identification of novelcompositetransposable element ...jb.asm.org/content/173/22/7077.full.pdf ·...
TRANSCRIPT

Vol. 173, No. 22
Identification of a Novel Composite Transposable Element, Tn5280,Carrying Chlorobenzene Dioxygenase Genes of
Pseudomonas sp. Strain P51JAN ROELOF VAN DER MEER,* ALEXANDER J. B. ZEHNDER,
AND WILLEM M. DE VOSDepartment of Microbiology, Wageningen Agricultural University, Hesselink van Suchtelenweg 4,
6703 CT Wageningen, The Netherlands
Received 18 June 1991/Accepted 16 September 1991
Analysis of one of the regions of catabolic plasmid pP51 which encode chlorobenzene metabolism ofPseudomonas sp. strain P51 revealed that the tcbA and tcbB genes for chlorobenzene dioxygenase anddehydrogenase are located on a transposable element, Tn5280. Tn5280 showed the features of a compositebacterial transposon with iso-insertion elements (IS1066 and IS1067) at each end of the transposon oriented inan inverted position. When a 12-kb Hindlll fragment of pP51 containing Tn5280 was cloned in the suicidedonor plasmid pSUP202, marked with a kanamycin resistance gene, and introduced into Pseudomonas putidaKT2442, TnS280 was found to transpose into the genome at random and in single copy. The insertion elementsIS1066 and IS1067 differed in a single base pair located in the inner inverted repeat and were found to be highlyhomologous to a class of repetitive elements of Bradyrhizobium japonicum and distantly related to IS630 ofShigella sonnei. The presence of the catabolic genes tcbA and tcbB on TnS280 suggests a mechanism by whichgene clusters can be mobilized as gene cassettes and joined with others to form novel catabolic pathways.
Chlorinated aromatic compounds have until now beenconsidered one of the most problematic categories of envi-ronmental pollutants which are nondegradable or slowlydegradable by microorganisms (36). Nevertheless, bacteriathat partially transform chlorinated aromatic compounds,such as biphenyls or other polyaromatic compounds, andeven completely mineralize chlorinated benzoates and ben-zenes (36, 38, 44) have been isolated. Metabolism of this lastclass of compounds is particularly interesting, because itrequires a specific combination of existing pathways, whicheach have their limitations with respect to chlorobenzenedegradation (31). The initial attack of the aromatic ring iscarried out in most cases by dioxygenase multienzymecomplexes and (substituted) catechols are formed as centralpathway intermediates (36). Chlorinated catechols whicharise as intermediates in chlorobenzene degradation can thenbe successfully metabolized only via a modified ortho cleav-age or chlorocatechol oxidative pathway (36). The metacleavage route will in this case lead to dead-end products (2,25). However, in microorganisms such as Pseudomonasputida Fl (15, 16, 56) that were shown to be able to oxidizechlorobenzenes, meta cleavage follows the initial conversionby the dioxygenase complex. The modified ortho cleavagepathway is the central degradation sequence used by micro-organisms that utilize 2,4-dichlorophenoxyacetic acid and3-chlorobenzoate (8, 9). However, these organisms fail toperform the initial oxidation of chlorobenzenes (31, 36).Pseudomonas sp. strain P51 was isolated from river sed-
iments and is able to use 1,2-dichloro-, 1,4-dichloro-, and1,2,4-trichlorobenzene, as the sole carbon and energy source(51). Strain P51 was found to harbor catabolic plasmid pP51of 110 kb (52). With the use of expression studies anddeletion analysis, two regions of pP51 that contain thegenetic information for the metabolism of chlorobenzenes
* Corresponding author.
have been identified and characterized (49, 52) (Fig. 1). Oneregion was found to contain a gene cluster, tcbCDEF, whichencoded a modified ortho cleavage pathway (49, 52). Theorganization of the tcbCDEF gene cluster was found to bevery similar to that of the tfdCDEF operon of Alcaligeneseutrophus JMP134 (8, 14, 34) and the clcABD operon of P.putida (11). The other region of pP51 contained the tcbABgene cluster which encoded a chlorobenzene dioxygenasecomplex and a dehydrogenase, catalyzing the conversion ofchlorobenzenes to chlorocatechols (52). This gene clusterexhibited size, organization, and function characteristicssimilar to those of other known bacterial dioxygenases, suchas todCIC2BA from P. putida Fl (16, 56), bphAB (12, 23,30), or xylXYZ (20). We were interested in analyzing themechanisms that could give rise to bacterial strains capableof metabolizing chlorobenzenes, such as Pseudomonas sp.strain P51, in order to allow a better understanding of theevolution of new pathways in bacteria. This article describesthe transposable nature of the tcbAB gene cluster of Pseu-domonas sp. strain P51, and our results support a mecha-nism for evolving novel, functional metabolic routes bycassettelike combination of existing pathways.
MATERIALS AND METHODS
Bacterial strains and plasmids. Escherichia coli DH5a[supE44 AlacUJ69 (d;80 lacZAM15) hsdRJ7 recAl endAlgyrA96 thi-J relAl] and TG1 [supE hsdA5 thi A(lac-proAB)F' (traD36 proAB+ lacIq lacZAM15)] (37) were routinelyused for cloning experiments with plasmids and M13 phages,respectively. E. coli S17-1 (41) was used as the donor strainfor mobilization of pSUP202-derived plasmids. Pseudomo-nas putida KT2442 (hsdRl hsdM+) (10) was the recipientstrain in mating experiments with E. coli S17-1 and was usedto study transposition events. The strain is a rifampin-resistant (Rif) plasmid-free derivative of strain P. putidamt-2. Plasmids pUC18 and pUC19 (55) were used as general
7077
JOURNAL OF BACTERIOLOGY, Nov. 1991, p. 7077-70830021-9193/91/227077-07$02.00/0Copyright © 1991, American Society for Microbiology
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

7078 VAN DER MEER ET AL.
FIG. 1. Physical maps of the two gene clusters and Tn5280 on
plasmid pP5l encoding chlorobenzene metabolism and of relevantclones used in this study. The map of pPS1 is shown for restrictionenzymes XhoI (9) and HindIII (?). The sizes of the relevant gene
clusters and their directions of transcription are shown in thephysical map of pP5l as solid arrows and below the detailed physicalmap. The upper pathway (pathway I) genes tcbA and tcbB mediatethe conversion of 1,2,4-trichlorobenzene to 3,4,6-trichlorocatechol;the lower pathway (pathway II) genes encode metabolism of 3,4,6-trichlorocatechol to 2,5-dichloromaleylacetic acid (52). Detailedanalysis of the genes and gene products of this region showed that itwas composed of at least three structural genes: tcbC, encodingchlorocatechol 1,2-dioxygenase; tcbD, encoding chloromuconatecycloisomerase, and tcbE, encoding dienelactone hydrolase. Thefunction of tcbF has not yet been revealed (49, 52). The tcbR gene
was found to be involved in regulation of the chlorocatecholoxidative pathway (50). The solid triangle indicates the position atwhich the Kmr gene cassette has been inserted in pTCB95R3.
cloning vectors. For DNA sequencing, we applied M13mpl8and M13mpl9 (55). Plasmid pSUP202 (41) is a pBR325-derived vector carrying the mob genes of plasmid RP4. It canbe mobilized to other gram-negative bacteria by the transfersystem of E. coli S17-1, but it does not replicate in P. putida.The plasmid was used to deliver TnS280 transposon con-
structs in P. putida KT2442. pTCB60 contains Tn5280 andthe tcbAB gene cluster on a 12-kb HindIII fragment ofcatabolic plasmid pP51 of Pseudomonas sp. strain P51 inpUC18 (52) (Fig. 1). pTCB78 contains a 2.8-kb PstI fragmentharboring IS1066, and pTCB93 contains a 3.2-kb BamHI-HindIII fragment with IS1067. Plasmid pTCB95 was con-
structed by cloning the 12-kb HindIII pP51 DNA fragment ofpTCB60 in pSUP202. This plasmid was then partially di-gested with BamHI and ligated with a DNA fragment con-
taining the kanamycin resistance (Kmr) gene of Tn903 (17),derived from plasmid pRME1 (19), with BamHI ends. Trans-formants of E. coli were subsequently analyzed for theirplasmid content, and a plasmid carrying the Kmr gene in theBamHI site on the tcbB locus was used (pTCB95R3) (Fig. 1).Plasmid pSUP202: :Km was created by cloning the Kmr gene
into the unique BamHI site present on pSUP202.Media and culture conditions. E. coli strains were culti-
vated at 37°C on LB (37), whereas P. putida strains were
grown at 30°C in the same medium. For mating experiments,
E. coli S17-1 and P. putida KT2442 strains were diluted20-fold from cultures grown overnight into fresh medium, towhich the relevant antibiotics were added, and grown to anA620 of approximately 1.0. Strains were then mixed in a 1:1ratio, filtered over a 0.2-,um-pore-size nitrocellulose filter,washed with 5 ml of sterile LB medium, and incubated onthe surface of an LB agar plate for 12 h at 30°C. After thisincubation time, the bacteria were washed from the filter,plated onto selective medium containing rifampin and kana-mycin, and incubated for at least 18 h at 30°C after whichtransconjugants were screened. Antibiotics were added asfollows: ampicillin, 50 ,ug/ml; kanamycin, 50 ,ug/mr; andrifampin, 50 ,ug/ml. When necessary, the medium was sup-plemented with 0.004% 5-bromo-4-chloro-3-indolyl-,3-D-ga-lactoside and 1.0 mM isopropyl-p-D-thiogalactopyranoside.DNA techniques and sequence analysis. Plasmid DNA
isolations, transformations, and other DNA techniques werecarried out as described elsewhere (49, 52) or by establishedprocedures (37). Total DNA was isolated from 1.5-ml cul-tures of P. putida transconjugants by the procedure ofMarmur (28). DNA sequencing was performed by cloningappropriate DNA fragments into M13 phages and carryingout sequencing reactions by a modified version (49) of thedideoxy-chain termination method of Sanger et al. (39).Computer analysis and processing of sequence informationwere done using the program PC/GENE (Genofit, Geneva,Switzerland) and the GCG package (7). Restriction enzymesand other DNA-modifying enzymes were obtained from LifeTechnologies Inc. (Gaithersburg, Md.) or Pharmacia LKBBiotechnology (Uppsala, Sweden).
Nucleotide accession number. The nucleotide sequencepresented in this article was deposited at GenBank underaccession number M61114.
RESULTS
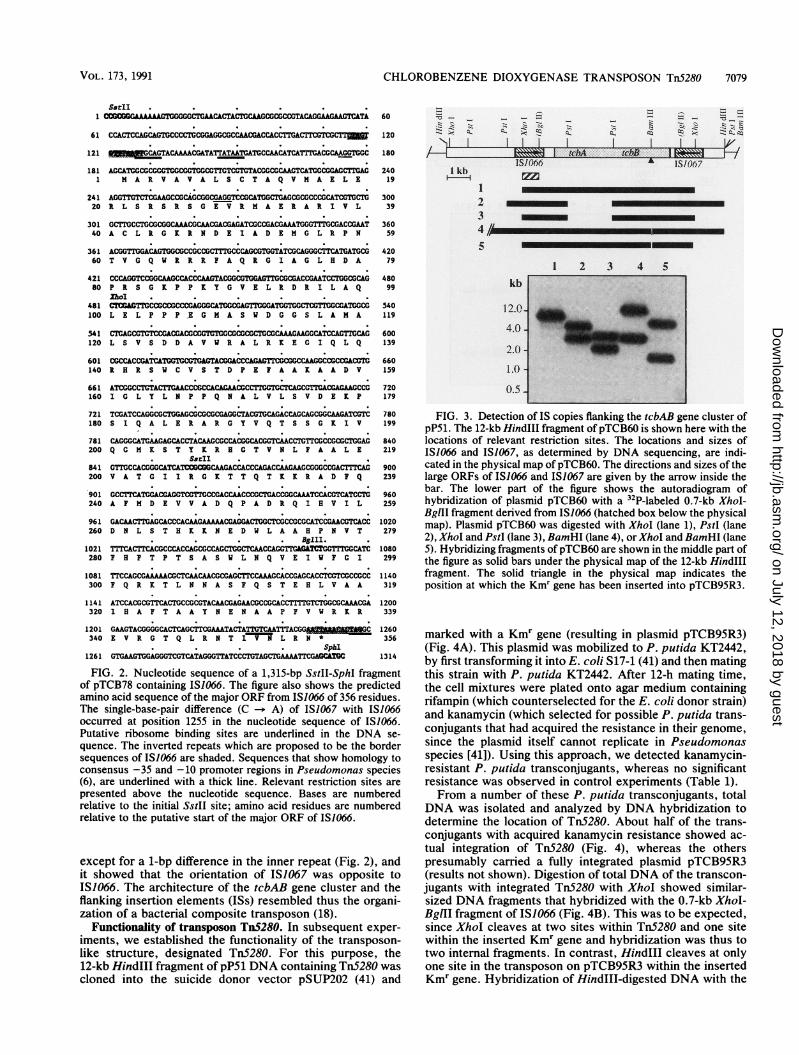
Identification and characterization of IS1066 and IS1067.Analysis of the start of the tebA chlorobenzene dioxygenasegene cluster of plasmid pP51 by DNA sequencing revealednot only the beginning of the gene encoding the large ironsulfur protein of the dioxygenase complex (48) but also alarge upstream open reading frame (ORF) (Fig. 2). This ORFand its flanking regions were compared with sequences in theGenBank data base and found to show considerable homol-ogy with presumed repetitive or insertion elements (seebelow). The 1,068-bp ORF was found to be flanked by shortinverted repeats of 13 bp (Fig. 2), giving the entire elementwithin these borders a size of 1,142 bp, which was tenta-tively designated IS1066. When translated, the ORF couldencode a polypeptide of 355 amino acids, with a calculatedmolecular mass of 39.7 kDa. We were interested to seewhether more copies of IS1066 occurred on plasmid pP51, sowe hybridized restriction fragments of pP51 generated withHindIII and XhoI with a 32P-labelled 0.7-kb XhoI-BglIIfragment of IS1066, derived from plasmid pTCB78. Thisrevealed a single hybridizing band with the 12-kb HindIlIand 7.5-kb XhoI fragments of pP51, containing the tcbABcluster (results not shown). However, fine mapping withinthe 12-kb HindIII fragment using hybridization with thesame DNA probe showed that another copy of IS1066 waspresent on the right side of the presumed tcbB locus (Fig. 3)(52). The DNA region that hybridized with the IS1066 probewas located on the 3.2-kb BamHI-HindIII fragment ofpTCB60. This region was further analyzed by DNA sequenc-ing and found to contain an iso-IS1066 copy, designatedIS1067. The sequence of this copy was identical to IS1066
J. BACTERIOL.
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
CHLOROBENZENE DIOXYGENASE TRANSPOSON Tn5280 7079
Seth .
CCACTCCAG-GTGCCCCuGCGGAGGCGCCA6kCG-CCkC
CAGTACAAAACGATATtATUTGATGCCAACATCAITTGACGCAAGGTGGC
AGCATGGCGCGGGTGGCGGTGGCGTTGTCGTGTACGGCGCAAGTCATGGCGGAGCTTGAGM A R V A V A L S C T A Q V M A E L E
241 AGGTTGTCTCGCCGCAGCGGCGAGGTCCGCATGGCTGGCGCGCCCGCATCGTGCTG20 R L S R S R S G E V R H A E R A R I V L
301 GCTTGCCTGCGCGGCAAACGCAACGACGAGATCGCCGACGAAATGGGTTTGCGACCGAAT40 A C L R G I R N D E I A D E M G L R P N
361 ACGGTTGGACAGTGGCGCCGCCGCTTTGCCCAGCGTGGTATCGCAGGGCTTCATGATGCG60 T V G Q W R R R P A Q R G I A G L B D A
P R S G X P P K Y G V E L R D R I L A QIhoICTGGTrGClCCGCCCGCAG;C G_C GL E L P P P -E G M A S W D G G S L A M A
CTGAGCGTGTCCGACGACGG CGCGCTGCGCAAAGAAGCATCCAGTTGCAGL S V S D D A V U R A L R R E G I Q L Q
CGCCACCG&TCATG GTGTGACTACGGACCCALADTTCGCGGCCAAGGCCGCCGCGTGR H R S U C V S T D P E 7 A A R A A D V
ATCGGCCTGTACTTGNACCCCCACAGAACGCCTGTGCCMCGTIGAG :CCGI G L Y L N P P Q N A L V L S V D E R P
TCGATCCAGGCGCTGGAGCGCGCGCGAGGCTACGTGCAGACCAGCAGCGGCA&GATCGTCS I Q A L E R A R G Y V Q T S S G R I V
CAGGGCATGAAGAGCACCTACAAGCGCCACGGCACGGTCAACCTGTTCGCCGCGCTGGAGQ G M R S T Y K R H G T V N L F A A L E
Stll .IGTTGCCACGGGCATCA7C0GCGCAAGACCACCCAGACCAAGAAGCGGGCCGACTTTCAGV A T G I I R G K T T Q T R R R A D P Q
GCCT'CATGGACGAGGTCEGTGCCGACCAACCCGCTGACCGGCAAATCCACGTCATCCTGA F M D E V V A D Q P A D R Q I B V I L
GACAACTTGAGCACCCACAAGAAAAACGMGACTGGCTCGCCGCGCATCCGAACGTCACCD N L S T H R K N E D W L A A H P N V T
BgII .TTTCACTTCACGCCCACCAGCGCCAGCTGGCTCAACCAGGTTGAOATCTGGTrTGGCATCP H P T P T S A S W L N Q V E I W P G I
TTCCAGCGAAAAACGCTCAACAACGCGAGCTTCCAAAGCACCGAGCACCTCGTCGCCGCCF Q R K T L N N A S F Q S T E H L V A A
60
120
180
24019
30039
36059
42079
48099
540119
600139
660159
720179
780199
840219
900239
960259
1020279
1080299
1140319
:6 -
-Z:z z.z -Z
>< C),
I 'NL..I-
I kb-
I
t4. x LCaX
W-/.0 *ItcA ic I I
1S1066 la 1S1067w-
e2
234/_5
kb
12.0
4.0
2.0 -
1.() -
0.5
1 2 3 4 5
.m
FIG. 3. Detection of IS copies flanking the tcbAB gene cluster ofpP51. The 12-kb HindIlI fragment of pTCB60 is shown here with thelocations of relevant restriction sites. The locations and sizes ofIS1066 and IS1067, as determined by DNA sequencing, are indi-cated in the physical map of pTCB60. The directions and sizes of thelarge ORFs of IS1066 and IS1067 are given by the arrow inside thebar. The lower part of the figure shows the autoradiogram ofhybridization of plasmid pTCB60 with a 32P-labeled 0.7-kb XhoI-BgIII fragment derived from IS1066 (hatched box below the physicalmap). Plasmid pTCB60 was digested with XhoI (lane 1), PstI (lane2), XhoI and PstI (lane 3), BamHI (lane 4), or Xhol and BamHI (lane5). Hybridizing fragments ofpTCB60 are shown in the middle part ofthe figure as solid bars under the physical map of the 12-kb Hindlllfragment. The solid triangle in the physical map indicates theposition at which the Kmr gene has been inserted into pTCB95R3.
1141 ATCCACGCGTTCACTGCCGCGTACAACGAGAACGCCGCACCTMTGTCTGGCGCAAACGA 1200320 I H A P T A A Y N E N A A P P V W R K R 339
1201340
1261
GAAGTACGGGGCACTCAGCTTCGAAATACTATEGTCA&TrTA cE V R G T Q L R N T I V N L R N *
SphlGTGAAGTGGAGGGTCGTCATAGGGTTATCCCTGTAGCTGAAAATTCGAOCA!C
FIG. 2. Nucleotide sequence of a 1,315-bp SstII-SphI fragmentof pTCB78 containing IS1066. The figure also shows the predictedamino acid sequence of the major ORF from IS1066 of 356 residues.The single-base-pair difference (C -* A) of IS1067 with IS1066occurred at position 1255 in the nucleotide sequence of IS1066.Putative ribosome binding sites are underlined in the DNA se-quence. The inverted repeats which are proposed to be the bordersequences of IS1066 are shaded. Sequences that show homology toconsensus -35 and -10 promoter regions in Pseudomonas species(6), are underlined with a thick line. Relevant restriction sites arepresented above the nucleotide sequence. Bases are numberedrelative to the initial SstII site; amino acid residues are numberedrelative to the putative start of the major ORF of IS1066.
except for a 1-bp difference in the inner repeat (Fig. 2), andit showed that the orientation of IS1067 was opposite toIS1066. The architecture of the tcbAB gene cluster and theflanking insertion elements (ISs) resembled thus the organi-zation of a bacterial composite transposon (18).
Functionality of transposon Tn5280. In subsequent exper-iments, we established the functionality of the transposon-like structure, designated TnS280. For this purpose, the12-kb HindIlI fragment of pP51 DNA containing TnS280 wascloned into the suicide donor vector pSUP202 (41) and
marked with a Kmr gene (resulting in plasmid pTCB95R3)(Fig. 4A). This plasmid was mobilized to P. putida KT2442,by first transforming it into E. coli S17-1 (41) and then matingthis strain with P. putida KT2442. After 12-h mating time,the cell mixtures were plated onto agar medium containingrifampin (which counterselected for the E. coli donor strain)and kanamycin (which selected for possible P. putida trans-conjugants that had acquired the resistance in their genome,since the plasmid itself cannot replicate in Pseudomonasspecies [41]). Using this approach, we detected kanamycin-resistant P. putida transconjugants, whereas no significantresistance was observed in control experiments (Table 1).From a number of these P. putida transconjugants, total
DNA was isolated and analyzed by DNA hybridization todetermine the location of TnS280. About half of the trans-conjugants with acquired kanamycin resistance showed ac-tual integration of TnS280 (Fig. 4), whereas the otherspresumably carried a fully integrated plasmid pTCB95R3(results not shown). Digestion of total DNA of the transcon-jugants with integrated Tn5280 with XhoI showed similar-sized DNA fragments that hybridized with the 0.7-kb XhoI-BgllI fragment of IS1066 (Fig. 4B). This was to be expected,since XhoI cleaves at two sites within TnS280 and one sitewithin the inserted Kmr gene and hybridization was thus totwo internal fragments. In contrast, HindIII cleaves at onlyone site in the transposon on pTCB95R3 within the insertedKmr gene. Hybridization of HindIll-digested DNA with the
1
61
121
181I
42180
481100
541120
601140
661160
721180
781200
841200
901240
961260
1021280
1081300
VOL. 173, 1991
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

7080 VAN DER MEER ET AL.
.......~~~~~~~~~~A A ....I_ztw r, I51:
t ffa* ¢ 8
Is -"P i *."s.:. I.-7 _ w-,
Ia
Q~~~~~~~~~~~~~~~~~~~~~~TfI
i
i-
pS.
** -0
FIG. 4. Analysis ofP. putida KT2442 transconjugants containingTnS280. (A) Physical map of plasmid pTCB95R3 which contains theKmr gene inserted into TnS280. (B and C) Autoradiograms of totalDNA and plasmid control DNA, digested with XhoI (B) or HindIII(C), and hybridized with the 32P-labelled 0.7-kb XhoI-BglII fragmentfrom IS1066 (hatched bar below the physical map in Fig. 4A).Plasmid pTCB95R3 DNA (lanes 1) and total DNAs from different P.putida KT2442 transconjugants with TnS280 insertions (lanes 2 to14) and from P. putida KT2442, the parent strain (lanes 15) areshown.
same IS1066 probe yielded two fragments which varied insize in different strains, indicating insertion of TnS280 indifferent sites of the genome (Fig. 4C). This analysis furthershowed that only a single copy of TnS280 had been inte-
TABLE 1. Frequencies of kanamycin and rifampin resistanceobtained by mating E. coli S17-1 harboring different suicide
constructs and P. putida KT2442
Strain Frequency ofresistancea
P. putida KT2442 (control) ..................................... <10-9E. coli S17-1(pTCB95R3) (control) ........................... <10-8E. coli S17-1(pSUP202::Km) ................................... <10-8P. putida KT2442 + E. coli S17-1(pTCB95R3) ........... 10-6-10`7P. putida KT2442 + E. coli S17-1(pSUP202::Km) ...... <10-9
a Frequencies are given per (recipient) cell, and the values were similar inthree independent experiments.
FIG. 5. FASTA alignment (33) of the deduced amino acid se-quences of putative proteins encoded by the major ORF of the ISsand repetitive elements IS1066, Riatl (43), Rhjsa9 and RhjsalO (24),and IS630 (29). Identical residues in two or more proteins areshaded. The N-terminal region of the polypeptides that was pre-dicted to contain a helix-turn-helix motif by using the algorithms ofthe GCG program (7) is boxed in the sequence. RhjsalO presents thepart of the translated first ORF of these repetitive elements that washomologous to the ORF of IS1066, and Rhjscx9 contains part of thesecond ORF. Gaps introduced in the sequence for optimal alignmentare indicated by hyphens.
grated into the genome in each case. Hybridization withDNA probes containing fragments of the tcbAB region orwith DNA fragments containing the Kmr gene gave the sameresults as those obtained with the IS1066 probe (not shown).No hybridization was detected with the vector pSUP202 (notshown). These results led us to the conclusion that TnS280introduced on a suicide delivery system is able to transposein P. putida KT2442 in single copy and apparently insertsrandomly.Homology of IS1066 with other insertion elements. A
TFASTA search (33) of the putative gene product of thelarge ORF of IS1066 with sequences present in the GenBankcatalogue revealed homology with one other IS (IS630 fromShigella sonnei) (29), with a class of repetitive elements fromBradyrhizobium japonicum (24), and with an uncharacter-ized ORF on the integrated Ri plasmid of Agrobacteriumrhizogenes (43) (Fig. 5). The homology of IS1066 was highestwith the Rhjsa elements of B. japonicum (61.5% DNAsequence identity in 1,010 bp). These elements, however, donot encode one large ORF but two smaller overlappingORFs (24). Percentages of identity of IS1066 with theseORFs on the amino acid level were 44.7% with ORF1 (in 141amino acids overlap) and 55.4 to 57.7% with ORF2 (in 182residues overlap). The ORF from A. rhizogenes could en-code a protein of 355 residues, and its identity to IS1066 wascalculated to be 36.4% in 338 amino acids overlap (53.1%DNA sequence identity in 727 bp). All these three types
i-
J. BACTERIOL.
_m
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CHLOROBENZENE DIOXYGENASE TRANSPOSON TnS280 7081
IS1066 TC arCGCLT <- 1116 bp--W AATP _ CGTGAAGr
IS1067 CAAAGGAT CrAIrAr 1116 bp-_. AAATGAACGATARiatl Tra7CTTAA 1113 bp- _ C(RTATGAAC
R8Ja9 cCT13CTCGA 1108 bp- (A A CCCCITQTRBJa7 CTIrTTI'CICG -.*- 1108 bp---O GGCPO IRAC
RSal MAGrAGN3GAT -ACC -A 1108 bp-_M TGCCTGCCOC00IS630 rrADOO '- 1130 bp-_ AGAM
FIG. 6. Comparison of sizes and border sequences of a class ofrepetitive elements and ISs homologous to IS1066 and IS1067. Thesizes of the inverted repeats differ from 9 to 14 bp. The boxedsequence indicates the duplicated CTAG found in some of thesequences. The single-base-pair difference between IS1066 andIS1067 is underlined in the inverted repeat of IS1067. Riatl, A.rhizogenes (43); RSJa9, RSJa7 and RSJa1O, B. japonicum (24);IS630, S. sonnei (29).
were slightly related to IS630 (identity of IS1066 with IS630was 21.8% in 312 amino acids overlap). The putative pro-
teins of all these ORFs were calculated to have an isoelectricpoint of between 10.6 and 11.6, which is a characteristic ofIS-encoded transposases (13). Further, secondary structurepredictions suggested a helix-turn-helix motif for all putativeproteins between residues 30 and 60 (Fig. 5), which could beinvolved in DNA binding and recognition. The sizes of theelements and their border sequences were similar (Fig. 6).The border sequences of the elements consisted of invertedrepeats of between 9 and 14 bp in size. A possible target siteduplication of 4 bp due to transposition with the sequence
CTAG could be observed in some of the inverted repeatswhich, however, could also be part of the inverted repeatitself.
DISCUSSION
This report describes the initial characterization of a novelbacterial transposable element, TnS280, which occurs on
catabolic plasmid pP51 of Pseudomonas sp. strain P51. Ourresults show that Tn5280 is a composite transposon, formedby two isocopies of an IS (IS1066) which are oriented inopposite directions. These elements, IS1066 and its twinwith a single-base-pair difference (IS1067), flank the com-
plete tcbAB gene cluster on pP51 which encodes chloroben-zene dioxygenase and dehydrogenase (52). We showed byanalyzing transposition events in P. putida KT2442 thatTn5280 is a functional transposon, transposing in single copyand inserting randomly into the genome.
The similarity of the ISs of Tn5280 with other repetitiveelements or ISs such as those from B. japonicum (24), A.rhizogenes (43), and S. sonnei (29) suggests that this class ofelements can be found in different bacterial species. Thelarge ORF present in IS1066 and related elements couldencode a putative transposase, with features similar to otherIS-encoded tranposases such as isoelectric point and helix-turn-helix motif (13). The DNA sequence of IS1066 upstreamof the putative start codon at position 184 further showed a
possible ribosome binding site and promoter sequences (Fig.2) (6). The end ofIS1066 also contained a sequence homol-ogous to the -35 promoter box TTGACA, facing outward(Fig. 2), which is a characteristic of other bacterial ISs as
well (13).Transposable elements and ISs are commonly found in
bacteria and are known to contribute to the spread of genes
and to trigger other genetic events by their insertion, such as
activation or inactivation of gene expression (1, 18). Inparticular, composite transposons, such as TnS (3) and TnJO(26), are examples of mobile genetic elements that promotethe spread of DNA (in this case, of antibiotic resistance
genes) that is captured between the ISs. In catabolic path-ways, transposable elements are presumed to play a role ingenerating rearrangements of existing routes (4, 5, 54).Probably the best-characterized transposable elements incatabolic pathways are the TOL transposons, Tn4361 andTn4362 (46, 47), which belong to the Tn3 family and containthe xyl operons which encode xylene and/or toluene metab-olism. Studies of different TOL plasmids have shown thatvarious combinations of the individual xyl operons arefound, as well as duplications, which may be caused by theactivity of these transposable elements or by general recom-bination events (32). In addition, the dehalogenase genes ofP. putida PP3 were discovered to be located on transposableelements (42). Finally, ISs were presumed to play a role inthe development of the catabolic pathway for 2,4,5-trichlo-rophenoxyacetic acid (45), and two functional ISs, IS931 andIS932, that were able to activate gene expression of adjacentgenes upon insertion and to mobilize adjacent DNA se-quences were characterized (21, 22).The characterization of the transposable nature of the
tcbAB gene cluster with its ISs, IS1066 and IS1067, is to ourknowledge the first documented example of the functionalityof a composite transposon carrying catabolic genes whichcode for degradation of aromatic compounds. The discoveryof the mobile nature of the tcbA and tcbB genes may help topropose a mechanism by which the pathway for chloroben-zene degradation has evolved in Pseudomonas sp. strainP51. Most existing routes of aromatic metabolism in bacteriawill not be able to successfully catalyze the complete con-version of chlorobenzenes but will either direct metabolites,such as chlorinated catechols into dead-end pathways (2,25), or fail to perform the initial oxidation of the chlorinatedsubstrate (35). This has led research groups to efforts tocombine two pathways operating in different strains into asingle progenitor, by using mating techniques to transfer onepathway, encoded on a catabolic plasmid, from one strain toa strain with the other pathway. In this manner, new strainsthat completely mineralized 1,4-dichlorobenzene and mono-chlorobenzene have been constructed, starting with strainsthat carry a toluene degradation pathway, such as P. putidaFl (15, 16, 56), or P. putida R5-3 (27) and strains that use3-chlorobenzoate or benzoate as the sole carbon and energysource, such as Pseudomonas sp. strain B13 (9, 31) or P.alcaligenes C-0 (27). The transconjugants in these experi-ments were all found to use the modified ortho cleavageroute to metabolize chlorinated catechols, although they stillcontained detectable catechol 2,3-dioxygenase activity (27,31). Our findings with Pseudomonas sp. strain P51 suggestthat in this strain two separate pathways were combined aswell. The chlorocatechol oxidative pathway is present in thisstrain, encoded by the tcbCDEF gene cluster that is locatedon plasmid pP51 (49, 52). The initial conversion of chlo-robenzenes to chlorocatechols is performed by the enzymesencoded by the tcbAB gene cluster (52) which are very muchakin to other dioxygenases, such as those encoded by thetodCIC2BA (16, 56), xylXYZ (20), or bphAB genes (12, 23,30). The tcbAB gene cluster of pP51 forms part of a com-posite transposon, which suggests that it was recruited on acatabolic plasmid that already contained the genes for thechlorocatechol oxidative pathway. The combination of thetwo gene clusters on a single plasmid may have beennecessary for a proper regulation of the two pathways.However, preliminary DNA binding experiments with theregulator of the chlorocatechol oxidative pathway, TcbR(50), and upstream regions of tcbA did not indicate anyinteraction (48), which suggests that both operons are not
VOL. 173, 1991
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

7082 VAN DER MEER ET AL. J. BACTERIOL.
regulated by a single regulatory gene. Furthermore, culturesof Pseudomonas sp. strain P51 or other chlorobenzene-degrading strains such as Alcaligenes sp. strain A175 (40)growing on chlorinated benzenes, showed in certain casesfor unknown reasons excretion of chlorinated intermediatesresulting in cell death (53), which suggests misrouting andimproper regulation of the two different pathways. Thismisrouting could be caused by the presence of a functionalgene for a catechol 2,3-dioxygenase which could be a rem-nant of the toluene dioxygenase pathway (15, 56), althoughits activity has so far not been detected in Pseudomonas sp.strain P51 (53).The recruitment of the tcbAB gene cluster in TnS280 raises
the interesting question of how the ISs became inserted neara dioxygenase gene cluster, e.g., present on the chromosomeas in the case of the todCJC2BA genes (16, 56), therebycreating a transposable element. The transposon could sub-sequently have been acquired by a passing mobilizablecatabolic plasmid that already contained a chlorocatecholoxidative gene cluster. If so, it seems as though nature'spatchwork assembly has worked in a similar manner to thegenetic engineering strategies we have developed at ourdesks and laboratory benches.
ACKNOWLEDGMENTSWe gratefully acknowledge the stimulating discussions and help-
ful comments of Rik Eggen and Johan Leveau.J.R.V.D.M was supported by a grant from the Netherlands
Integrated Soil Research Programme.
REFERENCES1. Ajioka, J. W., and D. L. Hartd. 1989. Population dynamics of
transposable elements, p. 939-959. In D. E. Berg and M. M.Howe (ed.), Mobile DNA. American Society for Microbiology,Washington, D.C.
2. Bartels, I., H.-J. Knackmuss, and W. Reineke. 1984. Suicideinactivation of catechol 2,3-dioxygenase from Pseudomonasputida mt-2 by 3-halocatechols. Appl. Environ. Microbiol.47:500-505.
3. Berg, D. E., J. Davies, B. Allet, and J.-D. Rochaix. 1975.Transposition of R factor genes to bacteriophage X. Proc. Natl.Acad. Sci. USA 72:3628-3632.
4. Chakrabarty, A. M., D. A. Friello, and L. H. Boff. 1978.Transposition of plasmid DNA segments specifying hydrocar-bon degradation and their expression in various microorgan-isms. Proc. Natl. Acad. Sci. USA 75:3109-3112.
5. Clarke, P. H. 1984. The evolution of degradative pathways, p.11-27. In D. T. Gibson (ed.), Microbial degradation of organiccompounds. Marcel Dekker, Inc., New York.
6. Deretic, V., W. M. Konyecsni, C. D. Mohr, D. W. Martin, andN. S. Hibler. 1989. Common denominators of promoter controlin Pseudomonas and other bacteria. Bio/Technology 7:1249-1254.
7. Devereux, J., P. Haeberli, and 0. Smithies. 1984. A comprehen-sive set of sequence analysis programs for the VAX. NucleicAcids Res. 12:387-395.
8. Don, R. H., A. J. Weightman, H.-J. Knackmuss, and K. N.Timmis. 1985. Transposon mutagenesis and cloning analysis ofthe pathways for degradation of 2,4-dichlorophenoxyacetic acidand 3-chlorobenzoate in Alcaligenes eutrophus JMP134(pJP4).J. Bacteriol. 161:85-90.
9. Dorn, E., M. HeDiwig, W. Reineke, and H.-J. Knackmuss. 1974.Isolation and characterization of a 3-chlorobenzoate degradingpseudomonad. Arch. Microbiol. 99:61-70.
10. Franklin, F. C. H., M. Bagdasarian, M. M. Bagdasarian, andK. N. Timmis. 1981. Molecular and functional analysis of theTOL plasmid pWWO from Pseudomonas putida and cloning ofthe genes for the entire regulated aromatic ring-cleavage path-way. Proc. Natl. Acad. Sci. USA 78:7458-7462.
11. Frantz, B., and A. M. Chakrabarty. 1987. Organization and
nucleotide sequence determination of a gene cluster involved in3-chlorocatechol degradation. Proc. Natl. Acad. Sci. USA 84:4460-4464.
12. Furukawa, K., and T. Miyazaki. 1986. Cloning of a gene clusterencoding biphenyl and chlorobiphenyl degradation in Pseudo-monas pseudoalcaligenes. J. Bacteriol. 166:392-398.
13. Galas, D. J., and M. Chandler. 1989. Bacterial insertion se-quences, p. 109-163. In D. E. Berg and M. M. Howe (ed.),Mobile DNA. American Society for Microbiology, Washington,D.C.
14. Ghosal, D., and L.-S. You. 1988. Nucleotide homology andorganization of chlorocatechol oxidation genes of plasmids pJP4and pAC27. Mol. Gen. Genet. 211:113-120.
15. Gibson, D. T., and V. Subramanian. 1984. Microbial degrada-tion of aromatic hydrocarbons, p. 181-252. In D. T. Gibson(ed.), Microbial degradation of organic compounds. MarcelDekker, Inc., New York.
16. Gibson, D. T., G. J. Zylstra, and S. Chauhan. 1990. Biotrans-formation catalyzed by toluene dioxygenase from Pseudomonasputida Fl, p. 121-133. In S. Silver, A. M. Chakrabarty, B.Iglewski, and S. Kaplan (ed.), Pseudomonas: biotransforma-tions, pathogenesis, and evolving biotechnology. American So-ciety for Microbiology, Washington, D.C.
17. Grindley, N. D., and C. M. Joyce. 1980. Genetic and DNAsequence analysis of the kanamycin resistance transposonTn903. Proc. Natl. Acad. Sci. USA 77:7176-7180.
18. Grindley, N. D. F., and R. R. Reed. 1985. Transpositionalrecombination in prokaryotes. Annu. Rev. Biochem. 54:863-896.
19. Harayama, S., R. A. Leppik, M. Rekik, N. Mermod, P. R.Lehrbach, W. Reineke, and K. N. Timnus. 1986. Gene order ofthe TOL catabolic plasmid upper pathway operon and oxidationof both toluene and benzyl alcohol by the xylA product. J.Bacteriol. 167:455-461.
20. Harayama, S., M. Rekik, and K. N. Timnis. 1986. Geneticanalysis of a relaxed substrate specificity aromatic ring dioxy-genase, toluate 1,2-dioxygenase, encoded by TOL plasmidpWWO of Pseudomonas putida. Mol. Gen. Genet. 202:226-234.
21. Haughland, R. A., U. M. X. Sangodkar, and A. M. Chakrabarty.1990. Repeated sequences including RS1100 from Pseudomonascepacia AC1100 function as IS elements. Mol. Gen. Genet.220:222-228.
22. Haughland, R. A., U. M. X. Sangodkar, P. R. Sferra, and A. M.Chakrabarty. 1991. Cloning and characterization of a chromo-somal DNA region required for growth on 2,4,5-T by Pseudo-monas cepacia AC1100. Gene 100:65-73.
23. Hayase, N., K. Taira, and K. Furukawa. 1990. Pseudomonasputida KF715 bphABCD operon encoding biphenyl and poly-chlorinated biphenyl degradation: cloning, analysis, and expres-sion in soil bacteria. J. Bacteriol. 172:1160-1164.
24. Kaluza, K., M. Hahn, and H. Hennecke. 1985. Repeated se-quences similar to insertion elements clustered around the nifregion of the Rhizobium japonicum genome. J. Bacteriol. 162:535-542.
25. Klecka, G. M., and D. T. Gibson. 1981. Inhibition of catechol2,3-dioxygenase from Pseudomonas putida by 3-chlorocate-chol. Appl. Environ. Microbiol. 41:1159-1165.
26. Kleckner, N., R. K. Chan, B.-K. Tye, and D. Botstein. 1975.Mutagenesis by insertion of a drug-resistance element carryingan inverted repetition. J. Mol. Biol. 97:561-575.
27. Kr6ckel, L., and D. D. Focht. 1987. Construction of chloroben-zene-utilizing recombinants by progenitive manifestation of arare event. Appl. Environ. Microbiol. 53:2470-2475.
28. Mannur, J. 1961. A procedure for the isolation of deoxyribo-nucleic acid from micro-organisms. J. Mol. Biol. 3:208-218.
29. Matsutani, S., H. Ohtsubo, Y. Maeda, and E. Ohtsubo. 1987.Isolation and characterization of IS elements repeated in thebacterial chromosome. J. Mol. Biol. 196:445-455.
30. Mondello, F. J. 1989. Cloning and expression in Escherichia coliof Pseudomonas strain LB400 genes encoding polychlorinatedbiphenyl degradation. J. Bacteriol. 177:1725-1732.
31. Oltmanns, R. H., H. G. Rast, and W. Reineke. 1988. Degrada-tion of 1,4-dichlorobenzene by enriched and constructed bacte-
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 173, 1991 CHLOROBENZENE DIOXYGENASE TRANSPOSON Tn5280 7083
ria. Appl. Microbiol. Biotechnol. 28:609-616.32. Osborne, D. J., R. W. Pickup, and P. A. Williams. 1988. The
presence of two homologous meta pathways operons on TOLplasmid pWW53. J. Gen. Microbiol. 134:2965-2975.
33. Pearson, D. J., and W. R. Lipman. 1988. Improved tools forbiological sequence analysis. Proc. Natl. Acad. Sci. USA 85:2444-2448.
34. Perkins, E. J., M. P. Gordon, 0. Caceres, and P. F. Lurquin.1990. Organization and sequence analysis of the 2,4-dichlo-rophenol hydroxylase and dichlorocatechol oxidative operonsof plasmid pJP4. J. Bacteriol. 172:2351-2359.
35. Reineke, W., and H.-J. Knackmuss. 1980. Hybrid pathway forchlorobenzoate metabolism in Pseudomonas sp. B13 deriva-tives. J. Bacteriol. 142:467-473.
36. Reineke, W., and H.-J. Knackmuss. 1988. Microbial degradationof haloaromatics. Annu. Rev. Microbiol. 42:263-287.
37. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual, 2nd ed. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
38. Sander, P., R.-M. Wittich, P. Fortnagel, H. Wilkes, and W.Francke. 1991. Degradation of 1,2,4-trichloro- and 1,2,4,5-tetra-chlorobenzene by Pseudomonas strains. Appl. Environ. Micro-biol. 57:1430-1440.
39. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
40. Schraa, G., M. L. Boone, M. S. M. Jetten, A. R. W. vanNeerven, P. J. Colberg, and A. J. B. Zehnder. 1986. Degradationof 1,4-dichlorobenzene by Alcaligenes sp. strain A175. Appl.Environ. Microbiol. 52:1374-1381.
41. Simon, R., U. Priefer, and A. Ptihler. 1983. A broad host rangemobilization system for in vivo genetic engineering: transposonmutagenesis in gram negative bacteria. Bio/Technology 1:784-791.
42. Slater, J. H., A. J. Weightman, and B. E. Hall. 1985. Dehaloge-nase genes of Pseudomonas putida PP3 on chromosomallylocated transposable elements. Mol. Biol. Evol. 2:557-567.
43. Slightom, J. L., M. Durand-Tardif, L. Jouanin, and D. Tepfer.1986. Nucleotide sequence analysis of TL-DNA of Agrobacte-rium rhizogenes agropine type plasmid. J. Biol. Chem. 261:108-121.
44. Spain, J. C. 1990. Metabolic pathways for biodegradation ofchlorobenzenes, p. 197-207. In S. Silver, A. M. Chakrabarty, B.Iglewski, and S. Kaplan (ed.), Pseudomonas: biotransforma-tions, pathogenesis, and evolving biotechnology. American So-
ciety for Microbiology, Washington, D.C.45. Tomasek, P. H., B. Frantz, U. M. X. Sangodkar, R. A. Haugh-
land, and A. M. Chakrabarty. 1989. Characterization and nu-cleotide sequence determination of a repeat element isolatedfrom 2,4,5-T degrading strain of Pseudomonas cepacia. Gene76:227-238.
46. Tsuda, M., and T. Iino. 1987. Genetic analysis of a transposoncarrying toluene degrading genes on a TOL plasmid pWWO.Mol. Gen. Genet. 210:270-276.
47. Tsuda, M., and T. Iino. 1988. Identification and characterizationof Tn4653, a transposon covering the toluene transposonTn4651 on TOL plasmid pWWO. Mol. Gen. Genet. 213:72-77.
48. van der Meer, J. R. Unpublished data.49. van der Meer, J. R., R. I. L. Eggen, A. J. B. Zehnder, and W. M.
de Vos. 1991. Sequence analysis of the Pseudomonas P51 tcbgene cluster encoding metabolism of chlorinated catechols:evidence for a specialization of catechol 1,2-dioxygenases forchlorinated substrates. J. Bacteriol. 173:2425-2434.
50. van der Meer, J. R., A. C. J. Frijters, J. H. J. Leveau, R. I. L.Eggen, A. J. B. Zehnder, and W. M. de Vos. 1991. Characteri-zation of the Pseudomonas sp. strain P51 gene tcbR, a LysR-type transcriptional activator of the tcbCDEF chlorocatecholoxidative operon, and analysis of the regulatory region. J.Bacteriol. 173:3700-3708.
51. van der Meer, J. R., W. Roelofsen, G. Schraa, and A. J. B.Zehnder. 1987. Degradation of low concentrations of dichlo-robenzenes and 1,2,4-trichlorobenzene by Pseudomonas sp.strain P51 in nonsterile soil columns. FEMS Microbiol. Ecol.45:333-341.
52. van der Meer, J. R., A. R. W. van Neerven, E. J. de Vries, W. M.de Vos, and A. J. B. Zehnder. 1991. Cloning and characteriza-tion of plasmid-encoded genes for the degradation of 1,2-dichloro-, 1,4-dichloro, and 1,2,4-trichlorobenzene of Pseudo-monas sp. strain P51. J. Bacteriol. 173:6-15.
53. van Neerven, A. R. W. Personal communication.54. Williams, P. A. 1982. Genetic interactions between mixed
microbial populations. Philos. Trans. R. Soc. London Biol. Sci.297:631-639.
55. Yanisch-Perron, C., J. Vieira, and J. Messing. 1985. ImprovedM13 phage cloning vectors and host strains: nucleotide se-quence of the M13mpl8 and pUC19 vectors. Gene 33:103-119.
56. Zylstra, G. J., W. R. McCombie, D. T. Gibson, and B. A.Finette. 1988. Toluene degradation by Pseudomonas putida Fl:genetic organization of the tod operon. Appl. Environ. Micro-biol. 54:1498-1503.
on July 12, 2018 by guesthttp://jb.asm
.org/D
ownloaded from