hydroxylation of prostaglandins a, and e, by liver ... · the journal of biowgical cwem~~ry vol....
TRANSCRIPT

THE JOURNAL OF BIOWGICAL CWEM~~RY Vol. 253, No. 8, Issue of April 25, pp. 2804-2911, 1979
Printed in U.S.A.
Hydroxylation of Prostaglandins A, and E, by Liver Microsomal Monooxygenase CHARACTERISTICS OF THE ENZYME SYSTEM IN THE GUINEA PIG*
(Received for publication, May 27, 1977)
DAVID KUPFER,$ JAVIER NAVARRO,§ AND DANIEL E. PICCOLO
From the Worcester Foundation for Experimental Biology, Shrewsbury, Massachusetts 01545
Guinea pig liver microsomes in the presence of NADPH catalyzed the hydroxylation of prostaglandin A, (PGA,) and prostaglandin E, (PGE,) primarily (>95%) at them-1 and to a minor extent (~5%) at the w position, yielding 19-hydroxy and 20-hydroxy derivatives. The identity of the 19-hydroxy metabolites was established in the form of 19-OH-PGB, after isolation with the help of high pressure liquid chro- matography (HPLC). The isolated product having charac- teristic UV absorption of PGB, (A,,, = 278 nm) was com- pared with authentic 19-OH-PGB,. The metabolites from PGA, and PGE, and authentic 19-OH-PGB, exhibited iden- tical retention times in HPLC as free acids and as the corresponding methyl esters. The corresponding t-butyldi- methylsilyl ethers-methyl esters also exhibited identical retention times in the gas chromatography (GC) and similar fragmentations in GC/mass spectrometry.
Both NADPH and NADH supported the hydroxylation of PGA, and PGE,, NADH being less effective. There was no synergism by NADH of the NADPH-supported hydroxyla- tion.
Inhibitors of microsomal monooxygenase such as SKF 525A, metyrapone, nicotinamide, and carbon monoxide in- hibited the hydroxylation of PGA, and PGE,; similarly cytochrome c and antibodies to NADPH-cytochrome c reduc- tase inhibited these reactions, indicating that the hydroxyl- ation of these prostaglandins is catalyzed by a typical monooxygenase system. The kinetic constants for the hy- droxylation of PGA, and PGE, were determined; the K, values were 2.1 x 10e4 M and 1.4 x 10e4 M, respectively. The V,,,,, values for the two prostaglandins were also similar, being for PGA, and PGE,, 21.8 and 16.7 nmol/h/mg of protein, respectively.
These findings are discussed in respect to the possibility that prostaglandins are natural substrates for the hepatic monooxygenase.
* This study was supported by Grant GM 22688 from the National Institute of General Medical Sciences, United States Public Health Service. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence and reprint requests should be ad- dressed.
8 Part of this work was in partial fulfillment of an M.Sc. degree. Present address, Universidad Peruana Cayetano, Heredia, Lima, Peru.
Previous studies showed that guinea pig liver microsomes catalyze the Co)- and (o-l)-hydroxylation of prostaglandin A,, yielding 20- and 19-hydroxy-PGA, (1, 2).’ However, under the same conditions these investigators observed little or no metabolism of PGE, (1). Recently we observed, under some- what different conditions,2 that guinea pig liver microsomes could catalyze also the metabolism of PGE,. Our studies also indicated that the oxidative metabolism of PGA, is catalyzed by the hepatic microsomal monooxygenase (4-6). However in these preliminary studies, we made no attempts to character- ize the metabolites of PGA, and PGE,.
In the present investigation on metabolism of PGA, and PGE, by guinea pig liver microsomes we aimed at the follow- ing objectives: (a) develop a rapid chromatographic separation of the w- and o-l-hydroxylated metabolites of PGA, and PGE,; (b) identify these metabolites; and (c) study the characteris- tics of the enzyme system catalyzing PGA, and PGE, hydrox- ylation.
EXPERIMENTAL PROCEDURES
Materials
L5,6-3H]Prostaglandin A,, 87 Cilmmol, was purchased from New England Nuclear (Boston, Mass.). [5,6-3HlProstaglandin E,, 89.5 Ci/ mmol and 40 Ci/mmol were purchased from New England Nuclear (Boston, Mass.) and from AmershamlSearle (Chicago, Ill.), respec- tively. Prostaglandins E, and A, were a gift from Upjohn Co., through the courtesy of Dr. John Pike and 19-hydroxy-PGE, methyl ester was a gift from Dr. John Sih (Upjohn Co.). Dr. Kazuo Sano (One Pharmaceutical Co.) kindly provided 20-OH-PGE,. Glucose-6- phosphate dehydrogenase (EC 1.1.1.49), n-glucose 6-phosphate mon- osodium salt, NADPH monosodium, and cytochrome c type III (horse heart) were purchased from Sigma Chemical Co. (St. Louis, MO.). SKF-525A. HCl (P-diethylaminoethyl diphenylpropylacetate hydro- chloride) was a gift from Smith Kline and French Co. Metyrapone (metopyrone) was obtained from CIBA Co. Thin layer coated plates (Silica Gel G; E. Merck A.G., Germany) were purchased from Brinkmann Instruments. Methanol and acetonitrile for HPLC were
’ The abbreviations used are: PGs, prostaglandins; PGA,, prosta- glandin A,; PGE,, prostaglandin E,; PGB,, prostaglandin B,; 19-OH- PGB,, 19-hydroxy-PGB,; 20-OH-PGB,, 20-hydroxy-PGB,; HPLC, high pressure liquid chromatography; tic, thin layer chromatogra- phy; GC, gas chromatography; MS, mass spectrometry; AUFS, absorbance units full scale.
2 In previous studies (l), nicotinamide was present in the incuba- tion medium, however, based on findings that nicotinamide inhibits monooxygenase activity (3) and on our observations that nicotina- mide inhibits PGA, metabolism (5) we excluded this compound from the incubation.
2804
by guest on August 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

purchased from Burdick and Jackson (Muskegan, Mich.). The sily- Gas Chromatography-Mass Spectrometry (GCIMS) lation reagent (t-butyldimethylsilyl chloride) was obtained from Applied Science Labs (State College, Pa.).
This was carried out using a Perkin-Elmer gas chromatograph model 990 with a flame ionization detector coupled to a Hitachi
Preparation of Authentic Prostaglandin Derivatives Perkin-Elmer mass spectrometer model RMU-6L. The GC was equipped with a silanized glass column (6 feet x 0.25 inch) packed
To obtain 19-OH-PGB,, we hydrolyzed the methyl ester of 19-OH- with 1% SE30 on Gas-chrom Q (100 to 120) and was operated under PGE, (Upjohn Co.); this was achieved by adding 0.1 ml of 10 N KOH the following conditions: column temperature was kept at 220” for 10 to a solution of the methyl ester in 6 ml of aqueous methanol (1:l) min after injection, followed by a 16”/min program to 300” or run and allowing the mixture to stand at room temperature for 19 h. The isothermally at 250”; injection port, 250”; interface, 270”; carrier gas, solution was acidified with 1 N HCl to pH 2, evaporated under a helium (30 ml/min). The mass spectrometer was operated under the stream of N, to 2 ml and extracted with ethyl acetate. The ethyl following conditions: ionizing voltage, 70 eV; accelerating voltage, acetate phase was evaporated to dryness under reduced pressure. 2.2 kV; source, 210”. The resulting residue was in the form of 19-OH-PGB, established with HPLC, and following esterification with diazomethane and Detection, Isolation, and Quantitation of Metabolites silylation with t-butyldimethylsilyl chloride, by GC/MS. 20-OH- PGE, (One Pharmaceutical Co.) was converted to 20-OH-PGB, by
Detection-Usually half of the evaporated ethyl acetate extract
exposure to base in methanol for about l/z h, followed by extraction containing PGB, and metabolites was dissolved in 2 to 3 ml of 0.1 N NaOH. The basic solution was extracted with 3 x 3 ml of chloroform
of the compound as described above for 19-OH-PGB,. and the organic phase was discarded (there was little or no radioac- tivity in the organic phase). The aqueous phase was then acidified
Animals with 1 N HCl to pH 1 to 2 and extracted with 3 x 4 ml of ethyl
Male Hartley strain albino guinea pigs weighing 550 to 600 g (350 acetate. The ethyl acetate phase was evaporated to dryness under a
to 400 g when so specified in text) were obtained from Elm Hill Farm stream of N, at room temperature. The residue was dissolved in
(Chelmsford, Mass.). methanol and the absorbance at 278 nm was measured to assess total amount of PGB, derivatives Q = 27.2 mM-’ cm-‘). The
Microsomes methanol was evaporated to dryness under N, at room temperature and the residue was dissolved in 40 to 100 ~1 of methanol. Further-
Liver microsomes were prepared as previously described (7) and more, the amount of polar metabolite(s) was determined by tic (see were used fresh or stored under layer of 1.15% KC1 at -70”. Under “Quantitation”). This information was utilized to choose an appro- these conditions of storage, activity remained unchanged for several priate aliquot for HPLC (see figures). Since PGB, derivatives have a weeks. Protein determinations were carried out by the Lowry A max = 278 nm, monitoring in HPLC was carried out at 280 and 254 procedure (81, using serum albumin as a standard. nm and peaks having a higher extinction at 280 nm than at 254 nm
were assumed to represent PGB, and PGB, derivatives. Particular
Incubation attention was paid to peaks with retention times corresponding to
One milliliter of a final volume contained sodium phosphate 19-OH-PGB, and 20-OH-PGB,. To establish that the UV-absorbing
buffer (pH 7.4, 50 mM), MgCl, (12.25 mM), microsomal preparation compounds were tritium-labeled PG metabolites, equal (5.0 ml) fractions were collected from the HPLC. The fractions were dried
(for amounts see text) and [5,6-3HlPGA, (0.2 mM, 0.25 &i) or [5,6- 3HlPGE, (0.10 to 0.13 mM; 0.4 to 1 FCi).3 The reaction was usually
under a stream of nitrogen and dissolved in 1 ml of methanol. To
started by adding the NADPH-generating system consisting of each 50-yl aliquot was added 5 ml of Liquifluor (New England
g!ucose 6-phosphate (38.4 mM), NADP (1.3 mM), and glucose-6- Nuclear) and the radioactivity was monitored in a Packard Tri-Carb
phosphate dehydrogenase (3 units). When so specified, NADPH or scintillation spectrometer.
NADH were used instead of the NADPH-generating system. The Isolation of Metabolites - To obtain sufficient quantities of metab-
incubations were carried out by shaking in a Dubnoff incubator at olites for identification purposes, extracts from similar incubations
37”C, in an atmosphere of air. To terminate the reaction and were first combined and processed by HPLC as described above;
simultaneously to achieve conversion of PGA,, PGE,, and metabo- however, since peaks of 19-OH-PGB, and 20-OH-PGB, do not sepa-
lites to the corresponding PGB, derivatives, 0.5-ml aliquot of rate completely, fractions of the peaks which do not overlap were
aqueous NaOH (4 N) was added; the resulting solution was kept at collected. The eluted fractions were evaporated to dryness under
37°C for an additional 30 min4 then was acidified with HCl to pH 2 to nitrogen flow at 20” and dissolved in methanol. The UV absorption
3. The acidified solutions were extracted with 6 volumes of ethyl spectrum was obtained in a Gilford spectrophotometer. To examine
acetate and the organic phase was removed and evaporated under a the purity of a given fraction, aliquots were again analyzed by HPLC.
stream of N, with mild heating. The recoveries of the radioactivity were usually 85 to 95%. In addition, there was no selectivity in
Identification of Metabolites - Aliquots of solutions of PG metabo-
recoveries of the two metabolites, 19-OH-PGB, and 20-OH-PGB,. lites, which were isolated as above, were co-injected with a sample
This was determined by adding known amounts of radiolabeled of an authentic 19-OH-PGB,. The appearance of a single peak
hydroxylated metabolites (isolated by HPLC as described below) to representing additive intensities was assumed to indicate identical
the complete incubation mixture, which was previously incubated retention times of the metabolite and the corresponding authentic
for 1 h, and following the extraction procedure as above; recoveries compound. Subsequently the isolated metabolite(s) were esterified
of 19-OH-PGB, and 20-OH-PGB, were 88 to 98%. with diazomethane (generated from N-methyl-N-nitrosourea) and their retention time in HPLC was compared with that of esterified 19-OH-PGB, and 20-OH-PGB,. Portions of the esteritied PGs (metab-
High Pressure Liquid Chromatography (HPLC) olites or authentic compounds) were silylated with t-butyldimethyl-
The high pressure liquid chromatography system (Waters Associ- silyl chloride in the presence of imidazole for 48 to 72 h at room
ates) was composed of ALC/GPC 204 liquid chromatograph equipped temperature. To extract the derivatized PGs, water was added
with dual wavelength UV detector (model 440), solvent delivery followed by three extractions with hexane. The hexane extract was
system (model 6000A), injector (model U6K), and solvent flow evaporated to dryness under N,, the residue was dissolved in
programmer (model 660). Reverse phase chromatography was car- methanol and recoveries were calculated using at 278 nm = 27.2
ried out with a “fatty acid” column (4 mm x 30 cm) (Waters rnM-‘.cm-’ (9). Aliquots of some of these solutions which were
Associates) using acetonitrile/l% aqueous acetic acid for the free subjected to HPLC, revealed a dramatic prolongation of retention
acids or acetonitrile/water for the methyl esters. time of the silylated prostaglandin derivatives over that of the corresponding methyl esters. This indicated conversion to highly nonpolar substances as would be expected from the corresponding
3 When L3H]PGE, or L3HIPGA, were not incubated but were silyl ethers. converted to PGB, by base treatment, the same chromatographic Quantitation - Residual extracts, which did not undergo HPLC peak was observed. There was only one detectable peak, demonstrat- (for work-up see “Incubation”), were dissolved in a few drops of ing radiochemical purity of 97+%. methanol and were chromatographed on thin layer plates (previ-
4 We determined that conversion of PGE, or PGA, to PGB,, as ously activated by heating at 110” for 30 min). The solvent system observed by the appearance of absorbance at 278 nm, was complete consisted of ethyl acetate/acetic acid/2,2,4-trimethylpentanelwater
. .~ wlthin 30 min.
. (11U:10:20:100). The plates were dried in air and were scanned on a
Hydroxylation of Prostaglandins by Liver Monooxygenase 2805
by guest on August 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
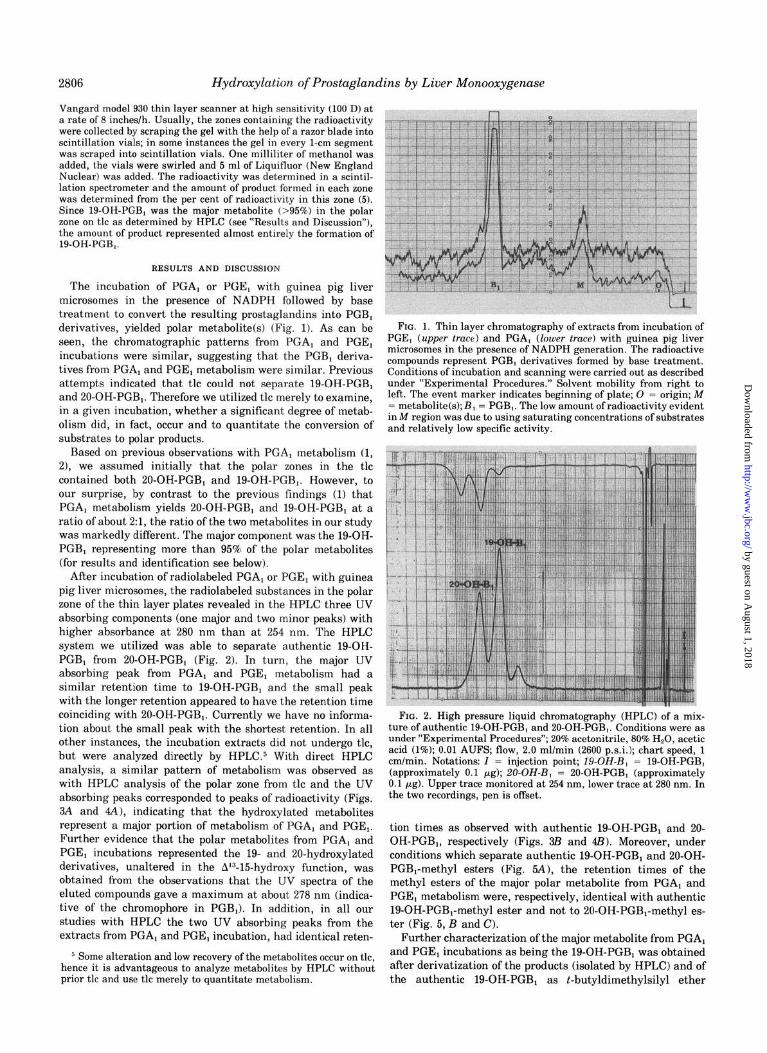
2806 Hydroxylation of Prostaglandins by Liver Monooxygenase
Vangard model 930 thin layer scanner at high sensitivity (100 D) at a rate of 8 inches/h. Usually, the zones containing the radioactivity were collected by scraping the gel with the help of a razor blade into scintillation vials; in some instances the gel in every l-cm segment was scraped into scintillation vials. One milliliter of methanol was added, the vials were swirled and 5 ml of Liquifluor (New England Nuclear) was added. The radioactivity was determined in a scintil- lation spectrometer and the amount of product formed in each sane was determined from the per cent of radioactivity in this zone (5). Since lQ-OH-PGB, was the major metabolite 1>95%) in the polar zone on tic as determined by HPLC (see “Results and Discussion”), the amount of product represented almost entirely the formation of 19-OH-PGB,.
RESULTS AND DISCUSSION
The incubation of PGA, or PGE, with guinea pig liver microsomes in the presence of NADPH followed by base treatment to convert the resulting prostaglandins into PGBl derivatives, yielded polar metabolite(s) (Fig. 11. As can be seen, the chromatographic patterns from PGA, and PGE, incubations were similar, suggesting that the PGB, deriva- tives from PGA, and PGE, metabolism were similar. Previous attempts indicated that tic could not separate 19-OH-PGB1 and 20-OH-PGB,. Therefore we utilized tic merely to examine, in a given incubation, whether a significant degree of metab- olism did, in fact, occur and to quantitate the conversion of substrates to polar products.
Based on previous observations with PGA, metabolism (1, 2), we assumed initially that the polar zones in the tic contained both 20-OH-PGB, and 19-OH-PGB,. However, to our surprise, by contrast to the previous findings (1) that PGA, metabolism yields 20-OH-PGBI and 19-OH-PGB, at a ratio of about 2:1, the ratio of the two metabolites in our study was markedly different. The major component was the 19-OH- PGBl representing more than 95% of the polar metabolites (for results and identification see below).
After incubation of radiolabeled PGA, or PGE, with guinea pig liver microsomes, the radiolabeled substances in the polar zone of the thin layer plates revealed in the HPLC three UV absorbing components (one major and two minor peaks) with higher absorbance at 280 nm than at 254 nm. The HPLC system we utilized was able to separate authentic 19-OH- PGBi from PO-OH-PGB, (Fig. 21. In turn, the major UV absorbing peak from PGA, and PGE, metabolism had a similar retention time to 19-OH-PGB1 and the small peak with the longer retention appeared to have the retention time coinciding with 20-OH-PGB1. Currently we have no informa- tion about the small peak with the shortest retention. In all other instances, the incubation extracts did not undergo tic, but were analyzed directly by HPLC.$ With direct HPLC analysis, a similar pattern of metabolism was observed as with HPLC analysis of the polar zone from tic and the UV absorbing peaks corresponded to peaks of radioactivity (Figs. 8A and 4A), indicating that the hydroxylated metabolites represent a major portion of metabolism of PGA, and PGE,. Further evidence that the polar metabolites from PGA, and PGE, incubations represented the 19- and 20-hydroxylated derivatives, unaltered in the A13-15-hydroxy function, was obtained from the observations that the UV spectra of the eluted compounds gave a maximum at about 278 nm (indica- tive of the chromophore in PGB,). In addition, in all our studies with HPLC the two UV absorbing peaks from the extracts from PGA, and PGE, incubation, had identical reten-
5 Some alteration and low recovery of the metabolites occur on tic, hence it is advantageous to analyze metabolites by HPLC without prior tic and use tic merely to quantitatc metabolism.
FIG. 1. Thin layer chromatography of extracts from incubation of PGE, (upper truce) and PGA, (lower truce) with guinea pig liver microsomes in the presence of NADPH generation. The radioactive compounds represent PGB, derivatives formed by base treatment. Conditions of incubation and scanning were carried out as described under “Experimental Procedures.” Solvent mobility from right to left. The event marker indicates beginning of plate; 0 = origin; M = metabolitefs); B, = PGB,. The low amount of radioactivity evident in M region was due to using saturating concentrations of substrates and relatively low specific activity.
FIG. 2. High pressure liquid chromatography (HPLC) of a mix- ture of authentic 19-OH-PGB, and 20-OH-PGB,. Conditions were as under “Experimental Procedures”; 20% acetonitrile, 80% H,O, acetic acid (1%); 0.01 AUF*, flow, 2.0 ml/min (2600 p.s.i.1; chart speed, 1 cm/min. Notations: I = injection point; 19-OH-B, = 19-OH-PGB, (approximately 0.1 gg); 20-OH-B, = PO-OH-PGB, (approximately 0.1 +g). Upper trace monitored at 254 nm, lower trace at 280 nm. In the two recordings, pen is offset.
tion times as observed with authentic 19-OH-PGB, and 20- OH-PGB1, respectively (Figs. 8B and 4Bl. Moreover, under conditions which separate authentic 19-OH-PGB1 and 20-OH- PGB,-methyl esters (Fig. 5A1, the retention times of the methyl esters of the major polar metabolite from PGA, and PGE, metabolism were, respectively, identical with authentic 19-OH-PGB,-methyl ester and not to 20-OH-PGB,-methyl es- ter (Fig. 5, B and C).
Further characterization of the major metabolite from PGA, and PGE, incubations as being the 19-OH-PGBi was obtained after derivatization of the products (isolated by HPLC) and of the authentic 19-OH-PGB, as t-butyldimethylsilyl ether
by guest on August 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
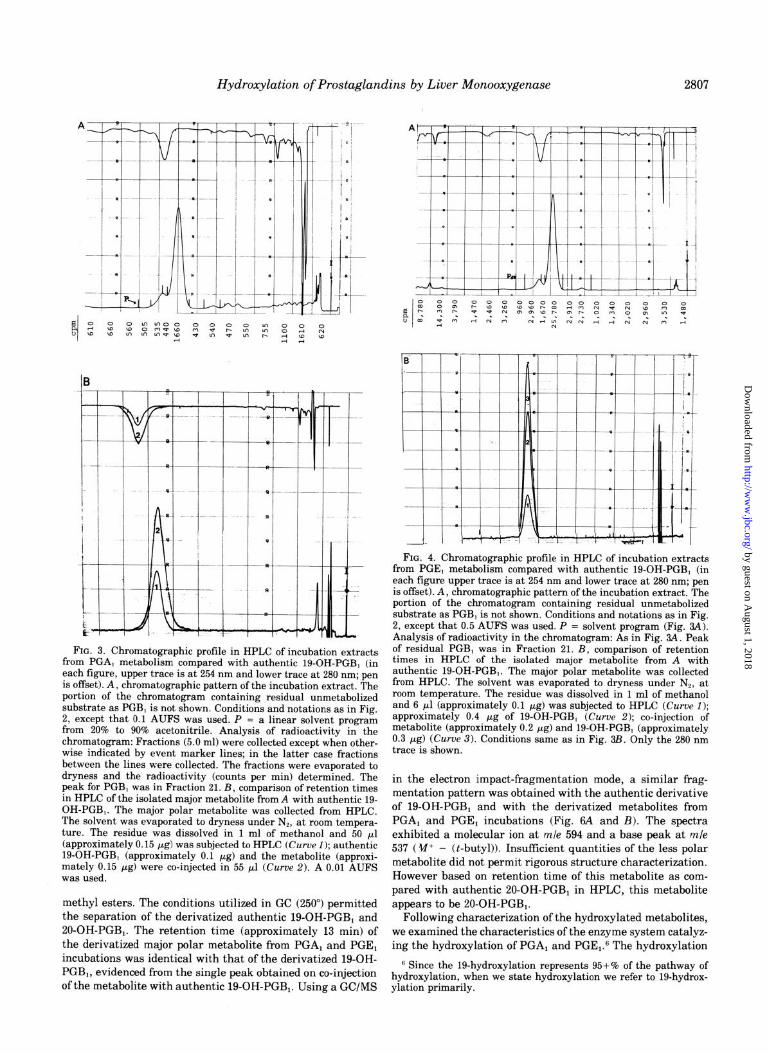
Hydroxylation of Prostaglandins by Liver Monooxygenase 2807
FIG. 3. Chromatographic profile in HPLC of incubation extracts from PGA, metabolism compared with authentic 19-OH-PGB, (in each figure, upper trace is at 254 nm and lower trace at 280 nm; pen is offset). A, chromatographic pattern of the incubation extract. The portion of the chromatogram containing residual unmetabolized substrate as PGB, is not shown. Conditions and notations as in Fig. 2, except that 0.1 AUFS was used. P = a linear solvent program from 20% to 90% acetonitrile. Analysis of radioactivity in the chromatogram: Fractions (5.0 ml) were collected except when other- wise indicated by event marker lines; in the latter case fractions between the lines were collected. The fractions were evaporated to dryness and the radioactivity (counts per min) determined. The peak for PGB, was in Fraction 21. B, comparison of retention times in HPLC of the isolated major metabolite from A with authentic 19- OH-PGB,. The major polar metabolite was collected from HPLC. The solvent was evaporated to dryness under N,, at room tempera- ture. The residue was dissolved in 1 ml of methanol and 50 ~1 (approximately 0.15 pg) was subjected to HPLC (Curve 1); authentic 19-OH-PGB, (approximately 0.1 pg) and the metabolite (approxi- mately 0.15 Fg) were co-injected in 55 ~1 (Curve 2). A 0.01 AUFS was used.
methyl esters. The conditions utilized in GC (250”) permitted the separation of the derivatized authentic 19-OH-PGB, and OO-OH-PGB,. The retention time (approximately 13 min) of the derivatized major polar metabolite from PGA, and PGE, incubations was identical with that of the derivatized 19-OH- PGB,, evidenced from the single peak obtained on co-injection ofthe metabolite with authentic 19-OH-PGBi. Using a GC/MS
FIG. 4. Chromatographic profile in HPLC of incubation extracts from PGE, metabolism compared with authentic 19-OH-PGB, (in each figure upper trace is at 254 nm and lower trace at 280 nm; pen is offset). A, chromatographic pattern of the incubation extract. The portion of the chromatogram containing residual unmetabolized substrate as PGB, is not shown. Conditions and notations as in Fig. 2, except that 0.5 AUFS was used. P = solvent program (Fig. 84). Analysis of radioactivity in the chromatogram: As in Fig. 84. Peak of residual PGB, was in Fraction 21. B, comparison of retention times in HPLC of the isolated major metabolite from A with authentic 19-OH-PGB,. The major polar metabolite was collected from HPLC. The solvent was evaporated to dryness under Nf, at room temperature. The residue was dissolved in 1 ml of methanol and 6 ~1 (approximately 0.1 pg) was subjected to HPLC (Curve 1); approximately 0.4 pg of 19-OH-PGB, (Curve 2); co-injection of metabolite (approximately 0.2 PgcLg) and 19-OH-PGB, (approximately 0.3 pg) (Curve 3). Conditions same as in Fig. 3B. Only the 280 nm trace is shown.
in the electron impact-fragmentation mode, a similar frag- mentation pattern was obtained with the authentic derivative of 19-OH-PGBi and with the derivatized metabolites from PGA, and PGEi incubations (Fig. 6A and B). The spectra exhibited a molecular ion at mle 594 and a base peak at mle 537 ( V+ - (t-butyl)). Insuffrcient quantities of the less polar metabolite did not permit rigorous structure characterization. However based on retention time of this metabolite as com- pared with authentic 20-OH-PGBi in HPLC, this metabolite appears to be 20-OH-PGBi.
Following characterization of the hydroxylated metabolites, we examined the characteristics of the enzyme system catalyz- ing the hydroxylation of PGA, and PGE,.G The hydroxylation
B Since the 19-hydroxylation represents 95+% of the pathway of hydroxylation, when we state hydroxylation we refer to 19-hydrox- ylation primarily.
by guest on August 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2808 Hydroxylation of Prostaglandins by Liver Monooxygenase
FIG. 5. Chromatographic profile of methyl esters of the major was observed that when 19-OH-B,-Me and 20-OH-B,-Me are individ- polar metabolites (PGB, derivatives) from incubations of PGE, and ually injected the 19-OH derivative has the shorter retention time. PGA, and comparison with methyl esters of 19-OH-PGB, and 20-OH- B, metabolite ester from PGA, incubation. Curve 1, methyl ester of PGB,. A, separation of methyl esters of 19-OH-PGB, (IS-OH-B&z) metabolite (approximately 0.3 Kg); Curve 2, co-injection of ester of and of 20-OH-PGB, (20-OH-B,-Me). The esters of 19-OH-PGB, (ap- metabolite (approximately 0.3 pg) and 19-OH-PGB,-methyl ester proximately 0.3 pgl and of 20-OH-PGB, (approximately 0.2 pg) were (approximately 0.3 pg). Conditions same as in A. C, metabolite injected in 13 ~1 of methanol. The solvent was acetonitrile (25%) and ester from PGE, incubation. Curue 1, methyl ester of metabolite H,O (75%); solvent flow = 2.0 ml/min (1700 p.s.i.1; 0.01 AUFS; chart (approximately 0.3 pg); Curve 2, co-injection of ester of metabolite speed 1 cm/min. Upper trace (254 nm) and lower trace (280 nm); pen (approximately 0.3 pg) and 19-OH-PGB,-methyl ester (approxi- is offset for the two tracings. In another experiment (not shown) it mately 0.3 kg). Conditions same as in A.
FIG. 6. Mass fragmentation pattern of the t-butyldimethylsilyl OH-PGB, and for the derivatized PGA, and PGE, metabolites. ether methyl ester derivatives of the authentic 19-OH-PGB, (A) and Furthermore the GC separated the derivatives of authentic 19-OH- of the pooled, HPLC-purified, metabolite from PGE, incubation (B). PGB, and 20-OH-PGB,. We also observed that the fragmentation The mass fragmentation of a similarly isolated and derivatised pattern of the trimethylsilyl ether of the methyl ester of the isolated metabolite from PGA, incubation (not shown) matched the mass base-treated metabolite from PGA, incubation (not shown) was spectrum of the authentic derivatized 19-OH-PGB,. The GCIMS, identical with the previously reported similarly derivatized 19-OH- described under “Experimental Procedures,” demonstrated identical PGB, (1). retention times (by co-injection) for the derivatized authentic 19-
by guest on August 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Hydroxylation of Prostaglandins by Liver Monooxygenase 2809
40 A
PROTEIN (mg)
FIG. 7 (left). Linearity of hydroxylation of PGA, (0) and PGE, (0) with varying microsomal protein concentrations and duration of incubation. A, product formation UCFSUS microsomal protein concen- tration; incubation time was 30 min. B, product formation uersus duration of time of incubation: A, (2.7 mg of protein), E, (3.0 mg of protein).
TIME (min) ‘h(M)
FIG. 8 (right). Lineweaver-Burk plots of PGA, (0-O) and PGE, (0- - -0) hydroxylation by guinea pig liver microsomes. PGA,-hydroxylation, 2.4 mg of microsomal protein per incubation; PGE,-hydroxylation, 2.9 mg of microsomal protein per incubation. V = 19-OH-PGB, formed (nanomoles per h per mg of protein); S = substrate in molar (M) concentrations.
of PGA, was linear up to 2.7 mg of microsomal protein and up
to 45 min of incubation (Fig. 7 A and B). The hydroxylation of PGE, demonstrated linearity to 3 mg of microsomal protein (with only minor deviation from linearity up to 5 mg of protein) and up to 30 min of incubation. Hydroxylation of PGA, and PGE, was supported by either NADPH or NADH (Table I); NADH was less effective than NADPH. However, NADH did not augment the hydroxylation rate supported by
saturating concentrations of NADPH, indicating that there is no synergism by NADH of PGA, and PGE, hydroxylation.
TABLE I
Hydrozylation of PGA, and PGE, by guinea pig liver microsomes supported by NADPH and NADH
Inhibition of the hydroxylation of PGA, and PGE, was observed by recognized inhibitors of microsomal monooxygen-
ase (Table II). The observation, in the presence of NADPH, that cytochrome c inhibited PGA, and PGEr hydroxylation suggested that the flavoprotein, NADPH-cytochrome c reduc- tase, was involved in the reaction. Additional evidence for the participation of this flavoprotein was obtained with antibodies specific for NADPH-cytochrome c reductase’ (Table III). As can be seen, the anti-rat NADPH-cytochrome c globulin inhibited the hydroxylation of PGA, and PGE, by guinea pig liver microsomes. Based on present and previous finding@ that antibodies to NADPH-cytochrome c reductase inhibit both the NADH- and NADPH-supported hydroxylation of PGA, and PGE,, it appears that the same flavoprotein is involved in both the NADPH and NADH-supported hydroxyl- ations. Such a conclusion was previously obtained by Prough and Burke (11) with monooxygenase activities in liver and lung of hamster. These investigators observed that antibodies to rat liver NADPH-cytochrome c reductase inhibited the NADH- and NADPH-sustained benzpyrene and biphenyl hy- droxylations and ethylmorphine demethylations. Previous in- vestigations showed that NADH causes synergism of the NADPH-mediated metabolism of various substrates of hepatic monooxygenase in the rat and it has been proposed that cytochrome b, is involved in donating the 2nd electron to cytochrome P-450 (12-15). It seems that in the guinea pig there is no apparent synergism by NADH (Table I), however whether cytochrome b, is involved in the hydroxylation of PGA, or PGE, cannot be concluded from these studies.
Experiments with PGA,. Experiment 1, 100% control = 7.7 nmol of productlmg of protein/45 min; microsomal protein = 5.9 mglml; incubation time was 45 min. Experiment 2, 100% control = 12.7 nmol of product/mg of protein/30 min; microsomal protein = 1.8 mg; incubation time 30 min. NADPH (2.5 mg) appeared saturating. Experiments with PGE,. Experiment 3, 100% = 3.6 nmol of product/ mg of protein/60 min; microsomal protein = 4.5 mglml; incubation time 60 min. Experiment 4, 100% = 9.4 nmol of product/mg of protein/30 min; microsomal protein = 1.8 mg; incubation time 30 min. NADPH (4.0 mg) appeared saturating. Concentrations of PGA, and PGE, were 0.2 mM, except in Experiment 3 when PGE, was at 0.1 mM. The rate of hydroxylation of benzpyrene into phenols (10) by guinea pig liver microsomes (using same batch of microsomes as above in Experiments 2 and 4) supported by NADH was only about 8% of the rate observed in the presence of NADPH, demonstrating that NADH is much more efficient in supporting prostaglandin hydroxylation than benzpyrene hydroxylation and indicating that the NADH-supported hydroxylation is not due to conversion of NADH into NADPH.
PGA, PGE,
Product in Ex- Product in Ex-
NADPH NADH periment NADPH NADH periment
1 2 3 4
w % control w % control
2.5 100 100 4.0 100 100 1.0 57 75 1.5 62 72
2.5 31 23 4.0 60 34 2.5 2.5 102 111 4.0 4.0 98 108 1.0 2.5 86 106 1.5 4.0 85 100
The unexpected finding that a-naphthoflavone inhibits PGA, hydroxylation (Table II) is of interest. Benzpyrene hydroxylation by liver microsomes from 3-methylcholan- threne-treated, but not from control or phenobarbital-treated,
’ In a preliminary experiment, we demonstrated that these anti- bodies inhibited the NADPH- but not the NADH-cytochrome c reductase.
* Unpublished observations.
animals is strongly inhibited by a-naphthoflavone (16, 17). Differences in susceptibilities to cY-naphthoflavone by cyto- chromes P-448 (P,-450) produced by 3-methylcholanthrene treatment uersus P-450s present in control or produced by phenobarbital treatment, is probably the cause. Whether the cytochrome P-450 in the guinea pig liver catalyzing PGA, hydroxylation mimics the catalytic characteristics of rat liver
by guest on August 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2810 Hydroxylation of Prostaglandins by Liver Monooxygenase
TABLE II
Effect of inhibitors of monooxygenase on PGA, and PGE, hydroxylation by guinea pig liver microsomes in presence of NADPH
Each value represents a mean of duplicate incubations (agreeing within less than 10%). In the absence of NADPH-generating system, there was no detectable metabolism of PGA, or PGE,. PGA,: Exper- iment 1, Column 1: 100% control was 7.6 nmol of product/mg of protein/45 min (5.5 mg of protein); Column 2: 100% control was 6.7 nmol/mg of protein/30 min (2.3 mg of protein per incubation). Experiment 2, Column 1: 100% control was 8.1 nmol of productlmg of protein/h (3.2 mg of protein); Column 2: same as in Experiment 1. Experiment 3, 100% control (0,/N, (1:9)1 was 9.2 nmol of product/mg of protein/h (5.5 mg of protein). PGE,: Experiment 1, Column 3: 100% control was 7.5 nmol of product/mg of protein/h (6.3 mg of protein); Column 4: 100% control was 7.5 nmol/mg of protein/30 min (2.1 mg of protein). Experiment 2, Column 3: 100% control was 6.3 nmol/mg of protein/h (3.2 mg of protein); Column 4: 100% control same as in Experiment 1. Experiment 3: 100% control (0,/N, (1:9)) was 9.1 nmol/mg of protein/h (2.8 mg of protein). Column 3: guinea pigs weighed 350 to 400 g (550 to 600 g in other columns).
PGA, PGE, Inhibitor
1 2 3 4 % control
Experiment 1 SKF 525A (1 mM) 44.4” 33.8 46.7 49.4 Metyrapone (5 mM) 38.0” 37.7 16.0 13.0 KCN (1 mM) -0 97.4 88.0 89.6 ANF (0.01 rnr# -b 39.0 -b -b
Cytochrome c (0.5 mM) 00 -b 0 -b
Experiment 2 Nicotinamide (10 mM) 54.0 59.7 44.0 66.2 Nicotinamide (30 mM) -b 40.3 -b 54.5
Experiment 3 COIO,/N2 (5:1:4) 18.8” 22.2
a Values which have been previously communicated by us (5). b -, not determined. c The inhibition by ANF (oc-naphthoflavonel appeared to be max-
imal at 0.01 mM since at 0.1 rnM similar level of inhibition was observed.
P-448 in aspects other than susceptibility to cw-naphthofla- vone, requires further investigation.
Previous studies showed that substrates of monooxygenase enhance the rate of NADPH oxidation by liver microsomes and in certain cases stoichiometry of NADPH oxidation with product formation was demonstrated (18-21). In the present study, both PGA, and PGE, appear to stimulate NADPH oxidation (Table IV). However, the stimulation of NADPH oxidation by prostaglandins was much lower than by benz- phetamine; this finding was expected from the much lower rate of hydroxylation of prostaglandins as compared with benzphetamine demethylation. Benzphetamine was demethy- lated at 4.6 ? 0.6 nmol of HCHO/min/mg of protein, which represents about 30-fold faster rate than hydroxylation of PGA, and PGE,. Carbon monoxide did not inhibit markedly NADPH oxidation in the absence of added substrates, however CO produced a marked inhibition of the stimulation of NADPH oxidation by these substrates, indicating that inhibi- tion of metabolism of PGA, and PGE, by CO (Table II) is probably the reason for the inhibition of the stimulation of NADPH oxidation by CO.
The kinetic constants for PGA, and PGE, hydroxylations were determined (Fig. 8). The K,,, values for PGA, and PGE, hydroxylation were 2.1 x 1O-4 and 1.4 x lo-” M, respectively, being similar to the previously reported K, value for PGA,, which was 1.7 x 10m4 M (4), suggesting that the K, reflects
binding of PGA, to the active site of the cytochrome. Such a speculation concerning the relation of K, to K,,, was proposed in the early studies on binding of drugs to rat liver microsomes (22, 23). However this proposition has been challenged due to observations that with certain compounds the K, and K, values differ widely (24). The V,,,,, values for PGA, and PGE, were also similar (21.8 and 16.7 nmol/h/mg of protein, respec- tively), indicating that the two compounds appear to act as alternate substrates for the same monooxygenase. Since the circulating levels of prostaglandins are probably several or- ders of magnitude lower than these K, values, V,,,,, velocities are probably never achieved in uivo under normal circum- stances. However, it is possible that the administration of foreign substances, such as drugs, which could serve as alternate substrates of the monooxygenase, might interfere with the metabolism of endogenous prostaglandins.
The dramatic difference in the ratio of IO-OH-PGB, to 19- OH-PGB, derived from metabolism of PGA, observed by us versus that reported by other investigators (1) is puzzling. It is possible that strain differences in guinea pigs used in the two respective studies may contribute to the marked quantitative differences in the ratio of the hydroxylated metabolites. It is also possible that nicotinamide at high concentrations (30 mM), utilized in the previous studies (1, 21, suppressed prefer- entially the 19-hydroxylation, yielding > high ratio of 20-OH-
TABLE III
Effects of anti-rut NADPH-cytochrome c rednctase globulin on PGA, and PGE, hydroxylation by guinea pig liuer microsomes
Experiment with PGA,: 0.8 mg of microsomal protein was used per incubation; 100% control = 8.5 nmol/h; NADPH was generated; 10.8 mg of globulin fraction was used. Experiment with PGE,: 2.8 mg of microsomal protein was used per incubation; 100% control = 6.9 nmol/h; NADPH (4 mg, 4.4 mM); 3.4 mg of globulin fraction was used.
Control + Nonimmune globulin + Immune globulin
PGA, PGE, % control
100 100 89 116 40 53
TABLE IV
Stimulation of NADPH oxidation by PGA, and PGE, and effect of carbon monoxide on NADPH oxidation
Each cuvette contained 2 mg of microsomal protein and 0.23 mg of NADPH in 3 ml containing potassium phosphate (pH 7.4, 0.053 M),
MgCl, (11.6 mM), and EDTA’2Na (0.2 mM). NADPH was assayed at 340 nm. The reaction was started by adding NADPH and allowed to proceed at 37”. When so indicated PGA, (1.2 x 1O-4 M), PGE, (1.3 x
10m4 M) and benzphetamine (3.3 x 1O-4 M) were present. NADPH oxi- Per cent stim-
d&ion ulation
nmollminlmg protein
Control 5.7 +PGA, 8.6 51 +PGE, 7.8 37 +Benzphetamine 19.7 246
Control +CO” 4.1 +CO + PGA, 4.8 17 (33Y +CO + PGE, 4.5 10 (27) +CO + benzphetamine 9.7 136 (85)
a The gas mixture was COIOJN, (5:1:4). * In parentheses, inhibition by CO of the stimulation of NADPH
oxidation given as per cent control without CO.
by guest on August 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Hydroxylation of Prostaglandins by Liver Monooxygenase 2811
PGB, to 19-OH-PGB,. In fact, we observed that nicotinamide inhibited significantly the overall hydroxylation of PGA, and PGE, (Table II), and preliminary observations suggest that 19-hydroxylation is inhibited more than 20-hydroxylation. Also of interest might be the finding that nicotinamide (12 mM) activates the o-hydroxylation but inhibits the (o-l)- hydroxylation of 4-chloropropionalide by rabbit liver micro- somes (25).
Studies in several species demonstrated that endogenous and exogenous prostaglandins are metabolized in vivo to w oxygenated derivatives (2, 26-28). The observations that the hepatic monooxygenase catalyzes the wl- and whydroxyl- ation of PGA, (1,2, 4-6) and the present findings on PGA, and PGE, hydroxylation suggest that this enzyme system may be involved in an essential step of the transformation of endoge- nous prostaglandins into hydrophilic urinary metabolites. If this were the case, then it would be of interest to establish whether inhibition of monooxygenase activity would alter the urinary excretion of endogenous or exogenous prostaglandins and could answer the question whether monooxygenase activ- ity is of prime importance in prostaglandin elimination. Such experiments will be the subject of future investigations.
AcknowZedgments -We greatly appreciate the help pro- vided by Dr. Warren Anderson in preparing 19-OH-PGB, (from human semen) and with some of the GC/MS studies. We thank Mr. Gregory Miranda for exceptional technical assistance. The generous supply of anti-rat NADPH-cyto- chrome c reductase globulin by Dr. Bettie Sue Siler Masters is gratefully acknowledged (B.S.S. M. was supported by National Institutes of Health Grant HL 13619.). We are extremely thankful to Doctors John Pike and John Sih (Upjohn Co.), Dr. Kazuo Sano (Ono Co.), and Dr. R. W. Kelly (MRC Reproduc- tive Biology Unit, Edinburgh) for supplying various prosta- glandin derivatives. We particularly wish to thank Dr. Cath- erine E. Costello of the National Institutes of Health mass spectrometry facility at MIT (Grant RR 00317, Division of Research Resources; K. Beimann, Principal Investigator) for her help with the GC/MS spectra.
REFERENCES
1. Israelsson, U., Hamberg, M., and Samuelsson, B. (1969)Eur. J. Biochem. 11, 390-394
2. Samuelsson, B., Granstriim, E., GrBen, K., and Hamberg, M. (1971) Ann. N. Y. Acad. Sci. 180, 138-163
3. Schenkman, J.B., Ball, J. A., and Estabrook, R. W. (1967) Biochem. Pharmacol. 16, 1071-1081
4. Kuufer. D. (1974) Life Sci. 15. 657-670 5. Kupfer; D., and Navarro, J. (1976) Life Sci. 18, 507-514 6. Kupfer, D., and Navarro, J. (1977) Proceedings of the 3rd
I&ernat&al Symposikon Microsomes and Drug &id&ion, Berlin, Germany, July 21-24, 1976, (Ullrich, V., Hilden- brandt. A.. Roots. I.. Estabrook. R. W.. and Connev. A. H.. eds) pp. 370-376, Periamon Press, Oxfoid
*,
7. Burstein, S. H.. and Kuofer. D. (1971) Ann. N. Y. Acad. Sci. 191, 61-67 _
8. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951) J. Biol. Chem. 193, 265-275
9. Jones, R. L. (1972) J. Lipid Res. 13, 511-518 Brown, J. E., and Kupfer, D. (1974) Chem.-Biol. Interact. 10,57-
70 10.
11.
12.
13.
Prough, R. A., and Burke, M. D. (1975) Arch. Biochem. Biophys. 170, 160-168
Hildenbrandt, A. G., and Estabrook, R. W. (1971) Arch. Bio- them. Biophys. 143, 66-79
Estabrook, R. W., Hildenbrandt, A. G., Baron, J., Netter, K. J., and Leibman, K. (1971) Biochem. Biophys. Res. Commun. 42, 132-139
14.
15.
16.
17.
18. 19.
20.
21.
22.
23. 24.
Baron, J., Hildenbrandt, A. G., Paterson, J. A., and Estabrook, R. W. (1973) Drug Metab. Disp. 1, 129-138
Sasame, H. A., Mitchell, J. R., Thorgeirsson, S. S., and Gillette, J. R. (1973) Drur! Metab. Disk. 1. 150-155
Wiebel, F. J., Le&z, J. C., Diambnd, L., and Gelboin, H. V. (1971) Arch. B&hem. B&whys. 144, 78
Goujon, F. M., Nebert, D.-W., and .Gielen, J. E. (1972) Mol. Pharmacol. 8, 667-680
Orrenius, S. (1965) J. Cell. Biol. 26, 713-723 Lu, A. Y. H., Strobel, H. W., and Coon, M. J. (1969) Biochem.
Biophys. Res. Commun. 36, 545-551 Buening, M. K., and Franklin, M. R. (1974) Mol. Pharmacol. 10,
999-1003 Jeffery, E., and Mannering, G. J. (1974) Mol. Pharmacol. 10,
1004-1008 Schenkman, J. B., Remmer, H., and Estabrook, R. W. (1967)
Mol. Pharmacol. 3, 113-123 Schenkman, J. B. (1970) Biochemistry 9, 2081-2091 Mannering, G. J. (1971) in Fundamentals of Drug Metabolism
and Drug Disposition (LaDu, B. N., Mandel, H. G., and Way, E. L., eds.) pp. 206-252, Williams & Wilkins Co., Baltimore, Md.
25. 26.
Lenk, W. (1975) Biochem. Sot. Trans. 3, 961-962 Svanborg, K., and Bygdeman, M. (1972) Eur. J. B&hem. 28,
127-135 27. 28.
Sun, F. F. (1974) Biochim. Biophys. Acta 348, 249-262 Sun, F. F., and Stafford, J. E. (1974) B&him. Biophys. Acta
369, 95-110
by guest on August 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D Kupfer, J Navarro and D E Piccolomonooxygenase. Characteristics of the enzyme system in the guinea pig.
Hydroxylation of prostaglandins A1 and E1 by liver microsomal
1978, 253:2804-2811.J. Biol. Chem.
http://www.jbc.org/content/253/8/2804.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/253/8/2804.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on August 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from