hsp70 bag3 interactions regulate cancer-related signaling...
TRANSCRIPT

Molecular and Cellular Pathobiology
Hsp70–Bag3 Interactions Regulate Cancer-RelatedSignaling Networks
TeresaA.Colvin1,2, Vladimir L.Gabai2, JianlinGong1,Stuart K.Calderwood3,HuLi4, SuryaramGummuluru5,Olga N. Matchuk6, Svetlana G. Smirnova6, Nina V. Orlova6, Irina A. Zamulaeva6, Mikel Garcia-Marcos2,Xiaokai Li7, Z.T. Young7, Jennifer N. Rauch7, Jason E. Gestwicki7, Shinichi Takayama8, andMichael Y. Sherman2
AbstractBag3, a nucleotide exchange factor of the heat shock protein Hsp70, has been implicated in cell signaling. Here,
we report that Bag3 interacts with the SH3 domain of Src, therebymediating the effects of Hsp70 on Src signaling.Using several complementary approaches, we established that the Hsp70–Bag3 module is a broad-actingregulator of cancer cell signaling by modulating the activity of the transcription factors NF-kB, FoxM1, Hif1a,the translation regulator HuR, and the cell-cycle regulators p21 and survivin. We also identified a small-moleculeinhibitor, YM-1, that disrupts the Hsp70–Bag3 interaction. YM-1mirrored the effects of Hsp70 depletion on thesesignaling pathways, and in vivo administration of this drug was sufficient to suppress tumor growth in mice.Overall, our results defined Bag3 as a critical factor in Hsp70-modulated signaling and offered a preclinical proof-of-concept that the Hsp70–Bag3 complex may offer an appealing anticancer target. Cancer Res; 74(17); 4731–40.�2014 AACR.
IntroductionThe major heat shock protein Hsp70 (HspA1A) has also been
implicated in cancer. Hsp70 is a stress-inducible molecularchaperone that participates in protection of the proteome fromaggregation and promotes refolding and degradation of dam-aged polypeptides (1, 2). The levels of this protein are highlyelevated in a variety of cancers, and correlate with tumor grade,metastasis, chemotherapy resistance, and subsequently, poorprognosis, suggesting that Hsp70 plays a specific role in cancer(3, 4). Importantly, although Hsp70 is critical for survival ofcancer cells, it is dispensable for viability ofnontransformedcells
(5). Accordingly, Hsp70 knockout mice are healthy (6), and theinactivation of Hsp70manifests serious problems only followinga challenge with stress (6, 7), or acute inflammation (8).
To leverage these observations into therapeutics, severalgroups have explored the mechanisms by which cancer cellsbecome addicted to Hsp70. On their path to tumorigenesis, cellsmust escape apoptosis (9), anoikis (10), and the harsh conditionsof the tumormicroenvironment (11). Although elevated levels ofHsp70 have strong antiapoptotic activity (12, 13), it is not yetclear whether this function is important for its role in cancer.
Unlike apoptosis, the finding that Hsp70 suppresses cellularsenescence has established a clear connection to cancer.Senescence is defined as irreversible cell growth arrest thatis associatedwith assorted cellular changes inmorphology andgene expression (14, 15). Certain oncogenes may trigger senes-cence, oncogene-induced senescence (OIS), which providesdefense against cancer. We have demonstrated that depletionof Hsp70 activates senescence in cells transformed by theoncogenes Her2, PIK3CA, and RAS, but that it has minimaleffects in normal cells (16, 17). Accordingly, Hsp70 knockoutmice did not develop breast cancer upon expression of Her2oncogene (5). Therefore, Hsp70 could be critical for the escapeof transformed cells fromOIS, a property that defines its role intumor initiation.
These properties of Hsp70 suggest that it could be used as adrug target. There have been several attempts to developinhibitors of Hsp70 for cancer treatment, including inhibitorsof substrate binding, for example, aptamers (18) or pifithrin m(19–22), or compounds that interact with the ATPase domain,such as VER155008 (23), MAL3-101 (24), and YK-5 (25). How-ever, development of these inhibitors has not reached theclinical trial stage.
1Department of Medicine, Boston University School of Medicine, Boston,Massachusetts. 2Department of Biochemistry, BostonUniversity School ofMedicine, Boston, Massachusetts. 3Molecular and Cellular RadiationOncology, Beth Israel DeaconessMedicalCenter, HarvardMedical School,Boston, Massachusetts. 4Center for Individualized Medicine, Departmentof Molecular Pharmacology & Experimental Therapeutics, Mayo Clinic,Rochester, Minnesota. 5Department of Microbiology, Boston UniversitySchool of Medicine, Boston, Massachusetts. 6Medical RadiologyResearch Center, Obninsk, Russia. 7Department of Pharmaceutical Chem-istry, Institute for Neurodegenerative Disease, University of California atSan Francisco, San Francisco, California. 8Department of Pathology,Boston University School of Medicine, Boston, Massachusetts.
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
T.A. Colvin and V.L. Gabai contributed equally to this article.
Corresponding Authors: Michael Y. Sherman, Department of Biochem-istry, Boston University School of Medicine, Boston, MA 02118. Phone:617-638-5971; Fax; 617-638-5339; E-mail: [email protected]; JasonGest-wicki, [email protected]; and Shinuchi Takayama,[email protected]
doi: 10.1158/0008-5472.CAN-14-0747
�2014 American Association for Cancer Research.
CancerResearch
www.aacrjournals.org 4731
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747

To tailor inhibitor development to cancer, specific mechan-isms underlying the effects of Hsp70 on tumor developmentshould be defined. In search for these mechanisms, we focusedon a nucleotide exchange factor Bag3. This protein containsthe Bag domain that binds to a motif in the ATPase domain ofHsp70 proteins (26), as well as PxxP and WW domains (26),which may connect it to SH3 domains and PPxY motifs ofsignaling proteins (27, 28). Bag3 has been implicated in macro-autophagy and aggresome formation (29–31). In both process-es, Bag3 was proposed to link complexes of Hsp70 with proteinaggregates to the autophagic and aggresome machineries.Although it is unknown how Bag3 interacts with the latter, inthe process of autophagy, Bag3 uses itsWWdomain to interactwith one of the organizers of the autophagic vacuole SYNPO2(30). In addition to its function in recruitment of the Hsp70-bound cargo, it was demonstrated that Bag3 can interact withsignaling factors via its PxxP motif (e.g., with PLCg ; ref. 32) orWW domain (e.g., with components of the Hippo pathwayLATS1 and AMOT1; ref. 30). Because Bag3 has been implicatedin cancer cell motility and invasion (33–35), there is a possi-bility that these effects result from regulation of signalingpathways by Bag3. Interestingly, in the previous studies, theeffects of Bag3 on signaling were not connected to effects ofHsp70 on these pathways. Here, we hypothesized that Bag3could serve as a scaffold with the potential for integratingHsp70 levels andmultiple cancer signaling pathways, and thusmediate effects of Hsp70 on cancer.
Materials and MethodsCell cultures
MCF10A, HEK293, MCF7, HCT116, B16-F10, and HeLawere from ATCC and initially frozen for storage uponreceipt. ATCC authenticates its cell lines using short tan-dem repeat analysis. Cells were resuscitated and passagedfor less than 6 months during use in experiments. MCF10Acells expressing PIK3CA or Her2 were as described before(5, 16, 17). MCF10A cells were cultivated in Dulbecco'sModified Eagle Medium (DMEM)-F12 medium supplemen-ted with 5% horse serum, hydrocortisone (500 ng/mL),insulin (10 mg/mL), cholera toxin (100 ng/mL), and epi-dermal growth factor (20 ng/mL). HEK293T cells weremaintained in DMEM supplemented with 10% heat-inacti-vated fetal bovine serum (FBS). B16-F10, MCF7, and HeLacells were cultivated in DMEM supplemented with 10%FBS. HCT116 cells were cultured in McCoy 5A mediumwith 10% FBS. All cultures contained 1% penicillin/streptomycin.
Mice and xenograftsFor xenografts, 500,000 MCF7 cells were mixed at a 1:1 ratio
withMatrigel, and 1million cells were injected subcutaneouslyin 6-week-old female NCR nude mice (Taconic). For the B16-F10 melanoma allografts, 200,000 cells were injected subcuta-neously in hind legs of 6-week-old C57BL6 female mice. Tumorgrowth was monitored every other day using caliper andcalculated according to the formula L � W2 � p/6, where Lis length and W is width.
Compounds and antibodiesMG132 was purchased from Biomol and puromycin from
Sigma. YM-1 was developed and supplied by Dr. J.E. Gestwicki(University of California at San Francisco, San Francisco, CA).The following antibodies were used: anti-Hsp70 was fromStressgen, anti-Bag3 was a gift from Dr. S. Takayama (BostonUniversity School of Medicine, Boston, MA), anti-p21 and anti-Hif1awere fromBDPharmingen, anti-b-actin, anti-Src, anti-p-Src (Tyr416), p-NF-kB (Ser536), anti-IkBa, p-paxillin (Tyr118),p-p130Cas (Tyr165, Tyr249, and Tyr410), and p130Cas werefrom Cell Signaling Technology, anti-FoxM1, anti-HuR, anti-survivin, and anti-Src were from Santa Cruz Biotechnology.
ImmunoblottingImmunoblotting and cell lysate preparations were done
according to ref. 5.
ResultsBag3 mediates effects of Hsp70 on Src activity
Touncover the role of Bag3 and its associationwithHsp70 incancer, we sought to identify SH3 domain-containing signalingfactors that can interact with the PxxP motif of Bag3. Accord-ingly, we performed pull-down experiments using a seriesof recombinant SH3 domains of various proteins fused to GST.GST-CD40 lacking the SH3 domain was used as a negativecontrol. Lysates with in vitro–translated S35-labeled Bag3were passed over beads carrying various SH3–GST fusionproteins, and bound Bag3 was detected. In the pull-downexperiment, Bag3 interacted with SH3 domains of PLCg , Src,c-Abl, c-Grap, Nck, Crk, RasGAP, n-Grab2, and c-Grab2(Fig. 1A). These proteins are components of cancer-relatedpathways, highlighting the potential of Bag3 as a masterregulator of cancer signaling. Although any of these signalingproteins may contribute to cancer development, to exploremechanistically howBag3 regulates signaling, we chose Src as amodel.
First, we confirmed that the full-length Src protein interactswith Bag3 in cells. FLAG-labeled Bag3 was coexpressed withSrc in 293T cells, precipitatedwith anti-FLAGantibody, and Srcwas probed on the blot with anti-Src antibody. Figure 1B, leftshows that Src is effectively associated with Bag3. A similarexperiment was carried out in HeLa cells to detect associationof Bag3 with endogenous Src (Fig. 1B, right).
Because Bag3 is a cofactor ofHsp70, we further assessed howthe ability to interact with Hsp70 influences association ofBag3 with the SH3 domain of Src. Various constructs of Bag3were expressed, cell lysateswere incubatedwith beads carryingrecombinant SH3 domain of Src, and bound Bag3 moleculeswere detected by immunoblotting. Besides a full-length Bag3construct, we tested deletions of the PxxP and Bag domains(referred to as FL, DPxxP, and DB, respectively), as well as anR480Amutation, which impairs Bag3 binding toHsp70 (Fig. 1C;ref. 36). The inability of the DB and R480A constructs to bind toHsp70 was confirmed in the pull-down experiment (Fig. 1D).The full-length Bag3 effectively interacted with the SH3domain of Src, and binding was strongly reduced by deletionof the PxxP domain, as expected (Fig. 1E). Importantly, bothDB
Colvin et al.
Cancer Res; 74(17) September 1, 2014 Cancer Research4732
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747
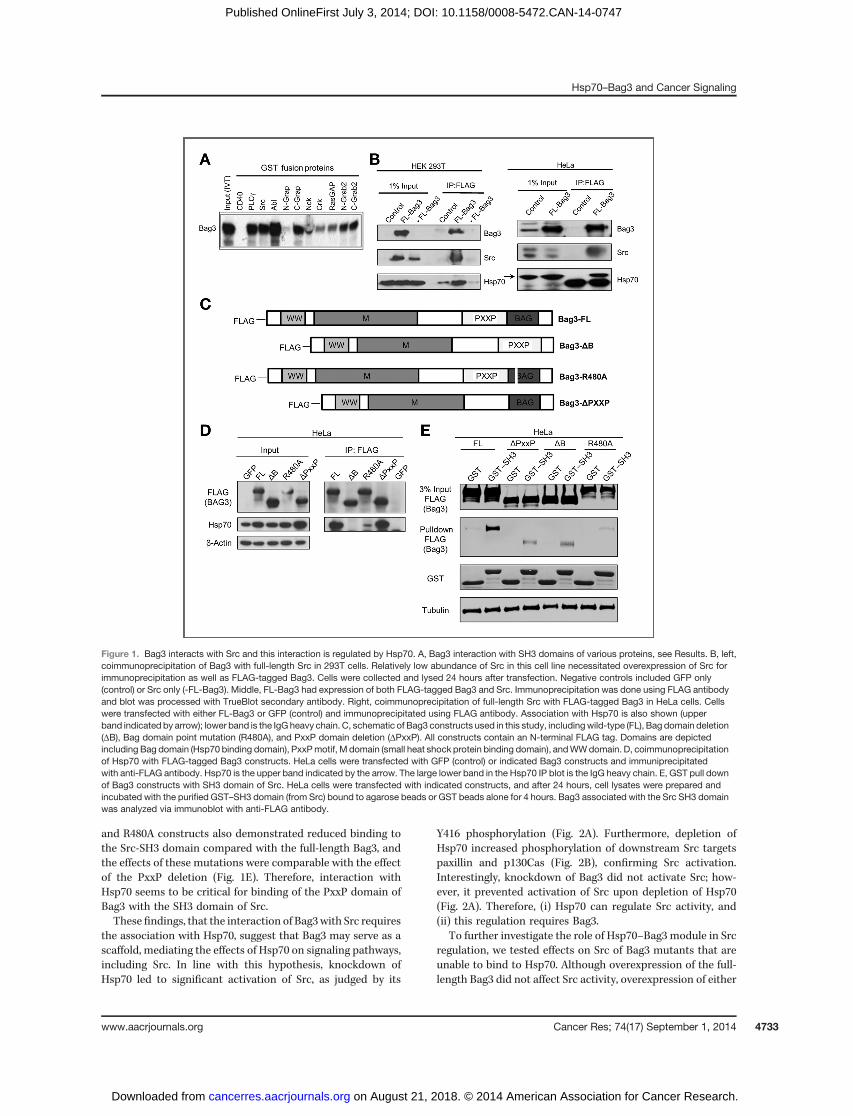
and R480A constructs also demonstrated reduced binding tothe Src-SH3 domain compared with the full-length Bag3, andthe effects of these mutations were comparable with the effectof the PxxP deletion (Fig. 1E). Therefore, interaction withHsp70 seems to be critical for binding of the PxxP domain ofBag3 with the SH3 domain of Src.These findings, that the interaction of Bag3with Src requires
the association with Hsp70, suggest that Bag3 may serve as ascaffold, mediating the effects of Hsp70 on signaling pathways,including Src. In line with this hypothesis, knockdown ofHsp70 led to significant activation of Src, as judged by its
Y416 phosphorylation (Fig. 2A). Furthermore, depletion ofHsp70 increased phosphorylation of downstream Src targetspaxillin and p130Cas (Fig. 2B), confirming Src activation.Interestingly, knockdown of Bag3 did not activate Src; how-ever, it prevented activation of Src upon depletion of Hsp70(Fig. 2A). Therefore, (i) Hsp70 can regulate Src activity, and(ii) this regulation requires Bag3.
To further investigate the role of Hsp70–Bag3 module in Srcregulation, we tested effects on Src of Bag3 mutants that areunable to bind to Hsp70. Although overexpression of the full-length Bag3 did not affect Src activity, overexpression of either
Figure 1. Bag3 interacts with Src and this interaction is regulated by Hsp70. A, Bag3 interaction with SH3 domains of various proteins, see Results. B, left,coimmunoprecipitation of Bag3 with full-length Src in 293T cells. Relatively low abundance of Src in this cell line necessitated overexpression of Src forimmunoprecipitation as well as FLAG-tagged Bag3. Cells were collected and lysed 24 hours after transfection. Negative controls included GFP only(control) or Src only (-FL-Bag3). Middle, FL-Bag3 had expression of both FLAG-tagged Bag3 and Src. Immunoprecipitation was done using FLAG antibodyand blot was processed with TrueBlot secondary antibody. Right, coimmunoprecipitation of full-length Src with FLAG-tagged Bag3 in HeLa cells. Cellswere transfected with either FL-Bag3 or GFP (control) and immunoprecipitated using FLAG antibody. Association with Hsp70 is also shown (upperband indicated by arrow); lower band is the IgG heavy chain. C, schematic of Bag3 constructs used in this study, includingwild-type (FL), Bag domain deletion(DB), Bag domain point mutation (R480A), and PxxP domain deletion (DPxxP). All constructs contain an N-terminal FLAG tag. Domains are depictedincluding Bag domain (Hsp70 binding domain), PxxPmotif, M domain (small heat shock protein binding domain), andWWdomain. D, coimmunoprecipitationof Hsp70 with FLAG-tagged Bag3 constructs. HeLa cells were transfected with GFP (control) or indicated Bag3 constructs and immuniprecipitatedwith anti-FLAG antibody. Hsp70 is the upper band indicated by the arrow. The large lower band in the Hsp70 IP blot is the IgG heavy chain. E, GST pull downof Bag3 constructs with SH3 domain of Src. HeLa cells were transfected with indicated constructs, and after 24 hours, cell lysates were prepared andincubated with the purified GST–SH3 domain (from Src) bound to agarose beads or GST beads alone for 4 hours. Bag3 associated with the Src SH3 domainwas analyzed via immunoblot with anti-FLAG antibody.
Hsp70–Bag3 and Cancer Signaling
www.aacrjournals.org Cancer Res; 74(17) September 1, 2014 4733
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747

theDB or R480A constructs significantly activated endogenousSrc in either HeLa or MCF7 cells (Fig. 2C), further supportingthe model.
Hsp70–Bag3 module affects NF-kBUpon establishing the general mechanistic principle that
Hsp70 and Bag3 can cooperate in direct regulation of signalingfactors, we further sought to identify a spectrum of cancer-controlling signaling factors regulated by the Hsp70–Bag3module. We used a combination of two general criteria forsuch regulation: the signaling factor should be affected (i) byHsp70 and Bag3 depletion, and (ii) by overexpression of the DBor R480A Bag3 constructs but not of the normal Bag3.
In this search, we used our prior data, where we usedmicroarray analysis to compare gene expression in controland Hsp70-depleted MCF7 cells (Supplementary Table S1),followed by gene set enrichment analysis (GSEA). The GSEA-based signal pathway analysis software was developed by theBroad Institute (Cambridge, MA). According to their manual,pathways that show a false discovery rate (FDR) less than 0.25are significant enough to warrant validation. Our analysisindicated that a large number of pathways were predicted tobe up- or downregulated by Hsp70. For example, the protea-some and ubiquitin pathways were strongly upregulated andreceived the highest scores (see Supplementary Fig. S1A).
Another pathway that received a high score was the NF-kB(Fig. 3A) and related pathways, such as the IL1R, or TNFpathways (Supplementary Fig. S1B). Indeed, using a luciferasereporter under the control of NF-kB, we confirmed that NF-kBactivity was increased in Hsp70-depleted cells (Fig. 3B). Also,levels of IkBwere reduced, whereas phosphorylation of the p65subunit of NF-kB was elevated (Fig. 3C).
Here, we tested if Bag3 is involved in regulation of NF-kBby Hsp70. Accordingly, Bag3 was depleted in MCF7 cellsusing shRNA, and the activity of NF-kB was assessed usingthe luciferase reporter. Unlike Hsp70, depletion of Bag3 hadlittle apparent effect on the activation of NF-kB (Fig. 3B) orthe levels of phosphorylated NF-kB p65 subunit (p-S536)and of IkB (Fig. 3C). On the other hand, in cells with Bag3depletion, the ability of the knockdown of Hsp70 to activateNF-kB was strongly reduced (Fig. 3B). Therefore, as with Srcregulation, the Hsp70–Bag3 module is critical for regulationof NF-kB.
Notably, it was previously reported that that Bag3 regulatesNF-kB by controlling IKKg stability (37), but we were unableto detect any change in IKKg levels following Bag3 depletion(Fig. 3C). On the other hand, a recent proteomics study (38)indicated that Bag3 can directly interact with IkB, which isconsistent with our finding that the Hsp70–Bag3 moduleaffects IkB levels (Fig. 3B and C).
Figure 2. Hsp70 affects Srcactivation in cells. A, Hsp70depletion activates Src in a Bag3-dependent manner. MCF7 cellswere infected with control,shHsp70, shBag3, or dualshHsp70 and shBag3 retrovirusesand immunoblotted for activatedSrc (p-Y416). Values under p-Srcblot represent relativephosphorylation compared withcontrol. Levels of p-Src and totalSrc were normalized to b-actin andthe ratio of normalized p-Src/Src isreported. B, effects of shHsp70and shBag3 on the downstreamSrc targets. Cells were treated for30 minutes with 1 mmol/L sodiumorthovanadate before lysis. Valuesunder p-Paxillin and p-130Casrepresent relative phosphorylationcompared with control determinedin a similar manner as p-Src. C,expression of Bag3 mutants thatcannot bind to Hsp70 leads toactivation of Src. Bag3 constructsor GFP were transfected into HeLaor MCF7 cells. Cells were lysedafter 24 hours and immunoblottedfor activated Src (p-Y416).Representative blots of twoexperiments for each cell line areshown.
Colvin et al.
Cancer Res; 74(17) September 1, 2014 Cancer Research4734
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747
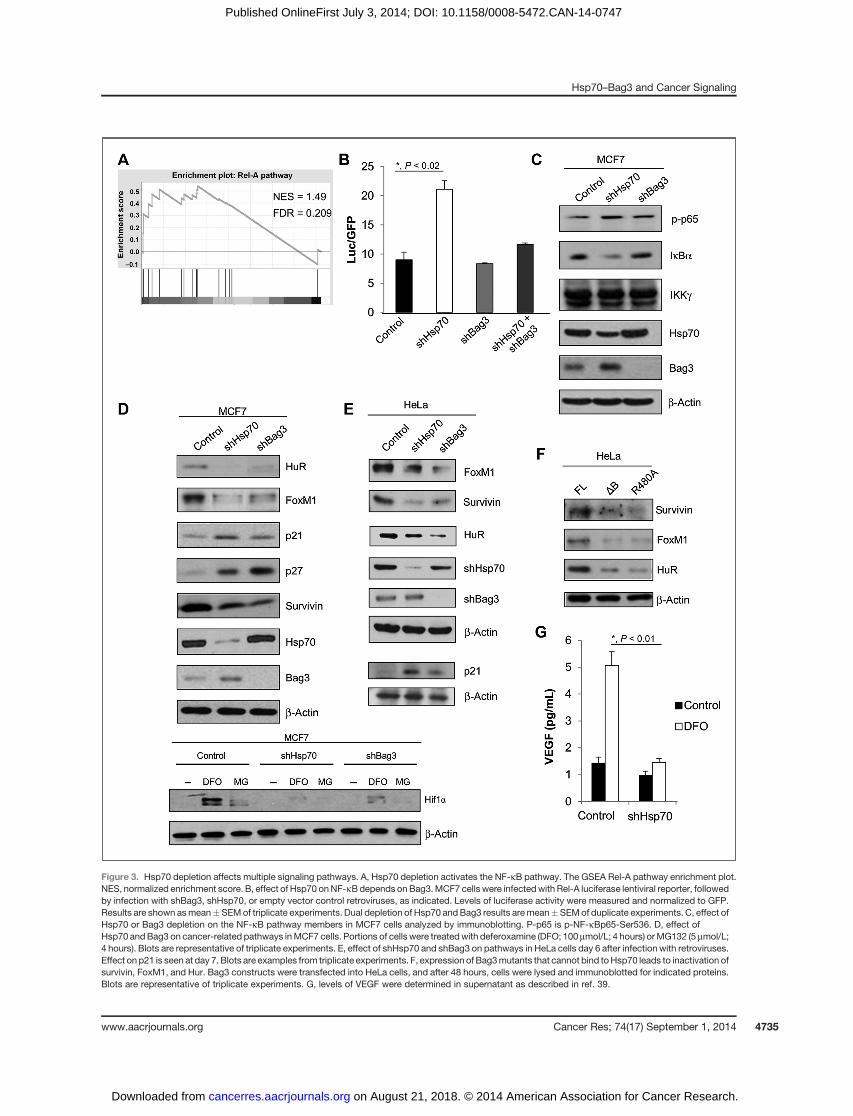
Figure 3. Hsp70 depletion affects multiple signaling pathways. A, Hsp70 depletion activates the NF-kB pathway. The GSEA Rel-A pathway enrichment plot.NES, normalized enrichment score. B, effect of Hsp70 onNF-kBdepends on Bag3.MCF7 cells were infectedwith Rel-A luciferase lentiviral reporter, followedby infection with shBag3, shHsp70, or empty vector control retroviruses, as indicated. Levels of luciferase activity were measured and normalized to GFP.Results are shown asmean�SEMof triplicate experiments. Dual depletion of Hsp70 and Bag3 results aremean�SEMof duplicate experiments. C, effect ofHsp70 or Bag3 depletion on the NF-kB pathway members in MCF7 cells analyzed by immunoblotting. P-p65 is p-NF-kBp65-Ser536. D, effect ofHsp70 andBag3 on cancer-related pathways inMCF7 cells. Portions of cells were treatedwith deferoxamine (DFO; 100 mmol/L; 4 hours) orMG132 (5 mmol/L;4 hours). Blots are representative of triplicate experiments. E, effect of shHsp70 and shBag3 on pathways in HeLa cells day 6 after infection with retroviruses.Effect on p21 is seen at day 7. Blots are examples from triplicate experiments. F, expression of Bag3mutants that cannot bind toHsp70 leads to inactivation ofsurvivin, FoxM1, and Hur. Bag3 constructs were transfected into HeLa cells, and after 48 hours, cells were lysed and immunoblotted for indicated proteins.Blots are representative of triplicate experiments. G, levels of VEGF were determined in supernatant as described in ref. 39.
Hsp70–Bag3 and Cancer Signaling
www.aacrjournals.org Cancer Res; 74(17) September 1, 2014 4735
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747

Effects of the Hsp70–Bag3 module on other cancer-related signaling factors
Because the unbiased approach with GSEA produced onlylimited information about signaling network regulated bythe Hsp70–Bag3 module, in further search for factors reg-ulated by this module, we used a more biased approachbased on hints obtained in our prior findings. Accordingly,we decided to test if the regulation of previously identifiedHsp70-controlled signaling factors also involved Bag3. Instudies of the OIS and tumor initiation, we reported thatdepletion of Hsp70 affects two important signaling proteinsthat control growth of cancer cells: the cell-cycle inhibitorp21 and the regulator of mitosis, survivin (5, 17). To test ifBag3 is also involved in control of these factors, we inves-tigated whether depletion of Bag3 in MCF7 using shRNAaffects levels of p21 and survivin. As with knockdown ofHsp70, Bag3 targeting resulted in decreased levels of survi-vin, and accumulation of p21 (Fig. 3D, top). Similar effects ofHsp70 and Bag3 depletion were seen in HeLa cells (Fig. 3E)and the colorectal carcinoma cell line, HCT116 (Supplemen-tary Fig. S2), demonstrating that these effects are related tomultiple cancer types. Bag3 depletion paralleled the effectsof Hsp70 knockdown in all three cell lines, further support-ing the idea that Hsp70 and Bag3 are coregulators of thesepathways. In line with this suggestion, overexpression of theDB and R480A Bag3 mutants, but not normal Bag3, in HeLacells mimicked effects of Hsp70 depletion on survivin (Fig.3F). In this experiment, we were unable to assess effects onBag3 mutants on p21, because levels of this protein in HeLacells were undetectable.
Because both p21 and survivin have a common transcrip-tional regulator, FoxM1, we tested whether the Hsp70–Bag3module can control this cancer-regulating factor. Indeed,depletion of either Hsp70 or Bag3 led to strong downregulationof FoxM1 (Fig. 3D, top) and upregulation of its distinct down-stream target, the cell-cycle inhibitor p27 (39). Furthermore,although the levels of FoxM1 were unaltered in normal Bag3and empty vector control conditions, the DB or R480A Bag3mutants caused strong downregulation of FoxM1 (Fig. 3F),indicating the importance of Hsp70–Bag3 interaction inexpression of this major transcription factor. Because FoxM1plays multiple roles in OIS, metastasis, and invasion (40), thesefindings further support a broad role for the Hsp70–Bag3module in tumorigenesis.
On the basis of our prior study that connected the stressresponse to the translation factor HuR, anothermajor player incancer development (41, 42), we also suggested that theHsp70–Bag3 module may affect this pathway. Depletion ofeither Hsp70 or Bag3 led to significant downregulation of HuR(Fig. 3D, top). Furthermore, although the levels of HuR wereunaltered in normal Bag3 and empty vector control conditions,the DB or R480A Bag3 mutants caused strong downregulationof HuR, indicating that interaction between Hsp70 and Bag3 isimportant for regulation of this major cancer regulator (Fig.3F). In addition, we considered whether one of HuR's trans-lational targets, Hif1a, which is yet another major transcrip-tion factor involved in cancer, might also be downregulated.Because levels of Hif1a depend on the availability of oxygen,
control, Hsp70-depleted, and Bag3-depleted cells were treatedwith either hypoxia mimetic deferoxamine, or proteasomalinhibitor MG-132, both of which stabilize Hif1a. Hsp70-deplet-ed or Bag3-depleted MCF7 cells had significantly reducedlevels of Hif1a compared with control cells (Fig. 3D, bottom).Furthermore, Hsp70 depletion strongly reduced expression of amajor Hif1 downstream target gene VEGF (Fig. 3G). Therefore,it seems that the entire signaling network that controls mul-tiple processes in cancer development, including growth,invasion, and angiogenesis, involving HuR, Hif1a, and VEGF,is regulated by Bag3 and Hsp70. It should be noted that onlywith Src did we identify a direct connection to the Hsp70–Bag3module biochemically. With other pathways, genetic data donot establish the direct interaction, and therefore these inter-actions could be mediated by additional factors that can bindto WW or PxxP motifs on Bag3.
In summary, the Hsp70–Bag3 module is involved in reg-ulating a complex network of pathways, including Src, NF-kB, FoxM1, survivin, p21, p27, HuR, Hif1a, and most likely,other pathways, all of which play a role in cancer develop-ment. When individually considered, these pathways couldhave cancer-promoting or cancer-repressing effects. Never-theless, we know that in breast cancer models, depletion ofHsp70 suppresses both tumor initiation and progression (5),indicating that the net outcome of this complex Hsp70-Bag3–controlled signaling network to be prooncogenic.
Disruption of the Hsp70–Bag3 complex by a smallmolecule mimics effects of Hsp70 depletion on signalingand cancer
Here, we predicted that blocking interaction betweenHsp70 and Bag3 could mimic depletion of Hsp70 and haveanticancer effects. We focused on an inhibitor of Hsp70,termed YM-1 (43, 44). To test if YM-1 might disrupt theHsp70–Bag3 interaction, we immobilized Hsp70 on beadsand measured its binding to fluorescently labeled Bag3 usinga flow cytometry protein interaction assay (FCPIA). YM-1strongly inhibited the interaction between Hsp70 and Bag3(IC50, �5 mmol/L; Fig. 4A). This activity was consistent withthe ability of YM-1 to bind to a conserved, allosteric pocketin Hsp70, which traps the chaperone in an ADP-bound formthat has weak affinity for Bag3 (43, 44). To test if YM-1 candisrupt the Hsp70–Bag3 interaction in cells, Hsp70 wasimmunoprecipitated from HeLa cells treated with YM-1 ora vehicle control, and immunoblotted for Bag3. YM-1 almostcompletely blocked binding of Hsp70 to Bag3 in cells (Fig.4B), indicating that YM-1 is a good chemical probe foraddressing the importance of the Hsp70–Bag3 interactionin cancer signaling.
Next, MCF7 and HeLa cells were treated with YM-1 and theactivities of signaling pathways were assessed. To determinethe appropriate dosage, a range of concentrations were tested(Supplementary Fig. S3). The concentrations of YM-1 used fortesting effects on signaling pathways are described in thelegend to Fig. 4, but represent the minimal dose necessary tosee an effect in the appropriate cell line. As predicted, YM-1diminished the FoxM1 and Hif1a pathways, while activatingSrc (Fig. 4C), further supporting a role for the Hsp70–Bag3
Colvin et al.
Cancer Res; 74(17) September 1, 2014 Cancer Research4736
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747
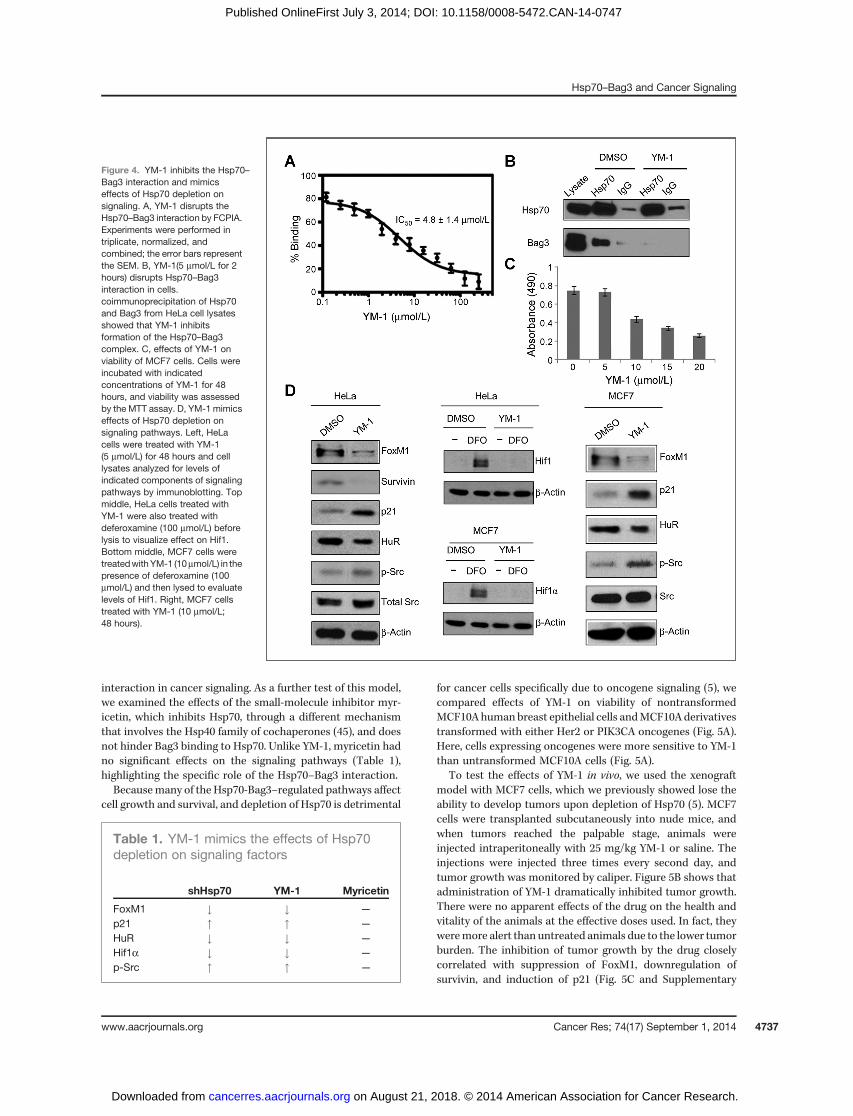
interaction in cancer signaling. As a further test of this model,we examined the effects of the small-molecule inhibitor myr-icetin, which inhibits Hsp70, through a different mechanismthat involves the Hsp40 family of cochaperones (45), and doesnot hinder Bag3 binding to Hsp70. Unlike YM-1, myricetin hadno significant effects on the signaling pathways (Table 1),highlighting the specific role of the Hsp70–Bag3 interaction.Becausemany of the Hsp70-Bag3–regulated pathways affect
cell growth and survival, and depletion of Hsp70 is detrimental
for cancer cells specifically due to oncogene signaling (5), wecompared effects of YM-1 on viability of nontransformedMCF10Ahumanbreast epithelial cells andMCF10Aderivativestransformed with either Her2 or PIK3CA oncogenes (Fig. 5A).Here, cells expressing oncogenes were more sensitive to YM-1than untransformed MCF10A cells (Fig. 5A).
To test the effects of YM-1 in vivo, we used the xenograftmodel with MCF7 cells, which we previously showed lose theability to develop tumors upon depletion of Hsp70 (5). MCF7cells were transplanted subcutaneously into nude mice, andwhen tumors reached the palpable stage, animals wereinjected intraperitoneally with 25 mg/kg YM-1 or saline. Theinjections were injected three times every second day, andtumor growth was monitored by caliper. Figure 5B shows thatadministration of YM-1 dramatically inhibited tumor growth.There were no apparent effects of the drug on the health andvitality of the animals at the effective doses used. In fact, theyweremore alert than untreated animals due to the lower tumorburden. The inhibition of tumor growth by the drug closelycorrelated with suppression of FoxM1, downregulation ofsurvivin, and induction of p21 (Fig. 5C and Supplementary
Figure 4. YM-1 inhibits the Hsp70–Bag3 interaction and mimicseffects of Hsp70 depletion onsignaling. A, YM-1 disrupts theHsp70–Bag3 interaction by FCPIA.Experiments were performed intriplicate, normalized, andcombined; the error bars representthe SEM. B, YM-1(5 mmol/L for 2hours) disrupts Hsp70–Bag3interaction in cells.coimmunoprecipitation of Hsp70and Bag3 from HeLa cell lysatesshowed that YM-1 inhibitsformation of the Hsp70–Bag3complex. C, effects of YM-1 onviability of MCF7 cells. Cells wereincubated with indicatedconcentrations of YM-1 for 48hours, and viability was assessedby the MTT assay. D, YM-1 mimicseffects of Hsp70 depletion onsignaling pathways. Left, HeLacells were treated with YM-1(5 mmol/L) for 48 hours and celllysates analyzed for levels ofindicated components of signalingpathways by immunoblotting. Topmiddle, HeLa cells treated withYM-1 were also treated withdeferoxamine (100 mmol/L) beforelysis to visualize effect on Hif1.Bottom middle, MCF7 cells weretreatedwithYM-1 (10mmol/L) in thepresence of deferoxamine (100mmol/L) and then lysed to evaluatelevels of Hif1. Right, MCF7 cellstreated with YM-1 (10 mmol/L;48 hours).
Table 1. YM-1 mimics the effects of Hsp70depletion on signaling factors
shHsp70 YM-1 Myricetin
FoxM1 # # —
p21 " " —
HuR # # —
Hif1a # # —
p-Src " " —
Hsp70–Bag3 and Cancer Signaling
www.aacrjournals.org Cancer Res; 74(17) September 1, 2014 4737
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747
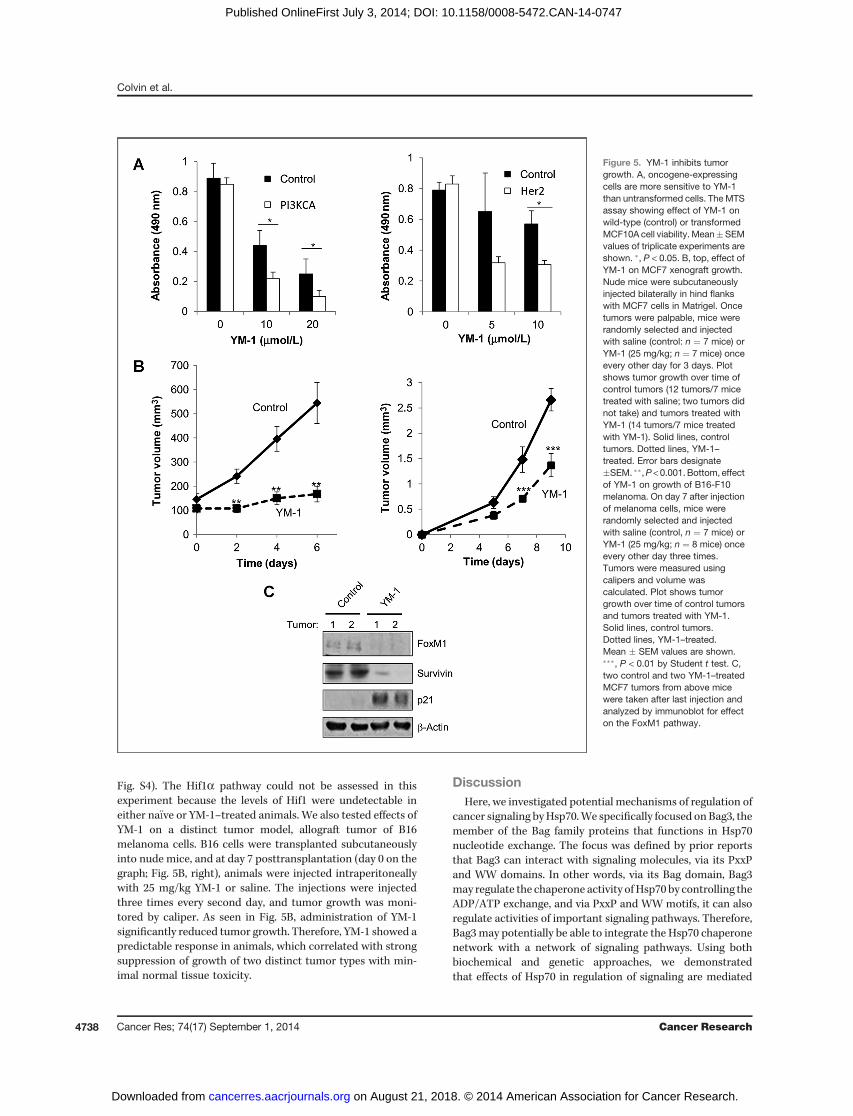
Fig. S4). The Hif1a pathway could not be assessed in thisexperiment because the levels of Hif1 were undetectable ineither na€�ve or YM-1–treated animals. We also tested effects ofYM-1 on a distinct tumor model, allograft tumor of B16melanoma cells. B16 cells were transplanted subcutaneouslyinto nude mice, and at day 7 posttransplantation (day 0 on thegraph; Fig. 5B, right), animals were injected intraperitoneallywith 25 mg/kg YM-1 or saline. The injections were injectedthree times every second day, and tumor growth was moni-tored by caliper. As seen in Fig. 5B, administration of YM-1significantly reduced tumor growth. Therefore, YM-1 showed apredictable response in animals, which correlated with strongsuppression of growth of two distinct tumor types with min-imal normal tissue toxicity.
DiscussionHere, we investigated potential mechanisms of regulation of
cancer signaling byHsp70.We specifically focused on Bag3, themember of the Bag family proteins that functions in Hsp70nucleotide exchange. The focus was defined by prior reportsthat Bag3 can interact with signaling molecules, via its PxxPand WW domains. In other words, via its Bag domain, Bag3may regulate the chaperone activity ofHsp70 by controlling theADP/ATP exchange, and via PxxP and WW motifs, it can alsoregulate activities of important signaling pathways. Therefore,Bag3 may potentially be able to integrate the Hsp70 chaperonenetwork with a network of signaling pathways. Using bothbiochemical and genetic approaches, we demonstratedthat effects of Hsp70 in regulation of signaling are mediated
Figure 5. YM-1 inhibits tumorgrowth. A, oncogene-expressingcells are more sensitive to YM-1than untransformed cells. The MTSassay showing effect of YM-1 onwild-type (control) or transformedMCF10A cell viability. Mean�SEMvalues of triplicate experiments areshown. �, P < 0.05. B, top, effect ofYM-1 on MCF7 xenograft growth.Nude mice were subcutaneouslyinjected bilaterally in hind flankswith MCF7 cells in Matrigel. Oncetumors were palpable, mice wererandomly selected and injectedwith saline (control: n ¼ 7 mice) orYM-1 (25 mg/kg; n ¼ 7 mice) onceevery other day for 3 days. Plotshows tumor growth over time ofcontrol tumors (12 tumors/7 micetreated with saline; two tumors didnot take) and tumors treated withYM-1 (14 tumors/7 mice treatedwith YM-1). Solid lines, controltumors. Dotted lines, YM-1–treated. Error bars designate�SEM. ��,P < 0.001. Bottom, effectof YM-1 on growth of B16-F10melanoma. On day 7 after injectionof melanoma cells, mice wererandomly selected and injectedwith saline (control, n ¼ 7 mice) orYM-1 (25 mg/kg; n ¼ 8 mice) onceevery other day three times.Tumors were measured usingcalipers and volume wascalculated. Plot shows tumorgrowth over time of control tumorsand tumors treated with YM-1.Solid lines, control tumors.Dotted lines, YM-1–treated.Mean � SEM values are shown.���, P < 0.01 by Student t test. C,two control and two YM-1–treatedMCF7 tumors from above micewere taken after last injection andanalyzed by immunoblot for effecton the FoxM1 pathway.
Colvin et al.
Cancer Res; 74(17) September 1, 2014 Cancer Research4738
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747

by Bag3. Surprisingly, whereas depletion of Bag3 had similareffects on some signaling pathways as depletion of Hsp70 (e.g.,on FoxM1 or Hif1a), in other cases, depletion of Bag3 did notaffect signaling but was necessary for Hsp70 effects (e.g., withNF-kB or Src). Therefore, in control of signaling pathways byBag3–Hsp70 pair, each individual target may be subject tospecific regulatory mechanisms. Although more work is need-ed, we suggest that these differences could be related tospecific ways in which the signaling modules interact withBag3. For example, whereas the Bag3 regulation of Src ismediated by the PxxP domain, the interaction with otherpathways could be through the WW or M domains.We undertook several unbiased and biased approaches to
dissect cancer-related pathways regulated by the Hsp70–Bag3module. We observed that Hsp70–Bag3 controls activities ofmultiple signaling factors that are involved in cancer develop-ment. The number of the factors was relatively large, consistentwith a broad "hub" role for Hsp70 in cancer. We validated asubset of these pathways, and found that Hsp70 depletiondownregulates Hif1a and FoxM1, whereas upregulating theNF-kB pathways. Interestingly, the former effects would pre-sumably be anticancerous, whereas the latter is likely to becancer-promoting. These disparate responses to the same stim-ulus should not be very surprising because similar phenomenahave been observed with other anticancer treatments. Forexample, radiation triggers the anticancer p53 pathway andalso cancer-promotingNF-kB and c-Abl (46, 47). In this case andin the case of Hsp70 depletion, the net effect is anticancerous.Previously, Hsp70 knockout was shown to prevent develop-
ment ofHer2-positive tumors in amousemodel by suppressingOIS (5), indicating that Hsp70 plays an important role at theearly stages of tumor development. Our findings that theHsp70–Bag3 module affects pathways regulating angiogenesisand invasion (e.g., Hif1a or FoxM1), suggest that Hsp70 mayplay a broader role in cancer development. Although it hasbeen reported that Hsp70 is seen at elevated levels in low-gradetumors (48) curiously, the levels of Hsp70 increase further inhigh-grade cancers (3, 4), consistent with distinct role(s) ofHsp70 in later stages of cancer progression, such as invasion.
Overall, this work pointed to a key role for the Hsp70–Bag3complex in cancer, and suggests that this pair may represent apromising drug target. Of the available Hsp70 inhibitors, YM-1is a unique probe because it specifically traps the ADP-boundform of Hsp70 (43). These observations suggested to us thatYM-1 might block the Hsp70–Bag3 interaction because Bag3has a weak affinity for the ADP-bound form of Hsp70 and itnormally favors client release (J.E. Gestwicki; manuscript inpreparation). Thus, although the known binding site for YM-1is more than 20 Å from the contact surface between Hsp70and Bag3, allosteric regulation of Hsp70 by YM-1 would bepredicted to interrupt nucleotide exchange factor binding.Accordingly, YM-1 has a mechanism of action that is uniqueamong the known Hsp70 inhibitors, which positions it todestabilize Hsp70–Bag3 activities in cancer cells, leading tosuppression of cancer-promoting signaling pathways. Consis-tently, YM-1 had anticancer activity in cells and in a xenograftmodel. These results validate the Hsp70–Bag3 complex as apotential anticancer target.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: T.A. Colvin, S.K. Calderwood, S. Takayama,M.Y. ShermanDevelopment of methodology: T.A. Colvin, V.L. GabaiAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): T.A. Colvin, V.L. Gabai, J. Gong, S. Gummuluru,O.N. Matchuk, S.G. Smirnova, N.V. Orlova, I. A. Zamulaeva, M. Garcia-Marcos,X. Li, J.N. Rauch, S. TakayamaAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): T.A. Colvin, V.L. Gabai, H. Li, X. Li, J.E. Gestwicki,S. TakayamaWriting, review, and/or revision of the manuscript: T.A. Colvin, V.L. Gabai,X. Li, Z.T. Young, J.E. Gestwicki, M.Y. ShermanAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): X. Li, Z.T. YoungStudy supervision: J.E. Gestwicki, M.Y. Sherman
The costs of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received March 20, 2014; revised May 9, 2014; accepted May 26, 2014;published OnlineFirst July 3, 2014.
References1. Michels AA, Kanon B, Konings AW, Ohtsuka K, Bensaude O,
Kampinga HH. Hsp70 and Hsp40 chaperone activities in the cyto-plasm and the nucleus of mammalian cells. J Biol Chem 1997;272:33283–9.
2. StegeGJ, Li GC, Li L, KampingaHH, Konings AW.On the role of hsp72in heat-induced intranuclear protein aggregation. Int J Hyperthermia1994;10:659–74.
3. Mosser DD, Morimoto RI. Molecular chaperones and the stress ofoncogenesis. Oncogene 2004;23:2907–18.
4. Calderwood SK, Khaleque MA, Sawyer DB, Ciocca DR. Heat shockproteins in cancer: chaperones of tumorigenesis. Trends Biochem Sci2006;31:164–72.
5. Meng L, Hunt C, Yaglom JA, Gabai VL, Sherman MY. Heat shockprotein Hsp72 plays an essential role in Her2-induced mammarytumorigenesis. Oncogene 2011;30:2836–45.
6. Hunt CR, Dix DJ, Sharma GG, Pandita RK, Gupta A, Funk M, et al.Genomic instability and enhanced radiosensitivity in Hsp70.1- andHsp70.3-deficient mice. Mol Cell Biol 2004;24:899–911.
7. Wang Z, Gall JM, Bonegio RG, Havasi A, Hunt CR, Sherman MY, et al.Inductionof heat shockprotein 70 inhibits ischemic renal injury. KidneyInt 2011;79:861–70.
8. McConnell KW, Fox AC, Clark AT, Chang NY, Dominguez JA, FarrisAB, et al. The role of heat shockprotein 70 inmediating age-dependentmortality in sepsis. J Immunol 2011;186:3718–25.
9. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation.Cell 2011;144:646–74.
10. Kim YN, Koo KH, Sung JY, Yun UJ, Kim H. Anoikis resistance: anessential prerequisite for tumor metastasis. Int J Cell Biol 2012;2012:306879.
11. Luo J, Solimini NL, Elledge SJ. Principles of cancer therapy: oncogeneand non-oncogene addiction. Cell 2009;136:823–37.
12. Garrido C, Brunet M, Didelot C, Zermati Y, Schmitt E, Kroemer G. Heatshock proteins 27 and 70: anti-apoptotic proteins with tumorigenicproperties. Cell Cycle 2006;5:2592–601.
13. Beere H, Wolf B, Cain K, Mosser DD, Mahboubi A, Kuwana T, et al.Heat shock protein 70 inhibits apoptosis by preventing recruitment
www.aacrjournals.org Cancer Res; 74(17) September 1, 2014 4739
Hsp70–Bag3 and Cancer Signaling
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747

of procaspase-9 to the Apaf-1 apoptosome. Nat Cell Biol 2000;2:469–75.
14. Itahana K, Campisi J, Dimri GP. Mechanisms of cellular senescence inhuman and mouse cells. Biogerontology 2004;5:1–10.
15. Shay JW, Roninson IB. Hallmarks of senescence in carcinogenesisand cancer therapy. Oncogene 2004;23:2919–33.
16. Yaglom JA, Gabai VL, Sherman MY. High levels of heat shock proteinHsp72 in cancer cells suppress default senescence pathways. CancerRes 2007;67:2373–81.
17. Gabai VL, Yaglom JA, Waldman T, Sherman MY. Heat shock proteinHsp72 controls oncogene-induced senescence pathways in cancercells. Mol Cell Biol 2009;29:559–69.
18. Rerole AL, Gobbo J, De Thonel A, Schmitt E, Pais de Barros JP,Hammann A, et al. Peptides and aptamers targeting HSP70: a novelapproach for anticancer chemotherapy. Cancer Res 2011;71:484–95.
19. Strom E, Sathe S, Komarov PG, Chernova OB, Pavlovska I, Shy-shynova I, et al. Small-molecule inhibitor of p53 binding to mitochon-dria protects mice from gamma radiation. Nat Chem Biol 2006;2:474–9.
20. Leu JI, Pimkina J, Frank A, Murphy ME, George DL. A smallmolecule inhibitor of inducible heat shock protein 70. Mol Cell2009;36:15–27.
21. Leu JI, Pimkina J, Pandey P,MurphyME, George DL. HSP70 inhibitionby the small-molecule 2-phenylethynesulfonamide impairs proteinclearance pathways in tumor cells. Mol Cancer Res 2011;9:936–47.
22. Miyata Y, Rauch JN, Jinwal UK, Thompson AD, Srinivasan S, DickeyCA, et al. Cysteine reactivity distinguishes redox sensing by the heat-inducible and constitutive forms of heat shock protein 70. Chem Biol2012;19:1391–9.
23. Massey AJ,Williamson DS, Browne H,Murray JB, Dokurno P, Shaw T,et al. A novel, small molecule inhibitor of Hsc70/Hsp70 potentiatesHsp90 inhibitor induced apoptosis in HCT116 colon carcinoma cells.Cancer Chemother Pharmacol 2010;66:535–45.
24. Huryn DM, Brodsky JL, Brummond KM, Chambers PG, Eyer B, IrelandAW, et al. Chemical methodology as a source of small-moleculecheckpoint inhibitors and heat shock protein 70 (Hsp70) modulators.Proc Natl Acad Sci U S A 2011;108:6757–62.
25. Rodina A, Patel PD, Kang Y, Patel Y, Baaklini I, Wong MJ, et al.Identification of an allosteric pocket on human hsp70 reveals a modeof inhibition of this therapeutically important protein. Chem Biol2013;20:1469–80.
26. Takayama S, Reed JC. Molecular chaperone targeting and regulationby BAG family proteins. Nat Cell Biol 2001;3:E237–41.
27. Ingham RJ, Colwill K, Howard C, Dettwiler S, Lim CS, Yu J, et al. WWdomains provide a platform for the assembly of multiprotein networks.Mol Cell Biol 2005;25:7092–106.
28. Li SS. Specificity and versatility of SH3 and other proline-recognitiondomains: structural basis and implications for cellular signal transduc-tion. Biochem J 2005;390(Pt 3):641–53.
29. Carra S, Seguin SJ, Lambert H, Landry J. HspB8 chaperone activitytoward poly(Q)-containing proteins depends on its association withBag3, a stimulator ofmacroautophagy. J Biol Chem2008;283:1437–44.
30. Ulbricht A, Eppler FJ, Tapia VE, van der Ven PF, Hampe N, Hersch N,et al. Cellular mechanotransduction relies on tension-induced andchaperone-assisted autophagy. Curr Biol 2013;23:430–5.
31. Gamerdinger M, Kaya AM, Wolfrum U, Clement AM, Behl C. BAG3mediates chaperone-based aggresome-targeting and selectiveautophagy of misfolded proteins. EMBO Rep 2011;12:149–56.
32. Doong H, Price J, Kim YS, Gasbarre C, Probst J, Liotta LA, et al. CAIR-1/BAG-3 forms anEGF-regulated ternary complexwith phospholipaseC-gamma and Hsp70/Hsc70. Oncogene 2000;19:4385–95.
33. Iwasaki M, Homma S, Hishiya A, Dolezal SJ, Reed JC, Takayama S.BAG3 regulatesmotility and adhesionof epithelial cancer cells. CancerRes 2007;67:10252–9.
34. Kassis JN, Guancial EA, Doong H, Virador V, Kohn EC. CAIR-1/BAG-3modulates cell adhesion and migration by downregulating activity offocal adhesion proteins. Exp Cell Res 2006;312:2962–71.
35. SuzukiM, IwasakiM, Sugio A, Hishiya A, TanakaR, EndoT, et al. BAG3(BCL2-associated athanogene 3) interacts with MMP-2 to positivelyregulate invasion by ovarian carcinoma cells. Cancer Lett 2011;303:65–71.
36. Gentilella A, Khalili K. BAG3 expression in glioblastoma cells promotesaccumulation of ubiquitinated clients in an Hsp70-dependent manner.J Biol Chem 2011;286:9205–15.
37. Ammirante M, Rosati A, Arra C, Basile A, Falco A, Festa M, et al. IKK{gamma} protein is a target of BAG3 regulatory activity in human tumorgrowth. Proc Natl Acad Sci U S A 2010;107:7497–502.
38. ChenY, Yang LN,Cheng L, Tu S,GuoSJ, LeHY, et al. Bcl2-associatedathanogene 3 interactome analysis reveals a new role in modulatingproteasome activity. Mol Cell Proteomics 2013;12:2804–19.
39. Wang IC, Chen YJ, Hughes D, Petrovic V, Major ML, Park HJ, et al.Forkhead box M1 regulates the transcriptional network of genesessential for mitotic progression and genes encoding the SCF(Skp2-Cks1) ubiquitin ligase. Mol Cell Biol 2005;25:10875–94.
40. Halasi M, Gartel AL. FOX(M1) news—it is cancer. Mol Cancer Ther2013;12:245–54.
41. Gabai VL, Meng L, Kim G, Mills TA, Benjamin IJ, Sherman MY. Heatshock transcription factor Hsf1 is involved in tumor progression viaregulation of hypoxia-inducible factor 1 andRNA-binding protein HuR.Mol Cell Biol 2012;32:929–40.
42. Abdelmohsen K, Gorospe M. Posttranscriptional regulation of cancertraits by HuR. Wiley Interdiscip Rev RNA 2010;1:214–29.
43. Rousaki A, Miyata Y, Jinwal UK, Dickey CA, Gestwicki JE, ZuiderwegER. Allosteric drugs: the interaction of antitumor compound MKT-077with human Hsp70 chaperones. J Mol Biol 2011;411:614–32.
44. Wang AM, Miyata Y, Klinedinst S, Peng HM, Chua JP, Komiyama T,et al. Activation of Hsp70 reduces neurotoxicity by promoting poly-glutamine protein degradation. Nat Chem Biol 2013;9:112–8.
45. Chang L, Miyata Y, Ung PM, Bertelsen EB, McQuade TJ, Carlson HA,et al. Chemical screens against a reconstituted multiprotein complex:myricetin blocks DnaJ regulation of DnaK through an allosteric mech-anism. Chem Biol 2011;18:210–21.
46. ChenX,ShenB, Xia L,KhaletzkiyA,ChuD,WongJY, et al. Activationofnuclear factor kappaB in radioresistance of TP53-inactive humankeratinocytes. Cancer Res 2002;62:1213–21.
47. Meltser V, Ben-YehoyadaM, Shaul Y. c-Abl tyrosine kinase in the DNAdamage response: cell death andmore. Cell Death Differ 2011;18:2–4.
48. Ciocca DR, Calderwood SK. Heat shock proteins in cancer: diagnos-tic, prognostic, predictive, and treatment implications. Cell StressChaperones 2005;10:86–103.
Cancer Res; 74(17) September 1, 2014 Cancer Research4740
Colvin et al.
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747

2014;74:4731-4740. Published OnlineFirst July 3, 2014.Cancer Res Teresa A. Colvin, Vladimir L. Gabai, Jianlin Gong, et al. Networks
Bag3 Interactions Regulate Cancer-Related Signaling−Hsp70
Updated version
10.1158/0008-5472.CAN-14-0747doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2014/07/21/0008-5472.CAN-14-0747.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/74/17/4731.full#ref-list-1
This article cites 48 articles, 19 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/74/17/4731.full#related-urls
This article has been cited by 11 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/74/17/4731To request permission to re-use all or part of this article, use this link
on August 21, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2014; DOI: 10.1158/0008-5472.CAN-14-0747