human cytomegalovirus persistently infects aortic ...jvi.asm.org/content/72/7/5661.full.pdftissue...
TRANSCRIPT

JOURNAL OF VIROLOGY,0022-538X/98/$04.0010
July 1998, p. 5661–5668 Vol. 72, No. 7
Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Human Cytomegalovirus Persistently Infects Aortic Endothelial CellsKENNETH N. FISH, CECILIA SODERBERG-NAUCLER,† LISA K. MILLS,
STEPHAN STENGLEIN,‡ AND JAY A. NELSON*
Department of Molecular Microbiology and Immunology, Oregon HealthSciences University, Portland, Oregon 97201
Received 15 January 1998/Accepted 8 April 1998
Endothelial cells (EC) have been implicated as constituting an important cell type in the pathogenesis ofhuman cytomegalovirus (HCMV). Microvascular and macrovascular EC exhibit different biochemical andfunctional properties depending on the organ of origin. Phenotypic differences between microvascular andmacrovascular EC may alter the ability of these cells to support HCMV replication. In this study, we comparedthe replication of HCMV in primary macrovascular aortic EC (AEC) with that in brain microvascular EC(BMVEC). An examination of IE72, pp65, and gB viral antigen expression in BMVEC and AEC by immuno-flourescence revealed similar frequencies of infected cells. Intracellular production of virus was 3 log unitsgreater in BMVEC than in AEC, while equal quantities of extracellular virus were produced in both cell types.HCMV infection of BMVEC resulted in rapid cellular lysis, while the virus was nonlytic and continuouslyreleased from HCMV-infected AEC for the life span of the culture. An examination of infected cells by electronmicroscopy revealed the formation of abundant nucleocapsids in both AEC and BMVEC. However, significantamounts of mature viral particles were only detected in the cytoplasm of BMVEC. These observations indicatethat levels of HCMV replication in EC obtained from different organs are distinct and suggest that persistentlyinfected AEC may serve as a reservoir of virus.
Human cytomegalovirus (HCMV) establishes a lifelong per-sistence in the host after primary infection. Although macro-phages from the peripheral blood have recently been identifiedas a site of HCMV latency in asymptomatically infected indi-viduals (21), another potential cell type latently or persistentlyinfected by HCMV is vascular endothelial cells (EC). Studiesof autopsy tissue from HCMV-seropositive transplant patientshave revealed that EC commonly harbor virus without obviouscytopathology (14). However, additional studies have indicatedthat EC constitute one of several cell types that exhibit cyto-megally in virus-infected tissues (6, 19, 24). An understandingof the role of EC in HCMV disease has been complicated bythe detection of viral DNA in arterial specimens from sero-positive individuals without active infection (12). These obser-vations have led to the speculation that EC may be a virusreservoir.
EC exhibit phenotypic differences that are dependent on theorigin (adult versus fetal), anatomical location, and vessel size(large vessel versus capillary) (16, 22). Human umbilical veinEC (HUVEC), which are commonly utilized for EC studies,are derived from fetal large-vessel tissue. These cells are func-tionally and biochemically distinct from EC derived from adulttissue such as large-vessel aortic EC (AEC) (16, 22). CapillaryEC not only display unique differences from large-vessel ECbut also demonstrate organ specificity. For example, humanbrain microvascular EC (BMVEC), which together with astro-cytes compose the blood-brain barrier, possess specific trans-porter systems that regulate the passage of specific metabolitesfrom the blood to the brain parenchyma (8, 13). These unique
properties differentiate BMVEC from EC in capillaries ofother tissues. The physiological and biochemical differencesbetween EC in different organs may affect the ability of HCMVto replicate in these cells.
HCMV infection of EC in vitro has been controversial. Earlystudies suggested that HCMV was unable to replicate in EC(4). However, others suggested that the virus could produc-tively infect a low percentage of cells in culture (20). In addi-tion, HCMV infectivity of EC was enhanced by serial passageof virus through these cells. Interestingly, viral infection ofHUVEC resulted in anchorage-independent growth and atransformed phenotype (23). An important consideration, how-ever, is that viral replication in HUVEC may not representviral replication in adult EC. In support of this hypothesis,other viruses have demonstrated exquisite cellular specificityconcerning their abilities to productively infect EC obtainedfrom different organs (13). Lathey et al. addressed this issuewhen they demonstrated that HCMV infected BMVEC moreefficiently than HUVEC, suggesting that the observed physio-logical differences between EC types may also affect viral rep-lication (10).
In the present study, we examined characteristics of HCMVreplication in AEC. We found that HCMV infection was notlytic and resulted in the accumulation of significant amounts ofextracellular but not intracellular virus. In addition, the cellcycle was not inhibited by HCMV and cells continuously re-leased infectious virus. These results contrast with those show-ing the rapid, lytic infection of BMVEC and HF cells. Theability of HCMV to infect AEC and to continuously produceextracellular virus and the absence of cytopathic effect are pre-requisites for establishing viral persistence. Therefore, HCMVinfection of AEC provides an ideal model to examine mecha-nisms of persistence in the human host.
MATERIALS AND METHODS
Culturing and infection of EC. BMVEC were a generous gift from AshleeMoses (Oregon Health Sciences University, Portland, Oreg.), and AEC werepurchased from Clonetics Normal Human Cell Systems (San Diego, Calif.).
* Corresponding author. Mailing address: Department of MolecularMicrobiology and Immunology, Oregon Health Sciences University,L220, 3181 S. W. Sam Jackson Park Rd., Portland, OR 97201. Phone:(503) 494-2434. Fax: (503) 494-6862. E-mail: [email protected].
† Present address: Karolinska Institute, Department for Biosciencesat Novum, Huddinge, Sweden.
‡ Present address: Abteilung fur Medizinische Virologie, Univer-sitat Tubingen, Tubingen, Germany.
5661
on June 3, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

BMVEC were cultured in Endo-SFM medium (GIBCO Laboratories, GrandIsland, N.Y.) containing 10% human AB serum (Sigma Chemical Co., St. Louis,Mo.), 1.0% penicillin-streptomycin solution (GIBCO), 1.0% glutamine (Sigma),heparin (40 mg/ml; Sigma), and EC growth factor (50 mg/ml; Sigma). AEC werecultured in the medium recommended by the manufacturer. Since BMVEC andAEC cultures represented single donors, each experiment was repeated in trip-licate with different donors. Low-passage-number (less than 5) BMVEC or AECwere plated in 35-mm2 Primaria culture dishes (Becton Dickinson, Lincoln Park,N.J.) or two-well coverslip bottom Lab-Tek chamber slides (Nunc, Inc., Naper-ville, Ill.) and allowed to grow at 37°C with 7% CO2 to 70% confluency prior toinfection with the HCMV laboratory strain Towne, a recent patient isolate Po(3), or the mutant laboratory strain AD169-pp65 (17). Heparin-free medium wasused at least 1 h prior to infection and throughout the infection time course.Supernatants from HCMV-infected HF cells were used as the source of the ECinoculum. For mock infections, HF supernatant virus was UV inactivated for 8 h.The virus titer was measured on HF cells by a plaque assay as previouslydescribed (3).
Immunofluorescence of HCMV IE and late antigens. HCMV-infected ECgrown on chamber slides were fixed for 20 min at room temperature in bufferedpicric acid-paraformaldehyde (2% paraformaldehyde and 15% buffered picricacid) and permeablized with 0.3% Triton X-100 in phosphate-buffered saline(PBS). Monolayers were blocked with 20% normal goat serum in PBS andincubated for 1 h at 37°C with one of the following antibodies raised againstHCMV gene products: a rabbit polyclonal immediate-early (IE)-specific anti-body (9), a monoclonal late antibody to pp65, or a monoclonal antibody againstgB (1). Cell surface gB antigen was detected prior to permeabilization of cells.The polyclonal von Willebrand’s factor (vWF) antibody was purchased fromDAKO Corp. (Carpinteria, Calif.). The binding of primary antibody was de-tected with fluorescein isothiocyanate-, tetramethyl rhodamine isocyanate-, orcyanine-5 (Biological Detection Systems, Inc., Pittsburgh, Pa.)-conjugated sec-ondary antibodies raised in the appropriate species and visualized on an uprightLeitz fluorescence microscope or a Leica confocal laser scanning microscopeequipped with a Leitz Fluorovert-FU microscope and an argon-krypton laser.The Slowfade antifade kit (Molecular Probes, Inc., Eugene, Oreg.) was utilizedto ensure minimal fluorescence fading.
FACS. Subconfluent primary AEC cultures at passage 7 were serum starvedfor 72 h. Starved cultures were mock infected or infected with either AD169 orTowne HCMV at a multiplicity of infection (MOI) of 5, followed by an addi-tional 12-h incubation period in serum-free media. During some experiments,phosphonoacetic acid (100 mg/ml) or 0.5 mM foscarnet was present in the mediafollowing infection. Cells were harvested both prior to adding serum to themedia, which was done at 12 h postinfection (hpi) and at intervals followingserum addition. Trypsinized cells were rinsed once in PBS and frozen at 270°Cin citrate-dimethyl sulfoxide (DMSO) buffer (40 mM citrate trisodium, 5%DMSO, 250 mM saccharose), pH 7.6 (18). Nuclei were isolated in nucleusisolation buffer (10 mM Tris-HCl [pH 7.5], 1 mM MgCl2, 0.2 mM phenylmeth-ylsulfonyl fluoride, 0.25 M sucrose, 0.5% Triton X-100) for 30 min on ice withoccasional vigorous vortexing, followed by a 5-min spin at 14,000 rpm in anEppendorf microcentrifuge at 4°C. This step was repeated two to four times untilgreater than 80% of the nuclei were clean. The nuclei were resuspended in 400ml of citrate stock solution (3.4 mM citrate trisodium, 0.1% IGEPAL CA-630(Sigma), 1.5 mM spermine tetrahydrochloride, 0.5 mM Tris-HCl) (18). One-halfof the nuclei were aliquoted into separate samples and stained with a 1:40dilution of the polyclonal IE rabbit antibody described above, followed by stain-ing with a goat anti-rabbit antibody conjugated with Cy2 (Amersham Life Sci-ences). All samples were resuspended in 200 ml of the citrate stock solution; then200 ml of a citrate stock solution containing 250 mg of RNase A per ml wasadded, and the samples were incubated for 10 min at room temperature. Finally,100 ml of a citrate stock solution containing 2.1 mg of propidium iodide per mlwas added, and the samples were incubated for 30 to 60 min at 4°C prior tofluorescence-assisted cell sorting (FACS) analysis. Becton Dickinson Cell QuestFACStation software (version 3.0.1) operating a Becton Dickinson FACSCaliburFACS scan instrument was used for analyzing the stained nuclei.
Tissue preparation for structural EM analysis. To examine virus-infected ECby electron microscopy (EM), uninfected and infected cells were harvested at 3,7, and 10 days after infection and fixed in 2% glutaraldehyde in 0.1 M sodiumcacodylate buffer (pH 7.4) at 4°C for 16 to 20 h. Fixed cells were collected byscraping and washed in 0.15 M sodium cacodylate (pH 7.4). Specimens werepostfixed in 1% osmium tetroxide in the same buffer for 1 h at 4°C, dehydratedin ethanol followed by acetone, and embedded in LX-112. Sections of uninfectedand infected cells on grids were washed and contrasted with uranyl acetate-oxalate for 5 min, embedded in 2% methyl cellulose containing 0.2% uranylacetate, and examined with a calibrated Philips 420 electron microscope at 80kV.
RESULTS
HCMV is lytic in BMVEC but not AEC. Since AEC arenaturally infected in vivo and represent a potential reservoir ofpersistent virus, we examined the ability of HCMV to infectprimary cultures of these large-vessel EC in vitro compared to
the ability of the virus to infect BMVEC. The EC cultures were.95% pure as determined by the presence of vWF (Fig. 1A).AEC and BMVEC subconfluent monolayers were infectedwith HCMV (Towne) at an MOI of 3 and examined at 3, 8, and14 days postinfection (dpi) by phase microscopy. HCMV in-fection of BMVEC resulted in the development of cytopathiceffect by 5 dpi and in lysis of 70% of the cells in culture at 14dpi (Fig. 1B and data not shown). Surprisingly, a cytopathiceffect was not observed in AEC infected with HCMV up to 30dpi, the in vitro life expectancy of these cells (Fig. 1B and 2Band data not shown).
To determine the frequency of HCMV infection, BMVECand AEC were fixed at various intervals postinfection andexamined for the presence of the HCMV IE antigen and eitherthe lower-matrix phosphoprotein pp65 or the major envelopeglycoprotein gB by double-label immunofluorescence (Fig. 2).By 3 dpi, 80% of the AEC and 50% of the BMVEC displayedthe presence of HCMV antigens. Although the frequency of IEantigen detection was greater in AEC, expression was delayedin these cells, with the first appearance of antigen at 24 hpostinfection (hpi) in contrast to 12 hpi in BMVEC (Fig. 2Aand B [red nuclei]). A minor delay in the expression of pp65and gB was also observed in AEC, in comparison to theirexpression in BMVEC. An examination of gB expression inBMVEC revealed the presence of antigen in intracellular vacu-oles and at the plasma membrane (PM) between 2 and 3 dpi(Fig. 2C [green and blue fluorescence, respectively]). In con-trast, gB was detected in infected AEC at 3 dpi in intracellularvacuoles but not at the PM (Fig. 2C [green fluorescence]).These results demonstrate that the frequency of infected cells,the kinetics of viral antigen expression, and the distribution ofviral antigens within the cell differ between HCMV-infectedBMVEC and AEC.
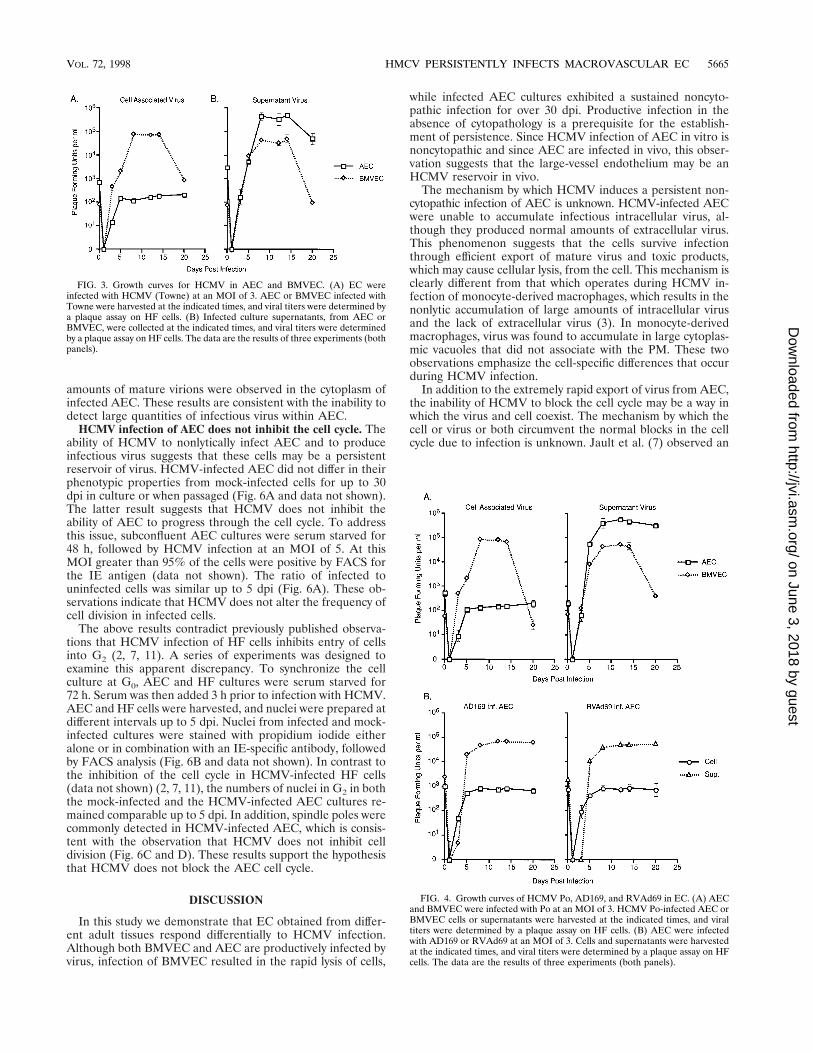
HCMV productively infects BMVEC and AEC. A one-stepgrowth curve was generated from HCMV-infected AEC andBMVEC cultures to assess the abilities of these cells to supportviral growth. An analysis of EC lysates revealed the productionof significant quantities of cell-associated virus in BMVEC butnot AEC (Fig. 3). Interestingly, similar amounts of virus weredetected in supernatants obtained from both the BMVEC andAEC cultures (Fig. 3). These observations suggest that infec-tious intracellular virus produced in AEC is rapidly exportedfrom infected cells in contrast to HCMV produced in infectedBMVEC.
To determine if the inability of AEC to accumulate intra-cellular HCMV is strain specific, EC cultures were infectedwith a recent isolate of HCMV (Po) (Fig. 4A), with AD169(Fig. 4B), and with the AD169 pp65 mutant (RVAd69) (17)(Fig. 4B). While strains Towne and Po are extremely lytic infibroblasts, AD169 and RVAd69 are more cell associated afterinfection. As seen in results obtained with the HCMV Townestrain, AEC infection by strains Po, AD169, and RVAd69resulted in modest intracellular production of infectious virusbut significant quantities of extracellular virus. Also consistentwith the growth of Towne in BMVEC, similar quantities ofintracellular and extracellular HCMV were produced by theother strains (Fig. 4A and data not shown). These observationsindicate that the inability of AEC to accumulate intracellularHCMV is not strain dependent.
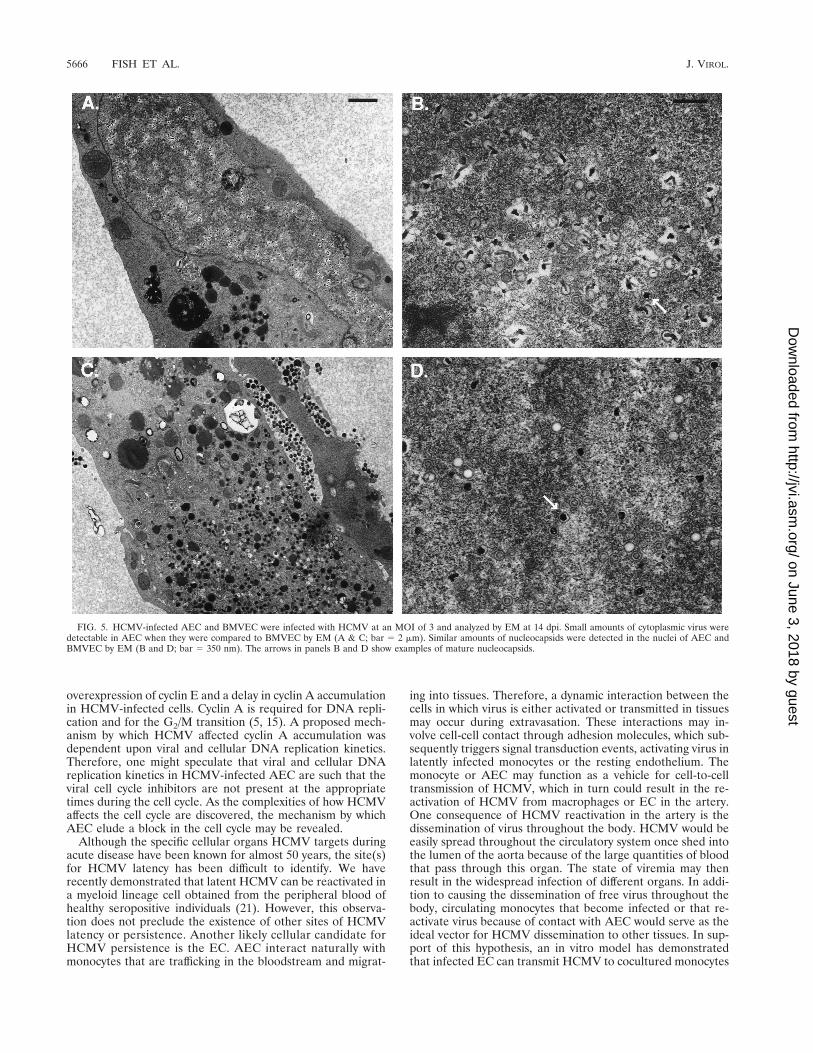
HCMV-infected AEC and BMVEC were analyzed by EM todetermine whether morphological differences occurred duringvirus assembly and egress. An examination of infected cellsrevealed that similar amounts of viral capsids were produced inthe nuclei of both AEC and BMVEC (Fig. 5). In contrast,while significant quantities of mature virus particles wereobserved within the cytoplasm of BMVEC, only minimal
5662 FISH ET AL. J. VIROL.
on June 3, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
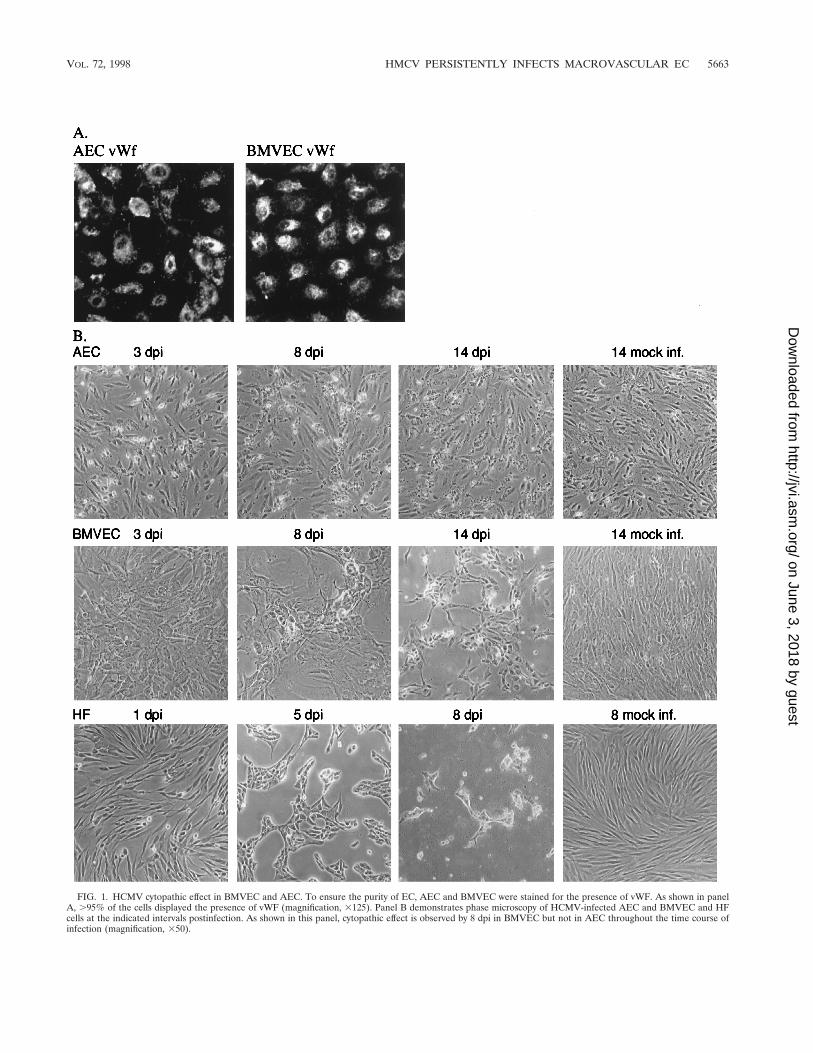
FIG. 1. HCMV cytopathic effect in BMVEC and AEC. To ensure the purity of EC, AEC and BMVEC were stained for the presence of vWF. As shown in panelA, .95% of the cells displayed the presence of vWF (magnification, 3125). Panel B demonstrates phase microscopy of HCMV-infected AEC and BMVEC and HFcells at the indicated intervals postinfection. As shown in this panel, cytopathic effect is observed by 8 dpi in BMVEC but not in AEC throughout the time course ofinfection (magnification, 350).
VOL. 72, 1998 HMCV PERSISTENTLY INFECTS MACROVASCULAR EC 5663
on June 3, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
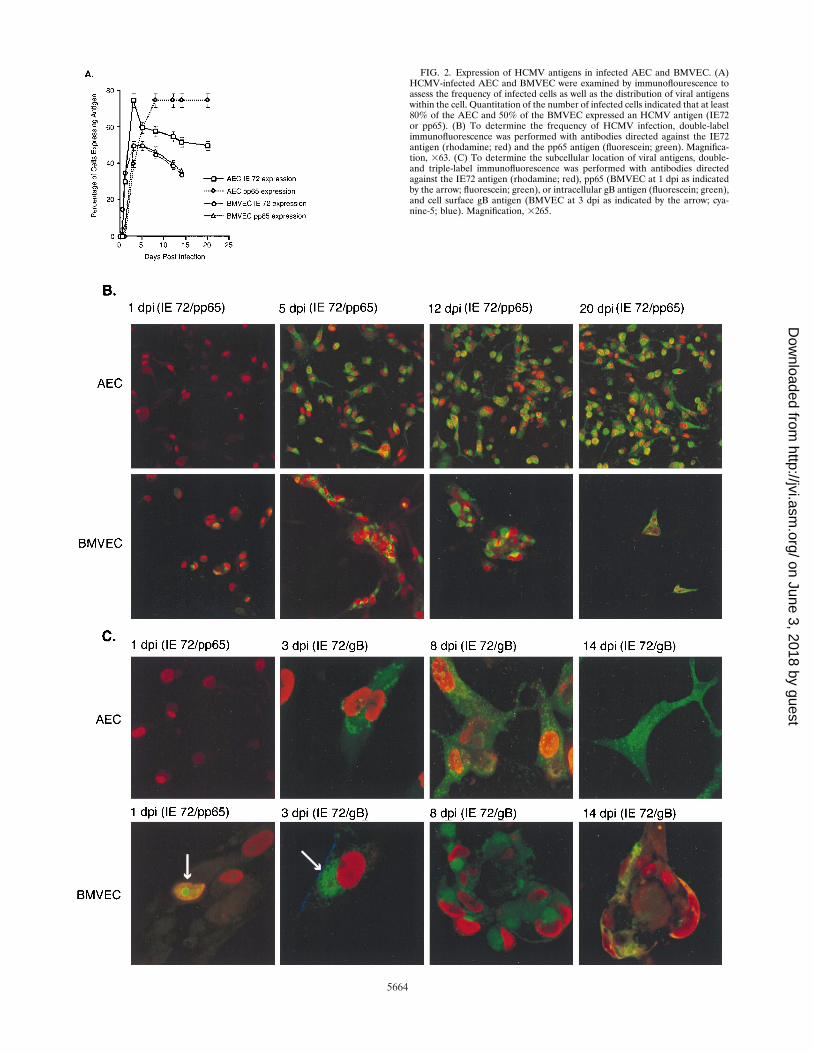
FIG. 2. Expression of HCMV antigens in infected AEC and BMVEC. (A)HCMV-infected AEC and BMVEC were examined by immunoflourescence toassess the frequency of infected cells as well as the distribution of viral antigenswithin the cell. Quantitation of the number of infected cells indicated that at least80% of the AEC and 50% of the BMVEC expressed an HCMV antigen (IE72or pp65). (B) To determine the frequency of HCMV infection, double-labelimmunofluorescence was performed with antibodies directed against the IE72antigen (rhodamine; red) and the pp65 antigen (fluorescein; green). Magnifica-tion, 363. (C) To determine the subcellular location of viral antigens, double-and triple-label immunofluorescence was performed with antibodies directedagainst the IE72 antigen (rhodamine; red), pp65 (BMVEC at 1 dpi as indicatedby the arrow; fluorescein; green), or intracellular gB antigen (fluorescein; green),and cell surface gB antigen (BMVEC at 3 dpi as indicated by the arrow; cya-nine-5; blue). Magnification, 3265.
5664
on June 3, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
amounts of mature virions were observed in the cytoplasm ofinfected AEC. These results are consistent with the inability todetect large quantities of infectious virus within AEC.
HCMV infection of AEC does not inhibit the cell cycle. Theability of HCMV to nonlytically infect AEC and to produceinfectious virus suggests that these cells may be a persistentreservoir of virus. HCMV-infected AEC did not differ in theirphenotypic properties from mock-infected cells for up to 30dpi in culture or when passaged (Fig. 6A and data not shown).The latter result suggests that HCMV does not inhibit theability of AEC to progress through the cell cycle. To addressthis issue, subconfluent AEC cultures were serum starved for48 h, followed by HCMV infection at an MOI of 5. At thisMOI greater than 95% of the cells were positive by FACS forthe IE antigen (data not shown). The ratio of infected touninfected cells was similar up to 5 dpi (Fig. 6A). These ob-servations indicate that HCMV does not alter the frequency ofcell division in infected cells.
The above results contradict previously published observa-tions that HCMV infection of HF cells inhibits entry of cellsinto G2 (2, 7, 11). A series of experiments was designed toexamine this apparent discrepancy. To synchronize the cellculture at G0, AEC and HF cultures were serum starved for72 h. Serum was then added 3 h prior to infection with HCMV.AEC and HF cells were harvested, and nuclei were prepared atdifferent intervals up to 5 dpi. Nuclei from infected and mock-infected cultures were stained with propidium iodide eitheralone or in combination with an IE-specific antibody, followedby FACS analysis (Fig. 6B and data not shown). In contrast tothe inhibition of the cell cycle in HCMV-infected HF cells(data not shown) (2, 7, 11), the numbers of nuclei in G2 in boththe mock-infected and the HCMV-infected AEC cultures re-mained comparable up to 5 dpi. In addition, spindle poles werecommonly detected in HCMV-infected AEC, which is consis-tent with the observation that HCMV does not inhibit celldivision (Fig. 6C and D). These results support the hypothesisthat HCMV does not block the AEC cell cycle.
DISCUSSION
In this study we demonstrate that EC obtained from differ-ent adult tissues respond differentially to HCMV infection.Although both BMVEC and AEC are productively infected byvirus, infection of BMVEC resulted in the rapid lysis of cells,
while infected AEC cultures exhibited a sustained noncyto-pathic infection for over 30 dpi. Productive infection in theabsence of cytopathology is a prerequisite for the establish-ment of persistence. Since HCMV infection of AEC in vitro isnoncytopathic and since AEC are infected in vivo, this obser-vation suggests that the large-vessel endothelium may be anHCMV reservoir in vivo.
The mechanism by which HCMV induces a persistent non-cytopathic infection of AEC is unknown. HCMV-infected AECwere unable to accumulate infectious intracellular virus, al-though they produced normal amounts of extracellular virus.This phenomenon suggests that the cells survive infectionthrough efficient export of mature virus and toxic products,which may cause cellular lysis, from the cell. This mechanism isclearly different from that which operates during HCMV in-fection of monocyte-derived macrophages, which results in thenonlytic accumulation of large amounts of intracellular virusand the lack of extracellular virus (3). In monocyte-derivedmacrophages, virus was found to accumulate in large cytoplas-mic vacuoles that did not associate with the PM. These twoobservations emphasize the cell-specific differences that occurduring HCMV infection.
In addition to the extremely rapid export of virus from AEC,the inability of HCMV to block the cell cycle may be a way inwhich the virus and cell coexist. The mechanism by which thecell or virus or both circumvent the normal blocks in the cellcycle due to infection is unknown. Jault et al. (7) observed an
FIG. 4. Growth curves of HCMV Po, AD169, and RVAd69 in EC. (A) AECand BMVEC were infected with Po at an MOI of 3. HCMV Po-infected AEC orBMVEC cells or supernatants were harvested at the indicated times, and viraltiters were determined by a plaque assay on HF cells. (B) AEC were infectedwith AD169 or RVAd69 at an MOI of 3. Cells and supernatants were harvestedat the indicated times, and viral titers were determined by a plaque assay on HFcells. The data are the results of three experiments (both panels).
FIG. 3. Growth curves for HCMV in AEC and BMVEC. (A) EC wereinfected with HCMV (Towne) at an MOI of 3. AEC or BMVEC infected withTowne were harvested at the indicated times, and viral titers were determined bya plaque assay on HF cells. (B) Infected culture supernatants, from AEC orBMVEC, were collected at the indicated times, and viral titers were determinedby a plaque assay on HF cells. The data are the results of three experiments (bothpanels).
VOL. 72, 1998 HMCV PERSISTENTLY INFECTS MACROVASCULAR EC 5665
on June 3, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
overexpression of cyclin E and a delay in cyclin A accumulationin HCMV-infected cells. Cyclin A is required for DNA repli-cation and for the G2/M transition (5, 15). A proposed mech-anism by which HCMV affected cyclin A accumulation wasdependent upon viral and cellular DNA replication kinetics.Therefore, one might speculate that viral and cellular DNAreplication kinetics in HCMV-infected AEC are such that theviral cell cycle inhibitors are not present at the appropriatetimes during the cell cycle. As the complexities of how HCMVaffects the cell cycle are discovered, the mechanism by whichAEC elude a block in the cell cycle may be revealed.
Although the specific cellular organs HCMV targets duringacute disease have been known for almost 50 years, the site(s)for HCMV latency has been difficult to identify. We haverecently demonstrated that latent HCMV can be reactivated ina myeloid lineage cell obtained from the peripheral blood ofhealthy seropositive individuals (21). However, this observa-tion does not preclude the existence of other sites of HCMVlatency or persistence. Another likely cellular candidate forHCMV persistence is the EC. AEC interact naturally withmonocytes that are trafficking in the bloodstream and migrat-
ing into tissues. Therefore, a dynamic interaction between thecells in which virus is either activated or transmitted in tissuesmay occur during extravasation. These interactions may in-volve cell-cell contact through adhesion molecules, which sub-sequently triggers signal transduction events, activating virus inlatently infected monocytes or the resting endothelium. Themonocyte or AEC may function as a vehicle for cell-to-celltransmission of HCMV, which in turn could result in the re-activation of HCMV from macrophages or EC in the artery.One consequence of HCMV reactivation in the artery is thedissemination of virus throughout the body. HCMV would beeasily spread throughout the circulatory system once shed intothe lumen of the aorta because of the large quantities of bloodthat pass through this organ. The state of viremia may thenresult in the widespread infection of different organs. In addi-tion to causing the dissemination of free virus throughout thebody, circulating monocytes that become infected or that re-activate virus because of contact with AEC would serve as theideal vector for HCMV dissemination to other tissues. In sup-port of this hypothesis, an in vitro model has demonstratedthat infected EC can transmit HCMV to cocultured monocytes
FIG. 5. HCMV-infected AEC and BMVEC were infected with HCMV at an MOI of 3 and analyzed by EM at 14 dpi. Small amounts of cytoplasmic virus weredetectable in AEC when they were compared to BMVEC by EM (A & C; bar 5 2 mm). Similar amounts of nucleocapsids were detected in the nuclei of AEC andBMVEC by EM (B and D; bar 5 350 nm). The arrows in panels B and D show examples of mature nucleocapsids.
5666 FISH ET AL. J. VIROL.
on June 3, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

FIG. 6. HCMV infection of AEC does not result in the arrest of the cell cycle. AEC were infected at an MOI of 5 or were mock infected. The culture doublingfrequency was determined throughout a time course of infection (A). Nuclei from infected and mock-infected AEC were stained with propidium iodide alone or incombination with IE antibody and used for FACS analysis (B). AEC were infected with HCMV, and at 5 dpi double-label immunofluorescence was performed withantibodies directed against the IE72 antigen (C) and microtubulin (D). Magnification (C and D), 3407. The arrow in panel D points to spindle poles.
VOL. 72, 1998 HMCV PERSISTENTLY INFECTS MACROVASCULAR EC 5667
on June 3, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

and that these monocytes can retransmit virus to uninfectedEC (23).
This study clearly indicates the importance of EC in thebiology of HCMV. Elucidating the mechanisms of HCMVreplication and virus assembly in AEC will be essential forunderstanding viral persistence and trafficking in the humanhost. The current studies indicate that HCMV replication inAEC differs from that in other naturally infected host cells.These observations emphasize the importance of examiningviral replication in biologically relevant cell types.
ACKNOWLEDGMENTS
We thank Rebecca Ruhl for technical assistance and Ashlee Mosesfor helpful discussion.
This work was supported by a Public Health Service grant from theNational Institutes of Health (AI 21640) (J.A.N.), the Molecular He-matology Training program NIH NRSA Training Award (K.N.F.), andthe Knut and Alice Wallenbergs Foundation (C.S.-N.). C.S.-N. is ascholar of the Wenner-Gren Foundation, Sweden.
REFERENCES
1. Britt, W. J., and L. G. Vugler. 1992. Oligomerization of the human cytomeg-alovirus major envelope glycoprotein complex gB (gp55-116). J. Virol. 66:6747–6754.
2. Dittmer, D., and E. S. Mocarski. 1997. Human cytomegalovirus infectioninhibits G1/S transition. J. Virol. 71:1629–1634.
3. Fish, K. N., A. S. Depto, A. V. Moses, W. Britt, and J. A. Nelson. 1995.Growth kinetics of human cytomegalovirus are altered in monocyte-derivedmacrophages. J. Virol. 69:3737–3743.
4. Friedman, H. M., E. J. Macarek, R. R. MacGregor, J. Wolfe, and N. A.Kefalides. 1981. Virus infection of endothelial cells. J. Infect. Dis. 143:266–273.
5. Girard, F., U. Strausfed, A. Fernandez, and N. J. C. Lamb. 1991. Cyclin A isrequired for the onset of DNA replication in mammalian fibroblasts. Cell 67:1169–1179.
6. Gnann, J., Jr., J. Ahlmen, C. Svalander, L. Olding, M. B. Oldstone, and J. A.Nelson. 1988. Inflammatory cells in transplanted kidneys are infected byhuman cytomegalovirus. Am. J. Pathol. 132:239–248.
7. Jault, F. M., J. M. Jault, F. Ruchti, E. A. Fortunato, C. Clark, J. Corbeil,D. D. Richman, and D. H. Spector. 1995. Cytomegalovirus infection induceshigh levels of cyclins, phosphorylated Rb, and p53, leading to cell cyclearrest. J. Virol. 69:6697–6704.
8. Joo, F. 1992. The cerebral microvessels in culture, an update. J. Neurochem.58:1–17.
9. Jupp, R., S. Hoffmann, A. Depto, R. M. Stenberg, P. Ghazal, and J. A.Nelson. 1993. Direct interaction of the human cytomegalovirus IE86 protein
with the cis repression signal does not preclude TBP from binding to theTATA box. J. Virol. 67:5595–5604.
10. Lathey, J. L., C. A. Wiley, M. A. Verity, and J. A. Nelson. 1990. Culturedhuman brain capillary endothelial cells are permissive for infection by humancytomegalovirus. Virology 176:266–273.
11. Lu, M., and T. Shenk. 1996. Human cytomegalovirus infection inhibits cellcycle progression at multiple points, including the transition from G1 to S. J.Virol. 70:8850–8857.
12. Melnick, J. L., E. Adam, and M. E. DeBakey. 1995. Cytomegalovirus andatherosclerosis. Bioessays 17:899–903. (Review.)
13. Moses, A. V., and J. A. Nelson. 1994. HIV infection of human brain capillaryendothelial cells—implications for AIDS dementia. Adv. Neuroimmunol. 4:239–247. (Review.)
14. Myerson, D., R. C. Hackman, J. A. Nelson, D. C. Ward, and J. K. McDougall.1984. Widespread presence of histologically occult cytomegalovirus. Hum.Pathol. 15:430–439.
15. Pagano, M., P. Pepperkok, F. Verde, W. Ansorge, and G. Draetta. 1992.Cyclin A is required at two points in the human cell cycle. EMBO J. 11:961–971.
16. Page, C., M. Rose, M. Yacoub, and R. Pigott. 1992. Antigenic heterogeneityof vascular endothelium. Am. J. Pathol. 141:673–682.
17. Schmolke, S., H. F. Kern, P. Drescher, G. Jahn, and B. Plachter. 1995. Thedominant phosphoprotein pp65 (UL83) of human cytomegalovirus is dis-pensable for growth in cell culture. J. Virol. 69:5959–5968.
18. Simbulan-Rosenthal, C. M., D. S. Rosenthal, H. Hilz, R. Hickey, L. Malkas,N. Applegren, Y. Wu, G. Bers, and M. E. Smulson. 1996. The expression ofpoly(ADP-ribose) polymerase during differentiation-linked DNA replicationreveals that it is a component of the multiprotein DNA replication complex.Biochemistry 35:11622–11633.
19. Sinzger, C., A. Grefte, B. Plachter, A. S. Gouw, T. H. The, and G. Jahn. 1995.Fibroblasts, epithelial cells, endothelial cells and smooth muscle cells aremajor targets of human cytomegalovirus infection in lung and gastrointesti-nal tissues. J. Gen. Virol. 76:741–750.
20. Smiley, M. L., E.-C. Mar, and E.-S. Huang. 1988. Cytomegalovirus infectionand viral induced transformation of human endothelial cells. J. Med. Virol.25:213–226.
21. Soderberg-Naucler, C., K. N. Fish, and J. A. Nelson. 1997. Reactivation oflatent human cytomegalovirus by allogeneic stimulation of blood cells fromhealthy donors. Cell 91:119–126.
22. Turner, R. T., J. H. Beckstead, R. A. Warnke, and G. S. Wood. 1987.Endothelial cell phenotypic diversity—in situ demonstration of immunologicand enzymatic heterogeneity that correlates with specific morphologic sub-types. Am. J. Clin. Pathol. 87:569–575.
23. Waldman, W. J., D. A. Knight, E. H. Huang, and D. D. Sedmak. 1995.Bidirectional transmission of infectious cytomegalovirus between monocytesand vascular endothelial cells: an in vitro model. J. Infect. Dis. 171:263–272.
24. Wiley, C. A., R. D. Schrier, F. J. Denaro, J. A. Nelson, P. W. Lampert, andM. B. Oldstone. 1986. Localization of cytomegalovirus proteins and genomeduring fulminant central nervous system infection in an AIDS patient.J. Neuropathol. Exp. Neurol. 45:127–139.
5668 FISH ET AL. J. VIROL.
on June 3, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from