head and neck squamous cell growth suppression using...
TRANSCRIPT

Vol. 3. 185-191, February 1997 Clinical Cancer Research 185
Head and Neck Squamous Cell Growth Suppression Using
Adenovirus-p53-FLAG: A Potential Marker for Gene
Therapy Trials’
S. Mark Overholt,2 Ta-Jen Liu,2
Dorothy L. Taylor, Mary Wang,
Adel K. El-Naggar, Ed Shillitoe,
Karen Adler-Storthz, Lisa St. John,
Wei-Wei Zhang, Jack A. Roth, and
Gary L. Clayman3
Departments of Head and Neck Surgery [S. M. 0.. T-J. L., D. L. 1.,
M. W.. G. L. Cl, Pathology [A. K. E-N.l, Section of Thoracic
Molecular Oncology. and Department of Thoracic and Cardiovascular
Surgery IW-W. Z.. J. A. Ri. The University of Texas M. D. Anderson
Cancer Center, Houston, Texas 77030; Department of Microbiology
and Immunology. State University of New York. Syracuse, New
York 13210 IE. 5.1: Department of Basic Sciences. The University of
Texas Health Science Center Dental Branch, Houston, Texas 77030
1K. A-SI: and University of Texas. Houston, Graduate School of
Biomedial Sciences, Houston. Texas 77030 IL. S. J.I
ABSTRACT
The recombinant wild-type p53 adenovirus has been
proven effective against the growth of human head and neck
squamous cell cancer (SCCHN) cell lines in vitro and in a
nude mouse model. The addition of a FLAG peptide se-
quence was used in this study, along with the p53 adenovirus
vector as a marker of the site of the gene therapy activity. It
provides clear evidence of the exogenous gene product
within the transduced carcinoma cells. No alterations in
transcription or translation of the p53 gene product were
noted with the addition of the FLAG sequence to the original
p53 adenovirus vector. Immunohistochemical analysis dis-
played simultaneous expression of the p53 and FLAG pro-
teins in the infected cells. The p53 protein remained local-
Received 9/16/96: revised I 1/7/96: accepted I 1/8/96.
The costs of publication of this article were defrayed in part by the
payment of page charges. This article must therefore be hereby marked
advertisement in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
I This work was supported in part by American Cancer Society Career
Development Award 93-9 (to G. L. C.): M. D. Anderson Cancer Center
Core Grant NIH-National Cancer Institute CA- I6672: National CancerInstitute Grant ROl CA-45l87 (to J. A. R.): Training Grant CA0961 I
(to J. A. R.); National Institute Grant RS5DE/OD10846 (to K. A-S.):National Institute Grant ROl DE-l0842 (to E. S.): National Science
Foundation Graduate Research Fellowship (to L. S. J.): gifts to the
Division of Surgery from Tenneco and Exxon for the Core Lab Facility:
and a generous gift from the Mathers Foundation.2 Contributed equally to the concept. investigation. and writing of this
work.
3To whom requests for reprints should be addressed. at Department ofHead and Neck Surgery. The University of Texas M. D. Anderson
Cancer Center, I 5 15 Holcombe Boulevard. Box 69, Houston, TX
77030. Fax: (713) 794-4662.
ized to the nucleus, whereas the FLAG protein was
additionally noted in the cytoplasm. In vitro growth suppres-
sion assays and in vivo microscopic residual tumor model
experiments in nude mice showed a similar tumoricidal
effect with the p53-FLAG adenovirus vector to that with the
previously studied p53 adenovirus vector without the addi-
tion of the FLAG sequence. We conclude that the addition of
the FLAG octapeptide sequence allows identification of
those cells that have been affected by the molecular therapy
independent of the endogenous gene expression of the cells.
This novel molecular tracer may prove useful in character-
izing infection efficiency and in gene therapy trials.
INTRODUCTION
Locoregional control of disease, despite improvement over
the past several decades, continues to impact significantly on
patients with SCCHN.4 Distant metastases fortuitously occur
late in the course of these tumors and. overall, occur in only
approximately I I % ( I ). Although long-term goals remain fo-
cused on the complicated sequence of molecular events leading
to carcinoma, the immediate goals that challenge researchers are
2-fold: (a) detect changes in epithelium before overt malignancy
occurs; and (b) develop strategies to eradicate local and regional
disease with minimal host morbidity. Brennan et ai. (2) have
recently described the use of p53 satellite mapping around the
resection margins of SCCHNs. They demonstrated that muta-
tions of the p53 gene in the margins of resection increased the
risk of local recurrence. Boyle et a!. (3) found that p53 muta-
tions precede invasion in SCCHN. Mutations of the p53 gene
are present in up to 60% ofSCCHNs (4-8). As new markers for
premalignancy develop with greater site and tumor specificity
and sensitivity. we may be able to predict early progression
toward malignancy.
Gene therapy strategies have shown promise in controlling
local and regional tumor burden in both in vitro and in animal
models (9-15). Specifically. in a head and neck squamous cell
carcinoma. growth suppression and cytotoxicity have been doe-
umented using the wild-type p53 gene delivered via a replica-
lion-defective adenovirus vector (9). A dose-dependent tumori-
cidal effect independent of the host endogenous p53 status has
additionally been shown. The mechanism of action appears to be
apoptosis (16. 17). Furthermore. a recent microscopic residual
disease model that mimics postsurgical resection situations in
humans has been described ( 10). Such a model lays the ground-
work for carryover application in a human trial.
4 The abbreviations used are: SCCHN, head and neck squamous cell
carcinoma: Ad, adenovirus: CMV. cytomegalovirus: pfu. plaque-form-
ing unit(s): TNF. tumor necrosis factor.
Research. on July 14, 2018. © 1997 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

186 p53-FLAG: Potential Marker for Gene Therapy Trials
As more promising therapies develop and we entertain the
application of gene therapy in the head and neck, the ability to
trace the sites affected by the therapy and identify which cells
have been affected by treatment becomes more important. De-
livery of a novel tracer would, therefore, provide definitive
evidence for infection efficiency and extent of effect, both
locally and distantly.
The FLAG biosystem (18), originally described in 1988,
allows one to tag and examine the eft’eet of an exogenously applied
treatment. Originally, it was used in the purification of large pro-
teins, but its design lends itself to other applications. This strategy
generates a fusion protein of the FLAG peptide and the gene of
interest. A monoclonal antibody against the polypeptide enables us
to detect the location of a fusion protein that would correspond to
clonal expression of the delivered gene. The polypeptide is eight
amino acids in length, and its small size should not disrupt the
expression of the delivered gene therapy protein. If the biological
activity remains unchanged, the ability to co-deliver such a marker
holds great implication for future human trials. The goal of this
investigation was to compare the biological effect of the previously
described wild-type pS3 adenovirus to that of the vector modified
with the FLAG octapeptide sequence. We demonstrated that the
biological effect was not changed by the addition of the FLAG
sequence to the adenovirus p53 vector (AdCMV-p53), and that by
using immunohistochemical techniques, we have the ability to trace
the location of the effect following the adenovirus administration.
MATERIALS AND METHODS
Cell Lines and Culture Conditions. Human SCCHN
cell lines Tu-l 38 and 686-LN were both established by the De-
partment of Head and Neck Surgery. The University of Texas
M. D. Anderson Cancer Center, and have been characterized pre-
viously (19, 20). These cells were grown in DMEM/Fl2 medium
supplemented with 10% heat-inactivated fetal bovine serum with
streptomycin/penicillin at 37#{176}Cand 5% carbon dioxide.
Recombinant Adenovirus Preparation and Infection.
The recombinant p53 adenovirus (AdCMV-p53) contains the
CMV promoter, wild-type p53 eDNA, and SV4O polyadenyl-
ation signal in a minigene cassette inserted into the El-deleted
region of modified AdS (21). Viral stocks were propagated in
293 cells. Cells were harvested 36-40 h after infection, pel-
leted, resuspended in PBS, and lysed. Cell debris was then
removed by subjecting the cells to CsCl gradient purification.
Concentrated virus was dialyzed, aliquoted, and stored at
-80#{176}C.Infection was carried out by the addition of virus to the
DMEMIFI2 medium with 10% fetal bovine serum to the cell
monolayers. The cells were incubated at 37#{176}Cfor 60 mm with
agitation. Then complete medium (DMEMIF12 with 10% fetal
bovine serum) was added to the cells in the appropriate volume
for the given Petri dish and incubated for the desired time.
Generation of the p53-FLAG Adenovirus. The p53
eDNA sequence was excised from the pCS3-SN [kindly pro-
vided by Dr. G. Lozano (The University of Texas M. D. Ander-
son Cancer Center, Houston, IX)1 by digestion with BwnHI and
cloned into the BainHI site of pGEM7Z. A recombinant plasmid
with the proper insert orientation was then digested with AccI
and Kpnl to remove 21 amino acids from the 3’ end of p53
eDNA. A linker with AccI-KjmI compatible ends containing the
sequence of the FLAG peptide including a stop codon was then
ligated into the digested plasmid to create the p53-FLAG fusion
gene. The resulting p53-FLAG fusion gene was then cloned into
an expression vector with the human CMV promoter and SV
polyadenylation signal. The final construct was subsequently
inserted into a shuttle vector pXCJL. I (2 1 ) to generate a recom-
binant p53-FLAG adenovirus.
Northern Blot Analysis. Total RNA was isolated by the
acid-guanidinium thiocyanate method of Chomczynski and Sacehi
(22). Northern analyses were performed on 20 �.tg oftotal RNA run
on a 1% agarose gel containing 2.2 M formaldehyde. The mem-
brane was hybridized overnight at 65#{176}Cwith a p53 eDNA probe
labeled by the random primer method in 5 X SSC, 5 X Denhardt’s
solution, 0.5% SDS denatured salmon sperm DNA (20 j.tg/ml).
Western Blot Analysis. Total cell lysates were prepared
by sonicating the cells 24 h after infection in RIPA buffer ( 150
mM NaC1, 1.0% NP4O, 0.5% sodium deoxycholate. 0.1% SDS,
and 50 filM Iris, pH 8.0). Fifty �.tg of protein from samples were
subjected to 10% PAGE and transferred to a Hybond-ECL
membrane (Amersham Corp.). The membrane was blocked with
Blotto/Tween (5% nonfat dry milk and 0.2% Tween 20 in PBS)
and probed with the primary antibodies. anti-FLAG M2 mono-
clonal antibody (IB13002), mouse anti-human p53 monoclonal
antibody (PAb I 80 1 ), and mouse anti-human B-actin mono-
clonal antibody (Amersham). The secondary antibody, horse-
radish peroxidase-conjugated goat anti-mouse IgG (Boehringer
Mannheim, Indianapolis, IN), was then exposed to the mem-
brane. The membrane was then processed and developed as the
manufacturer specified.
Cell Growth Assay. Cells were plated at a concentration
of 2 X l0� cells/mI in six-well plates in triplicate, and cells were
infected with AdCMV-p53-FLAG, AdCMV-p53. or the repli-
cation-defective adenovirus control (DL3I2), at an multiplicity
of infection of I 00. A mock infection was also used as a control,
against which to compare the effect of the replication-deficient
adenovirus and AdCMV-p53. Cells were then trypsinized and
counted at specified time points. Viability was checked using
trypan blue exclusion.
Immunohistochemical Staining of in Vitro Cell Layers.
Infected cell monolayers were fixed with 3.8% buffered forma-
lin and treated with H,O� in methanol for 5 mm. Immunohis-
tochemical staining was performed using the Vectastain Elite kit
(Vector Laboratories, Burlingame. CA). The primary antibody
was either the mouse anti-FLAG M2 monoclonal antibody
(IB13002) or mouse anti-human p53 monoclonal antibody
(PAbl8Ol), and the secondary antibody was an avidin-labeled
anti-mouse IgG (Vector). The biotinylated horseradish peroxi-
dase avidin-biotin complex reagent was used to detect the anti-
gen-antibody complex. Preadsorption controls were used in each
experiment. The cells were then counterstained with Harris
hematoxylin (Sigma Chemical Co., St. Louis, MO).
Immunohistochemical Staining of Tissue from in Vivo
Studies. Formalin-fixed, paraffin-embedded in vito animal
experimental tissues were cut at 4-5 p.si. dried at 60#{176}C.depar-
af’finized, and hydrated with distilled water. Sections were then
treated with 0.5% saponin in distilled water and rinsed in several
changes of distilled water; endogenous peroxidase activity was
blocked with 3% hydrogen peroxide in methanol, followed by
rinsing in several changes of distilled water. Sections were
Research. on July 14, 2018. © 1997 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

1234567 89
2.8 Kb
1.9 Kb
Endogenous
Exogenous
Clitiical Cancer Research 187
Fig. I Expressiots of exogenous p53
mRNA 24 h after infection with
AdCMV-p53 and AdCMV-p53-
FLAG. La,u’ I. RNA molecular
weight marker. LAUl(’S’ 2-5, Tv- I 38
(niutated PS3) cell line. Lane.s 6-9.
686-LN (wild-type PS3) cell line.
Laiies 2 and 6. mock infection. Lanes
3 and 7. DL3 I 2 infection. Lotus 4 and
8. AdCMV-p53 infection. Lanes’ 5 and
9. AdCMV-p53-FLAG infection.
microwave-irradiated in distilled water for 3 mm using a Sharp
model R9H8 I microwave oven operating at a frequency of 2450
MHz at 700 W. After cooling, sections were washed in several
changes of distilled water and placed in PBS; immunochemical
studies were performed by using the avidin-biotin-peroxidase
complex method of Hsu ci ai. (23) in the following manner.
Sections were blocked with normal horse serum and incubated
overnight at 4#{176}Cwith anti-FLAG M2 monoclonal antibody at a
concentration of 60 p.g/ml (International Biotechnologies, New
Haven, CT) and rabbit antihurnan p53 polyclonal antibody,
clone OM-1. 1:80 (Signet Laboratories, Denham. MA). An
antirabbit IgG Elite kit (Vector) was then used to apply bioti-
nylated antirabbit IgG and avidin-biotin-peroxidase complexes
that were incubated for 45 mm each. The immunostaining
reaction was visualized by using 0.5% dimethylaminoazoben-
zene in PBS containing 0.01% H,02 (pH 7.6), counterstained
with 0.01% toluidine blue. dehydrated. cleared. and mounted in
Permount.
In Vivo Microscopic Residual Disease Experiments. In
t’ivo experiments were performed in a defined pathogen-free
environment using athymic nude mice. The microscopic resid-
ual disease model described previously by our laboratory was
applied ( 10). Experiments were reviewed and approved by in-
stitutional review committees for both animal care and utiliza-
tion and the Biosafety Committee for recombinant DNA re-
search. Briefly, 4-7-week-old female nude mice were
anesthetized using i.p. ketamine/acepromazine at a dose of 70
mg/kg of body weight. The bodies were prepared with alcohol
wipes and incisions were made on four dorsal flanks. s.c. pock-
ets were elevated, and the desired number of tumor cells was
delivered in 100 pi of medium into the pocket. The flap was
then sealed with an interrupted horizontal mattress 5.0 nylon
suture, insuring a water-tight seal. At 48 h, the flaps were
reopened, and 100 ii of an appropriate concentration of virus
were delivered into the same bed inoculated previously with the
tumor cells. Two different sets of repeated experiments were
performed. The first was a dose-response experiment using the
AdCMV-p53-FLAG virus in three of the flaps at descending
concentrations ( l0�, l0�, 106 pfu). The fourth flap served as a
control and was randomized to either PBS or the replication-
defective adenovirus (DL312). The second study was performed
using l0� pfu of AdCMV-p53-FLAG. AdCMV-p53, and repli-
cation-defective adenovirus in three separate flaps. The fourth
flap was inoculated with the same volume ( 100 p.1) of sterile
PBS. Forty-eight h after treatment with virus, two animals were
sacrificed, and the flaps were harvested for immunohistochem-
ical analysis. The remaining animals were observed for 2 1 days
and then sacrificed. Tumor volumes were measured for corn-
parison using calipers.
RESULTS
Expression of mRNA after Infection with AdCMV-p53
and AdCMV-p53-FLAG Virus. Both To- I 38 and 686-LN
were examined for expression of p53 mRNA. Total RNA was
isolated after adenovirus infection. Northern blot analysis was
performed. Similar levels of exogenous AdCMV-p53 rnRNA
were detected between AdCMV-p53- and AdCMV-p53-FLAG-
infected cells. Fig. I shows the comparable levels of p53 mRNA
expression af’ter infection with AdCMV-p53 and AdCMV-p53-
FLAG in Lanes 4 and S for Iu-138 and in Lanes 8 and 9 for
MDA 686-LN. Variation of intensity is felt to be related to
loading dose. Endogenous expression of p53 rnRNA is seen in
Lanes 2 and 3 in the mutated p53 cell line, Iu-138. Lanes 6 and
7 show no endogenous p53 rnRNA in the 686-LN cell line,
which is wild-type for the p53 gene. These data suggest that the
AdCMV-p53-FLAG virus, like the AdCMV-p53 virus, is sue-
cessfully transduced and efficiently transcribed. Northern anal-
ysis did not reveal evidence of AdCMV-p53 DNA contarnina-
tion.
Expression of Exogenous p53 Protein in AdCMV-p53-
and AdCMV-p53-FLAG-infected SCCHN Cell Lines.
Western blot analysis was performed to compare the amount of
protein expressed by the AdCMV-p53- and AdCMV-p53-
FLAG-infected cells. Protein bands were identified using the
monospecifie p53 antibody (PAbl8Ol) and the anti-FLAG M2
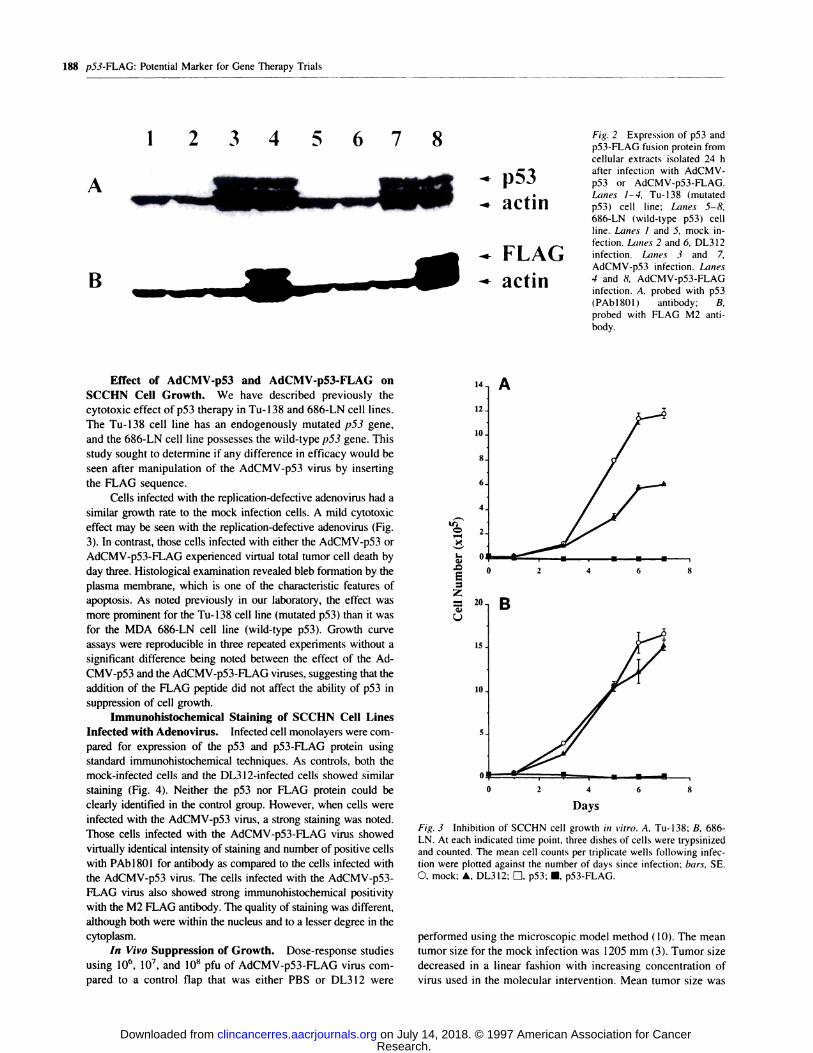
antibody (1B13002) on two simultaneously run gels. Fig. 2 (top)
showed, using the p53 antibody (PAbl8Ol ), a similar high level
of p53 protein expression in both cell lines that were infected
with the AdCMV-p53 and AdCMV-p53-FLAG. Lanes 3 and 7
correspond to the To- I 38 and MDA 686-LN cells infected with
AdCMV-p53, respectively. Lanes 4 and S of the blot represent
protein expression from those cells infected with the AdCMV-
p53-FLAG. No change in p53 protein expression was noted in
either the replication-defective, adenovirus-infeeted cells or in
the mock group. The bottom blot demonstrates a similarly
executed gel that was probed with the mouse anti-FLAG M2
antibody. The level of p53-FLAG protein expression appeared
to be similar to that expressed following p53 antibody probing.
but no detectable band was noted in those cells infected with the
AdCMV-p53 virus (Fig. 2. lanes 3 and 7). The mock and
DL3I2-infeeted cells exhibited no detectable level of the im-
munoreactive p53 protein in either cell line.
Research. on July 14, 2018. © 1997 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
A
0 2
B
6 8
tn
L
.nE
2:
LI
Days
Fig. 3 Inhibition of SCCHN cell growth in vitro. A, Tu-138; B, 686-
LN. At each indicated time point, three dishes of cells were trypsinized
and counted. The mean cell counts per triplicate wells following infec-
tion were plotted against the number of days since infection; bars, SE.
0, mock; A, DL312; EL PS3; U, p53-FLAG.
2 4 6 8
188 p53-FLAG: Potential Marker for Gene Therapy Trials
A
B
I 2345678
�*p53
,�. actin
.� FLAG
..�- actin
Fig. 2 Expression of p53 and
p53-FLAG fusion protein from
cellular extracts isolated 24 h
after infection with AdCMV-
p53 or AdCMV-p53-FLAG.
Lanes 1-4, Tu-138 (mutated
pS3) cell line; Lanes 5-8,
686-LN (wild-type p53) cell
line. Lanes I and 5, mock in-
fection. Lanes 2 and 6, DL3 12infection. Lanes 3 and 7,
AdCMV-p53 infection. Lanes
4 and 8, AdCMV-p53-FLAGinfection. A, probed with p53(PAbl8Ol) antibody; B,
probed with FLAG M2 anti-
body.
Effect of AdCMV-p53 and AdCMV-p53-FLAG on
SCCHN Cell Growth. We have described previously the
cytotoxic effect of p53 therapy in Tu-l38 and 686-LN cell lines.
The Iu-l38 cell line has an endogenously mutated p53 gene,
and the 686-LN cell line possesses the wild-type p53 gene. This
study sought to determine if any difference in efficacy would be
seen after manipulation of the AdCMV-p53 virus by inserting
the FLAG sequence.
Cells infected with the replication-defective adenovirus had a
similar growth rate to the mock infection cells. A mild cytotoxic
effect may be seen with the replication-defective adenovirus (Fig.
3). In contrast, those cells infected with either the AdCMV-p53 or
AdCMV-p53-FLAG experienced virtual total tumor cell death by
day three. Histological examination revealed bleb formation by the
plasma membrane, which is one of the characteristic features of
apoptosis. As noted previously in our laboratory, the effect was
more prominent for the Tu-138 cell line (mutated p53) than it was
for the MDA 686-LN cell line (wild-type p53). Growth curve
assays were reproducible in three repeated experiments without a
significant difference being noted between the effect of the Ad-
CMV-p53 and the AdCMV-p53-FLAG viruses, suggesting that the
addition of the FLAG peptide did not affect the ability of p53 in
suppression of cell growth.
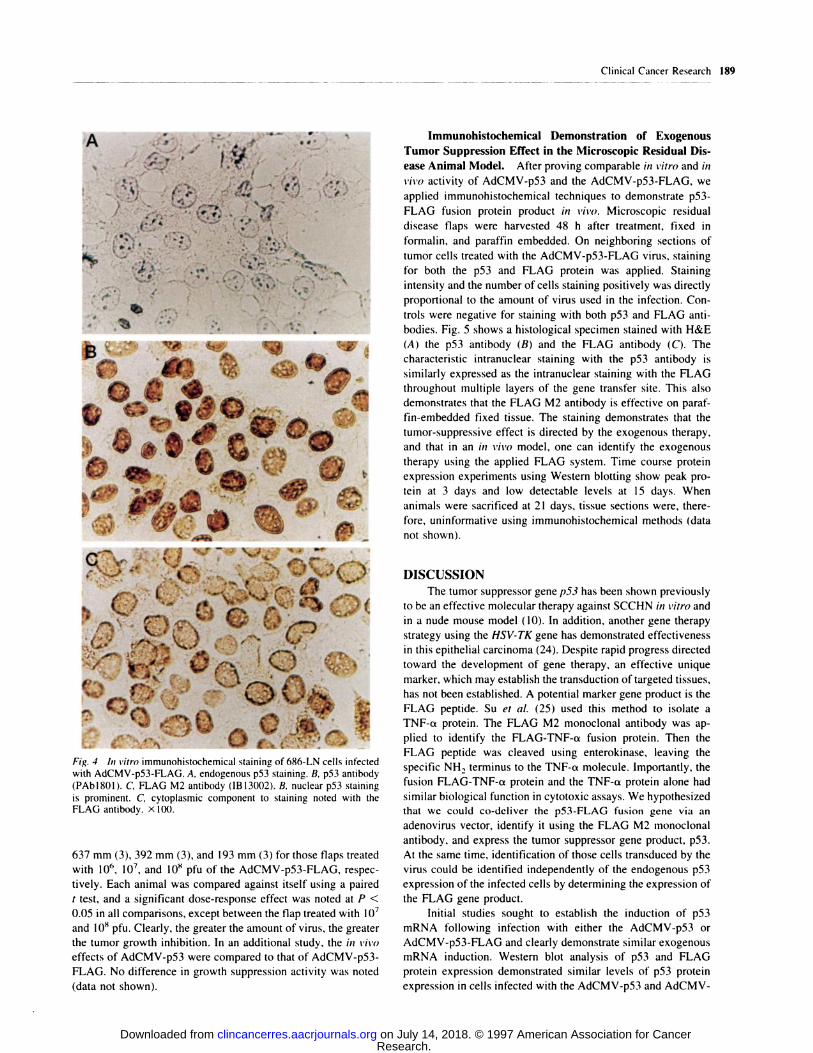
Immunohistochemical Staining of SCCHN Cell Lines
Infected with Adenovirus. Infected cell monolayers were corn-
pared for expression of the p53 arid p53-FLAG protein using
standard immunohistochemical techniques. As controls, both the
mock-infected cells and the DL3 12-infected cells showed similar
staining (Fig. 4). Neither the p53 nor FLAG protein could be
clearly identified in the control group. However, when cells were
infected with the AdCMV-p53 virus, a strong staining was noted.
Those cells infected with the AdCMV-p53-FLAG virus showed
virtually identical intensity of staining and number of positive cells
with PAbl8Ol for antibody as compared to the cells infected with
the AdCMV-p53 virus. The cells infected with the AdCMV-p53-
FLAG virus also showed strong immunohistochemical positivity
with the M2 FLAG antibody. The quality of staining was different,
although both were within the nucleus and to a lesser degree in the
cytoplasm.
In Vivo Suppression of Growth. Dose-response studies
using 106, l0�, and 108 pfu of AdCMV-p53-FLAG virus com-
pared to a control flap that was either PBS or DL3 12 were
performed using the microscopic model method (10). The mean
tumor size for the mock infection was 1205 mm (3). Tumor size
decreased in a linear fashion with increasing concentration of
virus used in the molecular intervention. Mean tumor size was
Research. on July 14, 2018. © 1997 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Clinical Cancer Research 189
A .. .‘ . 1’ ;-: .� #{149}..
‘, I � #{149}� � �‘ �, �, ‘ -
- � �A � �
� �‘ ,‘-.-�.. -- .. ,1 #{149}� � :‘,,.�‘T)I.,..
-‘. . I ‘�: � � �1
.(a. � � .‘s%�., ‘#{149}-.
. s-... .-‘
‘4 .% . : . L��#{149} #{149}‘:�� . �:
0#{149}0
0�
�
c� �
#{149}.:‘� � � g’:�t-��e .
Fig. 4 In vitro immunohistochemical staining of 686-LN cells infected
with AdCMV-p53-FLAG. A, endogenous p53 staining. B, p53 antibody
(PAbl8Ol). C, FLAG M2 antibody (1B13002). B, nuclear p53 staining
is prominent. C’, cytoplasmic component to staining noted with the
FLAG antibody. x 100.
637 mm (3), 392 mm (3), and 193 mm (3) for those flaps treated
with 106, l0�, and l0� pfu of the AdCMV-p53-FLAG, respec-
tively. Each animal was compared against itself using a paired
t test, and a significant dose-response effect was noted at P <
0.05 in all comparisons, except between the flap treated with l0�
and l0� pfu. Clearly, the greater the amount of virus, the greater
the tumor growth inhibition. In an additional study, the in vito
effects of AdCMV-p53 were compared to that of AdCMV-p53-
FLAG. No difference in growth suppression activity was noted
(data not shown).
Immunohistochemical Demonstration of Exogenous
Tumor Suppression Effect in the Microscopic Residual Dis-
ease Animal Model. After proving comparable in vitro and in
t,ivo activity of AdCMV-p53 and the AdCMV-p53-FLAG. we
applied immunohistoehemical techniques to demonstrate p53-
FLAG fusion protein product in vito. Microscopic residual
disease flaps were harvested 48 h after treatment, fixed in
formalin, and paraffin embedded. On neighboring sections of
tumor cells treated with the AdCMV-p53-FLAG virus, staining
for both the p53 and FLAG protein was applied. Staining
intensity and the number of cells staining positively was directly.� . proportional to the amount of virus used in the infection. Con-
trols were negative for staining with both p53 and FLAG anti-
bodies. Fig. S shows a histological specimen stained with H&E
(A) the p53 antibody (B) and the FLAG antibody (C). The
characteristic intranuclear staining with the p53 antibody is
similarly expressed as the intranuclear staining with the FLAG
throughout multiple layers of the gene transfer site. This also
demonstrates that the FLAG M2 antibody is effective on paraf-
fin-embedded fixed tissue. The staining demonstrates that the
tumor-suppressive effect is directed by the exogenous therapy,
and that in an in vito model, one can identify the exogenous
therapy using the applied FLAG system. lime course protein
expression experiments using Western blotting show peak pro-
tein at 3 days and low detectable levels at 15 days. When
animals were sacrificed at 21 days, tissue sections were, there-
fore, uninformative using immunohistoehemical methods (data
not shown).
DISCUSSION
The tumor suppressor gene p.53 has been shown previously
to be an effective molecular therapy against SCCHN in vitro and
in a nude mouse model (10). In addition, another gene therapy
strategy using the HSV-TK gene has demonstrated effectiveness
in this epithelial carcinoma (24). Despite rapid progress directed
toward the development of gene therapy, an effective unique
marker, which may establish the transduction oftargeted tissues,
has not been established. A potential marker gene product is the
FLAG peptide. So et ai. (25) used this method to isolate a
TNF-a protein. The FLAG M2 monoclonal antibody was ap-
plied to identify the FLAG-INF-a fusion protein. Then the
FLAG peptide was cleaved using enterokinase, leaving the
specific NH2 terminus to the TNF-a molecule. Importantly, the
fusion FLAG-TNF-a protein and the TNF-a protein alone had
similar biological function in cytotoxic assays. We hypothesized
that we could co-deliver the p53-FLAG fusion gene via an
adenovirus vector, identify it using the FLAG M2 monoclonal
antibody, and express the tumor suppressor gene product, p53.
At the same time, identification of those cells transduced by the
virus could be identified independently of the endogenous p53
expression of the infected cells by determining the expression of
the FLAG gene product.
Initial studies sought to establish the induction of p53
mRNA following infection with either the AdCMV-p53 or
AdCMV-p53-FLAG and clearly demonstrate similar exogenous
mRNA induction. Western blot analysis of p53 and FLAG
protein expression demonstrated similar levels of p53 protein
expression in cells infected with the AdCMV-p53 and AdCMV-
Research. on July 14, 2018. © 1997 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

B� “ “ . �.
. .
190 p53-FLAG: Potential Marker for Gene Therapy Trials
‘� “ ‘ ‘ .‘:“ “.- ‘ ‘ . . “ � ,�4 ‘
.. . . ..‘ .. . . .. :: : , . . � � t.1��P;t�
� � .‘ ‘�. �
‘ ‘ . .. - .: ,‘. �
�. �‘. I �‘ .: . ; , . ,.- ., .. � .‘-.� , C ��“�‘�-\ A
�;J�.1C.-.’ ‘ ‘ ‘ : , � . , , .
w
� :‘�;‘ � �N�I�’ � :
� .
� ,, . ,‘.
� ‘.- -�‘ . ...
� � ‘� � � , �
Fig. 5 Immunohistochemical staining of formalin-fixed, paraffin-em-bedded tissue from in vito Tu-I38 tumor treated with AdCMV-p53-
FLAG. A, H&E. B, p53 antibody (PAbl8Ol). C. FLAG M2 antibody
(IBI3002). B, strong nuclear staining noted with p53 antibody. C. strong
nuclear staining is also seen utilizing the FLAG antibody. X 100.
p53-FLAG, respectively. Furthermore, the FLAG antibody
demonstrated that the protein expressed by the cells infected
with the AdCMV-p53-FLAG was a unique fusion protein ex-
pressing the FLAG octapeptide moiety that could be differenti-
ated from the native p53 protein.
Northern analysis confirmed that the cells infected with
either the AdCMV-p53 or AdCMV-p53-FLAG exhibited simi-
lar exogenous mRNA induction. Our previous studies using the
AdCMV-p53 had demonstrated marked induction of apoptosis
among infected cancer cell lines ( 16, 26); however, nonmalig-
nant cell lines were spared. The next question was whether the
addition of the FLAG sequence to the putative p53 protein
would alter its inherent growth-inhibitory effect. To this end.
cell growth assays were performed, and no significant difference
could be demonstrated between AdCMV-p53 and AdCMV-p53-
FLAG. As we had described previously, induction of apoptosis
was not observed following mock or replication-defective virus
infection.
In vitro immunohistochemical studies were performed to
establish that the FLAG protein could be identified among
infected SCCHN cell lines. We were able to show a strong
nuclear staining for the p53 protein in those cells infected by
either the AdCMV-p53 or the AdCMV-p53-FLAG. Further-
more, using the FLAG antibody, the novel FLAG octapeptide
sequence was demonstrated in those cells that were infected
with the AdCMV-p53-FLAG only. There appeared to be a slight
qualitative difference in the staining with a probable cytoplas-
mic component when probing with the FLAG monoclonal
antibody.
in t’it’() experiments were carried out to further explore the
efficacy of FLAG immunohistochemical staining. Using the
microscopic residual disease model described previously. s.c.
tumor sites were infected with increasing concentrations of the
AdCMV-p53-FLAG vector. A dose-response curve was noted
with greater tumor kill as the virus concentration was increased.
Flaps infected with AdCMV-p53-FLAG could be effectively
immunohistoehemically analyzed for both the p53 and FLAG
proteins using formalin-fixed, paraffin-embedded tissues. The
large pS3 protein was noted to be primarily intranuclear. as
documented previously. However, the FLAG protein was noted
additionally within the cytoplasm. Because the nuclear translo-
cation signal was altered in the p53 3’-end deletion in this
FLAG construct, both nuclear and cytoplasniic localization of
the transgene product were found (27). We were not able to
demonstrate staining for the FLAG protein outside the region of
treatment, confirming the local expression of these virions and
lack of systemic expression when delivered in this model.
FLAG appears to be a potential marker for tracking protein
product expression in gene therapy. We currently have the
technology to use virus-specific promoters and PCR techniques
to analyze the presence ofdelivered virions. As a screening tool,
this would be an effective and sensitive test for identifyi�ig virus.
However, it does not answer the more important question: is the
protein product encoded by the delivered virus being expressed?
Herein lies the major advantage of the FLAG marker. Fixed
tissue can be analyzed for the presence of the delivered gene
product. This is particularly important in tumor biology, where
endogenous heterogeneity for the genes of interest might mask
locations where a specific gene therapy has been delivered.
Specifically, in tumors with a mutated p53 gene, exogenous
expression of the vector-delivered p53 can be differentiated
from the overexpression of the endogenous p53 of the tumor.
Such a marker would also demonstrate the expression and effect
imparted on normal bystander cells. Both of these lend them-
selves to carryover into human trials, where such questions must
be addressed.
Potential criticisms of FLAG as a marker for gene therapy
are 2-fold: (a) a minimal protein expression is required to
Research. on July 14, 2018. © 1997 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Clinical Cancer Research 191
immunohistochemically identify the gene product. For simple
documentation of the presence of virion, the sensitivity is far
less than that of PCR. However, the transduction efficiency of
the replication-defective adenovirus in SCCHN is excellent, and
p53 protein expression is high. The added advantage of docu-
menting expression of the viral product and its histological
location distinguishes this technique; (b) introduction of novel
genes has the potential to cause deleterious downstream effects.
Using a retroviral vector, these potential effects would be irre-
versible, but with the transient episomal expression of the ade-
novirus, this hazard is avoided. Lastly, the p53-FLAG and
p53-adenoviral vectors both show evidence of inflammatory
infiltrates in animal studies at viral doses of l0� pfu and greater.
Local inflammatory responses may, however, be beneficial in
the local tumor environment. The potential systemic immune
response from viral products and exogenous gene products will
require further investigations.
In conclusion, we feel that the co-delivery of the FLAG
protein along with the desired gene therapy offers potential
utility as a marker of gene therapy. We were able to show that
it was simultaneously promoted along with the p53 gene and
that expression of the mRNA and protein were not decreased.
More importantly, the biological activity of the delivered tumor
suppressor gene was not altered. For the first time, the FLAG
antibody was proven effective when immunohistoehemical
analysis was perf’ormed on formalin-fixed, paraffin-embedded
tissue. These factors suggest the utility of this novel protein as
a tracer in further gene therapy studies.
REFERENCES
1. Calhoun, K. H., Fulmer, P., Weiss, R., and Hokanson, J. A. Distant
metastases from head and neck squamous cell carcinomas. Laryngo-
scope, 104: 1199-1205, 1994.
2. Brennan, J. A., Mao, L., Hruban, R. H., Boyle, J. 0., Eby, Y. J..Koch, W. M., Goodman, S. N., and Sidransky, D. Molecular assessmentof histopathological staging in squamous-cell carcinoma of the head and
neck. N. EngI. J. Med., 332: 429-435. 1995.
3. Boyle. J. 0.. Hakim, J., Koch, W., van der Riet, P., Hruban, R. H.,
Roa, R. A., Correo, R., Eby, Y. J., Ruppert, M., and Sidransky, D. Theincidence of p53 mutation increases with progression of head and neck
cancer. Cancer Res., 53: 4477-4480, 1993.
4. Field, J. K.. Pavelic, Z. P., Spandidos, D. A.. Stambrook, P. J.. Jones.
A. S., and Gluckman. J. L. The role of the p53 tumor suppressor gene
in squamous cell carcinoma of the head and neck. Arch. Otolaryngol.
Head Neck Surg., 119: 1 1 18-1 122, 1993.
5. Watling, D. L., Gown, A. M., and Coltrera, M. D. Overexpression of
p53 in head and neck cancer. Head Neck, 14: 437-444, 1992.
6. Nylander. K., Stenling, R., Gustafsson, H., Zackrisson, B., and Root,G. p53 expression and cell proliferation in squamous cell carcinomas of
the head and neck. Cancer (Phila.), 75: 87-93, 1995.
7. Pavelic, Z. P.. Li, Y-Q., Stambrook, P. J., McDonald, J. S., Munck-
Wikland, E., Pavelic, K., Dacic, S., Danilovic, Z., Pavelic, L., Mugge,
R. E., Wilson, K., Nguyen, C.. and Gluckman, J. L. Overexpression of
p53 protein is common in premalignant head and neck lesions. Anti-
cancer Res., 14: 2259-2266, 1994.
8. Maestro, R., Doleetti, R., Gasparatto, D., Doglioni, C., Pelucchi, S.,
Barzan, L., Grandi, E., and Boiocchi, M. High frequency of p53 gene
alterations associated with protein overexpression in human squamous
cell carcinoma of the larynx. Oncogene, 7: 1 159-1 166, 1992.
9. Liu, T-J., Zhang, W-W., Taylor, D. L., Roth, J. A., Goepfert, H., and
Clayman. G. L. Growth suppression of human head and neck cancer
cells by the introduction of a wild-type p53 gene via a recombinant
adenovirus. Cancer Res., 54: 3662-3667, 1994.
10. Clayman, G. L., El-Naggar, A. K., Roth, J. A.. Zhang. W-W.,Goepfert. H.. Taylor. D. L., and Liu, 1-i. In vito molecular therapy withp53 adenovirus for microscopic residual head and neck squamous car-
cinoma. Cancer Res., 55: 1-6, 1995.
1 1 . Baker, S. J., Markowitz, S., Fearon, E. R., Wilson, J. R., and
Vogelstein, B. Suppression of human colorectal carcinoma cell growth
by wild-type p53. Science (Washington DC), 249.’ 912-915, 1990.
12. Mercer, W. E., Shields, M. 1., Amin, M., Sauve, G. J., Apella, E.,
Romano, J. W., and Ulrich, S. J. Negative growth regulation in aglioblastoma tumor cell line that conditionally expresses human wild-
Type p53. Proc. NatI. Acad. Sci. USA, 87: 6166-6170, 1990.
13. Diller, L., Kassel, J., Nelson, C. E., Gryka, M. A., Litwank, G.,
Gebhardt, M., Bressac, B., Ozturk, M., Baker, J. J., and Vogelstein, B.p53 functions as a cell cycle control protein in osteosarcoma. Mol. Cell.
Biol.. 10: 5772-5781. 1990.
14. Shaw, P., Bovey, R., Tardy, S., Sahli, R., Sordet, B., and Costa,
J. Induction of apoptosis by wild-type p53 in human colon tumor-
derived cell line. Proc. NatI. Acad. Sci. USA, 89: 4495-4499, 1992.
15. Fujiwara. 1., Grimm, E. A., Murkhopadhyay, 1., Cal, D. W.,Owen-Schaub, L. B., and Roth, J. A. A retroviral wild-type p53 expres-sion vector penetrates human lung cancer spheroids and inhibits growth
by inducing apoptosis. Cancer Res., 53: 4129-4133, 1993.
16. Liu, T-J., El-Naggar, A. K., McDonnell, 1. J., Steck, K. D., Wang,M., Taylor. D. L.. and Clayman. G. L. Apoptosis induction mediated by
wild-type p53 adenovirus gene transfer in squamous cell carcinoma ofthe head and neck. Cancer Res., 55: 31 17-3122, 1995.
I 7. Hartwell, L. H., and Kastan, M. B. Cell cycle control and cancer.
Science (Washington DC), 266: 1821-1828, 1994.
18. Hopp, 1. P., Prickett, K. S., Price, V., Libby, R. 1., March, C. J.,
Cerretti, P., Urdal, D. L., and Conlon, P. J. A short polypeptide marker
sequence useful for identification and purification of recombinant pro-
teins. Biotechnology, 7: 1205-1210, 1988.
19. Clayman, G. L., Wang, S. W., Nicholson, G. L., El-Naggar, A. K..Mazar, A.. Henkins, J., Blasi, F., Goepfert, H., and Boyd, D. D.
Regulation of urokinase-type plasminogen activator expression in squa-
mous-cell carcinoma ofthe oral cavity. Int. J. Cancer, 54: 73-80, 1993.
20. Sacks, P. G., Parnes, S. M., Gallick, G. E., Mansouri, Z., Lichtner,
R., Satya-Prakash. K. L.. Pathak, S., and Parsons, D. F. Establishmentand characterization of two new squamous cell carcinoma cell lines
derived From tumors of the head and neck. Cancer Res., 48: 2858-
2866, 1988.
21. Zhang. W-W.. Fang, X., Mazur, W., French, B. A., Georges, R. N.,
and Roth, J. A. High-efficiency gene transfer and high-level expression
of wild-type p53. I. Human lung cancer cells mediated by recombinant
adenovirus. Cancer Gene Ther., I: 1-10, 1994.
22. Chomczynski, P., and Sacchi, N. Single-step method of RNA iso-
lation by guanidinium thiocyanate-phenol-chloroform extraction. Anal.
Biochem., 162: 156-159, 1987.
23. Hsu, S. M., Raine, L., and Fanger, H. Use of avidin-biotin-perox-
idase complex (ABC) in immunoperoxidase techniques: a comparison
between ABC and unlabeled antibody (PAP) procedures. J. Histochem.Cytochem.. 29: 577-580, 1981.
24. O’Malley. B. W., Jr., Chen, S-H., Schwartz, M. R., and Woo,
S. L. C. Adenovirus-mediated gene therapy for human head and neck
squamous cell cancer in a nude mouse model. Cancer Res., 55: 1080-
1085, 1995.
25. So, X., Prestwood, A. K., and McGraw, R. A. Production of
recombinant porcine tumor necrosis factor a in a novel E. cohi expres-sion system. Biotechniques, /3: 756-761, 1992.
26. Wang. J., Bucana, C. D., Roth, J. A., and Zhang, W-W. Apoptosisinduced in human osteosarcoma cells is one of the mechanisms for the
cytocidal effect of AdCMV-p53. Cancer Gene Ther., 2: 1-9, 1995.
27. Shaulsky, G., Goldfinger, N., Ben-Ze’ev, A., and Rotter, V. Nuclear
accumulation of p53 protein is mediated by several localization signals
and plays a role in tumorigenesis. Mol. Cell Biol.. 10: 6565-6577, 1990.
Research. on July 14, 2018. © 1997 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

1997;3:185-191. Clin Cancer Res S M Overholt, T J Liu, D L Taylor, et al. trials.adenovirus-p53-FLAG: a potential marker for gene therapy Head and neck squamous cell growth suppression using
Updated version
http://clincancerres.aacrjournals.org/content/3/2/185
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/3/2/185To request permission to re-use all or part of this article, use this link
Research. on July 14, 2018. © 1997 American Association for Cancerclincancerres.aacrjournals.org Downloaded from