identification of a novel antiapoptotic protein, gam-1, encoded by
TRANSCRIPT

JOURNAL OF VIROLOGY,0022-538X/97/$04.0010
Apr. 1997, p. 3168–3177 Vol. 71, No. 4
Copyright q 1997, American Society for Microbiology
Identification of a Novel Antiapoptotic Protein, GAM-1,Encoded by the CELO Adenovirus
SUSANNA CHIOCCA,† ADAM BAKER, AND MATT COTTEN*
Institute for Molecular Pathology, 1030 Vienna, Austria
Received 27 August 1996/Accepted 11 December 1996
We have developed a simple screening method to identify genes that mimic bcl-2 or adenovirus E1B 19K inenhancing cell survival after transfection and have used this method to identify such a gene in the avianadenovirus CELO. The gene encodes a novel 30-kDa nuclear protein, which we have named GAM-1, thatfunctions comparably to Bcl-2 and adenovirus E1B 19K in blocking apoptosis. However, GAM-1 has nosequence homology to Bcl-2, E1B 19K, or any other known antiapoptotic proteins and thus defines a novelantiapoptotic function.
Apoptosis is a frequent response of eukaryotic cells to virusinfection. It is clearly of benefit to a multicellular organismwhen a single infected cell dies before a virus can replicate andinfect neighboring cells. However, many viruses possess genesthat retard or block the apoptotic response sufficiently to allowvirus replication. Antiapoptotic activities have been identifiedin a variety of viruses (46), including adenovirus (E1B 19K [15,41, 57, 60, 61]), Epstein-Barr virus (BHRF1 [53]), baculovirus(p35 and IAP [2, 11]), African swine fever virus (LMW5-HL[36]), cowpox virus (CrmA [21]), and human cytomegalovirus(IE1 and IE2 [66]). In an early description of apoptosis, theadenovirus deg or cyt phenotype (16, 18, 51) was due to E1B19K mutation (29, 39, 47, 50, 58, 61), and it is now clear thatapoptosis can be generated by expression of an E1A growthsignal in the absence of E1B (15, 41). The adenovirus E1B 19Kand E1B 55K proteins block this E1A-induced apoptosis, forc-ing host cells to survive long enough to allow completion ofviral replication (15). The E1B 19K protein functions similar tothe antiapoptotic mammalian protein Bcl-2 in several differentsettings (9, 15, 41, 49, 52).We recently reported the complete sequence of CELO
(chicken adenovirus type 1) (8). A striking feature of thisadenovirus genome is the lack of E1A and E1B regions iden-tifiable by sequence homology. We presumed that like mostsmall DNA viruses, CELO must possess genes, such as thoseencoded by the E1A region, which influence cellular prolifer-ation and genes such as E1B 55K and E1B 19K, which alter thecellular apoptotic response. Furthermore, CELO is capable oftransforming cells and inducing tumors in newborn hamsters(1, 27, 32, 33, 43), demonstrating that the virus does possesstransforming genes and could therefore possess functional ho-mologs to the mastadenovirus E1A and E1B regions. Lackingan obvious sequence homolog, we have established a functionassay to screen for a gene in CELO that slows the apoptoticresponse.Our method for identifying genes that block apoptosis is
based on an established adenovirus-mediated transfection sys-tem in which adenovirus particles, linked to polylysine-con-densed plasmid DNA, are used to transfer DNA into primaryhuman fibroblasts (54). Transcriptionally silent, psoralen-inac-
tivated virus particles (13) are used to avoid gene expressionfrom the carrier adenovirus. We obtain high levels of transientgene expression with this system, but in many cell types, theexpression levels decline rapidly over a 10-day period. Wedemonstrate here that this decline is slowed by the cotransfec-tion of bcl-2 or E1B 19K, two genes whose ability to blockapoptosis is well documented. We report here on the use ofthis transfection system to identify a novel gene in CELO thatmimics the functions of E1B 19K and bcl-2.
MATERIALS AND METHODS
CELO library. CELO was grown and purified as previously described (14).CELO genomic DNA was prepared from purified CELO by using a sodiumdodecyl sulfate-proteinase K digestion followed by banding in a CsCl densitygradient. To create the pXCELO libraries, purified CELO DNA was digestedwithHindIII or EcoRI and cloned downstream from the cytomegalovirus (CMV)enhancer/promoter in plasmid pX (48). Individual plasmids bearing each of theexpected fragments were isolated. Because of the structure of the adenovirusterminal fragments, the end fragments of the CELO genome were not obtained.The BssHII, SacII, SphI, and BglII GAM-1 mutations (see Fig. 3B) were
prepared by digesting pX9R1SmaI/HindIII (which encodes GAM-1) with theindicated restriction enzyme, generating blunt ends by treatment with Klenowenzyme, and ligating. Plasmids bearing the expected mutations were isolated andverified by DNA sequencing. The resulting peptide sequences of the mutantproteins are presented in Table 1. The leucine mutations (L 258, 265 P; L 258,265 A; and L 258, 265 G) were constructed using a PCR-based method; detailscan be supplied upon request.Additional plasmids. Plasmid pCMV-Bcl-2, encoding the bcl-2 cDNA (44)
driven by the CMV promoter/enhancer, was provided by Michael Buschle (In-stitute for Molecular Pathology). The CMV-driven green fluorescent protein(GFP) expression plasmid pCMV-GFP was derived from the GFP cDNA de-rived by Chalfie et al. (7). The CMV-driven luciferase expression plasmid pCLucwas described earlier (40). pCMV-E1B 19K and pCMVE1A (59) were providedby Eileen White. For the fluorescence-activated cell sorting (FACS) analysisshown in Table 2, the enhanced GFP plasmid pEGFP-C1 (Clontech) was used.The NF-kB-responsive luciferase reporter plasmid p3K-Luc was constructed asa derivative of pTK3kbB (2a). pTK3kbB was cleaved with XhoI and NcoI toremove most of the chloramphenicol acetyltransferase coding sequence, treatedwith Klenow enzyme, and ligated to a Klenow-treated HindIII/SspI fragmentfrom pRSVL (15a) encoding the luciferase sequence. This resulted in plasmidp3K-Luc, containing a triple binding site for the transcription factor NF-kB plusa thymidine kinase TATA box, driving the luciferase coding sequence.All plasmid preparations were purified and processed to remove lipopolysac-
charide as previously described (12).Transfections. E4-defective adenovirus type 5 dl1014 (4) was grown on W162
cells (56), purified, and processed for biotinylation and 8-methoxypsoralen inac-tivation (13). Transfection complexes containing biotinylated, psoralen-inacti-vated adenovirus dl1014 linked to streptavidin-polylysine (StrpL)–transferrin-polylysine-condensed plasmid DNA (usually 5 mg of a CMV-luciferase or CMV-GFP plasmid plus 1 mg of test plasmid, but see figure legends for details) wereprepared as described previously (13, 54). In brief, samples of biotinylated,psoralen-inactivated adenovirus dl1014 (8 ml, 1012 particles/ml) were diluted into150 ml of HEPES-buffered saline (HBS) and mixed with 1 mg of StrpL (54) in 150ml of HBS for 30 min at room temperature. Aliquots of 6 mg of plasmid DNA (6
* Corresponding author. Mailing address: Institute for MolecularPathology, Dr. Bohr Gasse 7, 1030 Vienna, Austria. Phone: 43 1 797 30526. Fax: 43 1 798 71 53. E-mail: [email protected].† Present address: Department of Experimental Oncology, Euro-
pean Institute of Oncology, 20141 Milan, Italy.
3168
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

mg of DNA) were diluted in 100 ml of HBS and then mixed with the adenovirus-StrpL solution for 30 min at room temperature. Finally, a 100-ml aliquot of HBScontaining 5 mg of transferrin-polylysine (55) was added to each sample, followedby incubation for 30 min at room temperature. Aliquots of the transfectioncomplexes were then supplied to cells (15 ml per 20,000 cells) in 500 ml ofDulbecco modified Eagle medium plus 2 mM glutamine, 100 IU of penicillin, 100mg of streptomycin per ml, and 2% horse serum. After 2 h at 378C, the mediumwas replaced with the same medium containing 10% fetal calf serum.Primary human skin fibroblasts (between passage 5 and 15) were used. In most
cases, both orientations of the CELO fragment (relative to the CMV promoter)were tested.TdT analysis of apoptosis. HeLa or A549 cells were transfected with the
cationic lipid Transfectam (1a) with 5 mg of pEGFP-C1 (Clontech) plus 1 mg ofeither pX, pX9R1SmaI/HindIII (GAM-1), or pCMV-E1B 19K. At 4 days post-transfection, cells (106) were harvested and fixed overnight in 1% paraformal-dehyde. The terminal deoxynucleotidyltransferase (TdT) reaction was then per-formed by resuspending the cells in 50 ml of TdT reaction mixture (200 mMpotassium cacodylate, 25 mM Tris-HCl [pH 6.6], 0.25 mg of bovine serumalbumin per ml, 1 to 2.5 mM CoCl2; 0.25 to 0.5 nmol of FluoroLink Cy5AP3-dCTP [Amersham], 12.5 U of TdT) and incubating them for 1 h at 378C. Cellswere then simultaneously analyzed for GFP expression and TdT staining by usinga FACS Vantage machine (Turbo Sort AC du SEM).PolyHEMA coating of cell culture plates. PolyHEMA [poly(2-hydroxyethyl
methacrylate] coating of cell culture plates was performed by a method similar tothat described by Folkman and Moscona (19) by applying 200 ml of polyHEMAsolution per well of a 24-well plate. Each well of normal tissue culture plates(24-well plates) was coated with 200 ml of solution (Sigma catalog no. P-3932; 10mg/ml in 95% ethanol). The ethanol was allowed to evaporate overnight at 378C,and this coating was repeated once. The wells were washed twice with HBS andonce with medium before the addition of cells. Dilutions of the original poly-HEMA stock (6 g of polyHEMA in 50 ml of 95% ethanol) were 1021, 1022, and1023.
RESULTS
Adenovirus-mediated transfection activates an apoptotic re-sponse. When a reporter gene (encoding luciferase or GFP)was transfected by using an adenovirus-polylysine transfectionsystem, we observed initial high levels of reporter gene expres-sion that declined over a 10-day period. Cotransfection of anantiapoptotic gene such as E1B 19K or bcl-2 markedly slowedthis decline (Fig. 1). This is demonstrated qualitatively in as-says using GFP expression to monitor the fate of transfectedcells (Fig. 1A). At day 1, comparable levels of GFP expressionare observed independent of the presence of a cotransfectedantiapoptotic gene, thus excluding the possibility that the co-transfected gene is directly influencing CMV-luciferase geneexpression by, for example, upregulating transcription of thereporter gene. By day 5, GFP-expressing cells in the controldeclined in frequency and displayed an apoptotic phenotype(cytoplasmic blebbing, partial substrate detachment, and nu-clear condensation and fragmentation [Fig. 1A]). Cotransfec-tion of the apoptosis-promoting adenovirus E1A gene en-hanced the incidence of the apoptotic phenotype (Fig. 1A). Incontrast, cells cotransfected with GFP and an antiapoptoticgene (bcl-2 or E1B 19K) maintain both gene expression andnormal morphology at later times. Quantitative analyses of
both GFP and luciferase expression over time (Fig. 1B and C)demonstrate similar declines in expression that are blocked byE1B 19K. A similar gene expression assay has been used toscreen the functions of bcl-2 variants (25, 26).This apoptosis after adenovirus-mediated transfection is in-
dependent of p53; the apoptosis is also observed in p532/2
fibroblasts, and no protection is obtained by cotransfection ofgenes that sequester p53 (Mdm-2 or temperature-sensitive p53at the restrictive temperature [results not shown]). This is animportant distinction because one activity of E1B 19K is toprevent a p53-dependent repression of transcription (45).Thus, after adenovirus-mediated transfection, gene expressiondeclines in a p53-independent apoptotic response that can beblocked by E1B 19K or Bcl-2. This transfection phenomenoncan therefore be used to identify functional homologs of Bcl-2and E1B 19K.Identification of an antiapoptotic CELO gene.We suspected
that CELO, like many other viruses, encoded an antiapoptoticfunction. Screening a library of CELO genomic fragments, weidentified two overlapping restriction fragments (theHindIII Dfragment in clone 7H3 and the EcoRI D fragment in clone 9R1[Fig. 2B]) that provided an enhancement of cell survival com-parable to that provided by Bcl-2 and E1B 19K when cotrans-fected with a luciferase reporter gene (Fig. 2A).The DNA sequence common to the two fragments (bp 1 to
3108 in Fig. 2D) includes several possible open reading frames(ORFs). A SmaI/HindIII restriction fragment bearing the larg-est ORF of 9R1 (Fig. 2D) exhibited the full activity of theentire 9R1 fragment (Fig. 3B and data not shown). Prematureframeshifts and stop codons were introduced at several sites inthe large 9R1 ORF. Analysis of these mutations demonstratedthat the protective function was not disrupted by an alterationoutside the ORF at the BssHII site, but alterations within theORF (at the SacII, SphI, and BglII sites [Fig. 2D]) disruptedthe activity and therefore clearly identified the protective ORFbetween bp 1600 and 2700 of the 9R1 fragment (Fig. 3B, lanes4 to 7). Furthermore, removing the amino-terminal 16 or 41amino acids impaired survival activity (Table 1), demonstratingthat these residues are essential for the protective function.GFP coexpression was used to monitor the morphology of
GAM-1-expressing cells. We find that like E1B 19K-expressingcells, GAM-1-expressing cells are nonapoptotic at times whencontrol and E1A expressing cells display predominantly anapoptotic morphology (Fig. 1A).There are no significant homologies between the protein
encoded by the 9R1 ORF (called hereafter GAM-1, for Gallusanti morte) and any of the known antiapoptotic proteins, in-cluding members of the Bcl-2 family, adenovirus type 5 E1B19K, the Bax family, the baculovirus IAP family, or the nem-atode protein Ced-3. In particular, GAM-1 does not possess
TABLE 1. Summary of GAM-1 mutations
Construct Description Antiapoptosis activity
pX9R1SmaI/HindIII Wild-type sequence YesMutant BssHII Wild-type sequence YesMutant SacII Wild type to S 93; adds GTQDPQstop NoMutant SphI Wild type to V 145; adds Pstop NoMutant BglII Wild type to D 195; adds RSEKNTSRTILQPIFstop NoMutant S2-3 Removes first 16 amino acids NoMutant S2-4 Removes first 41 amino acids NoGAM-1 (L 258, 265 P) Changes L 258 and L 265 to P NoGAM-1 (L 258, 265 G) Changes L 258 and L 265 to G NoGAM-1 (L 258, 265 A) Changes L 258 and L 265 to A No
VOL. 71, 1997 CELO ANTIAPOPTOSIS GENE 3169
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

the essential BH1, BH2, and BH3 motifs that define membersof the Bcl-2 family (10, 23, 26, 37, 38, 63, 64). CELO does notencode a gene with adenovirus E1B 19K homology and ap-pears to possess as a functional homolog the GAM-1 gene.
Mutational analysis of GAM-1. Analysis of the protein se-quence of GAM-1 revealed the presence of a series of spacedleucine residues in the carboxy-terminal region, with three ofthe four leucines in a seven-residue repeat (Fig. 3A) resem-
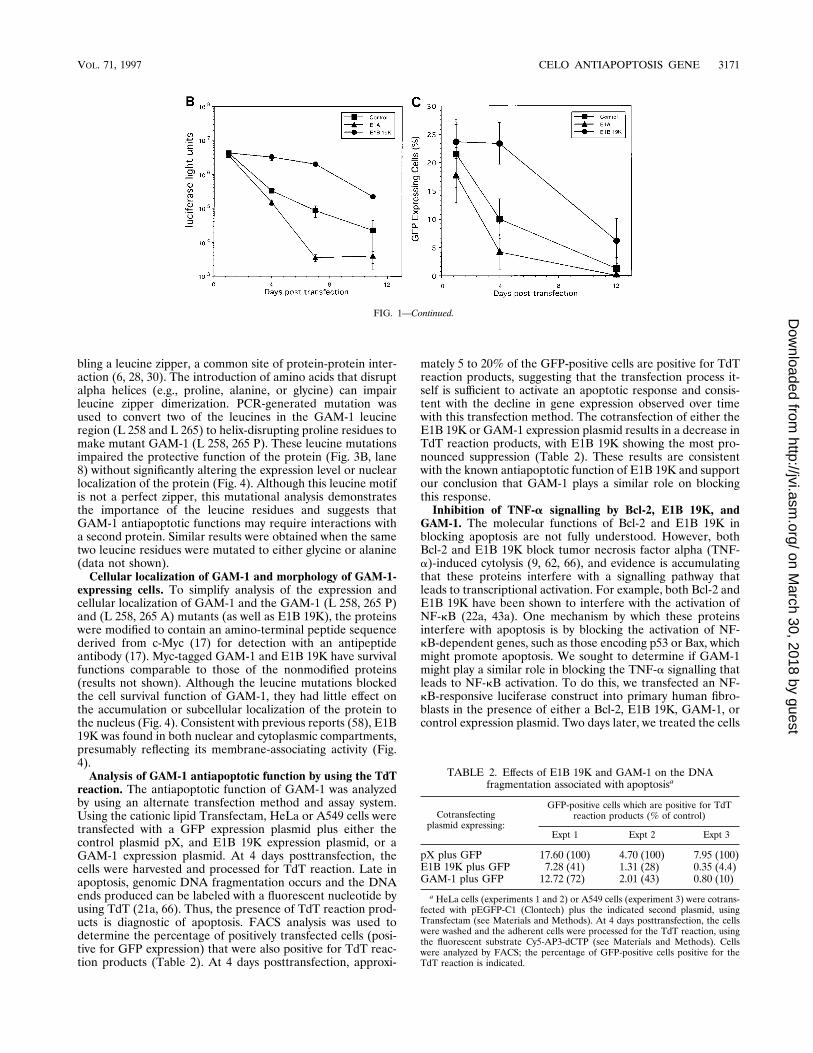
FIG. 1. Adenovirus transfection induces an apoptotic response. (A) Cotransfecting antiapoptotic genes enhances long-term gene expression. Primary fibroblastswere transfected with adenovirus-polylysine-DNA complexes containing pCMV-GFP plus control DNA pX or the indicated second plasmid (pCMV-E1A, pCMV-E1B19K, pCMV-Bcl-2, or pX9R1SmaI/HindIII [GAM-1]). At the indicated times posttransfection, the cells were fixed, stained with 49,6-diamidino-2-phenylindole (DAPI)to visualize DNA, and examined by fluorescence microscopy. Images were captured by a charge-coupled device camera and assembled with Adobe Photoshop. (B)Primary fibroblasts were transfected as described above with the luciferase-encoding vector pCMVLuc plus either control DNA (pX), pCMV-E1A, or pCMV-E1B 19K.At the indicated times posttransfection, cells were harvested and samples (standardized for protein content) were assayed for luciferase activity. Each point representsthe average of three transfection experiments with the standard deviation indicated. (C) Primary fibroblasts were transfected as described above and processed for GFPanalysis. At each time point, random microscope fields (10 to 30 cells per field) were examined. Percent GFP expressing cells 5 (GFP-positive cells/total cells in field)3 100. Each point represents the average value from at least 10 fields with the standard deviation indicated.
3170 CHIOCCA ET AL. J. VIROL.
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
bling a leucine zipper, a common site of protein-protein inter-action (6, 28, 30). The introduction of amino acids that disruptalpha helices (e.g., proline, alanine, or glycine) can impairleucine zipper dimerization. PCR-generated mutation wasused to convert two of the leucines in the GAM-1 leucineregion (L 258 and L 265) to helix-disrupting proline residues tomake mutant GAM-1 (L 258, 265 P). These leucine mutationsimpaired the protective function of the protein (Fig. 3B, lane8) without significantly altering the expression level or nuclearlocalization of the protein (Fig. 4). Although this leucine motifis not a perfect zipper, this mutational analysis demonstratesthe importance of the leucine residues and suggests thatGAM-1 antiapoptotic functions may require interactions witha second protein. Similar results were obtained when the sametwo leucine residues were mutated to either glycine or alanine(data not shown).Cellular localization of GAM-1 and morphology of GAM-1-
expressing cells. To simplify analysis of the expression andcellular localization of GAM-1 and the GAM-1 (L 258, 265 P)and (L 258, 265 A) mutants (as well as E1B 19K), the proteinswere modified to contain an amino-terminal peptide sequencederived from c-Myc (17) for detection with an antipeptideantibody (17). Myc-tagged GAM-1 and E1B 19K have survivalfunctions comparable to those of the nonmodified proteins(results not shown). Although the leucine mutations blockedthe cell survival function of GAM-1, they had little effect onthe accumulation or subcellular localization of the protein tothe nucleus (Fig. 4). Consistent with previous reports (58), E1B19K was found in both nuclear and cytoplasmic compartments,presumably reflecting its membrane-associating activity (Fig.4).Analysis of GAM-1 antiapoptotic function by using the TdT
reaction. The antiapoptotic function of GAM-1 was analyzedby using an alternate transfection method and assay system.Using the cationic lipid Transfectam, HeLa or A549 cells weretransfected with a GFP expression plasmid plus either thecontrol plasmid pX, and E1B 19K expression plasmid, or aGAM-1 expression plasmid. At 4 days posttransfection, thecells were harvested and processed for TdT reaction. Late inapoptosis, genomic DNA fragmentation occurs and the DNAends produced can be labeled with a fluorescent nucleotide byusing TdT (21a, 66). Thus, the presence of TdT reaction prod-ucts is diagnostic of apoptosis. FACS analysis was used todetermine the percentage of positively transfected cells (posi-tive for GFP expression) that were also positive for TdT reac-tion products (Table 2). At 4 days posttransfection, approxi-
mately 5 to 20% of the GFP-positive cells are positive for TdTreaction products, suggesting that the transfection process it-self is sufficient to activate an apoptotic response and consis-tent with the decline in gene expression observed over timewith this transfection method. The cotransfection of either theE1B 19K or GAM-1 expression plasmid results in a decrease inTdT reaction products, with E1B 19K showing the most pro-nounced suppression (Table 2). These results are consistentwith the known antiapoptotic function of E1B 19K and supportour conclusion that GAM-1 plays a similar role on blockingthis response.Inhibition of TNF-a signalling by Bcl-2, E1B 19K, and
GAM-1. The molecular functions of Bcl-2 and E1B 19K inblocking apoptosis are not fully understood. However, bothBcl-2 and E1B 19K block tumor necrosis factor alpha (TNF-a)-induced cytolysis (9, 62, 66), and evidence is accumulatingthat these proteins interfere with a signalling pathway thatleads to transcriptional activation. For example, both Bcl-2 andE1B 19K have been shown to interfere with the activation ofNF-kB (22a, 43a). One mechanism by which these proteinsinterfere with apoptosis is by blocking the activation of NF-kB-dependent genes, such as those encoding p53 or Bax, whichmight promote apoptosis. We sought to determine if GAM-1might play a similar role in blocking the TNF-a signalling thatleads to NF-kB activation. To do this, we transfected an NF-kB-responsive luciferase construct into primary human fibro-blasts in the presence of either a Bcl-2, E1B 19K, GAM-1, orcontrol expression plasmid. Two days later, we treated the cells
FIG. 1—Continued.
TABLE 2. Effects of E1B 19K and GAM-1 on the DNAfragmentation associated with apoptosisa
Cotransfectingplasmid expressing:
GFP-positive cells which are positive for TdTreaction products (% of control)
Expt 1 Expt 2 Expt 3
pX plus GFP 17.60 (100) 4.70 (100) 7.95 (100)E1B 19K plus GFP 7.28 (41) 1.31 (28) 0.35 (4.4)GAM-1 plus GFP 12.72 (72) 2.01 (43) 0.80 (10)
a HeLa cells (experiments 1 and 2) or A549 cells (experiment 3) were cotrans-fected with pEGFP-C1 (Clontech) plus the indicated second plasmid, usingTransfectam (see Materials and Methods). At 4 days posttransfection, the cellswere washed and the adherent cells were processed for the TdT reaction, usingthe fluorescent substrate Cy5-AP3-dCTP (see Materials and Methods). Cellswere analyzed by FACS; the percentage of GFP-positive cells positive for theTdT reaction is indicated.
VOL. 71, 1997 CELO ANTIAPOPTOSIS GENE 3171
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

3172 CHIOCCA ET AL. J. VIROL.
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

with TNF-a and 24 h later assayed luciferase expression as ameasure of NF-kB induction.The cells transfected with p3K-Luc alone respond with a
.6-fold induction of luciferase activity (Fig. 5A, lane 1). Con-trol experiments show that this construct is also responsive toa variety of standard NF-kB stimuli including interleukin-1,phorbol myristate acetate, and hydrogen peroxide (data notshown), demonstrating that the method adequately measuresNF-kB activation. Cotransfection of a Bcl-2 or E1B 19K ex-pression plasmid largely prevented the TNF-a induction of thispromoter (Fig. 5A, lanes 2 and 3). The expression of GAM-1also prevented the TNF-a induction, although the effect wasnot as complete as that observed with Bcl-2 and E1B 19K (Fig.5A, lane 4). Thus, it appears that one function of GAM-1 is inblocking a TNF-a signalling pathway which could lead to ap-optosis.E1B 19K, Bcl-2, and GAM-1 protect against detachment-
induced apoptosis. There can be signalling to the cell from theextracellular matrix. A number of experiments have shown thatthe appropriate extracellular environment is crucial for cellsurvival and that the absence of an appropriate environmentcan activate a death program (3, 5, 20, 34; reviewed in refer-ence 42). This may be important in establishing and maintain-ing proper cell positioning within an organism. It has beendemonstrated that blocking the substrate attachment of epi-thelial cells with polyHEMA (19) leads to apoptosis and thatthe expression of Bcl-2 can be protective against this detach-ment-induced apoptosis (20). We suspected that the decline ingene expression observed after transfection might be due to analtered matrix association, and one possible mechanism of theCELO antiapoptotic gene was an inhibition of this detach-ment-associated cell death. To test this idea, cells were trans-fected with a luciferase plasmid alone or with a plasmid en-coding E1B 19K, Bcl-2, or GAM-1. Two days later, the cellswere trypsinized and defined numbers of transfected cells weretransferred to tissue culture plates coated with a graded seriesof polyHEMA concentrations. Seven days later, the nonat-tached and attached cells were collected, and the surviving cellpopulation was determined by assaying the surviving luciferasegene expression.We find that with control cells (bearing only the luciferase
gene), a 20-fold decline in luciferase gene expression occurs ascells are plated on surfaces coated with increasing polyHEMAconcentrations (Fig. 5B, lanes 1 to 4). The surfaces coated withthe highest concentration of polyHEMA (1:10 dilution) causecomplete detachment of the cell population, while the 1:100and 1:1,000 dilutions of polyHEMA result in approximately 50and 10% detachment. All three antiapoptotic genes provideprotective activity. The bcl-2 gene appears to function moreefficiently than the E1B 19K or GAM-1 gene, with completesurvival of the luciferase expression at the highest polyHEMAconcentration (Fig. 5B, lanes 5 to 8). Although the GAM-1-loaded cells show a partial decline in survival when plated onthe highest polyHEMA concentration, the level of survival is
still comparable to that of the control cells on normal plastic(compare lanes 1 and 16).
DISCUSSION
We have developed an assay for identifying antiapoptoticgenes and have used this assay to identify and characterize anovel antiapoptotic gene, GAM-1.The apoptotic stimulus associated with the adenovirus trans-
fection methods used in this work has not yet been identified.There is substantial documentation of apoptosis stimulated byadenovirus gene expression. However, the adenovirus particlesused in this study were transcriptionally inactive. Thus, eithervirus particle entry itself or other components in the genetransfer system (polylysine or plasmid DNA) activate this celldeath response. One possibility is that at the relatively highmultiplicity of infection used for these transfections (ca. 1,000virus particles per cell), interference with cell attachment oc-curs and this partial detachment activates an apoptotic re-sponse. We can accelerate the apoptotic response by interfer-ing with cellular attachment (Fig. 5B). Furthermore, E1B 19K,
FIG. 3. (A) Amino acid sequence of GAM-1. The repetitive leucine residuesat the carboxy terminus are indicated in boldface. (B) Coexpression of CELOGAM-1 supports cell survival at levels comparable to those supported by ade-novirus E1B 19K. Primary human fibroblasts were transfected with pCLuc pluseither pX (control) or the indicated plasmids bearing E1B 19K, GAM-1, orvarious mutations (see Materials and Methods), using a mixture of 5 mg ofpCLuc plus 1 mg of test plasmid (Fig. 1). Luciferase expression at day 14posttransfection is shown. Each value represents the average of three transfec-tions with the standard deviation indicated.
FIG. 2. Identification of an antiapoptotic gene. (A) Strategy for identifying antiapoptotic genes. Primary fibroblasts are transfected with a mixture of a luciferaseor GFP expression plasmid and a plasmid encoding a possible protective gene. Apoptosis, stimulated by the transfection, results in cell death and a loss of reportergene expression. The expression of an antiapoptotic gene results in an enhanced reporter gene expression 5 to 14 days posttransfection. Dark cells are successfullytransfected cells. s/b, streptavidin/biotin linkage; TF, transferrin. (B) Screen of the CELO genome. pCLuc was cotransfected with the indicated CELO genomefragments (in plasmid pX). Double samples indicate that both orientations of the fragment (relative to the promoter) were tested. Luciferase expression was measuredat day 1, 4, 7, 14, and 21 posttransfection. In all cases, the 1-day luciferase expression levels were comparable among all samples. The relative expression at day 14 oftest samples (5 mg of pCLuc plus 1 mg of pXCELO, pCMV-E1B 19K or pCMV-Bcl-2) was compared to that for control transfection (5 mg pCLuc plus 1 mg of emptyplasmid pX). Averages of three transfection samples are shown. Enhancement of survival (indicated by a 3- to 10-fold enhancement of luciferase expression) wasobserved with pCMV-E1B 19K, pCMV-Bcl-2, pX7H3, and pX9R1. (C) Restriction map of the CELO genome based on the CELO sequence (8). (D) ORFs of greaterthan 200 bp (indicated by filled boxes) in the CELO 9R1 fragment. A 1,929-bp SmaI/HindIII fragment of the 9R1 fragment possessed full activity (Fig. 3B). Thelocations of relevant unique restriction sites in the SmaI/HindIII fragment are shown.
VOL. 71, 1997 CELO ANTIAPOPTOSIS GENE 3173
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Bcl-2, and GAM-1 can increase the survival of cells underthese detachment conditions. This finding suggests that onerole of the viral gene products is to enhance cell survival late ininfection when the cells have detached due to the shut down incellular protein synthesis (65). This might facilitate spreadingof infected cells and thus spreading of the virus to other areasof the host tissue.The conclusion that GAM-1 is the CELO counterpart to
mastadenovirus E1B 19K is based on the analysis of GAM-1and E1B-19K function in nonviral settings. We have demon-strated that GAM-1 functions like Bcl-2 and E1B 19K in block-
ing a TNF-a signalling pathway (Fig. 5A). Additional supportfor this conclusion will require complementation studies todetermine if GAM-1 will replace E1B 19K in the context of anormal adenovirus type 2 or type 5 replication. Furthermore,the consequence of a GAM-1 deletion on the replication ofCELOmust be determined. We do know that GAM-1 messageis detectable early and at more abundant levels late in CELOinfection (results not shown).The precise function of GAM-1 in blocking apoptosis is not
yet known. One possibility is that GAM-1 is a direct mimic ofBcl-2 and/or E1B 19K. This is unlikely since GAM-1 does not
FIG. 4. Subcellular localization of Myc-tagged versions of GAM-1, GAM-1(L 258, 265 P) (LL:PP), and E1B 19K. (A) Western blot analysis. Myc-taggedplasmids were generated by inserting the coding regions of each protein into thevector pSG9M (22). HeLa cells were transfected and 24 h later, cytoplasmic andnuclear fractions were prepared, resolved by sodium dodecyl sulfate-polyacryl-amide gel electrophoresis, and transferred to nitrocellulose. The Myc epitope-containing proteins were identified by using monoclonal antibody 9E10 (17),anti-mouse immunoglobulin-horseradish peroxidase, and enhanced chemilumi-nescence (Amersham). Lanes 1 and 6, extracts of cells transfected with pCLuc;lanes 2 and 7, Myc-tagged GAM-1; lanes 3, 4, 8, and 9, Myc-tagged GAM-1 (L258, 265 P); lanes 5 and 10: Myc-tagged E1B 19K. Sizes are indicated in kilo-daltons. (B) Immunofluorescence. Primary human fibroblasts were transfected asdescribed for Fig. 1. The fixed cells were stained with antibody 9E10 followed byanti-mouse immunoglobulin-fluorescein isothiocyanate (FITC). Nuclei were vi-sualized by DAPI staining.
3174 CHIOCCA ET AL. J. VIROL.
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

have the highly conserved BH1, BH2, and BH3 motifs found inBcl-2 family members (10, 23, 26, 37, 38, 63, 64). Furthermore,GAM-1 is found almost exclusively in the nucleus, with a pat-tern clearly distinct from those of E1B 19K, which is found in
both a cytoplasmic and a nuclear fraction (Fig. 4; see alsoreference 58), and Bcl-2, which is also membrane associated(31, 35). Another possibility is that GAM-1 interacts with ap-optosis-promoting proteins, such as Bax, to block apoptosis.However, GAM-1 lacks the Bax interaction domains which arethought to be the BH1, BH2, and BH3 domains (10, 23, 26, 37,38, 63, 64). GAM-1 could, like CrmA (21) and p35 (2, 11),function as an inhibitor of the proteases associated with apo-ptosis. However, there is no recognizable homology betweenGAM-1 and these protease inhibitors, and the GAM-1 nuclearlocalization would preclude any interaction with these cyto-plasmic proteases. A likely possibility is that GAM-1 functionsat the transcriptional level and blocks apoptosis by directlyupregulating the expression of protective genes. Other exam-ples of transcriptional activators that promote cell survivalinclude the Epstein-Barr virus LMP-1 protein, which directlyactivates Bcl-2 expression (24), and the CMV IE1 and IE2proteins, which function as transcriptional activators (66).GAM-1 synergizes with Bcl-2 in our transfection assay (datanot shown), suggesting that although the effects of Bcl-2 andGAM-1 are similar, the two proteins may influence apoptosisat distinct steps. Future research will identify the transcrip-tional consequences of GAM-1 expression.
ACKNOWLEDGMENTS
We thank Ernst Wagner for the abundant supply of transferrin-polylysine and StrpL and Max Birnstiel and Gerhard Christofori forcomments on the manuscript. We also thank Peter Steinlein for FACSanalysis and Kanaga Sabapathy for helpful suggestions on the TdTanalysis. We appreciate the excellent technical assistance of MediyhaSaltik and the sequencing help of Gotthold Schaffner, Robert Kurz-bauer, Ivan Botto, and Elisabeth Aigner. We thank Eileen White forpCMV-E1A-19K and Michael Buschle and Stanley Korsmeyer forpCMV-Bcl-2.
REFERENCES
1. Anderson, J., V. J. Yates, V. Jasty, and L. O. Mancini. 1969. In vitro trans-formation by an avian adenovirus (CELO). I. Hamster-embryo fibroblasticcultures. J. Natl. Cancer Inst. 42:1–7.
1a.Behr, J. P., B. Demeneix, J. P. Loeffler, and J. Perez-Mutul. 1989. Efficientgene transfer into mammalian primary endocrine cells with lipopolyamine-coated DNA. Proc. Natl. Acad. Sci. USA 86:6982–6986.
2. Birnbaum, M. J., R. J. Clem, and L. K. Miller. 1994. An apoptosis-inhibitinggene from a nuclear polyhedrosis virus encoding a polypeptide with Cys/Hissequence motifs. J. Virol. 68:2521–2528.
2a.Boehmelt, G., A. Walker, N. Kabrun, G. Mellitzer, H. Beug, M. Zenke, andP. J. Enrietto. 1992. Hormone-regulated v-rel estrogen receptor fusion pro-tein: reversible induction of cell transformation and cellular gene expression.EMBO J. 11:4641–4652.
3. Boudreau, N., C. J. Sympson, Z. Werb, and M. J. Bissell. 1995. Suppressionof ICE and apoptosis in mammary epithelial cells by extracellular matrix.Science 267:891–893.
4. Bridge, E., and G. Ketner. 1989. Redundant control of adenovirus late geneexpression by early region 4. J. Virol. 63:631–638.
5. Brooks, P. C., A. M. P. Montgomery, M. Rosenfeld, R. A. Reisfeld, T. Hu, G.Klier, and D. A. Cheresh. 1994. Integrin avb3 antagonists promote tumorregression by inducing apoptosis of angiogenic blood vessels. Cell 79:1157–1164.
6. Busch, S. J., and P. Sassone-Corsi. 1990. Dimers, leucine zippers and DNA-binding motifs. Trends Genet. 6:36–40.
7. Chalfie, M., Y. Tu, G. Euskirchen, W. W. Ward, and D. C. Prasher. 1994.Green fluorescent protein as a marker for gene expression. Science 263:802–805.
8. Chiocca, S., R. Kurzbauer, G. Schaffner, A. Baker, V. Mautner, and M.Cotten. 1996. The complete DNA sequence and genomic organization of thechicken adenovirus CELO. J. Virol. 70:2939–2949.
9. Chiou, S.-K., C.-C. Tseng, L. Rao, and E. White. 1994. Functional comple-mentation of the adenovirus E1B 19-kilodalton protein with Bcl-2 in theinhibition of apoptosis in infected cells. J. Virol. 68:6553–6566.
10. Chittenden, T., C. Flemington, A. B. Houghton, R. G. Ebb, G. J. Gallo, B.Elangovan, G. Chinnadurai, and R. J. Lutz. 1995. A conserved domain inBak, distinct from BH1 and BH2, mediates cell death and protein bindingfunctions. EMBO J. 14:5589–5596.
11. Clem, R. J., and L. K. Miller. 1994. Induction and inhibition of apoptosis by
FIG. 5. (A) Coexpression of Bcl-2, E1B 19K, and CELO GAM-1 interfereswith TNF-a-induced NF-kB activation. Primary human fibroblasts were trans-fected with p3K-Luc plus either pX (control) or the indicated plasmids bearingBcl-2, E1B 19K, or GAM-1 (pX9R1SmaI/HindIII), using a mixture of 4 mg ofp3K-Luc plus 2 mg of test plasmid (Fig. 1). At 2 days posttransfection, testcultures were stimulated with 25 ng of TNF-a per ml. Twenty-four hours later,luciferase expression was measured. All values are the averages of three trans-fection samples normalized for protein content. The values are expressed as foldinduction over control (same transfection, no TNF-a treatment) with the stan-dard deviation indicated. (B) Effects of Bcl-2, E1B 19K, and GAM-1 expressionon detachment-induced apoptosis. Primary human fibroblasts were transfected(as described for Fig. 1) with a luciferase marker plus either empty vector(control) or various test plasmids (5 mg of pCLuc plus 1 mg of either pSP65 or thepXCELO test plasmids). At 2 days posttransfection, the transfected cells weretrypsinized and counted, and defined numbers of cells (20,000 per well) wereplated, in triplicate, in 24-well plates which had been precoated with the indi-cated dilution of polyHEMA (see Materials and Methods). Seven days afterplating, the luciferase activity in the entire population of cells (nonadherent cellsplus adherent cells) was measured. Each value represents the average of threetransfections with the standard deviation indicated.
VOL. 71, 1997 CELO ANTIAPOPTOSIS GENE 3175
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

insect viruses, p. 89–110. In Apoptosis II: the molecular basis of apoptosis indisease. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
12. Cotten, M., A. Baker, M. Saltik, E. Wagner, and M. Buschle. 1994. Lipo-polysaccharide is a frequent contaminant of plasmid DNA preparations andcan be toxic to primary cells in the presence of adenovirus. Gene Ther.1:239–246.
13. Cotten, M., M. Saltik, M. Kursa, E. Wagner, G. Maass, and M. L. Birnstiel.1994. Psoralen treatment of adenovirus particles eliminates virus replicationand transcription while maintaining the endosomolytic activity of the viruscapsid. Virology 205:254–261.
14. Cotten, M., E. Wagner, K. Zatloukal, and M. L. Birnstiel. 1993. Chickenadenovirus (CELO virus) particles augment receptor-mediated DNA deliv-ery to mammalian cells and yield exceptional levels of stable transformants.J. Virol. 67:3777–3785.
15. Debbas, M., and E. White. 1993. Wild-type p53 mediates apoptosis by E1A,which is inhibited by E1B. Genes Dev. 7:546–554.
15a.De Wet, J., K. Wood, M. DeLuca, D. Helinski, and S. Subramani. 1987.Firefly luciferase gene: structure and expression in mammalian cells. Mol.Cell. Biol. 7:725–737.
16. D’Halluin, J., C. Allart, C. Cousin, P. Boulanger, and G. Martin. 1979.Adenovirus early function required for the protection of viral and cellularDNA. J. Virol. 32:61–71.
17. Evan, G. I., G. K. Lewis, G. Ramsay, and J. M. Bishop. 1985. Isolation ofmonoclonal antibodies specific for human c-myc proto-oncogene product.Mol. Cell. Biol. 5:3610–3616.
18. Ezoe, H., R. Lai Fatt, and S. Mak. 1981. Degradation of intracellular DNAin KB cells infected with cyt mutants of human adenovirus type 12. J. Virol.40:20–27.
19. Folkman, J., and A. Moscona. 1978. Role of cell shape in growth control.Nature 273:345–349.
20. Frisch, S. M., and H. Francis. 1994. Disruption of epithelial cell-matrixinteractions induces apoptosis. J. Cell Biol. 124:619–626.
21. Gagliardini, V., P. A. Fernandez, R. K. Lee, H. C. Drexler, R. J. Rotello,M. C. Fishman, and J. Yuan. 1994. Prevention of vertebrate neuronal deathby the crmA gene. Science 263:826–828.
21a.Gorczyca, W., J. Gong, and Z. Darzynkiewicz. 1993. Detection of DNAstrand breaks in individual apoptotic cells by the in situ terminal deoxynu-cleotidyl transferase and nick translation assays. Cancer Res. 53:1945–1951.
22. Green, S., I. Isseman, and E. Sheer. 1988. A versatile in vivo and in vitroeukaryotic expression vector for protein engineering. Nucleic Acids Res.16:369.
22a.Grimm, S., M. K. A. Bauer, P. A. Baeuerle, and K. Schulze-Osthoff. 1996.Bcl-2 down-regulates the activity of transcription factor NF-kB induced uponapoptosis. J. Cell Biol. 134:13–23.
23. Han, J., P. Sabbatini, D. Perez, L. Rao, D. Modha, and E. White. 1996. TheE1B 19K protein blocks apoptosis by interacting with and inhibiting thep53-inducible and death-promoting Bax protein. Genes Dev. 10:461–477.
24. Henderson, S., M. Rowe, C. Gregory, D. Croom-Carter, F. Wang, R. Long-necker, E. Kieff, and A. Rickinson. 1991. Induction of bcl-2 expression byEpstein-Barr virus latent membrane protein 1 protects infected B cells fromprogrammed cell death. Cell 75:1107–1115.
25. Hunter, J. J., and T. G. Parslow. 1996. A peptide sequence from Bax thatconverts Bcl-2 into an activator of apoptosis. J. Biol. Chem. 271:8521–8524.
26. Hunter, J. J., B. L. Bond, and T. G. Parslow. 1996. Functional dissection ofthe human Bcl2 protein: sequence requirements for inhibition of apoptosis.Mol. Cell. Biol. 16:877–883.
27. Jones, R. F., B. B. Asch, and D. S. Yohn. 1970. On the oncogenic propertiesof chicken embryo lethal orphan virus, an adenovirus. Cancer Res. 30:1580–1585.
28. Kouzarides, T., and E. Ziff. 1988. The role of the leucine zipper in the fos-juninteraction. Nature 336:646–651.
29. Lai Fatt, R. B., and S. Mak. 1982. Mapping of an adenovirus functioninvolved in the inhibition of DNA degradation. J. Virol. 42:969–977.
30. Landschulz, W. H., P. F. Johnson, and S. L. McKnight. 1988. The leucinezipper: a hypothetical structure common to a new class of DNA bindingproteins. Science 240:1759–1764.
31. Lithgow, T., R. Van Driel, J. F. Bertram, and A. Strasser. 1994. The proteinproduct of the oncogene bcl-2 is a component of the nuclear envelope, theendoplasmic reticulum and the outer mitochondrial membrane. Cell GrowthDiffer. 5:411–417.
32. Mancini, L. O., V. J. Yates, V. Jasty, and J. Anderson. 1969. Ependymomasinduced in hamsters inoculated with an avian adenovirus (CELO). Nature222:190–191.
33. Mancini, L. O., V. J. Yates, J. Anderson, and V. Jasty. 1970. CELO virus: anoncogenic virus. Arch. Gesamte Virusforsch. 30:257–260.
34. Meredith, J. E., B. Fazeli, and M. A. Schwartz. 1993. The extracellular matrixas a cell survival factor. Mol. Biol. Cell 4:953–961.
35. Monagan, P., D. Robertson, T. A. Amos, M. J. Dyer, D. Y. Mason, and M. F.Greaves. 1992. Ultrastructural localization of Bcl-2 protein. J. Histochem.Cytochem. 40:1819–1825.
36. Neilan, J. G., Z. Lu, C. L. Afonso, G. F. Kutish, M. D. Sussman, and D. L.Rock. 1993. An African swine fever virus gene with similarity to the proto-
oncogene bcl-2 and the Epstein-Barr virus gene BHRF1. J. Virol. 67:4391–4394.
37. Oltvai, Z. N., and S. J. Korsmeyer. 1994. Checkpoints of dueling dimers foildeath wishes. Cell 79:189–192.
38. Oltvai, Z. N., C. L. Milliman, and S. J. Korsmeyer. 1993. Bcl-2 heterodimer-izes in vivo with a conserved homolog, Bax, that accelerates programmed celldeath. Cell 74:609–619.
39. Pilder, S., J. Logan, and T. Shenk. 1984. Deletion of the gene encoding theadenovirus 5 early region 1B 21,000-molecular-weight polypeptide leads todegradation of viral and host cell DNA. J. Virol. 52:664–671.
40. Plank, C., K. Zatloukal, M. Cotten, K. Mechtler, and E. Wagner. 1992. Genetransfer into hepatocytes using asialoglycoprotein receptor mediated endo-cytosis of DNA complexed with an artificial tetra-antennary galactose ligand.Bioconjugate Chem. 3:533–539.
41. Rao, L., M. Debbas, P. Sabbatini, D. Hockenbery, S. Korsmeyer, and E.White. 1992. The adenovirus E1A proteins induce apoptosis which is inhib-ited by the E1B 19-kDa and Bcl-2 proteins. Proc. Natl. Acad. Sci. USA89:7742–7746.
42. Ruoslahti, E., and J. C. Reed. 1994. Anchorage dependence, integrins, andapoptosis. Cell 77:477–478.
43. Sarma, P. S., R. J. Huebner, and W. T. Lane. 1965. Induction of tumors inhamsters with an avian adenovirus (CELO). Science 149:1108.
43a.Schmitz, M. L., A. Indorf, F. P. Limbourg, H. Stadtler, E. B. M. Traenckner,and P. A. Baeuerle. 1996. The dual effect of adenovirus type 5 E1A 13Sprotein on NF-kB activation is antagonized by E1B 19K. Mol. Cell. Biol.16:4052–4063.
44. Seto, M., U. Jaeger, R. D. Hockett, W. Graninger, S. Bennett, P. Goldman,and S. J. Korsmeyer. 1988. Alternative promoters and exons, somatic mu-tation and deregulation of the Bcl-2-Ig fusion gene in lymphoma. EMBO J.7:123–131.
45. Shen, Y., and T. Shenk. 1994. Relief of p53-mediated transcriptional repres-sion by the adenovirus E1B 19-kDa protein or the cellular Bcl-2 protein.Proc. Natl. Acad. Sci. USA 91:8940–8944.
46. Shen, Y., and T. Shenk. 1995. Viruses and apoptosis. Curr. Opin. Genet.Dev. 5:105–111.
47. Subramanian, T., M. Kuppuswamy, J. Gysbers, S. Mak, and G. Chinnadu-rai. 1984. 19-kDa tumor antigen coded by early region E1b of adenovirus 2is required for efficient synthesis and for protection of viral DNA. J. Biol.Chem. 259:11777–11783.
48. Superti-Furga, G., G. Bergers, D. Picard, and M. Busslinger. 1991. Hor-mone-dependent transcriptional regulation and cellular transformation byFos-steroid receptor fusion proteins. Proc. Natl. Acad. Sci. USA 88:5114–5118.
49. Takayama, S., D. L. Cazals-Hatem, S. Kitada, S. Tanaka, T. Miyashita, L. R.Hovey, D. Huen, A. Rickinson, P. Veerapandian, S. Krajewski, K. Saito, andJ. C. Reed. 1994. Evolutionary conservation of function among mammalian,avian, and viral homologues of the Bcl-2 oncoprotein. DNA Cell Biol. 13:679–692.
50. Takemori, N., C. Cladaras, B. Bhat, A. Conley, and W. Wold. 1984. cyt geneof adenovirus 2 and 5 is an oncogene for transforming function in earlyregion E1B and encodes the E1B 19,000-molecular-weight polypeptide.J. Virol. 52:793–805.
51. Takemori, N., J. L. Riggs, and C. Aldrich. 1968. Genetic studies with tumor-igenic adenoviruses. I. Isolation of cytocidal (cyt) mutants of adenovirus type12. Virology 36:575–586.
52. Tarodi, B., T. Subramanian, and G. Chinnadurai. 1993. Functional similar-ities between adenovirus E1B 19K gene and Bcl-2 oncogene: mutant comple-mentation and suppression of cell death induced by DNA damaging agents.Int. J. Oncol. 3:467–472.
53. Tarodi, B., T. Subramanian, and G. Chinnadurai. 1994. Epstein-Barr virusBHRF-1 protein protects against cell death induced by DNA-damagingagents and heterologous viral infection. Virology 201:404–407.
54. Wagner, E., K. Zatloukal, M. Cotten, H. Kirlappos, K. Mechtler, D. Curiel,and M. L. Birnstiel. 1992. Coupling of adenovirus to transferrin-polylysine/DNA complexes greatly enhances receptor-mediated gene delivery and ex-pression of transfected cells. Proc. Natl. Acad. Sci. USA 89:6099–6103.
55. Wagner, E., M. Cotten, K. Mechtler, H. Kirlappos, and M. L. Birnstiel. 1991.DNA-binding transferrin conjugates as functional gene-delivery agents: syn-thesis by linkage of polylysine or ethidium homodimer to transferrin carbo-hydrate moiety. Bioconjugate Chem. 2:226–231.
56. Weinberg, D. H., and G. Ketner. 1983. A cell line that supports the growthof a defective early region 4 deletion mutant of human adenovirus type 2.Proc. Natl. Acad. Sci. USA 80:5383–5386.
57. White, E. 1994. Function of the adenovirus E1B oncogene in infected andtransformed cells. Semin. Virol. 5:341–348.
58. White, E., S. H. Blose, and B. Stillman. 1984. Nuclear envelope localizationof an adenovirus tumor antigen maintains the integrity of cellular DNA. Mol.Cell. Biol. 4:2865–2875.
59. White, E., and R. Cipriani. 1990. Role of adenovirus E1B proteins in trans-formation: altered organization of intermediate filaments in transformedcells that express the 19-kilodalton protein. Mol. Cell. Biol. 10:120–130.
3176 CHIOCCA ET AL. J. VIROL.
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

60. White, E., R. Cipriani, P. Sabbatini, and A. Denton. 1991. Adenovirus E1B19-kilodalton protein overcomes the cytotoxicity of E1A proteins. J. Virol.65:2968–2978.
61. White, E., T. Grodzicker, and B. W. Stillman. 1984. Mutations in the geneencoding the adenovirus early region 1B 19,000-molecular-weight tumorantigen cause the degradation of chromosomal DNA. J. Virol. 52:410–419.
62. White, E., P. Sabbatini, M. Debbas, W. S. M. Wold, D. I. Kusher, and L. R.Gooding. 1992. The 19-kilodalton adenovirus E1B transforming protein in-hibits programmed cell death and prevents cytolysis by tumor necrosis factoralpha. Mol. Cell. Biol. 12:2570–2580.
63. Williams, G. T., and C. A. Smith. 1993. Molecular regulation of apoptosis:genetic controls on cell death. Cell 74:777–779.
64. Yin, X. M., Z. N. Oltvai, and S. J. Korsmeyer. 1994. BH1 and BH2 domainsof Bcl-2 are required for inhibition of apoptosis and heterodimerization withBax. Nature 369:321–323.
65. Zhang, Y., and R. J. Schneider. 1994. Adenovirus inhibition of cell transla-tion facilitates release of virus particles and enhances degradation of thecytokeratin network. J. Virol. 68:2544–2555.
66. Zhu, H., Y. Shen, and T. Shenk. 1995. Human cytomegalovirus IE1 and IE2proteins block apoptosis. J. Virol. 69:7960–7970.
VOL. 71, 1997 CELO ANTIAPOPTOSIS GENE 3177
on March 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from