identification of gtp- binding proteins in myelin and oligodendrocyte membranes
TRANSCRIPT

Journal of Neuruscience Research 26: 16-23 (1990)
Identification of GTP-Binding Proteins in Myelin and Oligodendrocyte Membranes P.E. Braun, E. Horvath, V.W. Yong, and L. Bernier Departments of Biochemistry (P.E.B., E.H.) and Neurology and Neurosurgery (P.E.R., V.W.Y., L.B.), McGill University, Montreal, Quebec, Canada
Myelin membranes purified from mouse and rat brain are associated with alpha subunits of four sig- nal transducing guanosine triphosphate (CTP)- binding proteins: Go, Gi, G,, and ras. Four low-mo- lecular-weight (M,) GTP-binding proteins are also present, as demonstrated by the binding of GTP to proteins immobilized in nitrocellulose. This latter group is more prominent at early stages of myelina- tion and remains associated with isolated myelin membranes despite repetitive cycles of purification. At least one nonmyelin subcellular membrane frac- tion possesses the same proteins. The total membrane fraction of cultured oligodendrocytes is associated with both groups of GTP-binding proteins. None of the well-known myelin proteins bound GTP by the procedure described.
Key words: G proteins, GTP-binding proteins, ADP-ribosylation, myelin proteins, oligodendrocyte, myelinogenesis, muscarinic receptor
INTRODUCTION
A number of observations were recently made con- cerning thc presence in isolated myelin of cellular and catalytic components that do not accord with the conven- tional, accepted view of what role the myelin sheath has to play in the nervous system. Among these, we note the presence of cholinergic receptors and adenylate cyclase in highly purified central nervous system (CNS) myelin (Larocca et al., 1987), stimulation by acetylcholine (ACh) of phosphoinositide metabolism in CNS myelin (Kahn and Morell, 1988), evidence for muscarinic re- ceptor-stimulated metabolism of polyphosphoinositides in PNS myelin (Eichberg et al., 1989), regulated phos- phorylationidephosphorylation of myelin basic proteins (Ulmer et al., 1987), and myelin-associated glycoprotein (Edwards et al., 1988). Implicit in some or all these events is the possible indirect participation of guanosine triphosphate (GTP)-binding proteins, a potential mecha- nism that has been generally recognized by the contrib- utors of the above observations. The recent, unexpected discovery that myelin basic protein might serve as a
GTP-binding protein (Chan et al.. 1988) and that 2l.3'- cyclic nucleotide 3 '-phosphodiesterase (CNP) posses5es domains that share sequence homology with authentic G proteins (Bernier et al., 1989; Sprinkle and Hancock, 1989) has served to direct our attention to GTP-mediated events in myelination other than cell-surface-mediated signal transduction.
We report here a survey of the membrane associ- ated GTP-binding proteins that are present in purified brain myelin and in membranes of cultured oligodendro- cytes obtained from brains at an early stage of myelina- tion. We show that alpha-subunits of three major signal transducing G proteins as well as the proto-oncogene product ras are associated with these membranes. In ad- dition, we describe a class of low-molecular-weight pro- teins that bind GTP when they arc electroblotted onto nitrocellulose. Proteins of this type have been observed in other membranes (Bhullar and Haslam, 1987; 1988; Comerford et a l . , 1989) and may represent a group of GTP-binding proteins that subserve a role in the media- tion of vectorial transport processes (Bourne, 1988) in- tegral to membrane assembly and turnover. A prelimi- nary report of some of these findings has been presented (Braun and Bernier, 1989).
MATERIALS AND METHODS Identification of G, Proteins
Proteins that bind GTP when they are immobilized on nitrocellulose were detected by the procedure of Bh- ullar and Haslam ( 1 987) with minor modifications. Membrane proteins, separated by SDS-PAGE (Laemnili, 1970) in minigels of 10 or 15% acrylamide were elec- trophoretically transferred to nitrocellulose (Ni troscreen) for 60-70 min at 100 V. This electroblot was then incu- bated for 15 min in 50 mM Tris buffer, pH 7.5. contain- ing 50 mM MgCl,, 10 FM ATP (as a nonspecific
Recelled October 23, 1989, accepted November 9, 1989
Address reprint requests to Dr. Peter E Braun, Departments of Bio- chemistry and Neurology and Neurosurgery, McGill University. Mon- treal, Quebcc H3G 146, Canada.
G 1990 Wiley-Liss, Inc.
GTP-Binding Proteins in Myelin 17
blocking agent), and 0.3% Tween 20. It was transferred to the same buffer, with the addition of the isotope (2 pCi of a3’P-GTP; 4 pCi of y3’S-GTP), and incubated for 60 min with gentle shaking at room temp. It was then washed twice with the same buffer for 15 min each. In the case of y3’S-GTP, all buffers also contained 1 niM dithiothreitol. After air drying the blot was exposed for 24-48 hr to Kodak X-OMAT-AR film with a screen for 32P. a3*P = GTP (3,000 Ciimmol) and Y ~ ~ S - G T P (1 300 Ci/mmol) were purchased from New England Nuclear.
Identification of Proteins by Jmmunostaining A modification of the immunoelectroblotting pro-
cedure of Towbin et al. (1 979) was used to identify spe- cific proteins. We followed this protocol as detailed by Promega Corp. in their 1988 catalog, section 6, pp 15- 16. Antisera to CNP and MBP have been previously described (Bernier et al.. 1987; Ulmer and Braun. 1983). Polyclonal antibodies to alpha-subunits of G, , Gi, and G, were a generous gift from Dr. Allen M. Spiegel, Na- tional Institutes of Health, Bethesda. Anti-ras p 21 (pan) was provided by DuPont Corp.
Preparation of Membranes Mouse and rat brain myelin wa5 isolated by the
procedure of Norton and Poduslo (1973). Primary cul- tures of oligodendrocytes were prepared from myelinat- ing rat brain at d19 and purified by the procedure of Yong et al. (1988). More than 90% of the final cell preparations were oligodendrocytes as judged by antiga- lactocerebroside immunocytocheniistry. A crude mem- brane fraction of thesc cells was obtained as follows. After removal of culture medium 18 X lo6 cells in dishes were washed once with HEPES buffer (20 mM, pH 7.5) containing KCI ( 3 mM), MgCl, ( 3 mlz?) and protease inhibitors. Cells were then scraped into the same buffer (10 ml total) and homogenized by hand (15 strokes of the Dounce pestle). Large cell particulates and nuclei were removed by centrifugation at 3,000g, for 10 min, and total membranes were recovered from the su- pernatant by centrifugation for 30 min at 100,000g. The membrane-containing pellet was dispersed in sample buffer in preparation for SDS-PAGE.
ADP-Ribosylation of Membrane Proteins We used a modification of the procedure of Bokoch
et al. (1983). Briefly, membranes (200 pg protein) were incubated at 37°C with activated cholera toxin (2.5 pg) or pertussis toxin (2 pg) in 200 p1 of a buffer containing 100 mM potassium phosphate (pH 7.5) , 10 mM thymi- dine, 1 mM ADP, 0.1 mM GTP, 4 niM ADP-ribose, 2.5 mM MgCI,, 1 mM dithiothreitol and 0.05% Triton X-100. The reaction was initiated by the addition of
-29 29-
L 14-
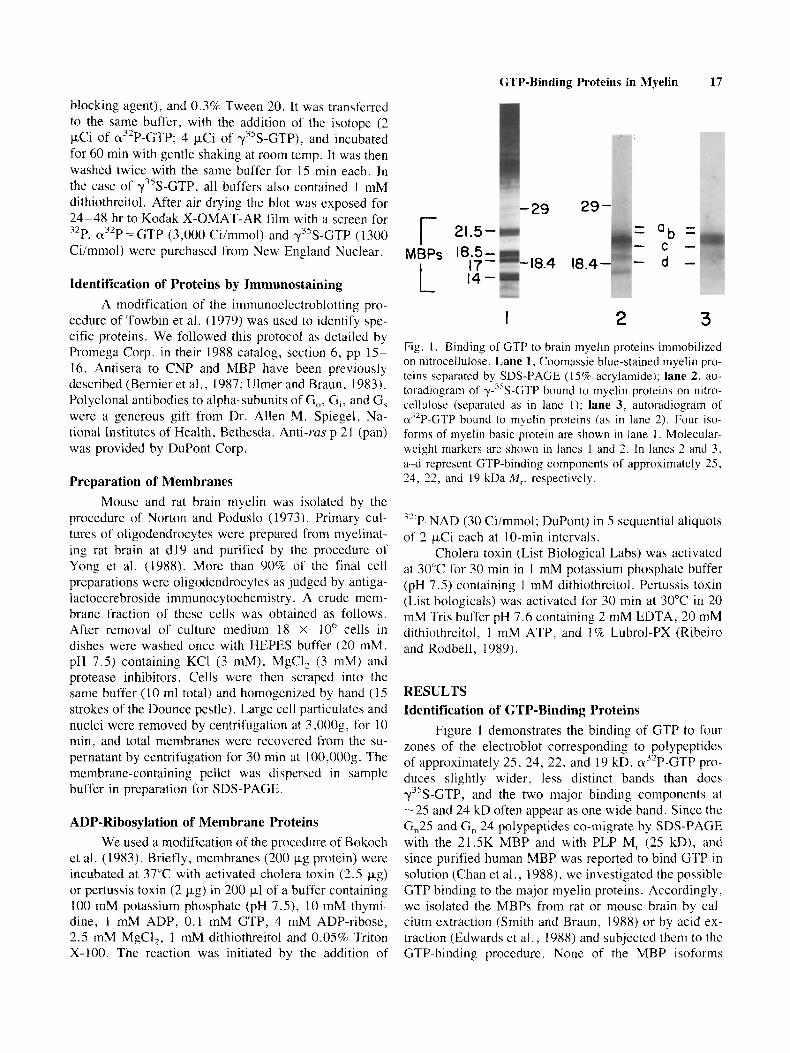
1 2 3 Fig. I . Binding of GTP to brain myelin proteins immobilized on nitrocellulose. Lane 1, Coomassie blue-stained myelin pro- teins separated by SDS-PAGE (15% acrylamide); lane 2, au- toradiograrn of y-3sS-GTP bound to myelin proteins on nitro- cellulose (separated as in lane 1); lane 3, autoradiogram of a”P-GTP bound to myelin proteins (as in lane 2). Four isu- forms of myelin basic protein are shown in lane 1 . Molecular- weight markers are shown in lanes 1 and 2. In lanes 2 and 3, a-cl represent GTP-binding components of approximately 25, 24, 22, and 19 kDa M,, respectively.
’*.P-NAD (30 Ci/mmol; DuPont) in 5 scquential aliquots of 2 FCi each at 10-min intervals.
Cholera toxin (List Biological Labs) was activated at 30°C for 30 min in 1 mM potassium phosphate buffer (pH 7.5) containing 1 mM dithiothreitol. Pertussis toxin (List bologicals) was activated for 30 min at 30°C in 20 mM Tris buffer pH 7 .6 containing 2 mM EDTA, 20 mM dithiothreitol, 1 mM ATP. and 1% Lubrol-PX (Ribeiro and Rodbell, 1989).
RESULTS Identification of CTP-Binding Proteins
Figure 1 demonstrates the binding of GTP to four zones of the electroblot corresponding to polypeptides of approximately 25, 24, 22, and 19 kD. a3’P-GTP pro- duces slightly wider, less distinct bands than does y”S-GTP, and the two major binding components at -2.5 and 24 kD often appear as one wide band. Since the G,25 and G, 24 polypeptides co-migrate by SDS-PAGE with the 21.5K MBP and with PLP M, (25 kD), and since purified human MBP was reported to bind GTP in solution (Chan et al., 1988), we investigated the possible GTP binding to the major myelin proteins. Accordingly, we isolated the MBPs from rat or mouse brain by cal- cium extraction (Smith and Braun, 1988) or by acid ex- traction (Edwards et al., 1988) and subjected them to thc GTP-binding procedure. None of the MBP isoforms
18 Braun et al.
43 -
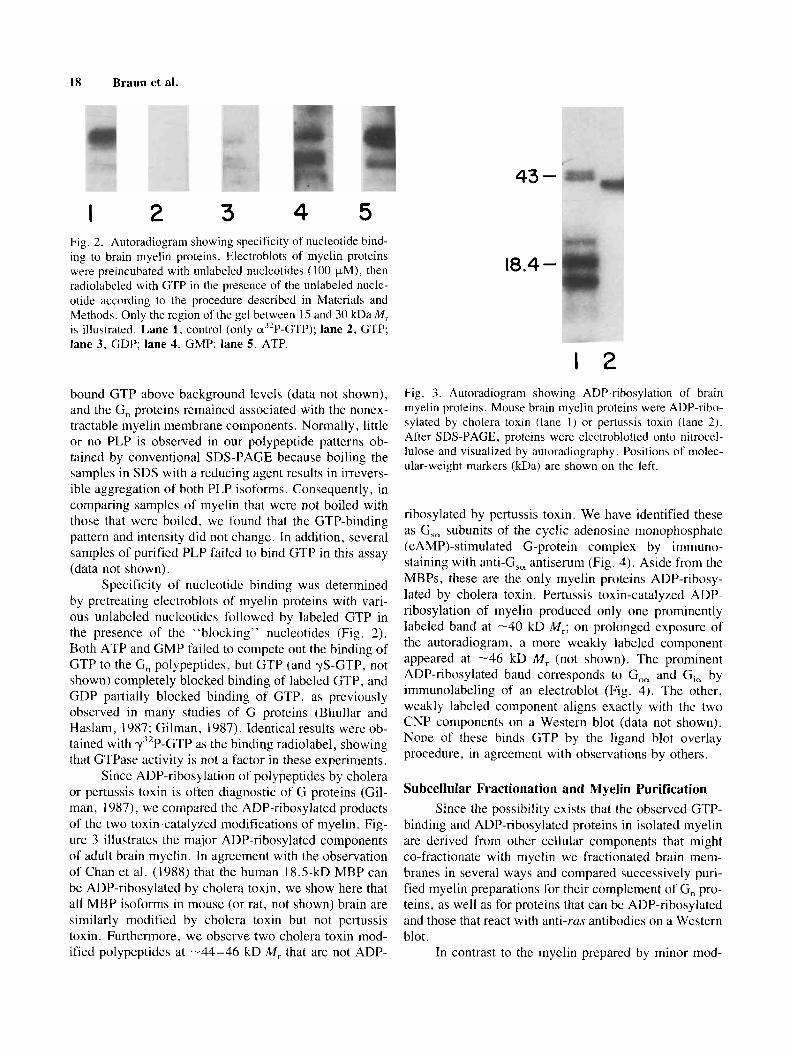
I 2 3 4 5 Fig. 2. Autoradiogram showing specificity of niicleotide bind- ing to brain myelin proteins. Electroblots of myelin proteins were preincubated with unlabeled nucleotjdes (100 pM), then radiolabeled with GTP in the prescnce of the unlabeled nucle- otide according to the procedure described in Materials and Methods. Only the region of the gel between 15 and 30 kDaM, is illustrated. Lane 1, contrul (only d2P-GTP); lane 2, GTP; lane 3, GDP: lane 4: GMP: lane 5 , ATP.
bound GTP above background levels (data not shown), and the G, proteins remained associated with the nonex- tractable myelin membrane components. Normally, little or no PLP is observed in our polypeptide patterns ob- tained by conventional SDS-PAGE because boiling the samples in SDS with a reducing agent rcsults in irrevers- ible aggregation of both PLP isoforms. Consequently, in comparing samples of myelin that were not boiled with those that were boiled, we found that the GTP-binding pattern and intensity did not change. In addition, several samples of purified PLP failed to bind GTP in this assay (data not shown).
Specificity of nucleotide binding was determined by pretreating electroblots of myelin proteins with vari- ous unlabeled nucleotides followed by labeled GTP in the presence of the “blocking” nucleotides (Fig. 2). Both ATP and GMP failed to compete out the binding of GTP to the G, polypeptides, but GTP (and yS-GTP, not shown) completely blocked binding of labeled GTP, and GDP partially blocked binding of GTP, as previously observed in many studies of G proteins (Bhullar and Haslam, 1987; Gilinan, 1987). Identical results were ob- tained with y’*P-GTP as the binding radiolabel, showing that GTPase activity is not a factor in these experiments.
Since ADP-ribosylation of polypeptides by cholera or pertussis toxin is often diagnostic of G proteins (Gil- man, 1987), we compared the ADP-ribosylated products of the two toxin-catalyzed modifications of myelin. Fig- ure 3 illustrates the major ADP-ribosylated components of adult brain myelin. In agreement with the observation of Chan et al. (1988) that the human 18.5-kD MBP can be ADP-ribosylated by cholera toxin, we show here that all MBP isoforms in mouse (or rat, not shown) br.. din are similarly modified by cholera toxin but not pertussis toxin. Furthermore, we observe two cholera toxin mod- ified polypeptides at -44-46 kD M, that are not ADP-
18.4 -
1 2 Fig. 3 . Autoradiogram showing ADP-ribosylation of brain myelin proteins. Mouse brain myelin proteins were ADP-ribo- sylated by cholera toxin (lane I ) or pertussis toxin (lane 2). After SDS-PAGE, proteins were electroblotted onto nitroccl- lulose and visualized by autoradiography . Positions of molec- ular-weight markers (kDa) are shown on the left.
ribosylated by pertussis toxin. We have identified these as G,, subunits of the cyclic adenosine monophosphatc (CAMP)-stimulated G-protein complex by immuno- staining with anti-G,, antiserum (Fig. 4). Aside from the MBPs, these are the only myelin proteins ADP-ribosy- lated by cholera toxin. Pertussis toxin-catalyzed ADP- ribosylation of myelin produced only one prominently labeled band at -40 kD iM,; on prolonged exposure of the autoradiogram! a more weakly labeled component appeared at -46 kD M , (not shown). The prominent ADP-ribosylated band corresponds to G,, and Gi, by immunolabeling of an electroblot (Fig. 4). The other, weakly labeled component aligns exactly with the two CNP Components on a Western blot (data not shown). None of these binds GTP by the ligand blot overlay procedure, in agreement with observations by others.
Subcellular Fractionation and Myelin Purification Since the possibility exists that the observed GTP-
binding and ADP-ribosylated proteins in isolated myelin are derived from other cellular components that might co-fractionate with myelin we fractionated brain mem- branes in several ways and compared successively puri- fied myelin preparations for their complement of G, pro- teins, as well as for proteins that can be ADP-ribosylated and those that react with anti-ras antibodies on a Western blot.
In contrast to the myelin prepared by minor mod-
GTP-Binding Proteins in Myelin 19
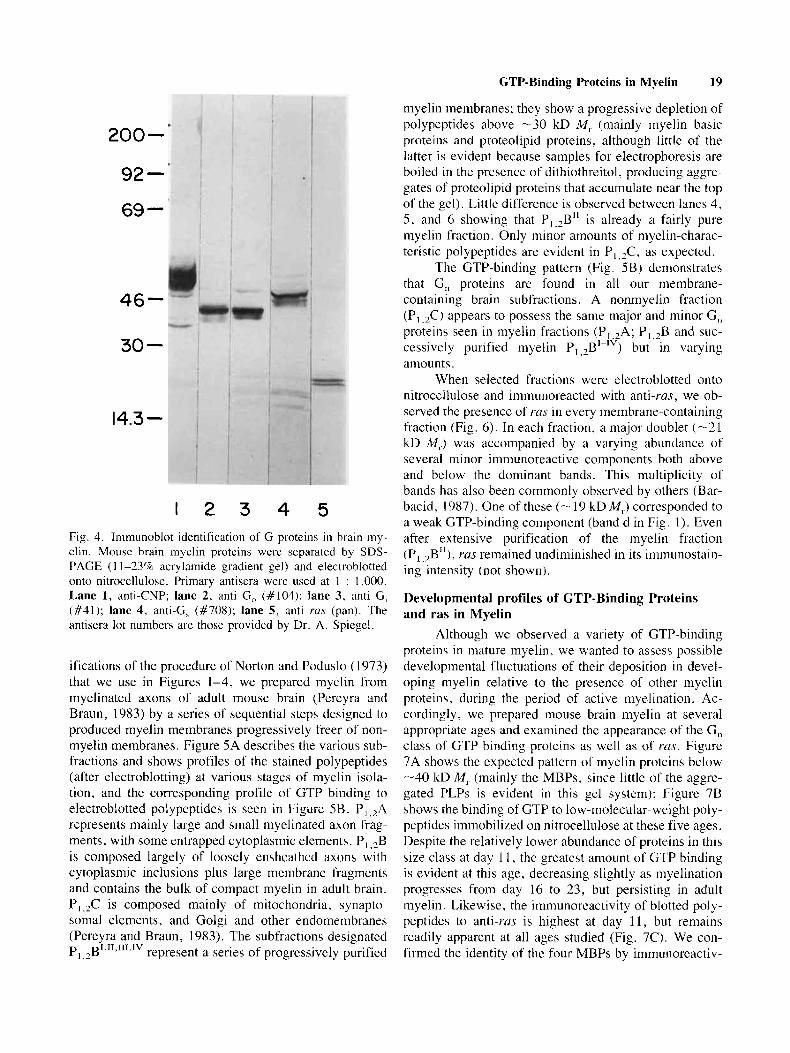
myelin membranes; they show a progressive depletion of polypcptides above -30 kD M , (mainly myelin basic proteins and proteolipid proteins, although little of the latter is evident because samples for electrophoresis are boiled in the presencc of dithiothreitol, producing aggrc- gates of proteolipid proteins that accumulate near the top of the gel). Littlc difference is observed between lanes 4, 5 , and 6 showing that Pl,2B" is already a fairly pure myelin fraction. Only minor amounts of niyelin-charac- teristic polypeptides are evident in P I $, as expectcd.
The GTP-binding pattern (Fig. SB) demonstrates that G,, proteins are found in all our membrane- containing brain subfractions. A nonmyelin fraction (PI ,&) appears to possess the same ma.jor and minor C, proteins seen in inyelin fractions (P,.,A; P,,,B and suc- cessivcly purified myelin P, , $ - I V ) but in varying amounts.
When selected fractions were electroblotted onto nitroccllulose and immunoreacted with anti-ras, we ob- served the prescncc of ras in every membrane-containing fraction (Fig. 6). In each fraction, a major doublet (-21 kI) M,.) was accompanied by a varying abundance of several minor imniunoreactive components both above and bclow the dominant bands. This multiplicity of bands has also been commonly observed by others (Bar- bacid, 1987). One of these (- 19 kD M , ) corresponded to a weak GTP-binding component (band d in Fig. 1). Even after extensive purification of the myelin fraction (P, ,2B11), ras remained undiminished in its imniunostain- ing intensity (not shown).
Developmental profiles of GTP-Binding Proteins and ras in Myelin
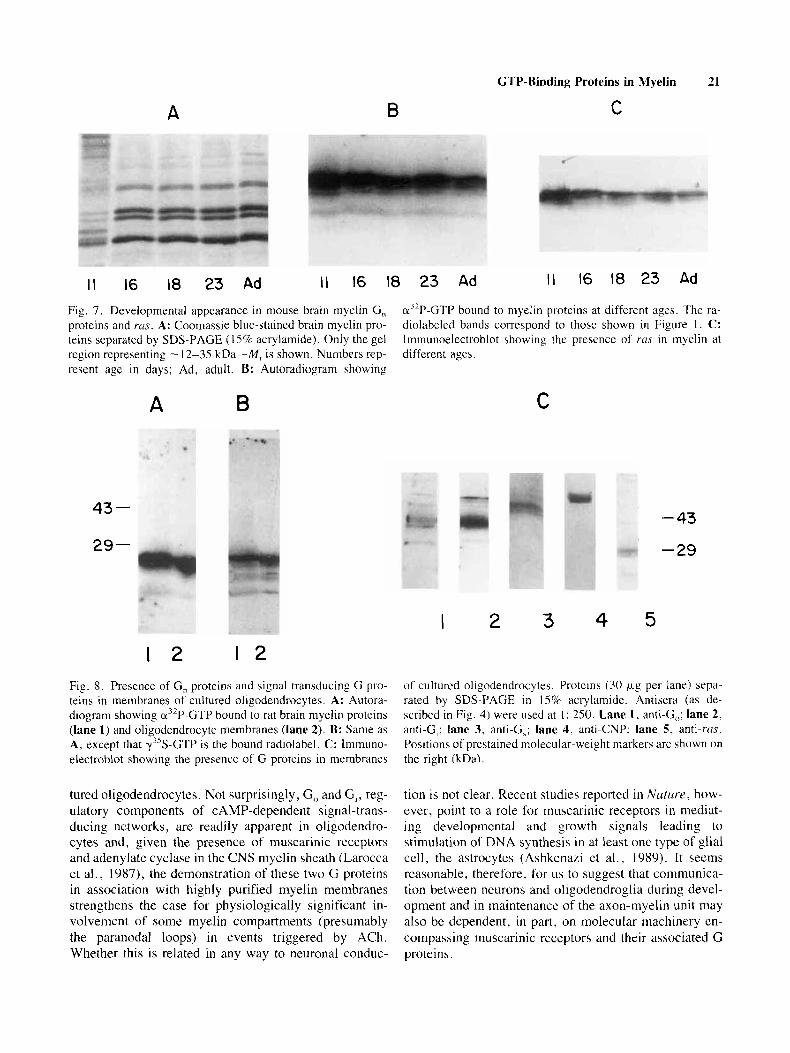
Although we observed a variety of GTP-binding proteins in mature myelin. we wanted to assess possible developmental fluctuations of their deposition in devel- oping myelin relative to the presencc of other myelin proteins, during the period of active myelination. Ac- cordingly, we prepared mouse brain myelin at several appropriate ages and examined thc appearance of the G, class of GTP-binding proteins as well as of ras. Figure 7A shows the expected pattern of myelin proleins below -40 kD M , (mainly the MBPs, since little of the aggre- gated PLPs is evident in this gel systcm); Figure 7B shows the binding of GTP to low-molecular-weight poly- peptides immobilized on nitrocellulose at these five ages. Despite the relatively lower abundancc of proteins in this size class at day 1 I , the greatest amount of GTP binding is evident at this age, decreasing slightly as myelination progresses from day 16 to 23, but persisting in adult myelin. Likewise, the immunoreactivity of blotted poly- peptides to anti-rus is highest at day 11, but remains readily apparent at all ages studied (Fig. 7C). We con- firmed the identity of the four MBPs by immunoreactiv-
200-
92-
69 -
46-
30 -
14.3 -
1 2 3 4 5 Fig. 4. Inimunoblot identification of G proteins in brain my- elin. Mouse brain myelin proteins were separated by SDS- PAGE ( 1 1-2370 acrylarnide gradient gel) and electrublotted onto nitrocellulose. Primary antisera were used at 1 : 1,000. Lane 1, anti-CNP; lane 2, anti G, (#104): lane 3, anti G, (#41); lane 4, anti-G, (#708); lane 5, anti rus (pan). The antisera lo t numbcrs are thosc provided by Dr. A. Spiegel.
ifications of the procedure of Norton and Poduslo (1973) that we use in Figures 1-4, we prepared myelin from myelhated axons of adult mouse brain (Pereyra and Braun, 1983) by a series of sequential steps designed to produced m y e h membranes progressively freer of non- myelin membranes. Figure 5A describes the various sub- fractions and shows profiles of the stained polypeptides (after electroblotting) at various stages of myelin isola- tion, and the corresponding profile of GTP binding to electroblotted polypeptides is seen in Figure SB. P, ,?A rcpresents mainly large and small myelinated axon frag- ments, with some entrapped cytoplasmic elements. PI ,*B is cornposed largely of loosely ensheathed axons with cytoplasmic inclusions plus large membrane fragments and contains the bulk of compact myelin in adult brain. P I ,,C is composed mainly of mitochondria, synapto- soma1 elements, and Golgi and other endomembranes (Pereyra and Braun, 1983). The subfractions designated
represent a series of progressively purified p ~','I,"l,lv 1.2
20 Braun et al.
A B
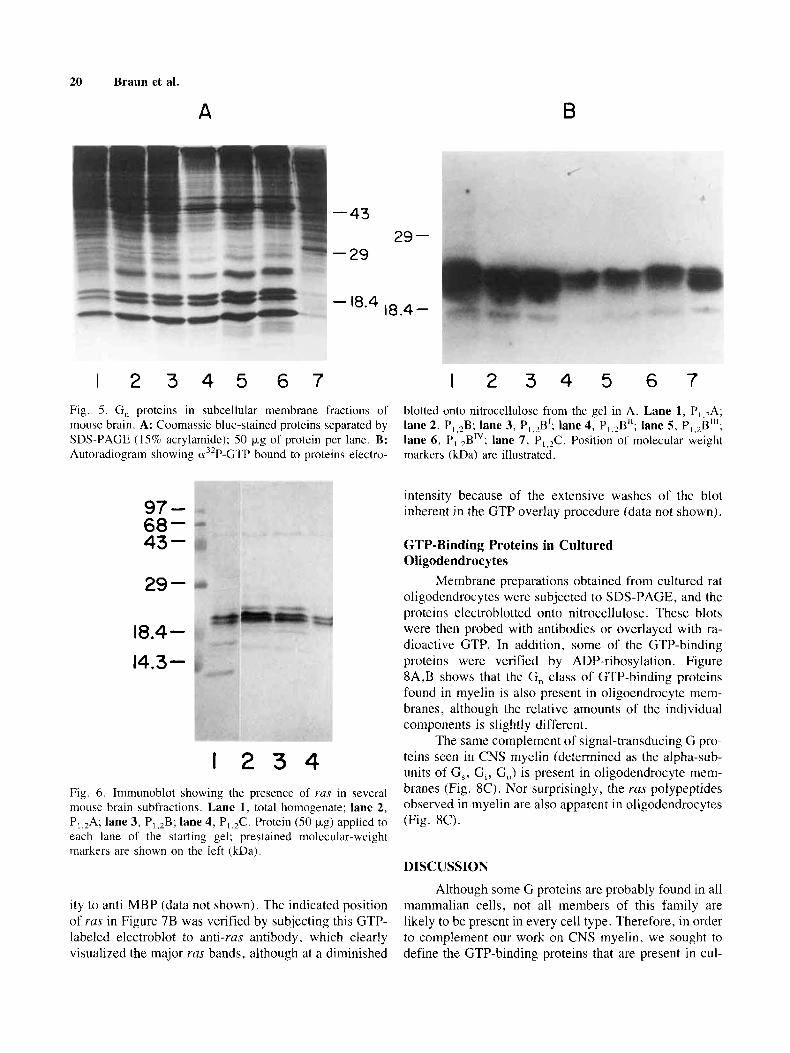
- 43
- 29 29 -
I 2 3 4 5 6 7 1 2 3 4 5 6 7 Fig. 5. G , proteins in subcellular membrane fractions of mouse brain. A: Coomassie blue-stained proteins separated by SDS-PAGE (15% acrylamide); 50 tJ.g of protein per lane. R: Autoradiogram showing u3'P-GTP bound to proteins electro-
blotted onto nitrocellulose from the gel in A. Lane 1, P,,,A; lane 2, P,,,B; lane 3, P,,,B1; lane 4, P,,,B"; lane 5, P,,2B11'; lane 6, P,,,BTV; lane 7. P,,,C. Position of molecular weight markers (kDa) are illustrated.
97 - 68 - 43 - 29 -
18.4 - 14.3-
I 2 3 4 Fig. 6. Immunoblot showing the presence of YUS in several mouse brain subfractions. Lane 1, total homogenate; lane 2, P,,2A; lane 3, P,,,B; lane 4, P,,,C. Protein (SO kg) applied to each lane of the starting gel; prestained molecular-weight markers are shown on the left (kDa).
intensity because of the extensive washes of the blot inherent in the GTP overlay procedure (data not shown).
GTP-Binding Proteins in Cultured Oligodendrocytes
Membrane preparations obtained from cultured rat oligodendrocytes were subjected to SDS-PAGE, and the proteins electroblotted onto nitrocellulose. These blots were then probed with antibodies or overlayed with ra- dioactive GTP. In addition, some of the CTP-binding proteins were verified by ADP-ribosylation. Figure XA,B shows that the G, class of GTP-binding proteins found in myelin is also present in oligoendrocyte mem- branes. although the relative amounts of the individual components is slightly different.
The same complement of signal-transducing G pro- teins seen in CNS myelin (determined as the alpha-sub- units of G,, G,, GJ is present in oligodendrocyte mem- branes (Fig. SC). Nor surprisingly, the ruus polypeptides observed in myelin are also apparent in oligodcndrocytes (Fig. SC).
IIISCUSSION
Although some G proteins are probably found in all mammalian cells, not all members of this family are likely to be present in every cell type. Therefore, in order to complement our work on CNS myelin, we sought to define the GTP-binding proteins that are present in cul-
ity to anti-MBP (data not shown). The indicated position of rus in Figure 7B was verified by subjecting this GTP- labeled electroblot to anti-ras antibody, which clearly visualized the major ras bands, although at a diminished
A
GTP-Binding Proteins in Myelin 21
C
II 16 18 23 Ad
B
II I6 18 23 Ad II I6 18 23 Ad
Fig. 7. Developmental appearance in mouse brain myelin G , proteins and ms. A: Coomassie blue-stained brain myelin pro- teins separated by SDS-PAGE ( 15% acrylamide). Only the gel region representing - 12-35 kDa-M, is shown. Numbers rep- resent age in days; Ad, adult. B: Autoradiogram showing
cr3’P-GTP bound to myelin proteins at different ages. The ra- diolabeled bands correspond to those shown in Figure 1. C: Imniunoelectroblot showing the presence of rus in myelin at different ages.
A B
43 -
29 -
I
C
2 3 4
- 43
-29
5 1 2 1 2
Fig. 8. Presence of G, proteins and signal transducing G pro- teins in membranes of cultured oligodendrocytes. A: Autora- diogram showing a3’P-GTP bound to rat brain myelin proteins (lane I) and oligodendrocyte membranes (lane 2). R: Same as A, except that y%GTP is the bound radiolabel. C: Immuno- electroblot showing the presence of G proteins in membranes
of cultured oligodendrocytes. Proteins (30 k*.g per lane) sepa- rated by SDS-PAGE in 15% acrylamide. Antisera (as de- scribed in Fig. 4) were used at 1 : 250. Lane 1, anti-G,,: lane 2, anti-G,; lane 3, anti-G,: lane 4. anti-CNP; lane 5, anti-ras. Positions of prestained molecular-weight markers are shown on the right (kDa).
tured oligodendrocytes. Not surprisingly, G, and G,, reg- ulatory components of CAMP-dependent signal-trans- ducing networks, are readily apparent in oligodendro- cytes and, given the presence of muscarinic receptors and adenylate cyclase in the CNS myelin sheath (Larocca et al., 1987), the demonstration of these two G proteins in association with highly purified myelin membranes strengthens the case for physiologically significant in- volvement of some myelin compartments (presumably the paranodal loops) in events triggered by ACh. Whether this is related in any way to neuronal conduc-
tion is not clear. Recent studies reported in N a t u r ~ . how- ever, point to a role for muscarinic receptors in mediat- ing developmental and growth signals leading to stimulation of DNA synthesis in at least one type of glial cell, the astrocytes (AshkcnaLi et al., 1989). It seems reasonable, therefore. for us to suggest that comniunica- tion between neurons and oligodendroglia during devel- opment and in maintenance of the axon-myelin unit may also be dcpendent, in part, on molecular machinery en- compassing muscarinic receptors and their associated G proteins.

22 Braun et al.
Although the function of G,, protein may overlap that of other G proteins (Spiegel, 1987), there are indi- cations that it may regulate ion channels (Freissmuth et al., 1989), in addition to other functions it might have. Since not all cell types possess this G protein (Gilman, 1987), we think it is significant that oligodendrocytes do and that the association of this protein with myelin mem- branes persists despite numerous procedures designed to rid myelin of adventitiously associated cellular elements. Participation of G, in thc regulation of ion channels and its putative linkage to the action of inositol phospholipids in signal transduction accords with current developments in studies of polyphosphoinositide metabolism in para- nodal loops (Kahn and Morell, 1988; Eichberg et al., 1989) and of ion channels in oligodcndroglia (Banes et al., 1988).
The proto-oncogene product, r m , is also thought to participate in the inositol phospholipid-linked mode of signal transduction (Barbacid, 1987; Kamata and Kung, 1988), although its prccisc role is not clear. The associ- ation of ras with isolated and purified myelin helps sup- port the argument for signal responsive receptors or ion channcls in paranodal compartments of the myelin sheath.
Numerous small (<27 kD M,) CTP-binding proteins have been described recently. Bhullar and Haslam ( I 987) referred to one group of thesc as G, because they are observed to bind GTP when the polypeptides are immobilized on nitrocellulose by electrophoretic transfer from gels. Using this GTP-overlay technique, we found that at least four polypeptides in both CNS myelin and oligodendrocyte membranes can be separated by SDS- PAGE and, when blotted onto nitroccllulose, they bind labeled GTP selectively, although GDP appears (by competition with GTP) to bind to some extent. The two most prominent of these at -24-25 kD M, may corre- spond to the major G, proteins in platelets (Bhullar and Haslam, 1987; 1988) or other G, proteins observed in various cell lines (Comerford et al., 1989). The persis- tence of these components in myelin, in apparently un- diminished abundance relative to other myelin proteins, judging by their GTP-binding capacity, leads us to sug- gest that they are associated with the myelin sheath in its native state. The major ras polypeptides that we ob- served at -2 1 kD M, did not bind appreciable amounts of GTP, although Comerford et al. (1989) and others report that the various YUS isoforms are indeed G,, proteins. One of the minor bands that was observed by immunochem- ical staining on an electroblot did, in fact, correspond to one of the minor GTP-binding polypeptides but identity cannot be assured. Neither pertussis nor cholera toxin catalyzed ADP-ribosylation of these G, proteins; botuli- num toxin was not tried, although some investigators have reported that this toxin modifies a small GTP-
binding protein from brain (Kikuchi et al., 1988). The relatively greater abundance of the G, proteins in myelin at an early stage of myelination (days 11 and 16) sug- gests that these proteins may have a role to play in the processes by which meylin membrane components are transported. targeted, and assembled into myelin. In this regard, it has becn suggested that a family of low-mo- lecular-weight GTP-binding proteins (with GTPase ac- tivity) may participate in the mediation of intracellular membrane traffic in exocytosis (Bourne, 1988) and en- docytosis (Mayorga et al., 1989). Other functions related to early stages of niyelinogenesis are not excluded. In other systems, e.g. , platelets, it has been suggested that the G, class or GTP-binding proteins may participatc in morphological alterations and cellular secretion (Nagota et al., 1989). The developmental pattern of ras appear- ance in myelin parallels that of the G, proteins and we surmise that it, too, may be relatively more important at early stages of myelinogcnesis.
We found that the myelin basic proteins failed to bind GTP specifically by the overlay approach. By con- tast, one of thc human MHP isoforins, in solution, was reported to bind 8-azido-CTP by photolabeling (Chan et al., 1988). Evidently, the binding domains do not sur- vive the denaturation that attends SDS-PAGE, a property also shared by some of the signal-transducing G proteins.
ACKNOWLEDGMENTS
This work was supported by grants from the Med- ical Rebearch Council and Multiple Sclerosis Society of Canada. L.B. is a fellow of the MS Society.
REFERENCES Ashkenazi A, Ramachandran J , Capon DJ ( 1989): Acetylcholine an-
alog stimulates DNA synthesis in brain-derived cells via spe- cific muscarinic receptor subtypes Nature (Lond) 340: 146-150.
Barbacid M (1987): ros gcncs. Annu Rev Biochem 56:779-828. Barres BA, Chun LLY, Corey DP (1988): Ion channel expression by
white matter glia: Type 2 astrocytes and oligodendrocytes. (ilia 1:10-30.
Bernier L, Alvarez F, Norgard EM, Raiblc DW, Mentabcrry A, Schembri JG. Sabatini DD, Colman DR (1987): Molecular cloning of a 2’. 3’ cyclic nucleolide 3’-phosphodiesterase: mR- NAs with different 5’-ends encode the same set of proteins in nervous and lymphoid tissues. J Neuroscience 7:2703 -27 10.
Bcrnicr L , Horvath E, Braun P ( 1 989): GTP binding proteins in CNS myclin. Trans Am Soc Neurochcm 20:254.
Bhullar RP. Haslam RJ (1987): Detection of23-27 kDa GT1’-binding proteins in platelets and othcr cclls. Biochem J 245:617-620.
Bhullar R, Haslam RJ (1988): Gn-proteins are distinct from rus p.21 and othcr known Low molecular mass GTP-binding proteins in the platelet. FERS Lctt 237:168-172.
Bokoch GM. Katada T, Northrup JK, Hewlett El2, Gilman AG (1983): Identification of the predominant substrate Tor ADP-

GTP-Binding Proteins in Myelin 23
Larocca JN, Ledeen RW, Dvorkin B, Makman MH (1987): Muscar- inic receptor binding and muscarinic receptor-mediated inhibi- tion of adenylate cyclase in rat brain myelin. J Neurosci 7: 3869-3876.
Mayorga LS, Diaz R, Stahl PD (1989): Regulatory role for GTP- binding proteins in endocytosis. Science 244:1475-1477.
Nagata K, Nagao S , Nozawa Y (1989): Low M, GTP-binding proteins in human platelets: Cyclic AMP-dependent protein kinase phospborylates m22KG(I) in membrane but not c21KG in cy- tosol. Biochem Biophys Res Commun 160:235-242.
Norton WT, Poduslo SE (1973): Myelination in rat brain: method of myelin isolation. J Neurochem 21 :749-757.
Pereyra PM, Braun PE (1983): Studies on subcellular fractions which are involved in myelin membrane assembly; isolation from de- veloping mouse brain and characterization by enzyme markers, electron microscopy, and electrophoresis. J Neurochem 41: 957-973.
Riheiro FAP, Rodbell M (1989): Pertussis toxin induces StrUCtUrdl
changes in Ga proteins independently of AFP-ribosylation. Proc Natl Acad Sci USA 86:2577.-2581.
Santos E, Nebreda AR ( 1989): Structural and functional properties of rus proteins. FASEB J 3:2151-2163.
Smith R, Braun PE (1988): Release of proteins from the surface of bovine CNS myelin by salts and phospholipases. J Neurochem 50:722-729.
Spiegel AM (1987): Signal transduction by guanine nucleotide binding proteins. Mol Cell Endocrinol 49: 1-16.
Sprinkle TJ, Hancock J (1989): Identification of pbosphoryl and nu- cleotide binding sites on CNPase. Trans Am SOC Neurochem 20:255.
Towbin H. Staehlin T, Gordon J (1979): Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Pro- cedure and some applications. Proc Natl Acad Sci USA 76: 4350-4354.
Ulmer JB, Braun PE (1983): In Vivo phosphorylation of myelin basic proteins in developing mouse brain: Evidence that phosphory- lation is an early event in myelin formation. Dev Neurosci 6:345-355.
Ulmer JB , Edwards AM, McMorris FA, Braun PE (1987): Cyclic AMP decreases the phosphorylation state of myelin basis pro- teins in rat brain cell cultures. J Biol Chem 262:1748-1755.
Yong VW, Sekiguchi S, Kim MW, Kim SU (1988): Phorbol ester enhances morphological differentiation of oligodendrocytes in culture. J Neurosci Res 19:187-194.
ribosylation by islet-activating protein. J Biol Chem 258:2072- 2075.
Bourne HR (1988): Do GTPases direct membrane traffic in secretion? Cell 53:669-671.
Braun PE, Bernier L (1989): GTP-Binding proteins associated with CNS myelin, in Cellular and Molecular Biology of Myelina- tion, Jeserich G, Althaus H, Waehneldt T, eds, NATO AS1 Series, Springer Verlag, in press.
Burgoyne RD (1987): Control of exocytosis. Nature (Lond) 328: I 12- 113.
Chan KC, Ranwani J , Moscarello MA (1988): Myelin basic protein binds GTP at a single site in the N-terminus. Biochem Biophys Res Commun 152:1468-1473.
Comerford JG, Gibson JR, Dawson AP, Gibson I (1989): RAS p21 and other Gn proteins are detected in mammalian cell lines by [y3'S]GTP yS binding. Biochem Biophys Res Commun 159:
Edwards AM, Arquint M, Braun PE, Roder JC, Dunn RJ , Pawson T, Bell JC (1988): Myelin associated glycoprotein, a cell adhesion molecule of oligodendrocytes, is phosphorylated in brain. Molec and Cell Biol 8:2655-2658.
Edwards, AM, Ross, NW, Ulmer JB , Braun PE (1989): Interaction of myelin basic protein and proteolipid protein. J Neurosci Res
Eichberg J, Berti-Mattera LN, Day S-F, Lowery J , Zhu X (1989): Basal and receptor-stimulated metabolism of polyphosphoi- nositides in peripheral nerve myelin. J Neurochem 52:524,
Freissmuth M, Casey PJ, Gilman AG (1989): G proteins control di- verse pathways of transmembranc signaling. FASEB J 3:2125- 2131.
Gilman AG ( 1987): G proteins: Transducers of receptor-generated signals. Annu Rev Biochem 56:615-650.
Kahn DW, Morel1 P (1988): Pbosphatidic acid and phosphoinositide turnover in myelin and its stimulation by acetylcholine. J Neu- rochem 50:1542-1550.
Kamata T, Kung HF (1988): Effects of ras-encoded proteins and plate- let-derived growth factor on inositol phospholipid turnover in NRK cells. Proc Natl Acad Sci (USA) 855799-5803.
Kikuchi A, Yamamoto K , Fujita T, Takai Y (1988): ADP-ribosylation of the bovine brain rho protein by botulinum toxin type C1. J Biol Chem 263: 16303-16308.
Laemmli UK (1970): Cleavage of structural proteins during the as- sembly of the head of bacteriophage T4. Nature 227:680-685.
1269-1 274.
22197-102.
Suppl.