hypericin suppresses osteoclast formation and wear particle-induced osteolysis via modulating erk...
TRANSCRIPT

Biochemical Pharmacology 90 (2014) 276–287
Hypericin suppresses osteoclast formation and wear particle-inducedosteolysis via modulating ERK signalling pathway
Zhengxiao Ouyang a,b,1, Zanjing Zhai a,1, Haowei Li a,1, Xuqiang Liu a, Xinhua Qu a,Xianan Li b, Qiming Fan a, Tingting Tang a, An Qin a,*, Kerong Dai a,*a Department of Orthopaedics, Shanghai Key Laboratory of Orthopaedic Implant, Shanghai Ninth People’s Hospital,
Shanghai Jiaotong University School of Medicine, Shanghai 200011, Chinab Department of Orthopaedics, Hunan Cancer Hospital and The Affiliated Cancer Hospital of Xiangya School of Medicine, Central South University,
Changsha, Hunan 410012, China
A R T I C L E I N F O
Article history:
Received 15 May 2014
Accepted 10 June 2014
Available online 17 June 2014
Keywords:
Prosthetic loosening
Hypericin
Osteoclast
Osteolysis
ERK
A B S T R A C T
Osteoclast-induced bone resorption and wear-particle-induced osteolysis leads to prosthetic loosening,
one of the most common causes of joint implant failure, resulting in revision surgery. Thus, inhibition of
osteoclastic bone resorption, which further prevents wear particle-induced osteolysis, is a potential
treatment strategy for prosthetic loosening. Here, we examined the therapeutic effect of hypericin (HP),
which was photosensitive, on osteoclastogenesis and wear particle-induced osteolysis in the absence of
visible light. HP inhibited RANKL-induced osteoclast differentiation in bone marrow macrophages
(BMMs) and RAW264.7 cell line without any evidence of cytotoxicity. The bone-resorbing activity of
mature osteoclasts was significantly inhibited by HP. As HP has been previously reported to inhibit
signalling pathway such as ERK and NF-kB in other cells, which is also important in osteoclast
differentiation. We thus examined the molecular mechanism and showed that HP significantly inhibited
the ERK/mitogen-activated protein kinase (MAPK) signalling pathway without affecting nuclear factor
kappaB (NF-kB), c-Jun N-terminal kinase (JNK) and p38 signalling in RANKL-stimulated BMMs. Further
in vivo studies revealed HP attenuated osteoclast formation and subsequently prevented wear particle-
induced bone erosion. Taken together, the results suggest that HP inhibits RANKL-mediated
osteoclastogenesis via affecting ERK signalling in vitro and suppresses wear particle-induced osteolysis
in vivo. We therefore conclude that HP may be an innovative and safe alternative treatment for osteoclast-related
prosthetic loosening.
� 2014 Elsevier Inc. All rights reserved.
Contents lists available at ScienceDirect
Biochemical Pharmacology
jo u rn al h om epag e: ww w.els evier .c o m/lo cat e/bio c hem p har m
1. Introduction
Total joint arthroplasty (TJA) is believed to be the majortreatment for severe joint diseases such as osteoarthritis andrheumatoid arthritis [1]. Every year, approximately 1.5 millioncases of TJA are carried out globally [2]. Great efforts have beenmade towards optimizing the quality and effectiveness of TJA.However, its long-term success is limited due to TJA failure such asaseptic prosthetic loosening [3]. Understanding these causes offailure in TJA and further conquer them in revision total hiparthroplasty procedures is essential for improving long-termpatient outcomes [3]. Aseptic prosthetic loosening occurs second-ary to a cascade of chronic inflammatory and osteoclast-mediated
* Corresponding authors.
E-mail addresses: [email protected] (A. Qin), [email protected] (K. Dai).1 These authors contributed equally to this work.
http://dx.doi.org/10.1016/j.bcp.2014.06.009
0006-2952/� 2014 Elsevier Inc. All rights reserved.
bone resorption in response to wear particles near the prostheses[4–7]. Wear particles are debris (polymeric, metallic, or ceramic)from joint replacements. They can recruit and activate osteoclasts(bone-resorbing cells) to the local site that subsequently showsprosthetic loosening. Given the fact that increased osteoclasticbone resorption plays a vital role in wear particle-inducedosteolysis, osteoclast is one of therapeutic targets towardsalleviating and even treating aseptic loosening after total jointreplacement.
Natural plant-derived compounds have increasingly attractedresearch interest due to their potential pharmacological activity inhuman diseases [8–10]. Hypericin (HP), a naphtodianthrone(Fig. 1A), is one of the major active components of St John’s Wort.It is available as an ‘‘over-the-counter’’ drug. Patients withmalignant diseases undergoing radiation therapy often use it forself-medication [11]. It is also shown to be clinically effective intreating mild to moderate forms of depression [12] and HIV

Fig. 1. Hypericin (HP) inhibited RANKL-induced osteoclast formation and bone resorption in a concentration-dependent manner without any cytotoxicity. (A) The structure of
HP. (B) BMMs were treated with various concentrations of HP followed by M-CSF (30 ng/mL) and RANKL (50 ng/mL) stimulation for five days. Cells were then fixed with 4%
PFA and subjected to TRAP staining. (C) TRAP-positive multinuclear cells and the area of them were counted as described in the methods. (D) BMMs were plated in 96-well
plates with 8000 cells/well and stimulated with M-CSF (30 ng/mL) and RANKL (50 ng/mL) in the presence of the indicated HP concentrations. Cell viability was then measured
using CCK-8 assays, as described in the Methods section, at 48, 72, and 96 h. (E) RAW264.7 cells were treated with various concentrations of HP followed by RANKL (50 ng/mL)
stimulation for five days. Cells were then fixed with 4% PFA and subjected to TRAP staining. (F) TRAP-positive multinuclear cells of RAW264.7 cells the areas of them were
counted as described in the methods. (G) RAW264.7 cells were plated in 96-well plates with 8000 cells/well and stimulated with RANKL (50 ng/mL) in the presence of the
indicated HP concentrations. Cell viability was then measured using CCK-8 assays, as described in the Methods section, at 48, 72, and 96 h. All experiments were carried out at
least three times, and the significance was determined as indicated in methods (*p < 0.05 versus the control).
Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287 277

Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287278
infection [13]. In particular, several phosphorylation of signallingpathways, such as extracellular signal-regulated kinases (ERK),nuclear factor kappaB (NF-kB) and protein kinase C (PKC), could besuppressed by HP in cultured cells [14–17].
Though the clinical usage of isolated HP has been wellstudied [18,19], its impact on bone biology, especially osteoclastbiology, has yet to be examined. It is generally known that theformation of osteoclasts is mainly depending on two key factors,macrophage colony-stimulating factor (M-CSF) and receptoractivator of nuclear factor kappa B ligand (RANKL) [20]. M-CSFplays an import part in osteoclast precursor cell proliferation[21], while the key factor for osteoclast differentiation is RANKL.Upon stimulation with RANKL, its receptor RANK binds to thesignalling adaptor molecule tumour necrosis factor (TNF)receptor-associated factor 6 (TRAF6) [22] and activates thetransforming growth factor (TGF) b-activated kinase-1 (TAK1)kinase complex, thereby resulting in the activation of down-stream signalling pathway such as nuclear factor kappaB (NF-kB) and mitogen-activated protein kinases (MAPK) includingp38, c-Jun N-terminal kinase (JNK) and extracellular signal-regulated kinases 1/2 (ERK 1/2) [23–25]. The activation of thesesignalling molecules induces transcription factors such as c-fos[26], activator protein 1 (AP1) and nuclear factor of activated T-cells, cytoplasmic 1 (NFATc1) [27], which are essential forosteoclast differentiation. Given the importance of NF-kB andMAPK signalling pathway in osteoclast formation, the effects ofHP on ERK and NF-kB suppression in other cell types, and thewide and safe clinical application of HP, we hypothesized thatHP may represent a novel treatment for osteoclast-relatedosteolytic diseases, thus prevent particle-induced peri-implantosteolysis. Therefore, in this study, we examined: (1) the effectsof HP on osteoclastogenesis, (2) the underlying mechanismsmediating the effects, and (3) and its effect in the treatment ofwear particle-induced peri-implant osteolysis.
2. Materials and methods
2.1. Cells, media, and reagents
RAW264.7 cells were purchased from the American TypeCulture Collection (Rockville, MD, USA); the Alpha modificationof Eagle medium (a-MEM), penicillin/streptomycin and foetalbovine serum (FBS) were purchased from Gibco-BRL (Gaithers-burg, MD, USA); and the cell counting kit (CCK-8) was obtainedfrom Dojindo Molecular Technology (Kumamoto, Japan). Re-combinant soluble mouse M-CSF and mouse RANKL wereobtained from R&D (R&D Systems, Minneapolis, MN, USA),and hypericin (HP) was purchased from Sigma Aldrich (St Louis,MO, USA), dissolved in dimethyl sulfoxide (DMSO) which waspurchased from Sigma Aldrich (St Louis, MO, USA) and stored at�20 8C and the experiments have been carried out in theabsence of visible light to prevent photosensitivity. The HP wasadded to cell culture medium such that the DMSO comprised<0.1% of the volume of the culture medium. Specific antibodiesagainst extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), p38, Nuclear factor of kappa lightpolypeptide gene enhancer in B-cells inhibitor alpha (IkBa),phospho-ERK (Thr202/Tyr204), phospho-JNK (Thr183/Tyr185),phospho-p38 (Thr180/Tyr182), c-fos, nuclear factor of activatedT–cells, cytoplasmic 1 (NFATc1), and glyceraldehyde 3-phos-phate dehydrogenase (GAPDH) were obtained from Cell Signal-ing Technology (Cambridge, MA, USA). The tartrate-resistantacid phosphatase (TRAP) staining kit and all other reagents werepurchased from Sigma Aldrich (St Louis, MO, USA), unless statedotherwise.
2.2. Mouse bone marrow macrophage preparation and osteoclast
differentiation
Male, four- to six-week-old C57BL/6 mice were used in thisstudy. Primary bone marrow macrophages (BMMs) were isolatedfrom the whole bone marrow, as described previously [8,10,28,29].Briefly, cells were isolated from the femoral and tibial bonemarrow and cultured in a T75 flask in a-minimum essentialmedium (a-MEM) supplemented with 10% FBS, 1% penicillin/streptomycin, and 10 ng/mL macrophage colony-stimulatingfactor (M-CSF) for 24 h. Non-adherent cells were then removed,and the adherent cells were cultured in a 37 8C, 5% CO2 incubatorfor another 3–4 days until cells were fully confluent. The BMMswere then seeded into a 96-well plate at a density of 8 � 103 cells/well in triplicate and in complete a-MEM supplemented with30 ng/mL M-CSF, 50 ng/mL RANKL, and different concentrations ofHP (0, 0.3, 0.6, 1.2, or 2.5 mM). Cell culture media were replacedevery 2 days until mature osteoclasts had formed. Cells were thenwashed twice with phosphate-buffered saline (PBS), fixed with 4%paraformaldehyde for 20 min, and stained for TRAP using theDiagnostic Acid Phosphatase kit. TRAP-positive cells with morethan three nuclei were counted under a microscope. Similarly, theeffect of HP on osteoclast differentiation was also tested on theRAW264.7 cell line.
2.3. Cell viability assay
The cytotoxic effects of HP were determined using a CCK-8 assayaccording to the manufacturer’s instructions. BMMs or RAW264.7cell were plated in 96-well plates at a density of 8 � 103 cells/well intriplicate, and cultured in complete a-MEM supplemented with30 ng/mL M-CSF for 24 h. Cells were then treated with differentconcentrations of HP (0, 0.3, 0.6, 1.2, 2.5, 5, 10, 20, 40 or 80 mM) for 2,3, or 4 days, respectively. Ten microliters of CCK-8 buffer was addedto each well, and cells were incubated at 37 8C for an additional 2 h.The absorbance was then measured at a wavelength of 450 nm(650 nm reference) on an ELX800 absorbance microplate reader(Bio-Tek, USA). Cell viability was calculated relative to the controlusing the following formula: (experimental group optimal density(OD) � zeroing OD)/(control group OD � zeroing OD).
2.4. Bone absorption assay
BMMs were seeded at a density of 2.4 � 104 cells/cm2 ontobovine bone slices with three replicates. Forty-eight hours later,cells were treated with 50 ng/mL RANKL, 30 ng/mL M-CSF, and 0,0.3, 0.6, or 1.2 mM HP until mature osteoclasts formed. Cells thathad adhered to bone slices were then removed by mechanicalagitation and sonication. Resorption pits were visualized under ascanning electron microscope (SEM, FEI Quanta 250), and the boneresorption area was quantified using Image J software (NationalInstitutes of Health, Bethesda, MD, USA).
2.5. F-actin ring immunofluorescence
For F-actin ring immunofluorescent staining, HP treatedosteoclasts cultured on bovine bone discs were fixed with 4%paraformaldehyde for 15 min and permeabilized for 5 min with0.1% (v/v) Triton X-100 (Sigma Aldrich, St Louis, MO, USA). Cellswere incubated with Alexa-Fluor 647 phalloidin (Invitrogen, SanDiego, CA, USA) diluted in 0.2% (w/v) BSA-PBS (Invitrogen, SanDiego, CA, USA) for 1 h, washed extensively with 0.2% (w/v) BSA-PBS and PBS, and finally mounted with ProLong Gold anti-fademounting medium (Invitrogen, San Diego, CA, USA). Detection offluorescence was carried out on the NIKON A1Si spectral detectorconfocal system (Nikion, Tokyo, Japan) equipped with 406(oil)

Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287 279
lenses. Fluorescence images were collected using the systems NIS-C Elements software and analyzed using Image J.
2.6. RNA extraction and quantitative PCR assay
Quantitative PCR was used to measure specific gene expressionduring osteoclast formation. BMMs were seeded in 24-well plates ata density of 10 � 104 cells/well and cultured in complete a-MEMsupplemented with 30 ng/mL M-CSF and 50 ng/mL RANKL. RANKLinduced osteoclastogenesis from mouse BMMs and was adminis-tered with either different doses of HP (0, 0.3, 0.6, or 1.2 mM) for 5days or 1.2 mM HP for 0–5 days. Total RNA was extracted using theQiagen RNeasy Mini kit (Qiagen, Valencia, CA, USA) following themanufacturer’s instructions, and cDNA was synthesized from 1 mgof total RNA using reverse transcriptase (TaKaRa Biotechnology,Otsu, Japan). Real-time PCR was performed using the SYBR PremixEx Tag kit (TaKaRa, Biotechnology, Otsu, Japan) and an ABI 7500Sequencing Detection System (Applied Biosystems, Foster City, CA,USA). The following cycling conditions were used: 40 cycles ofdenaturation at 95 8C for 5 s and amplification at 60 8C for 24 s.GAPDH was used as the housekeeping gene, and all reactions wererun in triplicate. The mouse primer sequences of GAPDH, CathepsinK, Calcitonin receptor (CTR), TRAP, V-ATPase d2, c-fos, and NFATc1were as follows:
Cathepsin K forward 50-CTTCCAATACGTGCAGCAGA-30 andreverse 50-TCTTCAGGGCTTTCTCGTTC-30;CTR forward 50-TGCAGACAACTCTTGGTTGG-30 and reverse 50-TCGGTTTCTTCTCCTCTGGA-30;TRAP forward 50-CTGGAGTGCACGATGCCAGCGACA-30 and re-verse 50-TCCGTGCTCGGCGATGGACCAGA-30;V-ATPase d2 forward 50-AAGCCTTTGTTTGACGCTGT-30 andreverse 50-TTCGATGCCTCTGTGAGATG-30;c-fos forward 50-CCAGTCAAGAGCATCAGCAA-30 and reverse 50-AAGTAGTGCAGCCCGGAGTA-30;NFATc1 forward 50-CCGTTGCTTCCAGAAAATAACA-30 and re-verse 50-TGTGGGATGTGAACTCGGAA-30;GAPDH forward 50-ACCCAGAAGACTGTGGATGG-30 and reverse50-CACATTGGGGGTAGGAACAC-30.
2.7. Western blotting
For examining the signalling pathway in which HP affect,RAW264.7 cells were seeded in 6-well plates at a density of5 � 105 cells/well. When the cells were confluent, they were pre-treated with or without 5 mM HP for 4 h. Cells were thenstimulated with 50 ng/mL RANKL for 0, 5, 15, or 30 min. Fordetermining the effect of HP with different concentrations on ERKsignalling pathway, RAW264.7 cells were seeded in 6-well platesat a density of 5 � 105 cells/well. When the cells were confluent,they were pre-treated with or without 0, 1, 5 mM HP for 4 h. Cellswere then stimulated with 50 ng/mL RANKL for 0, 5 min. Totalproteins were extracted from cultured cells using radioimmuno-precipitation assay (RIPA) lysis buffer (Sigma Aldrich, St Louis, MO,USA) containing 50 mM Tris–HCl, 150 mM NaCl, 5 mM EDTA, 1%Triton X-100, 1 mM sodium fluoride, 1 mM sodium vanadate, 1%deoxycholate, and protease inhibitor cocktail. Lysates werecentrifuged at 12,000 � g for 15 min, and the supernatants thatcontained the proteins were collected.
Protein concentrations were determined using a bicinchoninicacid (BCA, Thermo Fisher, MA, USA) assay. Thirty micrograms ofeach protein lysate was resolved using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE, Sigma Aldrich, StLouis, MO, USA) on 10% gels and transferred to polyvinylidenedifluoride membranes (Millipore, Bedford, MA, USA). Following
transfer, membranes were blocked with 5% skimmed milk in Tris-buffered saline-Tween (TBS-Tween; 0.05 M Tris, 0.15 M NaCl pH 7.5,and 0.2% Tween-20; Invitrogen, San Diego, CA, USA) for 1 h, and thenincubated with primary antibodies diluted in 1% (w/v) skimmedmilk powder in TBS-Tween overnight at 4 8C. Membranes were thenwashed and incubated with the appropriate secondary antibodiesconjugated with IRDye 800CW (molecular weight 1162 Da).Antibody reactivity was detected by exposure in an Odysseyinfrared imaging system (LI-COR, Nebraska, USA). Quantitativeanalysis of the band intensity was carried out by image J software.
2.8. Luciferase reporter gene activity assay
The effects of HP on RANKL-induced nuclear factor kappaB (NF-kB) activation were measured using RAW264.7 cells that had beenstably transfected with an NF-kB luciferase reporter construct, aspreviously described [30,31]. Briefly, cells were seeded into 48-well plates with three replicates and maintained in cell culturemedia for 24 h. Cells were then pretreated with or without theindicated concentrations of HP for 1 h, followed by addition ofRANKL (50 ng/mL) for 8 h. Luciferase activity was measured usingthe Promega Luciferase Assay System (Promega, Madison, WI, USA)and normalized to that of the vehicle control. Similarly, the effect ofHP on RANKL-induced NFATc1-dependent luciferase reporterassays was determined as described previously [32,33].
2.9. Titanium particle-induced calvarial osteolysis model
We next established a mouse calvarial osteolysis model todetermine the preventative effects of HP on osteolysis in vivo asdescribed previously [34]. All experimental procedures wereperformed in accordance with the Guide for the Care and Use ofLaboratory Animals as adopted and promulgated by the UnitedStates National Institutes of Health [35] and the Guide of the AnimalCare Committee of Shanghai Jiao Tong University. Briefly, male, 20healthy 8-week-old C57BL/J6 mice were assigned randomly to fourgroups: sham PBS control (sham), Ti particles with PBS (vehicle), andTi particles with low (HP-low) and high (HP-high) concentrations ofHP. The mice were anesthetized, and the cranial periosteum wasseparated from the calvarium by sharp dissection. Then, 30 mg of Tiparticles were embedded under the periosteum at the middle sutureof the calvaria [29]. Mice in the HP-low and HP-high groups wereinjected intraperitoneally with HP at 5 or 15 mg/kg/day, respective-ly, for 10 days. Mice in the sham and vehicle groups received PBSdaily. At the end of the experiment, the mice were sacrificed, and thecalvaria were excised and fixed in 4% paraformaldehyde (SigmaAldrich, St Louis, MO, USA) for micro-CT analysis.
2.10. Micro-CT scanning
The fixed calvaria were analyzed using a high-resolution micro-CT (Skyscan 1072; Skyscan, Aartselaar, Belgium). The scanningprotocol was set at an isometric resolution at 9 mm, and X-rayenergy settings of 80 kV and 80 mA. After reconstruction, a squareregion of interest (ROI) around the midline suture was selected forfurther qualitative and quantitative analysis. Bone volume/tissuevolume (BV/TV), the number of porosity, and percentage of totalporosity of each sample were measured, as reported previously [7].
2.11. Histological and histomorphometric analysis
After micro-CT scanning, the calvaria samples were decalcifiedin 10% EDTA (Sigma Aldrich, St Louis, MO, USA) for 3 weeks andthen embedded in paraffin. Histological sections were prepared forTRAP and H&E staining. The specimens were then examined andphotographed under a high quality microscope. The number of

Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287280
TRAP-positive multinucleated osteoclasts normalized with bonearea (OcN/BA) and percentage osteoclast area per bone area (OcA/BA, %) were assessed in each sample.
2.12. Statistical analysis
The data were expressed as means � standard deviation (SD).Experiments were conducted separately at least 3 times. Results wereanalyzed using the SPSS 13.0 software (SPSS Inc., USA). The analysis ofvariance (ANOVA) test, followed by Dunnett post hoc test was used tomake comparisons between groups. p < 0.05 indicated a significantdifference between groups.
3. Results
3.1. Hypericin inhibited RANKL-induced osteoclast differentiation
without cytotoxicity
To determine the effect of HP on osteoclast differentiation, wetreated primary BMMs with RANKL and M-CSF in the presence of
Fig. 2. Hypericin (HP) inhibited osteoclast bone resorption and F-actin ring structure. (A
(50 ng/mL) for three days. Later, cells were cultured in the presence of the indicated con
Scanning electron microscope (SEM) images of bone resorption pits are shown. (B) Reso
monocytes/macrophages (BMMs) were treated with various concentrations of HP follow
then fixed with 4% PFA and stained for the formation of F-actin ring with Alexa-Fluor 64
Number of actin ring was counted. All experiments were performed at least three times
control).
various concentrations of HP. In the control group, BMMsdifferentiated into mature TRAP-positive multinucleated osteo-clasts. However, HP significantly inhibited the formation of TRAP-positive multinucleated osteoclasts in a dose-dependent manner(Fig. 1B and C). The number of osteoclast was 15.17 � 3.54 per wellwith 1.2 mM HP, 56.91 � 7.22 per well with 0.6 mM HP, both of whichwere significantly lower than control group (p < 0.05). To exclude thepossibility that the inhibitory effect of HP on osteoclast differentiationis not due to the cytotoxic effect, cell viability the osteoclast precursorcells were tested. We showed that there are no cytotoxic effects of HPat the same doses that exert inhibitory effects on osteoclastdifferentiation (Fig. 1D). The inhibitory effect of HP on osteoclasto-genesis is further confirmed in RAW264.7 cell line (Fig. 1E–G)
3.2. Hypericin inhibited the bone-resorbing activity of mature
osteoclasts in vitro
Due to the impaired osteoclast differentiation by HP, wehypothesized that osteoclastic bone resorption would also beinhibited. To this end, BMMs were cultured on bone slices without
) BMM-derived pre-osteoclasts were stimulated with M-CSF (30 ng/mL) and RANKL
centrations of HP with M-CSF (30 ng/mL) and RANKL (50 ng/mL) for another 48 h.
rption pit areas were measured using Image J software. (C) Bone marrow-derived
ed by M-CSF (30 ng/mL) and RANKL (50 ng/mL) stimulation for five days. Cells were
7 phalloidin and imaged on NIKON confocal system with 406 oil emersion lens. (D)
, and the significance was determined as indicated in methods (*p < 0.05 versus the
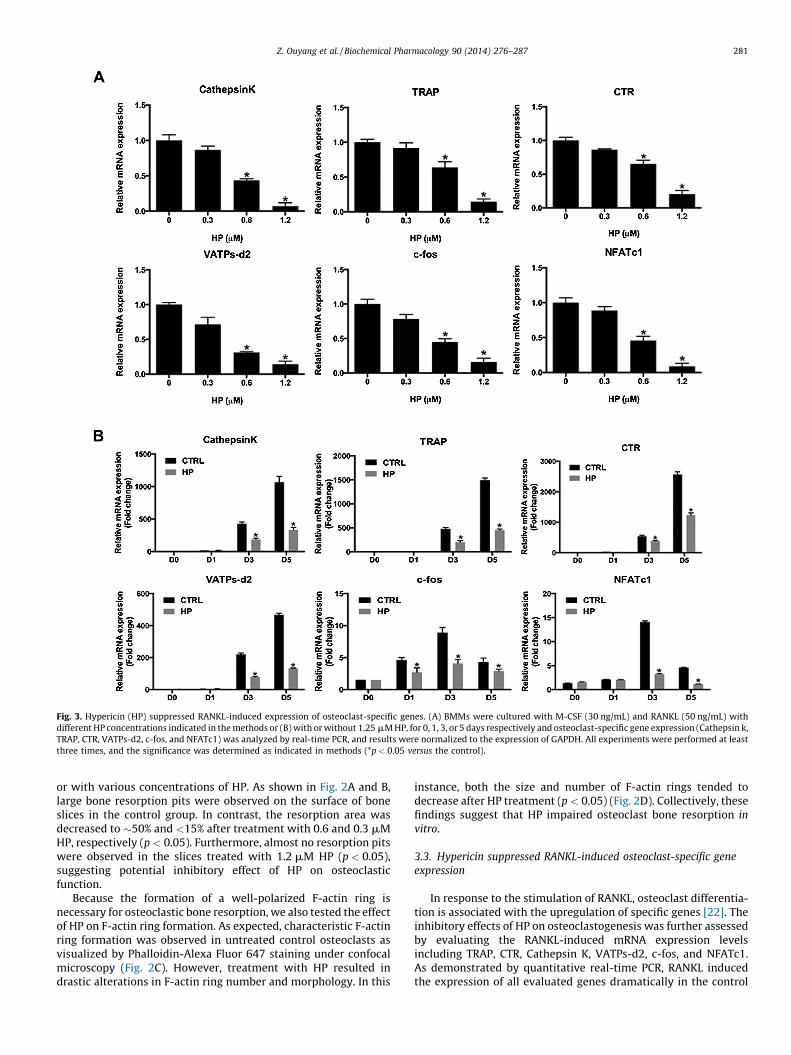
Fig. 3. Hypericin (HP) suppressed RANKL-induced expression of osteoclast-specific genes. (A) BMMs were cultured with M-CSF (30 ng/mL) and RANKL (50 ng/mL) with
different HP concentrations indicated in the methods or (B) with or without 1.25 mM HP, for 0, 1, 3, or 5 days respectively and osteoclast-specific gene expression (Cathepsin k,
TRAP, CTR, VATPs-d2, c-fos, and NFATc1) was analyzed by real-time PCR, and results were normalized to the expression of GAPDH. All experiments were performed at least
three times, and the significance was determined as indicated in methods (*p < 0.05 versus the control).
Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287 281
or with various concentrations of HP. As shown in Fig. 2A and B,large bone resorption pits were observed on the surface of boneslices in the control group. In contrast, the resorption area wasdecreased to �50% and <15% after treatment with 0.6 and 0.3 mMHP, respectively (p < 0.05). Furthermore, almost no resorption pitswere observed in the slices treated with 1.2 mM HP (p < 0.05),suggesting potential inhibitory effect of HP on osteoclasticfunction.
Because the formation of a well-polarized F-actin ring isnecessary for osteoclastic bone resorption, we also tested the effectof HP on F-actin ring formation. As expected, characteristic F-actinring formation was observed in untreated control osteoclasts asvisualized by Phalloidin-Alexa Fluor 647 staining under confocalmicroscopy (Fig. 2C). However, treatment with HP resulted indrastic alterations in F-actin ring number and morphology. In this
instance, both the size and number of F-actin rings tended todecrease after HP treatment (p < 0.05) (Fig. 2D). Collectively, thesefindings suggest that HP impaired osteoclast bone resorption in
vitro.
3.3. Hypericin suppressed RANKL-induced osteoclast-specific gene
expression
In response to the stimulation of RANKL, osteoclast differentia-tion is associated with the upregulation of specific genes [22]. Theinhibitory effects of HP on osteoclastogenesis was further assessedby evaluating the RANKL-induced mRNA expression levelsincluding TRAP, CTR, Cathepsin K, VATPs-d2, c-fos, and NFATc1.As demonstrated by quantitative real-time PCR, RANKL inducedthe expression of all evaluated genes dramatically in the control

Fig. 4. Hypericin (HP) specifically impaired the RANKL-induced ERK signalling. (A) RAW264.7 cells were pretreated with or without 5 mM HP for 4 h followed by 50 ng/mL
RANKL as indicated. Cell lysates were then analyzed using Western blotting with specific antibodies against phospho-JNK, JNK, phosphor-P38, P38, phosphor-ERK, ERK,
phospho-IkBa, IkBa, and GAPDH. (B) Average ratio of phospho-JNK, JNK, phosphor-P38, P38, phosphor-ERK, ERK, phospho-IkBa, IkBa relative to GAPDH. The bands’
intensity analysis was quantified by image J software. Each ratio was normalized to 0 min (*p < 0.05 versus the control). (C) RAW264.7 cells were pretreated with 0, 1, and
5 mM HP for 4 h followed by 0 or 50 ng/mL RANKL. Cell lysates were then analyzed using Western blotting with specific antibodies against phosphor-ERK, ERK. (D) Average
ratio of phosphor-ERK relative to ERK. The bands’ intensity analysis was quantified by image J software. Each ratio was normalized to 0 min (**p < 0.01 versus the group with
RANKL(+), HP(0 mM)). (E) Bone marrow-derived macrophages (BMMs) were cultured with 30 ng/mL M-CSF and 50 ng/mL RANKL with or without 1.2 mM HP for 0, 1, or 3
days. Cell lysates were then analyzed using Western blotting with antibodies against NFATc1, c-fos, and GAPDH. (F) Average ratio of NFATc1, c-fos relative to GAPDH. The
bands’ intensity analysis was quantified by image J software. Each ratio was normalized to day 0 (*p < 0.05 versus the control group). (G) RAW264.7 cells that had been stably
transfected with a NF-kB luciferase reporter construct were pretreated with the indicated concentrations of HP for 1 h, and then incubated in the absence or presence of
RANKL for 8 h. Luciferase activity was then determined using the Promega Luciferase Assay System. RAW264.7 cells that had been stably transfected with a NFATc1 luciferase
reporter construct were pretreated with the indicated concentrations of HP for 1 h and then incubated in the absence or presence of RANKL for 12 h. Luciferase activity was
then determined using the Promega Luciferase Assay System. All experiments were performed at least three times, and the significance was determined as indicated in
methods (*p < 0.05 versus the group with RANKL and HP (0 mM)).
Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287282
group. In contrast, HP substantially suppressed osteoclastic geneexpression in a time- and dose-dependent manner (p < 0.05)(Fig. 3A and B). Therefore, these data further confirmed that HPsuppressed osteoclast formation and gene expression in vitro.
3.4. Hypericin suppressing ERK signalling pathway during
osteoclastogenesis
Next, we used Western blotting and luciferase assays toinvestigate the main signalling pathways (NF-kB, MAPK) that areassociated with osteoclast differentiation. Previous researcheshave revealed that three major subfamilies of MAPKs (p38, ERK1/2,and JNK) play pivotal roles in the development of osteoclasts [36].In addition, the activation of the NF-kB pathway is crucial forosteoclast formation [23,24]. Thus, we investigated whether these
signalling pathways were involved in the suppression of osteo-clastogenesis by HP. We found that HP almost completelyattenuated ERK phosphorylation. In contrast, HP had no obviouseffect on p38 and JNK activation. In addition, in the control groupwith the RANKL’s stimulation, IkBa was degraded within 5 minand this trend was rescued in the following 15–30 min, which wasconsistent with previous study. In HP group, the process wassimilar to the control group, which indicated that the NF-kBsignalling pathway remains intact after HP treatment (Fig. 4A).Quantitative analysis confirmed these observations (p < 0.05)(Fig. 4B).
To further confirm the inhibitory effect of HP on ERK signallingpathway, we treated RAW264.7 cell with HP in differentconcentrations and found that HP indeed attenuated ERKsignalling pathway in a dose-dependent manner (Fig. 4C and D).

Fig. 5. Hypericin (HP) prevented titanium particle-induced mouse calvarial osteolysis. (A) Representative micro-computed tomography (CT) three-dimensional
reconstructed images from each group. (B) Bone volume/tissue volume (BV/TV), number of porosity, and the percentage of total porosity of each sample were measured. The
significance was determined as indicated in methods (*p < 0.05 versus vehicle group).
Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287 283
In addition to initial signalling activation upon RANKLsimulation, c-fos and NFATc1 induction are important for initiatingosteoclast differentiation, specifically, c-fos was regulated by ERKand is important for NFATc1 induction [26]. Thus we nextexamined the effect of HP on both c-fos and NFATc1 expression.As expected, Western blot analysis also showed that RANKL-induced expression of c-fos and NFATc1 was significantlydiminished by HP (Fig. 4E). This observation was also confirmedby quantitative analysis (p < 0.05) (Fig. 4F). More, luciferasereporter assay further supported that NFATc1 activation wasattenuated by HP and NF-kB activation was not (Fig. 4G). Takentogether, these data suggested that HP inhibited osteoclastogen-esis via the ERK/c-fos/NFATc1 pathway without affecting the NF-kB, p38, and JNK signalling pathways.
3.5. Administration of hypericin prevented wear particle-induced
bone loss in vivo
We further explored the protective effects of HP on pathologicalosteolysis. To achieve this, the effect of HP on titanium (Ti)-particle-induced osteolysis was examined in a murine calvarialmodel. We found that extensive bone resorption was observed inthe vehicle group by micro-CT. In the groups with HP treatment,reduced particle-induced osteolysis was observed in a dose-dependent manner, where bone resorption in mice treated with ahigh concentration was much lower than that with the lowconcentration (Fig. 5A). BV/TV, number of porosity, and thepercentage of total porosity in the ROI were measured from three-dimensional reconstruction images, as previously reported [7,37].Compared with the sham group, a significant osteolysis in thecalvaria was induced by the presence of Ti particles (p < 0.05).When HP was injected at 5 mg/kg and 10 mg/kg daily, osteolyticbone loss was prevented in different level compared with thevehicle group (p < 0.05) (Fig. 5B).
The histological and histomorphometric analysis furtherconfirmed the protection effect of HP on wear particle-inducedbone erosion (Fig. 6A). Multinucleated osteoclasts as well as theinflammatory infiltration of lymphocytes and macrophages wereinduced by the presence of Ti particles. TRAP staining revealed thatmultiple osteoclasts lined along the eroded bone surface in thevehicle group (arrow). The erosion surface was also reduced in theHP treatment groups, with the number of osteoclasts normalizedwith bone area and percentage osteoclast area per bone areadecreased in both groups (HP-low and HP-high) (p < 0.05)(Fig. 6B), which also indicates that HP inhibited osteoclastformation.
4. Discussion
TJA is an effective and commonly used treatment for severejoint diseases worldwide. However, periprosthetic osteolysis andsubsequent aseptic loosening remain the major causes ofarthroplasty failure [1,4,5]. Osteoclasts, which play an importantrole in the pathogenesis of diseases such as osteoporosis,rheumatoid arthritis, multiple myeloma, and metastatic cancers,are considered to be the logical and viable target for noveltherapeutic agents to treat wear particle-induced osteolysis [6,38].In the last two decades, much advancement has been made in thetreatment of osteoclast-related disease, such as oestrogen-replacement therapy [39], bisphosphonates (BPs), PTH, anddenosumab [40]; however, treatment options for aseptic pros-thetic loosening are few and far from ideal. Moreover, these drugsare also accompanied by adverse effects such as onset of breastcancer, stroke, heart attack, blood clots [41,42], osteonecrosis ofthe jaw (ONJ), and atypical fractures [40], and are therefore notindicated for long-term treatment [43]; additionally, they may notbe suitable as oral medication. As a result, further attempts todevelop better treatment options ought to be pursued.

Fig. 6. Assessment of titanium particle-induced mouse calvarial osteolysis prevention by hypericin (HP) using histological and histomorphometric analysis. (A) Haematoxylin
and eosin and tartrate-resistant acid phosphatase (TRAP) staining were performed for at least three sections per group. The sham section exhibited few inflammatory and
osteolytic changes. The vehicle group revealed an obvious inflammatory reaction and prominent osteolysis, whereas the HP-treated groups exhibited reduced inflammation
and osteolysis. The arrows points to TRAP positive osteoclasts. (B) The number of TRAP positive cells normalized with bone area (OcN/BA) and percentage osteoclast area per
bone area (OcA/BA, %) were counted. The significance was determined as indicated in methods (*p < 0.05 versus vehicle group).
Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287284
Natural plant-derived products have been recently reported toexert inhibitory effects on osteoclast differentiation and function,leading to decreased bone loss in vivo [8] [10]. Examples from thecurrent literature include sanguinarine [8] (an active ingredientderived from the roots of Sanguinaria canadensis) and androgra-pholide [10], (a diterpenoid lactone isolated from the traditionalChinese medicinal plant Andrographis paniculata).
Hypericum perforatum, also known as St. John’s Wort (SJW), isa popular herb, known for its antidepressant effects [44].Hypericin (HP), as the presumed active moiety within St. John’sWort, exhibits potent pharmacological effects that include anti-inflammatory [45], antibacterial [46], and antitumor activity [47].Additionally, it exhibits light-dependent antiretroviral activityagainst HIV infection [13] and photodynamic therapy (PDT) thatcauses oxidative damage to the target cells [48]. In particular, HPdemonstrated an inhibitory effect on several signalling pathwayssuch as ERK and NF-kB [14,15], which are also important forosteoclast differentiation and which enable us to hypothesize thatHP may also be able to inhibit osteoclast differentiation by thesemechanisms. Recently, Sorensen et al. reported that HP could actas a potent protein kinase C (PKC) inhibitor to inhibit osteoclasticacid secretion and bone resorption [49], which further support ourhypothesis. However, whether HP is a specific PKC inhibitor is notclear at present, [50] and its effect may not be strong enough for ananalysis of the relationship between PKC and osteoclast.Moreover, the effects of HP on RANKL and M-CSF functions, vitalto osteoclast differentiation, are not clarified. Thus, we have
focussed on its effects on osteoclast differentiation and subse-quently, function.
In our study, we provided evidence for the inhibitory effect ofHP on the differentiation of osteoclasts in vitro and prevention ofwear particle-induced bone loss in vivo. We also found that HPtreatment inhibited the expression of the genes related toosteoclast differentiation and function and therefore blocked thebone resorption activity of osteoclasts. Furthermore, our datasuggest that HP could attenuate osteoclastogenesis through itsability to suppress ERK activation.
In osteoclast differentiation of mammalian cells, RANKL bindsto its receptor RANK in the osteoclast precursor cell and activatesseveral key molecules including ERK, JNK, and p38, which consistthe MAPK signalling pathway. They phosphorylate serine andthreonine residues in response to extracellular stimuli andtransmit the stimuli from the cell surface to the nucleus [22].Previous studies reported that p38 regulates the microphthalmia-associated transcription factor that is important for osteoclastdifferentiation [51]. In addition, RANKL-induced osteoclastogen-esis could be prevented by JNK inactivation [52]. ERK, on the otherhand, induces c-fos for osteoclastogenesis [26]. The formation ofosteoclasts could be reduced by inhibition of ERK [53]. Further-more, upon stimulation with RANKL, RANK binds to the signallingadaptor molecule TRAF6 and activates the TAK1 complex, resultingin activation of the NF-kB pathway, and it is reported that the p50/p52 NF-kB double-knockout mouse exhibited severe osteopetro-sis; this indicates that NF-kB signalling could play a pivotal role in

Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287 285
osteoclast development [54,55]. The nuclear factor of kappa lightpolypeptide gene enhancer in the B-cells inhibitor, alpha (IkB), isattached to NF-kB preventing it from migrating into the nucleus,and in osteoclast differentiation phosphorylation with IkB kinase(IKK), it separates the two proteins, and IkBa is degraded. In ourresearch, the effect of HP on the NF-kB signalling pathway was notobserved as HP had no effect on the degradation of IkBa which isfurther confirmed by the luciferase assay. Consistent with previousstudies that report that HP could inhibit ERK signalling pathway inother cells [14], in our study, we found that HP specifically down-regulated ERK activation in osteoclast differentiation, evidenced bylittle ERK phosphorylation after HP treatment. This observation isfurther supported by the dose-dependent manner in which HPattenuated ERK activation. Additionally, the lower expression of c-fos in osteoclast precursors also support our finding becauseprevious studies have demonstrated that the transcription factor c-fos was regulated by ERK and is important for NFATc1 induction[26]. Thus, the suppressed expression of c-fos subsequently leadsto suppressed NFATc1 expression and nucleus translocation,resulting in failed osteoclast formation. Interestingly, phosphor-ylation of p38 and JNK were also examined and showed that HPhad no obvious effect on these signalling pathways. Takentogether, these results revealed that HP specifically inhibitedERK/c-fos/NFATc1 activation during osteoclast differentiationwithout affecting the p38, JNK and NF-kB pathways.
NFATc1 and c-fos are major regulators of osteoclastogenesis,which conducts the expression of osteoclast specific genes such asTRAP, CTR, Cathepsin K, VATPs-d2, all of which also participate inthe osteoclastic bone resorption process [22,56,57]. Here, we foundthat HP treatment inhibited the expression of these genes as wellas NFATc1 and c-fos, and therefore blocked the osteoclastdifferentiation and bone resorption activity. Additionally, inhibi-tion of NFATc1 and c-fos expression by HP further served asevidence of the mRNA level for the results from the Western blotassay.
We further examined the in vivo effect of HP on a wear particle-induced mouse calvarial osteolysis model. Both micro-CT andhistological and TRAP staining assessments demonstrated that HPsignificantly prevented Ti particle-induced osteolysis. As expected,the inflammatory reaction, bone destruction, and thus boneerosion area associated with particle-induced osteoclastogenesisand osteolysis were attenuated by HP treatment (Figs. 5 and 6).Collectively, these data demonstrate, for the first time, that HP canbe effectively used for the treatment of wear particle-inducedosteolysis.
The associated side effects of clinically used drugs constitute animportant drawback. In contrast, HP has a favourable tolerabilityand safety profile [58], which was further confirmed in our mousemodel by the absence of visible side effects. It is reported that HPpresents a low bioavailability in human plasma due to its poorwater solubility, and its elimination half-life (6–48.2 h) andconcentration in plasma (1.3–91 ng/mL) can vary widely [59].However, the dose of HP used (0.015 mg) in our study was largelylower than that required to induce antidepressant (1.80–2.7 mg/die of total HP) [44], analgesic, or antihyperalgesic effects (0.96 mgof total HP) [11], and also was not capable of producing significantclinical interactions [60]. Thus, we can suppose that the HP efficacyin the treatment of osteolysis in the animal model is not secondaryto its antidepressant or analgesic property and would be extremelysafe in humans.
This study, however, has limitations. As we focussed on theimpact of Ti particles in the local environment, there have alsobeen reports on systemic side effects of metal ions. For example,particle contamination with endotoxins might be responsible forproduction of IL-1b and thus cause some side-effects [61]. Themetal particles released into the tissues and circulation might also
contribute to proliferation and differentiation of T-lymphocytes toproduce excessive inflammatory cytokines, such as IL-1, IL-6, andTNF-a [62,63] ˙ These cytokines will trigger a series of systemicimmune responses, and in turn may contribute to osteoclastdifferentiation. However, we have taken a series of measures toremove adherent endotoxins as much as possible, according toprevious studies [64], and we did not observe any side-effects thatmight be triggered by the Ti particle. Further detailed investiga-tions on viscera are encouraged. In addition, Liu et al. mentioned inhis study limitations that 30 mg of Ti particles used in the animalmodel was higher than that in the human patient. For generatingthe murine calvarial osteolysis model, 15 mg [65], 20 mg [66], or30 mg [29,67] were previously reported. However, in patients,continuous particles will be liberated from the surface of theprosthesis. The concentration of particles retrieved from peripros-thetic tissues is about 0.1 mg/mL [64,68], which was thusobviously lower than in animal models. This difference was notedby several researchers and has been issued by von Knoch [69]. Inthe animal model, the particles were implanted in a single bolusrather than being continuously generated. Such an animal modelfocussed on the local bone erosion caused by the particles, but noton their accumulation mechanism. Therefore, it might be the mostreasonable animal model that simulates the particle-inducedosteolysis in patients, and continuous administration of lowerconcentration of Ti for the animal model might be better for futurestudies, but it is unrealistic at present.
In the present study, we demonstrated the inhibitory effects ofHP on osteoclastogenesis, which were achieved mainly throughinhibition of the ERK signalling pathway and downstreamtranscription factor including NFATc1 and c-fos. Furthermore,the in vivo efficacy of HP was confirmed with an animal model ofwear-particle induced osteolysis. We can therefore conclude thatHP, in a low dose, as in our study, represents an innovative and safetherapeutic alternative for the treatment of prosthetic loosening.
Conflicts of interest
None of the authors have any conflicts of interest to declare.
Acknowledgments
This study was supported by a major basic research grant fromthe Science and Technology Commission of Shanghai Municipality(Grant No. 11DJ1400303), a scientific research grant from theNational Natural Science Foundation for the Youth of China (GrantNo. 81201364), a Scientific Research Grant for Youth of Shanghai(Grant No. ZZjdyx 2097), a scientific research grant from 985project – Stem Cell and Regenerative Medicine Centre, and anInnovative Research Grant from the Shanghai Municipal EducationCommission (Grant No. 13YZ031) and Doctoral InnovationFoundation of Shanghai Jiaotong University School of Medicine(BXJ201330). The funding sources had no role in the study design,collection, analysis, data interpretation, or the writing of thereport.
References
[1] Harris WH. Wear and periprosthetic osteolysis: the problem. Clin Orthop RelatRes 2001;6:6–70.
[2] Teeny SM, York SC, Mesko JW, Rea RE. Long-term follow-up care recommen-dations after total hip and knee arthroplasty: results of the American Associa-tion of Hip and Knee Surgeons’ member survey. J Arthroplasty 2003;18:954–62.
[3] Bozic KJ, Kurtz SM, Lau E, Ong K, Vail TP, Berry DJ. The epidemiology ofrevision total hip arthroplasty in the United States. J Bone Joint Surg Am2009;91:128–33.

Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287286
[4] Greenfield EM, Bi Y, Ragab AA, Goldberg VM, Van De Motter RR. The role ofosteoclast differentiation in aseptic loosening. J Orthop Res: Off Publ OrthopRes Soc 2002;20:1–8.
[5] Holt G, Murnaghan C, Reilly J, Meek RM. The biology of aseptic osteolysis. ClinOrthop Rel Res 2007;460:240–52.
[6] Purdue PE, Koulouvaris P, Potter HG, Nestor BJ, Sculco TP. The cellular andmolecular biology of periprosthetic osteolysis. Clin Orthop Rel Res2007;454:251–61.
[7] Wedemeyer C, Xu J, Neuerburg C, Landgraeber S, Malyar NM, von Knoch F, et al.Particle-induced osteolysis in three-dimensional micro-computed tomogra-phy. Calcif Tissue Int 2007;81:394–402.
[8] Li H, Zhai Z, Liu G, Tang T, Lin Z, Zheng M, et al. Sanguinarine inhibits osteoclastformation and bone resorption via suppressing RANKL-induced activation ofNF-kappaB and ERK signaling pathways. Biochem Biophys Res Commun2013;430:951–6.
[9] Kim JY, Cheon YH, Oh HM, Rho MC, Erkhembaatar M, Kim MS, et al. Oleanolicacid acetate inhibits osteoclast differentiation by downregulatingPLCgamma2-Ca(2+)-NFATc1 signaling, and suppresses bone loss in mice. Bone2014;60:104–11.
[10] Zhai ZJ, Li HW, Liu GW, Qu XH, Tian B, Yan W, et al. Andrographolidesuppresses RANKL-induced osteoclastogenesis in vitro and prevents inflam-matory bone loss in vivo. Br J Pharmacol 2014;171:663–75.
[11] Galeotti N, Vivoli E, Bilia AR, Bergonzi MC, Bartolini A, Ghelardini C. Aprolonged protein kinase C-mediated, opioid-related antinociceptive effectof St John’s Wort in mice. J Pain: Off J Am Pain Soc 2010;11:149–59.
[12] Barnes J, Anderson LA, Phillipson JD. St John’s Wort (Hypericum perforatum L.):a review of its chemistry, pharmacology and clinical properties. J PharmPharmacol 2001;53:583–600.
[13] Piscitelli SC, Burstein AH, Chaitt D, Alfaro RM, Falloon J. Indinavir concentra-tions and St John’s Wort. Lancet 2000;355:547–8.
[14] Higuchi A, Yamada H, Yamada E, Jo N, Matsumura M. Hypericin inhibitspathological retinal neovascularization in a mouse model of oxygen-inducedretinopathy. Mol Vis 2008;14:249–54.
[15] Lavie G, Mandel M, Hazan S, Barliya T, Blank M, Grunbaum A, et al. Anti-angiogenic activities of hypericin in vivo: potential for ophthalmologic appli-cations. Angiogenesis 2005;8:35–42.
[16] Zhang W, Lawa RE, Hintona DR, Su Y, Couldwell WT. Growth inhibition andapoptosis in human neuroblastoma SK-N-SH cells induced by hypericin, apotent inhibitor of protein kinase C. Cancer Lett 1995;96:31–5.
[17] Galeotti N, Ghelardini C. St John’s Wort reversal of meningeal nociception: anatural therapeutic perspective for migraine pain. Phytomed: Int J PhytotherPhytopharmacol 2013;20:930–8.
[18] Liu CD, Kwan D, Saxton RE, McFadden DW. Hypericin and photodynamictherapy decreases human pancreatic cancer in vitro and in vivo. J Surg Res2000;93:137–43.
[19] Jacobson JM, Feinman L, Liebes L, Ostrow N, Koslowski V, Tobia A, et al.Pharmacokinetics, safety, and antiviral effects of hypericin, a derivative ofSt John’s Wort plant, in patients with chronic hepatitis C virus infection.Antimicrob Agents Chemother 2001;45:517–24.
[20] Novack DV. Role of NF-kappaB in the skeleton. Cell Res 2011;21:169–82.[21] Takahashi N, Udagawa N, Tanaka S, Suda T. Generating murine osteoclasts
from bone marrow. Methods Mol Med 2003;80:129–44.[22] Boyle WJ, Simonet WS, Lacey DL. Osteoclast differentiation and activation.
Nature 2003;423:337–42.[23] Keating SE, Maloney GM, Moran EM, Bowie AG. IRAK-2 participates in multiple
toll-like receptor signaling pathways to NFkappaB via activation of TRAF6ubiquitination. J Biol Chem 2007;282:33435–43.
[24] Takaesu G, Ninomiya-Tsuji J, Kishida S, Li X, Stark GR, Matsumoto K. Interleu-kin-1 (IL-1) receptor-associated kinase leads to activation of TAK1 by inducingTAB2 translocation in the IL-1 signaling pathway. Mol Cell Biol 2001;21:2475–84.
[25] Pearson G, Robinson F, Beers Gibson T, Xu BE, Karandikar M, Berman K, et al.Mitogen-activated protein (MAP) kinase pathways: regulation and physiolog-ical functions. Endocr Rev 2001;22:153–83.
[26] Monje P, Hernandez-Losa J, Lyons RJ, Castellone MD, Gutkind JS. Regulation ofthe transcriptional activity of c-Fos by ERK A novel role for the prolylisomerase PIN1. J Biol Chem 2005;280:35081–84.
[27] Takayanagi H, Kim S, Koga T, Nishina H, Isshiki M, Yoshida H, et al. Inductionand activation of the transcription factor NFATc1 (NFAT2) integrate RANKLsignaling in terminal differentiation of osteoclasts. Dev Cell 2002;3:889–901.
[28] Toro EJ, Zuo J, Ostrov DA, Catalfamo D, Bradaschia-Correa V, Arana-Chavez V,et al. Enoxacin directly inhibits osteoclastogenesis without inducing apopto-sis. J Biol Chem 2012;287:17894–904.
[29] Qin A, Cheng TS, Lin Z, Cao L, Chim SM, Pavlos NJ, et al. Prevention ofwear particle-induced osteolysis by a novel V-ATPase inhibitor saliphenylha-lamide through inhibition of osteoclast bone resorption. PLoS ONE2012;7:e34132.
[30] Wang C, Steer JH, Joyce DA, Yip KH, Zheng MH, XU J. 12-O-tetradecanoyl-phorbol-13-acetate (TPA) inhibits osteoclastogenesis by suppressing RANKL-induced NF-kB Activation. J Bone Miner Res 2003;18:2159–68.
[31] Zhai Z, Qu X, Yan W, Li H, Liu G, Liu X, et al. Andrographolide prevents humanbreast cancer-induced osteoclastic bone loss via attenuated RANKL signaling.Breast Cancer Res Treat 2014;1–13.
[32] Ikeda F, Nishimura R, Matsubara T, Tanaka S, Inoue J-i, Reddy SV, et al. Criticalroles of c-Jun signaling in regulation of NFAT family and RANKL-regulatedosteoclast differentiation. J Clin Invest 2004;114:475–84.
[33] Weber TJ, Markillie LM. Regulation of activator protein-1 by 8-iso-prostaglan-din E2 in a thromboxane A2 receptor-dependent and-independent manner.Mol Pharmacol 2003;63:1075–81.
[34] Liu X, Qu X, Wu C, Zhai Z, Tian B, Li H, et al. The effect of enoxacin onosteoclastogenesis and reduction of titanium particle-induced osteolysis viasuppression of JNK signaling pathway. Biomaterials 2014;35:5721–30.
[35] National Research Council Committee for the Update of the Guide for the C,Use of Laboratory A. The National Academies Collection: Reports funded byNational Institutes of Health. Guide for the care and use of laboratory animals.Washington (DC): National Academies Press (US) National Academy ofSciences; 2011.
[36] Stevenson DA, Schwarz EL, Carey JC, Viskochil DH, Hanson H, Bauer S, et al.Bone resorption in syndromes of the Ras/MAPK pathway. Clin Genet2011;80:566–73.
[37] Bouxsein ML, Boyd SK, Christiansen BA, Guldberg RE, Jepsen KJ, Muller R.Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J Bone Miner Res: Off J Am Soc Bone Miner Res2010;25:1468–86.
[38] Waddell J, Pritzker KP, Boynton EL. Increased cytokine secretion in patientswith failed implants compared with patients with primary implants. ClinOrthop Rel Res 2005;170–6.
[39] Cetinkaya MB, Kokcu A, Yanik FF, Basoglu T, Malatyalioglu E, Alper T. Com-parison of the effects of transdermal estrogen, oral estrogen, and oral estro-gen–progestogen therapy on bone mineral density in postmenopausalwomen. J Bone Miner Metab 2002;20:44–8.
[40] Verron E, Bouler JM. Is bisphosphonate therapy compromised by the emer-gence of adverse bone disorders? LID. pii: S1359-6446(13)00275-4,doi:10.1016/j.drudis.2013.08.010.
[41] Colditz GA. Hormone replacement therapy increases the risk of breast cancer.Ann N Y Acad Sci 1997;833:129–36.
[42] Rossouw JE, Anderson GL, Prentice RL, LaCroix AZ, Kooperberg C, Stefanick ML,et al. Risks and benefits of estrogen plus progestin in healthy postmenopausalwomen: principal results from the Women’s Health Initiative randomizedcontrolled trial. JAMA 2002;288:321–33.
[43] Cappuzzo KA, Delafuente JC. Teriparatide for severe osteoporosis. Ann Phar-macother 2004;38:294–302.
[44] Kasper S, Caraci F, Forti B, Drago F, Aguglia E. Efficacy and tolerability ofhypericum extract for the treatment of mild to moderate depression. EurNeuropsychopharmacol: J Eur Coll Neuropsychopharmacol 2010;20:747–65.
[45] Kang BY, Chung SW, Kim TS. Inhibition of interleukin-12 production inlipopolysaccharide-activated mouse macrophages by hpyericin, an activecomponent of Hypericum perforatum. Planta Med 2001;67:364–6.
[46] Fiebich BL, Heinrich M, Langosch JM, Kammerer N, Lieb K. Antibacterialactivity of hyperforin from St John’s Wort. Lancet 1999;354:777.
[47] Schempp CM, Simon-Haarhaus B, Termeer CC, Simon JC. Hypericin photo-induced apoptosis involves the tumor necrosis factor-related apoptosis-in-ducing ligand (TRAIL) and activation of caspase-8. FEBS Lett 2001;493:26–30.
[48] Pajonk F, Scholber J, Fiebich B. Hypericin-an inhibitor of proteasome function.Cancer Chemother Pharmacol 2005;55:439–46.
[49] Sorensen MG, Karsdal MA, Dziegiel MH, Boutin JA, Nosjean O, Henriksen K.Screening of protein kinase inhibitors identifies PKC inhibitors as inhibitors ofosteoclastic acid secretion and bone resorption. BMC Musculoskelet Disord2010;11:250.
[50] Kubin A, Wierrani F, Burner U, Alth G, Grunberger W. Hypericin—the factsabout a controversial agent. Curr Pharm Des 2005;11:233–53.
[51] Matsumoto M, Sudo T, Saito T, Osada H, Tsujimoto M. Involvement of p38mitogen-activated protein kinase signaling pathway in osteoclastogenesismediated by receptor activator of NF-kappa B ligand (RANKL). J Biol Chem2000;275:31155–61.
[52] Ikeda F, Nishimura R, Matsubara T, Tanaka S, Inoue J, Reddy SV, et al. Criticalroles of c-Jun signaling in regulation of NFAT family and RANKL-regulatedosteoclast differentiation. J Clin Invest 2004;114:475–84.
[53] Ang E, Liu Q, Qi M, Liu HG, Yang X, Chen H, et al. Mangiferin attenuatesosteoclastogenesis, bone resorption, and RANKL-induced activation of NF-kappaB and ERK. J Cell Biochem 2011;112:89–97.
[54] Iotsova V, Caamano J, Loy J, Yang Y, Lewin A, Bravo R. Osteopetrosis in micelacking NF-kappaB1 and NF-kappaB2. Nat Med 1997;3:1285–9.
[55] Franzoso G, Carlson L, Xing L, Poljak L, Shores EW, Brown KD, et al. Require-ment for NF-kappaB in osteoclast and B-cell development. Genes Dev1997;11:3482–96.
[56] Crotti TN, Flannery M, Walsh NC, Fleming JD, Goldring SR, McHugh KP. NFATc1regulation of the human beta3 integrin promoter in osteoclast differentiation.Gene 2006;372:92–102.
[57] Yavropoulou MP, Yovos JG. Osteoclastogenesis—current knowledge and futureperspectives. J Musculoskelet Neuronal Interact 2008;8:204–16.
[58] Rahimi R, Nikfar S, Abdollahi M. Efficacy and tolerability of Hypericum perfor-atum in major depressive disorder in comparison with selective serotoninreuptake inhibitors: a meta-analysis. Prog Neuropsychopharmacol Biol Psy-chiatry 2009;33:118–27.
[59] da Conceicao AO, Takser L, Lafond J. Effect of St John’s Wort standardizedextract and hypericin on in vitro placental calcium transport. J Med Food2010;13:934–42.
[60] Madabushi R, Frank B, Drewelow B, Derendorf H, Butterweck V. Hyperforin inSt John’s Wort drug interactions. Eur J Clin Pharmacol 2006;62:225–33.
[61] Bi Y, Seabold JM, Kaar SG, Ragab AA, Goldberg VM, Anderson JM, et al. Adherentendotoxin on orthopedic wear particles stimulates cytokine production and

Z. Ouyang et al. / Biochemical Pharmacology 90 (2014) 276–287 287
osteoclast differentiation. J Bone Miner Res: Off J Am Soc Bone Miner Res2001;16:2082–91.
[62] Hallab NJ, Caicedo M, Finnegan A, Jacobs JJ. Th1 type lymphocyte reactivity tometals in patients with total hip arthroplasty. J Orthop Surg Res 2008;3:6.
[63] Taki N, Tatro JM, Lowe R, Goldberg VM, Greenfield EM. Comparison of the rolesof IL-1, IL-6, and TNFalpha in cell culture and murine models of asepticloosening. Bone 2007;40:1276–83.
[64] Liu F, Zhu Z, Mao Y, Liu M, Tang T, Qiu S. Inhibition of titanium particle-inducedosteoclastogenesis through inactivation of NFATc1 by VIVIT peptide. Bioma-terials 2009;30:1756–62.
[65] Shin DK, Kim MH, Lee SH, Kim TH, Kim SY. Inhibitory effects of luteolin ontitanium particle-induced osteolysis in a mouse model. Acta Biomater2012;8:3524–31.
[66] Tsutsumi R, Hock C, Bechtold CD, Proulx ST, Bukata SV, Ito H, et al. Differentialeffects of biologic versus bisphosphonate inhibition of wear debris-inducedosteolysis assessed by longitudinal micro-CT. J Orthop Res 2008;26:1340–6.
[67] Goater JJ, O’Keefe RJ, Rosier RN, Puzas JE, Schwarz EM. Efficacy of ex vivo OPGgene therapy in preventing wear debris induced osteolysis. J Orthop Res2002;20:169–73.
[68] Jiang Y, Jia T, Gong W, Wooley PH, Yang SY. Titanium particle-challengedosteoblasts promote osteoclastogenesis and osteolysis in a murine model ofperiprosthestic osteolysis. Acta Biomater 2013;9:7564–72.
[69] von Knoch M, Jewison DE, Sibonga JD, Sprecher C, Morrey BF, Loer F, et al. Theeffectiveness of polyethylene versus titanium particles in inducing osteolysisin vivo. J Orthop Res 2004;22:237–43.