homocysteine interference in neurulation: a chick embryo model
TRANSCRIPT

Homocysteine Interference in Neurulation:A Chick Embryo Model
L.A. Afman,1,2* H.J. Blom,1 N.M.J. Van der Put,1 and H.W.M. Van Straaten2
1Department of Pediatrics, University Medical Center Nijmegen, The Netherlands2Department of Anatomy and Embryology, Maastricht University, The Netherlands
Received 15 April 2002; Accepted 6 December 2003
BACKGROUND: Periconceptional folic acid supplementation reduces the occurrence and recurrence risk of neural tube defects (NTD).Mothers of children with NTD have elevated plasma homocysteine levels. Administering homocysteine to chick embryos is reported tocause 27% NTD. Therefore, elevated plasma homocysteine levels per se or a disturbed homocysteine metabolism may be teratogenic to theembryo and may interfere with neural tube closure. Our aim was to obtain a chick embryo model to explore the interference ofhomocysteine in neural tube closure. METHODS: Homocysteine or saline was administered to chick embryos in ovo at 3 hr, 30 hr, and 60hr of incubation and harvested at 74 hr. Homocysteine was then applied to chick embryos in vitro at a defined time window of four to sixsomites and followed for 6 hr. RESULTS: Homocysteine administration to chick embryos in ovo resulted in several malformations but notin an increased number of NTDs. Homocysteine administration to chick embryos in vitro resulted in a transient, dose-dependent wideningof the anterior neuropore and closure delay of the rhombencephalic neuropore. After 16 hr of incubation the neural tube was closed.CONCLUSIONS: The in vitro chick embryo model appears a good model to explore the interference of a disturbed homocysteinemetabolism in neurulation. Birth Defects Research (Part A) 67:421–428, 2003. © 2003 Wiley-Liss, Inc.
Key words: neural tube closure; homocysteine; chick embryo; anterior neuropore
INTRODUCTION
Neural tube defects (NTD) are among the most commonbirth defects, which often result in infant mortality orserious disability. Anencephaly and spina bifida aperta arethe two major types of NTD and arise due to failure ofclosure of the neural tube. Both genetic and environmentalfactors are involved in the development of NTD. Onefactor that showed a clear preventive effect on NTD ispericonceptional folic acid supplementation. A 70% reduc-tion in recurrence risk for NTD in women supplementedwith folic acid was shown in the MRC trial (Medical Re-search Council Vitamin Study Research Group, 1991). Astudy of Czeizel and Dudas (1992) showed a completereduction of first occurrence NTD in women supple-mented with folic acid. Three mouse mutant NTD modelsare known for a preventive effect of folic acid on NTD . Theunderlying preventive mechanism of folic acid in human isunknown so far, but a part can be explained by the pres-ence of the 677 C�T polymorphism in the methylene tet-rahydrofolate (MTHFR) gene, which is shown to be a riskfactor for NTD (Van der Put et al., 1995, 1997a). The en-zyme MTHFR catalyzes the formation of 5-methyltetrahy-drofolate (5-MeTHF) out of 5,10-methylene THF. Subse-quently, 5-MeTHF is a substrate in the remethylation ofhomocysteine to methionine. Decreased 5-MeTHF levelswill therefore result in elevated homocysteine levels (Fig.1). In addition, several studies have shown that plasmahomocysteine levels were elevated in mothers of childrenwith NTD and that these elevated homocysteine levels
were reduced after folic acid supplementation (Steegers-Theunissen et al., 1994, Homocysteine Lowering Trialists’Collaboration, 1998; Brouwer et al., 1999, 2000), leading tothe hypothesis that the elevated plasma homocysteine lev-els per se may be teratogenic for the embryo. Rosenquist etal. (1996) tested this hypothesis by administering homo-cysteine to chick embryos before and during the stages ofneural tube closure; 27% of these embryos developed NTD.The authors suggested that this teratogenic effect might bedue to a growth factor-like effect influencing the expres-sion of genes involved in neural tube closure. In a laterexperiment the authors suggested that homocysteine wasinhibiting the activity of the N-methyl-d-aspartate receptor(Andaloro et al., 1998; Rosenquist et al., 1999).
The closure of the neural tube involves many processesthat are far from understood. Elevation and convergence ofthe neural folds is generated by intrinsic factors like apicalconstriction of actin microfilaments in the neuroepithelialcells, spatially distinct cell proliferation patterns, intercala-tion of neural plate cells, reshaping of neural plate cellsfrom columnar to ‘wedge’ shape leading to the formation
Grant sponsor: Prinses Beatrix Fonds, The Netherlands; Grant number: 00-108.N.M.J. Van der Put’s current address: Unilever Research Vlaardingen, Oliviervan Noortlaan 120, 3133 AT Vlaardingen, The Netherlands.*Correspondence to: L.A. Afman, UMC Nijmegen, Laboratory of Pediatrics andNeurology, PO Box 9101, 6500 HB Nijmegen, The Netherlands.E-mail: [email protected] online in Wiley InterScience (www.interscience.wiley.com).DOI: 10.1002/bdra.10040
© 2003 Wiley-Liss, Inc. Birth Defects Research (Part A) 67:421–428 (2003)

of the hinge points, as well as by extrinsic factors, whichinclude expansion of the mesoderm and medial expansionof the ectoderm. The closure of the neural folds includesapposition, adhesion and cell fusion (Copp et al., 1990;Schoenwolf and Smith, 1990). Disturbance of any of theseprocesses may result in NTD; the complexity and vulner-ability of neurulation is further illustrated by the presenceof many NTD mouse mutants, which nowadays regardgenetic variations of over 60 genes (Juriloff and Harris,2000).
It is unclear how homocysteine or a disturbed homocys-teine metabolism interferes with the process and timingof neural tube closure. Besides the hypothesis of the ter-atogenicity of homocysteine levels per se, another moreindirect effect of elevated homocysteine levels on NTD issuggested; homocysteine is formed of s-adenosylhomocys-teine (AdoHcy) (Fig. 1), but the equilibrium of this reactionfavors AdoHcy formation. Thus, elevated homocysteinelevels may lead to increased AdoHcy levels that will in-hibit most transmethylation reactions by s-adenosylmethi-onine (AdoMet). Elevated homocysteine levels may there-fore disturb methylation of many targets, like genes andproteins that are involved in neural tube closure (Coelhoand Klein, 1990; Van Aerts et al., 1994). In case of folatedeficiency, reduced 5-MeTHF levels result in a reducedremethylation of homocysteine to methionine and conse-quently, into a reduced methylation.
We developed a chick embryo model enabling us tounravel how elevated homocysteine levels may affect theprocess of neural tube closure. We first repeated the studyof Rosenquist et al. (1996) by administering homocysteineto chick embryos in ovo. We could not, however, confirmtheir results. We therefore used chick embryos in vitro andwithin a narrow time window to examine more specificallythe interference of homocysteine in the neurulation pro-cess. We selected the developmental process of closure ofthe anterior neuropore (ANP) and of the rhombencephalicneuropore (RNP) at the stages beginning at four to sixsomites because the morphogenesis of these processeshave been reported in detail (Van Straaten et al., 1996,1997).
MATERIALS AND METHODS
White Leghorn chicken eggs were obtained from a com-mercial breeding farm and stored at 12°C. At the start ofthe experiment, eggs were incubated at 38°C and 55%humidity (Polyhatch; Brinsea Products, Sandford, UK).
Homocysteine Application to Embryos In OvoThe experiments of Rosenquist et al. (1996) were re-
peated as described by the authors. At the start of theexperiment the egg was placed vertically with the air
chamber up and a hole of 2 mm was made in the outershell membrane, through which 50 �L 0.1 M l-homocys-teine thiolactone (Sigma, St. Louis, MO) in saline (Locke’ssolution: 154 mM NaCl, 6 mM KCl, 2 mM CaCl2, 10 mMd-glucose) or 50 �L saline was applied on the inner shellmembrane at Stages 2, 7, and 10 of Hamburger and Ham-ilton (1951). Empirically, Stages 2, 7, and 10 appeared tocorrespond with incubation times of 3, 30, and 60 hr,whereas Rosenquist et al. (1996) found 3, 24, and 48 hr forthese stages. The hole was subsequently sealed with tapeand incubation was continued. The embryos were har-vested 74 hr after initiation of incubation, and their devel-opmental stage was determined by counting the number ofsomite pairs. Embryos were screened morphologically forNTDs, other malformations, and degeneration. Severaleggs were taken into the experiment without any treat-ment, to control for an effect of hole drilling. Differences infrequency of NTD between homocysteine and saline treat-ment in the in ovo experiment were calculated with �2
analysis. Difference in mean number of somites were cal-culated with Student’s t-test.
Homocysteine Application to Cultured EmbryosEmbryos were cultured in vitro on a substrate of agar
(1% agar [Difco, Detroit, MI] in saline with egg white 1:1)in a Petri dish at 38°C. Eggs were carefully broken in abowl filled with saline of 38°C and the egg white wasremoved from the vitelline membrane. Only embryos withfour to six somite pairs were dissected and subsequentlyplaced on the substratum with the vitelline membrane up.Embryos were incubated for 30 min to adapt before one ofthe following solutions (in saline) were applied: 80 �L of 30mM l-homocysteine thiolactone, 10 or 30 mM l-homocys-teine,10 or 30 mM l-cysteine, 10 or 30 mM leucine or saline.l-homocysteine solution was freshly prepared from l-ho-mocysteine thiolactone (Sigma) by alkaline hydrolysis (5min, 37°C, 5 N NaOH) and subsequent neutralization.l-homocysteine is unstable because it oxidizes into its di-sulfide homocystine. Therefore, l-homocysteine thiolac-tone, which is more stable, was also used in these experi-ments. The average number of somites of the embryos atthe start of the experiments was not significantly differentbetween the various treatments. Embryos were initiallyanalyzed together to increase statistical power.
Digital images were made immediately before adminis-tration of a solution as well as every hour after the admin-istration up to 6 hr using a stereo microscope (Leica MZFLIII) equipped with a video camera (JVC KY-F 55B). Thedevelopmental stages at 0 and 6 hr were determined bycounting the number of somites.
The anterior neuropore (ANP) begins as a narrow slit atthe stage of zero somites, broadens subsequently till the
Figure 1. Homocysteine metabolism. AdoMet,s-adenosylmethionine; AdoHcy, s-adenosylho-mocysteine; SAHH, s-adenosylhomocysteinehydrolase; MS, methionine synthase; THF, tet-rahydrofolate.
422 AFMAN ET AL.
Birth Defects Research (Part A) 67:421–428 (2003)
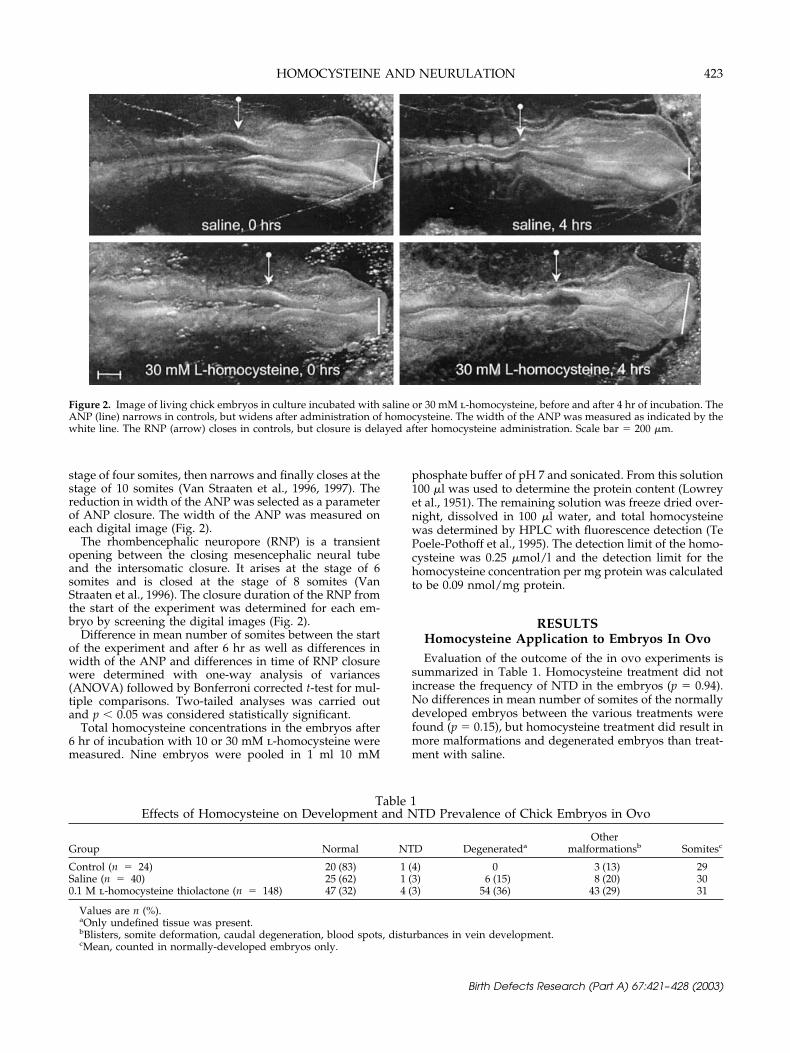
stage of four somites, then narrows and finally closes at thestage of 10 somites (Van Straaten et al., 1996, 1997). Thereduction in width of the ANP was selected as a parameterof ANP closure. The width of the ANP was measured oneach digital image (Fig. 2).
The rhombencephalic neuropore (RNP) is a transientopening between the closing mesencephalic neural tubeand the intersomatic closure. It arises at the stage of 6somites and is closed at the stage of 8 somites (VanStraaten et al., 1996). The closure duration of the RNP fromthe start of the experiment was determined for each em-bryo by screening the digital images (Fig. 2).
Difference in mean number of somites between the startof the experiment and after 6 hr as well as differences inwidth of the ANP and differences in time of RNP closurewere determined with one-way analysis of variances(ANOVA) followed by Bonferroni corrected t-test for mul-tiple comparisons. Two-tailed analyses was carried outand p � 0.05 was considered statistically significant.
Total homocysteine concentrations in the embryos after6 hr of incubation with 10 or 30 mM l-homocysteine weremeasured. Nine embryos were pooled in 1 ml 10 mM
phosphate buffer of pH 7 and sonicated. From this solution100 �l was used to determine the protein content (Lowreyet al., 1951). The remaining solution was freeze dried over-night, dissolved in 100 �l water, and total homocysteinewas determined by HPLC with fluorescence detection (TePoele-Pothoff et al., 1995). The detection limit of the homo-cysteine was 0.25 �mol/l and the detection limit for thehomocysteine concentration per mg protein was calculatedto be 0.09 nmol/mg protein.
RESULTSHomocysteine Application to Embryos In OvoEvaluation of the outcome of the in ovo experiments is
summarized in Table 1. Homocysteine treatment did notincrease the frequency of NTD in the embryos (p � 0.94).No differences in mean number of somites of the normallydeveloped embryos between the various treatments werefound (p � 0.15), but homocysteine treatment did result inmore malformations and degenerated embryos than treat-ment with saline.
Figure 2. Image of living chick embryos in culture incubated with saline or 30 mM l-homocysteine, before and after 4 hr of incubation. TheANP (line) narrows in controls, but widens after administration of homocysteine. The width of the ANP was measured as indicated by thewhite line. The RNP (arrow) closes in controls, but closure is delayed after homocysteine administration. Scale bar � 200 �m.
Table 1Effects of Homocysteine on Development and NTD Prevalence of Chick Embryos in Ovo
Group Normal NTD DegeneratedaOther
malformationsb Somitesc
Control (n � 24) 20 (83) 1 (4) 0 3 (13) 29Saline (n � 40) 25 (62) 1 (3) 6 (15) 8 (20) 300.1 M l-homocysteine thiolactone (n � 148) 47 (32) 4 (3) 54 (36) 43 (29) 31
Values are n (%).aOnly undefined tissue was present.bBlisters, somite deformation, caudal degeneration, blood spots, disturbances in vein development.cMean, counted in normally-developed embryos only.
423HOMOCYSTEINE AND NEURULATION
Birth Defects Research (Part A) 67:421–428 (2003)
Homocysteine Application to Cultured EmbryosAdministration of 30 mM l-homocysteine or 30 mM l-
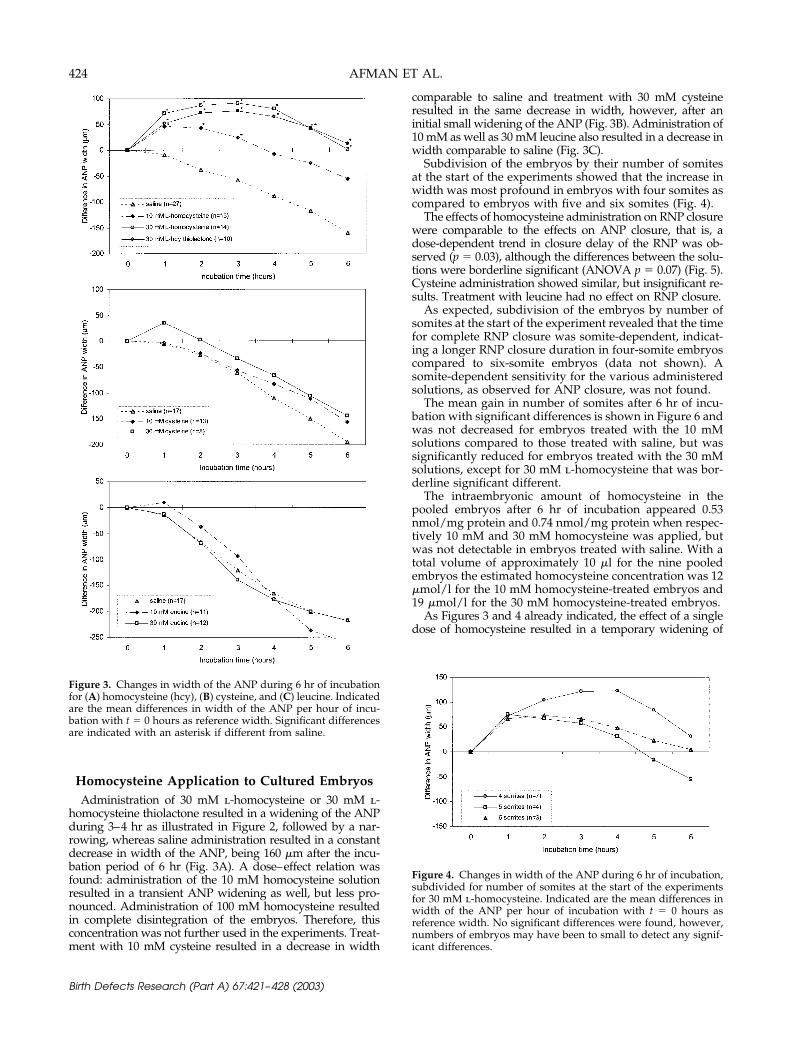
homocysteine thiolactone resulted in a widening of the ANPduring 3–4 hr as illustrated in Figure 2, followed by a nar-rowing, whereas saline administration resulted in a constantdecrease in width of the ANP, being 160 �m after the incu-bation period of 6 hr (Fig. 3A). A dose–effect relation wasfound: administration of the 10 mM homocysteine solutionresulted in a transient ANP widening as well, but less pro-nounced. Administration of 100 mM homocysteine resultedin complete disintegration of the embryos. Therefore, thisconcentration was not further used in the experiments. Treat-ment with 10 mM cysteine resulted in a decrease in width
comparable to saline and treatment with 30 mM cysteineresulted in the same decrease in width, however, after aninitial small widening of the ANP (Fig. 3B). Administration of10 mM as well as 30 mM leucine also resulted in a decrease inwidth comparable to saline (Fig. 3C).
Subdivision of the embryos by their number of somitesat the start of the experiments showed that the increase inwidth was most profound in embryos with four somites ascompared to embryos with five and six somites (Fig. 4).
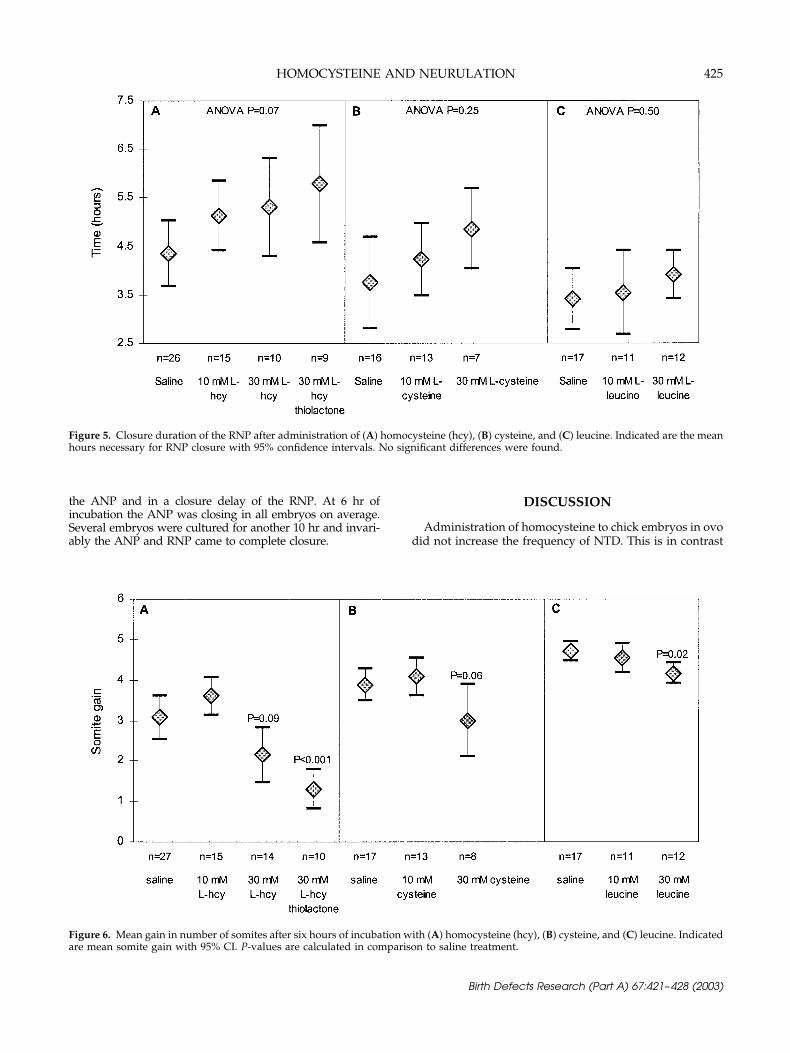
The effects of homocysteine administration on RNP closurewere comparable to the effects on ANP closure, that is, adose-dependent trend in closure delay of the RNP was ob-served (p � 0.03), although the differences between the solu-tions were borderline significant (ANOVA p � 0.07) (Fig. 5).Cysteine administration showed similar, but insignificant re-sults. Treatment with leucine had no effect on RNP closure.
As expected, subdivision of the embryos by number ofsomites at the start of the experiment revealed that the timefor complete RNP closure was somite-dependent, indicat-ing a longer RNP closure duration in four-somite embryoscompared to six-somite embryos (data not shown). Asomite-dependent sensitivity for the various administeredsolutions, as observed for ANP closure, was not found.
The mean gain in number of somites after 6 hr of incu-bation with significant differences is shown in Figure 6 andwas not decreased for embryos treated with the 10 mMsolutions compared to those treated with saline, but wassignificantly reduced for embryos treated with the 30 mMsolutions, except for 30 mM l-homocysteine that was bor-derline significant different.
The intraembryonic amount of homocysteine in thepooled embryos after 6 hr of incubation appeared 0.53nmol/mg protein and 0.74 nmol/mg protein when respec-tively 10 mM and 30 mM homocysteine was applied, butwas not detectable in embryos treated with saline. With atotal volume of approximately 10 �l for the nine pooledembryos the estimated homocysteine concentration was 12�mol/l for the 10 mM homocysteine-treated embryos and19 �mol/l for the 30 mM homocysteine-treated embryos.
As Figures 3 and 4 already indicated, the effect of a singledose of homocysteine resulted in a temporary widening of
Figure 4. Changes in width of the ANP during 6 hr of incubation,subdivided for number of somites at the start of the experimentsfor 30 mM l-homocysteine. Indicated are the mean differences inwidth of the ANP per hour of incubation with t � 0 hours asreference width. No significant differences were found, however,numbers of embryos may have been to small to detect any signif-icant differences.
Figure 3. Changes in width of the ANP during 6 hr of incubationfor (A) homocysteine (hcy), (B) cysteine, and (C) leucine. Indicatedare the mean differences in width of the ANP per hour of incu-bation with t � 0 hours as reference width. Significant differencesare indicated with an asterisk if different from saline.
424 AFMAN ET AL.
Birth Defects Research (Part A) 67:421–428 (2003)
the ANP and in a closure delay of the RNP. At 6 hr ofincubation the ANP was closing in all embryos on average.Several embryos were cultured for another 10 hr and invari-ably the ANP and RNP came to complete closure.
DISCUSSION
Administration of homocysteine to chick embryos in ovodid not increase the frequency of NTD. This is in contrast
Figure 5. Closure duration of the RNP after administration of (A) homocysteine (hcy), (B) cysteine, and (C) leucine. Indicated are the meanhours necessary for RNP closure with 95% confidence intervals. No significant differences were found.
Figure 6. Mean gain in number of somites after six hours of incubation with (A) homocysteine (hcy), (B) cysteine, and (C) leucine. Indicatedare mean somite gain with 95% CI. P-values are calculated in comparison to saline treatment.
425HOMOCYSTEINE AND NEURULATION
Birth Defects Research (Part A) 67:421–428 (2003)

to the experiments of Rosenquist et al. (1996) where 27%NTD were found. Several explanations for this discrepancyare considered. First, a variation in the chicken strain usedis likely present. The embryos used in our model appearedto have a lower developmental rate than those used byRosenquist et al. (1996). Second, an influence of nutritionalfactors can not be ruled out. Differences in diet supplied tothe chickens may change the composition of the egg yolk,supplying the embryos with a different set of factors in-volved in the metabolism of homocysteine. Besides thesevariations some other disadvantages of this model wereobserved. The amount of applied homocysteine reachingthe embryos likely varied considerably, because the posi-tion of the embryos was not always straight underneaththe injection hole. This can explain the finding of bothcompletely normally developed as well as degeneratedembryos after incubation with identical homocysteine con-centrations. Another disadvantage was that the actual de-velopmental stage of the embryos at time of administrationof homocysteine was unknown; only an estimate could bemade. As the developmental time window in which thehomocysteine is affecting neural tube closure may be nar-row, the age of the embryos at administration of homocys-teine is an important issue.
Recently, another study on chick embryos in ovoshowed an increased frequency of spina bifida after l-homocysteine thiolactone administration (Epeldegui et al.,2002). Although the administered homocysteine concentra-tion of 2.5 M was much higher than the 0.1 M used in ourin ovo experiments a low number of gross abnormalitieswas observed. The number of spinal NTDs of approxi-mately 10% in the control group indicates that the whiteleghorn strain used in their study may be more susceptiblefor spinal NTD than the white leghorn strain used in ourstudy.
By using the culture model we were able to control theamount of homocysteine to be administered as well as thedevelopmental stage of the embryos. Moreover, we wereable to monitor the influence of the homocysteine concen-trations on the developing embryo. Administration of ho-mocysteine to chick embryos of four to six somites resultedin a transient, dose-dependent widening of the ANP and adelayed closure of the RNP.
Comparison of the development of the embryos in theculture model with the in ovo situation (Van Straaten et al.,1996) demonstrated highly similar timing and presence ofdevelopmental processes. The mean reduction in width ofthe ANP of 160 �m observed in embryos treated withsaline in vitro corresponded to a similar reduction duringdevelopment of four somite embryos to seven somite em-bryos in ovo. The mean necessary time observed for com-plete RNP closure of 2.7 hr in an in vitro saline-treatedembryo of six somites implies that the embryo was sevento eight somites at time of completion of RNP closure. Thisis similar to the number of somites of seven to eight re-ported for completion of RNP closure in ovo (Van Straatenet al., 1996). The development of three to four somitesduring 6 hr of incubation was comparable to the in ovosituation as well. The differences in grow rate, as reflectedby the somite gain, observed for the saline treated embryoscan be explained by the season-sensitivity of chick em-bryos.
By using this culture model we found that homocysteinehas an dose–dependent effect on the closure of the ANP
and RNP in a narrow time window between four to sixsomites. Concerning ANP closure, the sensitivity of theembryos for homocysteine appeared to depend on theirdevelopmental age, the highest sensitivity being at foursomites. An explanation for the decreased sensitivity atlater stages may be that the process of closure of the neuralfolds had passed the stages of elevation of the neural walls.These findings indicate that not all phases of neural tubeclosure are susceptible for high homocysteine levels andthat the effect of homocysteine in ANP closure may evenoccur in a narrow time window.
A delay in development, determined by the gain insomites, occurred when 30 mM homocysteine was appliedbut was not seen in embryos administered 10 mM l-ho-mocysteine, whereas a transient widening of the ANP andclosure delay of the RNP occurred at both homocysteineconcentrations. This finding, including the reduced somitegain observed for 30 mM cysteine as well as for 30 mMleucine indicates that the 30 mM homocysteine concentra-tion is too high to attain normal development of the em-bryos, apart from the widening of the ANP and the closuredelay of the RNP. The normal gain in somites in embryostreated with 10 mM homocysteine suggest that the effectson ANP and RNP closure are specific and not general toxiceffects.
Homocysteine concentrations in embryos treated withhomocysteine were increased, demonstrating that the ho-mocysteine was internalized. Nevertheless, the concentra-tion of homocysteine as measured in the embryos was 1000times lower than that administered. We therefore suggestthat either the embryos absorbed only a small part of thehomocysteine or that the embryos metabolized or exportedthe homocysteine fast.
The recovery of 10 mM homocysteine and 30 mM ho-mocysteine-treated embryos indicates that one dose of ho-mocysteine might not be enough to achieve the maximumeffect on ANP closure. The identical ANP developmentcurves for 30 mM l-homocysteine and 30 mM l-homocys-teine thiolactone (Fig. 3) indicate that oxidation of homo-cysteine in the incubation medium is not the explanationfor the recovery. The recovery is further illustrated by thecomplete closure of the ANP after 16 hr of incubation.
Another point of discussion is the small widening of theANP during the first hour of incubation with 30 mMcysteine. Cysteine, which molecular structure closely re-sembles that of homocysteine, may affect ANP widening inthe same way as homocysteine but less profound. Bothhomocysteine and cysteine are able to split S–S bridges,which may be involved in neural tube closure. Presum-ably, cysteine is less harmful because in vivo cysteineconcentration in blood is about 20-fold higher than homo-cysteine concentrations. The higher osmolarity of the 30mM solution compared to the 10 mM solution and salinecan not explain this effect as 30 mM leucine treatment didnot result in a widening of the ANP. Taken together,although ultimately no final closure failure arose, this cul-ture model appeared very suitable to study the influence ofhomocysteine on the neurulation mechanism in a smalltime window.
A few other studies on the effect of homocysteine to invitro animal models are published. In cultured rat em-bryos, up to 6 mM homocysteine administration did notincrease the frequency of NTD (Van Aerts et al., 1994), butsome of the malformations they found: irregular somites
426 AFMAN ET AL.
Birth Defects Research (Part A) 67:421–428 (2003)

and blisters, correspond to the malformations found in ourin ovo experiments. Recently, Hansen et al. (2000) reportedthat NTD did neither occur in mice embryos cultured with1.3 mM d,l-homocysteine in the culture medium, nor inembryos cultured with approximately 78 mM l-homocys-teine thiolactone applied to the amniotic sac. The transientwidening of the ANP shown in our study may also havebeen present in the above mentioned experiments in rats(Van Aerts et al., 1994) and mice (Hansen et al., 2000) butno data on specific ANP closure were presented. Hansen etal. (2000) showed that 24 hr after administration of the 78mM l-homocysteine thiolactone solution the homocysteineconcentration in the embryos were not increased signifi-cantly. Van Aerts et al. (1994) found however significantlyelevated homocysteine levels in embryos cultured with 3mM l-homocysteine for 2 hr. We found elevated homocys-teine levels after 6 hr of incubation with l-homocysteine.The outcome of these studies may be explained by a pooruptake, a rapid metabolization, or a rapid export of appliedhomocysteine.
A possible mechanism for the effect of homocysteine inwidening of the ANP may regard the disturbance of meth-ylation. One of the proteins in which methylation is con-sidered important is actin. Methylation of the histidineresidue of actin occurs at a specific and highly conservedlocation on the molecule, which may be important in mi-crofilament function (Ueymura and Spudich, 1980). In neu-roepithelial cells of the neural groove apical constriction byactin microfilaments is proposed to be an important factorin the elevation and convergence of the neural folds, beingimportant especially in the cephalic region (Morriss-Kayand Tuckett, 1985; Ybot-Gonzalez and Copp, 1999; VanStraaten et al., 2002). A role for methylation in microfila-ment contraction during neural tube closure was sug-gested originally by Coelho et al. (1990). They found in-complete closure of the neural tube in rat embryos culturedon a methionine-deficient serum which has been con-firmed by Van Aerts et al. (1994). Moephuli et al. (1997)found that localization of actin and ��-tubulin in the basalcytoplasm did not occur in embryos cultured on a methi-onine-deficient serum. They also showed that these pro-teins are among the most excessively synthesized duringneural tube closure.
The aim of our study was to examine the effects of highhomocysteine concentrations on neurulation. When com-paring our in vitro experiments to the in vivo situation inhuman the following comment should be made. In moth-ers of NTD children homocysteine levels are slightly ele-vated to about 15 �mol/l, caused by an acquired or geneticfactors in the homocysteine and folate metabolism. Con-trarily, the embryos in our experiments were treated withone bolus of an unphysiologically high homocysteine con-centration of 10–30 mM, although in the embryos onlyhomocysteine levels of 12–19 �mol/l were found. In addi-tion, administration of one bolus of homocysteine resultedin a recovery of the effects on neural tube closure.
In conclusion, the present study shows that high homo-cysteine concentrations affect the process of neural tubeclosure at multiple sites in chick embryos. Whether this isdue to a changed methylation of DNA, RNA or proteinslike actin, caused by a high AdoHcy level or whether it isdue to teratogenicity of homocysteine itself is unclear.Further research is necessary to elucidate how a disturbed
homocysteine and folate metabolism interferes in neurula-tion.
ACKNOWLEDGMENTSDr. H.J. Blom is Established Investigator of The Nether-
lands Heart Foundation (D47.021). We thank M. Drittij andA. de Graaf-Hess for excellent support.
REFERENCESHomocysteine Lowering Trialists’ Collaboration. 1998. Lowering blood ho-
mocysteine with folic acid based supplements: meta-analysis of ran-domized trials. BMJ 316:894–898.
Andaloro VJ, Monaghan DT, Rosenquist TH. 1998. Dextromethorphan andother N-methyl-d-aspartate receptor antagonists are teratogenic in theavian embryo model. Pediatr Res 43:1–7.
Brouwer IA, van Dusseldorp M, Thomas CM, et al. 1999. Low-dose folicacid supplementation decreases plasma homocysteine concentrations:a randomized trial. Am J Clin Nutr 69:99–104.
Brouwer IA, van Dusseldorp M, Thomas CM, et al. 2000. Homocysteinemetabolism and effects of folic acid supplementation in patients af-fected with spina bifida. Neuropediatrics 31:298–302.
Carter M, Ulrich S, Oofuji Y, et al. 1999. Crooked tail (Cd) models humanfolate-responsive neural tube defects. Hum Mol Genet 8:2199–2204.
Coelho CN, Klein NW. 1990. Methionine and neural tube closure in cul-tured rat embryos: morphological and biochemical analyses. Teratol-ogy 42:437–451.
Copp AJ, Brook FA, Estibeiro JP, et al. 1990. The embryonic development ofmammalian neural tube defects. Prog Neurobiol 35:363–403.
Czeizel AE, Dudas I. 1992. Prevention of the first occurrence of neural-tubedefects by periconceptional vitamin supplementation. N Engl J Med327:1832–1835.
Epeldegui M, Pena-Melian A, Varela-Moreiras G, Perez-Miguelsanz J. 2002.Homocysteine modifies development of neurulation and dorsal rootganglia in chick embryos. Teratology 65:171–179.
Fleming A, Copp AJ. 1998. Embryonic folate metabolism and mouse neuraltube defects. Science 280:2107–2109.
Hamburger V, Hamilton H.G. 1951. A series of normal stages in the devel-opment of the chick embryo. J Morphol 88:49–92.
Hansen DK, Grafton TF, Melnyk S, James SJ. 2000. Lack of embryotoxicityof homocysteine thiolactone in mouse embryos in vitro. Reprod Toxicol15:239–244.
Juriloff DM, Harris MJ. 2000. Mouse models for neural tube closure defects.Hum Mol Genet 9:993–1000.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951. Protein measure-ment with the Folin phenol reagent. J Biol Chem 193:265–275.
Medical Research Council Vitamin Study Research Group. 1991. Preventionof neural tube defects: results of the Medical Research Council VitaminStudy. Lancet 338:131–137.
Mills JL, McPartlin JM, Kirke PN, et al. 1995. Homocysteine metabolism inpregnancies complicated by neural-tube defects. Lancet 345:149–151.
Moephuli SR, Klein NW, Baldwin MT, Krider HM. 1997. Effects of methi-onine on the cytoplasmic distribution of actin and tubulin duringneural tube closure in rat embryos. Proc Natl Acad Sci USA 94:543–548.
Morriss-Kay G, Tuckett F. 1985. The role of microfilaments in cranial neu-rulation in rat embryos: effects of short-term exposure to cytochalasinD. J Embryol Exp Morphol 88:333–348.
Rosenquist TH, Ratashak SA, Selhub J. 1996. Homocysteine induces con-genital defects of the heart and neural tube: effect of folic acid. ProcNatl Acad Sci USA 93:15227–15232.
Rosenquist TH, Schneider AM, Monogham DT. 1999. N-methyl-d-aspartatereceptor agonists modulate homocysteine-induced developmental ab-normalities. FASEB J 13:1523–1531.
Schoenwolf GC, Smith JL. 1990. Mechanisms of neurulation: traditionalviewpoint and recent advances. Development 109:243–270.
Steegers-Theunissen RP, Boers GH, Trijbels FJ, et al. 1994. Maternal hyper-homocystinemia: a risk factor for neural-tube defects? Metabolism43:1475–1480.
Te Poele-Pothoff M, van den Berg M, Franken DG, et al. 1995. Threedifferent methods for the determination of total homocysteine inplasma. Ann Clin Biochem 32:218–220.
Ueymura DG, Spudich JA. 1980. Biochemistry and regulation of non-muscleactins. In: Goldberger RF, editor. Biological regulations in develop-ment. New York: Plenum Press. pp. 327–328.
Van der Put NM, Eskes TK, Blom HJ. 1997a. Is the common 677C�Tmutation in the methylenetetrahydrofolate reductase gene a risk factorfor neural tube defects? A meta-analysis. QJM 90:111–115.
Van der Put NM, Steegers-Theunissen RP, Frosst P, et al. 1995. Mutated
427HOMOCYSTEINE AND NEURULATION
Birth Defects Research (Part A) 67:421–428 (2003)

methylenetetrahydrofolate reductase as a risk factor for spina bifida.Lancet 346:1070–1071.
Van der Put NM, Thomas CM, Eskes TK, et al. 1997b. Altered folate andvitamin B12 metabolism in families with spina bifida offspring. QJM90:505–510.
Van Straaten HW, Janssen HC, Peeters MC, et al. 1996. Neural tube closurein the chick embryo is multiphasic. Dev Dyn 207:309–318.
Van Straaten HW, Peeters MC, Szpak KF, Hekking JW. 1997. Initial closureof the mesencephalic neural groove in the chick embryo involves areleasing zipping-up mechanism. Dev Dyn 209:333–341.
Van Straaten HWM, Sieben I, Hekking JWM. 2002. Multistep role for actin
in initial closure of the mesencephalic neural groove in the chickembryo. Dev Dyn 224:103–108.
Van Aerts LA, Blom HJ, Deabreu RA, et al. 1994. Prevention of neural tubedefects by and toxicity of l-homocysteine in cultured postimplantationrat embryos. Teratology 50:348–360.
Ybot-Gonzalez P, Copp AJ. 1999. Bending of the neural plate during mousespinal neurulation is independent of actin microfilaments. Dev Dyn215:273–283.
Zhao Q, Behringer RR, de Crombrugghe B. 1996. Prenatal folic acid treat-ment suppresses acrania and meroanencephaly in mice mutant for theCart1 homeobox gene. Nat Genet 13:275–283.
428 AFMAN ET AL.
Birth Defects Research (Part A) 67:421–428 (2003)