human brain: proton diffusion mr spectroscopy' - meteore service
TRANSCRIPT

Human Brain: Proton DiffusionMR Spectroscopy’
Stefan Posse, PhD #{149}Charles A. Cuenod, MD #{149}Denis Le Bihan, MD, PhD
719
Diffusion of brain metabolites wasmeasured in 10 healthy volunteers byusing localized proton diffusion mag-netic resonance (MR) spectroscopy.Measurements were conducted witha clinical MR imager by using a stim-ulated-echo pulse sequence (3,000/60[repetition time msec/echo timemsecl, 200-msec mixing time) withadditional outside-volume suppres-sion. Motion artifacts due to macro-scopic brain movements were com-pensated by means of peripheralcardiac gating and separate collectionof individual spectroscopic acquisi-tions into a two-dimensional datamatrix. Phase errors due to macro-scopic motion were subsequently cor-rected in individual data traces priorto spectral averaging. Mean (±1 stan-dard deviation) apparent diffusioncoefficients of choline-containingcompounds ([0.13 ± 0.031 x i0� i�i.n�2/sec), creatine and phosphocreatine([0.15 ± 0.03] x iO� mm2!sec), andN-acetyl aspartate ([0.18 ± 0.02J x iO�mm2,/sec) were substantially smallerthan that of water and were consis-tent with recently published data ob-tamed in anesthetized and paralyzedanimals. Adequate diffusion sensi-tivity for metaboiites in the humanbrain can be obtained with clinicalwhole-body imagers despite macro-scopic head and brain movements.
Index terms: Brain, perfusion, 13.12144,
13.12145, 15.12144, 15.12145 #{149}Magnetic reso-
nance (MR), diffusion study, 13.12144,15.12144 #{149}Magnetic resonance (MR), spectnos-
copy, 13.12145, 15.12145 #{149}Magnetic resonance(MR), tissue characterization
Radiology 1993; 188:719-725
M AGNETIC resonance (MR) imag-
ing of water self-diffusion mayprovide useful information about tis-sue structure and function, as diffu-sion coefficients in vivo are related to
cellular parameters such as dimension-ality, compartmentation, and trans-
port processes (1-3). For instance, dif-fusion MR imaging provides valuable
information about white matter fiberorientation in the central nervous sys-tem (4,5) on the basis of anisotnopicdiffusion. Diffusion MR imaging alsoappears promising in the evaluationand management of brain infanctionsat a very early stage when findings ofother modalities, including Ti- andT2-weighted MR imaging, are stillnormal (6,7).
To date, however, diffusion mea-sunements in humans have been pen-
formed only on water, for reasons oftechnical feasibility. Water may not bethe most suitable molecule for diffu-sion measurements because of its
ubiquity and the permeability of mosttissue interfaces, such as membranes,
to water molecules. In this context,diffusion measurements of more com-
plex molecules that are tissue-specificappear to be promising for tissuecharacterization.
Such measurements are technically
more challenging, because such mote-
cules have a tower concentration than
that of water. Recently, initial diffu-sion measurements of selected metab-
otites in vivo have been reported in
brain and muscle tissue of anesthe-tized and paralyzed animals by usingboth localized phosphorus-31 andhydrogen-i spectroscopy (8-10). Themagnitudes and directional amsotropyof diffusion coefficients of molecules
such as choline-containing compounds
(Cho), creatine and phosphocreatine(Cn-PCn), and N-acetyl aspartate(NAA) were found to be consistent
with cell dimensions and orientations
(8,9).Applications of diffusion spectros-
copy in humans have been less suc-
cessful, probably because of the lackof adequate motion-compensation
methods. Macroscopic motion due toinvoluntary subject movements, nes-piration, and cerebrospinal fluid-re-
tated brain pulsations may lead to anoverestimation of diffusion coeffi-cients in humans (11). Gradient com-pensation schemes to reduce artifacts
from linear movements have beenproposed (12,13). They require, how-
even, substantially longer diffusiontimes to compensate for the loss in
diffusion sensitivity (13). This require-
ment may be disadvantageous if dif-
fusional restriction, which requiresshorten diffusion times, is to be inves-
tigated. In addition, signal losses dueto relaxation during the pulse se-
quence are increased. Shorten diffu-
sion times are achievable with small-
bore imagers, in which much stronger
gradient strengths are available.The purpose of this study was to
demonstrate the feasibility of spectro-scopic diffusion measurements of
Cho, Cn-PCr, and NAA in humanbrain tissue with a clinical whole-body imager. A stimulated-echo pulse
sequence with complete outen-vol-
ume presaturation (14) to improvevolume localization is designed toincrease diffusion sensitivity while
limiting motion artifacts from headmovements and cardiac-related brainpulsations. Signal losses due to mac-
I From the Department of Diagnostic Radiology, Warren Grant Magnuson Clinical Center (S.P.,D.L.B.), and Laboratory of Diagnostic Radiology Research (C.A.C.), National Institutes of Health,Bldg 10, Rm 1C660, Bethesda, MD 20892. From the 1992 RSNA scientific assembly. Received January14, 1993; revision requested March 4; revision received April 12; accepted April 16. S.P. supported bythe Swiss National Science Foundation. Address reprint requests to S.P.
C RSNA, 1993
Abbreviations: CHESS = chemical shift-selec-tive, Cho = choline-containing compounds,Cr-PCr = creatine and phosphocreatine, NAA =
N-acetyl aspartate, STEAM = stimulated-echoacquisition mode, TE = echo time, TM = mixingtime.

H
Rn
Gradient
TE/2
S.’�
TE/2
0’
g/2 �r/2 x/2
n� 0’Q 0’
TE/2 TE/2TM
C
noscopic motion are reduced by
means of cardiac gating and compen-sating motion-induced phase errors
in individual acquisitions. Resultsobtained in 10 healthy volunteers
demonstrate that adequate diffusion
sensitivity and motion artifact com-pensation can be achieved in con-
scious human subjects.
.0 0
8
Diffusion
720 #{149}Radiology September 1993
Figure 1. Pulse sequences for diffusion measurements and effects of � a
motion on the echo signal. a, Spin-echo sequence. b, Stimulated-echo
sequence. c, Diffusion gradients are applied symmetrically in the TE/2
(half echo time) periods. 6 is the duration of the diffusion gradients. � is
the time between the two diffusion gradients. The arrows in d-f depict
the spin magnetization in the transverse plane after excitation (left), af-ter the first diffusion gradient (center), and at the top of the echo (right).
d, With pure diffusion the signal is dephased, but the net phase is un- b
changed. C, In the case of purely linear motion along the direction of the
diffusion gradients, the signal amplitude is unchanged but the signal
phase changes.f, With other forms of motion, both signal amplitude and
phase changes occur.
MATERIALS AND METHODS
Theory
Conventional spin-echo and stimulated-
echo MR imaging can be sensitized to dif-
fusion by symmetrically applying gradient
pulses that attenuate the echo signal S
(Fig 1). Assuming isotropic and unre-
stricted diffusion, the diffusion coefficient,
D, can be determined by measuring the
signal attenuation as a function of the gra-
dient factor b, which is dependent on the
radio frequency and gradient pulse se-
quence and increases with increasing gra-
dient strength and pulse sequence timing
(3,15):
S = 5o � exp’�,
where S� is the signal intensity without
diffusion. In case of restricted diffusion
(eg, due to cellular compartmentation) the
measured diffusion coefficient becomes
dependent on the pulse sequence timing.The diffusion coefficient is related to theroot mean square displacement (�[x - x0]2)in a given spatial direction by �‘(x - x0)2 =
��2Dtd1ff, where td*ft �5 the diffusion time. For
the pulse sequences in Figure 1, td,ff is de-
fined as (� - 8/3). For unrestricted diffu-
sion (x - x,)2 increases linearly with tdjft. In
the case of motion restriction the relation-ship is nonlinear, with a leveling off where
(x - Xo)2 reaches the size of the restrictingcompartment while the measured diffu-
sion coefficient D decreases. The average
signal phase shift due to pure diffusion is
zero, since there is no net displacement ofthe spins under observation (Fig id).
By contrast, if net displacements of
spins in the presence of field gradients are
superimposed on diffusion, further signal
phase changes and amplitude losses may
occur in addition to amplitude losses due
to diffusion (Fig 1, e,f). The additional ef-fect of linear bulk motion (ie, along agiven spatial direction) on the signal
phase and amplitude can be determinedby integrating the spin signals at the time
UnearMotion
f
NonlinearMotion
Sof the echo formation (echo time [TE]) ne-
glecting relaxation and field inhomogene-
ity effects: S = fff p(r’)exp’#{176}�’1dr’, wherep(r’� is the local spin density and 0(r) =
-Yfo E G(t’)r”(t’)dt’.
The -y represents the gyromagnetic ra-
(1) tio. G(t’) is the effective gradient sequencein the direction of the movement as a
function of time. Gradients prior to the
refocusing pulse or pulses give negative
contributions. r”(t’) is the trajectory of the
spin during the pulse sequence. For sta-
tionary nondiffusing spins the pulse se-
quence is symmetric and the spins arecompletely refocused at the center of theecho. For linear bulk motion (coherent
translations along a given spatial direc-tion) the phase dispersion within the mov-ing object is one-dimensional. During thepulse sequence the relative phase differ-
ence between any two points in the objectis independent of the object position with
respect to the gradient fields. Thus, thephase dispersion is also completely nefo-
cused at the center of the echo. The net
phase of the total echo signal, however, isshifted in a way dependent on the applied
gradient strength, the pulse sequence tim-
ing, and the object velocity similar to that
in phase-contrast angiography. For very
small velocities this phase shift is directly
proportional to the object velocity. Thus,
the result of coherent translations is a netphase shift without signal losses (Fig 1, e).During signal averaging, which is usuallynecessary with MR spectroscopy to in-
crease the signal-to-noise ratio, such phase
shifts, usually incoherent from acquisition
ED
Before Phase Correction
Figure 2. Motion artifact compensation inlocalized spectroscopy. Top: Motion-related
phase errors in individual acquisitions, which
are usually incoherent, lead to signal loss ataveraging. Bottom: Phase correction of mdi-vidual data traces restores phase coherenceand avoids signal loss.
to acquisition, lead to signal losses. Bulkrotations, by contrast, lead to phase errorsand signal losses in individual acquisi-tions, since the gradient directions relativeto orientation of the volume of interestchange during the pulse sequence, lead-ing to incomplete refocusing of the phasedispersion imposed by the initial gradientpulses. Similarly, more complex forms of
macroscopic incoherent motion such as
turbulence and shear lead to phase errorsand signal losses as well (Fig 1,f). In vivo,such macroscopic motion-related signal
After Phase Correction
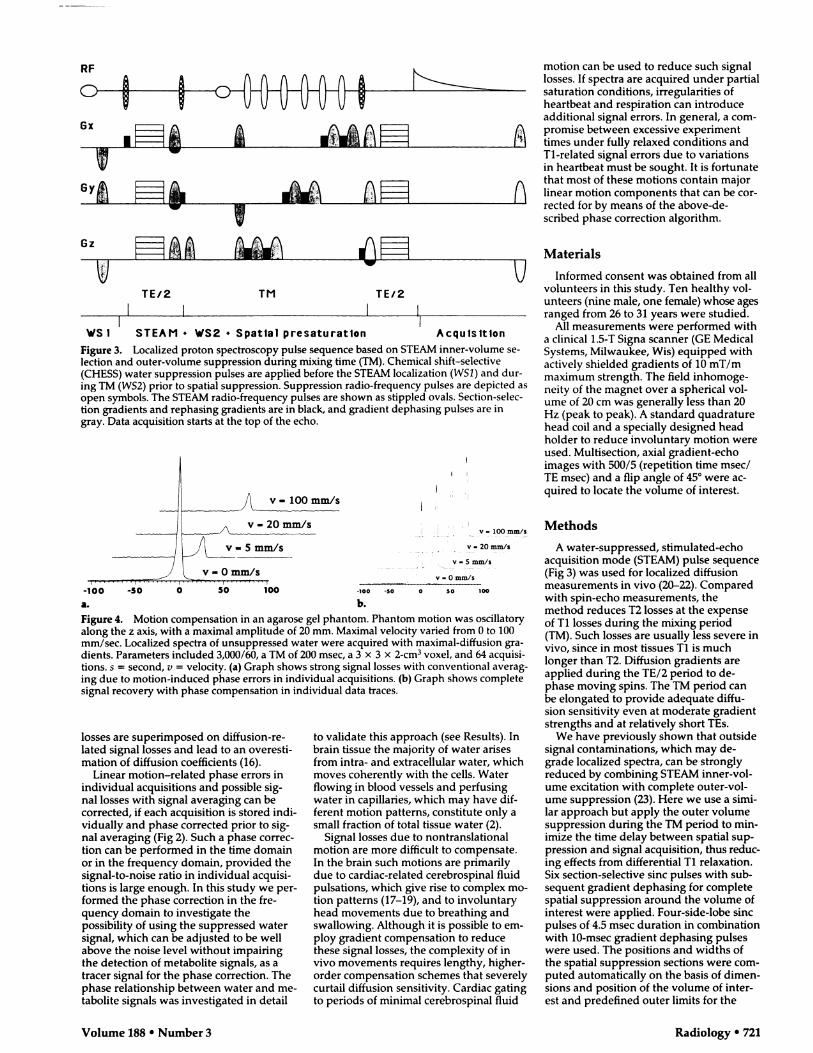
- � ,
Q\� r\
Gz
�) UTE/2 TM TE/2
.1 I I
ws 1 STEAM . WS2 #{149}Spatial presaturation Acquisition
Figure 3. Localized proton spectroscopy pulse sequence based on STEAM inner-volume se-
lection and outer-volume suppression during mixing time (TM). Chemical shift-selective(CHESS) water suppression pulses are applied before the STEAM localization (WSI) and dun-
ing TM (WS2) prior to spatial suppression. Suppression radio-frequency pulses are depicted as
open symbols. The STEAM radio-frequency pulses are shown as stippled ovals. Section-selec-tion gradients and rephasing gradients are in black, and gradient dephasing pulses are ingray. Data acquisition starts at the top of the echo.
v-lOOmm/s � � �
v-20m.m/s � � �-.---------------------- -� HV - 5 mm/s � � � � . � � Y 20 mm/s- � � �
-� v-Omm/s
-100 -50 0
V - 0 mm/s
5’O’ � � 100 -100 -SO 0 50 100
a. b.
Figure 4. Motion compensation in an aganose gel phantom. Phantom motion was oscillatoryalong the z axis, with a maximal amplitude of 20 mm. Maximal velocity varied from 0 to 100mm/sec. Localized spectra of unsuppressed water were acquired with maximal-diffusion gra-dients. Parameters included 3,000/60, a TM of 200 msec, a 3 x 3 x 2-cm3 voxel, and 64 acquisi-
tions. S = second, v = velocity. (a) Graph shows strong signal losses with conventional averag-
ing due to motion-induced phase errors in individual acquisitions. (b) Graph shows complete
signal recovery with phase compensation in individual data traces.
Volume 188 #{149}Number 3 Radiology #{149}721
Ox
losses are superimposed on diffusion-re-lated signal losses and lead to an overesti-mation of diffusion coefficients (16).
Linear motion-related phase errors inindividual acquisitions and possible sig-nat losses with signal averaging can becorrected, if each acquisition is stoned mdi-vidualty and phase connected prior to sig-nal averaging (Fig 2). Such a phase correc-tion can be performed in the time domainor in the frequency domain, provided thesignal-to-noise ratio in individual acquisi-tions is large enough. In this study we per-formed the phase correction in the fre-quency domain to investigate thepossibility of using the suppressed watersignal, which can be adjusted to be wellabove the noise level without impairingthe detection of metabolite signals, as atracer signal for the phase correction. Thephase relationship between water and me-tabotite signals was investigated in detail
to validate this approach (see Results). Inbrain tissue the majority of water arisesfrom intra- and extnaceltulan water, whichmoves coherently with the cells. Water
flowing in blood vessels and perfusingwater in capillaries, which may have dif-fenent motion patterns, constitute only asmall fraction of total tissue water (2).
Signal losses due to nontranslational
motion are more difficult to compensate.In the brain such motions are primarilydue to cardiac-related cerebrospinal fluidpulsations, which give rise to complex mo-tion patterns (17-19), and to involuntaryhead movements due to breathing andswallowing. Although it is possible to em-ploy gradient compensation to reducethese signal losses, the complexity of invivo movements requires lengthy, higher-order compensation schemes that severelycurtail diffusion sensitivity. Cardiac gatingto periods of minimal cerebrospinal fluid
motion can be used to reduce such signallosses. If spectra are acquired under partial
- saturation conditions, irregularities ofheartbeat and respiration can introduceadditional signal errors. In general, a com-
:�c promise between excessive experiment�t times under fully relaxed conditions and
Ti-related signal errors due to variationsin heartbeat must be sought. It is fortunatethat most of these motions contain major
linear motion components that can be con-rected for by means of the above-de-
scribed phase correction algorithm.
Materials
Informed consent was obtained from allvolunteers in this study. Ten healthy vol-
unteens (nine male, one female) whose ages
ranged from 26 to 31 years were studied.Alt measurements were performed with
a clinical 1.5-T Signa scanner (GE MedicalSystems, Milwaukee, Wis) equipped with
actively shielded gradients of 10 mT/rn
maximum strength. The field mnhornoge-neity of the magnet over a spherical vol-
ume of 20 cm was generally less than 20Hz (peak to peak). A standard quadraturehead coil and a specially designed headholder to reduce involuntary motion wereused. Muttisection, axial gradient-echoimages with 500/5 (repetition time msec/TE rnsec) and a flip angle of 45#{176}were ac-qumred to locate the volume of interest.
Methods
A water-suppressed, stimulated-echoacquisition mode (STEAM) pulse sequence(Fig 3) was used for localized diffusionmeasurements in vivo (20-22). Comparedwith spin-echo measurements, themethod reduces T2 losses at the expenseof TI losses during the mixing period(TM). Such tosses are usually less severe in
vivo, since in most tissues Ti is muchlonger than T2. Diffusion gradients areapplied during the TE/2 period to de-phase moving spins. The TM period canbe elongated to provide adequate diffu-sion sensitivity even at moderate gradientstrengths and at relatively short TEs.
We have previously shown that outsidesignal contaminations, which may de-grade localized spectra, can be stronglyreduced by combining STEAM inner-vol-ume excitation with complete outer-vol-ume suppression (23). Here we use a simi-lan approach but apply the outer volumesuppression during the TM period to mm-imize the time delay between spatial sup-pression and signal acquisition, thus reduc-ing effects from differential Ti relaxation.Six section-selective sinc pulses with sub-sequent gradient dephasing for completespatial suppression around the volume ofinterest were applied. Four-side-lobe sincpulses of 4.5 msec duration in combination
with 10-msec gradient dephasing pulseswere used. The positions and widths ofthe spatial suppression sections were com-puted automatically on the basis of dimen-sions and position of the volume of inter-est and predefined outer limits for the

4 3 2
ppm
Volunteer I
SCeC
S
C
U)
I 0#{176}
10#{176}
#{163}.-.4-_._
� #{149}
.,._e____e- .__.
e e #{149}���-_�._- C-S
-#{149} -Cho
- . -Cr/PCr
. -a.- NAA
500 1000 1500 2000 2500
bfactor (s/mn?)
722 #{149}Radiology September 1993
Figure 5. Axial GRASS localizer. Spectro-
scopic volume selection in white matter is
indicated by the box. The pulse sequence
parameters for this MR image were 600/5,
with a flip angle of 45#{176}.
Figure 6. Localized proton spectra of human brain tissue acquired
with 11 different diffusion gradient strengths. Spectra displayingmajor resonances from Cho (3.24 ppm), Cn-PCr (3.02 ppm), andNAA (2.01 ppm) are plotted with decreasing diffusion gradient
strengths (10-0 mT/m in 1 mT/m steps). Spectral artifacts are negli-
gible even with maximum diffusion gradients applied. Parameterswere 3,000/60, a TM of 200 msec, 3 x 3 x 2-cm3 voxel, and 64 ac-
quisitions.
presaturation sections. A gap of 5 mm be-
tween the suppression sections and thevolume of interest reduced signal lossesdue to imperfections of the suppressionsection profiles.
Water suppression was obtained bymeans of two CHESS pulses (24), oneprior to spatial localization and the secondduring TM. Sinc pulses of 75 Hz band-width without side lobes, filtered with aHanmng window, were used. The flip an-gle of the initial water suppression pulsewas larger than 90#{176}due to longitudinalrelaxation during the pulse sequence. Theduration of the dephasing gradient pulseswere 30 msec for the initial CHESS cycleand 10 msec for the CHESS cycle in theTM period.
Two-side-lobe sinc pulses of 1.6 msec
duration filtered with a Hanning windowwere used for the STEAM section selec-tion. The TE and TM dephasing gradientpulses had a duration of 4 and 6 msec, ne-spectively. Diffusion sensitizing gradientsof i8-msec duration were simultaneouslyapplied along the x, y, and z axes. Possibleresidual transverse magnetization afterdata acquisition was dephased with post-acquisition gradient pulses of 5-msecwidth. The pulse sequence was tuned forgradient slew rate effects by centering the
radio-frequency pulses with respect to thegradient envelope. No phase cycling wasused. All dephasing gradient pulses hada strength of 10 mT/m. Gradient rampswere 0.5 msec long.
The gradient switching scheme was op-timized to avoid unwanted gradient-ne-called echoes by choosing different gradi-ent crusher directions (25) for the spatialsuppression and for the STEAM volume
selection as shown in Fig 3. To reduce pos-sible artifacts due to residual eddy cur-
rents, sinusoidal crusher gradient wave-forms and additional gradient delays wereemployed before the STEAM section-selec-
tion pulses, diffusion gradients, and dataacquisition.
Spectroscopic measurements were per-
formed with 3,000/60 and a TM of 200msec. Spectra were acquired from a 3 x3 x 2-cm3 voxel in the right hemisphereabove the ventricles, containing mostlywhite matter. Peripheral gating was usedto compensate for cardiac-related brainand cerebrospmnal fluid pulsations. A timedelay of 300 msec between the systolicwave and the beginning of the pulse se-quence was found to minimize motion-induced signal amplitude variations (see
Results). Manual shimming on the se-lected voxel was performed with lineargradient shims by maximizing the absorp-tion peak height of the water resonance.We typically obtained absorption linewidths of 4-5 Hz (full-width at half-maxi-mum). Water suppression (approximately200-fold) was adjusted to obtain a residualwater signal that dominated the spectrumand served as a tracer signal for signalpostprocessing. The residual water signal
was undistorted except for a small phaseshift with respect to the metabolite reso-nances that did not vary substantially be-tween acquisitions (see Results).
Spectra were acquired randomly at 11different diffusion gradient values rangingfrom 0 to 10 mT/m with 1 mT/m incre-ments. The gradient factor b determinedby means of numeric integration (26)
ranged from 59 to 2,200 sec/mm2. Alt fieldgradients, G(t”), applied during the pulsesequence were taken into account. Sectionselection and TE dephasing gradient
pulses contributed greatly to the b factor,
both through cross terms with the diffu-sion gradients and through quadraticterms. For each diffusion gradient value,
64 acquisitions were individually recordedinto a two-dimensional data set. The spec-tra! width was 1 kHz, with a digital nesolu-
Figure 7. Signal intensities from Cho, Cr-
PCn, and NAA versus gradient factor b fordata from Figure 6. Dashed curves indicatecomputed exponential fits to Equation (1).
tion of I Hz. Total experiment time was 70minutes, including imaging, shimming,and water suppression.
Data processing of the two-dimensionaldata sets was performed with a Sun work-station and SAGE spectral analysis software(GE Medical Systems). It consisted of ex-ponential apodization with 2 Hz, zero fill-ing to 2,048 complex points, Fourier trans-formation, and automatic zero-orderphase correction for each individual signaltrace prior to spectral summation. Sincethe phase of the residual water signal wasslightly different from that of the metabo-tite signals (see Results), summed spectrawere manually zero-order phase connectedto optimize the absorption mode signal of
the metabolites. After correction for possi-ble residual baseline offsets, peak heightsof Cho, Cn-PCr, and NAA were obtainedby means of automatic peak picking and
fitted to single exponentials by usingEquation (1).

Volume 188 #{149}Number 3 Radiology e 723
RESULTS
Method Validation
Volume selection properties of thepulse sequence were investigated inphantoms by imaging the selectedvoxet with and without outer-volumesuppression. The voxel definition wasfound to be substantially improvedwith outer-volume suppression. Re-siduat outside signals were reducedby more than 100-fold, while the sig-
nat from the selected volume was re-duced by less than 5%.
In phantoms, spectral distortionwas insignificant and spectral line
widths increased by less than 1 Hzwhen maximal-diffusion gradientswere applied. Instrument stabilitywas assessed by observing the non-suppressed water resonance withmaximal-diffusion gradients applied.
Over periods of 10 minutes the ampli-tude was stable to within 3% and thephase was stable to within 5#{176}.
Compensation of linear motion ef-fects was verified with an agarose gelphantom that moved along the mag-net axis under software control. Themotion was oscillatory, with adjust-able amplitudes and maximal vetoci-ties reaching up to 100 mm/sec. The
voxet selection and experimental pa-rameters were the same as for the invivo studies performed with maximal-diffusion gradients. Figure 4a showsspectra of the unsuppressed waterresonance obtained with conven-tional averaging of 64 acquisitions.Strong signal tosses are evident, evenat table speeds as low as 5 mm/sec.With phase correction of individualacquisitions the signal is recovered fortable velocities of up to 100 mm/sec(Fig 4b).
Diffusion measurements of phan-toms with 10 mmol/L solutions ofCho and NAA were conducted tocompare the results with in vivo re-sults and to investigate the perfor-mance of the localization method (27).The in vitro metabolite diffusion coef-ficients at 37#{176}Cwere 1.3 x i0� mm2/
sec ± 0.03 x i0� (mean ± standarddeviation) for Cho and 0.9 x i0� mm2/
sec ± 0.03 x i0� for NAA. Water dif-
fusion at 21#{176}Cwas determined to be1.95 x io-� mm2/sec ± 0.05 x iO�,
which is in reasonable agreement with
literature values between 2.0 x i0�mm2/sec (3) and 2.09 x iO� mm2/sec(interpolated from reference 28).
Motion-induced signal errors invivo were investigated with the non-suppressed water signal acquired un-der the same conditions as for themetabolite measurements by usingmaximal diffusion gradients. Both
with and without peripheral gating,the signal phase was extremely unsta-ble and almost random. Without pe-nipheral gating, the signal amplitudein individual acquisitions exhibitedstrong variations, sometimes with at-most complete signal losses. With pe-
ripheral gating these signal amplitudevariations were strongly reduced. Byvarying the time delay between thesystolic wave and the pulse sequence
trigger from 0 to 600 msec, minimalamplitude fluctuations (usually lessthan 20%) were found at a delay timeof approximately 300 msec. Withoutperipheral gating or phase correction,
. diffusion coefficients were grosslyoverestimated.
The phase relationship between thewater and metabotite signals was in-vestigated by selecting a larger (38 x68 x 20 mm) volume, mostly in white
matter, to detect metabotite signalswith adequate signal-to-noise ratios inindividual acquisitions. The averagephase difference between water andNAA, as determined with maximal
diffusion gradients, was 25#{176}± 24#{176}.The residual phase fluctuations (stan-dard deviation, 24#{176})corresponded tothe noise level.
In Vivo Diffusion Coefficients
Spectroscopic volume localizationwas the same for all volunteers (Fig
5). As an example, Figure 6 shows a
typical series of spectra acquired withii different diffusion gradient ampli-tudes. Signals from NAA (2.01 ppm,
2.52 ppm), glutamine-glutamate (2.35
ppm, 3.8 ppm), Cr-PCr (3.02 ppm, 3.93ppm), Cho (2.24 ppm), and inositols(3.54 ppm) are detectable. Spectraloverlap from adjacent 1-coupled reso-nances (eg, glutamine-glutamate) atthe position of NAA was minimal, as
seen in Figure 6, because of the reta-tively tong TE chosen. Owing to sig-nat-to-noise constraints, only singletresonances from Cho, Cr-PCr, and
NAA were quantitated (Fig 7). Bestcurve-fitting results were usually ob-tamed for NAA, white data scatter forCr-PCr and Cho was slightly larger.Metabolite diffusion coefficients ob-tamed in 10 healthy volunteers aresummarized in the Table. For mostsubjects, with the exception of subject
7, the apparent diffusion coefficient ofNAA was slightly larger than that of
Cr-PCr or Cho. The cause for the de-viation in subject 7 is not known andmay be related to motion. Reproduc-ibility was verified in two volunteers(subjects 9 and 10), giving consistent
results within experimental errors.
The apparent diffusion coefficient
of the unsuppressed water resonance(0.65 x i0-� mm2/sec ± 0.02 x i0�)
obtained in separate experiments wassimilar to previously published results
obtained with diffusion-sensitive im-aging (3).
DISCUSSION
Localized diffusion spectroscopy invivo in the human brain appears to befeasible with adequate spectroscopic
localization and motion compensa-tion by means of phase correction of
individual acquisitions. While previ-ous measurements in vivo required
complete immobilization to avoid mo-
tion-induced signal losses, we wereable to investigate conscious human
subjects.
The spectral quality obtained in
human brain tissue, even with thestrongest diffusion gradients applied,was similar to that of conventional
non-diffusion-weighted spectroscopy,
indicating that eddy current-inducedtine broadening and spectral distor-tions were negligible despite the use
of long gradient pulses. The repro-ducibility of metabolite measurements
was largely dependent on their sig-nat-to-noise ratio, which favors NAA
over Cho and Cr-PCr. In addition,Cho and Cr-PCr, which are closer tothe water resonance, are more sensi-tive to small changes in water sup-pression with head motion and cardi-
ac-related brain pulsations, whichmight introduce baseline offsets.
At inspection of individual datatraces it became evident that signal
errors consisted mostly of phase en-rons rather than amplitude losses, in-
dicating that linear motion patterns(possibly from movements of the
whole head) dominated in measure-ments obtained with peripheral gat-ing. Signal errors due to linear mo-tion, along with small zero-orderphase shifts due to residual eddy cur-rents, could be completely removedby means of data postprocessing with
zero-order phase correction and didnot introduce spectral distortions. Asthe degree of amplitude errors variedslightly from subject to subject, de-
pending on factors such as breathingand swallowing, it was not possible tocompletely rule out the influence ofnonlinear motion on the measure-ments. This may be partly responsiblefor inter- and intraindividual mea-surement variations. Our own experi-ence with diffusion spectroscopy inanesthetized and paralyzed monkeys(Jordon EK, Posse 5, Cuenod CA, Le
Bihan D; unpublished data, 1992),which yields only slightly smaller me-

724 � Radiology September 1993
tabolite diffusion coefficients, mdi-
cates that such errors in human
measurements are probably less than
20%.
Metabolite diffusion coefficients
measured in 10 subjects were approxi-mately an order of magnitude smaller
than those previously reported in hu-
mans (11) and more consistent with
measurements in anesthetized and
paralyzed animals (9,10). For example,
Moonen and co-workers reported
NAA diffusion coefficients in cat brain
tissue that at a diffusion time of 200
msec ranged from 0.09 . iO� to 0.23�
iO� mm2/sec, depending on the gra-
dient direction relative to the fiber
orientation (interpolated from refer-
ence 9). Even at diffusion times as
short as 50 msec they found anisot-
ropy indicating bulk effects. By con-
trast, Merbotdt and co-workers ne-
cently reported isotropic metabotite
diffusion in rat brain tissue at a diffu-
sion time of 25 msec (10). They mea-
suned a slightly larger diffusion coeffi-
cient for Cho (0.3 . i0� mm2/sec)
than for NAA (0.25 . i0� mm2/sec).
In humans, apparent metabotitediffusion coefficients were consider-
ably smaller than that of water; this
finding may be attributable to differ-ences in molecular weight, to hydra-
tion layers around the metabolites,
and to hindrance by macnomolecutancompounds. Apparent metabolite dif-
fusion coefficients measured in vivo
were considerably smatter than those
measured in vitro, indicating domi-
nating influences of motion restriction
by membranes and hindrance due to
macnomoleculan compounds at the
long diffusion time used in this study.
This finding may also explain why the
diffusion coefficients of major metabo-
tites are similar in vivo, in contrast to
our in vitro measurements. At shorter
diffusion times for which motion ne-
striction is expected to be less severe,
more pronounced differences in me-
tabolite diffusion coefficients may be
observable, similar to those in rat
brain tissue reported by Merboldt and
co-workers (10).
When interpreting in vivo results, it
becomes evident that motion restric-
tion plays a major role, especially
with longer diffusion times, at whichapparent diffusion coefficients be-
come anisotopic. In the human study
presented here, in which diffusional
anisotnopy is expected to be similar to
that in cat brain tissue (9), the position
of the volume of interest and the ori-
entation of diffusion gradients mayhave influenced the measurement ofdiffusion coefficients. We are cur-
nently investigating metabotite diffu-
sion in relation to fiber orientation
and voxel position, for NAA in partic-ulan, to assess diffusional anisotropy.Experiments with different diffusiontimes are under development to eval-uate restriction effects. White it is fea-
sible to reach short diffusion times in
small-bone imagers with strong gradi-
ents, measurements with whole-body
imagers are limited by the available
gradient strengths. Local high-fluxgradient coils may help to reach
shorter diffusion times in human ex-
periments.Tissue-related magnetic field gradi-
ents, which were not taken into ac-
count during calculation of the gradi-ent factor b introduce the possibility
of overestimation of diffusion coeffi-cients. While macroscopic field gradi-
ents could be compensated by means
of localized shimming, the contribu-
tion of subvoxel field gradients is un-
known and remains to be addressed(29). Proton spectroscopy in the hu-
man brain at field strengths of 1-4 Tindicates that line widths in brain tis-
sue are dominated by microscopic
susceptibility effects rather than by
transverse relaxation (30,31). Local
differences in susceptibility due to
deoxyhemoglobin in small blood yes-sets, for example, can give rise to sub-
stantial local gradients. Our results at
1.5 T, however, are largely consistentwith measurements by Moonen and
co-workers at 4.7 T (9), with which
susceptibility effects are expected to
be much stronger. This indicates thatlocal gradients, on average, are small
compared with the applied diffusion
gradients.
At short echo times, a number ofrelatively weak resonances with com-
plex J-coupting or short transverserelaxation times are detectable (23,32)and may exhibit interesting diffusion
properties. To assess these resonances
at short echo times, it is necessary touse stronger local high-flux gradient
coils and more sensitive nadio-fre-
quency coils. In cortical regions, for
example, surface coils will stronglyimprove the signal-to-noise ratio.
With more complex spectral patterns,however, spectral overlap becomes aproblem and reliable data analysismethods must be developed to distin-guish small differences in overlapping
resonances.
As reported above, signal ampli-
tude variations in individual data
traces due to nontranslational motionmay lead to an overestimation of dif-fusion coefficients. To improve therobustness of the method, we are in-
vestigating statistical methods to ne-
ject individual data traces that exhibitexcessive signal tosses due to nonlin-
ear motion. We are also investigatingthe feasibility of diffusion spectro-
scopic imaging to simultaneously
measure diffusion in multiple voxets.
As expected, ghosting artifacts dueto the strong motion sensitization
proved to be a major problem. As afirst step, we developed a robust mo-
tion artifact compensation method for
diffusion-sensitive spectroscopic im-
aging (33).
The method presented in this anti-cle may be valuable for the investiga-
tion of metabolite mobility in normalhuman brain tissue similar to mea-surements in animal brain. By using
NAA as a neunonal marker, it may be
possible to investigate cell- and me-
tabolite-specific mechanisms of mo-tion restriction. Applications to dis-ease processes such as white matter
diseases, brain atrophy, and acute
stroke will be of particular interest.
Water diffusion in acute stroke, forexample, which is reduced in isch-

Volume 188 #{149}Number 3 Radiology � 725
emic lesions, is not yet fully under-stood. Intracellular and tissue-specificmarkers like Cho, Cr-PCr, NAA, andinositol may provide further informa-tion about physiologic changes inischemic tissue and the underlyingmechanisms of altered water diffu-
sion. Finally, diffusion represents anew degree of freedom to character-ize in vivo proton spectra that at ctini-cal field strengths suffer from denselypacked spectral information. If, forexample, several compounds withdifferent diffusion properties contrib-
ute to a single resonance tine, diffu-sion spectroscopy may help to distin-guish them. It is possible, for instance,to suppress rapidly diffusing com-pounds such as water and certain me-tabotites while retaining slowly dif-fusing compounds such as lipids andproteins (34). Diffusion spectroscopymay thus provide new opportunitiesfor tissue characterization and func-tionat studies. #{149}
Acknowledgments: We are indebted to Pierre
Germain, BS, for his help with implementingsoftware routines for automated data process-ing. We gratefully acknowledgeJoseph A.Frank, MD, for supporting this work
References1_ Le Bihan D, Breton E, Lallemand D, Grenier P,
Canabis PA, Lavel-Jeantet M. MR imaging ofintravoxel incoherent motion: applications todiffusion and perfusion in neurofogic disor-ders. Radiology 1986; 161:401-407.
2. Le Bihan D, ifreton E, Lallemand D, Aubin M,VignaudJ, Lavel-Jeantet M. Separation ofdiffusion and perfusion in intravoxel incoher-ent motion (IVIM) MR imaging. Radiology1988; 168:497-505.
3. Le Bihan D. Molecular diffusion nuclearmagnetic resonance imaging. Magn Reson Q1990; 7:1-30.
4. Moseley ME, Cohen Y, Mintorovitch J, et al.Diffusion-weighted MR imaging of water dif-fusion in cat central nervous system. Radiol-ogy 1990; 176:439-446.
5. Rutherford MA, Cowan FM, Manzur AY, et al.MR imaging of anisotropically restricted diffu-sion in the brain of neonates and infants.Comput Assist Tomogr 1991; 15:188-198.
6. Moseley M, Kucharczyk J, Kurhanewicz J,Norman D. Diffusion-weighted MR imagingof acute stroke: correlation with T2-weightedand magnetic susceptibility-enhanced MR rn-aging in cats. AJNR 1990; 11:423-429.
7_ Warach S, Chien D, Li W, Ronthal M, EdelmanRR. Fast magnetic resonance diffusion-weighted imaging of acute human stroke.Neurology 1992; 42:1717-1723.
8. Moonen CTW, van ZijI PCM, Le Bihan D,DesPres D. In vivo NMR diffusion spectros-copy: 31P applications to phosphorus metabo-lites in muscle. Magn Reson Med 1990; 13:467-477-
9. Moonen C�IW, van Gelderen P. van ZijI PCM,DesPres D, Olson A. The translational rnobil-ity of N-acetylaspartate in cat brain as mea-sured by localized proton NMR spectroscopy(abstr). In: Book of abstracts: Society of Mag-netic Resonance in Medicine 1991. Berkeley,Calif: Society of Magnetic Resonance in Medi-cine, 1991; 141.
10. Merboldt KD, HOstermann D, H#{228}nickeW,Bruhn H, Frahm J. Molecular self-diffusion ofintracellular metabolites in rat brain in vivoinvestigated by localized proton NMR diffu-sion spectroscopy. Magn Reson Med 1993; 29:125-129.
11. Moonen CTW, von Kienlin M, van ZijI PCM, etal. Comparison of single shot localizationmethods (STEAM and PRESS) for in vivo pro-ton NMR spectroscopy. NMR Biorned 1989;2:201-208.
12. Gyngell ML, Frahm J, Merboldt KD, H#{228}nickeW, Bruhn H. Motion rephasing in gradientlocalized spectroscopy. J Magn Reson 1988;77:596-598.
13. Klammler F, Kimmich R. Volume selectiveand spectroscopically resolved NMR investiga-tion of diffusion and relaxation in fertilizedhen eggs. Phys Med Biol 1990; 35:67-79.
14. Posse S�, Cuenod CA,Jordon EK, Le Bihan D.In vivo 1H diffusion spectroscopy in humanbrain (abstr). In: Book of abstracts: Society ofMagnetic Resonance in Medicine 1992. Berke-ley, Calif: Society of Magnetic Resonance inMedicine, 1992; 2141.
15. Stejskal EO, TannerJE. Spin diffusion mea-surements: spin echoes in the presence oftime-dependent field gradients. J Chem Phys1965; 42:288-292.
16. Chenevert T, Brunberg J, Pipe J. Anisotropicdiffusion within human white matter: demon-stration with NMR techniques in vivo. Radiol-ogy 1991; 177:401-405. .
17. Feinberg DA, Mark AS. Human brain motionand cerebrospinal fluid circulation demon-strated with MR velocity imaging. Radiology1987; 163:793-799.
18. Poncelet BP, Weeden VJ, Weisskopf RM, Co-hen Ms. Brain parenchyma motion: mea-surement with cine echo-planar MR imaging.Radiology 1992; 185:645-651.
19. Enzmann DR. PeIc NJ. Brain motion: mea-surement with phase-contrast MR imaging.Radiology 1992; 185:653-660.
20. Granot J. Selected volume excitation usingstimulated echoes (VEST): applications to spa-tially localized spectroscopy and imaging.Magn Reson 1986; 70:488-492.
21. Kimmich R, Hoepfel D. Volume selectivemultipulse spin echo spectroscopy. J MagnReson 1987; 72:379-384.
22. Frahm J, Merboldt KD, Haenicke W. Local-ized proton spectroscopy using stimulatedechoes. J Magn Reson 1987; 72:502-508.
23. Posse S, Schuknecht B, Smith ME, van ZijIPCM, Herschkowitz N, Moonen CTW. Shortecho time proton spectroscopic imaging.Comput Assist Tomogr 1993; 17:1-14.
24. Haase A, Frahm J, Haenicke W, Matthaei D.IH NMR chemical shift selective (CHESS) im-aging. Phys Med Biol 1985; 30:341-344.
25. Moonen CTW, van ZijI PCM. Highly effec-tive water suppression for in vivo protonNMR spectroscopy (DRYSTEAM). J MagnReson 1990; 88:28-41.
26. Le Bihan D, Cuenod CA, Posse S. Determina-tion of the gradient factor b in localized diffu-sion IH spectroscopy (abstr). In: Book of ab-stracts: Society of Magnetic Resonance inMedicine 1992. Berkeley, Calif: Society of Mag-netic Resonance in Medicine, 1992; 1218.
27. Cuenod CA, Posse 5, Le Bihan D. Relation-ship between diffusion and temperature forN-acetyl aspartate and choline: a localized IHNMR spectroscopy study (abstr). In: Book ofabstracts: Society of Ma�netic Resonance inMedicine. Berkeley, Calif: Society of MagneticResonance in Medicine, 1992; 1215.
28. Mills R. Self diffusion in normal and heavywater in the range 1-45#{176}.J Chem Phys 1973;77:685-688.
29. Zhong J, Kennan RI’, Gore JC. Effects of sus-ceptibility variations on NMR measurementsof diffusion. J Magn Reson 1991; 95:267-280.
30. Sauter R, LoefflerW, Bruhn H, Frahm J. Thehuman brain: localized H-I MR spectroscopyat 1.0 T. Radiology 1990; 176:221-224.
31. Barfuss H, Fischer H, Hentschel D, et al. Invivo magnetic resonance imaging and spec-troscopy of humans with a 4 Tesla whole-bodymagnet. NMR Biomed 1990; 3:31-45.
32. Frahm J, Bruhn H, Gyngell ML, Merboldt KD,H#{228}nickeW, Sauter R. Localized proton NMRspectroscopy in different regions of the humanbrain in vivo: relaxation times and concentra-tions of cerebral metabolites. Magn Reson Med1989; 11:47-63.
33. Posse 5, Cuenod CA, Le Bihan D. Motionartifact compensation in 1H spectroscopic im-aging by signal tracking. J Magn Reson (inpress).
34. Van ZijI PCM, Moonen CTW, Faustino P, Pe-karJ, Kaplan Ofer, Cohan J. Complete sepa-ration ofintracellular and extracellular infor-mation in NMR spectra of perfused cells bydiffusion-weighted spectroscopy. Proc NatIAcad Sri USA 1991; 88:3228-3232.