human cd4+ t cells stimulated by conserved adenovirus 5 hexon peptides recognize cells infected with...
TRANSCRIPT

Human CD4+ T cells stimulated by conserved adenovirus5 hexon peptides recognize cells infected with differentspecies of human adenovirus
Louise A. Veltrop-Duits1, Bianca Heemskerk1, Claudia C. Sombroek1,Tamara van Vreeswijk1, Sophie Gubbels1, Ren� E. M. Toes2,Cornelis J. M. Melief 3, Kees L. M. C. Franken3, Menzo Havenga4,Maarten J. D. van Tol1 and Marco W. Schilham1
1 Department of Pediatrics, Leiden University Medical Center, Leiden, The Netherlands2 Department of Rheumatology, Leiden University Medical Center, Leiden, TheNetherlands
3 Department of Immunohematology and Bloodtransfusion, Leiden University MedicalCenter, Leiden, The Netherlands
4 Crucell Holland BV, Leiden, The Netherlands
The immune response against human adenovirus (HAdV) has gained interest because ofthe application of HAdV-based vectors in gene therapy and the high incidence ofinfections in pediatric recipients of allogeneic stem cell grafts. Because antiviralmedication is frequently ineffective, the option of adoptive transfer of HAdV-specificdonor-derived T cells in these immunocompromised patients is investigated. Togenerate good manufacturing practice-compatible reagents, a panel of 63 long,overlapping, peptides of the hexon protein was screened for recognition by T cells. Fiveconserved peptides of 30 amino acids were identified that were recognized by themajority of adult donors. CD4+ T cells from long-term cultures of PBMC, stimulatedwith this set of five peptides, recognized cells infected with HAdV serotypes belonging todifferent species. These data demonstrate that adult human T cells preferentiallyrecognize conserved sequences of amino acid residues from a structural protein ofHAdV. In the context of gene therapy, this observation may limit the beneficial effect ofswitching to HAdV-based vectors derived from less common serotypes of HAdV in anattempt to circumvent pre-existing immunity. However, this cross-reactivity benefits theapplication of HAdV-specific T cells for adoptive immunotherapy in immunocompro-mised transplant recipients.
Introduction
Human adenovirus (HAdV) infections in the immuno-competent host are usually mild and self-limiting. Forthis reason, and because the virus is able to infect non-cycling cells, HAdV-based vectors are applied for genetherapy in humans. However, the presence of animmune response against HAdV in immunocompetentindividuals may limit the effectiveness of gene deliveryand/or gene expression. In severely immunocompro-mised patients, such as pediatric recipients of allogeneic
Immunity to infection
Correspondence: Dr. Marco W. Schilham, Department ofPediatrics, P3-P, Leiden University Medical Center, P.O. Box9600, 2300 RC Leiden, The NetherlandsFax: +31-71-526-6876e-mail: [email protected]
Received 15/12/05Revised 16/6/06
Accepted 20/7/06
[DOI 10.1002/eji.200535786]
Key words:Human � Peptides
� Stem cells � T cells� Viral
Abbreviations: AB: human AB serum � GMP: good manufac-turing practice � HAdV: human adenovirus � MB: methyleneblue � PCP: peptide culture pool � SFU: spot-forming units
Louise A. Veltrop-Duits et al. Eur. J. Immunol. 2006. 36: 2410–24232410
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

stem cell transplants (SCT), HAdV infection may causelife-threatening complications [1–6]. To date, 51serotypes of HAdV have been described, which areclassified into 6 species (A to F). Species A, B and C areresponsible for most of the infections in the pediatricSCT recipients [7–9]. In recent years, the incidence ofHAdV infection increased in these recipients [9–12]. In aprospective study performed at the pediatric stem celltransplantation unit in Leiden, HLA-mismatched trans-plants, immunosuppression with anti-thymocyte globu-lin (ATG) and/or Campath and delayed immunereconstitution were defined as risk factors for HAdVinfection [9, 12]. Together, these risk factors indicate arole for the adaptive immune system in viral clearance.Pharmacological options for treatment of HAdV infec-tions are limited. Ribavirin and cidofovir have been usedin immunocompromised patients, but these two drugshave not unequivocally been shown to be efficacious [8,13–17]. Therefore, immunotherapy by infusion ofHAdV-specific donor-derived T cells might be an optionfor treatment. Case reports have already suggested thatinfusion of donor T lymphocytes can contribute to theclearance of HAdV infection [18, 19]. Since infusion ofunselected donor lymphocytes bears the risk of inducinggraft-versus-host disease (GVHD) [18], it would bepreferable to infuse only HAdV-specific donor T cells.
Investigations of the immune response towardsHAdV have revealed that the major part of the cellularresponse in vitro consists of CD4+ T cells [20–24].Although CD8+ T cells specific for HAdV have beenreported [25, 26], we observed that the HAdV-specificT cells in HAdV-infected immunocompromised recipi-ents, who spontaneously cleared the infection, werelargely CD4+ [9]. Here, we report that even by using DCas antigen-presenting cells and viruses that lack thegenes E1 and E3, which regions contain sequencesencoding proteins known to down-regulate HLA class Imolecules, as the source of antigen, the majority of theHAdV-specific T cell response still appears to bemediated by CD4+ T cells. This leads to the conclusionthat, although HAdV-specific CD8+ T cells can bedetected, HAdV-specific CD4+ Tcells are more generallypresent in the human population.
For in vitro stimulation and expansion of T cells to beused in adoptive transfer protocols, a source of antigenthat is simpler and cheaper to produce under goodmanufacturing practice (GMP) conditions than purifiedHAdV is required. Relatively long (30-mer) peptides,preferentially containing multiple epitopes restricted byvarious HLA genotypes that can be processed physio-logically, would fulfill these criteria. To perform anunbiased study of major Tcell determinants in HAdV, wefirst investigated the recognition of several HAdV5proteins, i.e. the structural penton base and hexonproteins and the early protein E1A. Since the hexon
protein was recognizedmost frequently, this protein wassubsequently synthesized as a set of overlappingpeptides of 30 amino acid residues. Screening of PBMCfrom a panel of healthy donors revealed that a relativelylimited set of long peptides was able to evoke responsesin almost all adult donors. Without further preciselydefining HLA class II epitopes, this limited set of 30-amino acid peptides was used for culturing HAdV-specific T cells. The CD4+ T cells specific for thesepeptides recognized target cells pulsed with HAdV5 andother serotypes, leading to the conclusion that peptidesmay be appropriate for the generation of HAdV-specific,donor-derived, CD4+ T cells for infusion in patients atrisk for disseminated HAdV infection [27].
Results
Majority of T cells recognizing HAdV are CD4+
To investigate whether CD8+ and/or CD4+ T cellsrespond to HAdV-infected cells, PBMC from ten healthyadult donors were stimulated with infectious HAdV5.After 5 days of culture, CD4+ Tcells of all ten individualsand CD8+ T cells of seven of ten donors respondedaccording to our criteria of positivity. In the respondingT cell populations of those seven donors, an average of85% of the IFN-c-producing T cells were CD4+ and only15% were CD8+ (Table 1). It could be argued thatpeptides derived from the structural proteins of inputviral particles would preferentially be presented onMHCclass II molecules, thus biasing the response towardsCD4+ T cells. It has been described that viral early geneproducts encoded in the E1 and E3 region can down-regulate the expression of MHC class I molecules [28].Therefore, PBL from donors who lacked an HAdV-specific CD8+ T cell response when viral antigen waspresented by PBMC, were stimulated with DC infectedwith E1-E3-deleted HAdV5 in order to favor thepresentation of antigen on HLA class I molecules.Twelve days later, the responding cells were restimu-lated with autologous PBMC infected with wild-type orE1-E3-deleted HAdV5 in order to determine the IFN-c-producing T cells. Although significant numbers ofIFN-c+CD4+ T cells were observed, very few CD8+
T cells secreting IFN-c were detected after intracellularstaining (Fig. 1). These results suggest that the observedpredominance of CD4+ T cells recognizing antigenicdeterminants of structural proteins of HAdV5 is not dueto the culture conditions or antigenic stimulation, butreflects the true nature of the T cell mediated cellularresponse to this particular virus.
Eur. J. Immunol. 2006. 36: 2410–2423 Immunity to infection 2411
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Selecting HAdV5 peptides
To define which structural protein of HAdV contains themajority of antigenic determinants recognized by Tcells,we investigated the recognition of recombinant hexonprotein (constructed in four overlapping parts, i.e.
IIA1–273, IIB245–509, IIC479–743, IID720–952), penton base(III) and E1A by PBMC from healthy donors. Arepresentative reaction profile in IFN-c ELISPOT assay,measuring IFN-c spot-forming units (SFU)/105 PBMC, isshown in Fig. 2A, where PBMC of donor K responded torecombinant hexon part IIC479–743, IID720–952 and III
Table 1. IFN-c-producing CD4+ or CD8+ T cell population after 5 days of culturea)
No stimulus HAdV stimulated
Donor CD4+ CD8+ CD4+ CD8+
A 0.0 0.0 0.4 R 0.1 R
B 0.2 0.1 2.2 R 0.4 R
C 0.0 0.0 0.4 R 0.2 R
D 0.1 0.1 0.6 R 0.1 NR
E 0.0 0.0 1.3 R 0.1 R
F 0.1 0.1 1.9 R 0.4 R
G 0.2 0.1 0.4 R 0.1 NR
H 0.0 0.0 1.6 R 0.3 R
I 0.1 0.0 1.4 R 0.2 R
J 0.1 0.1 0.8 R 0.1 NR
Average 0.08 0.05 1.1 0.2
Range 0–0.2 0–0.1 0.4–2.2 0.1–0.4
a) PBMCwere infectedwithwild-type HAdV5 (MOI 200) and cultured for 5 days after which the IFN-c-producing CD4+ or CD8+ cellswere determined by flow cytometry as percentage of the CD3+ T cells. R: responding, NR: not responding..
Figure 1. Culture with E1-E3- deleted HAdV5 induces mainly CD4+ T cells. PBL were cultured with dendritic cells infected withE1-E3-deleted HAdV5 and tested at day 12 for recognition of autologous PBMC infected with wild-type HAdV5 or E1-E3- deletedHAdV5 (MOI 2000) by determining IFN-c-producing T cells after intracellular staining. Medium alone, referred to as no stimulus,was used as negative control. Cells shown are CD3+ gated lymphocytes. The% sign indicates the percentage of IFN-c+ T cells of theCD3+CD4+ or CD3+CD8+ lymphocytes. Results are from one donor and representative for the results obtained from two differentdonors.
Louise A. Veltrop-Duits et al. Eur. J. Immunol. 2006. 36: 2410–24232412
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

(Fig. 2A). The responses to IIA1–273, IIB245–509 and E1Awere below 25 spots/105 PBMC and were considerednegative.
In total, PBMC from 33 healthy donors, recognizingHAdV5, were tested. The C-terminal part of the hexonprotein, which includes recombinant proteins IIC479–743
(82%) and IID720–952 (82%), was recognized by moredonors than the N-terminal part of hexon protein,IIA1–273 (27%), IIB245–509 (55%), or the penton baseprotein (67%) (Fig. 2B). The recognition of the earlygene product E1A was low (20%). These data indicatethat the conserved part of hexon contains widelyrecognized T cell epitopes and it was decided to focuson recognition of the hexon protein.
To further define which regions of the hexon proteinwere recognized most frequently, overlapping peptidesof 30 amino acids with an overlap of 15 amino acidscovering the complete protein (peptide II4 to II66) weresynthesized. First, the responses of PBMC from healthyadult donors to small pools of these peptides were testedby measuring IFN-c secretion in culture supernatant byELISA. For example, PBMC of donor L contained T cells
that recognized peptide pools P7, P9 and P11 (Fig. 3A).Subsequently, PBMC of this donor were cultured withthe single peptides of each of the positive peptide pools,i.e. peptides II37-II41 of peptide pool P7, peptide II47-II51 of peptide pool P9 and peptides II57-II61 of peptidepool P11, and the response was measured by ELISPOTassay. Responses against HAdV5 or medium alone wereused as positive and negative control (the average of allnegative controls was 6.7 spots/105 PBMC), respec-tively. Positive responses for donor L were observedagainst peptides II37, II41, II51, II57, II60 and II61(Fig. 3B).
Twenty-one donors were tested accordingly. Mostresponses were observed against peptides from peptidepools at the C-terminal part of the hexon protein, inagreement with the results obtained using the recombi-nant hexon parts (Fig. 4A). Peptides from pools P8 (13donors), P9 (11 donors) and P11 (10 donors) wererecognized most frequently. At the level of singlepeptides peptide II42 (11 out of the 13 peptide poolP8 reactive donors), peptide II61, peptide II50 andpeptide II57 were the most frequently recognized
Figure 2. Recognition of HAdV5 proteins. (A) PBMC of healthy donor Kwere cultured for 4 dayswithout stimulus (-) or with HAdV5or recombinant hexon parts (IIA-IID), penton base protein (III) and E1A, after which IFN-c SFU/105 PBMC were measured. Dottedline represents threshold (25 spots/105 PBMC) above which the response is considered positive. Data are representative for theresults obtained from 33 adult donors. (B) Overview of healthy donors tested for their recognition of the recombinant hexonproteins (IIA-IID), penton base protein (III) and E1A. White bar represents the number of donors tested, black bar the number ofdonors responding to the recombinant protein.
Eur. J. Immunol. 2006. 36: 2410–2423 Immunity to infection 2413
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

peptides (Fig. 4B). Peptide II64 of peptide pool P12,containing epitope II910–924 [29] was recognized by fiveof the six donors reactive to this peptide pool. Thisscreening resulted in identification of five peptides ofthe hexon protein, which are frequently recognized byPBMC of healthy adult donors.
To culture PBMC with synthetic HAdV peptidescompatible with GMP requirements for adoptive im-munotherapy, the five most frequently recognized longpeptides [peptides II42, 50, 57 61 and 64 (Fig. 4B)] wereselected and combined to constitute a HAdV peptideculture pool (PCP). To investigate whether the PCP wasable to induce responses in the majority of donors, 18different healthy blood bank donors, responding toHAdV5, were tested against PCP and the individualpeptides. Seventeen (94%) of these donors recognizedone or more of the five peptides as well as PCP (Table 2).
HAdV5 PCP induced IFN-c production almostexclusively in the CD4+ T cell population, as was alsoobserved after culture with infectious HAdV5 virus(Table 1) [23]. This was concluded from IFN-c ELISPOTexperiments in which depletion of CD4+ T cellsabrogated the response, whereas depletion of CD8+
T cells had no effect (data not shown).
Peptide-generated T cells recognize HAdV-pulsedcells
To investigate whether PCP-reactive T cells alsorecognize cells infected with HAdV, PBL of six healthydonors (donors 13–18 from Table 2) were cultured withHAdV5 PCP. After 12 and 26 days of culture the HAdV-specific T cells recognizing autologous PBMC loadedwith methylene blue (MB)-inactivated HAdV5 wereenumerated by intracellular IFN-c staining, whichincreased from 3.0% (range 0.4–3.0%) of the CD4+
T cells at day 12 to 8.6% (range 1.0–10.0%) at day 26(Fig. 5). In the same cell lines, the percentage of CD4+
T cells recognizing PCP-loaded PBMC increased from7.4% (range 1.2–7.4%) at day 12 to 21.9% (range1.6–52.7%) at day 26 (Fig. 5). The responses to PCPwere usually a bit higher than to MB-inactivated HAdV,and individual donors differed considerably in percen-tages of cells responding after 26 days of culture.Furthermore, it is important for adoptive immunother-apy that the cultured T cells also recognize PBMCinfected with infectious HAdV. Ten percent (range0.7–12.2%) of the HAdV5 PCP cultured CD4+ T cellsproduced IFN-c in response to infectious HAdV5
Figure 3.Recognition ofHAdV5hexonpeptides. PBMCof healthy donor Lwere cultured for 4 days after stimulationwith (A)HAdV5hexon peptide pools (P1-P12), after which the amount of IFN-c secreted in culture supernatant was determined or (B) the singlepeptides of the selected peptide pools, after which the IFN-c SFUper 1.0x105 PBMCwasmeasured. CultureswithHAdV5 orwithoutstimulus (-) served as positive and negative control, respectively. Dotted line represented threshold above which the response isconsidered positive. Data are from 1 donor and representative for the results obtained from 21 donors.
Louise A. Veltrop-Duits et al. Eur. J. Immunol. 2006. 36: 2410–24232414
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

stimulation at day 26, confirming that these T cells notonly recognized MB-inactivated virus but also infectiousHAdV (Fig. 5, Table 3).
For these six donors, the response of PBMC to thesingle HAdV hexon peptides after 4 days of stimulationwith the single peptides (IFN-c ELISPOT assay at day 5)was compared with the response after 26 days ofstimulation with PCP (intracellular IFN-c staining).The response profiles after long-term culture with PCPare similar to those after short time culture with thesingle peptides, although sometimes responses topeptides that were recognized after 5 days culture werenot recognized after 26 days and vice versa (Table 3).Important to note is that the responses to PCP after26 days of culture were not dominated by only one ofthe five peptides.
Besides IFN-c production, the PCP-cultured T celllines also showed an antigen-specific proliferation. Themean proliferation of the peptide-specific T cells fromthe six donors at day 26 to cells pulsed with MB-inactivated HAdV5 (mean [range], 27 000[1000–98 000] cpm) and to PCP (mean [range],45 000 [1500–120 000] cpm) was elevated comparedto cells pulsed with medium alone (mean [range], 4900[300–14 000] cpm).
To investigate whether Tcells stimulated with the fiveHAdV5-derived peptides also recognized other HAdVserotypes, PBL of four donors (donors 15–18) culturedwith HAdV5 PCP were tested at day 26 againstautologous PBMC pulsed with one of eight differentMB-inactivated HAdV serotypes from four differentspecies. The CD4+ Tcells from these cultures responded
Figure 4. Overview of recognition of HAdV5 hexon peptides. (A) PBMC of 21 adult donors were first tested for their recognition ofhexon protein peptide pools (P1-P12). (B) Donors with a positive response to one or more of the peptide pools were then tested fortheir recognition of the single peptides from the respective pools.White bars represent the number of donors tested, black bar thenumber of donors recognizing (A) the indicated peptide pool or (B) the indicated single peptide. Peptides selected to constitute the"peptide culture pool" (PCP) are indicated by an arrow.
Eur. J. Immunol. 2006. 36: 2410–2423 Immunity to infection 2415
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Table 2. Recognition of 5 selected HAdV5 hexon peptides by 18 donorsa)
DonorPeptides of HAdV
II42 II50 II57 II61 II64 PCP
1 + + + + + +
2 + - + + + +
3 + + - - - +
4 - - + + + +
5 - - - - - -
6 - - - + - +
7 - + - - + +
8 + + - - - +
9 + - - - - +
10 + - + + + +
11 + - + + + +
12 - - - + - +
13 + - + + + +
14 + + + + + +
15 + - + + + +
16 + - - - - +
17 - + - - + +
18 + - - - + +
Positive 12 6 8 10 11 17
a) IFN-c SFU/105 PBMC: +: �25 spots/105 PBMC and exceeding the mean of the negative control + 3 times SD.
Figure 5. T cells cultured with HAdV5 PCP recognize MB-inactivated and infectious HAdV5. PBL were cultured for 26 days with0.4 ng/mL PCP and tested at days 12 and 26 for recognition of autologous PBMC pulsedwithMB-inactivated HAdV5 (MOI 2000) andinfectious HAdV5 (MOI 2000) by IFN-c production of the CD3+ gated lymphocytes. The % sign indicates the percentage of IFN-c+
T cells of the CD3+CD4+ lymphocytes. Medium alone, referred to as no stimulus, and PBMC pulsed with HAdV5 PCP were used asnegative and positive control, respectively. Results are from one donor and representative for the results obtained from sixdifferent donors tested.
Louise A. Veltrop-Duits et al. Eur. J. Immunol. 2006. 36: 2410–24232416
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

to all eight MB-inactivated HAdV serotypes from speciesA, B, C and D in IFN-c production and proliferation(Table 4), indicating that these peptide-cultured T cellsare indeed broadly reactive against different HAdVspecies. Two donors (16 and 17) were also tested atday 12 and showed an increase in the number of IFN-c-producing CD4+CD3+ T cells from day 12 to day 26 inrecognition of the eight MB-inactivated HAdV serotypes(data not shown).
It may be argued that all PCP-cultured T cell linesrecognize HAdV pulsed target cells, because thepreviously described HLA-DP restricted epitope,II910–924, is contained in peptide II64 [29, 30]. Todemonstrate that the other four peptides of the PCP alsocontain epitopes capable of stimulating virus-specificT cells, PBL from two donors (donor 16 and 17) werecultured with the combination of only the four peptidesII42, II50, II57 and II61, but not II64. Importantly, thesecell lines not only recognized MB-inactivated HAdV5,but also the seven otherMB-inactivated HAdV serotypes.Responses were detected by production of IFN-c andantigen-specific proliferation and were in the samerange as the responses for the T cell lines cultured in thepresence of PCP containing II64 (Table 4).
These results indicate that novel peptides wereidentified that can be used to generate HAdV-specificT cells. T cells stimulated with these peptides recognizedHAdV of various serotypes and may form a tool foradoptive immunotherapy in the setting of transfer ofdonor-derived HAdV-specific T cells to the immunocom-promised recipient of allogeneic stem cell transplants.
Discussion
Immunity to HAdV has only recently been investigatedin detail, as HAdV infections in immunocompetentpersons do not constitute a major health problem. Anintriguing observation is that, although HAdV-specificCD8+ T cells can be detected, the majority of HAdV-specific T cells in the blood of healthy adults as well aspatients who have just cleared an infection, are CD4+ [9,20, 23, 24]. This observation could have been biased bythe culture conditions used by the various investigators.Therefore, in an effort to favor presentation of HAdVantigen on HLA class I, DC were used as antigen-presenting cells and E1-E3-deleted HAdV as antigen.Even under these in vitro conditions, the majorresponding T cell population was CD4+, as was alsoobserved by others [31]. The CD4+ T cells could indeedrepresent the main HAdV-specific effector T cells in vivoas well and limit viral replication by (i) cytokine (i.e.IFN-c) secretion, protecting neighboring cells frominfection, (ii) providing help to B cells for the productionof neutralizing antibodies, (iii) inducing maturation ofDC, and, thereby, supporting differentiation of CD8+
T cells (which we have not been able to detect in highnumbers) or (iv) lysis of infected target cells.
This is the first unbiased study comparing T cellresponses to HAdV hexon peptides in the healthy humanpopulation. Until now, only one epitope for CD4+ Tcellshas been reported, which is restricted by HLA-DP4 [29,30]. For CD8+ T cells, five epitopes have been identifiedthat are restricted by HLA-A1, -A2, -A24 and -B7 [25].For the two latter studies, peptides were selected asepitopes by a predictive algorithm. The five mostfrequently recognized peptides identified in our studywere selected empirically without the aim to determinethe exact HLA-binding epitopes. The amino acid
Table 3. Recognition of HAdV5 hexon peptides after 26 days of culture
Donora) IFN-c SFUb) % of IFN-c+ CD4+ T cells after 26 days of culture with PCPc)
II42 II50 II57 II61 II64 No sti-mulus
MBHAdV5
PCP II42 II50 II57 II61 II64 infectiousHAdV5
13 + - + + + 0.2 8.6 21.9 0.6 0.1 3.6 12.9 2.6 10.0
14 + + + + + 0.0 2.5 5.8 0.2 2.8 0.1 1.6 0.1 4.8
15 + - + + + 3.1 10.0 52.7 23.5 2.2 4.5 48.8 4.6 9.2
16 + - - - - 0.3 1.2 12.8 10.5 0.2 0.5 0.8 2.0 1.1
17 - + - - + 0.1 12.4 12.1 1.3 11.6 0.7 2.3 4.5 12.2
18 + - - - + 0.6 1.0 1.6 1.3 0.6 0.5 0.8 0.4 0.7
a) Donor 13: HLA-DR17,15; donor 14: HLA-DR15, 7; donor 15: HLA-DR4, 17; donor 16: HLA-DR13, 15; donor 17: HLA-DR7, 12; donor18: HLA-DR15, 6.
b) IFN-c SFU: IFN-c ELISPOTassay at day 5: SFU/105 PBMC; +:�25 spots/105 PBMC and exceeding themean of the negative control +3 times SD.
c) % of IFN-c+ CD4+ T cells: % of IFN-c+ cells of the CD3+CD4+ lymphocytes. Underlined percentages indicate a positive response atday 5 in the IFN-c ELISPOT assay.
Eur. J. Immunol. 2006. 36: 2410–2423 Immunity to infection 2417
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

sequences of these peptides were compared betweendifferent HAdV serotypes from various species.Peptide II42, 50, 61 and 64 show minimal differencesbetween species (Table 5). Peptide II64 contains thedescribed HLA-DP restricted epitope II910–924, which isconserved between many human and other mamma-lian adenoviruses [29]. Peptides II42, 57 and 61 havenot been described before as containing major T cellepitopes, but are recognized at a high frequency (57,50 and 64%, respectively). The fact that these peptidesare conserved explains why other serotypes are alsoincreasingly recognized by T cells stimulated withthese peptides. No obvious correlation betweenrecognition of peptides and expression of HLA-DRalleles was observed. Explanations might be that the30-mers contain multiple T cell epitopes that arepresented by multiple HLA class II alleles [32] or thatthe hexon peptides are presented by certain HLA-DPalleles that occur very frequently in the population, asdescribed for the II910–924 peptide presented by HLA-DP4 [30]. Defining of HLA class II-binding epitopeswas not the aim of the present study and wouldrequire a much larger cohort of donors and thegeneration of large numbers of Tcell clones. The set offive long peptides forms a source of antigen that can besynthetically produced and from which variousepitopes can be naturally processed. The use of aset of long peptides to stimulate T cells is expected tomake this approach universally applicable to all stemcell donors irrespective of their HLA-type. By applyingpeptides that can easily be generated according toGMP conditions, the use of infectious wild-type HAdVor replication-deficient virus, which might recombinewith the wild-type virus in the patient, can be avoided.
The results from this study may also haveimplications for the use of recombinant adenoviralvectors for gene therapy and vaccination. Pre-existingimmunity against the adenoviral vector may limit theapplication of HAdV5 for both gene therapy protocolsand vaccine delivery. To circumvent the neutralizingantibody response to the HAdV-based vector, variousstrategies are being pursued including the use of otherhuman serotypes [33] and non-human adenoviruses[34]. Recent studies in mice suggest that T cells mayalso have a role in limiting the potency of HAdV5 forgene transfer and as a vehicle for vaccination [35]. Ourresults indicate that human HAdV-specific T cellsrecognize conserved peptides of the hexon proteinand, therefore, will recognize vectors derived fromother serotypes. In a murine model, no evidence forT cells cross-reactive between HAdV serotypes 5 and35 was found using artificial induction of immunityagainst HAdV [36]. However, the relevance of murinestudies for the situation in humans may be limited,since the human immune system has usually encoun-Ta
ble4.
Tce
llscu
lturedwithHAdV5PC
Preco
gnizeHAdV
from
differentsp
eciesa
)
CulturedwithPC
P(II42,
II50
,II57
,II61
,II64
)CulturedwithII42
,II50
,II57
,II61
Donor
1516
1718
1617
Spec
ies
IFN-c
Prol
IFN-c
Prol
IFN-c
Prol
IFN-c
Prol
IFN-c
Prol
IFN-c
Prol
Nostim
ulus
3.1
3.6�
0.7
0.3
3.2�
0.5
0.1
3.7�
1.0
0.6
5.0�
1.9
0.4
1.1�
0.0
0.0
4.4�
0.8
HAdV12
A8.8
8.3�
1.8
1.9
10.4
�0.8
9.6
77.3
�9.9
0.5
42.7
�3.3
2.5
6.9�
1.0
8.6
87.5
�13
.0
HAdV18
A9.7
9.9�
0.6
1.6
9.3�
2.0
9.5
68.8
�9.2
1.1
17.8
�2.7
2.6
12.7
�2.6
9.1
52.7
�8.1
HAdV16
B31
.029
.1�
1.4
1.9
8.9�
0.8
9.7
59.6
�6.6
1.0
11.9
�3.8
2.8
6.8�
1.1
8.7
99.5
�10
.1
HAdV35
B12
.516
.3�
1.6
1.1
1.9�
1.0
8.5
24.0
�2.9
1.3
14.4
�1.9
2.3
1.5�
0.3
8.3
50.8
�15
.4
HAdV2
C13
.517
.9�
1.8
2.9
24.6
�8.8
12.9
90.9
�27
.71.1
19.3
�1.9
3.4
26.5
�4.9
12.1
99.1
�6.9
HAdV5
C10
.011
.0�
0.8
1.2
7.4�
0.7
12.3
97.8
�15
.51.0
15.5
�1.8
2.7
3.5�
1.4
ND
80.0
�6.3
HAdV6
C12
.419
.1�
2.9
1.1
18.2
�3.5
11.1
169.6�
24.3
1.1
21.8
�4.5
2.9
17.5
�1.6
11.4
102.6�
25.5
HAdV39
D7.3
6.2�
1.1
1.4
13.8
�2.2
9.8
151.1�
24.4
0.9
21.9
�4.5
2.8
10.4
�2.1
9.5
158.1�
21.0
a)Cellsweretested
forreco
gnitionofa
utologo
usPB
MCpulsed
withMB-inac
tiva
tedHAdVfrom
differentsp
ecies(M
OI2
000)
byintrac
ellularIFN-c
staining[(IFN-c)%
IFN-c
+ce
llsof
theCD3+
CD4+
lym
phocy
tes]
andpro
liferation
[(Prol)m
ean�
SD
�10
00cp
m].Med
ium
alone,
referred
toas
nostim
ulus,
was
use
das
neg
ativeco
ntrol.ND,n
otdeterm
ined
.
Louise A. Veltrop-Duits et al. Eur. J. Immunol. 2006. 36: 2410–24232418
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
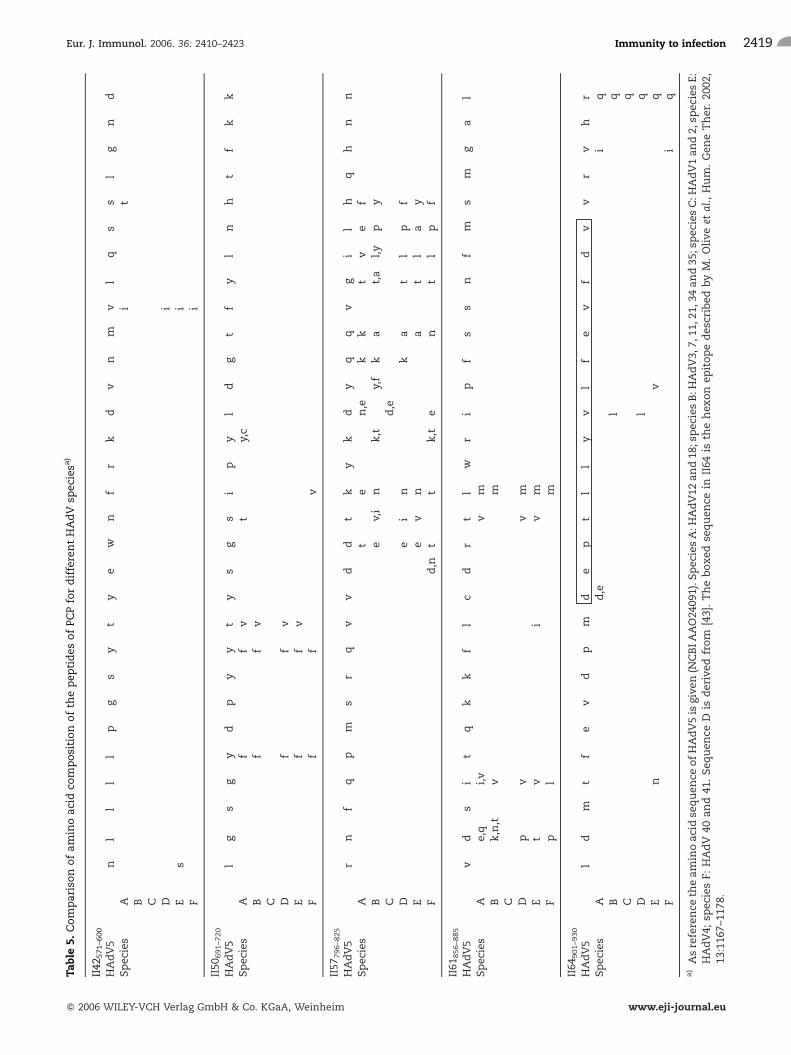
Table5.
Com
parisonofam
inoac
idco
mposition
ofth
epep
tides
ofPC
PfordifferentHAdV
spec
iesa
)
II42
571–6
00
HAdV5
nl
ll
lp
gs
yt
ye
wn
fr
kd
vn
mv
lq
ss
lg
nd
Spec
ies
Ai
tB C D
iE
si
Fi
II50
691–7
20
HAdV5
lg
sg
yd
py
yt
ys
gs
ip
yl
dg
tf
yl
nh
tf
kk
Spec
ies
Af
fv
ty,c
Bf
fv
C Df
fv
Ef
fv
Ff
fv
II57
796–8
25
HAdV5
rn
fq
pm
sr
qv
vd
dt
ky
kd
yq
qv
gi
lh
qh
nn
Spec
ies
At
en,e
kk
tv
ef
Be
v,i
nk,t
y,f
ka
t,a
l,y
py
Cd,e
De
in
ka
tl
pf
Ee
vn
at
la
yF
d,n
tt
k,t
en
tl
pf
II61
856–8
85
HAdV5
vd
si
tq
kk
fl
cd
rt
lw
ri
pf
ss
nf
ms
mg
al
Spec
ies
Ae,q
i,v
vm
Bk,n,t
vm
C Dp
vv
mE
tv
iv
mF
pl
m
II64
901–9
30
HAdV5
ld
mt
fe
vd
pm
de
pt
ll
yv
lf
ev
fd
vv
rv
hr
Spec
ies
Ad,e
iq
Bl
qC
qD
lq
En
vq
Fi
q
a)Asreference
theam
inoac
idse
quen
ceofHAdV5is
give
n(N
CBIA
AO24
091).S
pec
iesA:H
AdV12
and18
;spec
iesB:H
AdV3,
7,11
,21,
34an
d35
;spec
iesC:H
AdV1an
d2,
spec
iesE:
HAdV4;
spec
iesF:
HAdV
40an
d41
.Seq
uen
ceD
isderived
from
[43].Theboxed
sequen
cein
II64
isth
ehex
on
epitopedes
cribed
byM.Olive
etal.,Hum.Gen
eTher.20
02,
13:116
7–11
78.
Eur. J. Immunol. 2006. 36: 2410–2423 Immunity to infection 2419
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

tered multiple serotypes of HAdV and, therefore, cross-reactivity may occur more frequently. Detailed studiesare warranted to better understand the influence ofcross-reactive T cells on gene therapy procedures andvaccination potency.
In conclusion, a set of five long HAdV5 peptidesderived from the conserved region of the hexon proteinwas recognized by T cells of most adult healthy donors.With this peptide pool, HAdV-specific T cells weregenerated that not only respond to HAdV5 but also toother HAdV serotypes belonging to various species. Thedata suggest that these peptides are promising candi-dates for the generation of HAdV-specific T cells foradoptive immunotherapy, as peptides can easily beprepared under GMP conditions. Infusion of HAdV-specific T cells may be a valuable tool to decrease thehigh mortality rate due to HAdV viremia in immuno-compromised pediatric patients after allogeneic stemcell transplantation.
Materials and methods
PBMC
PBMC were isolated from blood of anonymous healthy blood-bank donors; these donors were typed for HLA-A, B, C andHLA-DR. Donors were asked for consent as approved by theInstitutional Review Board. The HLA types of the selecteddonors represented the normal distribution of HLA alleles inthe Dutch population.
Antigens
The HAdV5 hexon gene in plasmid pJM17 (kindly provided byProf. R. Hoeben, Department of Molecular Cell Biology,Leiden, The Netherlands) was amplified by PCR in fouroverlapping parts (Table 6). The PCR products were cloned byGateway technology (Invitrogen, San Diego, CA) in a bacterialexpression vector containing an N-terminal histidine tag. The
proteins were over expressed in Escherichia coli BL21 (DE3),purified, dissolved in DMSO and diluted in PBS [37]. The fouroverlapping parts are indicated by the numbers of first and lastamino acid in the protein (IIA1–273, IIB245–509, IIC479–743,IID720–952). Recombinant HAdV5 penton base and E1A proteinhad been produced before [37, 38].
Sixty-three peptides (30-mers with an overlap of 15 aminoacids) spanning the HAdV5 hexon protein are indicated as II4to II66. Peptides were synthesized by solid-phase strategiesusing a standard Fmoc-protocol on TentagelS AC resins usingan automated multiple peptide synthesizer (Syro II, Multi-syntech, Witten, Germany). Peptides were cleaved with TFA/water, 95/5, v/v, containing 5% ethanethiol (W-containingpeptides) and triethylsilane (C-containing peptides). Crudepeptides were purified by an optimized ether-precipitationprotocol. Peptides were dissolved in acetic acid/water 10/90,v/v, and lyophilized. Peptide purity was analyzed by reversedphase HPLC, indicating a purity of 90–95%, except for peptideII64 that contains also some parts of peptide II64. Peptideintegrity was analyzed by matrix-assisted laser desorption/ionization-time of flight mass spectrometry (MALDI-TOF MS)(Voyager DE Pro, ABI, Framingham, MA) using internalcontrols of known molecular mass for calibration. All peptideswere shown to have the expected molecular masses within anexperimental error of 0.5 Da. Peptide samples were dissolvedin DMSO and diluted in PBS. The 63 peptides were grouped in12 pools; each pool containing 5–6 adjacent peptides (e.g.peptide pool P1 consisted of peptide II4 to II9, pool P2 ofpeptides II10 to II15, etc).
Human adenovirus
Wild-type HAdV2, HAdV5, HAdV6, HAdV12, and HAdV35(RIVM, Bilthoven, The Netherlands) and HAdV16, HAdV18and HAdV39 (Crucell BV) were grown on HEp2 cells andpurified [9]. E1-E3- virus (AV1.0CMV/293-V142) was a kindgift of Prof. Dr. R. Hoeben. Methylene blue (MB) inactivationof virus was performed as previously described [39]. Viralparticle numbers were determined by measuring viral DNAand protein content at OD 260 nm, in which OD of 1.0corresponds to 1.1 � 1012 viral particles/mL. MOI is defined asratio of virus particles to cells.
Table 6. Primers for recombinant parts of the hexon protein
Recombinantprotein
Primer Nucleotide ofpolypeptide II gene
Specific part for cloning system Specific hexon sequence
IIA1–273 IIA-sense 225 - 244 50-ggggacaagtttgtacaaaaaagcaggctta- atggctaccccttcgat
IIA-antisense 1043 - 1026 50-ggggaccactttgtacaagaaagctgggtca- ggtcgcctcagtagttg
IIB245–509 IIB-sense 960 - 977 50-ggggacaagtttgtacaaaaaagcaggctta- gggcaaggcattcttgt
IIB-antisense 1751 - 1732 50-ggggaccactttgtacaagaaagctgggtca- caccactcgcttgttca
IIC479–743 IIC-sense 1659 - 1676 50-ggggacaagtttgtacaaaaaagcaggctta- ctgtatttgcccgacaag
IIC-antisense 2453 - 2436 50-ggggaccactttgtacaagaaagctgggtca- aaactcgttgggggtaag
IID720–952 IID-sense 2382 - 2400 50-ggggacaagtttgtacaaaaaagcaggctta- aaggtggccattaccttt
IID-antisense 3083 - 3066 50-ggggaccactttgtacaagaaagctgggtca- ttatgttgtggcgttgcc
Louise A. Veltrop-Duits et al. Eur. J. Immunol. 2006. 36: 2410–24232420
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

T cell stimulation
PBMC were cultured in RPMI 1640 (GibcoBRL, Grand Island,NY)medium supplemented with 10%human AB serum (AB) at2.0 � 106 cells/mL. Cells were stimulated with 5 lg/mLrecombinant protein or peptide, HAdV5 at MOI of 200, or withmedium alone as negative control. Cells were cultured for 4 or5 days either in a round-bottom 96-wells plate (CorningIncorporated, Life Sciences, Acton, MA) in order to measurethe amount of secreted IFN-c by ELISA or in a 48-wells plate(Corning Incorporated) to determine the IFN-c SFU/105 PBMCby ELISPOT or by intracellular cytokine staining and flowcytometry.
IFN-c levels in culture supernatant
To determine which peptide pools were recognized by PBMC ofhealthy donors, we measured the level of IFN-c secreted byPBMC in culture supernatants after 4 days. Secreted IFN-cwasdetermined using an ELISA kit (Sanquin, Amsterdam, TheNetherlands). The response to a distinct peptide pool wasconsidered positive if the IFN-c production to this pool was atleast 10% of the sum of IFN-c production in response to all 12peptide pools.
IFN-c ELISPOT
Recognition of the recombinant proteins or hexon peptides byT cells was analyzed by ELISPOT as described [23, 40]. After4 days of culture cells in each culture condition wereresuspended and transferred to three wells of an anti-IFN-cIgG antibody (MabTech, Nacha, Sweden) coated multiscreen96-wells plate (Millipore, Etten-Leur, The Netherlands), whichwas blocked for nonspecific binding by 10% FCS (GibcoBRL) inRPMI, at a concentration of 1.0 � 105 cells per well. After 16 hof incubation, SFU were visualized using anti-IFN-c IgGantibody MAb 7-B6-1-biotin (MabTech), Avidin-AP (Sigma-Aldrich, St. Louis, MO), followed by the substrate NBT-BCIP(NBT and 5-bromo-4-chloro-3-indolyl phosphate, Sigma-Aldrich) in 0.1 M Tris-HCl pH 9.5 and 5 mM MgCl2. Betweeneach detection step the plate was washed with PBS/0.05%Tween-20. Spots with a diameter >35 lmwere countedby computer-assisted video image analysis using KS ELISPOTsoftware release 4.1 (Carl Zeiss Vision GmbH, Hallbergmoos,Germany). The response was considered as positive if themeanof the number of spots in triplicate wells was >25 andexceeded the mean of the negative control + 3 SD.
Intracellular cytokine staining
To determine the phenotype of the IFN-c-producing cells, cellswere incubated at day 5 for 5 h with 5 lg/mL brefeldin A(BFA, Sigma-Aldrich) for intracellular IFN-c staining. Theprocedure was performed as described [41]. In short, cellswere washed with PBS containing 0.2% w/v NaN3 and fixedwith freshly made 4% paraformaldehyde (Sigma-Aldrich).Cells were permeabilized with PBS/NaN3 containing 0.1%saponin and 0.5% BSA, and nonspecific binding was blockedby incubating with PBS/NaN3/saponin/BSA/10% FCS. Cellswere stained with anti-CD3-PerCP-Cy5.5, anti CD4-FITC, anti-
IFN-c-PE (Becton Dickinson Biosciences, San Jose, CA) andanti-CD8-APC (Beckman Coulter, Miami, FL). Cells wereanalyzed on a FACSCalibur flow cytometer using CellQuestsoftware (Becton Dickinson).
Dendritic cell generation
For the generation of DC, PBMC were cultured at aconcentration of 2.0–3.0–106 cells/ml in a T75 flask in RPMIenriched with 10% FCS. After 2 h, the cells were washed andthe non-adherent fraction (PBL) was removed. The adherentfraction was cultured in RPMI with 10% FCS supplementedwith 800 U/mL recombinant GM-CSF (rGM-CSF, Leucomax,Novartis Pharma, Basel, Switzerland) and 40 ng/mL recombi-nant IL-4 (Peprotech, Rocky Hill, NJ) for 5 days. These cells,immature DC, were CD14– and CD1a+ as measured by FACSanalysis. The immature DCwere transferred to a round-bottom96-wells plate at a concentration of 1.0� 104 DC/well in RPMIenriched with 10%AB supplemented with 800 U/mL rGM-CSFand 40 ng/mL IL-4. After 24 h, the cells were matured byadding 0.25 lg/mL anti-CD40 (Mab89, Immunotech, Beck-man-Coulter, Miami, FL) and 500 U/mL IFN-c for anadditional 24 h. Mature DC were typically CD14– andCD83+, as assessed by FACS analysis [42].
T cell culture
Mature DC were loaded with peptides (range 1.0 lg/mL to0.08 ng/mL) in a volume of 25 lL/well of RPMI containing1 mM of Ca2+/Mg2+ at 37�C. The optimal concentration ofPCP was found to be 0.4 ng/mL. After 2 h of incubation,autologous PBL were added at a DC:PBL ratio of 1:20 in 200 lLRPMI/10%AB supplemented with 10 IU/mL of IL-2 (Proleu-kin, Chiron, Emeryville, CA) and 2 ng/mL of IL-7 (Peprotech).
At day 12, responding cells were restimulated usingautologous irradiated PBMC (30 Gray) loaded with the sameconcentration of peptides as used for loading the DC at theinitiation of the culture in RPMI/0.5%BSA at 1.0 � 107 cells/mL for 2 h. The T cells were co-cultured with the stimulatorcells in a T cell:stimulator ratio of 1:5 in a 24-wells plate inRPMI/10%AB in the presence of 0.1 lg/mL anti-CD40 and5 ng/mL IL-7. At day 19, 10 U/mL IL-2 and 5 ng/mL IL-7 wereadded. At days 12 and 26 cells were tested for specificity.
Proliferation and IFN-c production by peptide-specificT cells
To test T cell specificity, autologous irradiated PBMC(stimulator cells) were infected at day –1 with HAdV5 at aMOI of 2000 or loaded with 10 lg/mL peptide, or mediumalone as a negative control in RPMI/0.5%BSA at 1.0 � 107
cells/mL for 2 h and left overnight at 1.0 � 106 cells/mL inRPMI/10% AB.
For intracellular IFN-c production 2.0� 105 responder cellswere co-cultured with 2.0 � 105 stimulator cells in 400 lL in apolystyrene round-bottom tube (5 mL Falcon, Becton Dick-inson, Franklin Lakes, NJ). After 1 h, 5 lg/mL BFAwas added,and 16 h later intracellular staining was performed asdescribed above.
Eur. J. Immunol. 2006. 36: 2410–2423 Immunity to infection 2421
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

To determine the specific proliferative response, 2.0 � 104
responder cells were co-culturedwith 1.0�105 stimulator cellsin triplicate in a 96-wells round-bottom plate. At day 5,[3H]thymidine (0.5 lCi/well, Amersham International, Amer-sham, UK) was added for 16 h.
Acknowledgements: We would like to thank MartijnRabelink (Department of Molecular Cell Biology, Leiden)for virus purification and Dr. Jan-Wouter Drijfhout(Department of Immunohematology and Bloodtransfu-sion, Leiden) for synthesizing the peptides. This studiedwas supported by the Dutch Cancer Society (RUL-2001–2492) and the European Commission (FP5, QLK2-CT-2002–01432).
References
1 Flomenberg, P., Babbitt, J., Drobyski, W. R., Ash, R. C., Carrigan, D. R.,Sedmak, G. V., McAuliffe, T. et al, Increasing incidence of adenovirusdisease in bone marrow transplant recipients. J. Infect. Dis. 1994. 169:775–781.
2 Hierholzer, J. C., Adenoviruses in the immunocompromised host. Clin.Microbiol. Rev. 1992. 5: 262–274.
3 Carrigan, D. R., Adenovirus infections in immunocompromised patients.Am. J. Med. 1997. 102: 71–74.
4 Hale, G. A., Heslop, H. E., Krance, R. A., Brenner, M. A., Jayawardene, D.,Srivastava, D. K. and Patrick, C. C., Adenovirus infection after pediatricbone marrow transplantation. Bone Marrow Transplant. 1999. 23: 277–282.
5 Howard, D. S., Phillips II, G. L., Reece, D. E., Munn, R. K., Henslee-Downey, J., Pittard, M., Barker, M. and Pomeroy, C., Adenovirusinfections in hematopoietic stem cell transplant recipients. Clin. Infect.Dis. 1999. 29: 1494–1501.
6 Leen, A. M. and Rooney, C. M., Adenovirus as an emerging pathogen inimmunocompromised patients. Br. J. Haematol. 2005. 128: 135–144.
7 Lion, T., Baumgartinger, R., Watzinger, F., Matthes-Martin, S., Suda, M.,Preuner, S., Futterknecht, B. et al.,Molecular monitoring of adenovirus inperipheral blood after allogeneic bone marrow transplantation permits earlydiagnosis of disseminated disease. Blood 2003. 102: 1114–1120.
8 Bordigoni, P., Carret, A. S., Venard, V.,Witz, F. and Le Faou, A., Treatmentof adenovirus infections in patients undergoing allogeneic hematopoieticstem cell transplantation. Clin. Infect. Dis. 2001. 32: 1290–1297.
9 Heemskerk, B., Lankester, A. C., van Vreeswijk, T., Beersma,M. F., Claas,E. C., Veltrop-Duits, L. A., Kroes, A. C. et al., Immune reconstitution andclearance of human adenovirus viremia in pediatric stem-cell recipients. J.Infect. Dis. 2005. 191: 520–530.
10 Baldwin, A., Kingman, H., Darville, M., Foot, A. B., Grier, D., Cornish, J.M., Goulden, N. et al., Outcome and clinical course of 100 patients withadenovirus infection following bone marrow transplantation. Bone MarrowTransplant. 2000. 26: 1333–1338.
11 Runde, V., Ross, S., Trenschel, R., Lagemann, E., Basu, O., Renzing-Kohler, K., Schaefer, U.W. et al., Adenoviral infection after allogeneic stemcell transplantation (SCT): report on 130 patients from a single SCT unitinvolved in a prospective multi center surveillance study. Bone MarrowTransplant. 2001. 28: 51–57.
12 van Tol, M. J., Kroes, A. C., Schinkel, J., Dinkelaar, W., Claas, E. C., Jol-van der Zijde, C. M. and Vossen, J. M., Adenovirus infection in paediatricstem cell transplant recipients: increased risk in young children with adelayed immune recovery. Bone Marrow Transplant. 2005. 36: 39–50.
13 Lankester, A. C., Heemskerk, B., Claas, E. C., Schilham, M. W., Beersma,M. F., Bredius, R. G., van Tol, M. J. and Kroes, A. C., Effect of ribavirin onthe plasma viral DNA load in patients with disseminating adenovirusinfection. Clin. Infect. Dis. 2004. 38: 1521–1525.
14 Miyamura, K., Hamaguchi, M., Taji, H., Kanie, T., Kohno, A., Tanimoto,M., Saito, H. et al., Successful ribavirin therapy for severe adenovirushemorrhagic cystitis after allogeneic marrow transplant from close HLAdonors rather than distant donors. Bone Marrow Transplant. 2000. 25:545–548.
15 Hoffman, J. A., Shah, A. J., Ross, L. A. and Kapoor, N., Adenoviralinfections and a prospective trial of cidofovir in pediatric hematopoietic stemcell transplantation. Biol. Blood Marrow Transplant. 2001. 7: 388–394.
16 Ljungman, P., Ribaud, P., Eyrich, M., Matthes-Martin, S., Einsele, H.,Bleakley, M., Machaczka, M. et al., Cidofovir for adenovirus infectionsafter allogeneic hematopoietic stem cell transplantation: a survey by theInfectious Diseases Working Party of the European Group for Blood andMarrow Transplantation. Bone Marrow Transplant. 2003. 31: 481–486.
17 Leruez-Ville, M., Minard, V., Lacaille, F., Buzyn, A., Abachin, E., Blanche,S., Freymuth, F. and Rouzioux, C., Real-time blood plasma polymerasechain reaction for management of disseminated adenovirus infection. Clin.Infect. Dis. 2004. 38: 45–52.
18 Chakrabarti, S., Collingham, K. E., Fegan, C. D., Pillay, D. and Milligan,D. W., Adenovirus infections following haematopoietic cell transplantation:is there a role for adoptive immunotherapy? Bone Marrow Transplant. 2000.26: 305–307.
19 Hromas, R., Cornetta, K., Srour, E., Blanke, C. and Broun, E. R., Donorleukocyte infusion as therapy of life-threatening adenoviral infections afterT-cell-depleted bone marrow transplantation. Blood 1994. 84: 1689–1690.
20 Flomenberg, P., Piaskowski, V., Truitt, R. L. and Casper, J. T.,Characterization of human proliferative T cell responses to adenovirus. J.Infect. Dis. 1995. 171: 1090–1096.
21 Flomenberg, P., Piaskowski, V., Truitt, R. L. and Casper, J. T., Humanadenovirus-specific CD8+ T-cell responses are not inhibited by E3–19 K inthe presence of gamma interferon. J. Virol. 1996. 70: 6314–6322.
22 Smith, C. A., Woodruff, L. S., Kitchingman, G. R. and Rooney, C. M.,Adenovirus-pulsed dendritic cells stimulate human virus-specific T-cellresponses in vitro. J. Virol. 1996. 70: 6733–6740.
23 Heemskerk, B., Veltrop-Duits, L. A., van Vreeswijk, T., ten Dam, M. M.,Heidt, S., Toes, R. E., van Tol, M. J. and Schilham, M. W., Extensive cross-reactivity of CD4+ adenovirus-specific T cells: implications for immunother-apy and gene therapy. J. Virol. 2003. 77: 6562–6566.
24 Olive, M., Eisenlohr, L. C. and Flomenberg, P., Quantitative analysis ofadenovirus-specific CD4+ T-cell responses from healthy adults. ViralImmunol. 2001. 14: 403–413.
25 Leen, A.M., Sili, U., Vanin, E. F., Jewell, A.M., Xie,W., Vignali, D., Piedra,P. A. et al., Conserved CTL epitopes on the adenovirus hexon protein expandsubgroup cross-reactive and subgroup-specific CD8+ T cells. Blood 2004.104: 2432–2440.
26 Feuchtinger, T., Lang, P., Hamprecht, K., Schumm,M., Greil, J., Jahn, G.,Niethammer, D. and Einsele, H., Isolation and expansion of humanadenovirus-specific CD4+ and CD8+ T cells according to IFN-gammasecretion for adjuvant immunotherapy. Exp. Hematol. 2004. 32: 282–289.
27 Trivedi, D., Williams, R. Y., O'Reilly, R. J. and Koehne, G., Generation ofCMV-specific T lymphocytes using protein-spanning pools of pp65-derivedoverlapping pentadecapeptides for adoptive immunotherapy. Blood 2005.105: 2793–2801.
28 Mahr, J. A. and Gooding, L. R., Immune evasion by adenoviruses. Immunol.Rev. 1999. 168: 121–130.
29 Olive, M., Eisenlohr, L., Flomenberg, N., Hsu, S. and Flomenberg, P., Theadenovirus capsid protein hexon contains a highly conserved human CD4+
T-cell epitope. Hum. Gene Ther. 2002. 13: 1167–1178.
30 Tang, J., Olive, M., Champagne, K., Flomenberg, N., Eisenlohr, L., Hsu, S.and Flomenberg, P., Adenovirus hexon T-cell epitope is recognized by mostadults and is restricted by HLA DP4, the most common class II allele. GeneTher. 2004. 11: 1408–1415.
31 Leen, A. M., Sili, U., Savoldo, B., Jewell, A. M., Piedra, P. A., Brenner, M.K. and Rooney, C.M., Fiber-modified adenoviruses generate subgroupcross-reactive, adenovirus-specific cytotoxic T lymphocytes for therapeuticapplications. Blood 2004. 103: 1011–1019.
32 van der Burg, S. H., Kwappenberg, K. M., Geluk, A., van der Kruk,. M.,Pontesilli, O., Hovenkamp, E., Franken, K. L. et al., Identification of aconserved universal Th epitope in HIV-1 reverse transcriptase that is
Louise A. Veltrop-Duits et al. Eur. J. Immunol. 2006. 36: 2410–24232422
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

processed and presented to HIV-specific CD4+ T cells by at least fourunrelated HLA-DR molecules. J. Immunol. 1999. 162: 152–160.
33 Vogels, R., Zuijdgeest, D., van Rijnsoever, R., Hartkoorn, E., Damen, I.,de Bethune, M. P., Kostense, S. et al., Replication-deficient humanadenovirus type 35 vectors for gene transfer and vaccination: efficienthuman cell infection and bypass of preexisting adenovirus immunity. J.Virol. 2003. 77: 8263–8271.
34 Xiang, Z., Gao, G., Reyes-Sandoval, A., Cohen, C. J., Li, Y., Bergelson, J.M., Wilson, J. M. and Ertl, H. C., Novel, chimpanzee serotype 68-basedadenoviral vaccine carrier for induction of antibodies to a transgene product.J. Virol. 2002. 76: 2667–2675.
35 Sumida, S. M., Truitt, D. M., Kishko, M. G., Arthur, J. C., Jackson, S. S.,Gorgone, D. A., Lifton, M. A. et al., Neutralizing antibodies and CD8+ Tlymphocytes both contribute to immunity to adenovirus serotype 5 vaccinevectors. J. Virol. 2004. 78: 2666–2673.
36 Barouch, D. H., Pau, M. G., Custers, J. H., Koudstaal, W., Kostense, S.,Havenga, M. J., Truitt, D. M. et al., Immunogenicity of recombinantadenovirus serotype 35 vaccine in the presence of pre-existing anti-ad5immunity. J. Immunol. 2004. 172: 6290–6297.
37 Franken, K. L., Hiemstra, H. S., van Meijgaarden, K. E., Subronto, Y., denHartigh, J., Ottenhoff, T. H. and Drijfhout, J.W., Purification of his-taggedproteins by immobilized chelate affinity chromatography: the benefits fromthe use of organic solvent. Protein Expr. Purif. 2000. 18: 95–99.
38 Boonman, Z. F., van Mierlo, G. J., Fransen, M. F., Franken, K. L.,Offringa, R., Melief, C. J., Jager, M. J. and Toes, R. E., Intraocular tumor
antigen drains specifically to submandibular lymph nodes, resulting in anabortive cytotoxic T cell reaction. J. Immunol. 2004. 172: 1567–1574.
39 Li, Q. G., Lindman, K. and Wadell, G., Hydropathic characteristics ofadenovirus hexons. Arch. Virol. 1997. 142: 1307–1322.
40 van der Burg, S. H., Ressing, M. E., Kwappenberg, K. M., de Jong, A.,Straathof, K., de Jong, J., Geluk, A. et al., Natural T-helper immunityagainst human papillomavirus type 16 (HPV16) E7-derived peptide epitopesin patients with HPV16-positive cervical lesions: identification of 3 humanleukocyte antigen class II-restricted epitopes. Int. J. Cancer 2001. 91:612–618.
41 de Jong, A., van der Burg, S. H., Kwappenberg, K. M., van der Hulst, J.M., Franken, K. L., Geluk, A., van Meijgaarden, K. E. et al., Frequentdetection of human papillomavirus 16 E2-specific T-helper immunity inhealthy subjects. Cancer Res. 2002. 62: 472–479.
42 Kalinski, P., Hilkens, C. M., Wierenga, E. A. and Kapsenberg, M. L., T-cellpriming by type-1 and type-2 polarized dendritic cells: the concept of a thirdsignal. Immunol. Today 1999. 20: 561–567.
43 Ebner, K., Pinsker, W. and Lion, T., Comparative sequence analysis of thehexon gene in the entire spectrum of human adenovirus serotypes:phylogenetic, taxonomic, and clinical implications. J. Virol. 2005. 79:12635–12642.
44 Schagen, F. H., Moor, A. C., Cheong, S. C., Cramer, S. J., van Ormondt,H., van der Eb, A. J., Dubbelman, T. M. et al., Photodynamic treatment ofadenoviral vectors with visible light: An easy and convenient method forviral inactivation. Gene Ther. 1999. 6: 873–881.
Eur. J. Immunol. 2006. 36: 2410–2423 Immunity to infection 2423
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu