high affinity interaction between a … with a pestle in 50 ml of buffer ... glycan microarray...
TRANSCRIPT

HIGH AFFINITY INTERACTION BETWEEN A BIVALVE C-TYPE LECTIN AND A BIANTENNARY COMPLEX-TYPE N-GLYCAN REVEALED BY CRYSTALLOGRAPHY
AND MICROCALORIMETRY Jean-Philippe Gourdine‡, Gianluca Cioci¶, Laurence Miguet§1, Carlo Unverzagt║, Daniel Varón Silva║, Annabelle Varrot§, Catherine Gautier§, Emilie Juliette Smith-Ravin‡, and Anne Imberty§ From ‡Département de Biologie, Université des Antilles et de la Guyane, Pointe-à-Pitre, Guadeloupe,
France, ¶E.S.R.F. Experiments Division, Grenoble, France, §CERMAV-CNRS (affiliated to Université Joseph Fourier and belonging to ICMG), Grenoble, France, ║Bioorganische Chemie,
Universität Bayreuth, Germany. Running head: Crystal structure of Codakia orbicularis lectin
Address correspondence to: Anne Imberty, CERMAV-CNRS BP53, 38041 Grenoble cedex 09, France ; Fax: +33-476547203; E-Mail: [email protected]
Codakine is an abundant 14 kDa mannose-binding C-type lectin isolated from the gills of the sea bivalve Codakia orbicularis. Binding studies using inhibition of hemagglutination indicated specificity for mannose and fucose monosaccharides. Further experiments using a glycan array demonstrated, however, a very fine specificity for N-linked biantennary complex-type glycans. An unusually high affinity was measured by titration microcalorimetry performed with a biantennary Asn-linked nonasaccharide. The crystal structure of the native lectin at 1.3 Ǻ resolution revealed a new type of disulfide bridged homodimer. Each monomer displays three intramolecular disulfide bridges and contains only one calcium ion located in the canonical binding site that is occupied by a glycerol molecule. The structure of the complex between Asn-linked nonasaccharide and codakine has been solved at 1.7 Ǻ resolution. All residues could be located in the electron density map, except for the capping β1-4 linked galactosides. The α1-6 linked mannose binds to calcium by coordinating the O3 and O4 hydroxyl groups. The GlcNAc moiety of the α1,6 arm engages in several hydrogen bonds with the protein, while the GlcNAc on the other antenna is stacked against Trp108 forming an extended binding site. This is the first structural report for a bivalve lectin. Lectins are multivalent carbohydrate-binding proteins that play important roles in the social life of cells. A growing repertoire of lectins has been identified in invertebrates (1), where these molecules are involved in self/non-self recognition (2). For example lectins play a role in aggregation mechanisms in corals and sponges (3) or in sperm–egg recognition in
oysters (4). Lectin-mediation of symbiosis with algae or bacteria has been observed in coral (5) and nematodes (6). Nevertheless, the most common function assessed for lectins in marine invertebrates is their role in innate immunity by specific binding of polysaccharide-coated pathogenic bacteria (7,8). Different lectins have been identified in bivalves and they most frequently belong to the C-type lectin family. Proteins from this group of calcium-dependent lectins have been reported in oysters (9,10), scallops(11) and clams (12,13). C-type lectins are characterized by a carbohydrate recognition domain (CRD) with a conserved fold and the involvement of a calcium ion in carbohydrate binding (14). The crystal structure of mannose binding protein (MBP) was the first one to be described (15). The CRD belongs to a larger family sharing a common fold and is referred to as C-type lectin like domain (CTLD) (16).
Codakine is a 14 kDa C-type lectin purified from the gill of the tropical clam Codakia orbicularis (Linné 1758) by affinity chromatography on a mannose-agarose column (17). It forms homodimers and heterodimers with isoforms 1 (NCBI accession number AAX19697) and 2 (NCBI accession number ABQ40396) (13). A 19 amino acid peptide signal suggests that the lectin travels through a secretory pathway. The 129 amino acid sequence of the mature protein has significant sequence similarities to various fish lectins (33%). Sequence homologies have, moreover, been observed between codakine and mermaid nematodes (18). These results point to the probable role for the gill-located codakine in either antibacterial protection or in the recognition of sulfur-oxidizing bacteria, symbionts that are needed for the survival of Codakia orbicularis in sandy anaerobic environments (19).
1
http://www.jbc.org/cgi/doi/10.1074/jbc.M804353200The latest version is at JBC Papers in Press. Published on August 7, 2008 as Manuscript M804353200
Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

In this paper, we present data on the
specificity and affinity of codakine for various monosaccharides and oligosaccharides using inhibition of hemagglutination, glycan microarrays and microcalorimetry. The native crystal structure displays a new covalent dimerization mode. The structure of the complex with a high-affinity biantennary N-glycan displays a new oligosaccharide binding mode where both antennae are in contact with the protein.
EXPERIMENTAL PROCEDURES
Protein Purification The previously described protein
purification protocol (13) was used with minor modifications. Briefly, about 5g of nitrogen frozen gill tissue of Codakia orbicularis was crushed with a pestle in 50 ml of buffer composed of 20 mM Tris-HCl, 100 mM NaCl, 100 µM CaCl2, pH 7.4 (T buffer). After 10 min centrifugation at 10 000 rpm, the supernatant was dialysed overnight at 4°C against fresh T-buffer four times. Insoluble matter was pelleted by centrifugation as described above. The 0.25 µm filtered supernatant was loaded onto a mannose-agarose column pre-equilibrated with T-buffer. After washing with T-buffer containing 1 M NaCl codakine was eluted by 0.1M EDTA in T-buffer. The eluted fractions were pooled and dialysed extensively during 2 days at 4°C against T-buffer. The electrophoretic profile of eluted fractions was checked on 15% SDS-PAGE gel (20). The molar extinction coefficient and optical density at 280 nm were used to determine the concentration of codakine .
Hemagglutination Assays
Hemagglutination tests were performed using microtitre plates with U-bottom wells by two-fold serial dilution method. 25 µl of rabbit erythrocytes (2% in NaCl, Biomérieux France) were mixed with serially diluted codakine in T-buffer (described above), starting with a concentration of 1 mg/ml. After 30 min incubation at 37°C, the plates were read. One unit of hemagglutination activity was defined as the highest dilution of lectin giving a complete hemagglutination (HA unit μg/ml). For the inhibition tests, a dilution of codakine equivalent to four units of HA, corresponding to 15.6
μg/ml, was used. Inhibition of hemagglutination was assayed by two-fold serial dilutions of following sugars in T-buffer (D-Mannose, D-Galactose, D-Glucose, N-acetylglucosamine, L-fucose, N-acetylneuraminic acid, D-Rhamnose), each starting at a concentration of 100 mM.
Glycan Microarray Analysis
Purified codakine (1 mg/ml) was labeled with Alexa Fluor® 488 Protein Labeling Kit (Molecular Probes™, Invitrogen) according to the instructions of the manufacturer. The purification of Alexa-labeled codakine was performed by mannose-agarose chromatography as described above. Glycan microarray tests were carried out by the standard procedure of Core H of the Consortium for Functional Glycomics (http://www.functionalglycomics.org) (21). Preparation of Nonasaccharide-Asn 1
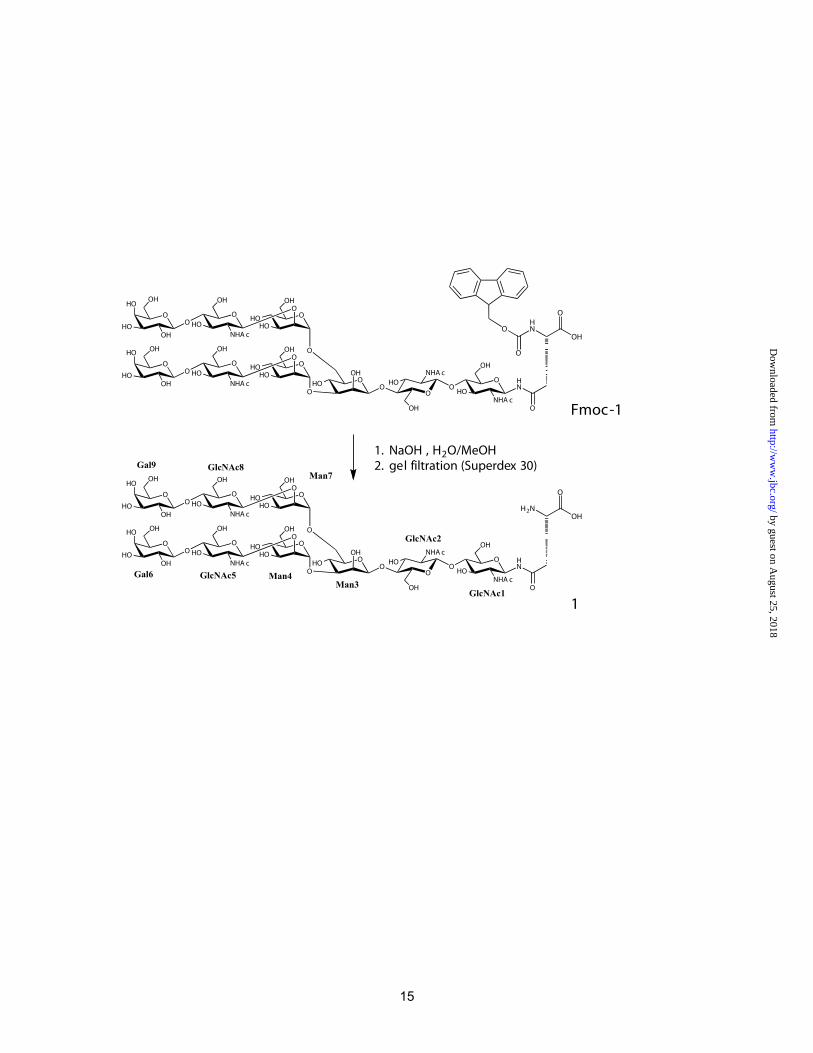
The protected compound Fmoc-1 was prepared from egg yolk by acid hydrolysis of the disialylated derivative of Fmoc-1 in analogy to the published procedures (22,23). For Fmoc deprotection (Scheme 1), a 5 mg portion of Fmoc-1 was dissolved in H2O/MeOH (75 μl, 30/9) and 0.5 M NaOH (20 μl) was added in four portions. After shaking for 10 min (TLC: 2-propanol/1M ammonium acetate), the suspension was acidified to pH 4-5 with 10% acetic acid. Subsequently, the suspension was diluted with 0.1 M NH4HCO3 to a final volume of 200 μl. The suspension was cleared by centrifugation and the supernatant was purified by gel filtration [Superdex 30 (16/60); flow rate: 1ml/min; eluent 0.1 M NH4HCO3; detection: 214 and 260 nm]. The peak eluting at 76 min was collected and lyophilized 5 times to yield 3.3 mg of nonasaccharide-Asn 1 (74.4 %). The purity of 1 (22) was confirmed by 360 MHz 1H-NMR in D2O (Figure S1 in suppl. Material). Titration Microcalorimetry
Thermodynamic parameters were estimated by isothermal titration calorimetry (ITC) using a Microcal (Northampton, MA) VP-ITC microcalorimeter. All experiments were carried out at 298K. Ligands and lectins were prepared in T-buffer (described above). Titration of codakine binding was performed in a cell with a volume of 1.447 ml by 30 injections of 10 µl of ligand with 5 min intervals while stirring at 310 rpm. Blank titration with buffer was used as
2
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

reference. The experimental data were fitted to a theoretical titration curve using software supplied by Microcal with ΔH (enthalpy change), Ka (association constant) and n (number of binding sites per monomer) as adjustable parameters, from the classical relationship (24). For each ligand, experiments were repeated two or three times.
Crystallization and Data Collection
Native codakine was concentrated to 15 mg/ml in 20 mM Tris-Cl pH 8, 5 mM CaCl2 and 5 mM α methyl-mannoside on Vivaspin 5 kDa (Vivascience). All crystals were obtained by the hanging-drop vapour diffusion method using 2 μl drops containing 50:50 (v/v) mix of protein and reservoir solution at 20°C. Initial crystallization screening was performed using the JCSG+ Suite (QIAGEN). After three months, crystals were obtained with a solution containing 0.2 M di-ammonium citrate pH 5, 20% w/v PEG 3350. A range of related conditions were used for co-crystallisation of the complex between codakine (22 mg/ml in the same buffer) and nona-Asn (10 mM). One large cube-shaped crystal was obtained with 0.2 M lithium sulfate, 100 mM sodium acetate pH 7.5 and 15% PEG 4000 from a well with a broken lamella.
Glycerol was added to 20% (v/v) to the crystallization solution as cryo-protectant prior to freezing crystals in a gaseous nitrogen stream at 100K. Native and complex data were collected at the European Synchrotron Radiation Facility (ESRF) on an ADSC Q4R CCD detector (Quantum Corp, Il, USA) on beamline ID14-1 and ADSC Q315R detector on beamline ID29, respectively. Diffraction images were processed using MOSFLM (25). All further computing was performed using the CCP4 suite (26), unless otherwise stated. Data processing statistics are presented in Table 1. Molecular Replacement and Structure Refinement
Attempts to solve the crystal structure of codakine by molecular replacement using the C-type lectin structures available in the Protein Data Bank were unsuccessful. A homology model was then constructed using FUGUE and ORCHESTRAR (Tripos Inc, St Louis). Among profile families identified by the FUGUE program, four templates with more than 25% identity with codakine were selected: human E-
selectin (27) (PDB code 1ESL), human lithosthatine (28) (1QDD), human tetranectin (29) (1TN3), and Hemitripterus americanus antifreeze protein (30) (2AFP). Structurally conserved regions were built by ORCHESTRAR and loops were modelled by ab initio approach. The conserved disulfide bridges and calcium ions were incorporated in the model that has been deposited in the Protein Model Data Base with accession number PM0074967 (http://mi.caspur.it/PMDB).
This model was then used for solving the codakine structure by molecular replacement with the program Phaser (31). The program Acorn (32) was subsequently used to improve the electron density. A few correctly placed segments were chosen from the molecular replacement solution and used as starting coordinates for Acorn phasing. Among the smallest fragments tested, Acorn was able to phase the structure starting from the positions of 8 sulfur atoms. The very high resolution (1.3 Ǻ) of the native crystals allowed the program to calculate an excellent electron density map where ARpWarp (33) built the complete model. In the carbohydrate binding site, clear density could be observed for one calcium atom. Two glycerol and one citrate molecules were also included in the model.
The structure in complex with the nona-Asn was solved by molecular replacement with Molrep (34) using the native codakine structure as search model. After the addition of a calcium ion and solvent molecules, clear residual electron density was visible for at least five sugar monomers. The oligosaccharide was docked manually into the electron density according to its chemical structure, no ambiguity at the glycosidic linkages was observed.
For the refinement of each structure, 5% of the observations were immediately set aside for cross validation analysis (35) and were used to monitor various refinement strategies. Manual corrections of the models using Coot (36) were interspersed with cycles of maximum-likelihood refinement with REFMAC (37). The two models were validated with the WhatIf suite (38) and deposited in the PDB with accession number 2vuv and 2vuz for the native codakine and the nonasaccharide complexes, respectively. The refinement statistics are listed in Table 1 Miscellaneous
Areas of buried surfaces were calculated with PISA (39). All figures were drawn with
3
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

PyMOL Molecular Graphics System, DeLano Scientific LLC, San Carlos, CA, USA unless otherwise stated.
RESULTS
Overall Structure of codakine In the presence of α-methyl-mannoside
(αMeMan), codakine crystallized in rock-shaped crystals of space group C2 that diffract to 1.3 Ǻ resolution. The asymmetric unit contains one monomer and a disulfide-bond dimer is generated by the 2 fold axis. The monomer consists of a typical C-type lectin CRD with two parts: the lower part contains the two helices and 3 strands and the upper part is composed of 4 strands and long loops (Figure 1A) (40). As predicted previously by bioinformatics, the mature protein lacks the 19 amino acids signal peptide present in the nucleotide sequence. The first two β-sheets and the last one run antiparallel and a salt bridge is observed linking the termini of the peptide chain. For the C-terminal amino acid, clear density for the backbone was found but not for the side chain. Apart from this, all amino acids could be modelled in the density map. Additional density was observed near Cγ of Pro91, which appears to be a 4-hydroxyproline.
Three intramolecular disulfide bonds are observed (Cys2-Cys13, Cys30-Cys124, Cys103-Cys116). The last two disulfide bridges are conserved in C-type lectins, linking the first helix to the last strand and one strand to a loop, respectively. The first bridge is present in long form CTLD (41). One free cysteine Cys53 is observed on the α2 helix. Cys44, the residue involved in the unique dimerization mode observed in this C-type lectin, is located at the beginning of the same helix. The dimer is further stabilized by hydrogen bonds from His51 and Glu55 to Asp11 and Gln8 on the other chain. Hydrophobic interactions were found for His51 stacking against Leu10 from the other chain and Leu10 against Phe9. Through the two fold symmetry, this creates a chain of contact between the 6 hydrophobic residues that extends to 20 Ǻ; this results in a dimmer interface area of about 400 Ǻ2.
C-type lectins usually contain two calcium ions in conserved binding sites referred to as site 1 and site 2 (40). In the present structure, only one calcium ion is present in site 2 that
corresponds to the monosaccharide binding site. The calcium is coordinated to the acidic oxygen of Glu93, Glu101 and Asp113, to the oxygen atoms from amide Asn95 and Asn112 and to the main chain oxygen of Asp113. Analysis of the electron density indicates that αMeMan is not present in the site but is instead replaced by a well-ordered glycerol molecule. Additional density was also observed and attributed to another glycerol molecule and a citrate molecule.
Analysis of the glycerol located in the carbohydrate binding site indicates that two oxygen atoms establish the contacts classically observed for the carbohydrate ligand. These two oxygens complete the coordination sphere of the calcium. The first oxygen shows hydrogen bonds to the side chains of Asn95, Glu93 and Asn112, the second oxygen to the side chains of Glu101 and Asn112 and to the backbone oxygen of Asp113 (Figure 1C). Affinity and Specificity for Monosaccharides
Hemagglutination inhibition assays were performed using rabbit erythrocytes. Codakine shows a hemagglutination activity for a minimum of 3.9 µg/ml (one unit of HA). Inhibition of interaction between the lectin and the red blood cells was assayed with a range of monosaccharides. The highest inhibition power (using 4 HA units of codakine) was observed for D-mannose and L-fucose) (25 mM). Glucose and N-acetylglucosamine (GlcNAc) have a weaker effect (100 mM) and no inhibition was observed for D-galactose, D-rhamnose, D-neuraminic acid even at the higher concentration. Codakine can therefore be confirmed as a mannose/fucose specific lectin, as was predicted from the EPN peptide signature in the carbohydrate binding site (40).
For a complete characterization of the interaction between codakine and the most efficient monosaccharides used in the hemagglutination assay, affinity and thermodynamic parameters were measured using isothermal microcalorimetry (ITC) (Table 2). Mannose and GlcNAc have a rather low affinity with a dissociation constant of 0.27 mM and 0.46 mM, respectively. No affinity could be measured for galactose. The dissociation constant obtained for αMeMan is 52 μM, which is about 20 times more potent than the interaction with mannose, an increase frequently observed in lectin-carbohydrate interactions. The
4
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

methyl fucoside (αMeFuc) also appears to be a good inhibitor as observed in hemagglutination assays. In all cases, the association is driven by enthalpy. The entropy term is weak and slightly favourable except for αMeMan. In this latter case, the strong enthalpy contribution (ΔH = -33.6 kJ/mol) is partly opposed by an entropy barrier (TΔS = -9.1 kJ/mol) as classically observed in protein-carbohydrate interactions (42). Affinity and Specificity for Oligosaccharides
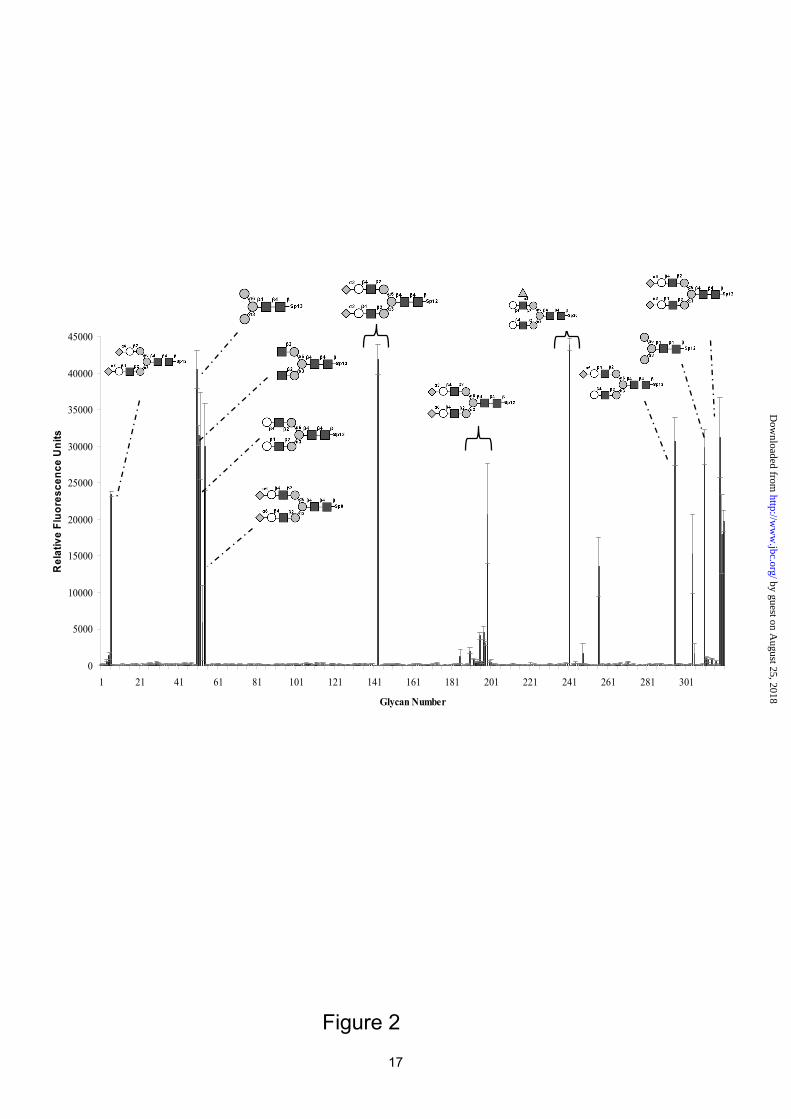
The specificity of the fluorescence labeled lectin was evaluated by binding to the 320 oligosaccharides present on the glycan array available at the Consortium for Functional Glycomics (Figure 2). Only 13 oligosaccharides are recognized with high affinity; they all represent complex-type biantennary N-glycans (complete data available as supplemental material). The shortest ligand is the core pentasaccharide, consisting of a chitobiose and a trimannoside with α1-3 and α1-6 linkage. Larger N-glycans elongated by successive addition of GlcNAc, galactose and sialic acid are also recognized by the lectin. Fucosylation at position 3 of the antennary GlcNAc moieties (Lewis x) is tolerated, but not fucosylation of the chitobiose core.
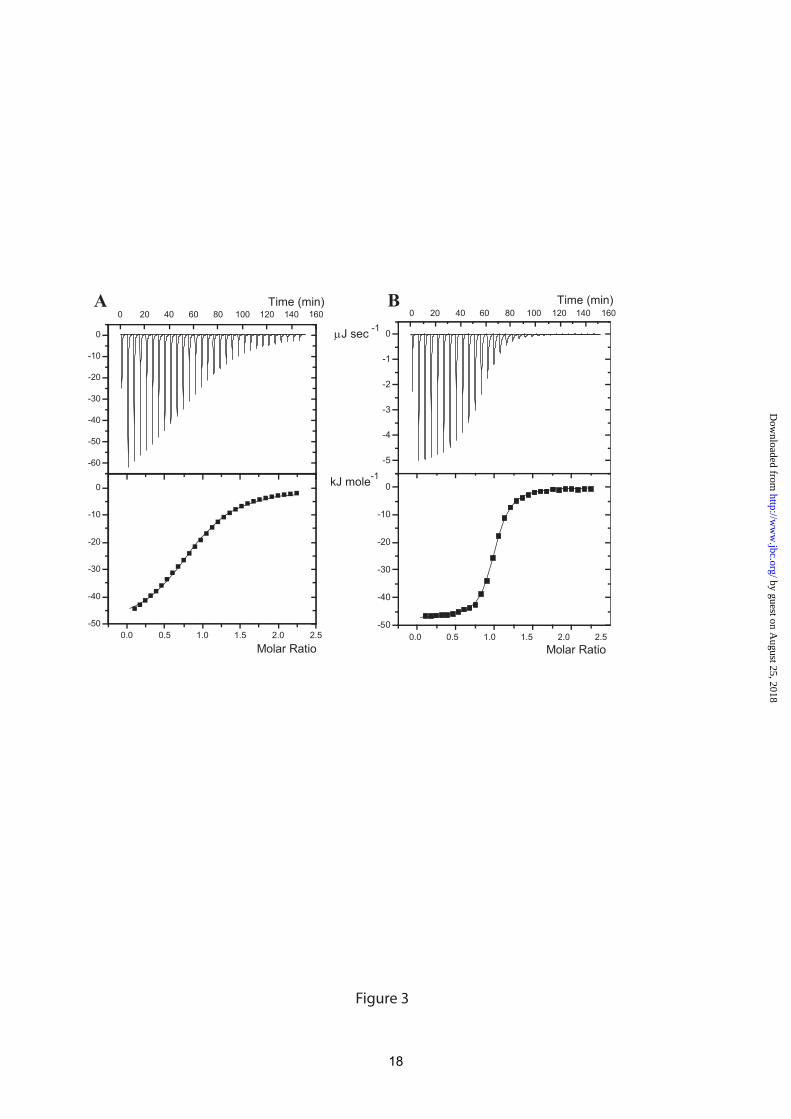
In order to characterize the thermodynamic and structural basis of this very narrow specificity, a biantennary nonasaccharide N-glycan linked to Asn (Scheme 1) was prepared from egg yolk. Codakine dissociation constants were measured for this compound and also for trimannoside in order to evaluate the influence of the internal chitobiose moiety on the high affinity (Figure 3). The affinity for the trimannoside (Kd= 80 μM) is of the same order of magnitude as the one measured for αMeMan (Table 2). In contrast, the affinity for nona-Asn is sub-micromolar (Kd= 432 nM). To our knowledge, this is the highest affinity reported for a C-type lectin, excluding multivalency effects. The 200 fold increase observed for nona-Asn compared to the trimannoside could be due to either the terminal N-acetyllactosamine (LaNAc) on each antenna, or to the presence of internal chitobiose. Since the core pentasaccharide Man3GlcNAc2 is efficiently recognized in the glycan array, it could be concluded that the internal chitobiose has a crucial effect on the affinity.
When analyzing the thermodynamic contributions, the binding of codakine to nona-Asn shows a higher enthalpy contribution (ΔH= -45.3 kJ/mol) than to trimannoside (ΔH= -41.9 kJ/mol). However, the difference in free energy (and therefore in affinity) is mainly due to the different entropy terms. Both oligosaccharides display an unfavorable entropy contribution, but the entropy for binding the trimannoside (TΔS= -18.5 kJ/mol) is much stronger than for the nonasaccharide (TΔS= -8.9 kJ/mol). Crystal Structure of Codakine/nona-Asn
Cocrystallization of codakine with nona-Asn yielded one rock-shaped crystal diffracting to 1.7 Ǻ in C2221 space group. The overall shape of each monomer is similar to what is observed in the structure of codakine complexed with glycerol. The intermonomer disulfide bridge also occurs, albeit with a longer S—S bond distance (2.33 Ǻ instead of 2.05 Ǻ). Such a deviation from normality might be due to radiation damage. This variation, associated with rotational flexibility of this linkage, induces a slightly different orientation of the second monomer with respect to the first one (Supplemental Figure S2).
Electron density could be readily identified for one calcium ion and five carbohydrate residues. After building the βGlcNAc12αMan13(βGlcNAc12αMan16) Man pentasaccharide, the two extra GlcNAc could be added at position 1 of the central βMan. The galactose residues capping the two antennae and the Asn moiety could not be located in the electron density (Figure 4A). The αMan of the 1-6 branch is located on the calcium ion in an orientation previously reported in DC-SIGN (43), with O3 and O4 coordinating the calcium ion. The OH3 group makes hydrogen bonds to the side chains of Glu93, Asn95, Glu101 and Asn112. The OH4 group binds to the Glu101 and Asn112 side chains and to the Asp 113 main chain oxygen, and the O6 hydroxyl to the Arg115 side chain (Table 3). The GlcNAc residue of the 1-6 branch interacts with Glu93 via OH6, with Asn95 via the ring oxygen O5, and with Ser97 via the acetyl oxygen. The branching βMan establishes two hydrogen bonds between O4 and the Asn107 and Trp108 side chains. The αMan of the 1-3 branch is not in contact with the protein but the antenna folds back and the terminal GlcNAc is perfectly stacked onto Trp108,
5
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

creating a strong hydrophobic interface. The two GlcNAc moieties of the chitobiose core do not make direct contact with the protein but the one linked to βMan interacts with the Ser61 side chain via a water molecule and establishes two intramolecular hydrogen bonds with the ring oxygens of βMan and αMan in the 1-6 branch (Figure 4B). The overall shape of the oligosaccharide is rather folded. The conformations of all the glycosidic linkages lie in energy minima that were identified previously in energy maps (44,45). In solution, the most flexible one would be the αMan1-6Man linkage that could generate several different overall shapes of the biantennary N-glycan. In the present case, the constraints created by the rather deep binding pocket result in a folded conformation, with the αMan of the 1-6 arm close to GlcNAc2 of the chitobiose core.
DISCUSSION
Comparison with other C-type Lectins The family of C-type lectins is characterized
by low sequence similarities despite a well conserved fold. Indeed, structural comparison by Secondary Structure Matching (SSM) (46) indicates high structural similarity with a range of vertebrate CTLDs such as the ones from scavenger receptor (47) (0.97 Ǻ for 122 amino acids), DC-SIGN (43) (1.29 Ǻ for 123 amino acids) and langerin (48) (1.09 Ǻ for 115 amino acids). Superimposition with DC-SIGN (Figure 1B) illustrates the high similarities in secondary structure and calcium site 2. Codakine lacks the calcium site 1, due to differences in the loop connecting strands β3 and β4. The loop is shorter and lacks acidic amino acids such as Glu324 in DC-SIGN. More importantly, Asp320 at the end of DC-SIGN β3-strand is replaced by a lysine (Lys70) that extends into the groove. The positively charged side chain occupies the calcium location in site 1 and establishes the equivalent interactions with conserved amino acids (Asp102 and Asn96 side chains and Glu101 backbone oxygen). Langerin is the only other structure of CRDs lacking calcium in site 1 and it also contains a lysine residue occupying the place of calcium. However, in langerin the connecting loop is longer and the resulting larger groove acts as a secondary carbohydrate binding site (48).
Molecular Basis for the High Affinity towards Oligosaccharides
Codakine displays millimolar affinity for monosaccharides (Kd = 0.27 mM for mannose) which is in agreement with ITC data previously obtained for other C-type lectins such as MBP with mannoside (49) and tunicate lectin TC14 with fucose (50). However, the strong affinity observed between codakine and the biantennary N-glycan (Kd = 0.432 μM) is very unique when compared to other C-type lectin/oligosaccharide interactions. The dissociation constant for E-selectin interacting with Sialyl-Lewis X has been estimated at 120 μM by fluorescence polarization (51) whereas DC-SIGN interaction with Man9GlcNAc2 was measured with a Kd of 26 μM by competition assays (52).
Indeed, the high affinity appears to be due to the extended binding site with numerous contacts between the oligosaccharide and the protein side chain. The stacking between Trp108 and the GlcNAc of the 3-arm also seems to be very favorable It results in a buried protein surface of 116 Ǻ2, almost as large as the surface buried by the mannose at the main binding site (133 Ǻ2). However, there are some discrepancies between this observation and the results derived from the glycan array data as the core pentasaccharide Man3GlcNAc2, which lacks the GlcNAc moieties on the antennae, is also a high affinity ligand of codakine.
When comparing the codakine/nona-Asn complex with the DC-SIGN/pentasaccharide βGlcNAc12αΜan13(βGlcNAc12Man16)Man (43), only the βGlcNAc12Man disaccharide segments on the calcium sites are similar (Figure 4C). In DC-SIGN, the rest of the pentasaccharide does not establish much contact with the protein and the binding site appears to be more open. The deeper character of the codakine binding site is mainly due to the longer loop between α2-helix and β3-strand (Figure 1D).
The inner chitobiose core seems to play a crucial role for the high affinity binding to codakine. From the glycan array data, the presence of these residues of the core is strictly needed. However, no direct contact is observed between the two GlcNAc and the protein. Interestingly, for DC-SIGN, a similar effect is observed since Man9GlcNAc2 has a 2-3-fold increase in affinity when compared with Man9. It was proposed that the inner GlcNAc residues
6
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

restrict the conformation of nearby sugar groups, resulting in lower entropy of binding. This hypothesis is confirmed by our work since the biantennary glycan with the chitobiose core has a significantly lower entropy barrier (-8 kJ/mol) than the trimannose compound (-18 kJ/mol). This is also correlated with the structural data since the GlcNAc residue linked to βMan establishes one hydrogen bond to the αMan residue on the 1-6 arm, which should significantly stabilize this otherwise very flexible linkage. StructureFunction Relationship in Invertebrate Lectins
Codakine is the first mannose-specific invertebrate lectin to be structurally characterized. Crystal structures were previously obtained for tunicate (Polyandrocarpa misakiensis) lectin TC14 complexed with galactose (50) and sea cucumber (Cucumaria echinata) CEL-I lectin complexed with N-acetyl-galactosamine (GalNAc) (53). In all cases, the lectins adopt a dimeric arrangement allowing for opposite face presentation of carbohydrate binding sites that would be more appropriate for aggregating cells than for avidity binding (Figure 5). All dimers are generated by 2-fold (or pseudo 2-fold) symmetry. TC14 is a non-covalent dimer with interaction through the α2 helix while codakine
and CEL-I dimerization involves an intermolecular disulfide bridge. The biological role and the natural ligands are not yet defined for the three lectins that have been crystallized. Invertebrate lectins are involved in innate immunity and are thought to bind to polysaccharides present at the surface of bacteria and parasites. Recently, the structural basis for the interaction between lung surfactant protein D, a C-type lectin and heptose residues present in bacterial lipopolysaccharide was demonstrated (54). It would be of interest to determine which bacterial LPS fragments could mimic the N-glycan conformation observed in the present structure and which could be recognized by codakine.
Additionally, codakine could play a role in the recognition of symbiotic sulfur-oxidizing bacteria (19). The presentation of opposite face binding sites resulting from dimeric association could result in the cross-linking of microorganisms to the invertebrate cell surface. GlcNAc-terminated biantennary glycans have been identified in invertebrates (55) and codakine would therefore attach to the gill surface through this ligand. Further investigation of the polysaccharides present on the sulfur-oxidizing bacteria should provide important clues regarding the role of invertebrate lectins in symbiotic associations that are often of primary importance in marine organisms.
7
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

REFERENCES
1. Vasta, G. R., Ahmed, H., and Odom, E. W. (2004) Curr. Opin. Struct. Biol. 14, 617-630 2. Vasta, G. R., Ahmed, H., Fink, N. E., Elola, M. T., Marsh, A. G., Snowden, A., and Odom, E.
W. (1994) Ann. NY Acad. Sci. 712, 55-73 3. Muller, W. E., Dorn, A., and Uhlenbruck, G. (1985) Acta Histochem. Suppl. 31, 37-46 4. Moy, G. W., Springer, S. A., Adams, S. L., Swanson, W. J., and Vacquier, V. D. (2008) Proc.
Natl. Acad. Sci. USA 105(6), 1993-1998 5. Jimbo, M., Koike, K., Sakai, R., Muramoto, K., and Kamiya, H. (2005) Biochem. Biophys.
Res. Commun. 330(1), 157-162 6. Bulgheresi, S., Schabussova, I., Chen, T., Mullin, N. P., Maizels, R. M., and Ott, J. A. (2006)
Appl. Environ. Microbiol. 72(4), 2950-2956 7. Iwanaga, S., and Lee, B. L. (2005) J. Biochem. Mol. Biol. 38(2), 128-150 8. Vasta, G. R. (1992) Invertebrate lectins: Distribution, synthesis, molecular biology and
function. In: Allen, H. J., and Kisailus, E. C. (eds). Glycoconjugates, Composition, Structure and Function, Marcel Dekker, New York
9. Minamikawa, M., Hine, M., Russell, S., Huber, P., Duignan, P., and Lumsden, J. S. (2004) Fish Shellfish Immunol. 17(5), 463-476
10. Yamaura, K., Takahashi, K. G., and Suzuki, T. (2008) Comp. Biochem. Physiol. B Biochem. Mol. Biol. 149(1), 168-175
11. Wang, H., Song, L., Li, C., Zhao, J., Zhang, H., Ni, D., and Xu, W. (2007) Mol. Immunol. 44(5), 722-731
12. Bulgakov, A. A., Park, K.-I., Choi, K.-S., Lim, H.-K., and Cho, M. (2004) Fish & Shellfish Immunology 16(4), 487-499
13. Gourdine, J.-P., and Smith-Ravin, E. J. (2007) Fish & Shellfish Immunology 22(5), 498-509 14. Drickamer, K. (1996) Biochem. Soc .Trans. 24(1), 146-150 15. Weis, W. I., Kahn, R., Fourme, R., Drickamer, K., and Hendrickson, W. A. (1991) Science
254(5038), 1608-1615 16. Drickamer, K. (1999) Curr. Opin. Struct. Biol. 9, 585-590 17. Gourdine, J.-P., and Smith-Ravin, E. J. (2002) Prep. Biochem. Biotechnol. 32(4), 341-353 18. Gourdine, J.-P., Markiv, A., and Smith-Ravin, J. (2007) Fish & Shellfish Immunology 23(4),
831-839 19. Berg, C. J., and Alatalo, P. (1984) Aquaculture 39(1-4), 165-179 20. Laemmli, U. K. (1970) Nature 227, 680-685 21. Blixt, O., Head, S., Mondala, T., Scanlan, C., Huflejt, M. E., Alvarez, R., Bryan, M. C.,
Fazio, F., Calarese, D., Stevens, J., Razi, N., Stevens, D. J., Skehel, J. J., van Die, I., Burton, D. R., Wilson, I. A., Cummings, R., Bovin, N., Wong, C. H., and Paulson, J. C. (2004) Proc. Natl. Acad. Sci. USA 101(49), 17033-17038
22. Kajihara, Y., Suzuki, Y., Yamamoto, N., Sasaki, K., Sakakibara, T., and Juneja, L. R. (2004) Chemistry 10, 971-985
23. Seko, A., Kotetsu, M., Nishizono, M., Enoki, Y., H. R. Ibrahim, Juneja, L. R., Kim, M., and Yamamoto, T. (1997) Biochim. Biophys. Acta 1335, 23– 32
24. Wiseman, T., Williston, S., Brandts, J. F., and Lin, L. N. (1989) Anal. Biochem. 179(1), 131-137
25. Leslie, A. G. W. (1992) Joint CCP4 + ESF-EAMCB Newsletter on Protein Crystallography 26
26. number4, C. c. p. (1994) Acta Crystallogr. (D50), 760-763 27. Graves, B. J., Crowther, R. L., Chandran, C., Rumberger, J. M., Li, S., Huang, K. S., Presky,
D. H., Familletti, P. C., Wolitzky, B. A., and Burns, D. K. (1994) Nature 367, 532-538 28. Gerbaud, V., Pignol, D., Loret, E., Bertrand, J. A., Berland, Y., Fontecilla-Camps, J. C.,
Canselier, J. P., Gabas, N., and Verdier, J. M. (2000) J. Biol. Chem. 275(2), 1057-1064 29. Kastrup, J. S., Nielsen, B. B., Rasmussen, H., Holtet, T. L., Graversen, J. H., Etzerodt, M.,
Thogersen, H. C., and Larsen, I. K. (1998) Acta Cryst. D Biol. Cryst. 54(Pt 5), 757-766
8
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

30. Gronwald, W., Loewen, M. C., Lix, B., Daugulis, A. J., Sonnichsen, F. D., Davies, P. L., and Sykes, B. D. (1998) Biochemistry 37(14), 4712-4721
31. McCoy, A. J., Grosse-Kunstleve, R. W., Adams, P. D., Win, M. D., Storoni, L. C., and Read., R. J. (2007) J. Appl. Cryst. 40, 658-674
32. Foadi, J., Woolfson, M. M., Dodson, E. J., Wilson, K. S., Jia-xing, Y., and Chao-de, Z. (2000) Acta Crystallogr. D Biol. Crystallogr. 56(Pt 9), 1137-1147
33. Perrakis, A., Morris, R., and Lamzin, V. S. (1999) Nat. Struct. Biol. 6(5), 458-463. 34. Vagin, A., and Teplyakov, A. (1997) J. Appl. Cryst. 30, 1022-1025 35. Brünger, A. (1992) X-PLOR Version 3.1: A System for Crystallography and NMR. In., Yale
University, New Haven, CT 36. Emsley, P., and Cowtan, K. (2004) Acta Crystallogr. D Biol. Crystallogr. 60, 2126-2132 37. Murshudov, G. N., A.A.Vagin, and E.J.Dodson. (1997) Acta Crystallogr. D53, 240-255 38. Vriend, G. (1990) J. Mol. Graph. 8, 52-56 39. Krissinel, E., and Henrick, K. (2007) J. Mol. Biol. 372, 774-797 40. Drickamer, K. (1993) Curr. Opin. Struct. Biol. 3(1), 393-400 41. Day, A. J. (1994) Biochemical Society transactions 22(1), 83-88 42. Dam, T. K., and Brewer, C. F. (2002) Chem. Rev. 102(2), 387-429. 43. Feinberg, H., Mitchell, D., Drickamer, K., and Weis, W. (2001) Science 294(5549), 2163-
2166 44. Imberty, A., Delage, M. M., Bourne, Y., Cambillau, C., and Pérez, S. (1991) Glyconconj. J. 8,
456-483 45. Imberty, A., Gerber, S., Tran, V., and Pérez, S. (1990) Glycoconj. J. 7, 27-54 46. Krissinel, E., and Henrick, K. (2004) Acta Cryst. D 60, 2256-2268 47. Feinberg, H., Taylor, M. E., and Weis, W. I. (2007) J. Biol. Chem. 282, 17250-17258 48. Chatwell, L., Holla, A., Kaufer, B. B., and Skerra, A. (2008) Mol. Immunol. 45, 1981-1994 49. Quesenberry, M. S., Lee, R. T., and Lee, Y. C. (1997) Biochemistry 36(9), 2724-2732 50. Poget, S. F., Legge, G. B., Proctor, M. R., Butler, P. J., Bycroft, M., and Williams, R. L.
(1999) J Mol Biol 290(4), 867-879 51. Jacob, G. S., Kirmaier, C., Abbas, S. Z., Howard, S. C., Steininger, C. N., Welply, J. K., and
Scudder, P. (1995) Biochemistry 34(4), 1210-1217 52. Feinberg, H., Castelli, R., Drickamer, K., Seeberger, P. H., and Weis, W. I. (2007) J Biol
Chem 282(6), 4202-4209 53. Sugawara, H., Kusunoki, M., Kurisu, G., Fujimoto, T., Aoyagi, H., and Hatakeyama, T.
(2004) J. Biol. Chem., 43, 45219-45225 54. Wang, H., Head, J., Kosma, P., Brade, H., Muller-Loennies, S., Sheikh, S., McDonald, B.,
Smith, K., Cafarella, T., Seaton, B., and Crouch, E. (2008) Biochemistry 47(2), 710-720 55. Gutternigg, M., Burgmayr, S., Poltl, G., Rudolf, J., and Staudacher, E. (2007) Glycoconj. J.
24(8), 475-489 56. Baker, N. A., Sept, D., Joseph, S., Holst, M. J., and Mc Cammon, J. A. Proc. Natl. Acad. Sci.
USA 98, 10037-10041
9
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FOOTNOTES
1 Present affiliation : Institut de Biologie Structurale, Grenoble, France 2 Abbreviations : GlcNAc, N-acetyl-glucosamine ; GalNAc, N-acetyl-galactosamine ; Man, Mannose; αMeMan, α−methyl-mannoside ; αMeFuc, α-methyl-fucoside ; EDTA, Ethylenediaminetetraacetate ; ITC, Isothermal titration microcalorimetry ; nona-Asn : asparagine-linked complex type nonasaccharide ; CRD, carbohydrate recognition domain ; MBP, mannose binding protein ; CTLD, C-type lectin like domain. 3Coordinates and structure factors have been deposited in the Protein Data Bank with accession Code 2vuv and 2vuz for native and ligand-bound codakine, respectively S The on-line version of this article (available at http://www.jbc.org) contains supplemental Figure and Table
ACKNOWLEDGMENTS
This work was supported by CNRS and French Ministry of Research. The glycan microarray analysis was provided by the Consortium for Functional Glycomics funded by the National Institute of General Medical Sciences grant GM62116. We thank the ESRF, Grenoble, for access to synchrotron data collection facilities.
10
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FIGURE LEGENDS
Figure 1. Crystal structure of codakine. A: Graphical representation of the codakine dimer with disulfide bonds displayed as orange sticks. B: Superimposition of the CRD of codakine (green) and DC-SIGN (blue). C: Glycerol in the binding site with labeling of amino acid of interest. D: Sequence alignment of codakine and DC-SIGN. Amino acids involved in monosaccharide binding are labeled with a blue star and intramolecular disulfide bridges are shown with orange lines. Figure 2. Glycan array analysis of codakine as measured by fluorescence intensity. Figure 3. Titration calorimetry results of codakine with oligosaccharides. A: Trimannoside (5 mM) in codakine (500 μM). B: nona-Asn (0.37 mM) in cokakine (37 μM). Top, data from 30 automatic injections of 10 μl oligosaccharide into the codakine-containing cell. Lower, plot of the total heat released as a function of ligand concentration for the titration shown above (squares). The solid line represents the best least-square fit for the obtained data. Figure 4. Crystal structure of codakine complexed with nona-Asn. A: Representation of the final maximum-likelihood weighted 2mFo-DFc electron density map (contoured at 1.0σ, 0.34 eÅ-3 for the nona-Asn oligosaccharide. B. Hydrogen bond network between oligosaccharide and codakine amino acids. Intramolecular hydrogen bonds of the nonasaccharide are represented by blue dash lines. C: Comparison of oligosaccharide binding site in codakine (left) and DC-SIGN (right)(43). The star indicates the position of the calcium ion. Electrostatic surface calculated with APBS (56). Figure 5. Comparison of dimerization modes for invertebrate lectins. Calcium ions are represented as dark spheres and intermolecular disulfide bridges, when present, are colored in black. A. Codakine complexed with Asn-nona. B. Tunicate (Polyandrocarpa misakiensis) lectin TL14 complexed with galactose (50). C. Sea cucumber (Cucumaria echinata) CEL-I lectin complexed with N-acetyl-galactosamine (53).
11
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Table 1. Data collection and refinement statistics.
Native nona-Asn Data collection statistics
Unit cell (Å) a = 82.91 b = 30.39 c = 67.09
β = 133.86
a=32.16 b=100.19 c=95.74
Beamline ID14-1 ID29 Spacegroup C2 C2221
Wavelength (Å) 0.931 0.976 Resolution limits (Å) 33.11-1.30 (1.37-1.30) 34.61-1.70 (1.74-1.70)
Total observations 87473 (4789)* 96802 (6992) Unique reflections 26746 (2230) 17204 (1232)
Completeness 89.7 (52.5) 98.6 (99.3) Multiplicity 3.3 (2.1) 5.6 (5.6)
<I>/<σI> 12.0 (2.1) 5.3 (2.0) Rmerge (%) 3.4 (32.3) 8.3 (37.7)
Wilson B-factor (Å2) 10.4 19.70 Refinement statistics
Used reflections 25381 16302 Rcryst 14.4 20.1 Rfree 16.1 25.8
RMS bonds 0.019 0.018 RMS angles 1.724 1.763
Cruickshank’s DPI 0.05 0.11 Protein atoms 1057 1028 Solvent atoms 176 121
Other atoms 26 109 B-factors
Overall 12.3 23.6 Protein atoms 10.1 21.3 Solvent atoms 25.6 33.7
Other atoms 16.0 34.9
PDB code 2VUV 2VUZ
* Values in parenthesis refer to the highest resolution shell. Rmerge = ∑|I−‹I›|/|∑‹I›| Rcryst = (∑||Fobs – Fcalc||)/(∑||Fobs||)
12
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Table 2 . Thermodynamic of the binding of codakine to carbohydrates. Standard deviations are in the range of 10%.
Sugar n Ka [×104 M-1] Kd [μM] ΔG [kJ/mol] ΔH [kJ/mol] -TΔS [kJ/mol] Mannose 1a 0 .37 270 -20.3 -18.7 -1.6 GlcNAc 1a 0.21 465 -19.0 -14.6 -4.4 αMeMan 1a 1.93 52 -24.5 -33.6 9.1 αMeFuc 1a 1.67 60 -24.1 -20.3 -3.8
Trimannnoside 0.9 1.27 80 -23.4 -41.9 18.5 nona-Asn 0.94 232 0.432 -36.2 -45.3 8.9
a Fixed value during fitting procedure
13
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Table 3. List of contact between nona-Asn and the proteins Atom 1 Atom 2 Distance (Ǻ) Buried prot.
surf.(Ǻ2) 1-6 branch – Man7 133 Man7.O3 Calcium 2.48 Glu93.OE2 2.82 Glu93.OE1 3.19 Asn95.ND2 2.78 Asn95.OD1 3.03 Asn112.OD1 3.33 GlcNAc8 O5 3.16 Man7.O4 Calcium 2.64 Glu101.OE2 2.48 Asp113.O 2.85 Asp112.ND2 2.98 Asn112.OD1 3.37 Man7.O6 Arg115.NH1 3.19 1-6 branch – GlcNAc8 106 GlcNAc8.O5 Ans95.ND2 2.84 GlcNAc8.O6 Glu93.OE2 2.83 GlcNAc8.O7 Ser97.OG 2.49 1-3 branch – Man4 38 Man4.O3 GlcNAc5 O5 3.24 1-3 branch – GlcNAc5 116 GlcNAc5 Trp108 stacking GlcNAc5.O6 Man3 O4 Water bridged Core – Man3 86 Man3.O4 Trp108.NE1 2.92 Asn107.ND2 2.92 Man3.O6 Asn107.ND2 3.30 Man3 O4 2.8 Core – GlcNAc2 37 GlcNAc2.O3 Man3.O5 2.83 GlcNAc2.O3 Man7 O5 3.26 Core – GlcNAc1 - GlcNAc1.O3 GlcNAc2 O5 2.88
14
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
O
O
OO O
OH
HN
HONHA c
OH
NHA cHO
OH
OHO
O
OH
NHA cHOO
O
OHO
HOHO
O
O
O
OH
OHHO
HO
O
OH
NHA c
HOOO
OHO
HOHOO
OH
OHHO
HO
HN
OH
O
O
O
1. NaOH , H2O/MeOH2. gel filtration (Superdex 30)
O
O
OO O
OH
HN
HONHA c
OH
NHA cHO
OH
OHO
O
OH
NHA cHOO
O
OHO
HOHO
O
O
O
OH
OHHO
HO
O
OH
NHA c
HOOO
OHO
HOHOO
OH
OHHO
HO
H2NOH
O
1
Fmoc-1
GlcNAc1
GlcNAc2
GlcNAc8
GlcNAc5Man3
Gal9
Gal6
Man7
Man4
15
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D
B
A
Figure 1
N95
N107
H105N112
E101
Hyp91
D113
E93
C
16
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
0
5000
10000
15000
20000
25000
30000
35000
40000
45000
1 21 41 61 81 101 121 141 161 181 201 221 241 261 281 301
Glycan Number
Rel
ativ
e Fl
uore
scen
ce U
nits
Figure 2
17
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
0.0 0.5 1.0 1.5 2.0 2.5-50
-40
-30
-20
-10
0
-60
-50
-40
-30
-20
-10
0
0 20 40 60 80 100 120 140 160Time (min)
���
��
Molar Ratio
��
���
����
��
���
�
µJ sec -1
kJ mole-1
0.0 0.5 1.0 1.5 2.0 2.5Molar Ratio
- 0
-40
-30
-20
-10
0
-5
-4
-3
-2
-1
0
���
��
��
���
����
��
���
�
20 40 60 80 100 120 140 1600
5
Time (min)A B
Figure 3
18
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

BA
C
Figure 4
**
19
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

C
B
A
Figure 5
20
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Anne ImbertyVaron Silva, Annabelle Varrot, Catherine Gautier, Emilie Juliette Smith-Ravin and Jean-Philippe Gourdine, Gianluca Cioci, Laurence Miguet, Carlo Unverzagt, Daniel
complex-type N-glycan revealed by crystallography and microcalorimetryHigh affinity interaction between a bivalve C-type lectin and a biantennary
published online August 7, 2008J. Biol. Chem.
10.1074/jbc.M804353200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2008/08/08/M804353200.DC1
by guest on August 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

O
O
OO O
OH
HN
HONHA c
OH
NHA cHO
OH
OHO
O
OH
NHA cHOO
O
OHO
HOHO
O
O
O
OH
OHHO
HO
O
OH
NHA c
HOOO
OHO
HOHOO
OH
OHHO
HO
HN
OH
O
O
O
1. NaOH , H2O/MeOH2. gel filtration (Superdex 30)
O
O
OO O
OH
HN
HONHA c
OH
NHA cHO
OH
OHO
O
OH
NHA cHOO
O
OHO
HOHO
O
O
O
OH
OHHO
HO
O
OH
NHA c
HOOO
OHO
HOHOO
OH
OHHO
HO
H2NOH
O
1
Fmoc-1
GlcNAc1
GlcNAc2
GlcNAc8
GlcNAc5Man3
Gal9
Gal6
Man7
Man4

D
B
A
Figure 1
N95
N107
H105N112
E101
Hyp91
D113
E93
C

0
5000
10000
15000
20000
25000
30000
35000
40000
45000
1 21 41 61 81 101 121 141 161 181 201 221 241 261 281 301
Glycan Number
Rel
ativ
e Fl
uore
scen
ce U
nits
Figure 2

0.0 0.5 1.0 1.5 2.0 2.5-50
-40
-30
-20
-10
0
-60
-50
-40
-30
-20
-10
0
0 20 40 60 80 100 120 140 160Time (min)
���
��
Molar Ratio
��
���
����
��
���
�
µJ sec -1
kJ mole-1
0.0 0.5 1.0 1.5 2.0 2.5Molar Ratio
- 0
-40
-30
-20
-10
0
-5
-4
-3
-2
-1
0
���
��
��
���
����
��
���
�
20 40 60 80 100 120 140 1600
5
Time (min)A B
Figure 3

BA
C
Figure 4
**

C
B
A
Figure 5