histamine and h3 receptor in alcohol-related behaviors
TRANSCRIPT

JPET#170928 1
Histamine and H3 receptor in alcohol-related behaviors*
Pertti Panula and Saara Nuutinen
Neuroscience Center and Institute of Biomedicine/Anatomy (PP, SN), University of Helsinki,
Haartmaninkatu 8, 00290 Helsinki. Finland
JPET Fast Forward. Published on September 23, 2010 as DOI:10.1124/jpet.110.170928
Copyright 2010 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 2
Running title: Histamine, H3 receptor and alcohol addiction
Corresponding author:
Pertti Panula
Neuroscience Center and Institute of Biomedicine/Anatomy
POB 63
00014 University of Helsinki
Finland
Tel: +358919125263
FAX: +358919125261
E-mail: [email protected]
The number of text pages: 24
The number of tables: 3
The number of figures: 3
The number of references: 76
The number of words in the Abstract: 249
The number of words in the Introduction: N/A
The number of words in the Discussion: N/A
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 3
A list of nonstandard abbreviations used in the paper
AA Alko Alcohol
ANA Alko Non-Alcohol
CPP conditioned place preference
Ctx cortex
DARPP-32 dopamine, cAMP-regulated phosphoprotein of 32,000 kDa
DID Drinking in the dark
HAP high alcohol-preferring
HDC Histidine decarboxylase
H1R Histamine H1 receptor
H2R Histamine H2 receptor
H3R Histamine H3 receptor
Hypot hypothalamus
LAP low alcohol-preferring
MHPG 3-methoxy-4-hydroxy-phenylglycol
NAcc nucleus accumbens
PFC prefrontal cortex
SD Sprague-Dawley
Thal thalamus
VP ventral pallidum
WT wild type
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 4
Abstract
Data from rat models for alcohol preference and histidine decarboxylase knockout (HDC KO) mice
suggest that brain histamine regulates alcohol-related behaviors. Histamine levels are higher in alcohol-
preferring than alcohol non-preferring rat brains, and expression of histamine H3 receptor is different in
key areas for addictive behavior. Histamine H3 receptor inverse agonists decrease alcohol responding in
one alcohol-preferring rat line. Conditioned place preference induced by alcohol is stronger in HDC KO
mice than control mice. The HDC KO mice display a weaker stimulatory response to acute alcohol than
the wild type (WT) mice. In male inbred C57BL/6 mice H3 receptor antagonist ciproxifan inhibits the
ethanol-evoked stimulation of locomotor activity. Ciproxifan also potentiates the ethanol reward, , but
does not alone result in the development of place preference. At least in one rat model developed to
study alcohol sensitivity, high histamine levels are characteristic of the alcohol insensitive rat line, and
lowering brain histamine with a HDC inhibitor increases alcohol sensitivity in the tilting plane test.
However, the motor skills of HDC KO mice do not appear to differ from those of the WT mice.
Current evidence suggests that the histaminergic system is involved in regulation of place preference
behavior triggered by alcohol, possibly through an interaction with the mesolimbic dopamine system.
Histamine may also interact with dopamine in regulation of the cortico-striato-pallido-thalamo-cortical
motor pathway and cerebellar mechanisms, which may be important in different motor behaviors
beyond alcohol-induced motor disturbances. Histamine H3 receptor ligands may have significant effects
on alcohol addiction.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 5
Rat models of alcohol-related behaviors
Several inbred and outbred rat lines with different alcohol preference have been developed and reported
(for a review, see Sinclair et al., 1989;Sommer et al., 2006). These include the high (HAP) and low
(LAP) alcohol-preferring (Kitanaka et al., 2004;Kitanaka et al., 2004), inbred alcohol-preferring (iP) and
non-preferring (iNP)(McBride et al., 2010) and outbred Alko alcohol (AA) and Alko, non alcohol
(ANA) rats (Sommer et al., 2006;Sinclair et al., 1989). A summary of rat and mouse models used in
alcohol research is shown in Table 1.
The AA (Alko, alcohol) and ANA (Alko, non-alcohol) rat lines were among the first lines produced
using a bidirectional selection method (Eriksson, 1968) for alcohol preference, and these rats have now
been maintained beyond the 100th generation (Sommer et al., 2006). The AA rats have higher levels of
dopamine in several brain regions including striatum and limbic forebrain than the ANA rats (Ahtee and
Eriksson, 1975), and tyrosine hydroxylase activity is also 42% higher in AA than ANA rats (Pispa et al.,
1986). The levels of noradrenaline are also higher in AA rats than in ANA rats in the cortex and limbic
areas (Ahtee and Eriksson, 1975). Noradrenaline turnover also appears higher in AA rats, as the levels of
3-methoxy-4-hydroxy-phenylglycol (MHPG) are higher in AA rats (Sommer et al., 2006). However,
there is no difference in the alcohol-induced release of dopamine between the lines, so that the
sensitivity to ethanol is not the major difference between the lines (Sommer et al., 2006). Serotonin
levels are higher in AA rats than in ANA rats in all brain regions studied (Ahtee and Eriksson, 1972),
but the metabolite levels are normal. Thus, there are differences in major catechol- and indolamine
neurotransmitter systems between the lines.
Histamine levels in AA rats are 20-170% higher in different brain regions in AA than in ANA rat brain
(Lintunen et al., 2001), Interestingly, in almost all brain areas histamine levels in Sprague-Dawley (SD)
rats are between the values seen in AA and ANA rats, suggesting correlation of histamine levels to
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 6
alcohol preference. Levels of tele-methylhistamine are generally 30-70% higher in AA than ANA rats,
with pons as the only exception where no difference is seen (Lintunen et al., 2001). Thus, there is a
distinct difference in histamine turnover between the rat lines. Significantly higher histamine and tele-
methylhistamine concentrations were found in several brain regions relevant for the alcohol preference,
including the frontal cortex, striatum, septum, hypothalamus and hippocampus. Immunohistochemical
analysis showed that this increased histamine resided in nerve fibers rather than mast cells. Higher
histamine-immunoreactive fiber densities in AA rats were found in e.g. medial and lateral septum,
nucleus accumbens and medial preoptic nucleus (Lintunen et al., 2001) (Fig. 1). Generally, histamine
levels in different rat strains are variable due to different numbers of mast cells found predominantly in
the thalamus and median eminence. In agreement with this concept, histamine levels in the thalamus of
SD rats were significantly higher than in either AA or ANA rats, whereas the levels in e.g.
hypothalamus, septum and hippocampus are about twice as high in AA as in ANA or SD rat brain
(Lintunen et al., 2001). Histamine H3 receptor regulates histamine synthesis and release (Arrang et al.,
1983), and in rat brain the different isoforms of this receptors are also differentially expressed (Drutel et
al., 2001). Due to its function, possible changes in H3R expression and/or receptor radiolingand binding
may be due to a primary difference in the histamine. On the other hand, primary changes in receptor
regulation may lie behind changes in histamine turnover. In AA rats, a distinct statistically significant
difference to ANA rats in H3R radioligand binding was found in motor cortex, nucleus accumbens and
CA1 area of the hippocampus, all areas which may be of relevance for the behavioral phenotype. H3R
radioligand binding was lower in AA rats than in ANA rats in only these areas, whereas there was no
difference in e.g. lateral septum or tuberomamillary nucleus (Lintunen et al., 2001). These differences
suggest no direct correlation between histamine levels and H3R radiolingand binding. Thus, the
observed differences in receptor binding are not merely due to higher histamine levels and release in
these areas, but may depend on other factors which are potentially important for the behavioral
differences. Interestingly, H1R expression was generally significantly lower or showed a tendency to
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 7
lower values in AA rats in all brain areas (Lintunen et al., 2001). This can be interpreted as a
downregulation due to high histamine level and release. Behaviorally, histamine H1R antagonist
mepyramine did not affect ethanol self-administration in AA rats, whereas two H3R inverse agonists,
thioperamide and clobenpropit, significantly and in a dose-dependent manner reduced self-
administration of ethanol (Lintunen et al., 2001). These differences in histaminergic system function
have not been tested similarly in the other rat line models of high/low alcohol consumption. However,
the histamine-stimulated phosphoinositide hydrolysis in the cerebral cortex is significantly lower in high
alcohol preferring HAP than in LAP rats with low alcohol preference (Takemura et al., 2003). Although
the histamine levels were not significantly different in that study, the mean of histamine concentration in
the cortex was more than twice as high as in LAP rats, and the lack of significance may be due to the
low number of experimental animals (n=3) (Kitanaka et al., 2004). The μ-opioid receptor antagonist
CTOP injected into the amygdala, and δ-opioid receptor antagonist naltrindole administered into the
nucleus accumbens or basolateral amygdala also decrease ethanol responding in a two-lever operant
system in both AA and Wistar rats (Hyytia, 1993;Hyytia and Kiianmaa, 2001). AA rats also show lower
spontaneous release of beta-endorphin in the hypothalamus (de Waele et al., 1994;Nylander et al., 1994)
and lower levels of proenkephalin peptides in the nucleus accumbens and prodynorphin peptides in the
VTA than ANA rats. Differences in pro-opiomelanocortin expression have been reported in both AA
and ANA rats (Gianoulakis et al., 1992) and HAP/LAP rats (Kinoshita et al., 2004), although in
somewhat different brain regions, which suggest that the same mediators are at least common
phenotypic signs of the selectively bred animals.
Rat lines have also been developed to study sensitivity to a moderate dose (2 g/kg) of alcohol (Rusi et
al., 1977), and in these alcohol-sensitive (alcohol non-tolerant, ANT) and alcohol-insensitive (alcohol
tolerant, AT) rats distinct differences have been observed in histamine levels, H1R mRNA expression,
H3R radioligand binding and H3R agonist-induced G protein activation (Lintunen et al., 2002). The
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 8
alcohol-sensitive rats have significantly reduced histamine levels in frontal cortex, septum,
hypothalamus and hippocampus, and increased H1R mRNA expression in several brain regions. In these
rats, brain histamine may be causally linked to the differences in alcohol sensitivity. Administration of
α-fluoromethylhistidine, a suicide inhibitor of HDC, induces a decline in both brain histamine levels and
motor coordination as measured on a tilting plane in alcohol-insensitive AT rats, which have high brain
histamine levels (Lintunen et al., 2002). Both histamine and immepip-evoked GTPγ[35S] binding is
higher in alcohol sensitive ANT rats in primary motor cortex, insula and caudate putamen (Lintunen et
al., 2002). These regions are important in motor disorders like Tourette’s syndrome, where functional
imaging has revealed abnormalities in a network involving e.g. prefrontal cortex, insula, caudate,
premotor and primary motor cortex (Stern et al., 2000). Tourette’s syndrome is also characterized by
increased dopamine release following amphetamine challenge both in striatal (Singer et al., 2002) and
extrastiatal (Steeves et al., 2010) sites. It is possible that histamine/dopamine interactions are widely
important in several motor disorders, as also in both Parkinson’s disease and experimental rat models the
H3R expression and radioligand binding are altered in substantia nigra and caudate putamen (Anichtchik
et al., 2000b;Anichtchik et al., 2000a;Anichtchik et al., 2001;Ryu et al., 1994). Thus, interactions of
histamine and dopamine are likely to span a wide range of disorders ranging from motor system to
addiction.
The alcohol-induced motor incoordination may also depend on cerebellar mechanisms. Indeed, the
diazepam-insensitive binding of the benzodiazepine [3H]Ro15-4513 is lacking in the granule cells of the
ANA rats, a phenomenon that is caused by a point mutation in the alpha6 subunit of the GABAA
receptor (Korpi et al., 1993). This mutation renders the alpha subunit benzodiazepine sensitive (Korpi et
al., 1993), suggesting that the altered GABAergic signaling may contribute to the behavior of the ANA
rats. The cerebellum receives a direct hypothalamocerebellar input from the histaminergic TMN neurons
both in rodents and humans (Panula et al., 1993;Panula et al., 1989), and fibers pass through the granule
cell layer and Purkinje cell layer perdendicularly, to then turn 90 degrees to make contact with Purkinje
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 9
cell dendrites (Panula et al., 1993). Thus, both extensive systems (basal ganglia and cerebellum) which
control the upper motor neuron functions are modulated by the histaminergic system in a manner that
may contribute to alcohol dependent motor incoordination.
Mouse strain properties and differences, methods used to study, pros and cons
Alcohol has strong behavioral effects in mice of which most studied are the reinforcing and rewarding
effects and the stimulation of locomotor activity. In addition, the motor impairing effect of alcohol is
well-known and can be studied in mouse behavioral models. The reinforcing and rewarding properties of
alcohol are studied using self-administration paradigms or conditioned place preference. Even though
both of these methods are thought to measure reinforcement it is possible that they do not measure
exactly same aspects of alcohol reinforcement, and the brain areas involved can be different (Green and
Grahame, 2008). Thus, a combination of the two methods is advisable. Self-administration of alcohol
has been largely done using a two-bottle choice test where mice have a chance to choose between water
or alcohol solution. However, most mouse strains do not drink alcohol initially at concentrations that are
pharmacologically relevant (Sanchis-Segura and Spanagel, 2006). Thus, procedures to train mice to
drink have been developed, such as ascending concentrations of alcohol and addition of a sweet flavor
agent which can progressively fade out or not. The recently described Drinking in the dark –method
(DID) (Rhodes et al., 2005) offers a faster option to screen the effects of drugs on alcohol drinking. The
DID method is based on the high voluntary alcohol consumption of C57BL/6J mice during their active
period of day and has received a lot of interest among alcohol researches as model for binge drinking.
Conditioned place preference (CPP) is a form of Pavlovian conditioning where environmental cues
become associated with alcohol administration (Tzschentke, 2007). CPP is routinely used to measure the
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 10
reinforcing and rewarding or aversive effects of drugs. The use of CPP in alcohol studies requires
careful planning of the study design since the alcohol dose, the length of the conditioning period and the
use of a biased vs. an unbiased protocol might lead to unexpected or misleading results (Cunningham et
al., 2006). The mice of the DBA/2J strain show strong alcohol CPP and are commonly used whereas
C57BL/6J mice do not develop alcohol-evoked CPP (Cunningham and Noble, 1992) or it is rather weak
(Gremel et al., 2006). Examples of different mouse strains used in alcohol studies are shown in Table 1.
Modifications of CPP and operant alcohol self-administration paradigms are used when other addictive
features beyond reinforcement, such as alcohol seeking and relapse are the focus of research.
The stimulatory effect of alcohol can be detected by measuring the horizontal locomotor activity in
response to alcohol administration. Typical to alcohol stimulation is that it occurs only with fairly low
doses and is rather short-lived. Sensitivity of different mice strains towards stimulatory effect of alcohol
vary to a great extent, the DBA/2 and FVB strains being most stimulated by alcohol and 129/Sv and
C57BL/6 strains showing moderate to complete loss of activation by alcohol (Crabbe et al., 1994)
(Crabbe et al. 1994; Crabbe et al. 2005). In a repeated alcohol administration some strains, but not all,
develop sensitization to the stimulatory effect of alcohol- a phenomenon that is commonly seen with
psychostimulants such as amphetamine and cocaine. The locomotor stimulation and the development of
psychomotor sensitization have been suggested to predict the addictive property of a drug (Wise and
Bozarth, 1987;Robinson and Berridge, 1993) respectively. However, these theories are vividly discussed
(Sanchis-Segura and Spanagel, 2006) and many studies including ours (Nuutinen et al., 2010) do not
support this view. Acute sedative effects of alcohol can be examined with various behavioral tests of
which rotating rod test is the most widely used. However, this test has been criticized to its lack of
sensitivity (Stanley et al., 2005). Different mouse strains differ in their sensitivity for ethanol also in
rotarod task (Rustay et al., 2003;Rustay et al., 2003). A more sensitive task to study alcohol-induced
ataxia and motor incoordination in mice is the balance beam walking test. However, finding the right
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 11
combination of alcohol and size of the beam that shows marked but not too strong impairment by
alcohol can be rather challenging and it is important to measure not only time to cross the beam but
more importantly how many foot slips the mouse makes while walking on the beam. In addition to these,
sedation can be studied using a grid or a dovel test or ataxia can be observer-rated. Very high doses of
alcohol (e.g. 4 g/kg) induce the loss of the righting reflex (LORR) which can be also a marker for
alcohol sedation. However, due to the high dose LORR tests measure different aspects of sedation than
the other motor function tests mentioned above.
Histamine in mouse models of addiction
Histamine has been suggested to have an inhibitory role in reward and reinforcement. This is supported
by the findings that cocaine-induced CPP is attenuated by increasing brain histamine levels with L-
histidine whereas inhibition of histamine synthesis by alpha-fluoromethylhistamine potentiates CPP by
cocaine (Suzuki et al., 1996). In line with this, the CPP induced by ethanol (Nuutinen et al., 2010) and
morphine (Gong et al., 2010) is stronger in HDC KO mice. However, no difference for cocaine reward
in CPP model was found between HDC KO and WT mice (Brabant et al., 2007). This discrepancy could
be explained by the different CPP designs. The cocaine study was carried out using a “biased” CPP
paradigm wheras an “unbiased” design was used for ethanol and morphine. In a biased model animal is
placed in the initially unpreferrred side of the conditioning apparatus following drug administration
(Tzschentke, 2007). In an unbiased paradigm, conditioning apparatus is designed so that there is no
initial preference for one of the conditioning compartments. The advantage of unbiased conditioning is
that the possible anxiolytic or anxiogenic effects of the drugs or e.g. anxious phenotypes of mice do not
affect the outcome of the study. Thus, the unbiased design is a more accurate model for measuring
reward than a biased design. Although the CPP responses differ for ethanol and cocaine, the HDC KO
mice show decreased locomotor stimulation in response to ethanol (Nuutinen et al., 2010) and to
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 12
cocaine (Brabant et al., 2007). These findings suggest that histamine is needed for the acute stimulation
by drugs of abuse whereas the reward and reinforcement might be inhibited by neuronal histamine.
Histamine’s inhibitory role in drug reward is further supported by the stronger methamphetamine-
induced CPP in H1 receptor knockout mice (Takino et al., 2009). Interestingly, histamine receptor triple
knockout mice (H1, H2, H3 KO) do not differ from wild type animals in place preference induced by
methamphetamine (Okuda et al., 2009).
Studies using histamine receptor ligands have resulted in variable findings concerning the behavioral
effects of abused drugs. Similar to what was shown in rat with first generation H1R antagonists (Suzuki
et al., 1996), diphenhydramine induces cocaine CPP with a cocaine dose that is not reinforcing alone
(Nguyen et al., 2010) in mice. However, this effect is probably due to the unspecific effect of
diphenhydramine on dopamine transporter (Tanda et al., 2008). Interestingly, a combination of
dihydrocodeine and the second generation antihistamine ebastine was found to evoke place preference
(Kamei et al., 2003) even though ebastine does not affect dopamine uptake (Fujisaki et al., 2002). Also
H2R antagonists can enhance reward in mice as was shown in the study by Suzuki et al. (Suzuki et al.,
1995) with zolantidine in combination with morphine.
Studies concerning the role of H3R in the effects of drugs of abuse are contradictory since
methamphetamine (Clapham and Kilpatrick, 1994;Morisset et al., 2002;Fox et al., 2005) and alcohol
stimulation are decreased by H3R antagonist but cocaine-induced hyperactivity is potentiated by H3R
inactivation (Brabant et al., 2006;Brabant et al., 2009). One reason for the different results can be the
ability of thioperamide to inhibit cocaine metabolism and increase plasma concentrations of cocaine
(Brabant et al., 2009). A CPP study using thioperamide showed that H3R inactivation resulted in
cocaine reward with a low dose of cocaine that is not rewarding alone (Brabant et al., 2006). In
agreement, we showed that ethanol-induced CPP was stronger in response to ciproxifan (H3R
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 13
antagonist/inverse agonist) pretreatment (Nuutinen et al., 2010). In contrast, H3R KO mice are not
different from the wild type mice in the methamphetamine-induced reward although they are less
stimulated by methamphetamine than the control animals (Okuda et al., 2009). Altogether these findings
suggest a role for histamine receptors, especially for H3Rs in the regulation of behavioral effects of
drugs of abuse. The H3R has a multifunctional role which includes the control of not only histamine
release but also the release of several other neurotransmitters and the recently described interaction of
postsynaptic H3Rs with dopamine receptors (Ferrada et al., 2008;Ferrada et al., 2009). Thus, more
studies are needed to clarify the actual role of histamine in addictive behaviors and to what degree the
effects seen with the H3R ligands are due to modulation of e.g. dopamine neurotransmission and
signaling. In this respect, and in chemical nature and pharmacological properties the different H3R
ligands (Table 3) have significantly different properties.
The studies described above have used either CPP paradigm or measured activation of locomotion by
drugs of abuse. Thus, it would be important to get data from self-administration paradigms to better
understand the role of histamine in addiction. Also it is worth noticing that an acute hyperactive
response is not a direct measure of reinforcement and reward drug. Indeed, findings from our laboratory
using HDC KO mice (Nuutinen et al., 2010) and e.g. in DARPP-32, the key dopamine signaling
molecule, knockout mice (Risinger et al., 2001) suggest that alcohol CPP and stimulatory response are
dissociated. Thus a combination of many behavioral methods examining different aspect of
reinforcement and other addictive behaviors are needed.
Are the models relevant for human alcoholism?
The basic structure and organization of the histaminergic system with all neurons in the posterior
hypothalamic tuberomamillary nucleus in human brain is very similar to that of all other studied
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 14
mammals (Airaksinen et al., 1991). Very little is known of the functions of brain histamine in human
alcoholics and other addicts. However, in one study on post-mortem brains of type 1 (late onset, often
females, low degree of association with violence) and type 2 (early onset, often males, high degree of
association with violence) alcoholics histamine levels were significantly higher in cortical grey matter of
type 1 alcoholics than normal control brains (Alakarppa et al., 2002). The levels of the first metabolite,
tele-methylhistamine, were significantly increased in type 2 alcoholics, indicating increased histamine
release and turnover (Alakarppa et al., 2002). This may mean that histamine synthesis and/or
metabolism are primarily altered in alcoholics, or that the possibly associated liver pathology lies behind
the abnormal findings. Indeed, histamine concentration is increased fourfold in the caudate-putamen and
significantly also in cortical regions of hepatic encephalopathy patients, and tele-methylhistamine levels
are also increased suggesting increased histamine turnover, whereas H3R radioligand binding is
decreased (Lozeva et al., 2003). A concomitant increase in densities (Bmax) of H1R in frontal cortex has
also been reported in patients with hepatic encephalopathy (Lozeva et al., 2001). Similar increases in
brain histamine have been reported in portocavally shunted rats (Fogel et al., 2002). Taken together,
changes in brain histamine, H1R and H3R are found in patients with hepatic failure, and these changes
coincide with sleep disturbances, abnormal circadian rhythm and other neuropsychiatric disturbances.
These findings, although they suggest that histaminergic drugs may be useful in treatment of alcohol-
related disorders associated with hepatic failure, do not imply that histamine alone would be necessary
or essential. Alterations in brain circuits containing multiple transmitters relevant for those in alcohol
addiction, including the hypothalamo-cortical or hypothalamo-striatal/accumbeal pathways may be
important.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 15
References
Ahtee L, Eriksson K (1972) 5-Hydroxytryptamine and 5-hydroxyindolylacetic acid content in brain of rat strains
selected for their alcohol intake. Physiol Behav 8: 123-126.
Ahtee L, Eriksson K (1975) Dopamine and noradrenaline content in the brain of rat strains selected for their
alcohol intake. Acta Physiol Scand 93: 563-565.
Airaksinen MS, Paetau A, Paljarvi L, Reinikainen K, Riekkinen P, Suomalainen R, Panula P (1991) Histamine
neurons in human hypothalamus: anatomy in normal and Alzheimer diseased brains. Neuroscience 44: 465-481.
Alakarppa K, Tupala E, Mantere T, Sarkioja T, Rasanen P, Tarhanen J, Tiihonen J, Tuomisto L (2002) Effect of
alcohol abuse on human brain histamine and tele-methylhistamine. Inflamm Res 51 Suppl 1: S40-S41.
Anichtchik OV, Huotari M, Peitsaro N, Haycock JW, Mannisto PT, Panula P (2000a) Modulation of histamine H-
3 receptors in the brain of 6-hydroxydopamine-lesioned rats. European Journal of Neuroscience 12: 3823-3832.
Anichtchik OV, Peitsaro N, Rinne JO, Kalimo H, Panula P (2001) Distribution and modulation of histamine H(3)
receptors in basal ganglia and frontal cortex of healthy controls and patients with Parkinson's disease. Neurobiol
Dis 8: 707-716.
Anichtchik OV, Rinne JO, Kalimo H, Panula P (2000b) An altered histaminergic innervation of the substantia
nigra in Parkinson's disease. Experimental Neurology 163: 20-30.
Arrang JM, Garbarg M, Schwartz JC (1983) Auto-inhibition of brain histamine release mediated by a novel class
(H3) of histamine receptor. Nature 302: 832-837.
Bonaventure P, Letavic M, Dugovic C, Wilson S, Aluisio L, Pudiak C, Lord B, Mazur C, Kamme F, Nishino S,
Carruthers N, Lovenberg T. (2007) Histamine H3 receptor antagonists: From target identification to drug leads.
Biochem Pharmacol 73:1084-1096.
Brabant C, Alleva L, Grisar T, Quertemont E, Lakaye B, Ohtsu H, Lin JS, Jatlow P, Picciotto MR, Tirelli E
(2009) Effects of the H3 receptor inverse agonist thioperamide on cocaine-induced locomotion in mice: role of the
histaminergic system and potential pharmacokinetic interactions. Psychopharmacology (Berl) 202: 673-687.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 16
Brabant C, Quertemont E, Anaclet C, Lin JS, Ohtsu H, Tirelli E (2007) The psychostimulant and rewarding
effects of cocaine in histidine decarboxylase knockout mice do not support the hypothesis of an inhibitory
function of histamine on reward. Psychopharmacology (Berl) 190: 251-263.
Brabant C, Quertemont E, Tirelli E (2006) Effects of the H3-receptor inverse agonist thioperamide on the
psychomotor effects induced by acutely and repeatedly given cocaine in C57BL/6J mice. Pharmacol Biochem
Behav 83: 561-569.
Clapham J, Kilpatrick GJ (1994) Thioperamide, the selective histamine H3 receptor antagonist, attenuates
stimulant-induced locomotor activity in the mouse. Eur J Pharmacol 259: 107-114.
Cowart M, Pratt JK, Stewart AO, Bennani YL, Esbenshade TA, Hancock AA. (2004) A new class of potent non-
imidazole H(3) antagonists: 2-aminoethylbenzofurans Bioorg Med Chem Lett 14:689-693.
Crabbe JC, Gallaher ES, Phillips TJ, Belknap JK (1994) Genetic determinants of sensitivity to ethanol in inbred
mice. Behav Neurosci 108: 186-195.
Crabbe JC, Metten P, Cameron AJ, Wahlsten D. (2005) An analysis of the genetics of alcohol intoxication in
inbred mice. Neurosci Biobehav Rev 28:785-802.
Crabbe JC, Phillips TJ, Belknap JK. (2010) The complexity of alcohol drinking: Studies in rodent genetic models
Behav Genet .In Press.
Cunningham CL, Gremel CM, Groblewski PA (2006) Drug-induced conditioned place preference and aversion in
mice. Nat Protoc 1: 1662-1670.
Cunningham CL, Noble D (1992) Conditioned activation induced by ethanol: role in sensitization and conditioned
place preference. Pharmacol Biochem Behav 43: 307-313.
de Waele JP, Kiianmaa K, Gianoulakis C (1994) Spontaneous and ethanol-stimulated in vitro release of beta-
endorphin by the hypothalamus of AA and ANA rats. Alcohol Clin Exp Res 18: 1468-1473.
Drutel G, Peitsaro N, Karlstedt K, Wieland K, Smit MJ, Timmerman H, Panula P, Leurs R (2001) Identification
of rat H3 receptor isoforms with different brain expression and signaling properties. Mol Pharmacol 59: 1-8.
Eriksson K (1968) Genetic Selection for Voluntary Alcohol Consumption in the Albino Rat. Science 159: 739-
741.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 17
Esbenshade TA, Fox GB, Krueger KM, Miller TR, Kang CH, Denny LI, Witte DG, Yao BB, Pan L, Wetter J,
Marsh K, Bennani YL, Cowart MD, Sullivan JP, Hancock AA. (2005)
Pharmacological properties of ABT-239 [4-(2-{2-[(2R)-2-methylpyrrolidinyl]ethyl}-benzofuran-5-
yl)benzonitrile]: I. potent and selective histamine H3 receptor antagonist with drug-like properties. J Pharmacol
Exp Ther 313:165-175.
Ferrada C, Ferre S, Casado V, Cortes A, Justinova Z, Barnes C, Canela EI, Goldberg SR, Leurs R, Lluis C,
Franco R (2008) Interactions between histamine H3 and dopamine D2 receptors and the implications for striatal
function. Neuropharmacology 55: 190-197.
Ferrada C, Moreno E, Casado V, Bongers G, Cortes A, Mallol J, Canela EI, Leurs R, Ferre S, Lluis C, Franco R
(2009) Marked changes in signal transduction upon heteromerization of dopamine D1 and histamine H3
receptors. Br J Pharmacol 157: 64-75.
Fogel WA, Michelsen KA, Granerus G, Sasiak K, Andrzejewski W, Panula P, Maslinski C (2002) Neuronal
storage of histamine in the brain and tele-methylimidazoleacetic acid excretion in portocaval shunted rats. Journal
of Neurochemistry 80: 375-382.
Fox GB, Esbenshade TA, Pan JB, Browman KE, Zhang M, Ballard ME, Radek RJ, Miner H, Bitner RS, Krueger
KM, Yao BB, Faghih R, Rueter LE, Komater VA, Drescher KU, Buckley MJ, Sullivan JP, Cowart MD, Decker
MW, Hancock AA (2005) Selective H3 receptor (H3R) blockade: broad efficacy in cognition and schizophrenia.
Inflamm Res 54 Suppl 1: S23-S24.
Fujisaki Y, Itoh Y, Oishi R (2002) In vivo evidence for a lack of central effect of ebastine, an antihistaminic
agent, in rats: a microdialysis study. Jpn J Pharmacol 90: 353-356.
Gianoulakis C, de Waele JP, Kiianmaa K (1992) Differences in the brain and pituitary beta-endorphin system
between the alcohol-preferring AA and alcohol-avoiding ANA rats. Alcohol Clin Exp Res 16: 453-459.
Gong YX, Zhang WP, Shou WT, Zhong K, Chen Z (2010) Morphine induces conditioned place preference
behavior in histidine decarboxylase knockout mice. Neurosci Lett 468: 115-119.
Green AS, Grahame NJ (2008) Ethanol drinking in rodents: is free-choice drinking related to the reinforcing
effects of ethanol? Alcohol 42: 1-11.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 18
Gremel CM, Gabriel KI, Cunningham CL (2006) Topiramate does not affect the acquisition or expression of
ethanol conditioned place preference in DBA/2J or C57BL/6J mice. Alcohol Clin Exp Res 30: 783-790.
Hyytia P (1993) Involvement of mu-opioid receptors in alcohol drinking by alcohol-preferring AA rats.
Pharmacol Biochem Behav 45: 697-701.
Hyytia P, Kiianmaa K (2001) Suppression of ethanol responding by centrally administered CTOP and naltrindole
in AA and Wistar rats. Alcohol Clin Exp Res 25: 25-33.
Ireland-Denny L, Parihar AS, Miller TR, Kang CH, Krueger KM, Esbenshade TA, Hancock AA. (2001) Species-
related pharmacological heterogeneity of histamine H(3) receptors Eur J Pharmacol 433:141-150.
Kamei J, Morita K, Miyata S, Onodera K (2003) Effects of second generation of histamine H1 antagonists,
cetirizine and ebastine, on the antitussive and rewarding effects of dihydrocodeine in mice. Psychopharmacology
(Berl) 166: 176-180.
Kiianmaa K. (1990) Neuronal mechanisms of ethanol sensitivity Alcohol Alcohol 25:257-262.
Kinoshita H, Harbuz MS, Nishiguchi M, Ouchi H, Minami T, Utsumi T, Motomura H, Hishida S (2004) High
alcohol preferring (HAP) and low alcohol preferring (LAP) rats show altered proopiomelanocortin (POMC)
messenger RNA expression in the arcuate nucleus. Alcohol Alcohol 39: 406-409.
Kitanaka N, Kitanaka J, Nishiguchi M, Kinoshita H, Ouchi H, Minami T, Hishida S, Takemura M (2004)
Decreased histamine-stimulated phosphoinositide hydrolysis in the cerebral cortex of a rat line selectively bred for
high alcohol preference. Neurochem Res 29: 1431-1436.
Kitbunnadaj R, Zuiderveld OP, De Esch IJ, Vollinga RC, Bakker R, Lutz M, Spek AL, Cavoy E, Deltent MF,
Menge WM, Timmerman H, Leurs R. (2003) Synthesis and structure-activity relationships of conformationally
constrained histamine H(3) receptor agonists J Med Chem 46:5445-5457.
Korpi ER, Kleingoor C, Kettenmann H, Seeburg PH (1993) Benzodiazepine-induced motor impairment linked to
point mutation in cerebellar GABAA receptor. Nature 361: 356-359.
Ligneau X, Perrin D, Landais L, Camelin JC, Calmels TP, Berrebi-Bertrand I, Lecomte JM, Parmentier R,
Anaclet C, Lin JS, Bertaina-Anglade V, la Rochelle CD, d'Aniello F, Rouleau A, Gbahou F, Arrang JM, Ganellin
CR, Stark H, Schunack W, Schwartz JC. (2007) BF2.649 [1-{3-[3-(4-chlorophenyl)propoxy]propyl}piperidine,
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 19
hydrochloride], a nonimidazole inverse agonist/antagonist at the human histamine H3 receptor: Preclinical
pharmacology. J Pharmacol Exp Ther 320:365-375.
Lintunen M, Hyytia P, Sallmen T, Karlstedt K, Tuomisto L, Leurs R, Kiianmaa K, Korpi ER, Panula P (2001)
Increased brain histamine in an alcohol-preferring rat line and modulation of ethanol consumption by H(3)
receptor mechanisms. FASEB J 15: 1074-1076.
Lintunen M, Raatesalmi K, Sallmen T, Anichtchik O, Karlstedt K, Kaslin J, Kiianmaa K, Korpi ER, Panula P
(2002) Low brain histamine content affects ethanol-induced motor impairment. Neurobiology of Disease 9: 94-
105.
Lozeva V, Tuomisto L, Sola D, Plumed C, Hippelainen M, Butterworth R (2001) Increased density of brain
histamine H(1) receptors in rats with portacaval anastomosis and in cirrhotic patients with chronic hepatic
encephalopathy. Hepatology 33: 1370-1376.
Lozeva V, Tuomisto L, Tarhanen J, Butterworth RF (2003) Increased concentrations of histamine and its
metabolite, tele-methylhistamine and down-regulation of histamine H3 receptor sites in autopsied brain tissue
from cirrhotic patients who died in hepatic coma. J Hepatol 39: 522-527.
McBride WJ, Kimpel MW, Schultz JA, McClintick JN, Edenberg HJ, Bell RL (2010) Changes in gene expression
in regions of the extended amygdala of alcohol-preferring rats after binge-like alcohol drinking. Alcohol 44: 171-
183.
Morisset S, Pilon C, Tardivel-Lacombe J, Weinstein D, Rostene W, Betancur C, Sokoloff P, Schwartz JC, Arrang
JM (2002) Acute and chronic effects of methamphetamine on tele-methylhistamine levels in mouse brain:
selective involvement of the D(2) and not D(3) receptor. J Pharmacol Exp Ther 300: 621-628.
Nguyen TL, You IJ, Oh S, Yang CH, Lee SY, Jang CG (2010) Assessment of the rewarding effects of
dimenhydrinate using the conditioned place preference paradigm in mice. Neurosci Lett 471: 38-42.
Nuutinen S, Karlstedt K, Aitta-Aho T, Korpi ER, Panula P (2010) Histamine and H3 receptor-dependent
mechanisms regulate ethanol stimulation and conditioned place preference in mice. Psychopharmacology (Berl)
208: 75-86.
Nylander I, Hyytia P, Forsander O, Terenius L (1994) Differences between alcohol-preferring (AA) and alcohol-
avoiding (ANA) rats in the prodynorphin and proenkephalin systems. Alcohol Clin Exp Res 18: 1272-1279.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 20
Okuda T, Zhang D, Shao H, Okamura N, Takino N, Iwamura T, Sakurai E, Yoshikawa T, Yanai K (2009)
Methamphetamine- and 3,4-methylenedioxymethamphetamine-induced behavioral changes in histamine H3-
receptor knockout mice. J Pharmacol Sci 111: 167-174.
Panula P, Pirvola U, Auvinen S, Airaksinen MS (1989) Histamine-immunoreactive nerve fibers in the rat brain.
Neuroscience 28: 585-610.
Panula P, Takagi H, Inagaki N, Yamatodani A, Tohyama M, Wada H, Kotilainen E (1993) Histamine-containing
nerve fibers innervate human cerebellum. Neurosci Lett 160: 53-56.
Pispa JP, Huttunen MO, Sarviharju M, Ylikahri R (1986) Enzymes of catecholamine metabolism in the brains of
rat strains differing in their preference for or tolerance of ethanol. Alcohol Alcohol 21: 181-184.
Rhodes JS, Best K, Belknap JK, Finn DA, Crabbe JC (2005) Evaluation of a simple model of ethanol drinking to
intoxication in C57BL/6J mice. Physiol Behav 84: 53-63.
Rhodes JS, Ford MM, Yu CH, Brown LL, Finn DA, Garland T,Jr, Crabbe JC. (2007) Mouse inbred strain
differences in ethanol drinking to intoxication Genes Brain Behav 6:1-18.
Risinger FO, Freeman PA, Greengard P, Fienberg AA (2001) Motivational effects of ethanol in DARPP-32
knock-out mice. J Neurosci 21: 340-348.
Robinson TE, Berridge KC (1993) The neural basis of drug craving: an incentive-sensitization theory of
addiction. Brain Res Brain Res Rev 18: 247-291.
Rusi M, Eriksson K, Maki J (1977) Genetic differences in the susceptibility to acute ethanol intoxication in
selected rat strains. Adv Exp Med Biol 85A: 97-109.
Rustay NR, Wahlsten D, Crabbe JC (2003) Influence of task parameters on rotarod performance and sensitivity to
ethanol in mice. Behav Brain Res 141: 237-249.
Ryu JH, Yanai K, Watanabe T (1994) Marked increase in histamine H3 receptors in the striatum and substantia
nigra after 6-hydroxydopamine-induced denervation of dopaminergic neurons: an autoradiographic study.
Neurosci Lett 178: 19-22.
Sanchis-Segura C, Spanagel R (2006) Behavioural assessment of drug reinforcement and addictive features in
rodents: an overview. Addict Biol 11: 2-38.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 21
Schnell D, Strasser A, Seifert R. (2010) Comparison of the pharmacological properties of human and rat
histamine H3-receptors Biochem Pharmacol .
Sinclair JD, Le AD, Kiianmaa K (1989) The AA and ANA rat lines, selected for differences in voluntary alcohol
consumption. Experientia 45: 798-805.
Singer HS, Szymanski S, Giuliano J, Yokoi F, Dogan AS, Brasic JR, Zhou Y, Grace AA, Wong DF (2002)
Elevated intrasynaptic dopamine release in Tourette's syndrome measured by PET. Am J Psychiatry 159: 1329-
1336.
Sommer W, Hyytia P, Kiianmaa K (2006) The alcohol-preferring AA and alcohol-avoiding ANA rats:
neurobiology of the regulation of alcohol drinking. Addict Biol 11: 289-309.
Stanley JL, Lincoln RJ, Brown TA, McDonald LM, Dawson GR, Reynolds DS (2005) The mouse beam walking
assay offers improved sensitivity over the mouse rotarod in determining motor coordination deficits induced by
benzodiazepines. J Psychopharmacol 19: 221-227.
Steeves TD, Ko JH, Kideckel DM, Rusjan P, Houle S, Sandor P, Lang AE, Strafella AP (2010) Extrastriatal
dopaminergic dysfunction in tourette syndrome. Ann Neurol 67: 170-181.
Stern E, Silbersweig DA, Chee KY, Holmes A, Robertson MM, Trimble M, Frith CD, Frackowiak RS, Dolan RJ
(2000) A functional neuroanatomy of tics in Tourette syndrome. Arch Gen Psychiatry 57: 741-748.
Suzuki T, Mori T, Takamori K, Onodera K, Misawa M (1996) Effects of H(1)-antagonists on discriminative
stimulus effects of cocaine and methamphetamine in rats. Behav Pharmacol 7: 111-118.
Suzuki T, Takamori K, Misawa M, Onodera K (1995) Effects of the histaminergic system on the morphine-
induced conditioned place preference in mice. Brain Res 675: 195-202.
Takemura M, Kitanaka N, Kitanaka J (2003) Signal transduction by histamine in the cerebellum and its
modulation by N-methyltransferase. Cerebellum 2: 39-43.
Takino N, Sakurai E, Kuramasu A, Okamura N, Yanai K (2009) Roles of the histaminergic neurotransmission on
methamphetamine-induced locomotor sensitization and reward: a study of receptors gene knockout mice. Int Rev
Neurobiol 85: 109-116.
Tanda G, Kopajtic TA, Katz JL (2008) Cocaine-like neurochemical effects of antihistaminic medications. J
Neurochem 106: 147-157.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 22
Tzschentke TM (2007) Measuring reward with the conditioned place preference (CPP) paradigm: update of the
last decade. Addict Biol 12: 227-462.
Wise RA, Bozarth MA (1987) A psychomotor stimulant theory of addiction. Psychol Rev 94: 469-492.
*Footnotes
The original studies have been financially supported by grants form the Academy of Finland [grant no
126744, 118547], Finnish Fund for Alcohol Research and Magnus Ehrnrooth’s Foundation
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 23
Legends for Figures
Figure 1
HA-immunoreactive nerve fibers in ANA (a,c,e,g) and AA (b,d,f,h) rat brain regions show distinct
differences in the medial septum (a,b), lateral septum (c,d), nucleus accumbens (e.f) and medial preoptic
nucleus. 3V, third ventricle; Aca, commissural anterior; acb, nucleus accumbens; LSV, lateral septum,
ventral part; LV, lateral ventricle; MPO, medial preoptic nucleus; Pe, periventricular nucleus. Scale bar
= 100 µm. Reproduced with permission from (Lintunen et al., 2001).
Figure 2 Lack of histamine synthesis leads to alterations in responses to alcohol. A) Male histidine decarboxylase
knockout (HDC KO) mice are not stimulated by acute alcohol (1.5 g/kg, i.p.). Data represents
cumulative distance moved within 30 min. ***p<0.001 from the baseline of wild type control mice. B)
Alcohol-induced reward is stronger in HDC KO mice than wild type control animals. An unbiased,
counterbalanced conditioned place preference paradigm was used where each animal received four 5-
min conditioning trials with alcohol and saline on alternating days. Grid+ stands for the conditioning
subgroup where metal grid floor was paired with alcohol (2 g/kg, i.p.) administration. In the Grid-group
plastic floor material was paired with alcohol injection. Place preference was indexed by comparing the
Grid+ and Grid- groups. ***p<0.001 from the corresponding Grid+ group. Two-way ANOVA yielded a
significant conditioning subgroup x genotype interaction indicating that the alcohol preference in HDC
KO mice was stronger that in control animals.
Figure 3
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 24
Distribution of the histaminergic neurons in the tuberomamillary nucleus of adult human posterior
hypothalamus in a frontal (A) and horizontal (B) sections. The schematic figures are based on camera
lucida drawings of two individuals (Airaksinen et al. 1991). Each dot represents a cell in a 30-40 um
thick section stained with an antibody against histamine. 3V, third ventricle; LH, lateral hypothalamic
area; Arc, arcuate nucleus, VMH, ventromedial hypothalamic nucleus, MM, medial mamillary nucleus;
f, fornix
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 25
Table 1 Examples of rat and mouse models and their suitability in the examination of alcohol-related
studies
Animal strain or line Description Characteristics
Rats
Sprague-Dawley Outbred line General multipurpose model , albino Wistar Outbred line General multipurpose model, albino AA and ANA rats Lines selectively bred for
high and low alcohol preference and consumption
Breeding program started in Finland in 1960s. Derived from Wistar and Sprague-Dawley background, Lewis and Brown Norwegian genotype was introduced later.
P and NP rats “ Breeding was initiated in 1970s in USA. Wistar background, inbred
sP and sNP rats “ Breeding started in 1981 in Sardinia, Wistar background AT and ANT rats Rats with genetic tolerance
or hypersensitivity towards alcohol-induced ataxia
Breeding started in 1970s in Finland. Several background lines.
Mice C57BL/6J Inbred strain, J refers to the
substrain from Jackson Laboratory
Multipurpose model. Used in ethanol drinking studies due to the high ethanol consumption. EtOH-induced CPP and stimulation low.
DBA/2J Inbred strain from Jackson Laboratory
Multipurpose model. Used e.g. in place preference studies. Do not drink ethanol voluntarily.
129/Sv Inbred strain, typically the background strain when generating knockout mice
Moderate to low responses to alcohol, high alcohol CPP, low spontaneous activity.
HAP/LAP High/low alcohol consumption
Derived from the HS/Igb 8-way inbred strain cross, USA
HDID-1, HDiD-2 High Drinking in the Dark Derived from the HS/Npt 8-way inbred strain cross, USA
For more information see: Crabbe et al., 1994, 2005, 2010; Kiianmaa 1990; Rhodes et al., 2007; Rustay et al., 2003; Tzschentke, 2007
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from
JPET#170928 26
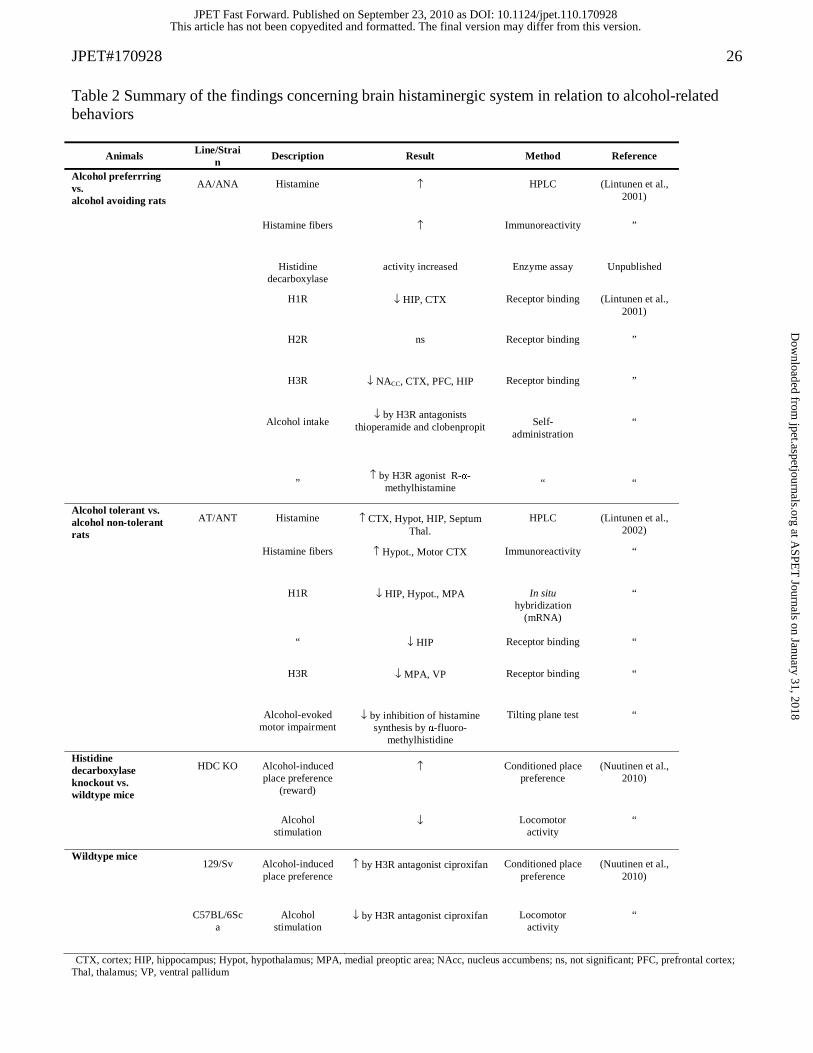
Table 2 Summary of the findings concerning brain histaminergic system in relation to alcohol-related behaviors
Animals Line/Strain
Description Result Method Reference
Alcohol preferrring vs. alcohol avoiding rats
AA/ANA Histamine ↑ HPLC (Lintunen et al., 2001)
Histamine fibers ↑ Immunoreactivity ”
Histidine
decarboxylase activity increased Enzyme assay Unpublished
H1R ↓ HIP, CTX Receptor binding (Lintunen et al.,
2001)
H2R ns Receptor binding ”
H3R ↓ NACC, CTX, PFC, HIP Receptor binding ”
Alcohol intake
↓ by H3R antagonists thioperamide and clobenpropit Self-
administration “
”
↑ by H3R agonist R-α-methylhistamine “ “
Alcohol tolerant vs. alcohol non-tolerant rats
AT/ANT Histamine ↑ CTX, Hypot, HIP, Septum Thal.
HPLC (Lintunen et al., 2002)
Histamine fibers ↑ Hypot., Motor CTX Immunoreactivity “
H1R ↓ HIP, Hypot., MPA
In situ hybridization
(mRNA)
“
“ ↓ HIP Receptor binding “
H3R ↓ MPA, VP Receptor binding “
Alcohol-evoked motor impairment
↓ by inhibition of histamine synthesis by α-fluoro-
methylhistidine
Tilting plane test “
Histidine decarboxylase knockout vs. wildtype mice
HDC KO Alcohol-induced place preference
(reward)
↑ Conditioned place preference
(Nuutinen et al., 2010)
Alcohol
stimulation ↓ Locomotor
activity “
Wildtype mice 129/Sv Alcohol-induced
place preference ↑ by H3R antagonist ciproxifan Conditioned place
preference (Nuutinen et al.,
2010)
C57BL/6Sc
a Alcohol
stimulation ↓ by H3R antagonist ciproxifan Locomotor
activity “
CTX, cortex; HIP, hippocampus; Hypot, hypothalamus; MPA, medial preoptic area; NAcc, nucleus accumbens; ns, not significant; PFC, prefrontal cortex; Thal, thalamus; VP, ventral pallidum
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET#170928 27
Table 3 Examples of brain penetrating histamine H3 receptor ligands
pKi=-logKi , the greater the pKi value → the higher the affinity towards H3Rs
Mechanism of action Human Rat Reference Agonists
Imetit agonist pKi= 9.2-9.7 pKi=9.4 (Ireland-Denny et al., 2001; Schnell et al., 2010)
Immepip partial agonist pKi=9.6 pKi=9.0 (Ireland-Denny et al., 2001) R-α-methylhistamine agonist pKi=8.9-9.2 pKi=8.6-8.7 (Ireland-Denny et al., 2001;
Schnell et al., 2010) Antagonists Imidazole-based
Ciproxifan Inverse agonist pKi=7.0 pKi=8.9-9.2 (Ireland-Denny et al., 2001; Schnell et al., 2010)
Thioperamide Inverse agonist pKi= 7.1-7.3 pKi= 7.9-8.1 ”
Clobenpropit Inverse agonist pKi=9.1-9.3 pKi=9.1-9.4 ”
Non-imidazole compounds
Pitolisant (BF2.649) Inverse agonist IC50=5.3 nM Ki=17 nM (Ligneau et al., 2007)
ABT-239 Inverse agonist pKi=9.4 pKi=8.9 (Cowart et al., 2004; Esbenshade et al., 2005)
JNJ-10181457 Neutral antagonist pKi=8.9 pKi=8.2 (Bonaventure et al., 2007)
VUF-5681 Neutral antagonist pKi=8.4 (Kitbunnadaj et al., 2003)
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from

This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on Septem
ber 23, 2010 as DO
I: 10.1124/jpet.110.170928 at ASPET Journals on January 31, 2018 jpet.aspetjournals.org Downloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 23, 2010 as DOI: 10.1124/jpet.110.170928
at ASPE
T Journals on January 31, 2018
jpet.aspetjournals.orgD
ownloaded from