ice recrystallisation inhibition by polyols: comparison of molecular and macromolecular inhibitors...
TRANSCRIPT

BiomaterialsScience
PAPER
Cite this: Biomater. Sci., 2013, 1, 478
Received 8th November 2012,Accepted 8th January 2013
DOI: 10.1039/c3bm00194f
www.rsc.org/biomaterialsscience
Ice recrystallisation inhibition by polyols: comparisonof molecular and macromolecular inhibitors and role ofhydrophobic units†
Robert C. Deller,a,b Thomas Congdon,a Mohammed A. Sahid,a Michael Morgan,a
Manu Vatish,c Daniel A. Mitchell,c Rebecca Notmana and Matthew I. Gibson*a
The ability of polyols to act as ice recrystallisation inhibitors (IRI), inspired by antifreeze (glyco)proteins
are studied. Poly(vinyl alcohol), PVA, a known IRI active polymer was compared to a panel of mono and
polysaccharides, with the aim of elucidating why some polyols are active and others show no activity.
When corrected for total hydroxyl concentration all the carbohydrate-based polyols displayed near iden-
tical activity with no significant influence of molecular weight. Conversely, PVAwas several orders of mag-
nitude more active and its activity displays significant dependence on molecular-weight implying that its
mechanism of action is not identical to that of carbohydrates. In a second step, the role of hydrophobicity
was studied and it is observed that monosaccharide IRI activity is enhanced by alkylation. Dye-quenching
assays demonstrated that PVA is able to present a hydrophobic surface without self-aggregation. There-
fore, the ability to present a hydrophobic domain is hypothesised to be essential to obtain high IRI
activity, which has many biotechnological applications.
Introduction
The development of technologies that inhibit or prevent thegrowth of ice is a key challenge facing modern science.1–3 Inparticular, new additives that improve both the long and short-term cryostorage of human cells, tissues and organs arerequired to meet the ever increasing demands of transplan-tation and regenerative therapies.4–7 Traditional cryopreserva-tion strategies rely on vitrification (a process of rapid coolingthat suppresses the formation of ice crystals) that requires veryhigh cooling rates, substantial amounts of cryopreservativeand is unsuitable for large sample volumes.8–11 Furthermorevitrification requires the use of organic solvents at high con-centrations that often have associated cytotoxicity at physio-logically relevant temperatures.12 Such cryoprotectants are alsochallenging to remove post thaw; a process which is incompa-tible with most clinical requirements.13,14 Analysis of the cryo-preservation process, however, shows that extensive cell
damage actually arises during the thawing process wherebysmall ice crystals grow in size in a process known as ice recrys-tallisation (IR), a form of Ostwald ripening.15,16 Antifreeze(glyco)proteins (AF(G)Ps)17 that have arisen by convergent evo-lution in polar fish species18 have been shown to be highlypotent ice recrystallisation inhibitors (IRIs).19 The extractionand purification of AF(G)Ps in commercially viable quantitieshas had limited success and transgenic approaches are com-plicated by the essential glycosylated unit.20,21 The total chem-ical synthesis of AF(G)Ps is also extremely challenging, withfew successful synthetic strategies reported.22,23 Studies havealso indicated AF(G)Ps to be cytotoxic24 and the introductionof non-native proteins may cause significant immunogenicityissues limiting their widespread application.25,26 Furthermore,the use of functionally similar antifreeze proteins (AFPs) forthe cryopreservation of human erythrocytes resulted inincreased cell death at higher AFP concentrations due to thesecondary effect of dynamic ice shaping (DIS)27 (and thermalhysteresis, TH)28 a property also exhibited by AF(G)Ps.Attempts at using AF(G)Ps as cryoprotectants have also failedto yield notable benefit in the cryopreservation of rat cardiactissue.29,30 However marginal benefits were observed with anAF(G)P analogue in improving the cryopreservation of rat isletcells.31
Considering the above, the design of (scalable) syntheticmimics capable of reproducing the desirable IRI propertiesbut without the potential toxicological and immunological
†Electronic supplementary information (ESI) available: This includes fullcharacterisation and synthetic details. See DOI: 10.1039/c3bm00194f
aDepartment of Chemistry, University of Warwick, Gibbet Hill Road, Coventry,
CV4 7AL, UK. E-mail: [email protected]; Fax: +44 (0) 2476 524112bMolecular Organisation and Assembly in Cells (MOAC) Doctoral Training Centre,
University of Warwick, CV4 7AL Coventry, UKcClinical Sciences Research Laboratories, University of Warwick, Clifford Bridge
Road, Coventry, CV2 2DX, UK
478 | Biomater. Sci., 2013, 1, 478–485 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
22
Janu
ary
2013
. Dow
nloa
ded
on 0
8/09
/201
4 16
:42:
36.
View Article OnlineView Journal | View Issue
side effect has attracted significant interest.32 Previous work,in particular that of Ben et al.,22,33–38 has focused on makingsmall glycopeptide mimics that have alternative glycoside link-ages as well as simpler monosaccharide moieties. Theseglycopeptides have definite IRI activity, without unwantedTH/DIS but are still challenging to synthesise and only availablein small quantities.33,39,40 Gibson et al. have shown that somesynthetic polymers can display significant IRI activity which isappealing due to their scalable syntheses and highly tunablestructure.41,42 Poly(vinyl) alcohol (PVA) in particular displayssignificant IRI activity at micromolar concentrations16,41,43 andis strongly molecular-weight dependant. This is remarkableconsidering the lack of structural similarities to native AF(G)Ps.It has also recently been shown that at relatively high con-centrations monosaccharides and disaccharides, which areessentially polyols, also have measurable, if weak, IRI activity.This was tentatively linked to their hydration index;35 thenumber and density of water molecules associated with a sac-charide. It is hypothesised that this ordering of water mole-cules disrupts the quasi-liquid layer (QLL) at the ice–waterinterface preventing the exchange of water molecules andhence ice crystal growth.34 This concept, however, does notexplain the high IRI activity of PVA, and in particular its mole-cular-weight dependence as the density of water moleculesper monomer unit will not vary with chain length. Recentwork has identified that carbohydrate-based surfactants havemeasurable IRI activity below their critical micelle concen-tration and analysis of several AF(G)Ps has shown them all tohave hydrophobic domains. Therefore, an additional contribu-tor to IRI activity would seem to be the repulsion of water mole-cules from an inhibitor bound/associated with the ice–waterinterface.36,44 The aim of this study was to undertake a com-parative investigation of PVA and a panel of carbohydrate-based polyols to determine if their IRI activities are linked bythe same mechanism of action, with a view to gaining agreater understanding of the IRI mechanism. The role ofhydrophobicity/amphiphilicity is also studied through chemi-cal modification of monosaccharides. This will aid in therational design of even more IRI active compounds as poten-tial cryoprotectants.
Results and discussion
To assess IRI activity a modified splat assay was used. Briefly, apoly-nucleated ice wafer made up of ice crystals with diameters<10 μm is annealed at −6 °C for 30 minutes before beingphotographed between crossed polarisers. This method separ-ates nucleation from growth process, the latter being of inter-est here. To assess the degree of recrystallisation (growth) themean largest grain size (MLGS) is measured from three indi-vidual wafers and expressed relative to the crystal size obtainedfrom a PBS control. It is important to note this measurementis slightly different from measuring the average grain area,which inevitably gives lower percentage values (due to
decreases in size scaling with radius2), but has the advantageof measuring all samples.37
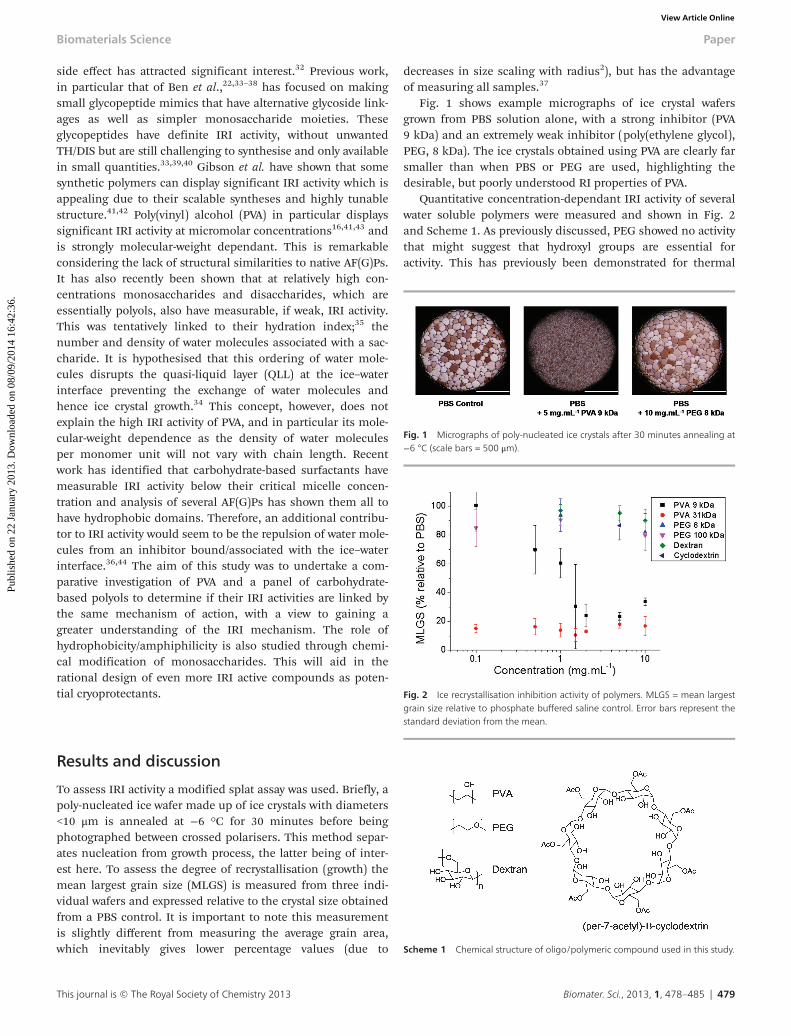
Fig. 1 shows example micrographs of ice crystal wafersgrown from PBS solution alone, with a strong inhibitor (PVA9 kDa) and an extremely weak inhibitor (poly(ethylene glycol),PEG, 8 kDa). The ice crystals obtained using PVA are clearly farsmaller than when PBS or PEG are used, highlighting thedesirable, but poorly understood RI properties of PVA.
Quantitative concentration-dependant IRI activity of severalwater soluble polymers were measured and shown in Fig. 2and Scheme 1. As previously discussed, PEG showed no activitythat might suggest that hydroxyl groups are essential foractivity. This has previously been demonstrated for thermal
Fig. 1 Micrographs of poly-nucleated ice crystals after 30 minutes annealing at−6 °C (scale bars = 500 μm).
Scheme 1 Chemical structure of oligo/polymeric compound used in this study.
Fig. 2 Ice recrystallisation inhibition activity of polymers. MLGS = mean largestgrain size relative to phosphate buffered saline control. Error bars represent thestandard deviation from the mean.
Biomaterials Science Paper
This journal is © The Royal Society of Chemistry 2013 Biomater. Sci., 2013, 1, 478–485 | 479
Publ
ishe
d on
22
Janu
ary
2013
. Dow
nloa
ded
on 0
8/09
/201
4 16
:42:
36.
View Article Online
hysteresis and dynamic ice shaping by AF(G)Ps, but does notprove hydroxyls are an essential feature in all IRI active com-pounds. The glucose-based polysaccharides dextran and acetyl-β-cyclodextrin were also evaluated for IRI but surprisinglyshowed very weak activity compared to PVA in the concentrationrange tested, which implies that presentation of multiplehydroxyl groups does not guarantee that a polymer will havepotent IRI properties. This is in agreement with our earlierwork using synthetic glycopolymers with pendant monosacchar-ides that had weak, but measurable, IRI properties.42
The above data highlights the lack of understanding aboutthe structural features essential for IRI activity and the chal-lenge associated with the design and application of novel RIactive compounds – put simply very few tested materialsdisplay any activity. Previous work has suggested preciselyspaced hydroxyl groups are essential for interactions with theice lattice, and hence RI activity.45,46 However, for carbo-hydrates the hydration index has also been implicatedsuggesting the local ordering of water, not ice-binding, iscrucial. This argument holds for monosaccharides, but failsfor disaccharides which have increased IRI activity, but lower
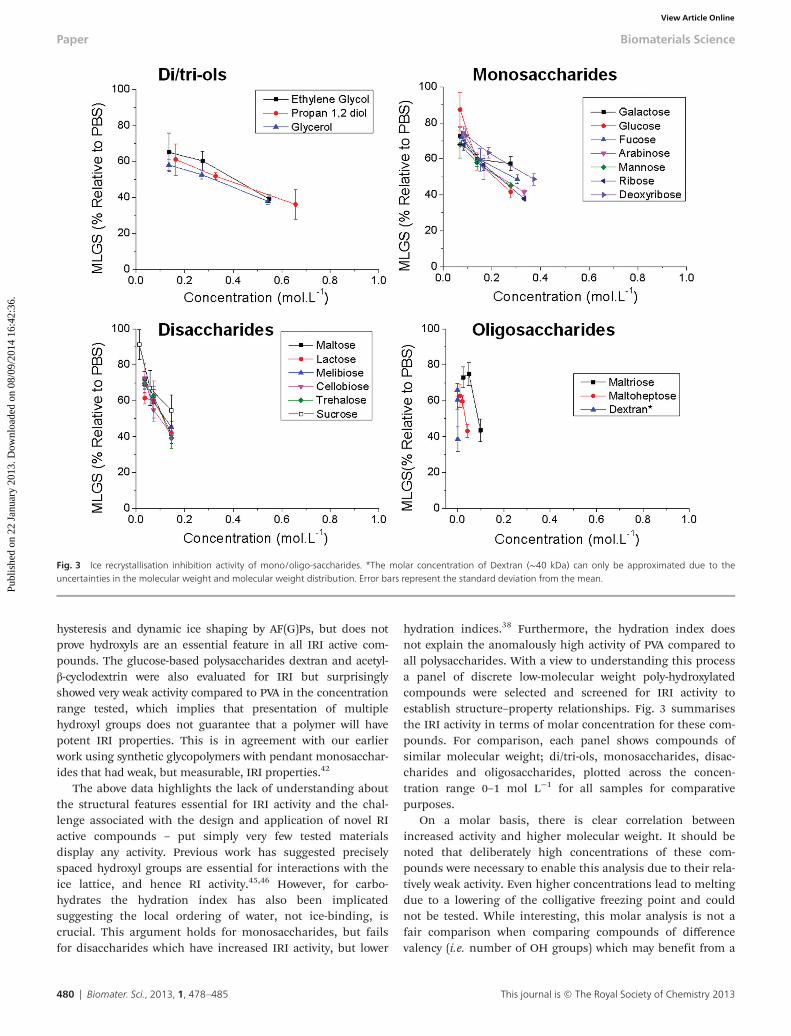
hydration indices.38 Furthermore, the hydration index doesnot explain the anomalously high activity of PVA compared toall polysaccharides. With a view to understanding this processa panel of discrete low-molecular weight poly-hydroxylatedcompounds were selected and screened for IRI activity toestablish structure–property relationships. Fig. 3 summarisesthe IRI activity in terms of molar concentration for these com-pounds. For comparison, each panel shows compounds ofsimilar molecular weight; di/tri-ols, monosaccharides, disac-charides and oligosaccharides, plotted across the concen-tration range 0–1 mol L−1 for all samples for comparativepurposes.
On a molar basis, there is clear correlation betweenincreased activity and higher molecular weight. It should benoted that deliberately high concentrations of these com-pounds were necessary to enable this analysis due to their rela-tively weak activity. Even higher concentrations lead to meltingdue to a lowering of the colligative freezing point and couldnot be tested. While interesting, this molar analysis is not afair comparison when comparing compounds of differencevalency (i.e. number of OH groups) which may benefit from a
Fig. 3 Ice recrystallisation inhibition activity of mono/oligo-saccharides. *The molar concentration of Dextran (∼40 kDa) can only be approximated due to theuncertainties in the molecular weight and molecular weight distribution. Error bars represent the standard deviation from the mean.
Paper Biomaterials Science
480 | Biomater. Sci., 2013, 1, 478–485 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
22
Janu
ary
2013
. Dow
nloa
ded
on 0
8/09
/201
4 16
:42:
36.
View Article Online
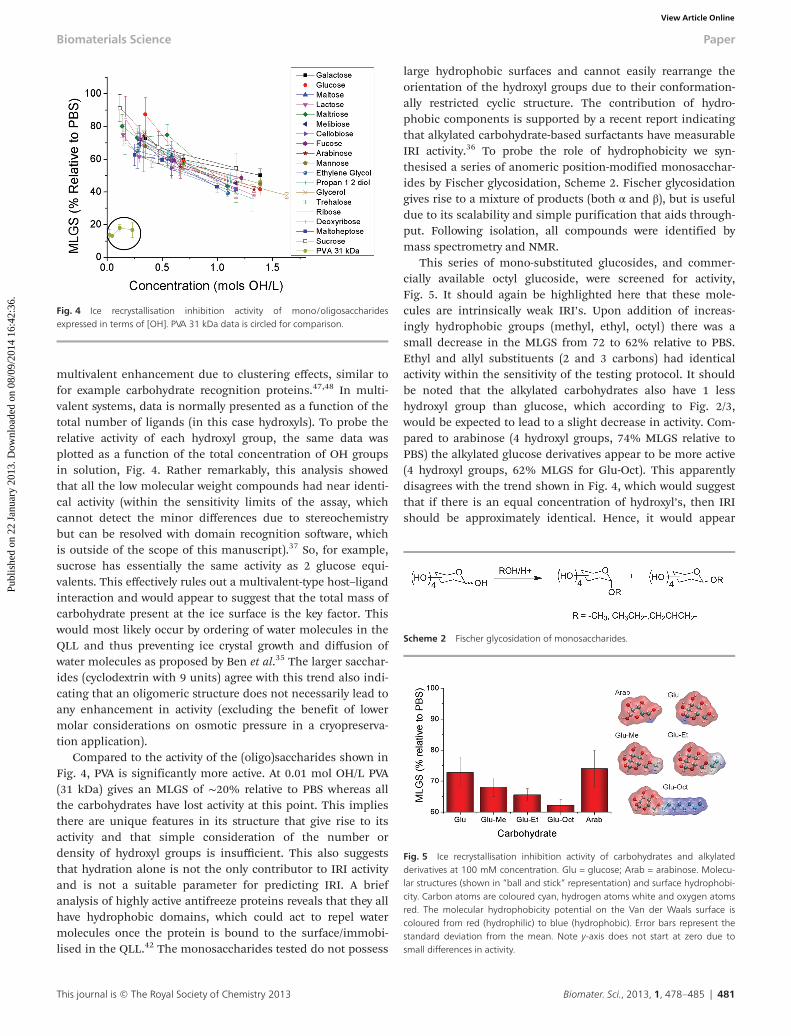
multivalent enhancement due to clustering effects, similar tofor example carbohydrate recognition proteins.47,48 In multi-valent systems, data is normally presented as a function of thetotal number of ligands (in this case hydroxyls). To probe therelative activity of each hydroxyl group, the same data wasplotted as a function of the total concentration of OH groupsin solution, Fig. 4. Rather remarkably, this analysis showedthat all the low molecular weight compounds had near identi-cal activity (within the sensitivity limits of the assay, whichcannot detect the minor differences due to stereochemistrybut can be resolved with domain recognition software, whichis outside of the scope of this manuscript).37 So, for example,sucrose has essentially the same activity as 2 glucose equi-valents. This effectively rules out a multivalent-type host–ligandinteraction and would appear to suggest that the total mass ofcarbohydrate present at the ice surface is the key factor. Thiswould most likely occur by ordering of water molecules in theQLL and thus preventing ice crystal growth and diffusion ofwater molecules as proposed by Ben et al.35 The larger sacchar-ides (cyclodextrin with 9 units) agree with this trend also indi-cating that an oligomeric structure does not necessarily lead toany enhancement in activity (excluding the benefit of lowermolar considerations on osmotic pressure in a cryopreserva-tion application).
Compared to the activity of the (oligo)saccharides shown inFig. 4, PVA is significantly more active. At 0.01 mol OH/L PVA(31 kDa) gives an MLGS of ∼20% relative to PBS whereas allthe carbohydrates have lost activity at this point. This impliesthere are unique features in its structure that give rise to itsactivity and that simple consideration of the number ordensity of hydroxyl groups is insufficient. This also suggeststhat hydration alone is not the only contributor to IRI activityand is not a suitable parameter for predicting IRI. A briefanalysis of highly active antifreeze proteins reveals that they allhave hydrophobic domains, which could act to repel watermolecules once the protein is bound to the surface/immobi-lised in the QLL.42 The monosaccharides tested do not possess
large hydrophobic surfaces and cannot easily rearrange theorientation of the hydroxyl groups due to their conformation-ally restricted cyclic structure. The contribution of hydro-phobic components is supported by a recent report indicatingthat alkylated carbohydrate-based surfactants have measurableIRI activity.36 To probe the role of hydrophobicity we syn-thesised a series of anomeric position-modified monosacchar-ides by Fischer glycosidation, Scheme 2. Fischer glycosidationgives rise to a mixture of products (both α and β), but is usefuldue to its scalability and simple purification that aids through-put. Following isolation, all compounds were identified bymass spectrometry and NMR.
This series of mono-substituted glucosides, and commer-cially available octyl glucoside, were screened for activity,Fig. 5. It should again be highlighted here that these mole-cules are intrinsically weak IRI’s. Upon addition of increas-ingly hydrophobic groups (methyl, ethyl, octyl) there was asmall decrease in the MLGS from 72 to 62% relative to PBS.Ethyl and allyl substituents (2 and 3 carbons) had identicalactivity within the sensitivity of the testing protocol. It shouldbe noted that the alkylated carbohydrates also have 1 lesshydroxyl group than glucose, which according to Fig. 2/3,would be expected to lead to a slight decrease in activity. Com-pared to arabinose (4 hydroxyl groups, 74% MLGS relative toPBS) the alkylated glucose derivatives appear to be more active(4 hydroxyl groups, 62% MLGS for Glu-Oct). This apparentlydisagrees with the trend shown in Fig. 4, which would suggestthat if there is an equal concentration of hydroxyl’s, then IRIshould be approximately identical. Hence, it would appear
Fig. 4 Ice recrystallisation inhibition activity of mono/oligosaccharidesexpressed in terms of [OH]. PVA 31 kDa data is circled for comparison.
Scheme 2 Fischer glycosidation of monosaccharides.
Fig. 5 Ice recrystallisation inhibition activity of carbohydrates and alkylatedderivatives at 100 mM concentration. Glu = glucose; Arab = arabinose. Molecu-lar structures (shown in “ball and stick” representation) and surface hydrophobi-city. Carbon atoms are coloured cyan, hydrogen atoms white and oxygen atomsred. The molecular hydrophobicity potential on the Van der Waals surface iscoloured from red (hydrophilic) to blue (hydrophobic). Error bars represent thestandard deviation from the mean. Note y-axis does not start at zero due tosmall differences in activity.
Biomaterials Science Paper
This journal is © The Royal Society of Chemistry 2013 Biomater. Sci., 2013, 1, 478–485 | 481
Publ
ishe
d on
22
Janu
ary
2013
. Dow
nloa
ded
on 0
8/09
/201
4 16
:42:
36.
View Article Online
that introducing hydrophobicity is a method to attenuate IRIactivity and that there are multiple structural features whichcan contribute to this effect. Representations of the moleculesin terms of hydrophilic/hydrophobic domain are also shown inFig. 5 to illustrate the relative hydrophobic (blue) to hydrophi-lic (red) within each molecule (see Experimental section forcomputational details).
As the data shown in Fig. 5 is not strictly statistically signifi-cant, the concentration dependence on the glucose-derivativeswas measured revealing the same trend of the hydrophobicgroups giving a decrease in the MLGS at any given concen-tration. Similarly, mannose and ethyl-mannose were compared,with the ethyl-derivative showing higher activity than freemannose. These changes are rather small though, and eachindividual value is not statistically significant, but still presentsa useful tool to aid understanding of IRI activity in our opinion.
Taken together the data presented above suggest thatpolyols do have measurable IRI activity and implies that theability to order a large number of water molecules (via hydro-gen bonding, and total number [OH]) does contribute to IRIactivity as previously suggested.35 However, the addition ofhydrophobic residues, at the expense of hydroxyl groups, mayenhance activity and may be the dominant factor in the activityof native proteins. Several AFPs, without any glycosylationsites, actually have charged hydrophilic surfaces49 but doretain a significant hydrophobic domain and therefore wouldnot be expected to function in the same manner as polyols. Alipo-polysaccharide has been identified from Upis ceramboides(Alaskan beetle) with potent antifreeze activity which appearsto require the hydrophobic lipid for activity, which also sup-ports our hypothesis.50
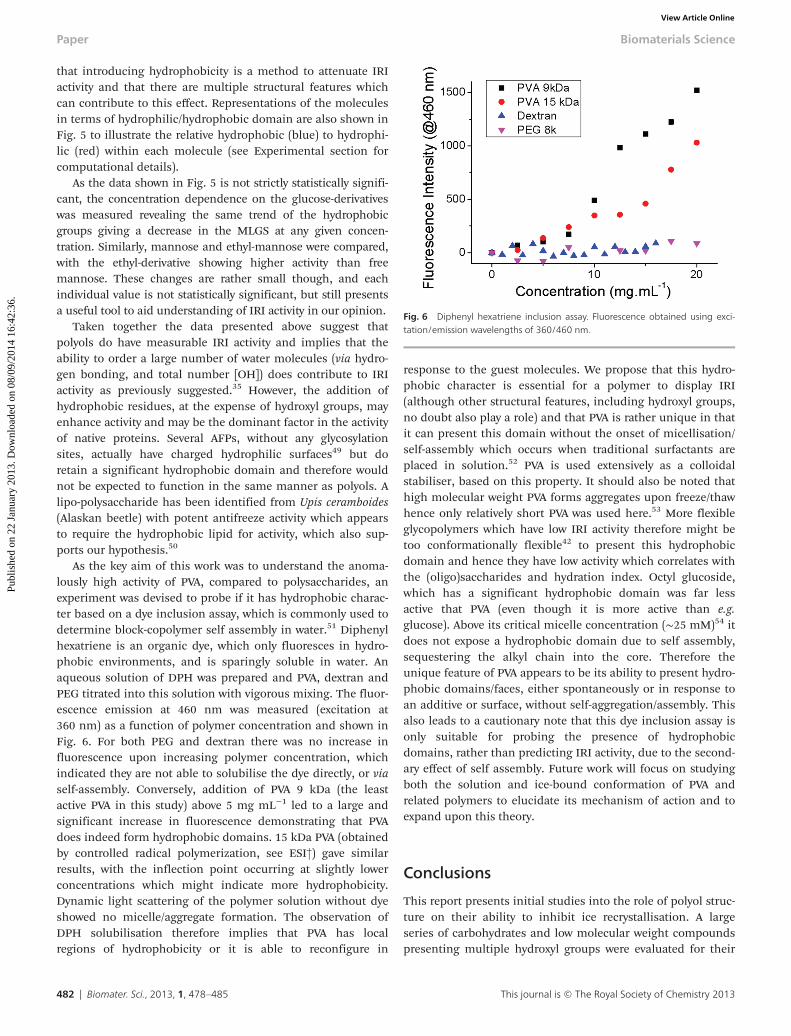
As the key aim of this work was to understand the anoma-lously high activity of PVA, compared to polysaccharides, anexperiment was devised to probe if it has hydrophobic charac-ter based on a dye inclusion assay, which is commonly used todetermine block-copolymer self assembly in water.51 Diphenylhexatriene is an organic dye, which only fluoresces in hydro-phobic environments, and is sparingly soluble in water. Anaqueous solution of DPH was prepared and PVA, dextran andPEG titrated into this solution with vigorous mixing. The fluor-escence emission at 460 nm was measured (excitation at360 nm) as a function of polymer concentration and shown inFig. 6. For both PEG and dextran there was no increase influorescence upon increasing polymer concentration, whichindicated they are not able to solubilise the dye directly, or viaself-assembly. Conversely, addition of PVA 9 kDa (the leastactive PVA in this study) above 5 mg mL−1 led to a large andsignificant increase in fluorescence demonstrating that PVAdoes indeed form hydrophobic domains. 15 kDa PVA (obtainedby controlled radical polymerization, see ESI†) gave similarresults, with the inflection point occurring at slightly lowerconcentrations which might indicate more hydrophobicity.Dynamic light scattering of the polymer solution without dyeshowed no micelle/aggregate formation. The observation ofDPH solubilisation therefore implies that PVA has localregions of hydrophobicity or it is able to reconfigure in
response to the guest molecules. We propose that this hydro-phobic character is essential for a polymer to display IRI(although other structural features, including hydroxyl groups,no doubt also play a role) and that PVA is rather unique in thatit can present this domain without the onset of micellisation/self-assembly which occurs when traditional surfactants areplaced in solution.52 PVA is used extensively as a colloidalstabiliser, based on this property. It should also be noted thathigh molecular weight PVA forms aggregates upon freeze/thawhence only relatively short PVA was used here.53 More flexibleglycopolymers which have low IRI activity therefore might betoo conformationally flexible42 to present this hydrophobicdomain and hence they have low activity which correlates withthe (oligo)saccharides and hydration index. Octyl glucoside,which has a significant hydrophobic domain was far lessactive that PVA (even though it is more active than e.g.glucose). Above its critical micelle concentration (∼25 mM)54 itdoes not expose a hydrophobic domain due to self assembly,sequestering the alkyl chain into the core. Therefore theunique feature of PVA appears to be its ability to present hydro-phobic domains/faces, either spontaneously or in response toan additive or surface, without self-aggregation/assembly. Thisalso leads to a cautionary note that this dye inclusion assay isonly suitable for probing the presence of hydrophobicdomains, rather than predicting IRI activity, due to the second-ary effect of self assembly. Future work will focus on studyingboth the solution and ice-bound conformation of PVA andrelated polymers to elucidate its mechanism of action and toexpand upon this theory.
Conclusions
This report presents initial studies into the role of polyol struc-ture on their ability to inhibit ice recrystallisation. A largeseries of carbohydrates and low molecular weight compoundspresenting multiple hydroxyl groups were evaluated for their
Fig. 6 Diphenyl hexatriene inclusion assay. Fluorescence obtained using exci-tation/emission wavelengths of 360/460 nm.
Paper Biomaterials Science
482 | Biomater. Sci., 2013, 1, 478–485 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
22
Janu
ary
2013
. Dow
nloa
ded
on 0
8/09
/201
4 16
:42:
36.
View Article Online

IRI activity. It was shown that the IRI activity could be corre-lated with the total number of hydroxyl groups and that therewere only small differences between individual carbohydrates.Polysaccharides also followed this trend. Replacement ofanomeric hydroxyl groups with hydrophobic alkyl unitsresulted in an enhancement in activity, even though the totalnumber of hydroxyl groups was decreased showing thatalthough hydration is important, the presentation of hydro-phobic domains is crucial to IRI activity. These results werecompared to that for PVA, which is a highly active IRI polymerwith no obvious hydrophobic regions. Dye incorporationassays show that PVA was capable of solubilising hydrophobicdyes but without the onset of micellisation, as judged by DLS,providing evidence that it can assume a conformation whichpresent hydrophobic surface and therefore a possible mechan-ism of action for IRI. These observations will aid the rationaledesign of new small molecule and macromolecular IRI com-pounds and also shed light on the fundamental mechanismsof actions of native AF(G)Ps that have many potential biotech-nological applications.
Experimental sectionMaterials
Phosphate-buffered saline (PBS) solutions were prepared usingpreformulated tablets (Sigma-Aldrich) in 200 mL of MilliQwater (>18.2 Ω mean resistivity) to give a buffered pH of 7.4.Dextran (40 kDa (obtained from Leuconostoc spp.)), poly(ethyl-ene glycol) (8 kDa, 100 kDa), poly(vinyl alcohol) (9 kDa (80%hydrolyzed), 31 kDa (98–99% hydrolysed)), 1,6-diphenyl-1,3,5-hexatriene, ethylene glycol, propan-1,2-diol, glycerol, galactose,fucose, glucose, octyl glucopyranoside, fructose, arabinose,mannose, ribose, 2-deoxyribose, maltose, sucrose, lactose,melibiose, cellobiose, trehalose, maltotriose, maltoheptoseand acetyl-β-cyclodextrin were purchased from Sigma-Aldrich.All other solvents and reagents were purchased from VWR orSigma-Aldrich and used without further purification. Icewafers were annealed on a Linkam Biological CryostageBCS196 with T95-Linkpad system controller equipped with aLNP95-Liquid nitrogen cooling pump, using liquid nitrogen asthe coolant (Linkam Scientific Instruments UK, Surrey, UK).
Physical and analytical methods1H and 13C NMR spectra were recorded on Bruker DPX-300and DPX-400 spectrometers using deuterated solventsobtained from Sigma-Aldrich. Mass spectral analyses wereobtained using Bruker MicroTOF or Bruker MaXis electrosprayinstruments using positive or negative electrospray mode. Themolecular ion and mass fragments are quoted and assigned.Fluorescence spectra were recorded using a Synergy HT multi-mode microplate reader (BioTek UK, Bedfordshire, UK) andclear, flat bottom 96-well plates. Dynamic light scattering wasconducted using a Nano-Zs from Malvern Instruments, UK.Scattered light was detected at 173° and the observed countrates recorded. Hydrodynamic radii (where appropriate) were
determined using the manufacturer’s software. Polymer stocksolutions were filtered through 0.45 μm filter before analysis.
Graphical representation of surface hydrophobicity
The chemical structures of a selection of molecules (arabinose,glucose, the alkylated glucose derivatives and PVA) were drawnthen energy minimised in vacuum using CHARMM force-fieldparameters. The surface hydrophobicity was calculated usingthe Platinum web server55 wherein a lipophilicity parameter isassigned to each atom and used to map out the molecularhydrophobicity potential on the van der Waals surface. Theimages were generated using Visual Molecular Dynamics.56
Note that these models are simply illustrative and by no meansclaim to represent the global energy minimum structures insolution.
Ice recrystallisation inhibition (splat) assay
Ice recrystallisation inhibition was measured using a modifiedsplay assay.43 A 10 μL sample of polymer dissolved in PBSbuffer (pH 7.4) was dropped 1.40 m onto a chilled glass cover-slip sat on a piece of polished aluminium placed on dry ice.Upon hitting the chilled glass coverslip, a wafer with diameterof approximately 10 mm and thickness 10 μm was formedinstantaneously. The glass coverslip was transferred onto theLinkam cryostage and held at −6 °C under N2 for 30 minutes.Photographs were obtained using an Olympus CX 41 micro-scope with a UIS-2 20×/0.45/∞/0-2/FN22 lens and crossedpolarizers (Olympus Ltd, Southend on sea, UK), equipped witha Canon DSLR 500D digital camera. Images were taken of theinitial wafer (to ensure that a polycrystalline sample had beenobtained) and after 30 minutes. Image processing was con-ducted using Image J, which is freely available.57 In brief, tenof the largest ice crystals were measured and the single largestlength in any axis recorded. This was repeated for at least threewafers and the average (mean) value was calculated to find thelargest grain dimension along any axis. The average of thisvalue from three individual wafers was calculated to give themean largest grain size (MLGS).
Measurement of critical micelle concentration
Critical micelle concentration was determined by fluorescencespectrometry using a Synergy HT multi-mode microplatereader (BioTek UK, Bedfordshire, UK). Aqueous 80 μL aliquotsof 9 kDa PVA, 40 kDa Dextran and 8 kDa PEG ranging from0–50 mg mL−1 and 20 μL of 0.01 mg mL−1 DPH were added toa clear flat-bottom 96-well plate and samples incubated at25 °C for a minimum of 3 minutes. The fluorescence of eachsample was then measured with excitation at 360 nm andemission at 460 nm. The critical micelle concentration wasdefined as the intercept between the two linear regions of thegraphs.
Synthesis of methyl-glucopyranoside
Concentrated hydrochloric acid (200 μL) was added to a stirredsolution of glucose (2.00 g, 11.1 mmol) in methanol (25 mL).The solution was left stirring at room temperature for 18 h,
Biomaterials Science Paper
This journal is © The Royal Society of Chemistry 2013 Biomater. Sci., 2013, 1, 478–485 | 483
Publ
ishe
d on
22
Janu
ary
2013
. Dow
nloa
ded
on 0
8/09
/201
4 16
:42:
36.
View Article Online

after which sodium carbonate (0.3 g, 2.8 mmol) was addedand the solution stirred for 20 min. The solution was filteredand then concentrated in vacuo. The residue was redissolved inmethanol (1 mL) and then precipitated into diethyl ethergiving a white solid and clear supernatant, which was thendecanted after centrifugation. Methyl-glucopyranoside wasobtained as a white solid after drying under a high vacuum for24 h. 1.13 g, 52.7%. 1H-NMR (400 MHz, D2O) 3.40 (s) (CH3O).13C-NMR (D2O, 100 MHz, 298 K) δ 109.0, 103.1, 95.9, 92.0,55.8, 55.2. MS (ESI) m/z calculated for C7H14O6 [M + H]+ 195.08found 195.05.
Synthesis of ethyl-glucopyranoside
The same procedure as for methyl-glucopyranoside was used,using ethanol (25 mL) as the reaction solvent. 0.18 g, 7.8%1H-NMR (400 MHz, D2O) 1.16 (m) (CH3CH2O).
13C-NMR(100 MHz, D2O) δ 107.8, 102.0, 95.9, 92.1, 14.1. MS (ESI) m/zcalculated for C8H16O6 [M + Na]+ 231.08 found 231.08.
Synthesis of allyl-glucopyranoside
The same procedure as for methyl-glucopyranoside was used,using glucose (0.2 g, 1.1 mmol) and allyl alcohol (3 mL) as thereaction solvent. 0.13 g, 78.3%. 1H-NMR (D2O, 400 MHz) δ 5.90(m), (CH2CH). MS (ESI) m/z calculated for C9H16O5 [M + Na]+
243.08 found 243.08.
Acknowledgements
Equipment used was supported by the Innovative Uses forAdvanced Materials in the Modern World (AM2), with supportfrom Advantage West Midlands (AWM) and part funded bythe European Regional Development Fund (ERDF). MIG is aBirmingham Science City Interdisciplinary Research Fellowfunded by the Higher Education Funding Council for England(HEFCE). This work was supported by a Research Project Grantfrom the Leverhulme Trust (RPG-144) and a Research Grantfrom the Royal Society. RD acknowledges the EPSRC for astudentship from the MOAC Doctoral Training Centre. RNacknowledges the Royal Society for a University ResearchFellowship.
Notes and references
1 J. B. Wareing, IEEE, 1997, 438, v3–v35.2 R. Carriveau, A. Edrisy, P. Cadieux and R. Mailloux,
J. Adhes. Sci. Technol., 2012, 26, 447–461.3 C. C. Ryerson, Cold Reg. Sci. Technol., 2011, 65, 97–110.4 R. A. Wolfe, E. C. Roys and R. M. Merion, Am. J. Transplant.,
2010, 10, 961–972.5 A. Atala, J. Tissue Eng. Regen. Med., 2007, 1, 83–96.6 J. Ringe, C. Kaps, G.-R. Burmester and M. Sittinger, Natur-
wissenschaften, 2002, 89, 338–351.7 J. O. M. Karlsson and M. Toner, Biomaterials, 1996, 17,
243–256.
8 C. Polge, A. U. Smith and A. S. Parkes, Nature, 1949, 164,666.
9 A. Rowe, E. Eyster and A. Kellner, Cryobiology, 1968, 5, 119–128.
10 G. M. Fahy, B. Wowk, J. Wu, J. Phan, C. M. Rasch, A. Changand E. Zendejas, Cryobiology, 2004, 48, 157–178.
11 S. Yavin and A. Arav, Theriogenology, 2007, 67, 81–89.12 J. M. Davis, S. D. Rowley, H. G. Braine, S. Piantadosi and
G. W. Santos, Blood, 1990, 75, 781–786.13 K. L. Scott, J. Lecak and J. P. Acker, Transfus. Med. Rev.,
2005, 19, 127–142.14 G. B. Quan, Y. Han, M. X. Liu and F. Gao, Cryobiology,
2009, 59, 258–267.15 P. Mazur, Am. J. Physiol., 1984, C125–C142.16 C. Budke, C. Heggemann, M. Koch, N. Sewald and T. Koop,
J. Phys. Chem. B, 2009, 113, 2865–2873.17 A. L. Devries, S. K. Komatsu and R. E. Feeney, J. Biol.
Chem., 1970, 245, 2901–2908.18 L. Chen, A. L. Devries and C. H. C. Cheng, Proc. Natl. Acad.
Sci. U. S. A., 1997, 94, 3817–3822.19 C. A. Knight, A. L. Devries and L. D. Oolman, Nature, 1984,
308, 295–296.20 Y. Wu, J. Banoub, S. V. Goddard, M. H. Kao and
G. L. Fletcher, Comp. Biochem. Physiol., B: Biochem. Mol.Biol., 2001, 128, 265–273.
21 J. W. Larrick and D. W. Thomas, Curr. Opin. Biotechnol.,2001, 12, 411–418.
22 B. L. Wilkinson, R. S. Stone, C. J. Capicciotti, M. Thaysen-Andersen, J. M. Matthews, N. H. Packer, R. N. Ben andR. J. Payne, Angew. Chem., Int. Ed., 2012, 51, 3606–3610.
23 Y. Tachibana, G. L. Fletcher, N. Fujitani, S. Tsuda,K. Monde and S.-I. I. Nishimura, Angew. Chem., Int. Ed.,2004, 116, 874–880.
24 J. Garner and M. M. Harding, ChemBioChem, 2010, 11,2489–2498.
25 A. S. De Groot and D. W. Scott, Trends Immunol., 2007, 28,482–490.
26 H. Schellekens, Clin. Ther., 2002, 24, 1720–1740.27 H. Chao, P. L. Davies and J. F. Carpenter, J. Exp. Biol., 1996,
199, 2071–2076.28 E. Kristiansen and K. E. Zachariassen, Cryobiology, 2005,
51, 262–280.29 T. Wang, Q. Zhu, X. Yang, J. R. Layne Jr. and A. L. Devries,
Cryobiology, 1994, 31, 185–192.30 J. A. Mugnano, T. Wang, J. R. Layne Jr., A. L. Devries and
R. E. Lee, Am. J. Physiol., 1995, 269, R474–479.31 S. Matsumoto, M. Matsusita, T. Morita, H. Kamachi,
S. Tsukiyama, Y. Furukawa, S. Koshida, Y. Tachibana,S.-I. I. Nishimura and S. Todo, Cryobiology, 2006, 52,90–98.
32 M. I. Gibson, Polym. Chem., 2010, 1, 1141–1152.33 S. Liu and R. N. Ben, Org. Lett., 2005, 7, 2385–2388.34 P. Czechura, R. Y. Tam, E. Dimitrijevic, A. V. Murphy and
R. N. Ben, J. Am. Chem. Soc., 2008, 130, 2928–2929.35 R. Y. Tam, S. S. Ferreira, P. Czechura, J. L. Chaytor and
R. N. Ben, J. Am. Chem. Soc., 2008, 130, 17494–17501.
Paper Biomaterials Science
484 | Biomater. Sci., 2013, 1, 478–485 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
22
Janu
ary
2013
. Dow
nloa
ded
on 0
8/09
/201
4 16
:42:
36.
View Article Online

36 C. J. Capicciotti, M. Leclere, F. A. Perras, D. L. Bryce,H. Paulin, J. Harden, Y. Liu and R. N. Ben, Chem. Sci.,2012, 3, 1408–1416.
37 J. Jackman, M. Noestheden, D. Moffat, J. P. Pezacki,S. Findlay and R. N. Ben, Biochem. Biophys. Res. Commun.,2007, 354, 340–344.
38 J. L. Chaytor, J. M. Tokarew, L. K. Wu, M. Leclere,R. Y. Tam, C. J. Capicciotti, L. Guolla, E. Moos,C. S. Findlay, D. S. Allan and R. N. Ben, Glycobiology, 2012,22, 123–133.
39 C. Heggemann, C. Budke, B. Schomburg, Z. Majer,M. Wissbrock, T. Koop and N. Sewald, Amino Acids, 2010,38, 213–222.
40 L. Nagel, C. Plattner, C. Budke, Z. Majer, A. L. Devries,T. Berkemeier, T. Koop and N. Sewald, Amino Acids, 2011,41, 719–732.
41 T. Inada and S.-S. Lu, Cryst. Growth Des., 2003, 3, 747–752.42 M. I. Gibson, C. A. Barker, S. G. Spain, L. Albertin and
N. R. Cameron, Biomacromolecules, 2009, 10, 328–333.43 C. A. Knight, J. Hallett and A. L. Devries, Cryobiology, 1988,
25, 55–60.44 A. D. J. Haymet, L. G. Ward and M. M. Harding, J. Am.
Chem. Soc., 1999, 121, 941–948.45 C. Budke and T. Koop, ChemPhysChem, 2006, 7, 2601–2606.
46 C. P. Garnham, R. L. Campbell and P. L. Davies, Proc. Natl.Acad. Sci. U. S. A., 2011, 108, 7363–7367.
47 Y. C. Lee and R. T. Lee, Acc. Chem. Res., 1995, 28, 321–327.48 S.-J. Richards, M. W. Jones, M. Hunaban, D. M. Haddleton
and M. I. Gibson, Angew. Chem., Int. Ed., 2012, 51, 7812–7816.
49 A. C. Doxey, M. W. F. Yaish, M. Griffith and B. J. McConkey,Nat. Biotechnol., 2006, 24, 852–855.
50 K. R. Walters, A. S. Serianni, T. Sformo, B. M. Barnes andJ. G. Duman, Proc. Natl. Acad. Sci. U. S. A., 2009, 106,20210–20215.
51 N. Ma, H. Zhang, B. Song, Z. Wang and X. Zhang, Chem.Mater., 2005, 17, 5065–5069.
52 R. Zana, Langmuir, 1996, 12, 1208–1211.53 M. Liu, R. Cheng and R. Qian, J. Polym. Sci., Part B: Polym.
Phys., 1995, 33, 1731–1735.54 K. Shinoda, T. Yamaguchi and R. Hori, Bull. Chem. Soc.
Jpn., 1961, 34, 237–241.55 T. V. Pyrkov, A. O. Chugunov, N. A. Krylov, D. E. Nolde and
R. G. Efremov, Bioinformatics, 2009, 25, 1201–1202.56 W. Humphrey, A. Dalke and K. Schulten, J. Mol. Graphics,
1996, 14, 33–38.57 M. Abràmoff, P. Magalhaes and S. J. Ram, Biophotonics Int.,
2004, 7, 36–44.
Biomaterials Science Paper
This journal is © The Royal Society of Chemistry 2013 Biomater. Sci., 2013, 1, 478–485 | 485
Publ
ishe
d on
22
Janu
ary
2013
. Dow
nloa
ded
on 0
8/09
/201
4 16
:42:
36.
View Article Online