heart failure - pdfs.semanticscholar.org · april j. beyer, ms; vijaya b. nareddy, ms; a. martin...
TRANSCRIPT

Heart Failure
Omega-3 Fatty Acids Prevent Pressure Overload–InducedCardiac Fibrosis Through Activation of Cyclic GMP/Protein
Kinase G Signaling in Cardiac FibroblastsJinghai Chen, PhD; Gregory C. Shearer, PhD; Quanhai Chen, MD; Chastity L. Healy, BS;
April J. Beyer, MS; Vijaya B. Nareddy, MS; A. Martin Gerdes, PhD; William S. Harris, PhD;Timothy D. O’Connell, PhD; Dajun Wang, MD
Background—Omega-3 polyunsaturated fatty acids (eicosapentaenoic acid and docosahexaenoic acid) from fish oilameliorate cardiovascular diseases. However, little is known about the effects of �-3 polyunsaturated fatty acids oncardiac fibrosis, a major cause of diastolic dysfunction and heart failure. The present study assessed the effects of �-3polyunsaturated fatty acids on cardiac fibrosis.
Methods and Results—We assessed left ventricular fibrosis and pathology in mice subjected to transverse aorticconstriction after the consumption of a fish oil or a control diet. In control mice, 4 weeks of transverse aortic constrictioninduced significant cardiac dysfunction, cardiac fibrosis, and cardiac fibroblast activation (proliferation and transfor-mation into myofibroblasts). Dietary supplementation with fish oil prevented transverse aortic constriction–inducedcardiac dysfunction and cardiac fibrosis and blocked cardiac fibroblast activation. In heart tissue, transverse aorticconstriction increased active transforming growth factor-�1 levels and phosphorylation of Smad2. In isolated adultmouse cardiac fibroblasts, transforming growth factor-�1 induced cardiac fibroblast transformation, proliferation, andcollagen synthesis. Eicosapentaenoic acid and docosahexaenoic acid increased cyclic GMP levels and blocked cardiacfibroblast transformation, proliferation, and collagen synthesis. Eicosapentaenoic acid and docosahexaenoic acidblocked phospho-Smad2/3 nuclear translocation. DT3, a protein kinase G inhibitor, blocked the antifibrotic effects ofeicosapentaenoic acid and docosahexaenoic acid. Eicosapentaenoic acid and docosahexaenoic acid increased phosphor-ylated endothelial nitric oxide synthase and endothelial nitric oxide synthase protein levels and nitric oxide production.
Conclusion—Omega-3 fatty acids prevent cardiac fibrosis and cardiac dysfunction by blocking transforming growthfactor-�1–induced phospho-Smad2/3 nuclear translocation through activation of the cyclic GMP/protein kinase Gpathway in cardiac fibroblasts. (Circulation. 2011;123:584-593.)
Key Words: cyclic GMP � protein kinase G � fatty acids � heart failure
Heart failure is the leading cause of cardiovascular mor-bidity and mortality worldwide.1 Up to 50% of heart
failure patients have diastolic dysfunction.2 One of the maincauses of diastolic dysfunction is cardiac fibrosis characterizedby a pathological accumulation of fibrillar collagen throughoutthe myocardium, which results in reduced cardiac musclecompliance, impaired filling, and ultimately heart failure.3,4
Clinical Perspective on p 593In the heart, collagen is produced primarily by cardiac
fibroblasts. In response to stress, profibrotic cytokines arereleased, leading to cardiac fibroblast proliferation and trans-
formation into myofibroblasts. Myofibroblasts have contrac-tile fibers containing �-smooth muscle actin (�-SMA) andare responsible for the excessive accumulation of extracellu-lar matrix under pathological conditions.3,5 Therefore, abro-gation of cardiac fibroblast transformation into myofibro-blasts is 1 strategy for suppressing cardiac fibroticremodeling, which can lead to heart failure.
Cardiac fibroblast transformation is induced primarily bytransforming growth factor (TGF)-�1,6,7 which, under patho-logical conditions, can result in excessive collagen produc-tion.8 TGF-�1 binds to type II (TGF�RII) and type I(TGF�RI) receptors, which phosphorylate TGF�RI-
Received January 25, 2010; accepted December 20, 2010.From the Cardiovascular Health Research Center at Sanford Research/University of South Dakota, Sioux Falls.Dr Chen’s current affiliation is Department of Cardiology, Cardiovascular Research Division, Children’s Hospital Boston, Harvard Medical School,
Boston, MA. Dr Gerdes’ current affiliation is Department of Biomedical Sciences, New York College of Osteopathic Medicine at New York Instituteof Technology, Old Westbury, NY. Dr Wang’s current affiliation is Aurora Sinai Medical Center, Milwaukee, WI.
The online-only Data Supplement is available with this article at http://circ.ahajournals.org/cgi/content/full/CIRCULATIONAHA.110.971853/DC1.Correspondence to Timothy D. O’Connell, PhD, Cardiovascular Health Research Center, Sanford Research/USD, 2301 E. 60th St N, Sioux Falls, SD
57104. E-mail: [email protected] or Dajun Wang, MD, Aurora Sinai Medical Center, 945 N 12th St, Milwaukee, WI 53201. [email protected]
© 2011 American Heart Association, Inc.
Circulation is available at http://circ.ahajournals.org DOI: 10.1161/CIRCULATIONAHA.110.971853
584
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2017http://circ.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2017http://circ.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2017http://circ.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2017http://circ.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2017http://circ.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2017http://circ.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2017http://circ.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2017http://circ.ahajournals.org/
Dow
nloaded from
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
by guest on A
ugust 31, 2017http://circ.ahajournals.org/
Dow
nloaded from
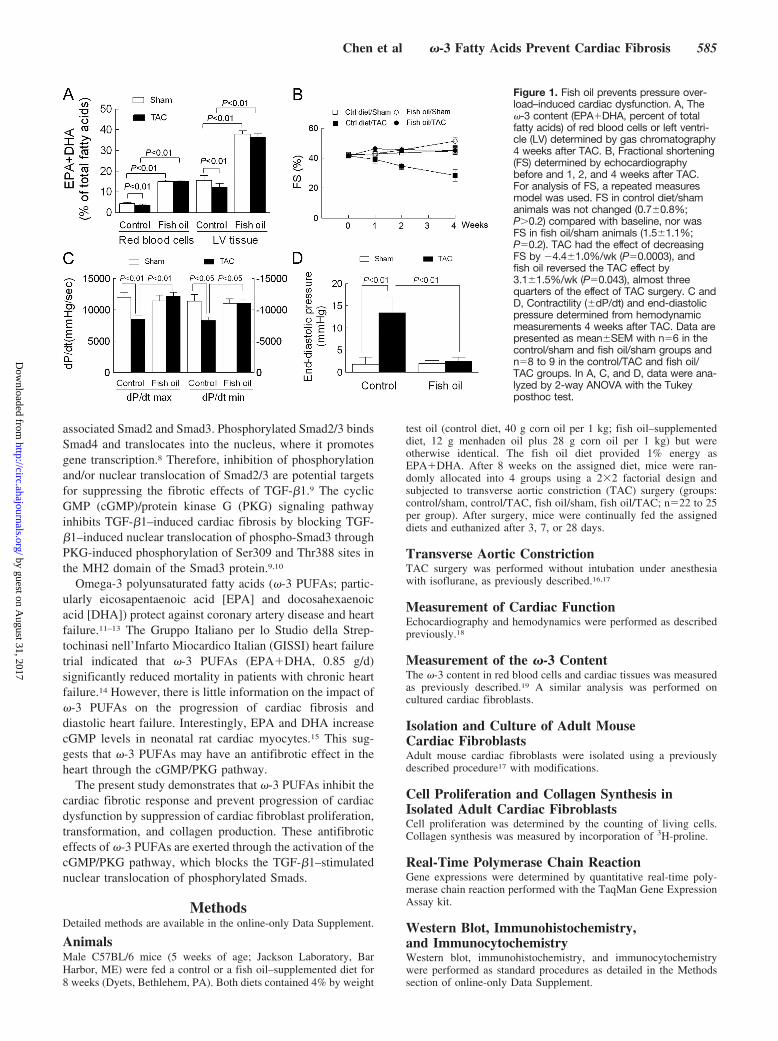
associated Smad2 and Smad3. Phosphorylated Smad2/3 bindsSmad4 and translocates into the nucleus, where it promotesgene transcription.8 Therefore, inhibition of phosphorylationand/or nuclear translocation of Smad2/3 are potential targetsfor suppressing the fibrotic effects of TGF-�1.9 The cyclicGMP (cGMP)/protein kinase G (PKG) signaling pathwayinhibits TGF-�1–induced cardiac fibrosis by blocking TGF-�1–induced nuclear translocation of phospho-Smad3 throughPKG-induced phosphorylation of Ser309 and Thr388 sites inthe MH2 domain of the Smad3 protein.9,10
Omega-3 polyunsaturated fatty acids (�-3 PUFAs; partic-ularly eicosapentaenoic acid [EPA] and docosahexaenoicacid [DHA]) protect against coronary artery disease and heartfailure.11–13 The Gruppo Italiano per lo Studio della Strep-tochinasi nell’Infarto Miocardico Italian (GISSI) heart failuretrial indicated that �-3 PUFAs (EPA�DHA, 0.85 g/d)significantly reduced mortality in patients with chronic heartfailure.14 However, there is little information on the impact of�-3 PUFAs on the progression of cardiac fibrosis anddiastolic heart failure. Interestingly, EPA and DHA increasecGMP levels in neonatal rat cardiac myocytes.15 This sug-gests that �-3 PUFAs may have an antifibrotic effect in theheart through the cGMP/PKG pathway.
The present study demonstrates that �-3 PUFAs inhibit thecardiac fibrotic response and prevent progression of cardiacdysfunction by suppression of cardiac fibroblast proliferation,transformation, and collagen production. These antifibroticeffects of �-3 PUFAs are exerted through the activation of thecGMP/PKG pathway, which blocks the TGF-�1–stimulatednuclear translocation of phosphorylated Smads.
MethodsDetailed methods are available in the online-only Data Supplement.
AnimalsMale C57BL/6 mice (5 weeks of age; Jackson Laboratory, BarHarbor, ME) were fed a control or a fish oil–supplemented diet for8 weeks (Dyets, Bethlehem, PA). Both diets contained 4% by weight
test oil (control diet, 40 g corn oil per 1 kg; fish oil–supplementeddiet, 12 g menhaden oil plus 28 g corn oil per 1 kg) but wereotherwise identical. The fish oil diet provided 1% energy asEPA�DHA. After 8 weeks on the assigned diet, mice were ran-domly allocated into 4 groups using a 2�2 factorial design andsubjected to transverse aortic constriction (TAC) surgery (groups:control/sham, control/TAC, fish oil/sham, fish oil/TAC; n�22 to 25per group). After surgery, mice were continually fed the assigneddiets and euthanized after 3, 7, or 28 days.
Transverse Aortic ConstrictionTAC surgery was performed without intubation under anesthesiawith isoflurane, as previously described.16,17
Measurement of Cardiac FunctionEchocardiography and hemodynamics were performed as describedpreviously.18
Measurement of the �-3 ContentThe �-3 content in red blood cells and cardiac tissues was measuredas previously described.19 A similar analysis was performed oncultured cardiac fibroblasts.
Isolation and Culture of Adult MouseCardiac FibroblastsAdult mouse cardiac fibroblasts were isolated using a previouslydescribed procedure17 with modifications.
Cell Proliferation and Collagen Synthesis inIsolated Adult Cardiac FibroblastsCell proliferation was determined by the counting of living cells.Collagen synthesis was measured by incorporation of 3H-proline.
Real-Time Polymerase Chain ReactionGene expressions were determined by quantitative real-time poly-merase chain reaction performed with the TaqMan Gene ExpressionAssay kit.
Western Blot, Immunohistochemistry,and ImmunocytochemistryWestern blot, immunohistochemistry, and immunocytochemistrywere performed as standard procedures as detailed in the Methodssection of online-only Data Supplement.
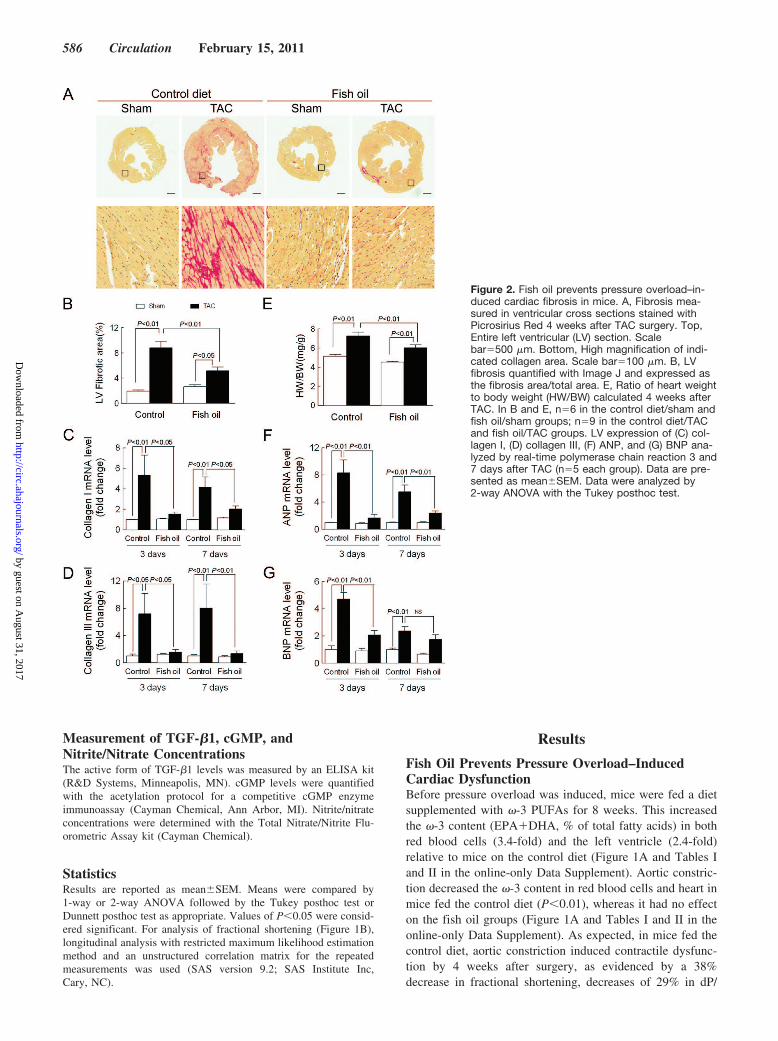
Figure 1. Fish oil prevents pressure over-load–induced cardiac dysfunction. A, The�-3 content (EPA�DHA, percent of totalfatty acids) of red blood cells or left ventri-cle (LV) determined by gas chromatography4 weeks after TAC. B, Fractional shortening(FS) determined by echocardiographybefore and 1, 2, and 4 weeks after TAC.For analysis of FS, a repeated measuresmodel was used. FS in control diet/shamanimals was not changed (0.7�0.8%;P�0.2) compared with baseline, nor wasFS in fish oil/sham animals (1.5�1.1%;P�0.2). TAC had the effect of decreasingFS by �4.4�1.0%/wk (P�0.0003), andfish oil reversed the TAC effect by3.1�1.5%/wk (P�0.043), almost threequarters of the effect of TAC surgery. C andD, Contractility (�dP/dt) and end-diastolicpressure determined from hemodynamicmeasurements 4 weeks after TAC. Data arepresented as mean�SEM with n�6 in thecontrol/sham and fish oil/sham groups andn�8 to 9 in the control/TAC and fish oil/TAC groups. In A, C, and D, data were ana-lyzed by 2-way ANOVA with the Tukeyposthoc test.
Chen et al �-3 Fatty Acids Prevent Cardiac Fibrosis 585
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
Measurement of TGF-�1, cGMP, andNitrite/Nitrate ConcentrationsThe active form of TGF-�1 levels was measured by an ELISA kit(R&D Systems, Minneapolis, MN). cGMP levels were quantifiedwith the acetylation protocol for a competitive cGMP enzymeimmunoassay (Cayman Chemical, Ann Arbor, MI). Nitrite/nitrateconcentrations were determined with the Total Nitrate/Nitrite Flu-orometric Assay kit (Cayman Chemical).
StatisticsResults are reported as mean�SEM. Means were compared by1-way or 2-way ANOVA followed by the Tukey posthoc test orDunnett posthoc test as appropriate. Values of P�0.05 were consid-ered significant. For analysis of fractional shortening (Figure 1B),longitudinal analysis with restricted maximum likelihood estimationmethod and an unstructured correlation matrix for the repeatedmeasurements was used (SAS version 9.2; SAS Institute Inc,Cary, NC).
Results
Fish Oil Prevents Pressure Overload–InducedCardiac DysfunctionBefore pressure overload was induced, mice were fed a dietsupplemented with �-3 PUFAs for 8 weeks. This increasedthe �-3 content (EPA�DHA, % of total fatty acids) in bothred blood cells (3.4-fold) and the left ventricle (2.4-fold)relative to mice on the control diet (Figure 1A and Tables Iand II in the online-only Data Supplement). Aortic constric-tion decreased the �-3 content in red blood cells and heart inmice fed the control diet (P�0.01), whereas it had no effecton the fish oil groups (Figure 1A and Tables I and II in theonline-only Data Supplement). As expected, in mice fed thecontrol diet, aortic constriction induced contractile dysfunc-tion by 4 weeks after surgery, as evidenced by a 38%decrease in fractional shortening, decreases of 29% in dP/
Figure 2. Fish oil prevents pressure overload–in-duced cardiac fibrosis in mice. A, Fibrosis mea-sured in ventricular cross sections stained withPicrosirius Red 4 weeks after TAC surgery. Top,Entire left ventricular (LV) section. Scalebar�500 �m. Bottom, High magnification of indi-cated collagen area. Scale bar�100 �m. B, LVfibrosis quantified with Image J and expressed asthe fibrosis area/total area. E, Ratio of heart weightto body weight (HW/BW) calculated 4 weeks afterTAC. In B and E, n�6 in the control diet/sham andfish oil/sham groups; n�9 in the control diet/TACand fish oil/TAC groups. LV expression of (C) col-lagen I, (D) collagen III, (F) ANP, and (G) BNP ana-lyzed by real-time polymerase chain reaction 3 and7 days after TAC (n�5 each group). Data are pre-sented as mean�SEM. Data were analyzed by2-way ANOVA with the Tukey posthoc test.
586 Circulation February 15, 2011
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from

dtmax and 27% in dP/dtmin, and a 6.5-fold increase in end-di-astolic pressure (all at least P�0.05 relative to sham). Con-versely, contractile dysfunction was prevented after aorticconstriction in mice fed the fish oil diet (Figure 1B through 1D).There was no significant difference in pressure gradients in-duced by aortic constriction between the control diet and fish oildiet groups (Table III in the online-only Data Supplement). Inshort, dietary supplementation with fish oil protects againstpressure overload–induced contractile dysfunction.
Fish Oil Prevents Pressure Overload–InducedCardiac Fibrosis in the Left VentricleTo determine whether dietary supplementation with fish oilprevents pressure overload–induced cardiac fibrosis, tissueswere stained with Picrosirius Red to detect collagen distributionin left ventricular cross sections 4 weeks after surgery. In micefed the control diet, aortic constriction induced significantfibrosis. The collagen content was increased 4.6-fold over sham(Figure 2A and 2B). This was accompanied by a significantincrease in collagen I and III expression at both 3 and 7 daysafter surgery (collagen I: 5.3-fold at 3 days, 4.2-fold at 7 days;collagen III: 7.2-fold at 3 days, 8.0-fold at 7 days; Figure 2C and2D). However, in mice fed the fish oil diet, fibrosis was
increased only 2.0-fold over sham, and there was no induction ofcollagen expression, resulting in 63% less fibrosis comparedwith the mice fed the control diet. In summary, fish oilconsumption prevents fibrosis induced by pressure overload.
A previous report indicated that dietary supplementation withfish oil reduced hypertrophy after aortic constriction in rats.20
Here, aortic constriction induced a hypertrophic response asevidenced by a 42% increase in the ratio of heart weight to bodyweight in mice fed the control diet and a 31% increase in theratio of heart weight to body weight in mice fed the fish oil diet.However, after adjustment for the effect of fish oil, aorticconstriction increased the ratio of heart weight to body weight by1.9�0.3 in both the control and fish oil groups (Figure 2E).Conversely, in mice fed the control diet, aortic constrictionsignificantly induced the expression of the hypertrophic markergenes atrial (ANP) and brain natriuretic peptide (BNP), whichwas not observed in mice fed the fish oil diet (Figure 2F and 2G).
Fish Oil Blocks Pressure Overload–InducedNonmyocyte Proliferation andMyofibroblast TransformationTo determine how fish oil feeding affected cardiac fibroblastproliferation and transformation after aortic constriction, we
Figure 3. Fish oil blocks pressure overload–in-duced nonmyocyte proliferation and myofibroblasttransformation. A, Fibroblast proliferation mea-sured in ventricular sections stained with an anti-body to Ki67 (green), phalloidin (red; labels cardiacmyocytes), and DAPI (blue; labels nuclei) 4 weeksafter TAC. Arrows show interstitial colabeling ofKi67 and DAPI. Scale bar�20 �m. B, Myofibro-blast transformation measured in ventricular sec-tions stained with an antibody to �-SMA 4 weeksafter TAC. Scale bar�200 �m. Quantification of (C)the number of Ki67-positive cells from 15 to 20fields per heart and (D) the percent area of�-SMA–positive staining in whole sections. Dataare presented as mean�SEM with n�4 per group.Data were analyzed by 2-way ANOVA with theTukey posthoc test.
Chen et al �-3 Fatty Acids Prevent Cardiac Fibrosis 587
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
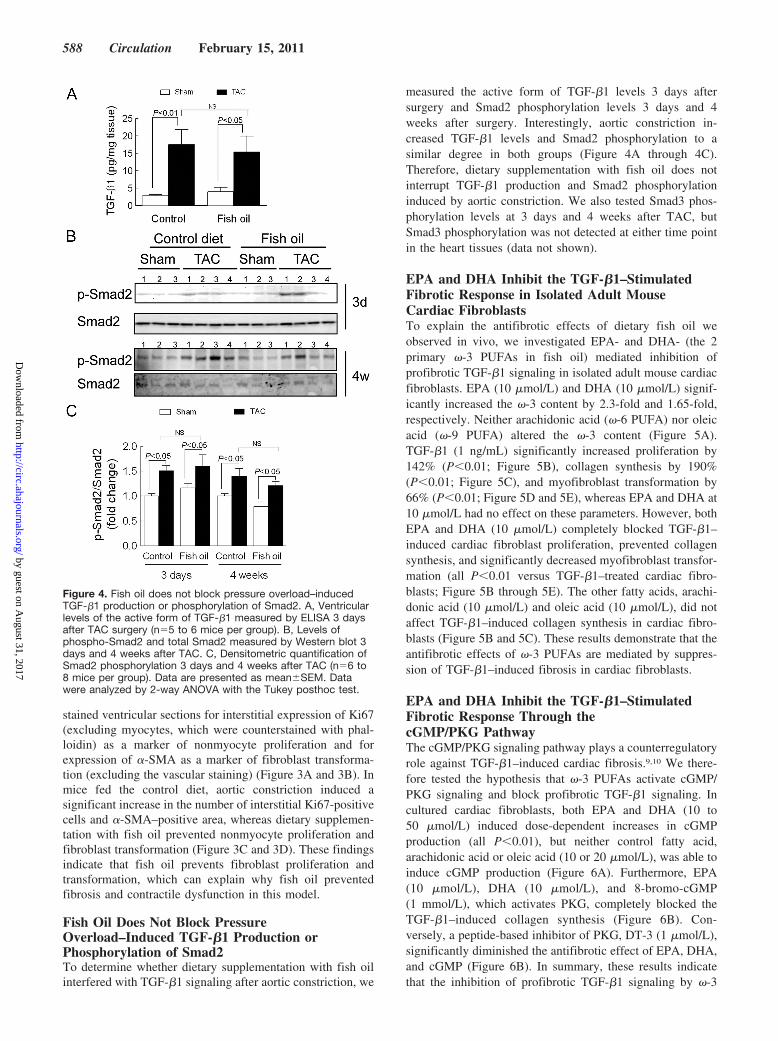
stained ventricular sections for interstitial expression of Ki67(excluding myocytes, which were counterstained with phal-loidin) as a marker of nonmyocyte proliferation and forexpression of �-SMA as a marker of fibroblast transforma-tion (excluding the vascular staining) (Figure 3A and 3B). Inmice fed the control diet, aortic constriction induced asignificant increase in the number of interstitial Ki67-positivecells and �-SMA–positive area, whereas dietary supplemen-tation with fish oil prevented nonmyocyte proliferation andfibroblast transformation (Figure 3C and 3D). These findingsindicate that fish oil prevents fibroblast proliferation andtransformation, which can explain why fish oil preventedfibrosis and contractile dysfunction in this model.
Fish Oil Does Not Block PressureOverload–Induced TGF-�1 Production orPhosphorylation of Smad2To determine whether dietary supplementation with fish oilinterfered with TGF-�1 signaling after aortic constriction, we
measured the active form of TGF-�1 levels 3 days aftersurgery and Smad2 phosphorylation levels 3 days and 4weeks after surgery. Interestingly, aortic constriction in-creased TGF-�1 levels and Smad2 phosphorylation to asimilar degree in both groups (Figure 4A through 4C).Therefore, dietary supplementation with fish oil does notinterrupt TGF-�1 production and Smad2 phosphorylationinduced by aortic constriction. We also tested Smad3 phos-phorylation levels at 3 days and 4 weeks after TAC, butSmad3 phosphorylation was not detected at either time pointin the heart tissues (data not shown).
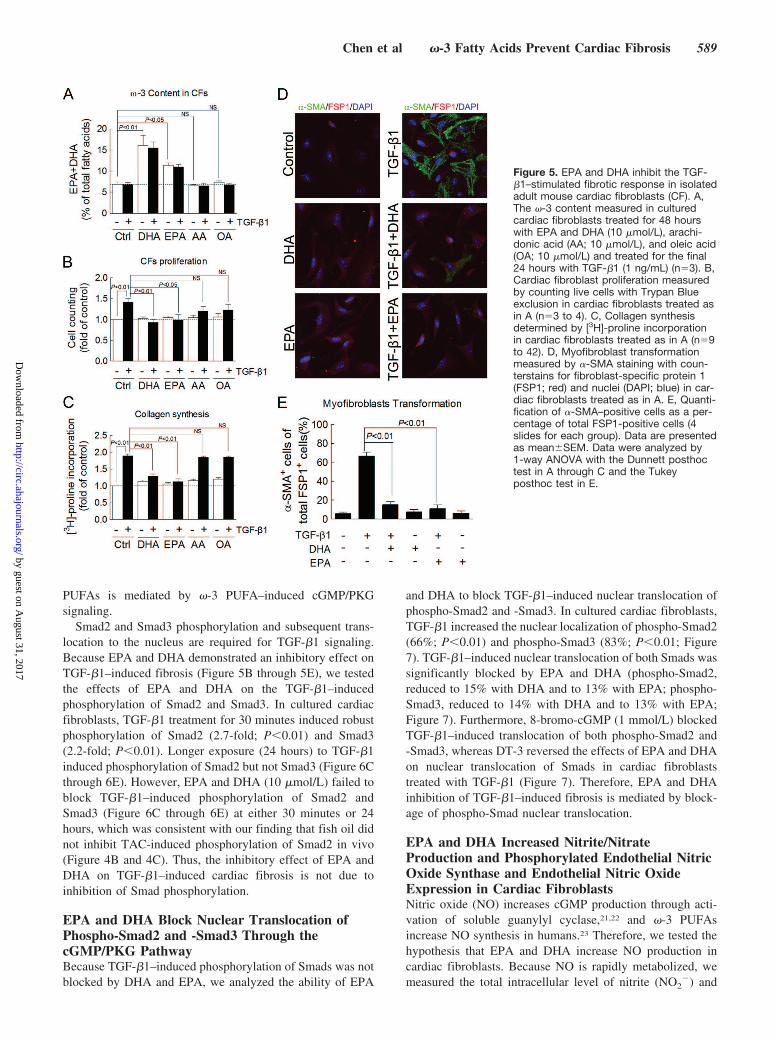
EPA and DHA Inhibit the TGF-�1–StimulatedFibrotic Response in Isolated Adult MouseCardiac FibroblastsTo explain the antifibrotic effects of dietary fish oil weobserved in vivo, we investigated EPA- and DHA- (the 2primary �-3 PUFAs in fish oil) mediated inhibition ofprofibrotic TGF-�1 signaling in isolated adult mouse cardiacfibroblasts. EPA (10 �mol/L) and DHA (10 �mol/L) signif-icantly increased the �-3 content by 2.3-fold and 1.65-fold,respectively. Neither arachidonic acid (�-6 PUFA) nor oleicacid (�-9 PUFA) altered the �-3 content (Figure 5A).TGF-�1 (1 ng/mL) significantly increased proliferation by142% (P�0.01; Figure 5B), collagen synthesis by 190%(P�0.01; Figure 5C), and myofibroblast transformation by66% (P�0.01; Figure 5D and 5E), whereas EPA and DHA at10 �mol/L had no effect on these parameters. However, bothEPA and DHA (10 �mol/L) completely blocked TGF-�1–induced cardiac fibroblast proliferation, prevented collagensynthesis, and significantly decreased myofibroblast transfor-mation (all P�0.01 versus TGF-�1–treated cardiac fibro-blasts; Figure 5B through 5E). The other fatty acids, arachi-donic acid (10 �mol/L) and oleic acid (10 �mol/L), did notaffect TGF-�1–induced collagen synthesis in cardiac fibro-blasts (Figure 5B and 5C). These results demonstrate that theantifibrotic effects of �-3 PUFAs are mediated by suppres-sion of TGF-�1–induced fibrosis in cardiac fibroblasts.
EPA and DHA Inhibit the TGF-�1–StimulatedFibrotic Response Through thecGMP/PKG PathwayThe cGMP/PKG signaling pathway plays a counterregulatoryrole against TGF-�1–induced cardiac fibrosis.9,10 We there-fore tested the hypothesis that �-3 PUFAs activate cGMP/PKG signaling and block profibrotic TGF-�1 signaling. Incultured cardiac fibroblasts, both EPA and DHA (10 to50 �mol/L) induced dose-dependent increases in cGMPproduction (all P�0.01), but neither control fatty acid,arachidonic acid or oleic acid (10 or 20 �mol/L), was able toinduce cGMP production (Figure 6A). Furthermore, EPA(10 �mol/L), DHA (10 �mol/L), and 8-bromo-cGMP(1 mmol/L), which activates PKG, completely blocked theTGF-�1–induced collagen synthesis (Figure 6B). Con-versely, a peptide-based inhibitor of PKG, DT-3 (1 �mol/L),significantly diminished the antifibrotic effect of EPA, DHA,and cGMP (Figure 6B). In summary, these results indicatethat the inhibition of profibrotic TGF-�1 signaling by �-3
Figure 4. Fish oil does not block pressure overload–inducedTGF-�1 production or phosphorylation of Smad2. A, Ventricularlevels of the active form of TGF-�1 measured by ELISA 3 daysafter TAC surgery (n�5 to 6 mice per group). B, Levels ofphospho-Smad2 and total Smad2 measured by Western blot 3days and 4 weeks after TAC. C, Densitometric quantification ofSmad2 phosphorylation 3 days and 4 weeks after TAC (n�6 to8 mice per group). Data are presented as mean�SEM. Datawere analyzed by 2-way ANOVA with the Tukey posthoc test.
588 Circulation February 15, 2011
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from
PUFAs is mediated by �-3 PUFA–induced cGMP/PKGsignaling.
Smad2 and Smad3 phosphorylation and subsequent trans-location to the nucleus are required for TGF-�1 signaling.Because EPA and DHA demonstrated an inhibitory effect onTGF-�1–induced fibrosis (Figure 5B through 5E), we testedthe effects of EPA and DHA on the TGF-�1–inducedphosphorylation of Smad2 and Smad3. In cultured cardiacfibroblasts, TGF-�1 treatment for 30 minutes induced robustphosphorylation of Smad2 (2.7-fold; P�0.01) and Smad3(2.2-fold; P�0.01). Longer exposure (24 hours) to TGF-�1induced phosphorylation of Smad2 but not Smad3 (Figure 6Cthrough 6E). However, EPA and DHA (10 �mol/L) failed toblock TGF-�1–induced phosphorylation of Smad2 andSmad3 (Figure 6C through 6E) at either 30 minutes or 24hours, which was consistent with our finding that fish oil didnot inhibit TAC-induced phosphorylation of Smad2 in vivo(Figure 4B and 4C). Thus, the inhibitory effect of EPA andDHA on TGF-�1–induced cardiac fibrosis is not due toinhibition of Smad phosphorylation.
EPA and DHA Block Nuclear Translocation ofPhospho-Smad2 and -Smad3 Through thecGMP/PKG PathwayBecause TGF-�1–induced phosphorylation of Smads was notblocked by DHA and EPA, we analyzed the ability of EPA
and DHA to block TGF-�1–induced nuclear translocation ofphospho-Smad2 and -Smad3. In cultured cardiac fibroblasts,TGF-�1 increased the nuclear localization of phospho-Smad2(66%; P�0.01) and phospho-Smad3 (83%; P�0.01; Figure7). TGF-�1–induced nuclear translocation of both Smads wassignificantly blocked by EPA and DHA (phospho-Smad2,reduced to 15% with DHA and to 13% with EPA; phospho-Smad3, reduced to 14% with DHA and to 13% with EPA;Figure 7). Furthermore, 8-bromo-cGMP (1 mmol/L) blockedTGF-�1–induced translocation of both phospho-Smad2 and-Smad3, whereas DT-3 reversed the effects of EPA and DHAon nuclear translocation of Smads in cardiac fibroblaststreated with TGF-�1 (Figure 7). Therefore, EPA and DHAinhibition of TGF-�1–induced fibrosis is mediated by block-age of phospho-Smad nuclear translocation.
EPA and DHA Increased Nitrite/NitrateProduction and Phosphorylated Endothelial NitricOxide Synthase and Endothelial Nitric OxideExpression in Cardiac FibroblastsNitric oxide (NO) increases cGMP production through acti-vation of soluble guanylyl cyclase,21,22 and �-3 PUFAsincrease NO synthesis in humans.23 Therefore, we tested thehypothesis that EPA and DHA increase NO production incardiac fibroblasts. Because NO is rapidly metabolized, wemeasured the total intracellular level of nitrite (NO2
�) and
Figure 5. EPA and DHA inhibit the TGF-�1–stimulated fibrotic response in isolatedadult mouse cardiac fibroblasts (CF). A,The �-3 content measured in culturedcardiac fibroblasts treated for 48 hourswith EPA and DHA (10 �mol/L), arachi-donic acid (AA; 10 �mol/L), and oleic acid(OA; 10 �mol/L) and treated for the final24 hours with TGF-�1 (1 ng/mL) (n�3). B,Cardiac fibroblast proliferation measuredby counting live cells with Trypan Blueexclusion in cardiac fibroblasts treated asin A (n�3 to 4). C, Collagen synthesisdetermined by [3H]-proline incorporationin cardiac fibroblasts treated as in A (n�9to 42). D, Myofibroblast transformationmeasured by �-SMA staining with coun-terstains for fibroblast-specific protein 1(FSP1; red) and nuclei (DAPI; blue) in car-diac fibroblasts treated as in A. E, Quanti-fication of �-SMA–positive cells as a per-centage of total FSP1-positive cells (4slides for each group). Data are presentedas mean�SEM. Data were analyzed by1-way ANOVA with the Dunnett posthoctest in A through C and the Tukeyposthoc test in E.
Chen et al �-3 Fatty Acids Prevent Cardiac Fibrosis 589
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from

nitrate (NO3�), stable NO metabolites, as an index of total
NO production. Twenty-four hours of treatment with EPAand DHA (10 �mol/L) significantly increased nitrite andnitrate levels in cardiac fibroblasts (70% in EPA and 65% inDHA). We also examined whether NO production wasinduced with an increase in intracellular levels of cGMP. Asexpected, the cGMP analog 8-bromo-cGMP (0.1 and1 mmol/L) did not induce NO production in cardiac fibro-blasts (Figure 8A). This demonstrates that the increase of NOis not a result of increased intracellular cGMP.
To determine whether EPA and DHA enhance expressionof phosphorylated endothelial nitric oxide synthase (eNOS)and eNOS, we measured phospho-eNOS and eNOS proteinlevels in cardiac fibroblasts. Treatment with 10 �mol/L DHAor EPA significantly increased phospho-eNOS and eNOSprotein levels in cardiac fibroblasts (Figure 8B through 8D).These results demonstrate that EPA and DHA increasephospho-eNOS and eNOS expression, thereby promoting NOproduction in cardiac fibroblasts.
To examine other possible mechanisms involved in cGMPproduction and degradation,21,22 we tested the effects of EPAand DHA on particulate guanylyl cyclase activity, cGMP-specific phosphodiesterases activity, and messenger RNA(mRNA) levels of ANP and BNP in cardiac fibroblasts. Wefound that EPA (10 �mol/L) and DHA (10 �mol/L) didnot increase particulate guanylyl cyclase activity and did not
inhibit cGMP-specific phosphodiesterases activity (data notshown). We also found that EPA (10 �mol/L) and DHA(10 �mol/L) significantly decreased ANP mRNA and did notchange the BNP mRNA in cardiac fibroblasts (data notshown).
DiscussionIn the present study, we investigated the effects of �-3PUFAs on the cardiac fibrotic response. Fischer et al24
reported that a diet supplemented with �-3 PUFAs inhibitsleft ventricle perivascular fibrosis in rats carrying humanrennin and angiotensinogen transgenes, and Medeiros et al25
reported that �-3 PUFAs and palm oil (which is devoid of�-3 PUFAs) reduce cardiac fibrosis in diabetic spontaneouslyhypertensive rats. However, in the above hypertension mod-els, �-3 PUFAs significantly decreased the systemic arterialblood pressure. Therefore, from the above studies, we cannotconclude that the antifibrotic effect of �-3 PUFAs is inde-pendent of blood pressure. At the same time, both studiescompared the group fed an �-3 PUFA–supplemented diet anda control group fed a diet containing less total fatty acids,which makes the results more difficult to interpret. In thepresent study, which used an isocaloric control diet contain-ing equal levels of total fatty acids, �-3 PUFAs decreasedpressure overload–induced cardiac fibrosis by 63% and con-sequently preserved cardiac function without changing the
Figure 6. EPA and DHA inhibit the TGF-�1–stimulated fibrotic response throughthe cGMP/PKG pathway. A, cGMP levelsmeasured by ELISA in cultured cardiacfibroblasts treated for 48 hours with EPAand DHA (10 to 50 �mol/L) with arachi-donic acid (AA; 10 to 20 �mol/L) andoleic acid (OA; 10 to 20 �mol/L) (n�3 to7) as control. B, Collagen synthesis deter-mined by [3H]-proline incorporation in car-diac fibroblasts treated for 48 hours withEPA (10 �mol/L), DHA (10 �mol/L),8-bromo-cGMP (1 mmol/L), and/or theguanylyl cyclase inhibitor DT-3 (1 �mol/L)and for the final 24 hours with TGF-�1 (1ng/mL) (n�4 to 20). C, Smad2 andSmad3 phosphorylation (Smad2, Ser465/467; Smad3, Ser423/425) detected byWestern blot in cultured cardiac fibro-blasts treated for 24 hours with EPA orDHA (10 �mol/L) and for an additional 30minutes or 24 hours with fatty acids andTGF-�1 (1 ng/mL). D and E, Quantifica-tion of Smad2 or Smad3 phosphorylationrelative to total Smad2/3 (n�3). Data arepresented as mean�SEM. Means werecompared by paired Student t test in B,1-way ANOVA with the Dunnett posthoctest in A, and the Tukey posthoc test in Dand E.
590 Circulation February 15, 2011
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from

blood pressure or pressure gradients (Table III in the online-only Data Supplement). Therefore, our results demonstrate anantifibrotic effect of �-3 PUFAs that is independent ofafterload.
We also identified a potential in vivo mechanism for thisantifibrotic effect. Here, we found that in the heart, aorticconstriction significantly increased the levels of activeTGF-�1 protein, phosphorylation of Smad2, transformationof cardiac fibroblasts, nonmyocyte proliferation, and thelevels of collagen I and III mRNA. We also found that �-3PUFAs prevented the increase in transformation of cardiacfibroblasts, nonmyocyte proliferation, and collagen I and IIImRNA without altering the levels of active TGF-�1 proteinand phosphorylation of Smad2. Therefore, our findings indi-cate that �-3 PUFAs inhibit pressure overload–induced car-diac fibrosis by preventing cardiac fibroblast transformation,proliferation, and consequently collagen I and III geneexpression. Our findings also indicate that �-3 PUFAs do notinhibit TGF-�1 fibrotic signaling at the level of the TGF-�1ligand, TGF-�1 receptors, or phosphorylation of Smad2.
Using cultured adult mouse cardiac fibroblasts, we furtheridentified the antifibrotic signaling of �-3 PUFAs and itsinteraction with the fibrotic signaling of TGF-�1. In our invitro study, we saw increased �-3 content and cGMP levels inDHA- and EPA-treated cardiac fibroblasts. These findings
indicate that EPA and DHA can be incorporated into cardiacfibroblasts and activate cGMP signaling in cardiac fibro-blasts. We also observed that EPA and DHA preventedTGF-�1–induced proliferation, transformation, and collagensynthesis in cardiac fibroblasts and TGF-�1–induced Smad-responsive promoter activity (Figure I in the online-only DataSupplement). We also found that DT3, a highly selectivePKG inhibitor, blocked the inhibitory effects of EPA andDHA on TGF-�1–induced cardiac fibroblast proliferation,transformation, and collagen synthesis in cardiac fibroblasts.These findings suggest that EPA and DHA act through thecGMP/PKG pathway to achieve their antifibrotic effect incardiac fibroblasts.
cGMP has an inhibitory effect on the TGF-�1–inducedfibrotic response.9,10,26 Li10 reported that disruption of nucleartranslocation of Smad3 accounts for the antifibrotic effect ofcGMP/PKG signaling against the TGF-�1 signaling pathway.In our observations, EPA and DHA increased cGMP levels incardiac fibroblasts in a dose-dependent manner. EPA, DHA,and 8-bromo-cGMP prevented TGF-�1–induced nucleartranslocation of phospho-Smad2 and phospho-Smad3 withoutaffecting Smad2 and Smad3 phosphorylation in cardiacfibroblasts. In addition, DT3 reversed the inhibitory effects of8-Br-cGMP, EPA, and DHA on the TGF-�1–induced nucleartranslocation of phospho-Smad2 and phospho-Smad3. These
Figure 7. EPA and DHA block nucleartranslocation of phosphorylated Smad2and Smad3 through the cGMP/PKG path-way. A, Phospho-Smad localizationdetected by staining for phospho-Smad2(green; first row), Smad4 (red; secondrow), or phospho-Smad3 (green; fourthrow) with a nuclear counterstain (DAPI;merged images; third and fifth rows) incultured cardiac fibroblasts treated for 24hours with EPA (10 �mol/L) (images notshown), DHA (10 �mol/L), 8-bromo-cGMP(1 mmol/L), and/or the guanylyl cyclaseinhibitor DT-3 (1 �mol/L) and for an addi-tional 30 minutes with fatty acids andTGF-�1 (1 ng/mL). B and C, Quantifica-tion of the percent of nuclei positive forphospho-Smad2 and -Smad3 (counting50 to 200 cells per field; n�5 to 33 in B,and n�4 to 31 in C). Data are presentedas mean�SEM. Data were analyzed by1-way ANOVA with the Tukey posthoctest.
Chen et al �-3 Fatty Acids Prevent Cardiac Fibrosis 591
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from

findings indicate that in cardiac fibroblasts, EPA and DHAblock TGF-�1–induced nuclear translocation of phospho-Smad2 and phospho-Smad3 through activation of the cGMP/PKG pathway (Figure II in the online-only Data Supplement).
In the cell, NO-sensitive soluble guanylyl cyclase andnatriuretic peptide–sensitive particulate guanylyl cyclase pro-duce cGMP, whereas cGMP-specific phosphodiesterases hy-drolyze cGMP.21,22 DHA increases NO synthesis andphospho-eNOS and eNOS expression levels.27 NO promotescGMP production through activation of NO-sensitive solubleguanylyl cyclase.21,22 In this study, we found that EPA andDHA significantly increased the expression levels ofphospho-eNOS and eNOS and increased nitrite/nitrate levelsin cardiac fibroblasts. We also found that EPA and DHA didnot increase particulate guanylyl cyclase activity or ANP/BNP production. Additionally, EPA and DHA did not inhibitcGMP-specific phosphodiesterase activity. Therefore, wepostulate that EPA and DHA increase intracellular levels ofcGMP by increasing phospho-eNOS and eNOS protein levelsand NO production in cardiac fibroblasts.
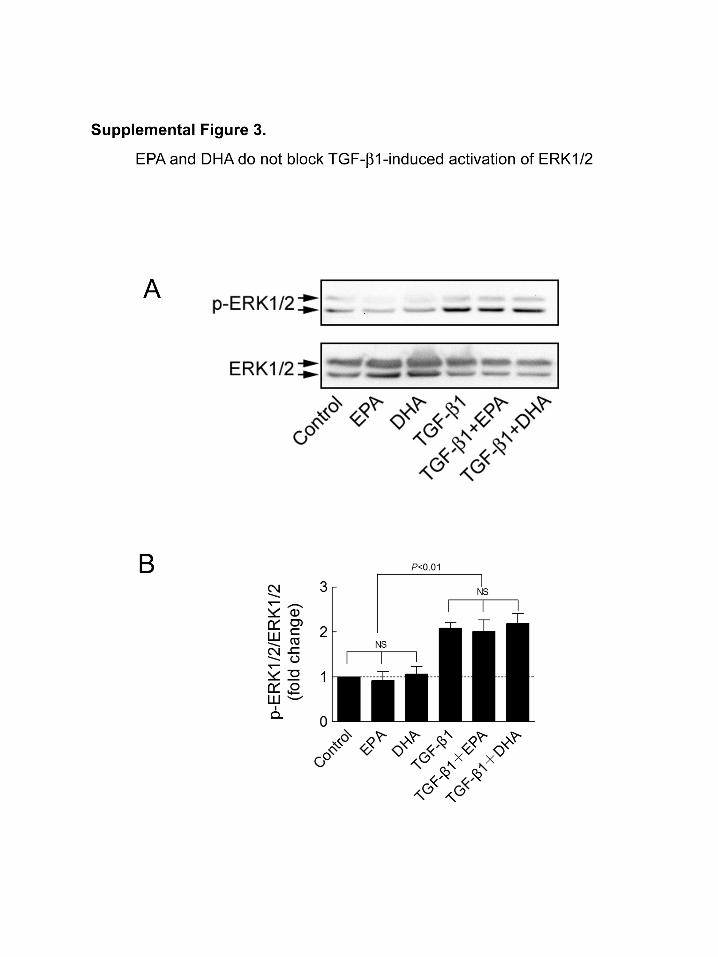
TGF-�1–induced phosphorylation of ERK1/2 is alsoinvolved in the TGF-�1–induced fibrotic response in fibro-blasts.28 To determine whether EPA and DHA affect TGF-
�1–induced phosphorylation of ERK1/2, we tested phospho-ERK levels in cardiac fibroblasts after 1 hour of treatment ofTGF-�1 with or without pretreatment of EPA and DHA.TGF-�1 (1 ng/mL) induced significant phosphorylation ofERK1/2 in cardiac fibroblasts, whereas EPA and DHA didnot block this effect (Figure III in the online-only DataSupplement).
In summary, we discovered an antifibrotic effect of EPAand DHA and established that EPA and DHA exert theirantifibrotic effect through activation of the cGMP/PKGpathway. We extended the current understanding of themechanism of the antifibrotic effects of cGMP by demon-strating that activation of PKG blocks TGF-�1–inducednuclear translocation of phospho-Smad2 in cardiac fibro-blasts. Our results also suggest that �-3 PUFAs increasecGMP levels in cardiac fibroblasts by increasing phospho-eNOS and eNOS protein levels and NO production.
About 50% of heart failure cases are due to diastolicdysfunction.2 One of the main causes of diastolic heart failureis cardiac fibrosis, and at present, no therapies are available toprevent or treat cardiac fibrosis.4,8,29 Given the clinicallyproven safety and tolerance of oily fish, fish oil, and theDHA�EPA preparations, a new clinical study of the thera-peutic effect of �-3 PUFAs on cardiac fibrosis and diastolicheart failure is warranted.
Study LimitationsThis study examined the preventive effects of �-3 PUFAs ona pressure-overloaded animal model that mimics only someaspects of the disease as it occurs in human. In this animalmodel, pressure overload induced an acute cardiac injury witha rapid progression that is more severe compared with humanhypertension conditions. Another limitation of this study isthat even though we fed �-3 PUFAs at a reasonable level (1%energy), the red blood cell levels achieved were much highercompared with humans taking 2 g/d EPA and DHA.30
Therefore, our findings should be explored with lower �-3PUFA doses.
AcknowledgmentsWe thank the Physiology and Cell Culture Cores at SanfordResearch/University of South Dakota for technical support andJames V. Pottala for his assistance with the longitudinal analysis.
Sources of FundingThis study was supported by National Institutes of Health NationalCenter for Research Resources grant 2P20RR017662–06A1 and bythe South Dakota 2010 Initiative Research Centers Program.
DisclosuresDrs Harris and Shearer have relationships with companies that sell�-3 products, are on the speaker’s bureau for GlaxoSmithKline, andhave received grants from GlaxoSmithKline. Dr Harris is a scientificadvisor to GlaxoSmithKline and is a consultant for and has receivedresearch grants from Monsanto. In addition, Dr Harris is the ownerof a company that offers blood �-3 tests (OmegaQuant Analytics).The other authors report no conflicts.
References1. Braunwald E, Bristow MR. Congestive heart failure: fifty years of
progress. Circulation. 2000;102(suppl):IV-14–IV-23.2. Gary R, Davis L. Diastolic heart failure. Heart Lung. 2008;37:405–416.
Figure 8. EPA and DHA increase nitrite/nitrate (NOx) productionand phospho-eNOS and eNOS expression in cardiac fibro-blasts. A, The levels of NOx in the cell were determined with theTotal Nitrate/Nitrite Fluorometric method in cultured cardiacfibroblasts treated for 24 hours with EPA (10 �mol/L), DHA(10 �mol/L), 8-bromo-cGMP (0.1 mmol/L), or 8-bromo-cGMP(1 mmol/L) (n�4 to 14). B, Phospho-eNOS and eNOS detectedby Western blot in cultured cardiac fibroblasts treated for 24hours with EPA or DHA (10 �mol/L). C and D, Quantification ofphospho-eNOS or eNOS relative to GAPDH (n�7 to 9). Data arepresented as mean�SEM. Data were analyzed by 1-wayANOVA with the Tukey posthoc test.
592 Circulation February 15, 2011
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from

3. Berk BC, Fujiwara K, Lehoux S. ECM remodeling in hypertensive heartdisease. J Clin Invest. 2007;117:568–575.
4. Rossi MA. Pathologic fibrosis and connective tissue matrix in left ven-tricular hypertrophy due to chronic arterial hypertension in humans.J Hypertens. 1998;16:1031–1041.
5. Brown RD, Ambler SK, Mitchell MD, Long CS. The cardiac fibroblast:therapeutic target in myocardial remodeling and failure. Annu RevPharmacol Toxicol. 2005;45:657–687.
6. Kai H, Kuwahara F, Tokuda K, Imaizumi T. Diastolic dysfunction inhypertensive hearts: roles of perivascular inflammation and reactive myo-cardial fibrosis. Hypertens Res. 2005;28:483–490.
7. Kuwahara F, Kai H, Tokuda K, Kai M, Takeshita A, Egashira K,Imaizumi T. Transforming growth factor-beta function blocking preventsmyocardial fibrosis and diastolic dysfunction in pressure-overloaded rats.Circulation. 2002;106:130–135.
8. Leask A. TGFbeta, cardiac fibroblasts, and the fibrotic response. Car-diovasc Res. 2007;74:207–212.
9. Buxton IL, Duan D. Cyclic GMP/protein kinase G phosphorylation ofSmad3 blocks transforming growth factor-beta-induced nuclear Smadtranslocation: a key antifibrogenic mechanism of atrial natriuretic peptide.Circ Res. 2008;102:151–153.
10. Li P, Wang D, Lucas J, Oparil S, Xing D, Cao X, Novak L, Renfrow MB,Chen YF. Atrial natriuretic peptide inhibits transforming growth factorbeta-induced Smad signaling and myofibroblast transformation in mousecardiac fibroblasts. Circ Res. 2008;102:185–192.
11. Levitan EB, Wolk A, Mittleman MA. Fish consumption, marine omega-3fatty acids, and incidence of heart failure: a population-based prospectivestudy of middle-aged and elderly men. Eur Heart J. 2009;30:1495–1500.
12. Mozaffarian D, Bryson CL, Lemaitre RN, Burke GL, Siscovick DS. Fishintake and risk of incident heart failure. J Am Coll Cardiol. 2005;45:2015–2021.
13. Yamagishi K, Iso H, Date C, Fukui M, Wakai K, Kikuchi S, Inaba Y,Tanabe N, Tamakoshi A. Fish, omega-3 polyunsaturated fatty acids, andmortality from cardiovascular diseases in a nationwide community-basedcohort of Japanese men and women the JACC (Japan CollaborativeCohort Study for Evaluation of Cancer Risk) study. J Am Coll Cardiol.2008;52:988–996.
14. Tavazzi L, Maggioni AP, Marchioli R, Barlera S, Franzosi MG, Latini R,Lucci D, Nicolosi GL, Porcu M, Tognoni G. Effect of n-3 polyunsat-urated fatty acids in patients with chronic heart failure (the GISSI-HFtrial): a randomised, double-blind, placebo-controlled trial. Lancet. 2008;372:1223–1230.
15. Picq M, Dubois M, Grynberg A, Lagarde M, Prigent AF. Specific effectsof n-3 fatty acids and 8-bromo-cGMP on the cyclic nucleotide phospho-diesterase activity in neonatal rat cardiac myocytes. J Mol Cell Cardiol.1996;28:2151–2161.
16. O’Connell TD, Ishizaka S, Nakamura A, Swigart PM, Rodrigo MC,Simpson GL, Cotecchia S, Rokosh DG, Grossman W, Foster E, SimpsonPC. The alpha(1A/C)- and alpha(1B)-adrenergic receptors are requiredfor physiological cardiac hypertrophy in the double-knockout mouse.J Clin Invest. 2003;111:1783–1791.
17. O’Connell TD, Swigart PM, Rodrigo MC, Ishizaka S, Joho S, Turnbull L,Tecott LH, Baker AJ, Foster E, Grossman W, Simpson PC. Alpha1-
adrenergic receptors prevent a maladaptive cardiac response to pressureoverload. J Clin Invest. 2006;116:1005–1015.
18. Tang YD, Kuzman JA, Said S, Anderson BE, Wang X, Gerdes AM. Lowthyroid function leads to cardiac atrophy with chamber dilatation,impaired myocardial blood flow, loss of arterioles, and severe systolicdysfunction. Circulation. 2005;112:3122–3130.
19. Duda MK, O’Shea KM, Tintinu A, Xu W, Khairallah RJ, Barrows BR,Chess DJ, Azimzadeh AM, Harris WS, Sharov VG, Sabbah HN, StanleyWC. Fish oil, but not flaxseed oil, decreases inflammation and preventspressure overload-induced cardiac dysfunction. Cardiovasc Res. 2009;81:319–327.
20. Duda MK, O’Shea KM, Lei B, Barrows BR, Azimzadeh AM, McElfreshTE, Hoit BD, Kop WJ, Stanley WC. Dietary supplementation withomega-3 PUFA increases adiponectin and attenuates ventricularremodeling and dysfunction with pressure overload. Cardiovasc Res.2007;76:303–310.
21. Ritchie RH, Irvine JC, Rosenkranz AC, Patel R, Wendt IR, Horowitz JD,Kemp-Harper BK. Exploiting cGMP-based therapies for the preventionof left ventricular hypertrophy: NO* and beyond. Pharmacol Ther. 2009;124:279–300.
22. Tsai EJ, Kass DA. Cyclic GMP signaling in cardiovascular pathophys-iology and therapeutics. Pharmacol Ther. 2009;122:216–238.
23. Harris WS, Rambjor GS, Windsor SL, Diederich D. N-3 fatty acids andurinary excretion of nitric oxide metabolites in humans. Am J Clin Nutr.1997;65:459–464.
24. Fischer R, Dechend R, Qadri F, Markovic M, Feldt S, Herse F, Park JK,Gapelyuk A, Schwarz I, Zacharzowsky UB, Plehm R, Safak E, Heuser A,Schirdewan A, Luft FC, Schunck WH, Muller DN. Dietary n-3 polyun-saturated fatty acids and direct renin inhibition improve electricalremodeling in a model of high human renin hypertension. Hypertension.2008;51:540–546.
25. Medeiros FJ, Mothe CG, Aguila MB, Mandarim-de-Lacerda CA.Long-term intake of edible oils benefits blood pressure and myocardialstructure in spontaneously hypertensive rat (SHR) and streptozotocindiabetic shr. Prostaglandins Other Lipid Mediat. 2005;78:231–248.
26. Takimoto E, Champion HC, Li M, Belardi D, Ren S, Rodriguez ER,Bedja D, Gabrielson KL, Wang Y, Kass DA. Chronic inhibition of cyclicGMP phosphodiesterase 5A prevents and reverses cardiac hypertrophy.Nat Med. 2005;11:214–222.
27. Stebbins CL, Stice JP, Hart CM, Mbai FN, Knowlton AA. Effects ofdietary docosahexaenoic acid (DHA) on eNOS in human coronary arteryendothelial cells. J Cardiovasc Pharmacol Ther. 2008;13:261–268.
28. Liu X, Sun SQ, Hassid A, Ostrom RS. Camp inhibits transforming growthfactor-beta-stimulated collagen synthesis via inhibition of extracellularsignal-regulated kinase 1/2 and Smad signaling in cardiac fibroblasts. MolPharmacol. 2006;70:1992–2003.
29. Beltrami CA, Finato N, Rocco M, Feruglio GA, Puricelli C, Cigola E,Quaini F, Sonnenblick EH, Olivetti G, Anversa P. Structural basis ofend-stage failure in ischemic cardiomyopathy in humans. Circulation.1994;89:151–163.
30. Harris WS, Von Schacky C. The omega-3 index: a new risk factor fordeath from coronary heart disease? Prev Med. 2004;39:212–220.
CLINICAL PERSPECTIVEHeart failure is the leading reason for hospital admissions and is the most expensive Medicare expenditure. About half ofheart failure cases are due to diastolic dysfunction. One of the main causes of diastolic dysfunction is cardiac fibrosis, andno therapies are available to prevent or treat cardiac fibrosis. Transforming growth factor-�1–induced cardiac fibroblasttransformation and proliferation are the key events leading to cardiac fibrosis. This study shows that �-3 polyunsaturatedfatty acids prevent pressure overload–induced cardiac fibrosis and subsequent cardiac dysfunction. This study alsodemonstrates that in cardiac fibroblasts, eicosapentaenoic acid and docosahexaenoic acid increase cyclic GMP levels byincreasing phosphorylated endothelial nitric oxide synthase and endothelial nitric oxide synthase protein levels and nitricoxide production, and they exert their antifibrotic effect through activation of the cyclic GMP/protein kinase G pathwayand subsequent blocking of transforming growth factor-�1–induced nuclear translocation of phospho-Smad2 andphospho-Smad3. This study defines the beneficial effects of �-3 polyunsaturated fatty acids on cardiac fibrosis and cardiacdysfunction and clarifies the underlying mechanisms. In addition, this study provides the basis for extending the applicationof �-3 polyunsaturated fatty acids, which appear to be exceptionally safe and well tolerated, to the prevention of cardiacfibrosis.
Chen et al �-3 Fatty Acids Prevent Cardiac Fibrosis 593
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from

Nareddy, A. Martin Gerdes, William S. Harris, Timothy D. O'Connell and Dajun WangJinghai Chen, Gregory C. Shearer, Quanhai Chen, Chastity L. Healy, April J. Beyer, Vijaya B.
Activation of Cyclic GMP/Protein Kinase G Signaling in Cardiac FibroblastsInduced Cardiac Fibrosis Through−Omega-3 Fatty Acids Prevent Pressure Overload
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2011 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/CIRCULATIONAHA.110.971853
2011;123:584-593; originally published online January 31, 2011;Circulation.
http://circ.ahajournals.org/content/123/6/584World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circ.ahajournals.org/content/suppl/2011/02/03/CIRCULATIONAHA.110.971853.DC1Data Supplement (unedited) at:
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialCirculationin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on August 31, 2017
http://circ.ahajournals.org/D
ownloaded from

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
SUPPLEMENTAL MATERIAL
ω-3 fatty acids prevent pressure overload-induced cardiac fibrosis through activation
of cGMP/PKG signaling in cardiac fibroblasts
by Jinghai Chen, PhD; Gregory C. Shearer, PhD; Quanhai Chen, MD; Chastity L. Healy, BS; April J.
Beyer, MS; Vijaya B. Nareddy, MS; A. Martin Gerdes, PhD; William S. Harris, PhD; Timothy D.
O’Connell, PhD; Dajun Wang, MD
Supplemental Abbreviations
Commonly used abbreviations
PUFAs Polyunsaturated fatty acids
DHA Docosahexaenoic acid
EPA Eicosapentaenoic acid
TGF-β1 Transforming growth factor beta1
TAC Transverse aortic constriction
cGMP Cyclic guanosine monophosphate
PKG cGMP-dependent protein kinase G
α-SMA alpha-smooth muscle actin
1

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
Supplemental Methods
Animals
The use of animals in this study conformed to the Public Health Service Guide for
Care and Use of Laboratory Animals and was approved by Sanford Research/USD
Institutional Animal Care and Use Committee.
Male C57BL/6 mice were obtained from Jackson Laboratory at 5 weeks of age. All
mice were fed a control or a fish oil supplemented diet for eight weeks (Dyets;
Bethlehem, PA). Both diets contained 4% by weight test oil (control diet: 40 g corn oil
per kg; fish oil supplemented diet: 12 g menhaden oil plus 28 g corn oil per kg), but
were otherwise identical. The fish oil diet provided 1% energy as EPA+DHA, similar to
the amounts that have been shown to be protective in human studies (i.e., about 2
g/d).1 After 8 weeks on the assigned diet, mice were randomly allocated into 4 groups
using a 2x2 factorial design and subjected to transverse aortic constriction (TAC)
(groups: control/sham, control/TAC, fish oil/sham, fish oil/TAC; n=22-25/group).
Following surgery, mice were continually fed the assigned diets and sacrificed after 3,
7, or 28 days.
Transverse Aortic Constriction
Transverse aortic constriction (TAC) surgery was performed without intubation
under anesthesia with isoflurane, as previously described.2-3
Measurement of the Omega-3 Content
The omega-3 content is expressed as the percentage of EPA and DHA of total fatty
acids. Omega-3 content in red blood cells and cardiac tissues was measured as
previously described.4 A similar analysis was performed on cultured cardiac
fibroblasts.
2

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
Measurement of Cardiac Function
Echocardiography and Hemodynamics were performed as described previously.5
Echocardiographic measurements were performed under anesthesia (3%
isoflurane induction, 1% maintenance) using a Visual Sonics Vevo 660
High-Resolution Imaging System (Visual Sonics, Toronto, Canada) with a 30-MHz
linear array transducer (model RMV-707). Left ventricular dimensions and heart rate
were measured from 2-D short-axis M-mode tracings at the level of the papillary
muscle. Left ventricular mass and functional parameters were calculated using the
above primary measurements.
Left ventricular hemodynamics were measured under anesthesia (3% isoflurane
induction, 1% maintenance) using a Millar catheter inserted into the left ventricle via
the carotid artery. After stabilization, heart rate (HR) as well as left ventricular
end-systolic and end-diastolic pressure were measured from the left ventricular
pressure waveform, and ±dP/dt and tau were calculated.
Isolation and Culture of Cardiac Fibroblasts
Adult mouse cardiac fibroblasts were isolated using a previously described
procedure3 with the following modifications. Briefly, following perfusion and digestion
of the heart with collagenase II (Worthington Biochemical Corp, Lakewood, NJ),
dissociated cells (myocytes and non-myocytes) were sedimented by gravity, and the
supernatant, rich in cardiac fibroblasts, was collected and centrifuged for 5 min at
1000 rpm. Fibroblasts were resuspended in Dulbecco's Modification of Eagle's
Medium (DMEM; Fisher Scientific, Pittsburgh, PA) with 15% fetal bovine serum (FBS;
Fisher Scientific) and plated on laminin-coated 60 mm dishes (Becton Dickinson,
Franklin Lakes, NJ). After one hour, the culture medium was changed, which removed
weakly adherent cells, including any myocytes and endothelial cells. After 24 hours,
the culture medium was changed again, and the fibroblast cultures were grown to 85%
confluence, at which point the medium was replaced with DMEM containing 0.1% FBS.
After 24 hours in reduced serum medium, fibroblast cultures were treated with
3

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
fatty-acids or other agonist/antagonists as indicated.
Fatty Acids Preparation
Free fatty acids (DHA, EPA, arachidonic acid, oleic acid; Sigma) for in vitro studies
were dissolved in 100% ethanol to make a stock solution (100 mg/ml). Aliquots (5
μl/tube) of the stock solution were stored in PCR tubes under a blanket of nitrogen and
sealed with parafilm and stored at -80 ºC. Before use, the aliquots were diluted with
0.2% BSA/PBS and then further diluted in culture medium.
Measurement of Cell Proliferation in Isolated Adult Cardiac Fibroblasts
Cardiac fibroblasts (3X105 cells/dish) were cultured and treated with TGF-1 as
above. After 48 hours, fibroblasts were harvested by trypsinization and cell number
was determined by trypan blue exclusion.
Measurement of Collagen Synthesis in Isolated Adult Cardiac Fibroblasts
Collagen synthesis was measured by incorporation of 3H-proline. Cardiac
fibroblasts were cultured in 24-well plates (2.5X104 cells/well), and after 24 hours of
culture in reduced serum medium, fibroblasts were treated with TGF-1 (1ng/ml) to
induce collagen synthesis. After 8 hours, 3H-proline (1 μCi/ml, Perkin Elmer, Waltham,
MA) was added. After 40 hours, fibroblasts were rinsed three times with PBS and fixed
with ice-cold 10% TCA for 30 min. Cell precipitates were washed with
phosphate-buffered saline and solubilized in 0.2 N NaOH at room temperature for one
hour. Incorporation of 3H-proline was determined by liquid scintillation spectrometry
(Beckman LS 6500, Fullerton, CA).
Real-time Quantitative PCR for Gene Expression of LV after Surgery
Gene expression was measured by the Genomic-Microarray/qPCR Core at The
Burnham Institute for Medical Research (La Jolla, CA). Briefly, RNA from the left
ventricle (3 and 7 days post-surgery) was extracted using the RNeasy Fibrous Tissue
4

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
Mini Kit (QIAGEN, Valencia, CA). Oligo (dT) primed cDNA synthesis was performed
using Superscript III (Invitrogen, Carlsbad, CA). Expression of collagen I, collagen III,
atrial natriuretic peptide (ANP), brain natriuretic peptide (BNP), PAI-1 and
glyceraldehyde 3-phosphate dehydrogenase (GAPDH) transcripts were measured
using the TaqMan Gene Expression Assay kit using validated primers (Applied
Biosystems, Foster City, CA).
Measurement of TGF-β1 and Cyclic GMP
TGF-β1 levels were measured by ELISA (R&D Systems, Minneapolis, MN) as
directed by the product insert. Absorbance was measured at 450 nm on a ThermoMax
microplate reader (Molecular Devices) and data collected using SoftMax (Molecular
Devices) software for data analysis.
Cyclic GMP levels were quantified using the acetylation protocol for a competitive
Cyclic GMP EIA kit (Cayman Chemical, Ann Arbor, MI). All procedures were
performed as directed in the product insert with modifications as indicated. Briefly,
samples were diluted 1:1 with EIA buffer included in the kit. Absorbance was
measured at 405 nm on a SpectraMax Plus 384 microplate reader (Molecular Devices)
and data collected using SoftMax Pro 5.3 (Molecular Devices).
Measurement of Nitrite/Nitrate Concentrations in Adult Cardiac Fibroblasts
Confluent cultures of cardiac fibroblasts in 6-cm collagen-coated plates were
incubated in the presence or absence of the indicated DHA, EPA or 8-bromo-cGMP
for 24 hours. The levels of nitrite/nitrate (NOx) in the cell were determined using a
Total Nitrate/Nitrite Fluorometric Assay kit (Cayman Chemical, Ann Arbor, MI). All
procedures were performed as directed in the product insert with modifications as
indicated. Briefly, after indicated treatments, fibroblasts were rinsed three times with
PBS, collected and sonicated in the Assay buffer included in the kit. Samples were
centrifuged for 30 min at 10,000g at 4oC. The supernatant was filtered through a
10,000 Dalton micropore filter (Millipore Billerica, MA) prior to the assay to remove
5

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
high molecular weight proteins. A sample volume of 50 µl per well was used.
Fluorescence was measured with an excitation of 365 nm and emission of 430 nm
using a Synergy Mx microplate reader (BioTek, Winooski, VT) and Gen5 software
(BioTek).
Immunohistochemistry and Immunocytochemistry
Hearts were excised, cannulated through the aorta and perfused with buffer (1%
adenosine, 2% BDM, and 0.04% heparin in PBS). Hearts were fixed in formalin
overnight, embedded in paraffin and sectioned (5 m) by the Histology Core at The
Burnham Institute for Medical Research (La Jolla, CA). To measure fibrosis, sections
were stained with picro-sirius red. High resolution images were obtained through
Aperio Scanscope software (Burnham Institute, La Jolla, CA) and cardiac fibrosis was
quantified using Image J (NIH website).
α-SMA staining was performed on formalin-fixed, paraffin-embedded left
ventricular sections (5 μm).
After deparaffinization, rehydration, heat-induced epitope retrieval and blocking of
endogenous peroxidase activity, sections were incubated with antibodies to α-SMA
(1:100, Dako, Carpinteria, CA) and an HRP-labeled secondary antibody. Dako
EnVision+ System-HRP (DAB) was used to visualize positive α-SMA staining.
Sections were counterstained with hematoxylin to visualize nuclei. Images were
captured using an inverted microscope (Olympus IX71). The positive area of α-SMA
was quantified by Image-Pro plus (Media Cybernetics, Bethesda, MD).
Ki67 staining for proliferation was performed on formalin-fixed, paraffin-embedded
left ventricular sections (5 μm). After deparaffinization, re-hydratation and
heat-induced epitope retrieval, sections were incubated with antibodies to Ki67 (1:200,
Abcam, Cambridge, MA) and a goat anti-rabbit AlexaFluor 488 secondary
antibody(1:400, Invitrogen). Images were captured using confocal microscopy
(FV1000, Olympus). Quantitative data were obtained by measuring co-localization of
4',6-diamidino-2-phenylindole DAPI (nuclear staining) with Ki67 in the interstitial area
6

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
of the left ventricle.
Staining for fibroblast-specific protein-1 (FSP-1), SMA or Smad was performed
on cultured cardiac fibroblasts fixed with 4% paraformaldehyde and permeabilized
with ice-cold methanol. After blocking with 5% normal serum from the same species as
the secondary antibody, fibroblasts were incubated with primary antibody
[phospho-Smad3 (Ser423/425) (1:400, Cell Signaling Technology, Danvers, MA);
phospho-Smad2 (ser465/467) (1:500, Millipore, Billerica, MA); Smad4 (1:100, Santa
Cruz Biotechnology, Inc, Santa Cruz, CA); FSP1 (1:100, Abcam); and α-SMA (1:200,
Dako)] and fluorochrome-conjugated secondary antibodies. Fibroblasts were
counterstained with DAPI to visualize nuclei. Images were captured using confocal
microscopy (FV1000, Olympus).
Western Blot Analysis
Samples from the left ventricle and cultured adult cardiac fibroblasts were
homogenized and incubated in Tissue Extraction Reagent I (Invitrogen, Carlsbad, CA)
and Cell Extraction Buffer (Invitrogen) with protease inhibitors cocktail (Sigma, Saint
Louise, MO) and 1mM PMSF (phenylmethylsulfonyl Fluoride) on ice for 15 minutes.
The lysates were centrifuged at 13,000g for 10 minutes at 4°C. The supernatant was
collected, aliquoted, and stored at -80°C until time of use. Protein concentration was
determined by Bio-Rad Protein Assay (Bio-Rad Laboratories, Hercules, CA). Samples
were mixed with Laemmli buffer containing 5% β-mercaptoethanol and were evenly
loaded onto SDS-PAGE gels. Proteins were transferred to PVDF membranes.
Membranes were incubated overnight at 4°C with primary antibodies,
phospho-Smad2, phospho-Smad3, Smad2/3, phospho-eNOS, total eNOS,
phospho-ERK1/2, total ERK1/2 ( all from Cell Signaling Technology, Inc., Danvers,
MA) in 5% BSA. A horseradish peroxidase conjugated secondary antibody was
incubated for 1 h at room temperature in 5% milk and processed for chemiluminescent
detection using an ECL Advanced Western Blotting Kit (GE Healthcare, Piscataway,
NJ). Protein abundance on Western blots was quantified by densitometry with the
7

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
Quantity One program from Bio-Rad.
Luciferase Smad Reporter Assay
Passage one cardiac fibroblasts were seeded into a 96-well tissue culture plate (5
x 103 cells/well) and were transduced with commercially available lentivirus particles
(SABiosciences, Frederick, MD) expressing the firefly luciferase gene under the
control of a minimal (m) CMV promoter and tandem repeats of the SMAD
transcriptional response element (AGCCAGACA). After 24 hours of culture in reduced
serum medium, fibroblasts were pretreated with vehicle or fatty acids for 24 hours,
followed by TGF-β (1 ng/ml) treatment for 48 hours. Fibroblasts were rinsed with PBS
and lysed prior to measurement of luciferase activity using a Luciferase Assay System
(Promega, Madison, WI) and luminometer (Zylux, Huntsville, AL).
References
1. Harris WS. N-3 fatty acids and serum lipoproteins: Human studies. Am J Clin Nutr.
1997;65:1645S-1654S
2. O'Connell TD, Ishizaka S, Nakamura A, Swigart PM, Rodrigo MC, Simpson GL,
Cotecchia S, Rokosh DG, Grossman W, Foster E, Simpson PC. The alpha(1a/c)- and
alpha(1b)-adrenergic receptors are required for physiological cardiac hypertrophy in
the double-knockout mouse. J Clin Invest. 2003;111:1783-1791
3. O'Connell TD, Swigart PM, Rodrigo MC, Ishizaka S, Joho S, Turnbull L, Tecott LH,
Baker AJ, Foster E, Grossman W, Simpson PC. Alpha1-adrenergic receptors prevent
a maladaptive cardiac response to pressure overload. J Clin Invest.
2006;116:1005-1015
4. Duda MK, O'Shea KM, Tintinu A, Xu W, Khairallah RJ, Barrows BR, Chess DJ,
8

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
9
Azimzadeh AM, Harris WS, Sharov VG, Sabbah HN, Stanley WC. Fish oil, but not
flaxseed oil, decreases inflammation and prevents pressure overload-induced cardiac
dysfunction. Cardiovasc Res. 2009;81:319-327
5. Tang YD, Kuzman JA, Said S, Anderson BE, Wang X, Gerdes AM. Low thyroid
function leads to cardiac atrophy with chamber dilatation, impaired myocardial blood
flow, loss of arterioles, and severe systolic dysfunction. Circulation.
2005;112:3122-3130

Chen et al., ω-3 fatty acids prevent cardiac fibrosis
10
Legends for Supplemental Figures
Supplemental figure 1. EPA and DHA reduce the TGF-β1-induced
Smad-responsive promoter activity. (A) Left ventricular mRNA expression of PAI-1
analyzed by real-time PCR 3 days and 7 days after TAC. Data are presented as
mean ± SEM (n=5 per group). Data were analyzed by two-way ANOVA with Tukey’s
post-hoc test. (B) Cardiac fibroblasts were transfected with VSV-g pseudotyped
lentivirus particles expressing the firefly luciferase gene under the control of a CMV
promoter and tandem repeats of the Smad transcriptional response element (TRE).
TGF-β1 (1 ng/ml) increased the luciferase activity in transfected cardiac fibroblasts;
24hrs pretreatment of DHA and EPA reduced TGF-β1-induced increase of luciferase
activity. Data are presented as mean ± SEM (n=5-6 per group). Data were analyzed
by one-way ANOVA with Tukey’s post-hoc test.
Supplemental Figure 2. Proposed anti-fibrotic mechanism of ω-3 PUFAs in
cardiac fibroblast.
Supplemental Figure 3. EPA and DHA do not block TGF-b1-induced
activation of ERK1/2. (A) Phosphorylated ERK1/2 (Thr202/Tyr204) and total ERK1/2
were detected by Western blot in cultured cardiac fibroblasts treated for 24 hr with EPA
or DHA (10 µM) and for an additional 1hr of TGF-β1 (1 ng/ml) with fatty acids. (B)
Quantification of Phosphorylated ERK1/2 relative to total ERK1/2. Data are presented
as mean ± SEM (n=5 per group).Data were analyzed by one-way ANOVA with Tukey’s
post-hoc test.
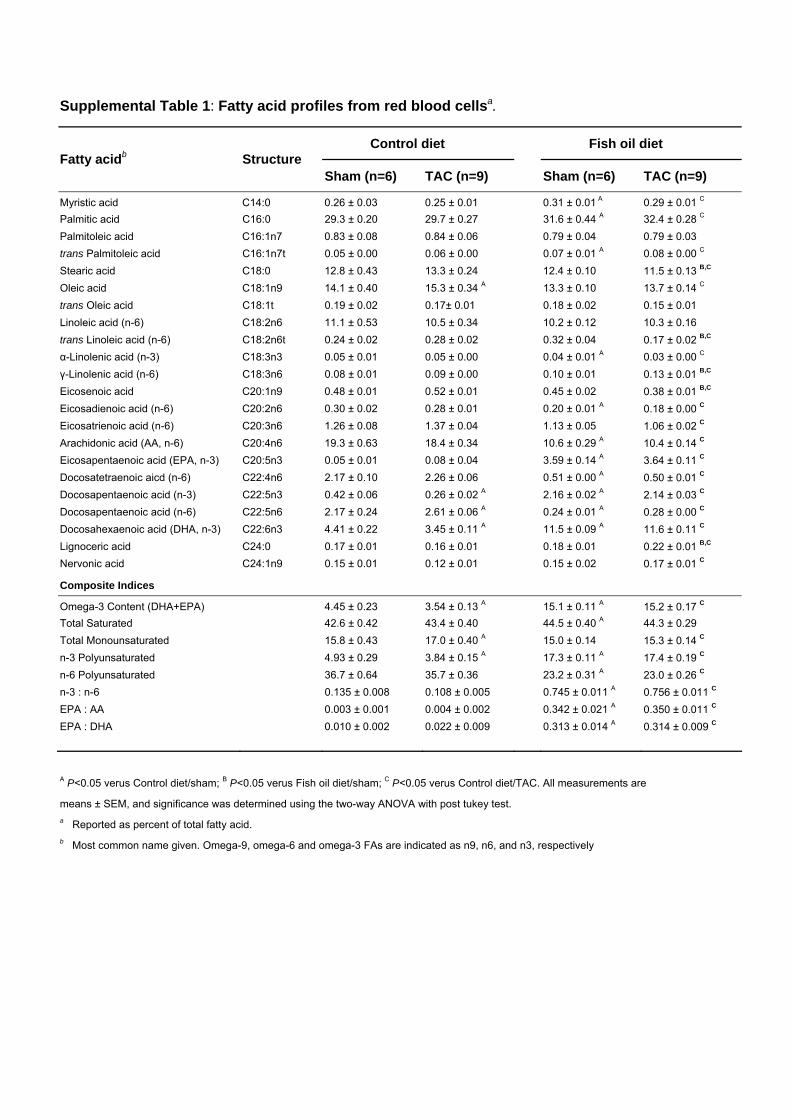
Supplemental Table 1: Fatty acid profiles from red blood cellsa.
A P<0.05 verus Control diet/sham; B P<0.05 verus Fish oil diet/sham; C P<0.05 verus Control diet/TAC. All measurements are
means ± SEM, and significance was determined using the two-way ANOVA with post tukey test. a Reported as percent of total fatty acid. b Most common name given. Omega-9, omega-6 and omega-3 FAs are indicated as n9, n6, and n3, respectively
Control diet Fish oil diet Fatty acidb Structure
Sham (n=6) TAC (n=9) Sham (n=6) TAC (n=9)
Myristic acid C14:0 0.26 ± 0.03 0.25 ± 0.01 0.31 ± 0.01 A 0.29 ± 0.01 C Palmitic acid C16:0 29.3 ± 0.20 29.7 ± 0.27 31.6 ± 0.44 A 32.4 ± 0.28 C Palmitoleic acid C16:1n7 0.83 ± 0.08 0.84 ± 0.06 0.79 ± 0.04 0.79 ± 0.03 trans Palmitoleic acid C16:1n7t 0.05 ± 0.00 0.06 ± 0.00 0.07 ± 0.01 A 0.08 ± 0.00 C Stearic acid C18:0 12.8 ± 0.43 13.3 ± 0.24 12.4 ± 0.10 11.5 ± 0.13 B,C Oleic acid C18:1n9 14.1 ± 0.40 15.3 ± 0.34 A 13.3 ± 0.10 13.7 ± 0.14 C trans Oleic acid C18:1t 0.19 ± 0.02 0.17± 0.01 0.18 ± 0.02 0.15 ± 0.01 Linoleic acid (n-6) C18:2n6 11.1 ± 0.53 10.5 ± 0.34 10.2 ± 0.12 10.3 ± 0.16 trans Linoleic acid (n-6) C18:2n6t 0.24 ± 0.02 0.28 ± 0.02 0.32 ± 0.04 0.17 ± 0.02 B,C α-Linolenic acid (n-3) C18:3n3 0.05 ± 0.01 0.05 ± 0.00 0.04 ± 0.01 A 0.03 ± 0.00 C γ-Linolenic acid (n-6) C18:3n6 0.08 ± 0.01 0.09 ± 0.00 0.10 ± 0.01 0.13 ± 0.01 B,C Eicosenoic acid C20:1n9 0.48 ± 0.01 0.52 ± 0.01 0.45 ± 0.02 0.38 ± 0.01 B,C Eicosadienoic acid (n-6) C20:2n6 0.30 ± 0.02 0.28 ± 0.01 0.20 ± 0.01 A 0.18 ± 0.00 C Eicosatrienoic acid (n-6) C20:3n6 1.26 ± 0.08 1.37 ± 0.04 1.13 ± 0.05 1.06 ± 0.02 C Arachidonic acid (AA, n-6) C20:4n6 19.3 ± 0.63 18.4 ± 0.34 10.6 ± 0.29 A 10.4 ± 0.14 C Eicosapentaenoic acid (EPA, n-3) C20:5n3 0.05 ± 0.01 0.08 ± 0.04 3.59 ± 0.14 A 3.64 ± 0.11 C Docosatetraenoic aicd (n-6) C22:4n6 2.17 ± 0.10 2.26 ± 0.06 0.51 ± 0.00 A 0.50 ± 0.01 C Docosapentaenoic acid (n-3) C22:5n3 0.42 ± 0.06 0.26 ± 0.02 A 2.16 ± 0.02 A 2.14 ± 0.03 C Docosapentaenoic acid (n-6) C22:5n6 2.17 ± 0.24 2.61 ± 0.06 A 0.24 ± 0.01 A 0.28 ± 0.00 C Docosahexaenoic acid (DHA, n-3) C22:6n3 4.41 ± 0.22 3.45 ± 0.11 A 11.5 ± 0.09 A 11.6 ± 0.11 C Lignoceric acid C24:0 0.17 ± 0.01 0.16 ± 0.01 0.18 ± 0.01 0.22 ± 0.01 B,C Nervonic acid C24:1n9 0.15 ± 0.01 0.12 ± 0.01 0.15 ± 0.02 0.17 ± 0.01 C
Composite Indices
Omega-3 Content (DHA+EPA) 4.45 ± 0.23 3.54 ± 0.13 A 15.1 ± 0.11 A 15.2 ± 0.17 C
Total Saturated 42.6 ± 0.42 43.4 ± 0.40 44.5 ± 0.40 A 44.3 ± 0.29 Total Monounsaturated 15.8 ± 0.43 17.0 ± 0.40 A 15.0 ± 0.14 15.3 ± 0.14 C n-3 Polyunsaturated 4.93 ± 0.29 3.84 ± 0.15 A 17.3 ± 0.11 A 17.4 ± 0.19 C n-6 Polyunsaturated 36.7 ± 0.64 35.7 ± 0.36 23.2 ± 0.31 A 23.0 ± 0.26 C n-3 : n-6 0.135 ± 0.008 0.108 ± 0.005 0.745 ± 0.011 A 0.756 ± 0.011 C EPA : AA 0.003 ± 0.001 0.004 ± 0.002 0.342 ± 0.021 A 0.350 ± 0.011 C EPA : DHA 0.010 ± 0.002 0.022 ± 0.009 0.313 ± 0.014 A 0.314 ± 0.009 C
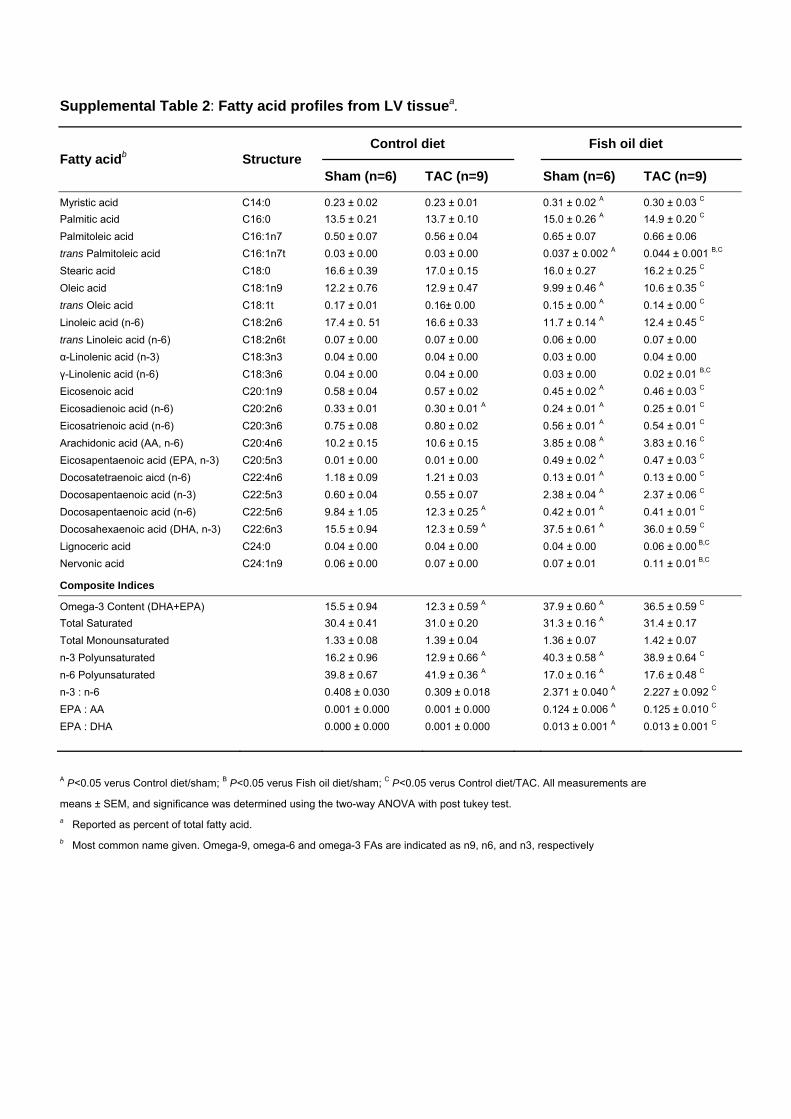
Supplemental Table 2: Fatty acid profiles from LV tissuea.
A P<0.05 verus Control diet/sham; B P<0.05 verus Fish oil diet/sham; C P<0.05 verus Control diet/TAC. All measurements are
means ± SEM, and significance was determined using the two-way ANOVA with post tukey test. a Reported as percent of total fatty acid. b Most common name given. Omega-9, omega-6 and omega-3 FAs are indicated as n9, n6, and n3, respectively
Control diet Fish oil diet Fatty acidb Structure
Sham (n=6) TAC (n=9) Sham (n=6) TAC (n=9)
Myristic acid C14:0 0.23 ± 0.02 0.23 ± 0.01 0.31 ± 0.02 A 0.30 ± 0.03 C Palmitic acid C16:0 13.5 ± 0.21 13.7 ± 0.10 15.0 ± 0.26 A 14.9 ± 0.20 C Palmitoleic acid C16:1n7 0.50 ± 0.07 0.56 ± 0.04 0.65 ± 0.07 0.66 ± 0.06 trans Palmitoleic acid C16:1n7t 0.03 ± 0.00 0.03 ± 0.00 0.037 ± 0.002 A 0.044 ± 0.001 B,C Stearic acid C18:0 16.6 ± 0.39 17.0 ± 0.15 16.0 ± 0.27 16.2 ± 0.25 C Oleic acid C18:1n9 12.2 ± 0.76 12.9 ± 0.47 9.99 ± 0.46 A 10.6 ± 0.35 C trans Oleic acid C18:1t 0.17 ± 0.01 0.16± 0.00 0.15 ± 0.00 A 0.14 ± 0.00 C Linoleic acid (n-6) C18:2n6 17.4 ± 0. 51 16.6 ± 0.33 11.7 ± 0.14 A 12.4 ± 0.45 C trans Linoleic acid (n-6) C18:2n6t 0.07 ± 0.00 0.07 ± 0.00 0.06 ± 0.00 0.07 ± 0.00 α-Linolenic acid (n-3) C18:3n3 0.04 ± 0.00 0.04 ± 0.00 0.03 ± 0.00 0.04 ± 0.00 γ-Linolenic acid (n-6) C18:3n6 0.04 ± 0.00 0.04 ± 0.00 0.03 ± 0.00 0.02 ± 0.01 B,C Eicosenoic acid C20:1n9 0.58 ± 0.04 0.57 ± 0.02 0.45 ± 0.02 A 0.46 ± 0.03 C Eicosadienoic acid (n-6) C20:2n6 0.33 ± 0.01 0.30 ± 0.01 A 0.24 ± 0.01 A 0.25 ± 0.01 C Eicosatrienoic acid (n-6) C20:3n6 0.75 ± 0.08 0.80 ± 0.02 0.56 ± 0.01 A 0.54 ± 0.01 C Arachidonic acid (AA, n-6) C20:4n6 10.2 ± 0.15 10.6 ± 0.15 3.85 ± 0.08 A 3.83 ± 0.16 C Eicosapentaenoic acid (EPA, n-3) C20:5n3 0.01 ± 0.00 0.01 ± 0.00 0.49 ± 0.02 A 0.47 ± 0.03 C Docosatetraenoic aicd (n-6) C22:4n6 1.18 ± 0.09 1.21 ± 0.03 0.13 ± 0.01 A 0.13 ± 0.00 C Docosapentaenoic acid (n-3) C22:5n3 0.60 ± 0.04 0.55 ± 0.07 2.38 ± 0.04 A 2.37 ± 0.06 C Docosapentaenoic acid (n-6) C22:5n6 9.84 ± 1.05 12.3 ± 0.25 A 0.42 ± 0.01 A 0.41 ± 0.01 C Docosahexaenoic acid (DHA, n-3) C22:6n3 15.5 ± 0.94 12.3 ± 0.59 A 37.5 ± 0.61 A 36.0 ± 0.59 C Lignoceric acid C24:0 0.04 ± 0.00 0.04 ± 0.00 0.04 ± 0.00 0.06 ± 0.00 B,C Nervonic acid C24:1n9 0.06 ± 0.00 0.07 ± 0.00 0.07 ± 0.01 0.11 ± 0.01 B,C
Composite Indices
Omega-3 Content (DHA+EPA) 15.5 ± 0.94 12.3 ± 0.59 A 37.9 ± 0.60 A 36.5 ± 0.59 C Total Saturated 30.4 ± 0.41 31.0 ± 0.20 31.3 ± 0.16 A 31.4 ± 0.17 Total Monounsaturated 1.33 ± 0.08 1.39 ± 0.04 1.36 ± 0.07 1.42 ± 0.07 n-3 Polyunsaturated 16.2 ± 0.96 12.9 ± 0.66 A 40.3 ± 0.58 A 38.9 ± 0.64 C n-6 Polyunsaturated 39.8 ± 0.67 41.9 ± 0.36 A 17.0 ± 0.16 A 17.6 ± 0.48 C n-3 : n-6 0.408 ± 0.030 0.309 ± 0.018 2.371 ± 0.040 A 2.227 ± 0.092 C EPA : AA 0.001 ± 0.000 0.001 ± 0.000 0.124 ± 0.006 A 0.125 ± 0.010 C EPA : DHA 0.000 ± 0.000 0.001 ± 0.000 0.013 ± 0.001 A 0.013 ± 0.001 C

Supplemental Table 3: Echocardiographic and hemodynamic assessment of cardiac
structure and function after 4 weeks of TAC.
AP<0.05 verus Control diet/sham;
BP<0.05 verus Fish oil diet/sham;
CP<0.05 verus Control diet/TAC.
All measurements are means ± SEM, and significance was determined using the two-way ANOVA with
post tukey test. BW, body weight; HW, heart weight; IVSth, intraventricular septal thickness; LVPWth,
left ventricular posterior wall thickness; LVEDD, left ventricular end-diastolic dimension; LVESD, left
ventricular end-systolic dimension; FS, fractional shortening; HR, heart rate; ESP, end-systolic
pressure; EDP, end-diastolic pressure; PG, pressure gradient.
Control diet Fish oil diet
Sham TAC Sham TAC
n=6 n=9 n=6 n=9
BW (g) 26 ± 0.5 27 ± 0.5 26 ± 0.5 26 ± 0.4
HW (mg) 133 ± 7 197 ± 12A 118 ± 2 160 ± 10
B,C
HW/BW (mg/g) 5.1 ± 0.2 7.3 ± 0.4A 4.5 ± 0.1 6.0 ± 0.3
B,C
Echocardiography n=6 n=9 n=6 n=9
HR 610 ± 19 608 ± 22 640 ± 9 641 ± 10
IVSth (mm) 0.87 ± 0.03 1.21 ± 0.04A 0.88 ± 0.02 1.11 ± 0.05
B
LVPWth (mm) 0.86 ± 0.01 1.27 ± 0.06A 0.86 ± 0.01 1.05 ± 0.03
B,C
LVEDD (mm) 3.37 ± 0.08 3.79 ± 0.19A 3.31± 0.06 3.66 ± 0.09
LVESD (mm) 1.87 ± 0.11 2.76 ± 0.28A 1.62 ± 0.12 2.01 ± 0.11
C
FS(%) 45 ± 2 28 ± 4A 51 ± 3 45 ± 3
C
Hemodynamics n=6 n=8 n=6 n=8
ESP (mmHg) 110 ± 2 186 ± 7A 118 ± 6 206 ± 9
B
EDP (mmHg) 1.8 ± 1.6 13.4 ± 3.5A 2.0 ± 0.7 2.5 ± 0.9
C
dP/dT max (mmHg/sec) 11979 ± 753 8485 ± 538A 11450 ± 897 12144 ± 631
C
dP/dT min (mmHg/sec) -11380 ± 1101 -8340 ± 549A -11038 ± 764 -11088 ± 654
C
Tau_w 3.9 ± 0.9 8.5 ±1.6A 7.0 ± 1.9 4.2 ± 0.9
C
PG(mmHg) 67.2 ± 8.8 76.4 ± 12.0