heteromorphic life histories of certain marine algae as adaptations
TRANSCRIPT

Heteromorphic Life Histories of Certain Marine Algae as Adaptations toVariations in Herbivory
Jane Lubchenco; John Cubit
Ecology, Vol. 61, No. 3. (Jun., 1980), pp. 676-687.
Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28198006%2961%3A3%3C676%3AHLHOCM%3E2.0.CO%3B2-H
Ecology is currently published by Ecological Society of America.
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available athttp://www.jstor.org/about/terms.html. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtainedprior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content inthe JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained athttp://www.jstor.org/journals/esa.html.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
The JSTOR Archive is a trusted digital repository providing for long-term preservation and access to leading academicjournals and scholarly literature from around the world. The Archive is supported by libraries, scholarly societies, publishers,and foundations. It is an initiative of JSTOR, a not-for-profit organization with a mission to help the scholarly community takeadvantage of advances in technology. For more information regarding JSTOR, please contact [email protected].
http://www.jstor.orgMon Mar 24 17:02:11 2008

Ecoloyy, 61(3), 1980, pp. 676-687 1980 by the Ecological Society of America
HETEROMORPHIC LIFE HISTORIES OF CERTAIN MARINE ALGAE AS ADAPTATIONS TO
VARIATIONS IN HERBIVORY1
JANE LUBCHENCO~'~ Depurtment of Zoology, Oregon State University,
Corvullis, Oregon 97331 USA
A N D
JOHNCUBIT^'^ Depurtment of Biology, University of Oregon,
Eugene, Oregon 97403 USA
Abstract. Many of the annual or ephemeral algae of the mid to high intertidal zones have het- eromorphic life histories, existing as upright morphs during seasonal algal blooms and as crustose or boring morphs during other portions of the year. Experimental removal of herbivores on the coasts of New England and Oregon resulted in the occurrence of the upright morphs in the times of year when they were normally absent (summer in our areas), demonstrating that such uprights can survive the summertime physical regime (contrary to earlier speculation). We suggest that the upright and crustose or boring stages of these algae represent mutually exclusive adaptations to fluctuations in grazing pressure: the upright stages are adapted for high rates of growth and reproduction when grazing pressure is low, and the crustose and boring stages are adapted for surviving through times of high grazing pressure. We predict isomorphic species of algae would predominate in these sorts of habitats if grazing pressure were more constant.
Key words: algae; Bangia; Codiolum; Conchocelis; herbivores; heteromorphology; life histories; Petalonia; Porphyra; Ralfsia; Scytosiphon; Ulothrix; Urospora.
established, stable life cycles involving several eco- Heteromorphic algae exhibit a high degree o f inde- logically distinct stages. To these we add certain het-
pendence and differentiation among the stages o f their eromorphic algae, which by Istock's (1967) criteria life cycles. Most o f the particular heteromorphic algae should be particularly evolutionarily unstable: the discussed in this paper have two separate, in some stages are often found under different growing condi- cases self-propagating, and ecologically distinct phas- tions, and some, rather than being larva-like devel- es which are so dissimilar in appearance that until re- opmental stages, are capable o f independent self-prop- cently they had been classified as separate species, agation that allows them to persist indefinitely in the and, in some cases, had been placed in separate fam- absence o f any other stage. ilies or orders. Algae with various types o f heteromorphic life
Istock (1967) has proposed that evolutionarily such cycles are both phylogenetically and ecologically wide- life cycles should be inherently unstable: as selection spread: heteromorphic algae occur in the three major acts independently on the separate stages, one stage divisions o f macroalgae (Chlorophyta, Phaeophyta, should eventually be eliminated or reduced in favor o f and Rhodophyta) and are found in a variety o f habi- the other. He cites as examples the loss or reduction tats. However, in this paper we concentrate on those o f larval or adult stages in some insects, cnidarians, species o f heteromorphic algae which comprise much and amphibians. Another more pervasive set o f ex- o f the macroalgal portions o f seasonal or ephemeral amples is the loss o f the free-living, haploid, gameto- blooms in the mid to high intertidal zones o f rocky phyte stage from the life cycles o f most plants and shores. These species are in the following divisions and animals. genera: Chlorophyta: Ulothrix and Urospora; Phae-
However, Istock (1967, 1970) points out there are ophyta: Petalonia and Scytosiphon; Rhodophyta: a number o f counterexamples with apparently long- Bangia and Porphyra. Although the algae in this group
are taxonomically quite different, their life cycles and morphologies have much in common. Each species
' Manuscript received 14 February 1979; accepted 1 June has two primary stages: the stage which appears in the 1979; final version received 29 August 1979. periodic algal blooms is an upright filament, tube, or
Order of authorship determined by coin toss. In previous publications, J . Lubchenco Menge. blade; the other is a nonupright crust or boring stage. Present address: Smithsonian Tropical Research Insti- ( A third stage, a small filamentous tuft, is also known
tute, APO Miami, Florida 34002 USA. for Petalonia and Scytosiphon.) The convprgence in

677 June 1980 HERBIVORY AND ALGAL LIFE HISTORIES
the patterns of life cycles o f these algae suggests their forms and life histories may be adaptations to a com- mon set o f selective factors; thus, these plants provide a system to examine the mechanisms by which com- plex life cycles are selected and maintained. The main question addressed in this paper is the following: what is the adaptive significance of the heteromorphic life cycles o f these intertidal algae?
An explanation already proposed is that the non- upright morphs are perennating stages which survive through the physically harsh seasons when the upright morphs are killed by such stresses as desiccation, in- solation, and high temperatures (Conway et al. 1976).
A second hypothesis occurred to each o f us during our independent studies on the effects o f herbivores on benthic marine algae. W e observed that in several heteromorphic species the upright morphs could sur- vive in physically harsh seasons i f protected from her- bivores, and that the nonupright morphs were either themselves grazer-resistant or specifically exploit mi- crohabitats which we infer protect them from grazing.
In this paper we suggest the hypothesis that spatial and temporal variations in grazing play a major role in the selection and continued maintenance o f the dif- ferent morphologies in these life cycles. In the follow- ing sections we present the results o f experiments test- ing this hypothesis on the east and west coasts o f the United States: in New England (studies o f J.L.) and in Oregon (studies of J.C.). Alternate hypotheses and ways to test them are suggested in the discussion sec- tion. I f our interpretations are correct, we predict that in less variable environments algae with heteromor- phic life cycles would not be maintained in such abun- dance and, instead, algae with isomorphic life cycles would increase in relative proportion.
New England studies Description of sites and species studied.-The het-
eromorphic algal species treated in the studies o f the rocky shores o f New England are the brown algae Petulonia fascia (0.F . Miill.) 0. Kuntze and Scyto-siphon lomentaria (Lyngb.) Link, the red algae Bangia fuscopurpurea (Dillw.) Lyngb., Porphyra miniata (C. Ag.) C. Ag., P. linearis Grev., and P. umbilicalis (L . ) J. Ag., and the possibly heteromorphic green alga Ulo-thrix flucca (Dillw.) Thur. in LeJolis. Observations were made on all o f these species, but experiments focused on the two brown algae Petulonia and Scy-tosiphon. These plants were studied at four study areas in Massachusetts (MA) and Maine (ME) from fall 1971 until summer 1977. The areas are nonestua- rine and range in exposure to wave action from very protected to very exposed as follows: Canoe Beach Cove, Nahant, MA; Grindstone Neck, ME; East Point, Nahant, MA; and Pemaquid Point, ME. All areas are described in detail in J . Lubchenco Menge (1975) and B. Menge (1976).
SEASONAL CYCLES OF :
HERBIVORE ACTIVITY
ULOTHRIX k m
B m A
PORPHYRA %-'Lv O N D J F M A M J J A S O
FIG. 1 . Subjective evaluations of percent of gastropod herbivore individuals active and of the relative abundance of the upright morphs of five heteromorphic algal species on the New England coast during the year. The herbivores are pri- marily Littorinu littorea, but also include other snails: L. ob-tusuta, L . saxatilis, Lucunu vinctu, Murgurites helicinu, and Acmaea testudinulis. Herbivores not included are isopods, amphipods, and diptera. Each algal species was recorded as being common (C), present (P), or absent (A) at each study area at least monthly for 3 yr. Study areas where gastropod herbivores are common (protected to intermediate in expo- sure to wave action) are indicated by the solid portions. Areas where gastropod herbivores are rare or absent (ex- posed sites) are indicated by cross-hatching. Scytosiphon, Petuloniu, and Porphyra are able to persist longer in exposed sites. Crustacean herbivores are often abundant at more ex- posed sites. Their effect on algae is yet to be determined.
The life histories of these algae are as follows: 1 ) Petalonia fascia (0.F . Miill.) 0. Kuntze and
Scytosiphon lamentaria (Lyngb.) Link (Phaeophyta: Scytosiphonaceae).
The upright morphs of both of these algae are "win- ter annuals" in New England (Fig. 1 , Taylor 1957, Kingsbury 1969). The upright blades of Petalonia (7.5-45 cm long) and upright tubes o f Scytosiphon (15-70 cm long) appear in tide pools and on the shore from October-November until March-May. Both species probably alternate (but not in an obligate sense) with tar-like crusts previously thought to be species of Ralf-sia (Phaeophyta: Ralfsiaceae) and perhaps with small filamentous tufts, as has been shown for these species elsewhere (Edelstein et al. 1970, Rhodes and Connell 1973, review by Wynne and Loiseaux 1976). The sea- sonal occurrence o f the ralfsioid crust morph is diffi- cult to assess accurately because these crusts are dif- ficult to distinguish in the field from other, valid Ralfsia species. The crustose morph, hereafter termed "Ralfsia," appears to be more abundant in the sum- mer than in the winter in New England.
Petalonia and Scytosiphon differ somewhat in the specifics o f their life cycles, in particular in the ploidy levels. According to Nakamura and Tatewaki (1975), Scytosiphon tubes are haploid and produce gametes. These gametes do one of three things: ( 1 ) develop par- thenogenetically into more haploid tubes, (2 ) develop parthenogenetically into haploid "Ralfsiu" crusts or tufts or (3) fuse, with the zygote developing into a

678 JANE LUBCHENCO AND JOHN CUBIT Ecology, Vol. 61, No. 3
diploid "Ralfsia" crust or tuft. Both haploid and dip- loid crusts or tufts produce zooids which develop into upright tubes. Meiosis occurs only when diploid crusts or tufts form zooids. Wynne (1969) found that his "Ralfsia" could also produce more "Ralfsia" crusts. Thus in this species the upright morph is haploid while the other can be either diploid or haploid, indicating a partial decoupling o f the morphological and genetic components.
This decoupling is complete in Petalonia, where all stages (crusts, tufts, and blades) appear to be of the same ploidy number. No evidence o f sexuality has been reported. "Ralfsia" crusts or tufts produce zooids which develop into either more "Ralfsia," or tufts or Petalonia blades. Blades produce swarmers which develop into either crusts, tufts , or blades (Wynne 1969, Wynne and Loiseaux 1976).
Neither of these species represents the classical pic- ture o f a strict alternation o f generations (haploid with diploid). More significantly, both species appear to possess tremendous morphological flexibility, each morph being able to produce either more individuals like itself or the other alternate morph. In other words, neither genetic nor morphological alternation is obli- gate. Why then are there two distinct morphs, each with this plasticity?
Proximate factors affecting which morph is pro- duced have been investigated in the laboratory. In nu- merous culture studies, blades o f Petalonia and tubes o f Scytosiphon were found to be produced mainly un- der simulated "wintertime conditions ," i.e., short daylength photoperiods and cool temperatures. The "Ralfsia" crusts or tufts were obtained under labo- ratory "summertime conditions," i.e., long daylength photoperiods and warm temperatures (Wynne 1969, Roeleveld et al. 1974, Dring and Luning 1975, Naka- mura and Tatewaki 1975).
It seems significant that in these laboratory culturing experiments there was usually some variation in the response o f plants to temperature and photoperiod; that is, a small percentage o f the progeny became the "wrong" morph for a particular set o f conditions (Wynne 1969, Roeleveld et al. 1974). For example, in three o f Wynne's experiments with Petalonia, 5-20%, 5-10%, and 0% o f the progeny developed into blades under summertime conditions (18"-1YC, 16 h light-8 h dark). The potential significance o f this variation will be discussed below.
Both species' laboratory behavior generally corre- sponds to their seasonal occurrence in New England: uprights are produced under wintertime conditions, and crusts appear primarily under summertime con- ditions.
2) Bangia fuscopurpurea (Dillw.) Lyngb. (Rho- dophyta: Bangiaceae).
This species has two very different phases in its life history: upright, macroscopic plants which are unise- riate or multiseriate filaments, and an endolithic fila-
mentous form previously known as "Conchocelis." Upright plants are dioecious and produce two of three possible types of spores: ( 1 ) "monospores" or "neu- tral spores" which develop into more upright filaments and one of the following types o f spores (2) "carpo- spores" or "alpha spores" which germinate to give rise to the "Conchocelis" phase, or (3) "spermatia" or "beta spores" that presumably act as male gametes and fuse with the cells that develop into carpospores. The "Conchocelis" phase produces two types o f spores: ( 1 ) "monospores" which develop into more ' 'Conchocelis" plants, and (2) ' 'conchospores' ' which develop into the macroscopic upright phase (Sommer- feld and Nichols 1970). Both photoperiodism (Rich- ardson and Dixon 1968, Dixon and Richardson 1970, Richardson 1970) and temperature (Sommerfeld and Nichols 1973) have been shown to control the forma- tions and release o f spores o f both phases o f the life history in the laboratory. In New England, the upright Bangia filaments measure =lo-20 mm in length and 0.15 mm in diameter and usually appear in December- February and persist through March-May (Fig. 1 ) . The seasonal occurrence o f the "Conchocelis" phase is not known; however, the plants have been found in intertidal mollusc and barnacle shells (Bird 1973). The "Conchocelis" phase is thought to be perennating.
3) Porphyra umbilicalis (L . ) J . Ag., P. miniata (C. Ag.) C. Ag., and P . linearis Grev.
As in Bangia, the life history o f these species in- volves two different phases with different morpholo- gies. The macroscopic Porphyra thalli discussed here are leafy sheets usually ranging in size from 3 to 15 cm long, are haploid, and alternate with a "Conchocelis" phase (as described for Bangia) which is usually dip- loid and which bores into and lives in calcareous sub- strates.
There exists considerable variation among the species with respect to seasonal occurrence and spe- cifics o f the life history (Conway 1964, Edelstein and McLachlan 1966, Chen et al. 1970, Bird et al. 1972, Bird 1973, Bold and Wynne 1978, Hawkes 1978). The foliose phase o f P. linearis is a high intertidal winter annual. Since no neutral spores are formed by the blades and the basal portion does not perennate, blades must come solely from conchospores. Con-chospores are released only at 13°C in culture, but can be found throughout the year in the intertidal. The "Conchocelis" phase has been found in subtidal mus- sel (Modiolus modiolus) shells (Bird et al. 1972, Bird 1973). The foliose thallus o f P. miniata occurs in the mid and low intertidal and subtidal regions from spring to summer. All blades apparently come from concho- spores. Conchospore release is triggered in the labo- ratory at temperatures <13"C (Chen et al. 1970). P. umbilicalis has several upright forms: ( 1 ) perennial ro- settes present in the high zone (Edelstein and Mc-Lachlan 1966), (2) an elongated morph that appears in the winter in mid zones, and (3) a persistent blade

679 June 1980 HERBIVORY AND ALGAL LIFE HISTORIES
phase that occurs in the low intertidal and reproduces by neutral spores to form more blades (Conway 1964, Chen et al. 1970).
In the areas reported in this paper, foliose uprights usually appeared in October-November and persisted through May-June where herbivores were present or occurred throughout the year where herbivores were absent or rare, for example, at exposed areas (Fig. 1 ) . Species o f Porphyra could not always be distinguished and are lumped together here.
4) Ulothrix flacca (Dillw.) Thur. in LeJolis (Chlo- rophyta: Ulotrichaceae).
Both isomorphic and heteromorphic life cycles have been described for U . flacca elsewhere. Perrot (1972) suggests that in France it is really two separate species, one isomorphic and occurring in the very high intertidal zone and the other heteromorphic and oc-curring lower. The isomorphic species, termed "Form A" by Perrot, has upright, filamentous gametophytes and sporophytes. Each stage reproduces to form either more plants o f the same or the alternate stage. The heteromorphic species, "Form B," has dioecious, up- right, filamentous gametophytes which produce two types o f cells: ( 1 ) zoospores which develop directly into more upright gametophytes and (2) gametes which fuse to form a zygote that develops into a discoid spo- rophyte resembling the green prostrate Codiolum. This Codiolum-like stage produces aplanospores which develop directly into upright gametophytes. Both photoperiod and temperature are involved as cues in this nonobligate alternation.
The life histories o f the New England U . flacca have not been investigated, but circumstantial evidence suggests that at least the heteromorphic form is pres- ent. Ulothrix occurs as long, slender filaments (10-70 mm long, 0.15 mm in diameter) forming dense mats which are common throughout the intertidal region from October-November to March-May (Fig. 1 ) . When Ulothrix disappears during the spring, "Co-diolum" appears and persists throughout the summer and fall.
Herbivore occurrence and algal preference.-The most abundant herbivore in the New England rocky intertidal zone is the common periwinkle snail Litto-rina littorea which occurs in tide pools and on emer- gent substrata where these heteromorphic algae are found ( J . Lubchenco Menge 1975, Lubchenco 1978). Littorina is usually active from spring (March-May) to late fall (October-November, Fig. I) , at just the time when the upright morphs o f Scytosiphon, Peta- lonia, Bangia, Porphyra, and Ulothrix are absent (Fig. 1).
Laboratory preference experiments indicate that Littorina will readily eat the upright morphs of all o f these species, but not the crustose forms (Lubchenco 1978). Crusts like these are not totally herbivore re- sistant, but are much less preferred than are the up- right morphs or many other nonheteromorphic upright species.
6. Roof p--X, '0°1 x--K'x "t;kLp, I I 'x--f
C. h. w69 enclosure 0-0 Petalonia 0-,,o S@asiphm
50 x---x Ephememl u ;\ ...,,
Algae W X-- A,
lo]0 x,., ; , , I, , \; , , , ,
U
X p--x--x--x, X
'\ , \ x--xI i
! I
, D. Herbivore 'x,, ;x' "\x-.X" 5 0
'x' ,* Exclusion I x' p..
O . . , - . . c . A M J J A S O N D J F M A M J J A
1 9 7 3 1 9 7 4
FIG.2. Effect of herbivores on the occurrence of Peta-loniu and Scytosiphon in the high intertidal zone at Grind- stone Neck, Maine. The numbers below each treatment title indicate the mean number of grazers present and 95% con-fidence interval. L.1. = Littorina littoreu, L.o. = L. obtusata, L.s. = L. saxutilis. Ephemeral algae are those plants that usually persist for short periods of time during the year. Pe-tuloniu and Scytosiphon are ephemeral species by this defi- nition, but are separated from the rest of the ephemeral algae since they are the focus of this experiment. The other ephem- eral species appearing in this experiment include some het- eromorphic and some nonheteromorphic uprights: Bangia fuscopurpureu, Dumontia incrussata, Porphyra spp., Rhi-zoclonium tortuosum. Spongomorpha sp., and Ulothrixjuc-ca. Separate percent cover values for each species were usually taken; occasionally some species were mixed togeth- er so thoroughly that it was necessary to lump them together (as for Scytosiphon and Petulonia in A and B). The approx- imate period of seasonal inactivity of the herbivore, 1,ittorinu littoreu, is indicated by the bar at the top of the figure. See text for details of treatments.
Field herbivore exclusions: design.-During the course of a general investigation o f the effects of rocky intertidal herbivores on algae in New England, a series o f 10 x 10 x 3 or 10 x 10 x 5 cm stainless steel mesh cages were attached to the rock and used to exclude or enclose various herbivores (see J. Lubchenco
JANE LUBCHENCO AND JOHN CUBIT Ecology, Vol. 61, No. 3
Herbiwres inactive
A. CONTROL D HERBIVORE EXCL.. E!lQ& REMOVAL
C. L LITTOREA ENCLOSURE F HERB. EXCL., m S 8 EPHEM REM.
'"1 4 LI 1 ad*- *,J- /\o
-- --x--x-.." X --,* 0 0 a *-80 . . " , '
J J A S O N O J F M A M J J A J J A S O N D J F M A M J J A
0... . .O Scytosiphar *-a A.-.A x- - -x Ephememl Algoe
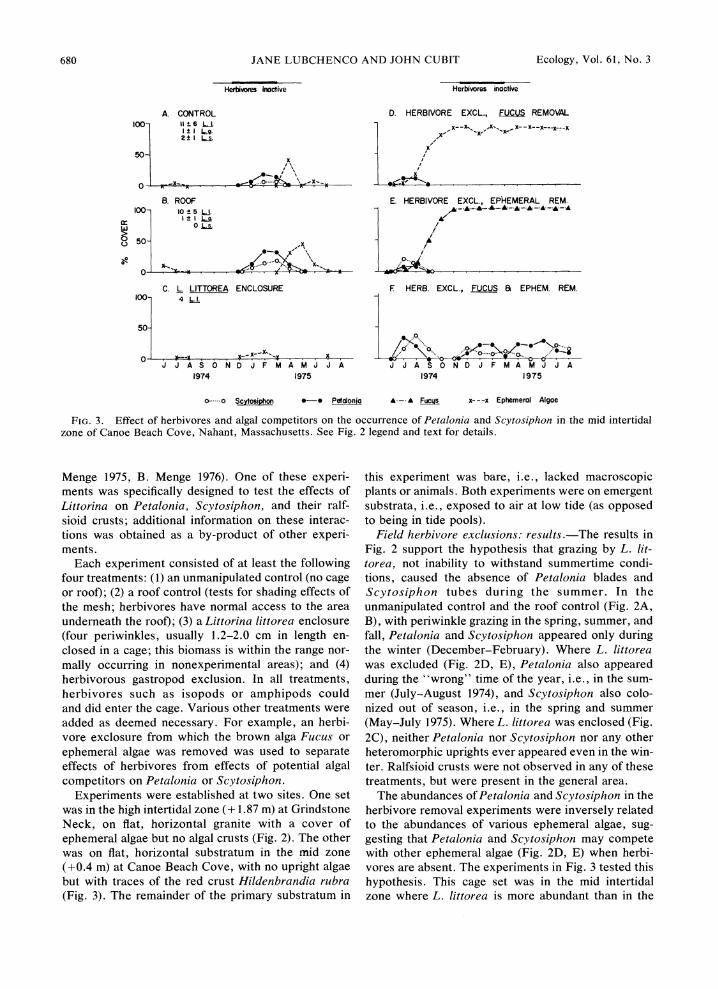
FIG.3. Effect of herbivores and algal competitors on the occurrence of Petalonia and Scytosiphon in the mid intertidal zone of Canoe Beach Cove, Nahant, Massachusetts. See Fig. 2 legend and text for details.
Menge 1975, B. Menge 1976). One of these experi-ments was specifically designed to test the effects of Littorina on Petalonia, Scytosiphon, and their ralf-sioid crusts; additional information on these interac-tions was obtained as a by-product of other experi-ments.
Each experiment consisted of at least the following four treatments: (1) an unmanipulated control (no cage or roof); (2) a roof control (tests for shading effects of the mesh; herbivores have normal access to the area underneath the roof); (3) a Littorina littorea enclosure (four periwinkles, usually 1.2-2.0 cm in length en-closed in a cage; this biomass is within the range nor-mally occurring in nonexperimental areas); and (4) herbivorous gastropod exclusion. In all treatments, herbivores such as isopods or amphipods could and did enter the cage. Various other treatments were added as deemed necessary. For example, an herbi-vore exclosure from which the brown alga Fucus or ephemeral algae was removed was used to separate effects of herbivores from effects of potential algal competitors on Petalonia or Scytosiphon.
Experiments were established at two sites. One set was in the high intertidal zone (+1.87 m) at Grindstone Neck, on flat, horizontal granite with a cover of ephemeral algae but no algal crusts (Fig. 2). The other was on flat, horizontal substratum in the mid zone (+0.4 m) at Canoe Beach Cove, with no upright algae but with traces of the red crust Hildenbrandia rubra (Fig. 3). The remainder of the primary substratum in
this experiment was bare, i.e., lacked macroscopic plants or animals. Both experiments were on emergent substrata, i.e., exposed to air at low tide (as opposed to being in tide pools).
Field herbivore exclusions: results.-The results in Fig. 2 support the hypothesis that grazing by L. lit-torea, not inability to withstand summertime condi-tions, caused the absence of Petalonia blades and Scytosiphon tubes during the summer. In the unmanipulated control and the roof control (Fig. 2A, B), with periwinkle grazing in the spring, summer, and fall, Petalonia and Scytosiphon appeared only during the winter (December-February). Where L. littorea was excluded (Fig. 2D, E), Petalonia also appeared during the "wrong" time of the year, i.e., in the sum-mer (July-August 1974), and Scytosiphon also colo-nized out of season, i.e., in the spring and summer (May-July 1975). Where L. littorea was enclosed (Fig. 2C), neither Petalonia nor Scytosiphon nor any other heteromorphic uprights ever appeared even in the win-ter. Ralfsioid crusts were not observed in any of these treatments, but were present in the general area.
The abundances of Petalonia and Scytosiphon in the herbivore removal experiments were inversely related to the abundances of various ephemeral algae, sug-gesting that Petalonia and Scytosiphon may compete with other ephemeral algae (Fig. 2D, E) when herbi-vores are absent. The experiments in Fig. 3 tested this hypothesis. This cage set was in the mid intertidal zone where L. littorea is more abundant than in the

June 1980 HERBIVORY A N D ALGAL LIFE HISTORIES 68 1
high zone experiments of Fig. 2. Here again, Petalonia and Scytosiphon occurred in the control and roof only during the wintertime when the snails are usually less active (Fig. 3A, B). Where L. littorea was enclosed in cages (Fig. 3C), it prevented most algae (including Petalonia and Scytosiphon) from becoming estab- lished, even in the winter. L. littorea usually retreat to crevices during the winter and seldom (but occa- sionally) forage. Snails enclosed in cages do feed, but at reduced rates as compared with the summer. Thus caged snails can evidently prevent even wintertime occurrence of most algae, unless very dense settle- ment of algae (swamping) occurs. Where herbivores were excluded (Fig. 3D, E , F), algae were more abun- dant. The particular kind of alga that comes in is prob- ably a function of the plants which are available to colonize and of competitive interactions between those plants which do settle (J. Lubchenco Menge 1975, Lubchenco and B. Menge 1978, J. Lubchenco, personal observation).
In all three exclosures (Fig. 3D, E, F) Petalonia blades and sometimes Scytosiphon tubes appeared in July, the "wrong" time of year for blades and tubes. Where Fucus and ephemeral algae were continually removed, Petalonia and Scytosiplzon continually col- onized (Fig. 3F). In herbivore exclusion cages where ephemeral species but not Fucus were removed, Fu- cus took over the cage (Fig. 3E) and neither Petalonia nor Scytosiphon occupied primary space. (EAperi- ments on competitive interactions between Fucus and various ephemeral species will be reported elsewhere.)
These results indicate that it is physiologically pos- sible for Petalonia blades and Scytosiphon tubes to exist during the summer months. These plants appear to be normal and healthy, judging by their color and size. Thus warmer temperatures or longer daylengths neither kill nor stunt them. Similar results have oc- curred in a number of other experiments designed to investigate other algae. Upright forms of Petalonia and Scytosiphon appeared out of season in 33% of 15 herbivore exclusion cages where free space was available (i.e., unutilized by other upright algae or an- imals; out of a total of 53 herbivore exclusion cages in place for 1-5 yr at the four study sites). Moreover, upright Petalonia and Scytosiphon can sometimes be found in nonexperimental (uncaged) areas at the "wrong" time of year. In the summer these plants were generally in the mid and low zones, both in tide pools and on emergent substrata. J.L. has observed this only a few times in 5 yr, at more wave-exposed areas, e.g., East Point and Pemaquid Point, where her- bivores such as littorinids are less abundant and/or less effective, and waves remove potential competi- tors more frequently. Other investigators have also reported Petalonia and Scytosiphon occurring during summer and fall months in New England at exposed sites (Lamb and Zimmerman 1964, Mathieson and
Fralich 1972, 1973, M. H. Zimmerman,personal com- munication).
Thus the general pattern of Petalonia and Scytosi- phon upright occurrence in the field corresponds to laboratory culturing results: uprights occur during the winter. The occurrence of a few uprights out of season (either in herbivore exclusion cages or exposed sites where herbivores are rare or ineffective) may be a result of the small variation in plant response to pho- toperiod and temperature reported by Wynne (1969; see above).
In similar herbivore exclusion and sometimes in competitor removal cages, the other heteromorphic species also appeared out of season. Uprights of Ulo- thrix, Bangia, and Porphyra were all observed during June, July, and August when they are normally absent (Fig. 1).
Oregon studies
Description of the sites and species studied.-The Oregon studies were performed at two sites near Coos Bay: South Cove of Cape Arago and Sunset Bay. The study plots at each site were above the mean high tide level on the wave-exposed sandstone rocks of the out- er coast. (The level of the study areas corresponds to "Zone 1," the uppermost intertidal zone, in the scheme of Ricketts et al. 1969.)
At the tidal level of these study areas there are wet and dry seasons which result from seasonally changing weather conditions and tidal cycles. In the wet season (late autumn, winter, and early spring) air tempera- tures are cooler, precipitation is greater, tidal levels are higher during the daylight hours, and the rocks receive more spray and wash from the waves gener- ated by winter storms. During this season the high rocks are almost continually wet, even at low tide. In the dry season (late spring, summer, early fall) the high intertidal rocks often dry out at low tide; this pattern is modified in some years by cool, wet fogs in the late spring and early summer (Cubit 1975).
In the areas of the study plots, the most abundant herbivores in terms of biomass per unit area were the acmaeid limpets, nearly all of which were Collisella (=Acmaea) digitalis. Chironomid flies, gammarid am- phipods, and littorinid snails were also occasionally abundant in certain areas.
The abundance of algae at these sites varied with the seasons. In the winter wet season there is a bloom of microalgae (diatoms and blue-greens) and macro- algae. Nearly all of the macroalgae comprising this bloom are in the genera treated in this paper: Bangia, Porphyra, and Urospora. Most of these algae disap- pear from the high intertidal zone during the drier con- ditions of summer. These plants and their life cycles are described below. (Bangia fuscopurpurea has been described in the previous section.) All of these species have heteromorphic life cycles.

682 JANE LUBCHENCO AND JOHN CUBIT Ecology, Vol. 61, No. 3
1) Porphyra perforata J. Ag., P. pseudolanceolata Krishnamurthy , and P. schizophylla Hollenberg (Rho- dophyta: Bangiaceae).
The general features of the life histories of these species of Porphyra are similar to those described above for this genus in New England. Most of the Porphyra plants in the Oregon study areas were <5 cm wide and <10 cm long. Size and shape vary some- what among the species. For these species, alterna- tions of generations between the upright stages and the "Conchocelis" stages may be obligatory, since no other types of reproduction have yet been reported (Mumford 1975, Conway et al. 1976).
2) Urospora perlicilliformis (Roth) Aresch. (Chlo- rophyta: Acrosiphoniaceae).
The upright, macroscopic plants are filamentous, reaching a maximum size of 30-40 mm long and 0.06 mm in diameter. Reproduction in the macroscopic stage is by at least four different reported methods: (1) fragmentation, (2) asexual production of quadrifla- gellate zoospores, (3) asexual production of akinetes (nonmotile spores), and (4) sexual reproduction. In the last, the zygote develops into a free-living "Codiol-urn" stage that penetrates the encrusting red alga Pet-rocelis (Abbott and Hollenberg 1976). Chapman and Chapman (1976) also report "Gomontia" as the alter- nate stage of Urospora.
Experiments and observations.-To investigate sea- sonal variations in the effects of limpet grazing on populations of high intertidal algae, sets of limpet ex- closures and controls (4-5 of each per set) were established at 3-4 mo intervals staggered over a period of 2.5 yr. A total of 46 exclosures and 45 controls was set out in this series of experiments. Each set of study plots was randomly selected from a much larger group of plots that had been chosen earlier for their relative similarity. Within each set of study plots, exclosures and controls were again designated randomly. The size and shape of a plot was determined by its topography: the average area of the study plots was 1651 cm2. There was no significant difference between the mean areas of the control and exclosure plots (P > .4).
Cageless methods were used to exclude the limpets since in this high intertidal habitat cages themselves would be expected to reduce the physical stresses of desiccation, insolation, and high temperatures, as well as reduce grazing. A continuous strip of copper paint kept the limpets out of the exclosure plots; a discon- tinuous strip of the paint was applied around controls. This paint was effective only in the exclusion of lim- pets; littorinid snails and arthropod grazers were not prevented from entering the exclosures. Further de- tails of this exclosure technique are reported in Cubit (1975).
Algal abundance was measured as the percent cov- erage of each genus and was estimated by projecting a stratified-random array of points onto color trans-
parencies of the plots, a method similar to that of Con- nell(1970). In the cases where the filamentous Bangia and Urospora grew so closely intermingled that they could not be separated their coverage was measured as a Bangia-Urospora mixture. The relative abun- dances of these algae are reported here only for the first 6 mo following the establishment of each study plot. In the longer term the succession of barnacles, perennial algae, and other organisms altered the sub- strate and other growing conditions within the exclo- sures so that for the purposes of this paper they were no longer comparable to the controls.
At all times of year, including summer, there was an immediate increase in algal cover following the ex- clusion of limpets. Urospora and Bangia were gener- ally the first macroalgae to appear in the exclosures, often in mixed stands, and were followed by Porphyra and the isomorphic green alga Ulva. The percent cov- erage data for the exclosures and controls are sum- marized by month for the 2.5-yr period in Table 1. The number of plots in which each alga was present is also given as a measure of the extent to which the alga occurred over the study areas. In summer the algal covers within the exclosures were much greater than those in the controls for the same months and were comparable to the natural algal blooms of the winter months.
From March through October Bangia, Urospora, and Porphyra formed higher percent covers and oc- curred in more plots of the exclosures than in the con- trols. With the possible exception of Porphyra, these genera were continually present throughout the year in the exclosures as compared to being ephemerally present in the controls. September was the only month in which no Porphyra was recorded in an exclosure. The probable explanation for this is that a total of only six exclosures was censused in this month, and that these six exclosures were probably too new for Por-phyra to have established. Three other, older exclo- sures not censused in September, but censused shortly before and after (28, 29 August 1972 and 5 October 1972), contained Porphyra on both dates, suggesting that this alga was present in September as well.
The months of lowest abundance of algae in the con- trols were April and October. In April none of the upright morphs was found in the 13 controls censused, while Bangia and Urospora were found in 12 and 11 of the 13 exclosures, forming up to 26 and 23% covers, respectively, with both averaging 3% overall in pure stands. In mixed stands, Bangia and Urospora ranged up to 51% cover with an average over all the exclo- sures of 15% cover. Porphyra was more rare, but oc- curred in five of the 14 plots. Its percent cover aver- aged over all exclosure plots was 2% and up to 1% in individual plots.
In October Urospora was the only alga of this group found in the controls: it occurred in trace amounts in

683 June 1980 HERBIVORY AND ALGAL LIFE HISTORIES
TABLE1. Monthly comparisons of algal covers in limpet exclosures and controls on the Oregon coast. Treatments were set up over a 2.5-yr period and monitored for 6 mo. Percent covers shown are overall means for each month followed by the minima and maxima, respectively, in parentheses. Percent covers of pure and mixed stands of Bangia and Urospora are presented separately (Bangia, Urospora, and B-U mix). The figures for the number of plots in which an alga occurred include plots in which there were only trace amounts (<0.5% cover) of the alga. These occurrence data for Bangia and Urospora are totals for all the plots in which the alga was present regardless of its being in mixed or pure stands. See text for further explanation.
Exclosures Controls
Por- B- U Por-N Bangia Urospora B-U mix phyra Total N Bangia Urospora mix phyra Total
Jan. % cover No. plots with
this alga
Feb. % cover No. plots with
this alga
Mar. % cover No. plots with
this alga
Apr. % cover No. plots with
this alga
May% cover No. plots with
this alga
June % cover No. plots with
this alga
July % cover No. plots with
this alga
Aug. % cover No. plots with
this alga
Sept. % cover No. plots with
this alga
Oct. % cover No. plots with
this alga
Nov. % cover No. plots with
this alga
Dec. % cover No. plots with
this alga
two of the 20 controls. In the exclosures, however, Porphyra was also present in the exclosures in Oc-Urospora and Bangia were abundant in mixed stands, tober: it was present in six of the 21 exclosures, av- occurring together in 12 of the 21 exclosures, aver- eraging 2% cover overall, and ranging up to 25% cov- aging 25% cover overall and ranging up to 97% cover er. in individual exclosures. The extent and species composition of the algal cov-

684 JANE LUBCHENCO AND JOHN CUBIT Ecology, Vol. 61, No. 3
ers varied considerably among the exclosures; obser- vations of colonization patterns within the exclosures suggested that this variation resulted from competition among the algal species (as in the New England stud- ies) as well as from grazing by littorinid snails and other small herbivores which were not excluded in these experiments. For instance, those exclosures with a higher degree of structural complexity (crev- ices, holes, barnacles, etc.) harbored higher densities of nonlimpet grazers and developed lower percent covers of algae. This effect varied from plot to plot according to the amount of shelter there. Observations that Porphyra was more common in those portions of the exclosures where light grazing by littorinid snails and other herbivores had reduced the densities of fil- amentous species of algae suggested that the abun- dance of Porphyra was negatively affected by com- petition, but favored by some grazing.
The results of the experiments presented here in- dicate that herbivory plays a substantial role in con- trolling the seasonal abundance of the upright thalli of certain common annual and ephemeral algae of the intertidal coasts of New England and Oregon. At the high algal densities that occurred within the herbivore exclosures, competition apparently also affected the abundances of some species.
In New England, the upright thalli of Petalonia, Scytosiphon, Ulothrix, Bangia, and Porphyra are nor- mally winter annuals, but in the experiments where they were protected both from herbivores and com- petitors, they were present in other seasons of the year. In the field experiments protecting them only from herbivores, these algae still occurred out of sea- son, but to a lesser extent than when competitors were also removed. In the laboratory experiments, the up- right thalli of these algae are all highly preferred food of the common grazers (Lubchenco 1978). Thus graz- ing is probably important in ultimately determining the seasonal abundance of the upright morphs.
On the high intertidal rocks in Oregon, the upright morphs of Bangia, Porphyra, and Urospora are also primarily winter annuals. Although individuals of these species may be found at other times of year at lower intertidal levels (and occasionally at higher levels) the bulk of the populations in the high intertidal zone occur during the winter (Table 1 "controls"). In the protection of the limpet exclosures these algae were present and common throughout the year. Den- sities of possibly competing species were not experi- mentally manipulated in the Oregon studies. However, observations of successional sequences and other growth patterns within the limpet exclosures suggest that interspecific competition also may have affected the relative abundances of these algae.
Areas yet to be investigated are the demography and other aspects of the ecology of the naturally oc-
curring crustose and boring "Ralfsia," "Conchocel-is," and "Codiolum" stages such as the study by Paine et al. (1979) for the crustose red alga Petrocelis middendorfJi. The crustose "Ralfsia" morphs in New England are known to be more grazer resistant than are the upright morphs (Lubchenco 1978). From our field observations we infer that the Oregon crustose morphs and all of the boring morphs are also less vulnerable to being removed by herbivores. In the course of our studies we have observed that the crus- tose algae in general are more abundant in, if not en- tirely restricted to, areas where herbivores are com- mon and active. There is indication from other studies that the establishment of at least some species of crus- tose algae may require the removal of their upright competitors by herbivores (Vine 1974, Adey and Vas- sar 1975, Wanders 1977).
There are few field studies applicable to the algae which penetrate or burrow into crustose algae, shells, and wood; however, we presume that within such sub- strata the boring morphs are sufficiently protected from herbivores that their survival exceeds that of the upright morphs through seasons of, and in habitats of, greater exposure to herbivory (Mumford 1973). (Some of these boring morphs may live within the shells of the grazers themselves.) As with the crustose forms, herbivory may also be necessary to maintain the bor- ing morphs. If the exteriors of the substrata occupied by these algae were to become covered with other algae, the algae below the substrate could be smoth- ered or shaded out.
The preceding experiments and observations indi- cate that increases of grazing intensity in summer can prevent the year-round survival of the upright stages of the algae we studied. Thus we suggest that a pri- mary adaptive value of the nonupright crustose and boring stages of these species is both in their ability to persist through times when the uprights are re-moved by grazers and to exist in areas of persistently heavy grazing where the upright stages themselves cannot survive. We suggest, however, that under con- ditions of light grazing the upright stages are compet- itively superior and have the additional selective advan- tages of rapid establishment, fast growth to reproductive maturity, and subsequent production of large numbers of propagules. We suggest that for plants such as these in mid to high intertidal habitats, a single type of plant cannot serve both functions well, because the adap- tations which confer protection from herbivores are mutually exclusive with those required for competitive superiority, rapid colonization, and high rates of re- production as discussed below. If this is correct, in habitats of spatially and temporarily heterogeneous patterns of grazing, a heteromorphic alga will be more successful than one which is isomorphic.
Among the adaptations of the nonupright stages which may provide protection from herbivores are the following: (1) In the crustose forms muth of the thallus

685 June 1980 HERBIVORY AND ALGAL LIFE HISTORIES
TABLE2. Predictions of the predominant life history for mid to high intertidal ephemeral algae under different grazing regimes, according to the definitions and constraints indicated in the footnotes.
If grazing pressure* is: Then we predict:
a. Light Isomorphology: competitively superior morphs 1. Constant and (uprights) predominate
b. Heavy Isomorphology: grazer-resistant morphs (e.g. crusts and borers) predominate
a . Predictable$ Heteromorphology: alternation of production and 2. Variable? and predominance of morphs (e.g. seasonally)
b. Unpredictable Heteromorphology: continuous production, but not survival, of both morphs
* The probability that a given individual alga will b e removed by herbivory In a given period of time. 7 Grazing pressure fluctuates with the condition that periods of high and low grazing pressure exceed generation tlrnes of nonupright and upright rnorphs, respectively. $ Fluctuations in grazing pressure can be forecast (e g., by correlat~ons with time or other cues from the environment).
adheres tightly to the substratum; thus the whole thal- gae may be under bimodal seiection pressures, simul- lus is not lost if an herbivore removes a small portion taneously evolving toward opposing ends of the r and at the base. (2) The thallus is tough, formed of many K continuum of selection. However, if our arguments layers of cells compacted together, and thus may be above are correct, the upright stages exhibit both r more difficult to graze. (3) Some Ralfsia are reported and K characteristics: rapid growth, early maturity, to contain tannins (Conover and Sieburth 1966) which and high reproductive output (r) as well as competitive may be herbivore deterrents. (4) The burrowing forms superiority (K), and the bimodality of selection exists are presumably protected from herbivores by being between this combination and the ability to withstand within other crustose algae or hard substrata such as grazing. shells and wood. In the following discussion we define the term graz-
Among the adaptations of the uprights that might ing pressure as the probability with which a given in- contribute to higher growth rates at the expense of dividual alga will be removed by herbivory in a given resistance to herbivory are the following: (1) Since period of time. If the preceding explanations are cor- most of the surface of an upright thallus neither ad- rect we would predict that the relative proportions of heres to the substratum nor is buried in it, and the heteromorphic and isomorphic algae should vary from thallus is either filamentous or only a few cells thick, habitat to habitat according to the seasonal fluctua- the ratio of exposed surface to internal volume of the tions of grazing in those places. In areas where grazing plant is greater than in the nonupright forms. This al- pressure does not fluctuate, we would expect iso- lows potentially higher growth rates through more rap- morphic algae (either upright or nonupright) to pre- id assimilation of nutrients (Odum et al. 1958, Fogg dominate, the type of morph being determined by 1965) and a higher ratio of photosynthetic area per unit grazing intensity (Table 2). If grazing pressure were biomass. (2) Upright forms presumably allocate less constantly low, the upright morphs should outcompete energy and material for attaching themselves to the the nonupright morphs (Table 2, la). If grazing pres- substratum or burrowing through it. (3) With small sure were constantly high, the uprights should be re- points of attachment, the uprights require less space moved and the nonuprights survive in their place (Ta- per unit biomass than the crustose forms, an adapta- ble 2, lb). Under such conditions, algae with obligate tion that is important in space-limited habitats of the alternations of heteromorphic generations should intertidal zone. (4) The uprights are probably not as eventually disappear as one morph or the other is elim- restricted in substrate requirements as are the burrow- inated by competition or grazing. Those algae with ing forms. alternations of heteromorphic generations that are not
As noted earlier it has been suggested that the crus- obligate may have one morph survive, but it should be tose and boring stages of heteromorphic algae serve at a lower relative fitness than similar, but isomorphic, as perennating phases through the seasons when the species in which no reproductive efforts are wasted on upright stages are unable to tolerate the physical producing the morphs which do poorly in these con- stresses of the habitat (Wynne 1969, Conway et al. ditions. 1976). Low surface-to-volume ratios and living within In contrast, where grazing pressure does fluctuate perforations of the substrate probably do render these from season to season, conditions should alternately nonupright forms more resistant to desiccation and the favor upright, then nonupright morphs, resulting in damaging effects of insolation and high temperatures. selection for seasonal alternation of these morphs We agree that the nonupright stages serve as peren- (Table 2, 2a). Greater fitness should accrue to those nating phases, but suggest that grazing and perhaps heteromorphic algae which allocate their resources competition should be considered as important factors into the production of nonuprights and uprights in syn- preventing the upright stages of these algae from being chrony with the grazing changes. Just prior to the on- perennial. set of the seasons of increased grazing it should be to
Vadas (1979) has suggested that heteromorphic al- the advantage of an upright alga to convert its re-

686 JANE LUBCHENCO AND JOHN CUBIT Ecology, Vol. 61, No. 3
sources to the grazer-resistant morph rather than have these resources consumed by herbivores. Similarly, selection should favor those algae concentrating their production of uprights in the seasons of reduced her- bivory (Table 2, 2a).
Finally, if grazing pressure fluctuates in an unpre- dictable manner, both morphs should be present con- tinually (Table 2, 2b). Each morph will be favored at some time, but prediction of precisely when is not possible. Thus each plant should be most fit by pro- ducing offspring of both morphs.
According to this scheme, the New England and Oregon studies reported above are examples of Table 2, 2a in which uprights are favored during the winter months and nonuprights during summer months. In our field studies, decreases in grazing pressure occur in winter and increases occur in summer. Numerous factors in the physical environment are correlated with the grazing changes and could be used by an alga as cues to change from one morph to another. These fac- tors include temperature, insolation, photoperiod, desiccation, wave force, and salinity. Laboratory cul- turing experiments have demonstrated that the switch from upright to nonupright morphs or vice versa is under photoperiodic or temperature control or both for Petalonia, Scytosiphon, Bangia, and Porphyra (Dring 1967, Wynne 1969, Bixon and Richardson 1970, Sommerfeld and Nichols 1973, Roeleveld et al. 1974). In the Oregon study areas species of Porphyra were observed to liberate spores during the first series of daytime low tides and warm weather in the spring.
For the algae discussed here, it makes little differ- ence which cues are used for timing, providing the cues are closely correlated with the seasonal increases and decreases in the probabilities of being consumed by herbivores. The strength of selective pressure to respond to these cues should depend on the closeness of the correlation. Conceivably there are environ-ments in which the seasonal variations in grazing pres- sure are the reverse of those in our study areas; in such areas we would expect opposite responses to the same cues, all other factors being equal. The mecha- nisms which we suspect cause the variations of grazing pressure in New England and Oregon might follow other seasonal patterns on other coasts. In New En- gland the winter reduction in grazing is apparently the result of wave action causing littorinids to decrease their grazing activities during the winter months (J. Lubchenco Menge 1975). Evidently this same phe- nomenon also occurs on parts of the Oregon coast where storm-generated waves decrease the densities andlor activities of littorinids (Castenholz 1961, Beh- rens 1974, Cubit 1975). In the particular Oregon study areas described in this paper the winter decrease in grazing pressure apparently results from a swamping of the grazing capacities of the limpet populations by increased algal productivity rather than a reduction of the grazing activities of the limpets (Cubit 1975). In other habitats factors such as seasonal removal of her-
bivores by predators, or seasonal influx of algal drift may also operate to reduce grazing at other times of year and change the selective regime for the timing of production of the various morphs of the heteromor- phic algae. In other words, rather than the seasonal abundance of these different morphs being dependent on the timing responses of the plants, the timing re- sponses are probably dependent on the seasonal sur- vival of these morphs.
Our experiments do not address factors involved in the origin of heteromorphology; they demonstrate only that variation in the grazing regime may maintain this adaptation in the species investigated.
The hypotheses and predictions put forth in this pa- per are not intended to explain the occurrence of het- eromorphology in all algae. Rather, we have focused on one of the mechanisms that may be involved in selection for heteromorphology of common species that have annual or ephemeral upright thalli in the mid and higher littoral zones. Since the morphologies of stages and the patterns of life cycles in other hetero- morphic algae differ from those we have studied, the selective mechanisms probably also differ. Moreover, the hypotheses presented in this paper are subject to further testing, particularly regarding the ecology of the nonupright stages. For example, more information is needed on (1) the demography and phenology of the crustose and boring stages, (2) the relative competitive abilities of uprights vs. nonuprights, and (3) effects of other factors, e.g., seasonal burial and scour by sand which might act similarly or in addition to grazing in selecting for heteromorphology.
We thank the following people for field assistance or com- ments on versions of this manuscript: L. Ashkenas, R. W. Day, P. K. Dayton, S. D. Gaines, M. E. Hay, J. A. Kilar, E. G. Leigh, B. A. Menge, T. F. Mumford, Jr., J . N. Norris, R. T. Paine, W. P. Sousa, J . A. West, S. D. Williams, and two anonymous reviewers. J. Lubchenco is grateful to N. W. Riser for use of Northeastern University's Marine Science Institute, Nahant, Massachusetts. This paper is contribution 51 from that laboratory. J. Lubchenco's research was sup- ported by National Science Foundation Grants GA-40003 to J. Lubchenco and GA-35617 and DES72-01578-A01 to B. Menge. J. Cubit thanks P. Frank and S. Cook for helpful discussion, T. F. Mumford, Jr. for identifying specimens of Porplzyra, and Sigma Xi for a grant-in-aid of research.
Abbott, I. A., and G. J. Hollenberg. 1976. Marine algae of California. Stanford University Press, Stanford, California, USA.
Adey, W. H., and J. M. Vassar. 1975. Colonization, succes- sion, and growth rates of tropical crustose coralline algae (Rhodophyta, Cryptonemiales). Phycologia 14:55-69.
Behrens, S. 1974. Ecological interactions of three Littorina (Gastropoda, Prosobranchia) along the west coast of North America. Dissertation. University of Oregon, Eugene, Or- egon, USA.
Bird, C. J. 1973. Aspects of the life history and ecology of Porphyra linearis (Bangiales, Rhodophyceae) in nature. Canadian Journal of Botany 51:2371-2379.
Bird, C. J., L. C.-M. Chen, and J. McLachlan. 1972. The

- -
687 June 1980 HERBIVORY ,4ND A1 -GAL LIFE HISTORIES
culture of Porphyrn litlc~clris (Bangiales, Rhodophyceae). Canadian Journal of Botany 50: 1859-1863.
Bold, H . C. , and M. J. Wynne. 1978. Introduction to the algae. Prentice-Hall, Englewood Cliffs, New Jersey, US.4.
Castenholz, R . W. 1961. The effect of grazing on marine littoral diatom populations. Ecology 42.783-794.
Chapman, V. J . , and D. J. Chapman. 1976. Life forms in the algae. Botanica Marina 19:65-74.
Chen, L. C.-M., T . Edelstein, E. Ogata, and J . McLachlan. 1970. The life history of Porphyrcl ~nitliertcr. Canadian Jour- nal of Botany 48:385-389.
Connell, J . H . 1970. A predator-prey system in the marine intertidal region. I. Balnrziis glcirzduln and several predatory species of Thczis. Ecological Monographs 40:49-78.
Conover, J. T. , and J . Sieburth. 1966. Effect of tannins ex- creted from Phaeophyta on planktonic animal survival in tidepools. Proceedings of the International Seaweed Sym- posium 5:99-100.
Conway, E. 1964. Autecological studies of the genus Por-phyrcl: 11. Porphyri~ rrtnbilici~lis (L.) J. Ag. British Phyco- logical Bulletin 2:349-363.
Conway, E. , T . F. Mumford, Jr., and R. F . Scagel. 1976. The genus Porphyrcl in British Columbia and Washington. Syesis 8: 185-244.
Cubit, J. 1975. Interactions of seasonally changing physical factors and grazing affecting high intertidal communities on a rocky shore. Dissertation. University of Oregon, Eu- gene, Oregon, USA.
Dixon. P. S.. and W. N. Richardson. 1970. Growth and reproduction in red algae in relation to light and dark cycles. Annals of the New York Academy of Sciences 175:764-777.
Dring, M. J . 1967. Effects of daylength on growth and re- production of the Conchocelis-phase of Porphyra terlern. Journal of the Marine Biological Association of the United Kingdom 47:501-510.
Dring, M. J., and K. Luning. 1975. .4 photoperiodic re-sponse mediated by blue light in the brown algae Scytosi-photi lotrzet~tciricz. Planta 125:25-32.
Edelstein, R . L . , L . C.-M. Chen, and J . McLachlan. 1970. The life cycle of Rrrlfsii~ c.lnvcltn and R. hortletii. Canadian Journal of Botany 48:527-531.
Edelstein, T., and J . McLachlan. 1966. Winter observations on species of Porphyrcz from Halifax County, Nova Scotia. Proceedings of the International Seaweed Symposium 5:117-122.
Fogg, G. E . 1965. Algal cultures and phytoplankton ecology. University of Wisconsin Press, Madison, Wisconsin, USA.
Hawkes, M. W. 1978. Sexual reproduction in Porphyra gczrdtieri (Smith et Hollenberg) Hawkes (Bangiales, Rho- dophyta). Phycologia 17:329-353.
Istock, C. A. 1967. The evolution of complex life cycle phe- nomena: an ecological perspective. Evolution 21:592-605. - 1970. Natural selection in ecologically and geneti-
cally deferred populations. Behavioral Science 15:lOl-115. Kingsbury, J. M. 1969. Seaweeds of Cape Cod and the Is-
lands. Chatham Press, Chatham, Massachusetts, USA. Lamb, I . M., and M. H. Zimmerman. 1964. Marine vege-
tation of Cape Ann, Essex County, Massachusetts. Pho- dora 66:217-154.
Lubchenco, J . 1978. Plant species diversity in a marine in- tertidal community: importance of herbivore food prefer- ence and algal competitive abilities. American Naturalist 112:23-39.
Lubchenco, J . , and B. A. Menge. 1978. Community devel- opment and persistence in a low rocky intertidal zone. Ecological Monographs 48:67-94.
Mathieson. A . C . , and R. .4. Fralich. 1972. Investigations of New England marine algae. V. The algal vegetation of the Hampton-Seabrook Estuary and the open coast near Hampton, New Hampshire. Rhodora 74:406-435.
Mathieson, A. C. , and R. A. Fralich. 1973. Benthic algae
and vascular plants of the lower Merrimack River and ad- jacent shoreline. Rhodora 75:52-64.
Menge, B. 1976. Organization of the New England rocky intertidal community: role of predation, competition, and temporal heterogeneity. Ecological Monographs 46:355- 393.
Menge, J. Lubchenco. 1975. Effect of herbivores on com- munity structure of the New England rocky intertidal re- gion: distribution, abundance, and diversity of algae. Dis- sertation. Harvard University, Cambridge, Massachusetts, USA.
Mumford, T . F . , Jr. 1973. Observations on the taxonomy and ecology of some species of Porphyrcl from Washington and Vancouver Island, British Columbia. Dissertation. University of Washington, Seattle, Washington, USA.
. 1975. Observations on the distribution and seasonal occurrence of Porphyra sc~hizoplzylle~Hollenberg, Por-plzyri~ torta Krishnamurthy, and Porplqra brumcllis sp. nov. (Rhodophyta, Bangiales). Syesis 8:321-332.
Nakamura, Y., and M. Tatewaki. 1975. The life history of some species of Scytosiphonales. Scientific Papers, Insti- tute of Algological Research, Hokkaido University 6:57- 93.
Odum, E. P., E. J. Kuenzler, and M. X. Blunt. 1958. Uptake of P3? and primary productivity in marine algae. Limnology and Oceanography 3:340-345.
Paine, R. T . , C. J . Slocum, and D. 0 . Duggins. 1979. Growth and longevity in the crustose red alga Petrocelis rniddetldotfji. Marine Biology 51:185-192.
Perrot, Y. 1972. Les U l o t h r i ~marins d e Roscoff et le prob- Ieme de leur cycle de reproduction. Societe Botanique de France, Memoires: 67-74.
Rhodes, R. G., and M. U. Connell. 1973. The biology of brown algae on the Atlantic Coast of Virginia. 11. Pemlot~ i i~ fizscia and Scvtosi~hotl lotnerztaricl. Chesapeake Science . . 14:211-215.
Richardson, W. N. 1970. Studies on the ~hotobiology of Bnrzgin friscopirrpirrec~. Journal of Phycology 6:216-219.
Richardson, W. N. , and P. S. Dixon. 1968. Life history of Beltzgin ,firsco~purprtrea (Dillw.) Lyngb. in culture. Nature 218:496-497.
Ricketts, E. F. , J. Calvin, and J. W. Hedgpeth. 1969. Be- tween Pacific tides. Stanford University Press, Stanford, California, USA.
Roeleveld, J . G. , M. Duisterof, and M. Vroman. 1974. On the year cycle of Peti~lotlii~.fosc.ii~ in the Netherlands. Netherlands Journal of Sea Research 8:410-426.
Sommerfeld, M. R., and H. W. Nichols. 1970. Develop-mental and cytological studies of Bnrzgicz,fiiscoprrrprtren in culture. American Journal of Botany 57:640-648.
Sommerfeld, M. R. , and H . W. Nichols. 1973. The life cycle of Bntzgici frtscoplrrpuren in culture. I. Effects of temper- ature and photoperiod on the morphology and reproduction of the Botigin phase. Journal of Phycology 9:205-210.
Taylor, W. R. 1957. Marine algae of the Northeastern Coast of North America. University of Michigan Press, Ann Ar- bor, Michigan, USA.
Vadas, R. L. 1979. Seaweeds: an overview: ecological and economic importance. Experientia 35:429-433.
Vine, P. J . 1974. Effects of algal grazing and aggressive behavior of the fishes Potnclcetltrus li\,idus and Acatlthurus sohczl on coral reef ecology. Marine Biology 24: I? 1-136.
Wanders, J . B. W. 1977. The role of benthic algae in the shallow reef of C u r a ~ a o (Netherlands Antilles) 111: The significance of grazing. Aquatic Botany 3:357-390.
Wynne, M. J . 1969. Life history and systematic studies of some Pacific North American Phaeophyceae (brown algae). University of California Publications in Botany 50:l-88.
Wynne, M. J.. and S. Loiseaux. 1976. Recent advances in life history studies of the Phaeophyta. Phycologia 15:435- 452.

You have printed the following article:
Heteromorphic Life Histories of Certain Marine Algae as Adaptations to Variations inHerbivoryJane Lubchenco; John CubitEcology, Vol. 61, No. 3. (Jun., 1980), pp. 676-687.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28198006%2961%3A3%3C676%3AHLHOCM%3E2.0.CO%3B2-H
This article references the following linked citations. If you are trying to access articles from anoff-campus location, you may be required to first logon via your library web site to access JSTOR. Pleasevisit your library's website or contact a librarian to learn about options for remote access to JSTOR.
Literature Cited
The Effect of Grazing on Marine Littoral Diatom PopulationsRichard W. CastenholzEcology, Vol. 42, No. 4. (Oct., 1961), pp. 783-794.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28196110%2942%3A4%3C783%3ATEOGOM%3E2.0.CO%3B2-D
A Predator-Prey System in the Marine Intertidal Region. I. Balanus glandula and SeveralPredatory Species of ThaisJoseph H. ConnellEcological Monographs, Vol. 40, No. 1. (Winter, 1970), pp. 49-78.Stable URL:
http://links.jstor.org/sici?sici=0012-9615%28197024%2940%3A1%3C49%3AAPSITM%3E2.0.CO%3B2-G
The Evolution of Complex Life Cycle Phenomena: An Ecological PerspectiveConrad A. IstockEvolution, Vol. 21, No. 3. (Sep., 1967), pp. 592-605.Stable URL:
http://links.jstor.org/sici?sici=0014-3820%28196709%2921%3A3%3C592%3ATEOCLC%3E2.0.CO%3B2-C
http://www.jstor.org
LINKED CITATIONS- Page 1 of 2 -

Plant Species Diversity in a Marine Intertidal Community: Importance of Herbivore FoodPreference and Algal Competitive AbilitiesJane LubchencoThe American Naturalist, Vol. 112, No. 983. (Jan. - Feb., 1978), pp. 23-39.Stable URL:
http://links.jstor.org/sici?sici=0003-0147%28197801%2F02%29112%3A983%3C23%3APSDIAM%3E2.0.CO%3B2-4
Community Development and Persistence in a Low Rocky Intertidal ZoneJane Lubchenco; Bruce A. MengeEcological Monographs, Vol. 48, No. 1. (Winter, 1978), pp. 67-94.Stable URL:
http://links.jstor.org/sici?sici=0012-9615%28197824%2948%3A1%3C67%3ACDAPIA%3E2.0.CO%3B2-L
Organization of the New England Rocky Intertidal Community: Role of Predation,Competition, and Environmental HeterogeneityBruce A. MengeEcological Monographs, Vol. 46, No. 4. (Autumn, 1976), pp. 355-393.Stable URL:
http://links.jstor.org/sici?sici=0012-9615%28197623%2946%3A4%3C355%3AOOTNER%3E2.0.CO%3B2-R
Uptake of P32 and Primary Productivity in Marine Benthic AlgaeEugene P. Odum; Edward J. Kuenzler; Marion Xavier BluntLimnology and Oceanography, Vol. 3, No. 3. (Jul., 1958), pp. 340-345.Stable URL:
http://links.jstor.org/sici?sici=0024-3590%28195807%293%3A3%3C340%3AUOAPPI%3E2.0.CO%3B2-Z
The Biology of Brown Algae on the Atlantic Coast of Virginia. II. Petalonia fascia andScytosiphon lomentariaRussell G. Rhodes; Mary U. ConnellChesapeake Science, Vol. 14, No. 3. (Sep., 1973), pp. 211-215.Stable URL:
http://links.jstor.org/sici?sici=0009-3262%28197309%2914%3A3%3C211%3ATBOBAO%3E2.0.CO%3B2-G
Developmental and Cytological Studies of Bangia fuscopurpurea in CultureMilton R. Sommerfeld; H. Wayne NicholsAmerican Journal of Botany, Vol. 57, No. 6. (Jul., 1970), pp. 640-648.Stable URL:
http://links.jstor.org/sici?sici=0002-9122%28197007%2957%3A6%3C640%3ADACSOB%3E2.0.CO%3B2-M
http://www.jstor.org
LINKED CITATIONS- Page 2 of 2 -