hyaluronan induces monocyte chemoattractant protein...
TRANSCRIPT

J Am Soc Nephrol 9: 2283-2290, 1998
Hyaluronan Induces Monocyte Chemoattractant Protein-i
Expression in Renal Tubular Epithelial Cells
BEATRICE BECK�SCHIMMER,*t BEAT OERTLI,*� THOMAS PASCH,t and
RUDOLF P. WUTHRICH*�*physiological Institute, University of Zllrich-lrchel; �Department ofAnesthesiology and �Division of
Nephrology, Department of Medicine, University Hospital, Zurich, Switzerland.
Abstract. Hyaluronan (HA) is a nonsulfated glycosaminogly-
can that accumulates in the renal interstitium in immune-
mediated kidney diseases. The functional significance of such
HA deposition in the kidney has not been elucidated. Several
studies have suggested that HA may exhibit proinflammatory
effects. Since chemokines such as monocyte chemoattractant
protein- 1 (MCP- 1) play an important robe in the recruitment of
beukocytes in renal injury, this study tested whether HA and its
fragments could promote MCP- 1 production by renal paren-
chymal cells. Mouse cortical tubular cells were stimulated with
fragmented HA or with high molecular weight HA (Heabon) in
vitro and were examined for MCP-l expression. Fragmented
HA, but not Heabon, increased MCP-l mRNA within 30 mm
with a peak after 2 h. In addition, a 10-fold increase of MCP- 1
protein in the supernatant was found after a 6-h stimulation
with fragmented HA. The enhanced MCP-l mRNA and pro-
tein expression in response to HA was dose-dependent between
1 and 100 �sg/mb. Upregubation of MCP-l protein production
could be blocked by preincubation with actinomycin D or
cycboheximide, suggesting that MCP-l mRNA and protein
expression in response to HA are based on de novo synthesis.
The HA-stimulated MCP-l production was also inhibited with
anti-CD44 antibodies, suggesting that MCP-l is upregubated at
least in part by signaling through CD44. In summary, frag-
mented HA markedly stimulates renal tubular MCP-l produc-
tion by mechanisms that involve binding to the HA receptor
CD44. It is hypothesized that the accumulation of HA in
immune renal injury could participate in the recruitment and
activation of inflammatory cells in vivo through production of
MCP-1.
The synthesis and the degradation of components of the extra-
cellular matrix (ECM) represent important steps in chronic
inflammation. One of the constituents of the altered ECM in
inflammation is hyaluronan (HA), a nonsulfated gbycosamino-
glycan that is composed of repeating units of (f31-4)-o-glucu-
ronic acid-(�3b-3)-N-acetyl-D-glucosamine (1). Under normal
conditions, HA is synthesized as a high molecular weight
extracellubar pobysaccharide (2). In inflammatory conditions,
high molecular weight HA may undergo degradation into low
molecular weight components, presumably under the influence
of oxygen free radicals or inflammation-specific hyaluroni-
dases (3-5).
High and low molecular weight forms of hyaburonan exert
different biological effects on tissues and cells. In macro-
phages, for example, which express the main HA receptor
CD44 abundantly, high molecular weight HA has no prom-
flammatony action, but it acts as an inhibitor of phagocytosis
(6), whereas low molecular weight HA induces proinflamma-
tory genes, growth factors, and nitric oxide synthase (7-10).
Depending on the balance of synthesis and degradation, HA
Received February 24, 1998. Accepted June 1, 1998.Correspondence to Dr. Rudolf P. WUthnich, Division of Nephrobogy, Univer-
sity Hospital. Rhmistrasse 100. 8091 ZUrich, Switzerland.
1046-6673/0901 2-2283$03.00/0
Journal of the American Society of Nephrology
Copyright 0 1998 by the American Society of Nephrology
might therefore play an important role in macrophage activa-
tion in vivo.
Very little HA is present in the normal kidney. In inflam-
matory conditions, however, HA markedly accumulates in the
kidney cortex, particularly in crescentic gbomerubonephritis
( 1 1 ), tububointerstitial injury ( I 2, 1 3), and albograft rejection
(14,15). In immune renal injury, HA accumulation is most
prominent at sites where the HA receptor CD44 is also in-
duced, e.g. , in glomerubar crescents and particularly in the
peritubular interstitium adjacent to injured proximal tubular
epithebial cells (13).
The functional significance of HA accumulation in the kid-
ney cortex has not been elucidated. Given its proinflammatory
effects, we were interested in examining the effects of HA and
its fragments on the expression of chemokine genes in the
kidney. In a previous study, we have shown that fragments of
HA can upregulate the expression of the adhesion molecules
intercellular adhesion molecule- 1 (ICAM- 1 ) and vascular cell
adhesion molecule-b (VCAM-b) in cultured kidney tubular
epithebiab cells ( 16). These studies have suggested indirectly
that HA accumulation in immune-mediated renal injury could
play a robe in beukocyte recruitment and adhesion.
Chemokines represent a family of structurally rebated che-
motactic cytokines produced by a variety of cells, including
kidney proximal tubular cells (17-19). Monocyte chemoattrac-
tant protein- 1 (MCP-l) is a particularly important chemokine
that is ovenexpressed in many immune-mediated renal diseases
and also in abbograft rejection (1 7). It has been suggested that

2284 Journal of the American Society of Nephrobogy J Am Soc Nephrol 9: 2283-2290, 1998
MCP-l may play a key role in the perpetuation of tububoin-
terstitial inflammation by attracting mainly monocytes and T
cells (20-22). Since HA fragments are known to induce prom-
flammatory genes, we have now examined whether HA in-
duces MCP- 1 upregubation in kidney tubular epithelial cells. In
this study, we demonstrate that fragmented HA is a potent
stimulatory factor for MCP- 1 expression by cultured tubular
cells. The HA-induced stimulation of MCP- 1 is mediated at
least in part by interaction with the CD44 molecule. Therefore,
HA-induced signaling through CD44 and subsequent MCP-l
gene transcription could be an important event in the genera-
tion and maintenance of a chemoattractant gradient in immune
renal injury.
Materials and MethodsChemicals and Reagents
Actinomycin D (ACTD), cycboheximide (CHX), and lipopoly-
sacchanide (LPS) were purchased from Sigma Chemical Co. (St.
Louis, MO). Fragmented HA derived from human umbilical cord
(molecular weight approximately 0.08 to 0.8 X 106 daltons) was
purchased from Fluka (Buchs, Switzerland). The HA samples were
checked for significant LPS contamination with a Limulus ame-
bocyte lysate assay (COATEST#{174} Endotoxin kit, Endosafe, Inc.,
Charleston, SC). LPS concentrations of all HA preparations were<20 pg/mb at the maximum concentration of HA used to stimulate
cells, a concentration that did not stimulate MCP-l per se. High
molecular weight HA (Heabon#{174}, >106 Da) was a generous gift
from Phanmacia (Uppsala, Sweden).
Cell Lines and Cell Culture
The simian virus 40-transformed mouse cortical tubular (MCT) cell
line was grown to confluence in Dulbecco’s modified Eagle’s medium
with Glutamax-l (DMEM) containing 10% fetal bovine serum (FBS),
10 mM Hepes, 100 U/mb penicillin, and 100 �.rg/ml streptomycin (all
from Life Technologies, Gaithensburg, MD) at 37#{176}Cwith 5% CO,
(23). The medium was then changed to DMEM containing 1% FBS
for 18 h to rest the cells. Cells were subsequently stimulated with HA
fragments or Heabon at different concentrations in the presence or
absence of inhibitors. HA fragments and Heabon were not toxic at
concentrations up to I mg/mb as assessed by morphology and trypan
blue exclusion. The KM8 1 hybridoma producing the rat anti-munine
CD44 IgG monocbonal antibody (mAb) was obtained from American
Type Culture Collection (Rockville, MD) (24). The hybridoma was
grown in RPMI containing 20% FBS. The control IgG mAb FF-l0targeting the Thy-l molecule has been described (25). Purified mAb
were generated from hybnidoma supernatants using affinity chnoma-tography with protein 0-Sepharose (Pharmacia).
RNA Extraction and Northern Blot Analysis
Total RNA was isolated from MCT cells using the Tnizob#{174}reagent
(Life Technologies) and phenol/chloroform extraction as described
previously (26,27). For Northern blot analysis, b0-p�g abiquots of total
RNA were size-fractionated by electrophoresis on 1 % aganose-form-
abdehyde gels and transferred to Zeta-Probe#{174} blotting membranes
(Bio-Rad, Hercules, CA). Methylene blue staining to detect the 18Sand 285 ribosomal RNA (rRNA) bands was performed to ensure
equal loading. After prehybridization overnight at 65#{176}Cin a solution
containing 1 mM ethybenediaminetetra-acetic acid, 7% sodium dode-
cyl sulfate in 0.5 M Na,HPO4, pH 7.2, the blots were hybridized
overnight with an MCP-b eDNA fragment previously labeled with
32P(dCTP). The MCP-l cDNA probe was generated by reverse tran-scniption-PCR, using the following primers: 5’-CCC AAT GAG TAO
OCT OGA GA-3’ (forward primer), 5’-AAA ATO GAT CCA CAC
CTT GC-3’ (reverse primer), yielding a 435-bp MCP-l eDNA fnag-ment (28). Blots were washed at 65#{176}Cin 40 mM sodium phosphate
buffer with decreasing concentrations of sodium dodecyb sulfate.
Autoradiography was performed using X-OMAT AR film (Kodak,
Rochester, NY). Blots were rehybnidized with an 800-bp eDNA probe
for the housekeeping gene �3-actin.
Enzyme-Linked Immunosorbent Assay for MCP-1
Protein
MCP- 1 protein production was examined in the supernatant ofstimulated MCT cells, using a standard sandwich enzyme-linked
immunosorbent assay (ELISA) technique as described elsewhere (29).
The capturing antibody was a purified hamster anti-mouse MCP-l
IgG mAb (clone 2H5) and the detecting antibody was a biotinylated
hamster anti-mouse MCP-l IgO mAb (clone 4E2/MCP), both pun-
chased from Phanmingen (San Diego, CA). Recombinant mouse
MCP- 1 was used as a standard (Pharmingen).Antibodies, standards, and samples were diluted in phosphate-
buffered saline (PBS) containing 10% FBS and 0.05% Tween. All
washing steps were performed 3 times with 200 �tl, using PBS
containing 0.05% Tween. Then, 96-well plates (Costar) were coated
overnight with purified 2H5 anti-mouse-MCP- 1 mAb (2 p.g/ml, 50
�rl/well) in 0. 1 M Na2HPO4 binding buffer, pH 9.0. After a blocking
step with PBS/l0% FBS for 20 mm at 37#{176}C,standards and samples
(100 j.d per well) were incubated for I h at 37#{176}C,followed by the
incubation with the biotinylated 4E2/MCP hamster anti-mouse-
MCP-1 mAb (I j�g/ml, 100 pJ per well) for 45 mm at 37#{176}C.Strepta-vidin-conjugated horseradish penoxidase (Southern Biotechnology
Associates, Birmingham, AL) at a dilution of I :6000 was then added
to the plate. After washing, the plates were developed with 200 �rl of
an o-phenylenediamine dihydrochlonide solution (Sigma). The reac-
tion was stopped by adding 50 jrl of 95% sulfuric acid to each well.
Optical density was determined with an ELISA reader at 492 nm.
Statistical Analyses
All samples were run in triplicate, and the results (mean ± SEM)were calculated. Experiments were performed at beast three times, and
results from typical experiments are shown in the figures.
ResultsTime- and Dose-Dependent Upregulation of MCP-1
mRNA by HA
The effect of HA on MCP- 1 gene expression by MCT cells
was studied by Northern blot analysis. MCT cells were slim-
ulated with fragmented HA for various time periods, and total
RNA was extracted. Figure 1A shows that the steady-state
transcript levels for MCP- 1 increased markedly in response to
short-term stimulation with HA compared with unstimulated
cells that express MCP-l constitutively. MCP-l mRNA levels
peaked between 1 .5 and 3 h. Although MCP- I mRNA levels
also increased slightly in unstimulated MCT cells when exam-
med for up to 72 h, the HA-stimulated cells always displayed
higher steady-state mRNA bevels for MCP-1 when compared
with the corresponding control (Figure 1B).
To examine the effect of increasing HA doses on MCP- 1
mRNA expression, MCT cells were stimulated with HA at

f3-actin 2.2 kb
time(h) 0 1 1.5 2 3 6A
I control ,� HA
MCP-1
methylene blue
time (h) 24 48 72 24 48 72B lh 1.5h 2h 3h 6h
timeFigure 1. Steady-state mRNA bevels for monocyte chemoattractant
protein- 1 (MCP-b) are time dependently upregubated by fragmented
hyaburonan (HA) when compared with �-actin. Mouse cortical tubular
(MCT) cells were stimulated with HA (100 p.g/mb) for 0 to 6 h.Specific MCP-l mRNA is at a maximum level between 1.5 and 3 h
(A). Extending the stimulation oven 72 h demonstrates upregulation ofMCP-l mRNA at all time points oven control, with only a slight
increase in control cells (B). Methylene blue reveals undegraded total
RNA.
0
C00 HA
50
40
30
20
10
0
A
100
90
80
70‘I)
.� 60
(00
x
� A0
30
20
10
0
B
Figure 3. The MCP-l protein is time dependently upregulated in
response to HA. Supernatants from HA-stimulated (100 pg/mb) MCT
cells were collected, and MCP- 1 protein determination was performed
by enzyme-linked immunosonbent assay (ELISA). The amount of
MCP-l above control in the supernatants of stimulated cells at dif-fenent time points increased steadily within 6 h (A). Prolonged incu-
bation with HA (100 �g/ml) oven 72 h reveals that the rate of MCP-b
production is greatest in the first 6 h. The concentration of MCP-l is
always greater in HA-stimulated plates compared with unstimulated
plates. *p < 001 (comparisons made within each time point) (B). The
figure represents one of three similar experiments.
MCP-1 0.8 kb
0 0.1 1_�_10 100 1000 dose �.tg/ml
�-actin
Oh 6h 24h 48h 72h
time
112.2
Figure 2. MCP-l mRNA is dose dependently upregubated by HA.MCT cells were stimulated with various HA concentrations (0. 1 to1000 p�g/mb for 2 h), and RNA was extracted and analyzed by
Northern blotting for MCP-1 and (3-actin. Transcript levels for MCP-b
increase from 0.1 to 100 j.tg/ml, with a peak at 100 �.rg/ml. With an
HA concentration of 1000 p�g/ml, the mRNA bevel is decreased again.
J Am Soc Nephrol 9: 2283-2290, 1998 Hyaluronan Upregulates MCP-l 2285
concentrations between 0. 1 and 1000 j.tg/mb. Figure 2 shows
tflill M#{128}T-I mRNAupr�u1�tion wii� � �t’t’�’�n1 and 100 j.tg/mb, peaking at 100 �tg/ml. At 1000 pjg/mb, the
MCP- 1 transcript bevels were slightly lower compared with
100 j.sg/ml HA.
MCP-1
0C..
0Or
HA
0.8 kb
Time- and Dose-Dependent Upregulation of MCP-1
f3r�t(n(?yttuiWe then examined MCP-l protein synthesis in response to
HA by measuring MCP- 1 concentrations in the supernatant of
HA-stimulated MCT cells with a sensitive and specific ELISA.
Figure 3A represents a time course over 6 h, showing a marked
increase of MCP-l protein within 6 h. Compared with the
mRNA bevels for MCP- 1 , the MCP- 1 protein concentrations
increased with a 2- to 3-h delay, suggesting that the de novo
transcription precedes MCP-l mRNA translation. Stimulation
with HA over 72 h showed that MCP- 1 levels increased the
most within the first 6 h, with very little further increase after
U)
a,
U
0.8kb ‘�‘C
40
30
U)
U
‘Do 20‘C
10
0
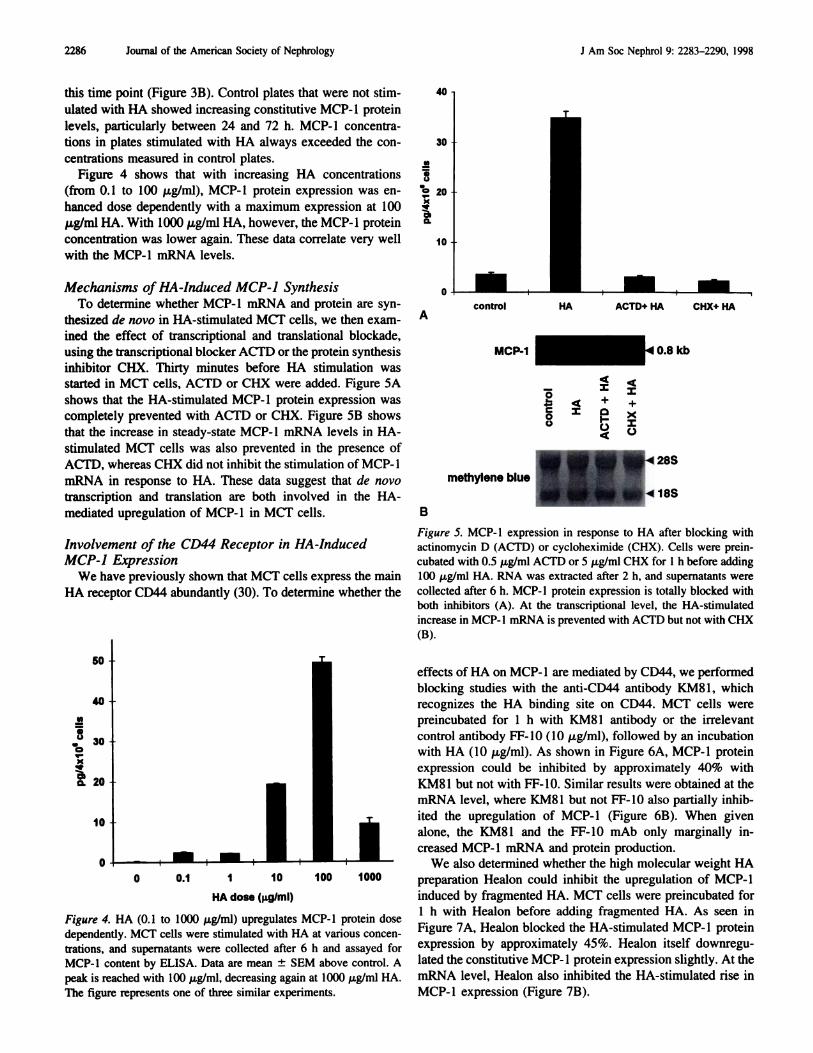
Acontrol HA ACTD+ HA CHX+ HA
MCP-1
methylene blue
4- x
0� 4 +� I a
0C)4
4I+
IC.)
28S
18S
50
40
30
20
Cl)
4)U
0‘C
a)0.
10
0
2286 Journal of the American Society of Nephrology J Am Soc Nephrol 9: 2283-2290, 1998
this time point (Figure 3B). Control plates that were not stim-
ulated with HA showed increasing constitutive MCP- 1 protein
bevels, particularly between 24 and 72 h. MCP- 1 concentra-
tions in plates stimulated with HA always exceeded the con-
centrations measured in control plates.
Figure 4 shows that with increasing HA concentrations
(from 0. 1 to 100 p�gImb), MCP- 1 protein expression was en-
hanced dose dependently with a maximum expression at 100
pg/mb HA. With 1000 �g/ml HA, however, the MCP- 1 protein
concentration was bower again. These data correlate very well
with the MCP-l mRNA levels.
Mechanisms of HA-Induced MCP-1 Synthesis
To determine whether MCP-l mRNA and protein are syn-
thesized de novo in HA-stimulated MCT cells, we then exam-
med the effect of transcriptional and translational blockade,
using the transcriptional blocker ACTD or the protein synthesis
inhibitor CHX. Thirty minutes before HA stimulation was
started in MCT cells, ACTD or CHX were added. Figure SA
shows that the HA-stimulated MCP-b protein expression was
completely prevented with ACTD or CHX. Figure SB shows
that the increase in steady-state MCP- 1 mRNA levels in HA-
stimulated MCT cells was also prevented in the presence of
ACTD, whereas CHX did not inhibit the stimulation of MCP-l
mRNA in response to HA. These data suggest that de novo
transcription and translation are both involved in the HA-
mediated upregubation of MCP-l in MCT cells.
involvement of the CD44 Receptor in HA-inducedMCP-1 Expression
We have previously shown that MCT cells express the main
HA receptor CD44 abundantly (30). To determine whether the
0 0.1 1 10
HA dose (jsglml)
Figure 4. HA (0.1 to 1000 �tg/ml) upregulates MCP-b protein dose
dependently. MCT cells were stimulated with HA at various concen-
trations, and supernatants were collected after 6 h and assayed for
MCP- 1 content by ELISA. Data are mean ± SEM above control. A
peak is reached with bOO j.tg/ml, decreasing again at 1000 jrg/ml HA.
The figure represents one of three similar experiments.
B
Eu0.8 kb
Figure 5. MCP-l expression in response to HA after blocking withactinomycin D (ACTD) or cycloheximide (CHX). Cells were prein-
cubated with 0.5 pg/mb ACTD or 5 jtg/mb CHX for 1 h before adding
100 �tg/ml HA. RNA was extracted after 2 h, and supennatants were
collected after 6 h. MCP-l protein expression is totally blocked with
both inhibitors (A). At the transcriptional level, the HA-stimulated
increase in MCP-l mRNA is prevented with ACTD but not with CHX
(B).
effects of HA on MCP- 1 are mediated by CD44, we performed
blocking studies with the anti-CD44 antibody KM81, which
recognizes the HA binding site on CD44. MCT cells were
preincubated for 1 h with KM8 1 antibody on the irrelevant
control antibody FF- 10 (10 j�g/ml), followed by an incubation
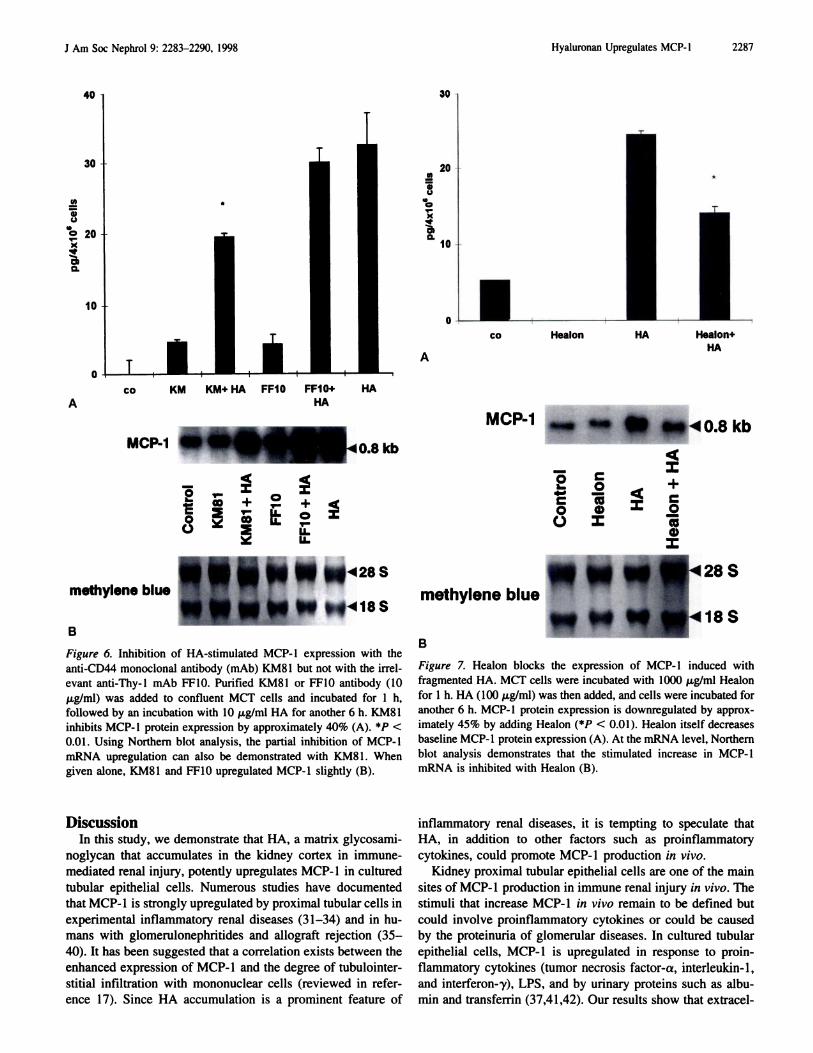
with HA (10 .tg/ml). As shown in Figure 6A, MCP-l protein
expression could be inhibited by approximately 40% with
KM8 1 but not with FF- 10. Similar results were obtained at the
mRNA bevel, where KM81 but not FF-l0 also partially inhib-
ited the upregubation of MCP-l (Figure 6B). When given
alone, the KM81 and the FF-b0 mAb only marginally in-
creased MCP-l mRNA and protein production.
We also determined whether the high molecular weight HA100 1000 preparation Heabon could inhibit the upregulation of MCP-b
induced by fragmented HA. MCT cells were preincubated for
1 h with Heabon before adding fragmented HA. As seen in
Figure 7A, Heabon blocked the HA-stimulated MCP- 1 protein
expression by approximately 45%. Heabon itself downregu-
bated the constitutive MCP- 1 protein expression slightly. At the
mRNA bevel, Heabon also inhibited the HA-stimulated rise in
MCP-l expression (Figure 7B).
40
30
30
10
0
A
0
Co KM KM+ HA FF10 FFIO+ HAHA
A
MCP1
Co Healon
MCP-1
0.8 kb
HA
HA
0.8 kb
<
<
�
���u-L1-
�
<x
0‘-
LI.
Z
.�
��..
0C)
C
.i0Q)
I
<�
+
0#{149}�
.I
I Am Soc Nephrol 9: 2283-2290. 1998 Hyaluronan Upregulates MCP- I 2287
Cl)
4)U
IDo 20
‘C
C)
0.
methylene blue
B
28 S
185
Figure 6. Inhibition of HA-stimulated MCP-b expression with the
anti-CD44 monocbonal antibody (mAb) KM8I but not with the irreb-
evant anti-Thy-l mAb FF10. Purified KM81 or FF10 antibody (10
p�g/mb) was added to confluent MCT cells and incubated for I h,
followed by an incubation with 10 p.g/mb HA for another 6 h. KM81
inhibits MCP-l protein expression by approximately 40% (A). *P <
0.01. Using Northern blot analysis, the partial inhibition of MCP-l
mRNA upregulation can also be demonstrated with KM8 1 . When
given alone, KM8I and FF10 upregulated MCP-l slightly (B).
DiscussionIn this study, we demonstrate that HA, a matrix glycosami-
nogbycan that accumulates in the kidney cortex in immune-
mediated renal injury, potently upregulates MCP- 1 in cultured
tubular epitheliab cells. Numerous studies have documented
that MCP-l is strongly upregubated by proximal tubular cells in
experimental inflammatory renal diseases (31-34) and in hu-
mans with gbomerubonephritides and albognaft rejection (35-
40). It has been suggested that a correlation exists between the
enhanced expression of MCP- 1 and the degree of tububointen-
stitial infiltration with mononuclear cells (reviewed in refer-
ence 17). Since HA accumulation is a prominent feature of
20Cl)
4,U
0
‘C
C)0. io
methylene blue
B
28S
18S
Figure 7. Heabon blocks the expression of MCP-l induced withfragmented HA. MCT cells were incubated with 1000 p.g/ml Healon
for 1 h. HA (100 j.�g/mb) was then added. and cells were incubated for
another 6 h. MCP-l protein expression is downregulated by approx-
imately 45% by adding Heabon (*� < 0.01 ). Healon itself decreases
baseline MCP-l protein expression (A). At the mRNA level, Northern
blot analysis demonstrates that the stimulated increase in MCP-l
mRNA is inhibited with Healon (B).
inflammatory renal diseases, it is tempting to speculate that
HA, in addition to other factors such as proinfiammatory
cytokines, could promote MCP- I production in vivo.
Kidney proximal tubular epithebiab cells are one of the main
sites of MCP- 1 production in immune renal injury in vivo. The
stimuli that increase MCP- 1 in vivo remain to be defined but
could involve proinflammatony cytokines on could be caused
by the proteinunia of gbomerubar diseases. In cultured tubular
epithelial cells, MCP-l is upnegubated in response to prom-
flammatory cytokines (tumor necrosis factor-a, interbeukin- 1,
and interferon-y), LPS, and by urinary proteins such as albu-
mm and transfenrin (37,41,42). Our results show that extracel-

2288 Journal of the American Society of Nephrology J Am Soc Nephrol 9: 2283-2290. 1998
bubar matrix components such as HA have to be included in the
list of factors capable of promoting MCP- 1 production in
kidney tubular cells. Our data are in agreement with experi-
mental research in macrophages demonstrating that HA pro-
motes the synthesis of cytokines, chemokines, and growth
factors in these cells (7,8,43,44).
The effects of HA on MCP- 1 expression were dose-depen-
dent between 1 and 100 �g/ml. These concentrations could be
pathophysiobogicabby relevant, as the renal cortical accumula-
tion of HA in inflammatory renal diseases can be impressive.
In rejecting rat renal abbografts, for example, 300 to 400 p�g
HA/g dry weight can be detected in the kidney (45), and
concentrations > 1 jtg/mb can be found in the serum or in the
peritoneab diabysate of patients with end-stage renal disease
(46,47). There is no obvious explanation why higher concen-
trations of HA (up to 1 mg/mb) stimulated MCP-l productionless than bower doses (100 p�g/ml). Possible mechanisms could
include less efficient interaction with cell surface receptors for
HA or a downregulating effect of high HA doses on HAreceptor-mediated signaling events.
Earlier work has suggested that high molecular weight HA
inhibits inflammatory responses (6), and it has only recently
been recognized that fragments of HA display different effects
which are proinflammatory (48). In cultured MCT cells,
MCP-b was only induced with fragmented HA but not with
high molecular weight HA (Healon). In fact, high molecular
weight HA inhibited the effect of low molecular weight HA,
suggesting that the HA fragments were competed away with
high molecular weight preparations of HA. This is further
evidence of specificity of the HA-mediated effect on MCP-l
expression.
Our studies also demonstrate that the HA receptor CD44 is
involved in MCP- 1 production by cultured tubular cells, be-
cause anti-CD44 mAb inhibited the stimulation with HA. It
was not possible, however, to block the HA-stimulated MCP- 1
production completely. This could be due to several reasons.
First, relatively high concentrations of HA had to be used to
stimulate MCP- I expression in MCT cells; to compete away a
barge amount of HA, one needs to use also large amounts of
antibody. It is possible that the high antibody concentration
could have a partial agonist activity that negates the blocking
effect. Similar considerations were made previously by other
groups (43,44). Second, it may be intrinsically problematic to
block the binding of disaccharide moieties (HA) to protein
(CD44) with the use of protein (anti-CD44 antibodies). Third,
we cannot exclude that additional HA receptors could be
involved in the HA-stimulated generation of MCP-l by tubular
epitheliab cells. CD44 is very abundant on MCT cells, but
additional, less well characterized HA binding proteins such as
RHAMM could also be present (49,50). Additional studies will
be required to examine in detail whether HA promotes MCP-l
production solely via CD44 or whether additional HA recep-
tons are also involved.
Our in vitro study raises important new questions regarding
the interaction of matrix components with kidney tubular epi-
thebial cells in interstitial renal diseases. In a normal situation,
the tubular basement membrane separates the proximal tubules
from the penitubular interstitial space, shielding anatomically
the proximal tubule. The situation is different, however, in
renal interstitial inflammation, where the tubular basement
membrane can be disrupted, as, for example, in albograft re-
jection. In such an inflammatory situation, HA accumulates in
the renal cortex, and it could interact with the HA receptor
CD44 on tubular cells. This interaction, and perhaps the bind-
ing of additional matrix breakdown products, could then trig-
ger proximal tubules to express proinflammatory genes such as
chemokines, which participate in the recruitment of leukocytes.
HA needs to bind to its receptors before it can produce
proinflammatory effects in proximal tubular cells. It is known
that the main HA receptor CD44 must be in an activated state
to bind HA (S I). In a previous study, we have shown that the
CD44 molecule is in this activated state in MCT cells, binding
the HA molecule abundantly (30). Not all cells that express
CD44 are capable of binding HA constitutively, however. T
cell lines, for example, are strongly CD44-positive but they
vary markedly in their capacity to bind HA, and certain lines
do not bind HA at all (52). Previously, we have shown that
MCT cells, but not primary cultures of mouse tubular epithebiab
cells, can be stimulated with HA fragments to express adhesion
molecules, presumably because of limited HA-binding by
CD44 in primary cultures (16). Primary cultures were also only
marginally responsive to HA in stimulating MCP-l (unpub-
lished observation). It is not excluded that CD44-positive prox-
imal tubular cells are in an active binding state in vivo in
inflammatory renal diseases, but this needs to be examined
further.
In summary, we have shown that the ECM component HA
markedly stimulates the expression of the chemokine MCP- 1
by cultured kidney tubular epithelial cells. HA needs to be in
fragmented form to promote MCP- 1 production since a high
molecular weight HA preparation did not stimulate MCP-l.
The stimulatory effect of fragmented HA is transduced at least
in part by the CD44 cell surface receptor on tubular epithebiab
cells. De novo transcription and translation are involved in the
HA-mediated upregubation of MCP- 1 production. We specu-
late that fragmented HA, in concert with other more short-lived
stimuli such as proinflammatory cytokines, could produce sus-
tamed expression of MCP-l and therefore contribute to the
activation and recruitment of mononuclear cells in the kidney
in immune renal injury.
AcknowledgmentsThis study was supported by the Swiss National Science Founda-
tion (Grants 32-40390.94 and 32-5072 1 .97 to Dr. WUthnich), the
Olga-Mayenfisch Foundation, the Hartmann-MUller Foundation, and
the Research Foundation of the University of ZUrich. Dr. Beck-
Schimmer is the recipient of a grant by the Federal Foundation for the
Advancement of Young Scientists and Scholars. Dr. Oentli is the
recipient of a Postgraduate Fellowship from the University of Zurich
and is supported by the Swiss National Science Foundation and the
Maurice E. Muller Foundation. Dr. W0thnich is the recipient of a
Physician Scientist Award (Grant 32-38821.93) from the Swiss Na-
tional Science Foundation. We thank C. Gasser for the illustrations.

J Am Soc Nephrol 9: 2283-2290, 1998 Hyaluronan Upregulates MCP-1 2289
References1 . Laurent TC, Fraser IRE: Hyaluronan. FASEB J 6: 2397-2404,
1992
2. Prehm P: Hyaluronate is synthesized at plasma membranes.
Biochem J 220: 597-600, 1984
3. Saani H: Oxygen derived free radicals and synovial fluid hyal-
uronate. Ann Rheum Dis 50: 389-392, 1991
4. Schenck P, Schneider 5, Miehlke R, Prehm P: Synthesis and
degradation of hyaluronate by synovia from patients with rheu-
matoid arthritis. J Rheu,natol 22: 400-405, 1995
5. Kreil 0: Hyaluronidases: A group of neglected enzymes. Protein
Sci 4: 1666-1669, 1995
6. Forrester IV, Babazs EA: Inhibition of phagocytosis by high
molecular weight hyalunonate. mmniuizolog�’ 40: 435-446, 1980
7. Noble PW, Lake FR, Henson PM, Riches DW: Hyabunonate
activation of CD44 induces insulin-bike growth factor-b expres-
sion by a tumor necrosis factor-a-dependent mechanism in mu-
nine macrophages. J Cliii mns’est 91 : 2368-2377, 1993
8. Lake FR, Noble PW, Henson PM, Riches DW: Functionalswitching of macnophage responses to tumor necrosis factor-a
(TNF-a) by interfenons: Implications for the pbeiotropic activities
of TNF-a. J C/in Ins’est 93: 1661-1669, 1994
9. McKee CM, Lowenstein CI, Horton MR. Wu J, Bao C, Chin BY,
Choi AMK, Noble PW: Hyaluronan fragments induce nitric-
oxide synthase in murine macrophages through a nuclear factor
RB-dependent mechanism. J Biol Chein 272: 8013-8018, 1997
10. Rockey DC, Chung II, McKee CM, Noble PW: Stimulation of
inducible nitric oxide synthase in rat liven by hyaburonan frag-
ments. Hepatology 27: 86-92, 1998
1 1. Nishikawa K, Andres 0, Bhan AK, McCluskey RT, Collins AB,
Stow IL, Stamenkovic I: Hyaburonate is a component of cres-cents in rat autoimmune gbomerubonephnitis. Lob m�ivest 68: 146-
153, 1993
12. Wu I, Fan 0, Kitazawa K, Sugisaki T: The relationship of
adhesion molecules and leukocyte infiltration in chronic tububo-
interstitial nephnitis induced by puromycin aminonucbeoside in
Wistar rats. C/in hn,nunol mtnmiinopathol 79: 229-235, 1996
13. Sibalic V, Fan X, Loffing I, Wuthrich RP: Upregulated renal
tubular CD44, hyaluronan and osteopontin in kdkd mice with
interstitial nephnitis. Nephrol Dial Transplant 12: 1344-1353,
1997
14. Wells AF, Lansson E, Tengblad A, Fellstrom B, Tufveson 0,
Kbareskog L, Laurent TC: The localization of hyalunonan in
normal and rejected human kidneys. Transplantation 50: 240-
243, 1990
15. Wells A, Larsson E, Hanas E, Laurent T, Hhllgnen R, Tufveson
0: Increased hyaluronan in acutely rejecting human kidney
grafts. Transplantation 55: 1346-1349, 1993
16. Oentli B, Beck-Schimmer B, Fan X, Wuthrich RP: Mechanisms
of hyaluronan-induced upregulation of ICAM-l and VCAM-l
expression by murine kidney tubular epithelial cells: Hyaluronan
triggers cell adhesion molecule expression through a mechanism
involving activation of nuclear facton-[K]B and activating pro-
tein-!. J mmnzunol 161: 3431-3437, 1998
17. Schl#{246}ndorffD, Nelson P1, Luckow B, Banas B: Chemokines andrenal disease. Kidney mnt 51: 610-621, 1997
18. Adams DH, Lloyd AR: Chemokines: Leucocyte recruitment and
activation cytokines. Loncet 349: 490-495, 1997
19. Luster AD: Chemokines: Chemotactic cytokines that mediate
inflammation. N Engi J Med 338: 436-445, 1998
20. Eddy AA: Experimental insights into the tububointerstitial dis-
ease accompanying primary gbomerulan lesions. J Am Soc Neph-
rol 5: 1273-1287, 1994
2 1 . Schl#{246}ndorff D: The robe of chemokines in the initiation andprogression of renal disease. Kidney mnt Suppi 49: 544-547,
1995
22. Banas B, Wenzeb U, Stahl RA, Schl#{246}ndorff D: Role of chemo-
kines in glomerular diseases. Kidney Blood Press Res 19: 270-
280, 1996
23. Haverty TP, Kelly CI, Hines WH, Amenta PS, Watanabe M,Harper RA, Kefalides NA, Neilson EG: Characterization of a
renal tubular epitheliab cell line which secretes the autobogous
target antigen of autoimmune experimental interstitial nephnitis.
J Cell Biol 107: 1359-1368, 1988
24. Miyake K, Medina KL, Hayashi 5, Ono 5, Hamaoka T, Kincade
PW: Monocbonal antibodies to Pgp-l/CD44 block lympho-he-mopoiesis in long-term bone marrow cultures. J Exp Med I 7 1:
477-488, 1990
25. Fan X, Wuthrich RP: ICAM- I -, -2- and LFA- 1-independent
homotypic T cell aggregation induced by a novel activating
monoclonal antibody targeting the murine Thy-I molecule. mn-flammation 20: 401-41 1, 1996
26. Chomczynski P, Sacchi N: Single-step method of RNA isolation
by acid guanidinium thiocyanate-phenol-chborofonm extraction.
AnalBiochem 162: 156-159, 1987
27. Benz PS, Fan XH, W0thnich RP: Enhanced tubular epithelial
CD44 expression in MRL-lpr lupus nephnitis. Kidney mnt 50:
156-163, 1996
28. Rollins BI, Morrison ED, Stiles CD: Cloning and expression of
JE, a gene inducible by platelet-derived growth factor and whose
product has cytokine-like properties. Proc Nail Acad Sci USA 85:
3738-3742, 1988
29. Beck-Schimmen B, Schimmen RC, Warner RL, Schmal H, Nor-
dbbom 0, Fbory CM, Lesch ME, Friedb HP, Schnien Di, Ward PA:
Expression of lung vascular and airway ICAM- 1 after exposure
to bacterial bipopolysaccharide. Am J Respir Cell Mo! Biol 17:
344-352, 1997
30. Oentbi B, Fan X, Wuthnich RP: Charactenisation of CD44-medi-
ated hyalunonan binding by renal tubular epithelial cells. Nephro!
Dial Transplant 13: 271-278, 1998
3 1 . Stahl RAK, Thaiss F, Disser M, Hebmchen U, Hona K, Schl#{246}n-
dorff D: Increased expression of monocyte chemoattractant pro-
tein- I in anti-thymocyte antibody-induced gbomerulonephnitis.
Kidneylnt44: 1036-1047, 1993
32. Tang WW, Qi M, Warren IS: Monocyte chemoattnactant protein1 mediates gbomerular macrophage infiltration in anti-GBM Ab
ON. Kidney hit 50: 665-671, 1996
33. Zoja C, Liu XH, Donadelli R, Abbate M, Testa D, Corna D,
Tanaboletti 0, Vecchi A, Dong QG, Rollins BI, Bertani T,
Remuzzi 0: Renal expression of monocyte chemoattnactant pro-
tein-l in lupus autoimmune mice. JAm Soc Nephrol 8: 720-729,
1997
34. Tang WW, Qi M, Warren IS, Van GY: Chemokine expression in
experimental tubulointerstitiab nephritis. J mm,nu,zol 159: 870-
876, 1997
35. Rovin BH, Rumancik M, Tan L, Dickerson I: Olomenulan ex-
pression of monocyte chemoattractant protein- 1 in expenimental
and human glomenulonephritis. Lob mnvest 7 1 : 536-542, 1994
36. Prodjosudjadi W, Gemtsma IS, van Es LA, Daha MR. Bnuijn
IA: Monocyte chemoattnactant protein- I in normal and diseasedhuman kidneys: An immunohistochemical analysis. C!i,z Neph-
rol44: 148-155, 1995
37. Prodjosudjadi W, Gemtsma IS, KIar-Mohamad N, Oemtsen AF,

2290 Journal of the American Society of Nephrology I Am Soc Nephrol 9: 2283-2290, 1998
Bruijn JA, Daha MR. van Es LA: Production and cytokine-mediated regulation of monocyte chemoattractant protein- I by
human proximal tubular epithelial cells. Kidney mnt 48: 1477-
1486, 199538. Prodjosudjadi W, Daha MR. Gerritsma IS, Flonijn KW, Baren-
dregt IN, Bruijn IA, van der Woude Fl, van Es LA: Increasedurinary excretion of monocyte chemoattnactant protein- 1 during
acute renal allograft rejection. Nephrol Dial Transplant 1 1:
1096-1103, 1996
39. Gesualdo L, Grandaliano 0, Ranieri E, Monno R, Montinaro V.
Manno C, Schena FP: Monocyte recruitment in cryogbobubine-mic membranoprolifenative glomenulonephritis: A pathogenetic
role for monocyte chemotactic peptide-b. Kidney mnt 51 : 155-
163, 1997
40. Grandaliano 0, Gesualdo L, Ranieri E, Monno R, Stallone 0,
Schena FP: Monocyte chemotactic peptide- 1 expression and
monocyte infiltration in acute renal transplant rejection. Trans-
plantation 63: 414-420, 1997
41. Schmouder RL, Stnieter RM, Kunkeb SL: Interfenon-y regulation
of human renal cortical epithebiab cell-derived monocyte chemo-
tactic peptide-l. Kidney mnt 44: 43-49, 1993
42. Wang Y, Chen I, Chen L, Tay YC, Rangan OK, Harris DC:
Induction of monocyte chemoattractant protein- 1 in proximaltubule cells by urinary protein. JAm Soc Nephrol 8: 1537-1 545,
1997
43. McKee CM, Penno MB, Cowman M, Burdick MD, Strieten RM,
Bao C, Noble PW: Hyaluronan (HA) fragments induce chemo-kine expression in alveolar macrophages: The robe of HA size
and CD44. J Clin mnvest 98: 2403-2413, 1996
44. Hodge-Dufour J, Noble PW, Horton MR, Bao C, Wysoka M,
Burdick MD, Strieter RM, Tninchieri 0, Pure E: Induction of
IL- I 2 and chemokines by hyalunonan requires adhesion-depen-
dent priming of resident but not elicited macrophages. J Immunol
159: 2492-2500, 1997
45. Hallgren R, Gerdin B, Tufveson 0: Hyaluronic acid accumula-
tion and redistribution in rejecting rat kidney graft: Relationship
to the transplantation edema. J Exp Med 17 1 : 2063-2076, 1990
46. Turney IH, Davison AM, Forbes MA, Cooper EH: Hyaluronic
acid in end-stage renal failure treated by haemodiabysis: Clinical
correlates and implications. Nephro! Dial Transplant 6: 566-570, 1991
47. Lipkin OW, Forbes MA, Cooper EH, Turney IH: Hyaluronicacid metabolism and its clinical significance in patients treated
by continuous ambulatory peritoneal dialysis. Nephrol Dial
Transplant 8: 357-360, 1993
48. Aruffo A: CD44: One ligand, two functions. J Clin Invest 98:2191-2192, 1996
49. Yang B, Yang BL, Savani RC, Turley EA: Identification of a
common hyalunonan binding motif in the hyaluronan bindingproteins RHAMM, CD44 and link protein. EMBO J 13: 286-
296, 1994
50. Entwistbe J, Hall CL, Turbey EA: HA receptors: Regulators of
signalling to the cytoskeleton. J Cell Biochem 61 : 569-577, 1996
51. Lesley I, Hyman R, Kincade PW: CD44 and its interaction with
extracebbular matrix. Adv Immunol 54: 271-335, 1993
52. Lesley I, Schulte R, Hyman R: Binding of hyaburonic acid tolymphoid cell lines is inhibited by monocbonal antibodies againstPgp-l. Exp Cell Res 187: 224-233, 1990