identification and characterization of serine proteinase inhibitors from neospora caninum
TRANSCRIPT

Molecular & Biochemical Parasitology 136 (2004) 101–107
Identification and characterization of serine proteinaseinhibitors fromNeospora caninum
Susana Brunoa, Vilma G. Duschakb, Bibiana Ledesmaa, Marcela Ferellab,c,Björn Anderssonc, Eduardo A. Guarneraa, Sergio O. Angela,d,∗
a Departamento de Parasitolog´ıa, ANLIS Dr. Carlos G. Malbran, Ciudad de Buenos Aires, Buenos Aires, Argentinab Instituto Nacional de Parasitolog´ıa, Dr. Mario Fatala Chabén, Ministerio de Salud, Ciudad de Buenos Aires, Buenos Aires, Argentina
c Center for Genomics and Bioinformatics, Karolinska Institutet, Stockholm, Swedend Laboratorio de Parasitolog´ıa Molecular, UB2, IIB-INTECH, CONICET-UNSAM, Camino de Circunvalación Laguna Km. 6 CC 164,
B7130IWA, Chascomús, Prov. de Buenos Aires, Buenos Aires, Argentina
Received 4 September 2003; received in revised form 9 March 2004; accepted 11 March 2004
Abstract
Two cDNA clones obtained from theNeospora caninumExpressed Sequence Tag project were selected by their homology with theToxoplasma gondiiserine proteinase inhibitor (serpin) gene, TgPI-1 and TgPI-2. One of them, named NcPI-H, showed several premature stopcodons. The other cDNA, named NcPI-S, encoded a 79 amino acid protein containing a putative signal peptide and only one non-classicalKazal domain. Two otherN. caninumEST sequences (NcEST1 and NcEST2) and one fromEimeria tenella(EtPI-S) were retrieved fromthe database. Amino acid sequence analysis suggested that NcEST1 and NcEST2 might be theN. caninumcounterparts of TgPI-1 andTgPI-2, respectively. EtEST-S, as NcPI-S, is a single domain serpin. The open reading frame encoding the mature version of NcPI-S wasexpressed as recombinant protein, fused to a 6 histidine tag inEscherichia coli. Specific rabbit antiserum generated against the recombinantNcPI-S was used in immunoblot assays. Bands of 20, 30, 40, and 66-kDa were detected by SDS-PAGE of whole parasite homogenate. Inaddition, when an anti-TgPI-1 serum was used, bands of 25 and 35-kDa were detected indicating that there is no cross-reactivity betweenboth serpins, and showing as well, the presence of another putative serpin inN. caninum. The recombinant protein NcPI-S, inhibitedbacterial subtilisin completely, and showed lower inhibitory capacity on human neutrophil elastase, animal trypsin, and chymotrypsin,suggesting differences in effectiveness.© 2004 Elsevier B.V. All rights reserved.
Keywords: Neospora caninum; Serine proteinase inhibitor; Kazal
1. Introduction
Neosporaspecies are coccidian parasites, closely relatedto Toxoplasma gondii, Hammondia hammondiandH. hey-dorni [1]. Up to now, two species were detected in the genusNeospora: N. caninumandN. hughesi[1–3]. N. caninumisrecognized as a major cause of abortion in cattle worldwideand it is also responsible for clinical disease in dogs, itsdefinitive host[4]. Neosporainfections were also reportedin horses and sheeps[4].
Serpins are members of a super family of proteinsfrom a wide range of organisms, with a characteristicwell-conserved tertiary structure, that regulates complex
∗ Corresponding author. Tel.:+54-2241-430323;fax: +54-2241-424048.
E-mail address:[email protected] (S.O. Angel).
cellular functions and also includes proteins which lackproteinase inhibitor activity such as chicken ovoalbumin,angiotensinogen, etc.[5,6]. Recently, three serine pro-teinase inhibitors (serpins), belonging to the non-classicalKazal-type family, were characterized inT. gondii. Twoof them, TgPI-1 and TgPI-2, have a four-domain structure[7–10], while the third one, TgTI, has only one domain[11]. Notably, the TgPI-2 nucleotide sequence showed tobe highly similar to TgTI, especially at the N-terminusregion. The divergence is at the protein level, after aminoacid 59, due to single nucleotide differences that alterthe reading frame[10]. TgPI-1 has shown to be a potenttrypsin, chymotrypsin, and neutrophil elastase inhibitor,whereas, the inhibition of tryptase, pancreatic elastase andthrombin was less specific[7–9]. In contrast, TgPI-2 andTgTI have shown to inhibit only trypsin[10,11]. TgPI-1was shown to be a dense granule protein that is releasedin the parasitophore vacuole[8,9]. Despite the fact that
0166-6851/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.molbiopara.2004.03.005

102 S. Bruno et al. / Molecular & Biochemical Parasitology 136 (2004) 101–107
immunolocalization was unsuccessful, TgPI-2 could be lo-cated at the excreted–secreted antigen fraction, suggestingdense granule localization[10].
It was hypothesized that serpins can protect parasites fromproteinase-rich environments during ingesta, modulate thehost inflammatory response, and/or modulate the action ofproteinases secreted by the parasites themselves[5–9]. Inan attempt to find similar proteins, we selected two cDNAs(NcPI-SandNcPI-H) from theNeosporaEST database withsimilarity to TgPI-1. Sequencing showed only one Kazal do-main in NcPI-S protein, whereas, the nucleotide sequenceof another EST (NcPI-H) clone did not show an open read-ing frame, although hypothetical Kazal domains could bedetected by Blastx analysis. In addition, two other EST en-coding putative serpins were found in theN. canimumESTdatabase and one single domain serpin was also identified intheEimeria tenellagenome project. The NcPI-S open read-ing frame (ORF) was expressed inEscherichia coliand itsserine proteinase inhibition activity was studied. In addition,a comparative study was performed between TgPI-1 andNcPI-S at the biochemical and immunocrossreactive levels.
2. Materials and methods
2.1. Source of the cDNA clone, excision of insert andsequencing
cDNAs with high homology to the TgPI proteinwere identified in theNeosporaexpressed sequence tag(EST) database (http://genome.wustl.edu/est/index.php?neospora=1). The cDNAs described here are the cloneBF248641 (NcPI-H) and AI579851 (NcPI-S) obtainedfrom theN. caninumEST project (Genome System, USA).Bluescript plasmids were excised from the lambda ZAPvector using the ExAssist kit (Stratagene). Both strandsof recombinant plasmids were sequenced by using T7-,Sp6-promoter, and internal primers. Gel electrophoresiswas carried out on an ABI 377 Automated DNA Sequencer(Perkin-Elmer) and a MegaBACE 1000 sequencer. Nu-cleotide sequence data ofNcPI-H andNcPI-Sare availablein the GenBankTM database under the accession numberAY368155 and AF179480.
2.2. Host cells and parasite cultures
N. caninumNc-1 strain tachyzoites were grown in hu-man foreskin fibroblasts monolayers, with Eagle’s mini-mum essential media (Gibco) containing 1% fetal calf serum(Gibco). Parasites were purified from infected monolayersby filtration through 3�m polycarbonate filters (Nuclepore).
2.3. Sequence analysis
Database searches and sequence comparisons were per-formed using blastn, blastx, and blast two sequences pro-
grams (www.ncbi.nlm.nih.gov/Blast). Preliminary Toxo-plasmagenomic and/or cDNA (TgESTzy) sequence datawere accessed viahttp://ToxoDB.org and/or http://www.tigr.org/tdb/tgondii/. Other apicomplexan genomic data-base used were:www.plasmodb.org, www.nbi.nlm.nih.gov/projects/malaria/blastindex.html, www.parvum.mic.vcu.eduandwww.sanger.ac.uk. EST databases were also searched byusingwww.ncbi.nlm.nih.gov/Blast. Bioedit and Phrap pro-grams were used for multiple alignment and sequence anal-ysis. BCM search launcher programs (http://searchlauncher.bcm.tmc.edu) were used to find putatives open readingframes. TgPI-1 was described in Pszenny et al.[7] andMorris et al.[9]; TgPI-2 in Morris and Carruthers[10].
Neighbor-joining trees were constructed with win-PAUP4b-10 based on a pairwise comparison of two se-quences according to Kimura’s[12] two-parameter model.Phylogenetic inferences had to be restricted to networkanalysis without outgroup rooting due to the lack of suitableoutgroups.
2.4. Subcloning, expression, and purification of therecombinant proteins
The AF179480 cDNA was amplified with NcPISF(5′ ggatccaaggaaaatgaagatcagggatg 3′) and NcPISK (5′
ggtacctcaggatgggcgacaacttc 3′) primers and the PCR prod-uct was digested withBamHI andKpnI restriction enzymes(underlined in primer sequences), purified from agarose gelby Qiaex II (Qiagen), cloned into the corresponding sites ofthe pQE32 plasmid (Qiagen), and sequenced. The plasmidpQE-NcPI-Sproduced a recombinant protein with a 6-Histag at the N-terminus (rNcPI-S). rNcPI-S corresponds tothe mature version of native NcPI-S protein.
To express only the domains 1 and 2 of TgPI-1 (r-NT), theTgPI-1nucleotide sequence was digested from pQE-TgPI-1[7] with BamHI and SalI. BamHI is the cloning site ofpQE-TgPI-1, whereas,SalI site is in position 711 onTgPI-1 cDNA sequence. The expressed protein presentedonly the domains 1 and 2 complete, whereas, the third isdisrupted by the cloning. To express only the domains 3and 4 of TgPI-1 (r-CT), the region from nucleotide 628to 1135 of TgPI-1 cDNA was amplified with TgPI-1CF(5′ ggatccggatccgaatctgagaacgatgctctg 3′) and TgPI-1CR(5′ ggtaccagaagatcaacgggacagcac 3′) primers and the PCRproduct was digested withBamHI and KpnI restrictionenzymes (underlined in primer sequences), purified fromagarose gel by Qiaex II (Qiagen), cloned into the corre-sponding sites of the pQE plasmid (Qiagen), and sequenced.
Expression and purification under non-denaturing condi-tions were performed as previously described[7].
2.5. Production of anti-rNcPI-S serum and Western blotanalysis
A rabbit was immunized with 0.5 mg of affinity purifiedrNcPI-S emulsified with Freund’s complete adjuvant. Two

S. Bruno et al. / Molecular & Biochemical Parasitology 136 (2004) 101–107 103
subsequent boosters were given at a 2-week intervals with0.2 mg of rNcPI-S emulsified in Freund’s incomplete adju-vant. Serum was collected 1 week after the last booster.
SDS-PAGE on 15 and 20% polyacrylamide gels was per-formed in the Mini–Protean system (Bio-Rad) using stan-dard conditions. After electrophoresis, proteins were trans-ferred to a nitrocellulose membrane (Mini–Protean–Blotsystem, Bio-Rad), and the membrane was blocked with 5%non-fat dried milk powder in PBS-0.5% Tween 20 (block-ing solution). The nitrocellulose filters were probed with thefirst antiserum diluted in blocking solution. A peroxidaseimmunoconjugate (Jackson Immuno Research Laboratories)was used as the secondary antibody (1:5000), and specificbinding was detected by ECL Western blot kit (Amersham).
2.6. Proteinase inhibitory activity of rNcPI-S
Inhibition activity towards different serin proteinaseswas determined by measuring the residual hydrolytic ac-tivity after incubation of each of them with the recombi-nant inhibitory protein rNcPI-S (1.5�g/ml) in comparisonwith rTgPI-1 (1�g/ml) [7], the recombinant amino andcarboxi-terminal halves of TgPI-1, r-NT (1.5�g/ml) andr-CT (3.3�g/ml), respectively, and rROP2 (10�g/ml) [13]used as controls, by chromogenic assays in the condi-tions given below. The respective enzyme were incubatedwith increasing concentrations of each recombinant protein(0.1–5× [E0]) for a specified interval; then, after additionof substrate to the reaction mixture in a final volume of500�l, the release ofp-nitroaniline from the peptidyl paran-itroanilide substrate was monitored spectrophotometricallyat 410 nm for an appropriate length of time. Taking into ac-count the molecular weights of the enzymes and recombinantinhibitors tested in each case molar ratios were calculated.
Porcine trypsin: 8 nM, 50 mM Tris–HCl, 100 mM NaCl,0.25 mM Bz-Pro-Phe Arg-pNA, pH 8.0, 0◦C, 5 min; bovinepancreas chymotrypsin: 8 nM enzyme, 100 mM Tris–HCl,10 mM CaCl2, 0.15 mM Suc-Ala-Ala-Pro-Phe-pNA, pH7.4, 0◦C, 1 min; human tryptase, 5 nM, 10 mM Tris–HCl,200 mM NaCl, Bz-Arg-pNA, pH 6.8, 15 min, 37◦C; bovinethrombin: 5 nM, 50 mM Tris–HCl, 100 mM NaCl, 0.25 mMBz-Phe-Val-Arg-pNA, pH 8.0, 10 min, 37◦C; human neu-trophil elastase: 3 nM enzyme, 50 mM Tris–Hcl, 0.25 mMSuc-Ala-Ala-Ala-pNA, pH 7.4, 10 min, 37◦C; subtilisinfrom Bacillus licheniformis: 3.6 nM, 100 mM Tris–HCl,150 mM NaCl, 10 mM CaCl2, Triton X-100 0.005% v/v,0.15 mM CBZ-Gly-Gly-Leu-pNA, pH 8.0, 10 min, 37◦C.All the enzymes and substrates were purchased to SigmaChemical Co., St. Louis, MO, USA.
3. Results
3.1. Sequence analysis of NcPI-S cDNA
The nucleotide sequence showed thatNcPI-S cDNA is831 bp in length.NcPI-ScDNA showed an ORF encoding
Fig. 1. (A) Structure of NcPI-S protein. Numbers indicate amino acidposition. (B) Alignment of putative signal peptides (SP) of deducedN.caninumandT. gondii serpins. Signal peptide sequence was obtained byusing Psort II program. Black indicates identical residues; grey indicatesconservative residues. Gaps are indicated by dashes and were introducedto improve the alignment.
a 79-amino acid protein (Fig. 1). The N-terminus seems tocontain a signal peptide as defined by the PSORT II programanalysis with a putative cleavage site located after position24 (Fig. 1A), suggesting that NcPI-S, like TgPI-1 and TgPI-2[8–10] is a secreted protein. Thus, the mature protein wouldhave 55 amino acids with a theoretical mass of 6.01 kDaand a pI of 4.60. Neither a transmembrane glycosylation nora glycosylphosphatidylinositol recognition site was found.Blastp analysis using the entire predicted protein sequence ofNcPI-ScDNA showed that NcPI-S is a single non-classicalKazal domain serpin (Figs. 1A and 2B).
3.2. Searching the Apicomplexa genome project databasesand sequence analysis
In order to extend the identification and analysis to re-lated serpins, a complete search in Apicomplexa databasewas carried out. New non-classical Kazal domains serpinswere detected only forN. caninum(NcEST1 and NcEST2)and for E. tenella(EtEST-S). The deducedE. tenellaser-pin (EtEST-S) showed an only single domain. No suchnon-classical Kazal domain related serpins were detectedneither inCryptosporidium parvumnor in Plasmodium spp.database.
To gain a better understanding of the relationship amongserpins, a Blast 2 analysis was carried out. The deducedamino acid sequence of NcEST1 showed high similarity(46% of similarity and 53% of positives) with TgPI-1,whereas, the deduced amino acid sequence of NcEST2showed high similarities with TgPI-2 (68% of similarity and80% of positives) and TgTI (47% of similarity and 60% ofpositives). NcPI-S showed a maximal identity and positivesvalues when compared with NcEST1 deduced protein (40and 50%, respectively). AnotherN. caninumEST (NcPI-H)was sequenced (accesion number: AY368155). Despite

104 S. Bruno et al. / Molecular & Biochemical Parasitology 136 (2004) 101–107
Fig. 2. Domains analysis. (A) Basic structure of Kazal signatureand sequence of Kazal-type inhibitory domains, aligned according tocysteines. The arrowhead indicates P1, reactive site residue. NcPI-Snon-classical Kazal domain, NcEST1, NcEST2, and EtPI-S putativedomains were aligned with otherT. gondii non-classical Kazal do-mains: TgPI-1 (domains 1–4), TgPI-2 (domains 1–4). Sequences wereordered in three separate alignment according a NJ tree results (seeB), to better observe the degree of similarity among them. Gaps areindicated by dashes and were introduced to improve the alignment.(B) NJ tree of the serpin domains. To generate NcEST1, NcEST2,and EtEST-S contigs, sharing sequences were obtained from databaseas follows: NcEST1 = NcEST3c94a05.y2, NcEST3d06h11.y2, andNcEST3b88g06.y1; NcEST2 = NcEST3c68a08.y1, NcESTqab35f08.y1,and NcESTqab49e09.y1; and EtEST-S = etsHEST00405.
Blastx analysis showed similarity withT. gondii Kazalinhibitors, NcPI-H did not show any ORF, presenting pre-mature stop codon in the first hypothetical domain as wellas along the mRNA (data not shown).NcPI-H (1421 bp)and NcEST1(1574 bp) shared 98% of identity only in the
second half of their sequence (nucleotides 656–1404 and808–1544, respectively), whereas, in the first half, theyshowed 63% of similarity inside a short region, suggestingthat they are encoded by different genes. EtPI-S showedidentity with the other serpins just at the domain region.Since the similarity of domains could be favoring some re-lations among serpins, similar studies were done using thepredicted signal peptide sequences (Fig. 1B). The analysisshowed a similar relationship among signal peptide regionthan the observed by using the complete protein.
3.3. Relationship among putative non-classical Kazaldomains
Fig. 2A shows the alignment of the domains of NcPI-S,TgPI-1 (domains 1–4)[7,9] and TgPI-2 (domains 1–4)[10],TgTI [11], as well as those deduced from other serpin cD-NAs of N. caninumandE. tenelladetected in database. Thealignment showed that almost all of them showed the samestructure, except NcEST2.2 which presented two amino acidinsertion, like TgPI-2.2. All deduced domains belong tonon-classical Kazal family[7].
The NJ tree topology showed three defined groups, one ofthem formed only by NcPI-S domain, another group cluster-ing TgPI-1 and NcEST1 domains and a third group cluster-ing TgPI-2, TgTI, NcEST2 and EtEST-S domains (Fig. 2B).
3.4. Expression of the recombinant protein andidentification of the native protein
The cDNA region encoding the mature version of NcPI-Swas expressed as recombinant protein (rNcPI-S) inE. coli. Itwas soluble and migrated with an apparent molecular massof ∼16-kDa (Fig. 3A), which resulted to be greater than theexpected, in a similar way than it was observed on TgPI-1and TgPI-2 proteins[7,9,10]. To identify native NcPI-S pro-tein, antibodies were generated by rabbit immunization. Theimmunoblot analysis showed that the anti-rNcPI-S serumrecognized rNcPI-S (lane 1,Fig. 3B). Several bands of ap-proximately 20, 30, 40, and 66-kDA in the wholeN. caninumtachyzoite homogenate were recognized by the same anti-body (lane 3,Fig. 3B). Similar pattern was obtained by us-ing 20% SDS-PAGE either under reducing or non-reducingconditions (data not shown). This polyclonal serum did notshow reactivity against rTgPI (lane 2,Fig. 3B). Western blotanalysis with an anti-rTgPI serum[7] showed no reactiv-ity with rNcPI-S but it recognized itself and two bands ofapproximately 25 and 35-kDa inN. caninumlysate (Fig.3C). Anti-rNcPI-S sera did not recognize any protein ofT. gondii lysate (Fig. 3D). Pre-immune sera did not reactagainst rNcPI-S, rTgPI, or parasite lysate (data not shown).
3.5. Inhibitory activity of rNcPI-S
The relative inhibitory activity of rNcPI-S in compari-son with rTgPI-1 and rROP2 versus the serin proteinases
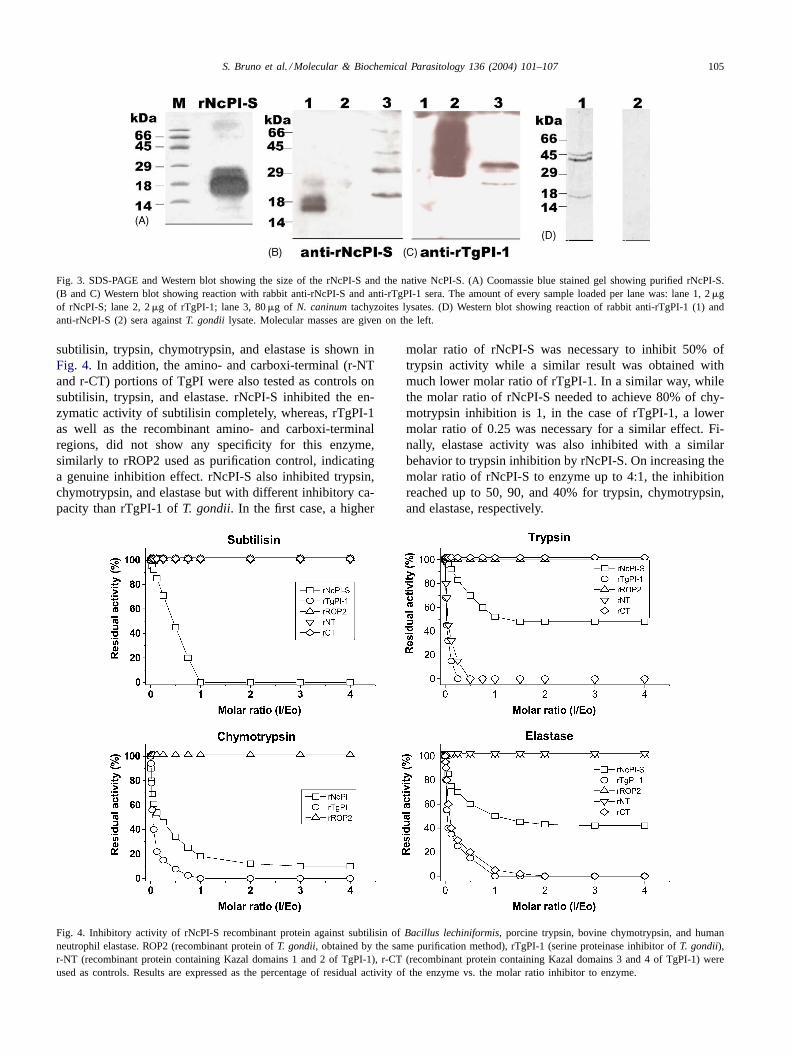
S. Bruno et al. / Molecular & Biochemical Parasitology 136 (2004) 101–107 105
Fig. 3. SDS-PAGE and Western blot showing the size of the rNcPI-S and the native NcPI-S. (A) Coomassie blue stained gel showing purified rNcPI-S.(B and C) Western blot showing reaction with rabbit anti-rNcPI-S and anti-rTgPI-1 sera. The amount of every sample loaded per lane was: lane 1, 2�gof rNcPI-S; lane 2, 2�g of rTgPI-1; lane 3, 80�g of N. caninumtachyzoites lysates. (D) Western blot showing reaction of rabbit anti-rTgPI-1 (1) andanti-rNcPI-S (2) sera againstT. gondii lysate. Molecular masses are given on the left.
subtilisin, trypsin, chymotrypsin, and elastase is shown inFig. 4. In addition, the amino- and carboxi-terminal (r-NTand r-CT) portions of TgPI were also tested as controls onsubtilisin, trypsin, and elastase. rNcPI-S inhibited the en-zymatic activity of subtilisin completely, whereas, rTgPI-1as well as the recombinant amino- and carboxi-terminalregions, did not show any specificity for this enzyme,similarly to rROP2 used as purification control, indicatinga genuine inhibition effect. rNcPI-S also inhibited trypsin,chymotrypsin, and elastase but with different inhibitory ca-pacity than rTgPI-1 ofT. gondii. In the first case, a higher
Fig. 4. Inhibitory activity of rNcPI-S recombinant protein against subtilisin ofBacillus lechiniformis, porcine trypsin, bovine chymotrypsin, and humanneutrophil elastase. ROP2 (recombinant protein ofT. gondii, obtained by the same purification method), rTgPI-1 (serine proteinase inhibitor ofT. gondii),r-NT (recombinant protein containing Kazal domains 1 and 2 of TgPI-1), r-CT (recombinant protein containing Kazal domains 3 and 4 of TgPI-1) wereused as controls. Results are expressed as the percentage of residual activity of the enzyme vs. the molar ratio inhibitor to enzyme.
molar ratio of rNcPI-S was necessary to inhibit 50% oftrypsin activity while a similar result was obtained withmuch lower molar ratio of rTgPI-1. In a similar way, whilethe molar ratio of rNcPI-S needed to achieve 80% of chy-motrypsin inhibition is 1, in the case of rTgPI-1, a lowermolar ratio of 0.25 was necessary for a similar effect. Fi-nally, elastase activity was also inhibited with a similarbehavior to trypsin inhibition by rNcPI-S. On increasing themolar ratio of rNcPI-S to enzyme up to 4:1, the inhibitionreached up to 50, 90, and 40% for trypsin, chymotrypsin,and elastase, respectively.

106 S. Bruno et al. / Molecular & Biochemical Parasitology 136 (2004) 101–107
r-NT and r-CT showed a similar behavior to completeTgPI-1 on trypsin and elastase, respectively, but the doublemolar ratio inhibitor to enzyme was necessary for completeinhibition with the recombinant halves of TgPI-1 in bothcases (Fig. 4). rNcPI-S did not inhibit neither tryptase northrombin (data not shown).
4. Discussion
A cDNA (NcPI-S) obtained fromN. caninumEST projectwas characterized, finding that it encodes for single domainserpin of the non-classical Kazal family. In addition, otherapicomplexan serpins were sequenced (NcPI-H, probablya pseudo-gene) or detected in the database, three fromN.caninumand one fromE. tenella. Despite that sequencesobtained from databases need to be confirmed by addi-tional sequencing, the analysis of deduced proteins, domainsand signal peptides suggested that NcEST1 and NcEST2could be theN. canimuncounterpart of TgPI-1 and TgPI-2,respectively. In this case, these data would be suggestingthat the presence of these two genes preceded theN. can-inum andToxoplasmadivergence. NJ analysis showed thatTgPI-1 and NcEST1 domains clustered together as well asTgPI-2, NcEST2, TgTI, and EtEST-S domains, whereas,NcPI-S domain arose as a separate branch. Interestingly,rTgPI-2 proteins showed only specificity for trypsin[10,11],whereas, rTgPI-1 presented strong inhibitory activities to-wards trypsin, chymotrypsin, tryptase and human neutrophilelastase[8,9], but none of them showed specificity for sub-tilisin. In contrast, rNcPI-S presented a unique specificity forsubtilisin, but also showed capacity to inhibit human neu-trophil elastase, trypsin, and chymotrypsin. We wonder ifNcEST1 and NcEST2 have also the same specificity, behav-ior, and role than their putativeT. gondii counterparts. Ex-periments addressing analysis of inhibitory specificity andexon–intron sequence evolution among these genes and pro-teins are carrying out to verify the suggested hypothesis re-lated to divergence and relationship betweenN. caninumandT. gondiiserpin genes.
It is known that the inhibitory specificity of a domainis largely determined by the residues at the P1-P1′ posi-tion within the reactive site loop region which acts as apseudo-substrate for the target proteinase[14] and varieswidely among Kazal inhibitors. Inhibitors with P1 Lysand Arg tend to inhibit trypsin and trypsin-like enzymes,while those with P1 Leu and Met are strong inhibitors ofelastase. In agreement with this rule, domains 1 and 2 ofTgPI-1 (r-NT, P1: Arg) lost the elastase specificity, whereas,domains 3 and 4 of TgPI-1 (rCT, P1: Leu) lost trypsinspecificity. AmongN. caninum, T. gondii, and E. tenellaserpins, as well as if hypothetical serpins are considered,NcPI-S is the only protein that showed Met in P1 position.Recent studies indicated that the functional classificationof serpins relies on their protein structure and not on se-quence similarity[15]. Therefore, the presence of Met in
P1 and/or structural features around the reactive site mightbe responsible for the differences in specificity.
Despite TgTI also showed to be a single domain serpin[11], based on sequence analysis it showed to be more re-lated to TgPI-2[10] than NcPI-S protein. NcPI-S domain isa short polypeptide that comprises practically the entire pro-tein structure, and resembles the bikazin family. This familyof Kazal-type inhibitors were isolated from sub-mandibularglands of some carnivores like bagger, mink, snow leopard,fox, and in extremely large amounts in dogs[16,17]. Theyshowed, unlike NcPI-S, a double-headed structure insteadof a single headed one, consisting of two tandem Kazal-typedomains linked by a short connecting peptide, which inhibitstrypsin (domain with Arg in P1), chymotrypsin, subtilisin,and elastase (domain with Met in P1), the latter with simi-larity to NcPI-S.
The immunoblot analysis, using an anti-rNcPI-S poly-clonal antibody, showed multiple bands pattern. This couldbe indicating cross reacting proteins, complexes with otherproteins (in general, serpin–protease interaction has tightbinding properties), and/or formation of large homod-imer complexes. The majority of serpin proteins wouldbe single-chain polypeptides, but homodimer complexes,shown to be functional, were also observed[18]. On theother hand, several studies linked the multimerization pro-cess to a misfolded serpin that showed enhanced liability[19,20]. Further studies should be done to know if nativeNcPI-S are forming multimeric forms of NcPI-S and, if thisis the case, it should be probed if they are active or not.
It is noteworthy that, an anti-rTgPI-1 polyclonal serumdetected another cross reactive protein, suggesting the pres-ence of another putative serpin inN. caninum, different toNcPI-S but related to TgPI-1. Since the NcEST-1 and TgPI-1showed high similarity at the amino acid level, it might beexpected that anti-TgPI-1 antibodies recognize the NcEST-1protein. Experiments to elucidate this are being carried out.
The presence of at least three distinct serpin genes inN.caninum, might be suggesting a fine homeostatic regulationof either parasite proteases and/or environmental proteases.Regarding the role of NcPI-S, it could be modulating theaction of aN. caninumsubtilisin-like serine protease, whichhas been recently described[21]. In addition, NcPI-S couldhave an interference role in proinflamatory response by in-hibition of elastase proteins[22,23], as well as it could pro-tect the parasite from the action of host proteinases (trypsin,elastase, subtilisin).
Acknowledgements
This work was supported by an ANPCyT grant(BID802/OC-AR-PICT 05-04831) and INEI (ANLIS Dr.Carlos G. Malbran). S.O.A. and V.G.D. are researchers ofNational Council Research (CONICET). We acknowledgeDr. Celeste Pérez, Dr. Fernando Landreau, Dr. Adrián Lewis,and Dr. Mónica Tous (cell culture provision and assistance).

S. Bruno et al. / Molecular & Biochemical Parasitology 136 (2004) 101–107 107
References
[1] Dubey JP, Dolores EH, Lindsay DS, Jenkins MC, Uggla A, SpeerCA. Neospora caninumandHammondia heydoniare separate species.Trends Parasitol 2002;18:66–9.
[2] Dubey JP, Hattel AL, Lindsay DS, Topper MJ. NeonatalNeosporacaninum infection in dogs: isolation of the causative agent andexperimental transmission. J Am Vet Med Assoc 1988;193:1259–63.
[3] Anderson ML, Blanchard PC, Barr BC, Dubey JP, Hoffman RL, Con-rad PA.Neospora-like protozoan infection as a major cause of abo-rtion in California dairy cattle. Am Vet Med Assoc 1991;198:241–4.
[4] Dubey J P. Recent advances inNeosporaand neosporosis. Vet Par-asitol 1999;84:349–67.
[5] Roberts RM, Mathialagan N, Duffy JY, Smith GW. Regulation andregulatory role of proteinase inhibitors. Crit Rev Eukaryot Gene Expr1995;5:385–436.
[6] Gettings PG. Serpin structure, mechanism and function. Chem Rev2002;102:4751–803.
[7] Pszenny V, Angel SO, Duschak VG, Paulino M, Ledesma B, YaboMI, et al. Molecular cloning, sequencing and expression of a serineproteinase inhibitor gene formToxoplasma gondii. Mol BiochemParasitol 2000;107:241–9.
[8] Pszenny V, Ledesma BE, Matrajt M, Duschak VG, Bontempi EJ,Dubremetz JF, et al. Subcellular localization and post-secretory tar-geting of TgPI, a serine proteinase inhibitor fromToxoplasma gondii.Mol Biochem Parasitol 2002;121:283–6.
[9] Morris MT, Coppin A, Tomavo S, Carruthers V. Functional anal-ysis of Toxoplasma gondiiprotease inhibitor 1. J Biol Chem2002;277:45259–66.
[10] Morris MT, Carruthers V. Identification and partial characterizationof a second Kazal inhibitor inToxoplasma gondii. Mol BiochemParasitol 2003;128:119–22.
[11] Lindh JG, Botero-Kleiven S, Arboleda JI, Wahlgren M. A proteaseinhibitor associated with the surface ofToxoplasma gondii. MolBiochem Parasitol 2001;116:137–45.
[12] Kimura M. A simple method for estimating evolutionary rates of basesubstitution through comparative studied of nucleotide sequences. JMol Evol 1980;16:111–20.
[13] Mart́ın V, Arcavi M, Amendoeira MRR, de Souza Neves E, SantillánG, Guarnera E, et al. Detection of human anti-Toxoplasmaspecificimmunoglobulins A, M and G by a recombinantToxoplasma gondiiRop2 protein. Clin Diagn Lab Immunol 1998;5:627–31.
[14] Kato I, Kohr WJ, Laskowski Jr M. Regulatory proteolytic enzymesand their inhibitors. In: Magnusson S, Ottesen M, Foltmann B, DanoK, Neurath H, editors. Proceedings of the 11th FEBS Meeting.Oxford: Pergamon; 1978. p. 197–206.
[15] Van Gent D, Sharp P, Morgan K, Kalsheker N. Serpins: struc-ture, function and molecular evolution. Int J Biochem Cell Biol2003;35:1536–47.
[16] Homandberg G. Interaction of exopeptidases with specific proteinsfrom ascaris lumbricoides. In: Peeters H, editor. Protides of theBiological Fluids, XXIII. Pergamon; 1976. p. 267–70.
[17] Hochstrasser K, Wachtter E, Reisinger PW, Greim M, AlbrechtGJ, Gebhard W, Amino acid sequences of mammalian Kazal-typeproteinase inhibitors from salivary glands . Comp Biochem PhysiolB 1993;106:103–8.
[18] Nar H, Bauer M, Stassen JM, Lang D, Gils A, Declerck PJ. Plas-minogen activator inhibitor 1. Structure of the native serpin, compar-ison to its other conformers and implications for serpin inactivation.J Mol Biol 2000;297:683–95.
[19] Yu MH, Lee KN, Kim J. The Z type variation of human al-pha 1-antitrypsin causes a protein folding defect. Nat Struct Biol1995;2:363–7.
[20] Bos IG, Lubbers YTP, Roem D, Abrahamsb JP, Hack CE, Elder-ingd E. The functional integrity of the serpin domain of C1-Inhibitordepends on the unique N-terminal domain, as revealed by a patho-logical mutant. J Biol Chem 2003;278:24963–70.
[21] Louie K, Conrad PA. Characterization of a cDNA encoding asubtilisin-like serine protease (NC-p65) ofNeospora caninum. MolBiochem Parasitol 1999;103:211–23.
[22] Shepherd JC, Aitken A, McManus DP. A protein secreted in vivoby Echinococcus granulosusinhibits elastase activity and neutrophilchemotaxis. Mol Biochem Parasitol 1991;44:81–90.
[23] Maizels RM, Gomez-Escobar N, Gregory WF, Murray J, ZangX. Immune evasion genes from filarial nematodes. Int J Parasitol2001;31:889–98.