sites to TAD.We are grateful to Doreen Smith, Walter Earle,Margaret Graham, Bill Jensen, Gene Cooley, RockyThompson, John Herrick, Ralph Johnson, and MarciaJohnson, for information about and access to Califor-nia hawthorn collecting sites. The Siskiyou FieldInstitute & Deer Creek Center, Selma, Oregon, kindlypermitted us to collect on their property. Data on pollensize and stainability were collected by undergraduateproject students Yi-Min Chun, Jason Lee, JenniferShiller, and Jurgena Tusha. Adam Dickinson did theinitial GIS comparison of climate variables at C.douglasii and C. suksdorfii collecting sites. Many thanksto the Royal Botanical Gardens Science Department,especially Natalie Iwanycki, for their support of thisproject. Financial support from a Natural Sciences andEngineering Research Council of Canada Undergrad-uate Student Research Award to KC and DiscoveryGrant A3430 to TAD, the Department of Ecology &Evolutionary Biology of the University of Toronto, andthe Royal Ontario Museum is gratefully acknowledged.
LITERATURE CITED
ANGERT, A. L. AND D. W. SCHEMSKE. 2005. Theevolution of species’ distributions: reciprocal trans-plants across the elevation ranges of Mimuluscardinalis and M. lewisii. Evolution 59:1671–1684.
BEAULIEU, J. M., I. J. LEITCH, S. PATEL, A.PENDHARKAR, AND C. A. KNIGHT. 2008. Genomesize is a strong predictor of cell size and stomataldensity in angiosperms. New Phytologist 179:975–986.
BROWN, J. H., G. C. STEVENS, AND D. M. KAUFMAN.1996. The geographic range: size, shape, boundar-ies, and internal structure. Annual Review ofEcology and Systematics 27:597–623.
BRUNSFELD, S. J. AND F. D. JOHNSON. 1990. Cytolog-ical, morphological, ecological and phenologicalsupport for specific status of Crataegus suksdorfii(Rosaceae). Madrono 37:274–282.
BUECHLER, W. K. 2000. Estimating polyploidy levelsusing cell size in Salix leaves. American Journal ofBotany 87:238.
CAMP, W. 1942. The Crataegus problem. Castanea7:51–55.
CHEN, G., W.-B. SUN, AND H. SUN. 2009. Morpho-logical characteristics of leaf epidermis and sizevariation of leaf, flower and fruit in different ploidylevels in Buddleja macrostachya (Buddlejaceae).Journal of Systematics and Evolution 47:231–236.
CHEN, S.-L. AND P. S. TANG. 1945. Studies oncolchicine-induced autotetraploid barley. III. Phys-iological studies. American Journal of Botany32:177–179.
COUGHLAN, J. 2012. Mechanisms for range size anddistribution variation in the polyploid complexblack-fruited hawthorn (Crataegus series Dougla-sianae): biogeographic implications for the mainte-nance of cytotype diversity. M.Sc. thesis, Univer-sity of Toronto, Canada.
COYNE, J. A. AND H. A. ORR. 2004. Speciation.Sinauer Associates, Sunderland, MA.
CURTIS, A. B. 1986. Upper Elk Meadows ResearchNatural Area. USDA, Forest Service, Pacific North-west Forest and Range Experiment Station 1–20.
DENG, B., W. DU, C. LIU, W. SUN, S. TIAN, AND H.DONG. 2011. Antioxidant response to drought,cold and nutrient stress in two ploidy levels oftobacco plants: low resource requirement conferspolytolerance in polyploids? Plant Growth Regu-lation 66:37–47.
DICKINSON, T., S. BELAOUSSOFF, R. LOVE, AND M.MUNIYAMMA. 1996. North American black-fruitedhawthorns: I. Variation in floral construction,breeding system correlates, and their possibleevolutionary significance in Crataegus sect. Dou-glasii Loudon. Folia Geobotanica et Phytotaxono-mica 31:355–371.
DICKINSON, T. A. AND J. PHIPPS. 1985. Studies inCrataegus L. (Rosaceae: Maloideae). XIII. Degreeand pattern of phenotypic variation in Crataegussect. Crus-galli in Ontario. Systematic Botany10:322–337.
——— AND ———. 1986. Studies in Crataegus(Rosaceae: Maloideae) XIV. The breeding systemof Crataegus crus-galli sensu lato in Ontario.American Journal of Botany 73:116–130.
———, E. Y. Y. LO, N. TALENT, AND R. M. LOVE.2008. Black-fruited hawthorns of western NorthAmerica — one or more agamic complexes?Botany 86:846–865.
DRAY, S. AND P. LEGENDRE. 2008. Testing the speciestraits-environment relationships: the fourth-cornerproblem revisited. Ecology 89:3400–3412.
GANEVA, T., K. UZUNOVA, AND D. KOLEVA. 2009.Comparative leaf epidermis investigation in speciesof genus Crataegus L. (Rosaceae) from Bulgaria.Feddes Repertorium 120:169–184.
GRANT, V. 1981. Plant Speciation, 2nd ed. ColumbiaUniversity Press, New York, NY.
HADLEY, A. 2010. Combine Z. http://www.hadleyweb.pwp.blueyonder.co.uk/CZP/News.htm
HARGROVE, W. AND R. Luxmoore (eds.). 1998. A newhigh-resolution national map of vegetation eco-regions produced empirically using multivariatespatial clustering. Oak Ridge National Laboratory.Website http://www.geobabble.org/,hnw/esri98/(accessed August 2011).
HORANDL, E. 2006. The complex causality of geo-graphical parthenogenesis. New Phytologist171:525–538.
———. 2008. Evolutionary implications of self-com-patibility and reproductive fitness in the apomicticRanunculus auricomus polyploid complex (Ranun-culaceae). International Journal of Plant Sciences169:1219–1228.
JOACHIMIAK, A. AND A. GRABOWSKA-JOACHIMIAK.2000. Stomatal cell length and ploidy level in fourtaxa belonging to the Phleum sect. Phleum. ActaBiologica Cracoviensia Series Botanica 42:103–107.
JOLY, S. AND A. BRUNEAU. 2007. Delimiting speciesboundaries in Rosa sect. Cinnamomeae (Rosaceae)in eastern North America. Systematic Botany32:819–836.
KREBS, C. 2001. Ecology: the experimental analysis ofdistribution and abundance. Benjamin Cummings,San Francisco, CA.
2014] MCGOEY ET AL.: PLOIDY LEVEL AND STOMATA SIZE IN CRATAEGUS 191

LEGENDRE, P., R. GALZIN, AND M. L. HARMELIN-VIVIEN. 1997. Relating behavior to habitat: solu-tions to the fourth-corner problem. Ecology78:547–562.
LEVIN, D. A. 1983. Polyploidy and novelty in floweringplants. American Naturalist 122:1–25.
———. 2002. The role of chromosomal change in plantevolution. Oxford University Press, New York,NY.
LEWIS, W. H. 1980. Polyploidy in species populations.Pp. 103–144 in W. H. Lewis (ed.), Polyploidy:biological relevance. Proceedings of the Interna-tional Conference on Polyploidy, Biological Rele-vance, held at Washington University, St. Louis,Missouri, May 24–27, 1979. Plenum PublishingCorp., New York, NY.
LI, W., G. BERLYN, AND P. ASHTON. 1996. Polyploidsand their structural and physiological characteris-tics relative to water deficit in Betula papyrifera(Betulaceae). American Journal of Botany 83:15–20.
LO, E. Y. Y., S. STEFANOVIC, AND T. A. DICKINSON.2007. Molecular reappraisal of relationships be-tween Crataegus and Mespilus (Rosaceae, Pyr-eae)—Two genera or one? Systematic Botany32:596–616.
———, ———, AND ———. 2009. Population geneticstructure of diploid sexual and polyploid apomictichawthorns (Crataegus; Rosaceae) in the PacificNorthwest. Molecular Ecology 18:1145–1160.
———, ———, AND ———. 2010a. Reconstructingreticulation history in a phylogenetic frameworkand the potential of allopatric speciation driven bypolyploidy in an agamic complex in Crataegus(Rosaceae). Evolution 64:3593–3608.
———, ———, AND ———. 2013. Geographicalparthenogenesis in Pacific Northwest hawthorns(Crataegus; Rosaceae). Botany 91:107–116.
———, ———, K. RITLAND, AND T. A. DICKINSON.2010b. Fine-scale comparisons of genetic variabil-ity in seed families of asexually and sexuallyreproducing Crataegus (Hawthorn; Rosaceae).American Journal of Botany 97:1014–1024.
LOVE, R. M. AND M. FEIGEN. 1978. Interspecifichybridization between native and naturalizedCrataegus (Rosaceae) in western Oregon. Madrono25:211–217.
LOWRY, E. AND S. E. LESTER. 2006. The biogeographyof plant reproduction: potential determinants ofspecies’ range sizes. Journal of Biogeography 33:1975–1982.
MAHERALI, H., A. E. WALDEN, AND B. C. HUSBAND.2009. Genome duplication and the evolution ofphysiological responses to water stress. NewPhytologist 184:721–731.
MANZANEDA, A., P. REY, J. BASTIDA, C. WEISS-LEHMAN, E. RASKIN, AND T. MITCHELL-OLDS.2012. Environmental aridity is associated withcytotype segregation and polyploidy occurrence inBrachypodium distachyon (Poaceae). New Phytolo-gist 193:797–805.
MARSHALL, H. H. 1978. Identification of Crataegusspecies native to Manitoba. The Canadian Field-Naturalist 92:321–326.
MARTIN, S. L. AND B. C. HUSBAND. 2009. Influence ofphylogeny and ploidy on species ranges of NorthAmerican angiosperms. Journal of Ecology 97:913–922.
MCINTYRE, P. J. 2012. Polyploidy associated withaltered and broader ecological niches in theClaytonia perfoliata (Portulacaceae) species com-plex. American Journal of Botany 99:655–662.
MCNEILL, J., F. R. BARRIE, W. R. BUCK, V. DEMOULIN,W. GREUTER, D. L. HAWKSWORTH, P. S.HERENDEEN, S. KNAPP, K. MARHOLD, J. PRADO,W. F. PRUD’HOMME VAN REINE, G. F. SMITH, J. H.WIERSEMA, AND N. J. TURLAND. 2012. Internation-al Code of Nomenclature for algae, fungi, and plants(Melbourne Code) adopted by the EighteenthInternational Botanical Congress Melbourne, Aus-tralia, July 2011. Regnum Vegetabile 154. KoeltzScientific Books, Koenigstein, Germany.
MEACHAM, C. AND T. DUNCAN. 1991. MorphoSys: anautomated morphometric system. Version 1.29.University of California, Berkeley, Regents of theUniversity of California, Berkeley, CA.
METCALFE, C. AND L. CHALK. 1979. Anatomy of thedicotyledons, 2nd ed. Clarendon Press, Oxford,U.K.
NOGLER, G. A. 1984. Gametophytic apomixis. Pp. 475–518 in B. M. Johri (ed.), Embryology of angio-sperms. Springer-Verlag, Berlin.
OTTO, S. P. 2007. The evolutionary consequences ofpolyploidy. Cell 131:452–462.
PARISOD, C., R. HOLDEREGGER, AND C. BROCHMANN.2010. Evolutionary consequences of autopolyploi-dy. New Phytologist 186:5–17.
PHIPPS, J. 2012. Crataegus, in Jepson Flora Project(eds.), Jepson eFlora. Website http://ucjeps.berkeley.edu/cgi-bin/get_IJM.pl?tid591319 (accessed on 9June 2013).
——— AND R. O’KENNON. 1998. Three new species ofCrataegus (Rosaceae) from western North Amer-ica: C. okennonii, C. okanagenensis, and C. phippsii.Sida 18:169–191.
——— AND ———. 2002. New taxa of Crataegus(Rosaceae) from the northern Okanagan-south-western Shuswap diversity center. Sida 31:355–371.
PHIPPS, J. B., K. R. ROBERTSON, P. G. SMITH, AND
J. R. ROHRER. 1990. A checklist of the subfamilyMaloideae (Rosaceae). Canadian Journal of Bota-ny 68:2209–2269.
POTTER, D., T. ERIKSSON, R. C. EVANS, S. OH, J. E. E.SMEDMARK, D. R. MORGAN, M. KERR, K. R.ROBERTSON, M. ARSENAULT, T. A. DICKINSON,AND C. S. CAMPBELL. 2007. Phylogeny andclassification of Rosaceae. Plant Systematics andEvolution 266:5–43.
PRISM CLIMATE GROUP. 2012. PRISM Data Explor-er. Oregon State University. Website http://www.prism.oregonstate.edu/ (accessed August 2011).
R DEVELOPMENT CORE TEAM. 2004. R: A languageand environment for statistical computing. RFoundation for Statistical Computing. Websitehttp://www.r-project.org/ (accessed 15 April 2011).
RAMSEY, J. 2011. Polyploidy and ecological adaptationin wild yarrow. Proceedings of the NationalAcademy of Sciences of the United States ofAmerica 108:7096–7101.
SALTONSTALL, K., K. GLENNON, A. BURNETT, R. B.HUNTER, AND K. L. HUNTER. 2007. Comparisonof morphological variation indicative of ploidylevel in Phragmites australis (Poaceae) from easternNorth America. Rhodora 109:415–429.
SHORTHOUSE, D. P. 2010. SimpleMappr, an online toolto produce publication-quality point maps. Web-
192 MADRONO [Vol. 61

site http://www.simplemappr.net (accessed on 9June 2013).
SOLTIS, D. E., R. J. A. BUGGS, J. J. DOYLE, AND P. S.SOLTIS. 2010. What we still don’t know aboutpolyploidy. Taxon 59:1387–1403.
STEBBINS, G. L. 1950. Variation and evolution inplants. Columbia University Press, New York,NY.
TAL, M. AND I. GARDI. 1976. Physiology of polyploidplants: water balance in autotetraploid and diploidtomato under low and high salinity. PhysiologiaPlantarum 38:257–261.
TALENT, N. AND T. A. DICKINSON. 2005. Polyploidy inCrataegus and Mespilus (Rosaceae, Maloideae):evolutionary inferences from flow cytometry ofnuclear DNA amounts. Canadian Journal ofBotany 83:1268–1304.
——— AND ———. 2007. Endosperm formation inaposporous Crataegus (Rosaceae, Spiraeoideae,tribe Pyreae): parallels to Ranunculaceae andPoaceae. New Phytologist 173:231–249.
VASCAN. 2013. Crataegus gaylussacia A. Heller.Database of Vascular Plants of Canada (VAS-CAN). Website http://data.canadensys.net/vascan/taxon/20019 (accessed on 7 June 2013).
WANG, H. AND J. CLARKE. 1993. Genotypic, intra-plant, and environmental variation in stomatalfrequency and size in wheat. Canadian Journal ofPlant Science 73:671–678.
ZARREI, M., N. TALENT, J. COUGHLAN, S. STEFANOVIC,AND T. A. DICKINSON. 2012. Nuclear ribosomal ITS2DNA sequences illuminate hybridization in a diploid-polyploid agamic complex of Crataegus (Rosaceae).Presented at Botany 2012, Columbus, OH.
2014] MCGOEY ET AL.: PLOIDY LEVEL AND STOMATA SIZE IN CRATAEGUS 193