igf-ii promotes neural stemness igf-ii and igf-ii analogs with
TRANSCRIPT

1
IGF-II Promotes Neural Stemness
IGF-II and IGF-II Analogs with Enhanced Insulin Receptor-A Binding Affinity Promote Neural Stem
Cell Expansion*
Amber N Ziegler1, Shravanthi Chidambaram1, Briony E. Forbes2, Teresa L. Wood1^ and Steven
W. Levison1^
1. Department of Neurology and Neuroscience, Rutgers Biomedical and Health Sciences, Newark, New
Jersey 07103-1709
2. Discipline of Medical Biochemistry/Centre for Neuroscience, Flinders University, Adelaide, Australia,
5001
*Running title: IGF-II Promotes Neural Stemness
To whom correspondence should be addressed: Steven W Levison, Department of Neurology and
Neuroscience, Rutgers Biomedical and Health Sciences, 205 South Orange Ave, Cancer Center H1226,
Newark, NJ 07103. Tel.: (973) 972-5162; Fax: (973) 972-2668; E-mail: [email protected]
^Denotes co-senior authors.
Keywords: Stem cells, Central Nervous System, Cell proliferation, Self-renewal, Insulin receptor, Neurodevelopment, Insulin, Receptor Tyrosine Kinase
Background: IGF-II promotes neural stem cell (NSCs) proliferation and self-renewal. Results: IGF-II analogs are useful for elucidating the receptors responsible for NSC expansion. Conclusion: F19A expands NSCs via IR-A. Significance: IGF-II promotes stemness of NSCs via the IR-A and not through activation of either the IGF-1R or the IGF-2R.
ABSTRACT The objective of this study was to employ genetically engineered IGF-II analogs to establish which receptor(s) mediate the stemness promoting actions of IGF-II on mouse
subventricular zone neural precursors. Neural precursors from the subventricular zone were propagated in vitro in culture media supplemented with IGF-II analogs. Cell growth and identity were analyzed using sphere-generation and further analyzed by flow cytometry. F19A, an analog of IGF-II that does not bind the IGF-2R, stimulated an increase in the proportion of neural stem cells (NSCs) while decreasing the proportion of the later stage progenitors at a lower concentration than IGF-II. V43M, which binds to the IGF-2R with high affinity but which has low binding affinity to the IGF-1R and to the A isoform of the insulin receptor (IR-A) failed to promote NSC
http://www.jbc.org/cgi/doi/10.1074/jbc.M113.537597The latest version is at JBC Papers in Press. Published on January 7, 2014 as Manuscript M113.537597
Copyright 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
growth. The positive effects of F19A on NSC growth was unaltered by the addition of a functional blocking antibody to the IGF-1R. Altogether, these data lead to the conclusion that IGF-II promotes stemness of NSCs via the IR-A and not through activation of either the IGF-1R or the IGF-2R.
The IGF peptides and their receptors comprise an evolutionarily conserved family of signaling molecules that are essential for normal stem cell proliferation and brain development; however, the individual roles of the ligands are only beginning to be realized. Indeed, the IGF system has been largely overlooked as a critical signaling pathway that is required to propagate neural stem cells (NSCs)1 in culture. Recent studies have determined IGF-II is involved in NSC proliferation and maintenance in the subventricular zone (SVZ) (1,2).
The most commonly used method to study neural stem and progenitor (NSP) self-renewal, growth and differentiation is the neurosphere assay (NSA) (3-6). For this assay, the tissue to be evaluated is microdissected and dissociated into a single cell suspension. When grown in a biochemically defined medium with specific growth factors under non-adherent conditions, self-renewing cells proliferate and form a free-floating colony of cells called a neurosphere. A key aspect of the NSA is that each sphere arises from the proliferation of a single cell. A limitation of this method is that NSCs, progenitors and lineage-restricted progenitors are all capable of forming self-renewing spheres. Another limitation is that as the spheres grow they become cellularly heterogeneous. Each neurosphere contains a few stem cells with the bulk of the sphere comprised of progenitors and lineage-restricted cells. These limitations have made it difficult to evaluate which
1 Abbreviations: Neural stem cells (NSCs), Insulin receptor (IR), Subventricular Zone (SVZ), Neural stem and progenitors (NSPs), Neurosphere assay (NSA), mannose-6-phosphate (M6P), Insulin like growth factor (IGF), IGF type 1 receptor (IGF-1R), multipotential progenitor (MP), Platelet derived growth factor (PDGF), Fibroblast Growth factor (FGF), PDGF FGF responsive MP (PFMP), cerebral spinal fluid (CSF), Subgranular zone (SGZ)
population of cells is altered with different treatment conditions in the NSA.
A different approach to the NSA is to exploit the unique molecular identities of the NSPs, enabling cell populations to be quantified by flow cytometry. Buono et al. (2012) recently combined 4 cell surface markers for flow cytometry to reveal the existence of a NSC and 7 types of intermediate progenitors (7). Not surprisingly, no single marker was sufficient to label the NSCs (7). Nestin, sox2, aldehyde dehydrogenase and Hoechst exclusion label multipotential progenitors as well as NSCs, and did not select for neurosphere forming cells. Many sets of markers have been used to classify NSCs (for a list see Pastrana et al. (2011) (8)). The difficulty of classifying these cells by their molecular identity can be attributed to the complex lineage of the cells and the fact that the cells within the neurosphere are heterogeneous. The assay developed by Buono et al., 2012 defined a combination of CD133, CD140a, NG2 and LeX to distinguish multipotential progenitors and NSCs (7).
Our prior studies revealed a role for IGF-II in promoting neural stemness in the NSA (1). However, IGF-II has high affinity to three different receptors: the IGF-2R/mannose-6-phosphate (M6P) receptor, the IGF type 1 receptor (IGF-1R) and an isoform of the insulin receptor known as the IR-A (9-11). Our prior findings in neurospheres supported the hypothesis that IGF-II promotes stemness through the insulin receptor, however the precise makeup of the neurospheres was not examined in that study. Moreover, another study reported IGF-II promotes expansion of neural progenitor cells via activating the IGF-1R (2). Here, we evaluate the effectiveness of three genetically engineered analogs of IGF-II (Table 1), which have different binding affinities for the receptors, to stimulate NSC expansion in culture using both the widely used NSA and the recently established flow cytometry protocol to test the hypothesis that IGF-II promotes neural stemness specifically by activating IR-A (12,13).
Experimental Procedures Analog production and receptor and IGFBP binding assays- For production of IGF-II analogs, purification and binding assays using IGF-1R, IR-
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
A and domains 10-13 of the IGF2R see Delaine et al. (2007) (12). IGFBP-2 binding affinities were derived from BIAcore analyses performed as described by Carrick et al. (2001) (14).
Fibroblast stimulation and Western blot analysis- Fibroblasts that lack IGF-1R and express IR-A, obtained from Andrea Morrione (Thomas Jefferson University), were starved in phenol-red-free DMEM with 0.1% BSA for 24 hours (15). Cells were then stimulated with IGF-II or F19A for 20 min at 37 C. Cells were lysed and protein extracted in RIPA buffer prior to Western blot analysis. Antibodies used: phospho IR/IGF-1R (3024S), IR (3020S), IGR-1R (3027S), phospho AKT Ser473 (4060S), AKT (4056S), phospho-S6 (4848S) S6 (2217S) (all from Cell Signaling used at 1:1000) and beta-actin (A5441 Sigma, 1:5000). Secondary DAM-HRP and DAR-HRP (Jackson ImmunoResearch 1:5000) were used with Western Lightning Plus ECL (Perkin Elmer) to visualize bands on a UVP Bioimaging System.
Neurosphere propagation and quantification- The periventricular region of C57Bl/6 pups (P4-5) was enzymatically dissociated for 15 min at 37°C using 0.25% trypsin/EDTA (Invitrogen) and 100 units Papain (Sigma) in MEM-HEPES with 250 µg/mL of DNase I (Sigma) and 3 mM MgSO4. The tissue was triturated in 0.01% trypsin inhibitor (Sigma) in Pro-N media (16). The cells were plated into wells at 1x 105 cells/ml in media supplemented with 20 ng/ml recombinant human epidermal growth factor (EGF) (PreproTech). Cultures were fed every 2 days by replacing half of the media with an equal volume of fresh media. Neurospheres were collected after 8-9 days in vitro by centrifugation at 200 x g for 5 min. They were dissociated for 5 min at 37°C in Acutase (Millipore) and plated into 12-well plates at 5 x 104 cells/ml in Pro-N media with either no (zero), 4.4 nM or 4.4 µM insulin (Sigma) supplemented with 20 ng/ml EGF and either mouse recombinant IGF-II (R&D Systems Inc) or an IGF-II analog (supplied by B.E. Forbes) at either 30 nM, 15 nM or 8 nM (8x KD, 4x KD or 2x KD of IGF-II for IGF-1R). In some experiments the functional blocking antibody, IMC-A12, a monoclonal antibody against IGF-1R was used (provided by
ImClone Systems, a wholly owned subsidiary of Eli Lilly and Co).
Neurospheres were quantified as described previously (16). Neurosphere volume was determined from phase contrast images using a Zeiss Axiovision Observer.Z1 (Carl Zeiss) microscope under 10x or 40x magnification. Axiovision software was used to measure neurosphere diameters of a minimum of 50 randomly selected neurospheres per condition.
Flow cytometry- Secondary spheres were dissociated by incubating them in 0.2 Wünsch unit (WU)/ml of Liberase DH (Roche) and 250 μg of DNase1 (Sigma) in PGM solution (PBS with 1 mM MgCl2 and 0.6% dextrose) at 37°C for 5 min with gentle shaking. An equal volume of PGM was added and the spheres were placed onto a shaker (LabLine) at 225 rpms at 37°C for 15 min. After enzymatic digestion, Liberase DH was quenched with 10 ml of PGB (PBS without Mg2+ and Ca2+ with 0.6% dextrose and 2 mg/ml fraction V of BSA (Fisher Scientific, BP1600-100) and cells were collected by centrifugation for 5 min at 200 x g. Cells were dissociated by repeated trituration, collected by centrifugation, counted using ViCell (Beckman Coulter, Miami, FL) and diluted to at least 106 cells per 50 μl of PGB. All staining was performed in 96 V-bottom plates using 150 μl/well. For surface marker analysis, cells were incubated in PGB for 25 min with antibodies against Lewis-X (1:20, LeX/CD15 FITC, MMA; BD Bioscience), CD133-APC (1:50,13A4; eBioscience), CD140a (1:400, APA5; BioLegend) and NG2 Chondroitin Sulfate Proteoglycan (1:50, AB5320; Millipore). Cells were washed with PGB by centrifugation at 278 x g. Goat anti-rabbit IgGAlexa Fluor 700 (1:100; Invitrogen) was used for NG2. Cells were then incubated in LIVE/DEAD fixable Violet (Invitrogen) for 20 min for dead cell exclusion. Cells were washed with PGB by centrifugation at 278 x g. They were fixed with 1% ultrapure formaldehyde (50000; Polysciences, Inc) for 20 min, collected by centrifugation for 9 min at 609 x g, resuspended in PBS w/o Mg2+ and Ca2+ and stored at 4°C for next day analysis. All sample data were collected on the BD LSR II (BD Biosciences Immunocytometry Systems). Matching isotype controls were used for all
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
antibodies and gates were set based on these isotype controls. Post-acquisition analysis was performed using FlowJoX (Tree Star Inc, Ashland, OR).
Quantitative real-time PCR- Q-PCR was performed as described in Ziegler et al. (2012), with beta-actin and Quantitech real-time primers for BMI-1, Hes5, Id2 and Axin 2 (QT00165298, QT00268044, QT01038870, 01547672)(1).
Statistics- Data are expressed as means ± SEM and analyzed using one-way ANOVA followed by Tukey’s post-hoc test (Graphpad Prism 4 software).
RESULTS
IGF-2R stimulation fails to promote neurosphere growth
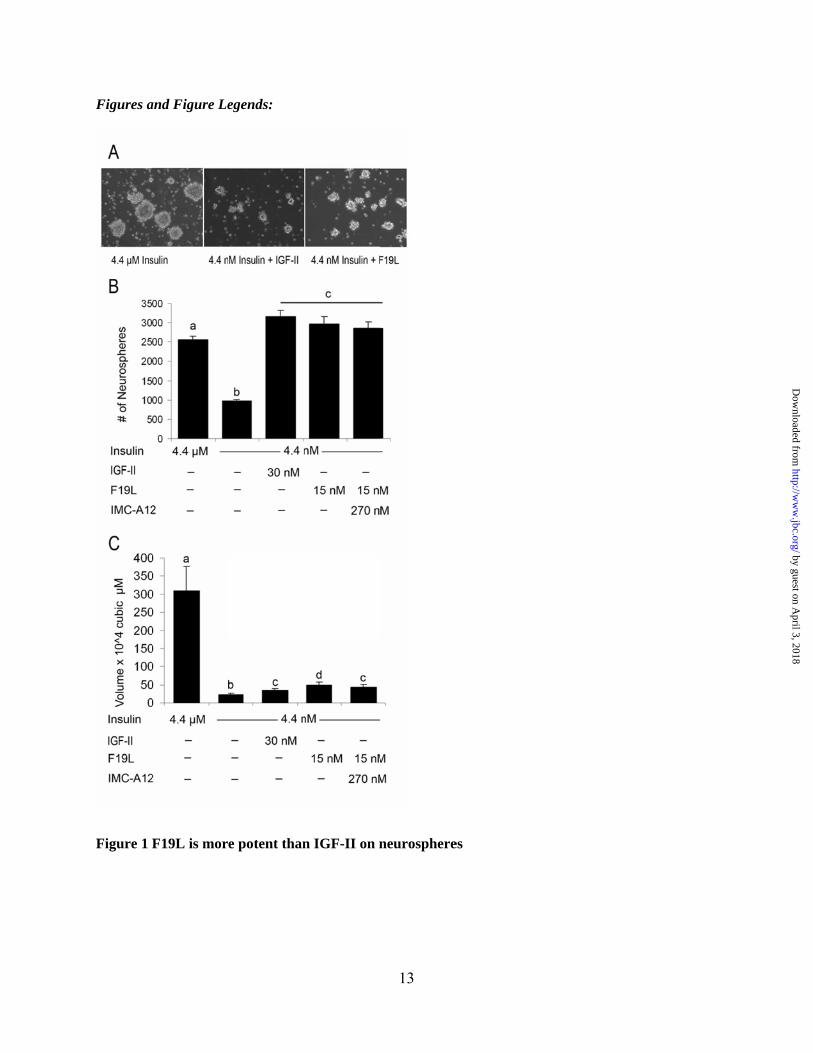
The IGF-2R functions as a scavenger receptor for IGF-II, reducing ligand bioavailability (17). A role for this receptor in IGF-II signaling has been controversial. A recent study suggested that IGF-2R promotes memory consolidation in vivo (18); which is connected to the maintenance of hippocampal neurogenesis. In our prior study defining IGF-II function in NSC self-renewal, we investigated effects of IGF-II through IR or IGF-1R but did not analyze potential IGF-II actions through the IGF-2R (1). Thus, we first tested the F19L analog, which has a phenylalanine substitution at amino acid 19 for leucine and has greater than 6 fold higher binding affinity for IGF-2R than wild type IGF-II while binding the IGF-1R and IR-A with affinities similar to IGF-II (Table 1). Reproducing our earlier studies, primary neurospheres were cultured under standard growth conditions and then dissociated into single cells and grown in control media (4.4 µM insulin), 4.4 nM insulin, or 4.4 nM insulin with IGF-II, F19L or F19L + A12 antibody (an IGF-1R blocking antibody). Similar to our prior studies, when the level of insulin was reduced to 4.4 nM (to abrogate activation of the IGF-1R), the neurosphere size and number were greatly reduced. Previously we determined that 15 nM of IGF-II produced a modest effect and 30 nM IGF-II was able to restore neurosphere volume (1). Addition of F19L or IGF-II restored neurosphere number, an indication of self-renewal (Figure 1). F19L produced the same effect as IGF-II at half the
concentration. The addition of the IGF-1R blocking antibody, A12, did not alter the effectiveness of F19L, indicating its actions are independent of IGF-1R. This is consistent with our previous studies showing the ability of IGF-II to promote NSP self-renewal independent of IGF-1R.
To further address the possibility that IGF-2R binding contributed to IGF-II-induced NSC growth, we used V43M, valine substituted to methionine at amino acid residue 43, an analog of IGF-II that has very low affinity to IGF-1R and to IR-A and high affinity binding to the IGF-2R (0.72 fold compared to wild type IGF-II; Table 1). Few spheres formed when the medium contained V43M (Table 2). In fact, cells cultured in V43M were similar to cells grown without insulin indicating that IGF-2R stimulation fails to promote the growth of neural precursors. An IGF-II analog with increased neurosphere-forming ability
Next, spheres were grown in medium supplemented with a variety of other recombinant IGF-II peptides. The relative affinities of these ligands to the IGF-1R, IGF-2R and IGFBP2 are provided in Table 1. F19A, like F19L, binds with high affinity to IGF-1R and has slightly reduced affinity to the IR. However, F19A, where amino acid 19 has been substituted with alanine, has virtually no binding to the IGF-2R and reduced binding to IGFBP-2. Therefore, we hypothesized that F19A would be effective at a lower concentration than F19L in in situ neurosphere formation as it would be more available to bind the target receptors. We tested this using a concentration of the IGF-II, F19L and F19A that was equal to 2x the KD of IGF-II to IGF-1R. Only F19A was effective at stimulating neurosphere formation at this concentration, and it produced the greatest number of primary neurospheres (Table 2). F19A produced more spheres than control, or insulin containing media, at a lower concentration than either wild-type IGF-II or F19L. As seen in Table 2, F19A enriched the proportion of neurosphere forming cells to 46% of the population compared to 20% when using IGF-II. Moreover, F19A increased the total population of neural precursors by 2 fold versus IGF-II (at the same concentration).
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
Transcription factor, Id2, associated with stemness, is induced by IGF-II
Since we observed an increase in the number of neurospheres grown in 4.4 nM insulin with IGF-II or F19A, we next examined the expression of a group of transcription factor genes responsible for maintaining stemness. Cells were grown in 4.4 μM insulin or 4.4 nM insulin with IGF-II or F19A. Gene expression analysis was performed by QPCR. The expression was normalized to cells grown in 4.4 μM insulin. In cells grown in 4.4 nM insulin with IGF-II or F19A, the expression of BMI-1, Hes 5 and Axin2 were not significantly different from cells grown in 4.4 μM insulin. However the expression of Id2 was 2.7 fold greater in cells cultured in media supplemented with IGF-II (P<0.05) (Table 3). Flow cytometry reveals IGF-II and F19A promote expansion of NSCs
The nervous system, like the hematopoietic system, contains a variety of precursors at different stages of developmental restriction. While the neurosphere assay is a widely used technique for propagating and evaluating NSC, it fails to distinguish the stem cells from neural progenitor cells. Buono et al. (2012) recently combined 4 cell surface markers for flow cytometry to reveal the existence of a NSC and 7 types of intermediate progenitors. Moreover, each of these 8 types of precursors was competent to form a neurosphere (7). Therefore, an increase in neurosphere number does not distinguish NSCs from intermediate progenitors. We used the same set of cell markers, CD133, LeX, CD140a and NG2 used by Buono et al., 2012 (Table 4) to establish which precursors were present within the neurospheres grown under standard NSA conditions versus those grown in a biochemically defined medium containing IGF-II or the F19A analog (7). Neurospheres were cultured in control medium or in medium with reduced insulin (4.4 nM insulin) or in medium with reduced insulin and supplemented with IGF-II (8x KD) or F19A (2x KD). The concentration of each ligand was selected to be the lowest concentration at which neurosphere number was restored to the numbers observed in control media.
Cells were grown for 6 days in vitro and then dissociated for flow cytometric analyses. Gates were set for forward and side scatter and only live cells, based on live/dead dye staining, were analyzed (Figure 2). NSCs were defined as NG2/CD140a double negative and CD133/LeX double positive. These flow cytometric analyses revealed that F19A and IGF-II treatment almost tripled the proportion of NSCs while nearly reducing by 2-fold the multipotential progenitors (MP) termed the “PFMPs” (PDGF FGF responsive MPs) (Figure 2). Other progenitor populations such as MP1, MP2 and MP4 were not affected. Thus, F19A and IGF-II promote the expansion of early NSCs and decrease the number of later multipotential progenitors, such as PFMPs. Modified ligand, F19A, is biologically active
Having established that F19A and IGF-II promote the expansion of early NSCs via IR-A, we wanted to determine whether the modified ligand activates the IR signaling pathway. Since NSCs express IR-A, IR-B and IGF-1R (1), we chose to use a cell line that expresses IR-A in high copy number and does not express IR-B or IGF-1R, enabling us to selectively examine the activation of IR-A via F19A. We performed Western blots to examine IR phosphorylation and activation of downstream signaling, P-AKT (ser473) and P-S6 (Figure 3). Fibroblasts engineered to express only the IR-A form of the IR were treated with IGF-II or F19A for 20 min. This time-point was chosen based on time course activation studies of IGF-II on this cell type (15). IGF-II and F19A produced similar activation of signaling pathways downstream of the IR-A indicating that F19A is stimulating IR-A.
DISCUSSION Stem cells throughout the body are found
within specialized niches. The NSC niche is complex and is elegantly organized to favor specific cell-cell interactions, as well as access to the cerebral microvasculature, extracellular matrix components, meninges and cerebral spinal fluid (CSF). Whereas the lineage of NSCs within the SVZ, as well as the generation and migration of NSP progeny has been an area of intense focus, the niche microenvironment has only recently been studied in detail. The notion that the CSF
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
and IGFs, in particular, play an integral role in the NSC niche has only garnered attention recently (1,2,19). The CSF stimulates the growth of neurospheres in an IGF-II-dependent manner. In fact, IGF-II in the CSF binds to primary cilia protruding from the wall of the lateral ventricles in vivo (2). Our previous study showed that IGF-II could increase the number of cultured postnatal SVZ neurospheres and promoted self-renewal based on differentiation, Q-PCR gene profiling, limiting dilution analysis and transplantation studies (1). Additionally, we demonstrated that IR-A was more highly expressed than IGF-1R on the medial SVZ as opposed to lateral SVZ by laser capture microdissection and Q-PCR. IR-A was also the most highly expressed isoform on neurospheres and, as cells became more lineage restricted, IR-A decreased. Thus, this work supported the idea that the in vitro effects of IGF-II were via IR-A and independent of IGF-1R. Taken together, these studies strongly support a role for IGF-II in the neural stem cell niche. Here we have shown that in vitro simulation of SVZ derived neurospheres with IGF-II or a modified form of IGF-II, F19A increases the proportion of NSCs as quantified using a new flow cytometry analysis, and that these actions are independent of the IGF-2R and IGF-1R. In extending our analyses of stemness genes activated by IGF-II, here we show that IGF-II promotes the expression of the Id2 gene which has been shown to facilitate self-renewal and proliferation of NSCs (20).
Studies of the precursors in the other neurogenic region of the brain suggest that IGF-II promotes the maintenance of hippocampal NSCs in the subgranular zone (SGZ) (19). FACs sorted, microdissected hippocampal cells expressing GFP driven off a sox2 promoter were used in a microarray analysis to identify genes that were induced in these putative stem cells; a gene identified from this study was IGF-II. The importance of IGF-II was further evaluated in the hippocampus using lentiviral shRNA knockdown of IGF-II expression, which resulted in decreased hippocampal precursor proliferation (19). Other papers support a role for IGF-II and neurogenesis in the SVZ (1,2) Together, these studies lend support for a role for IGF-II in the promotion of neurogenesis in both SVZ and SGZ NSCs.
IGF-II consists of 4 domains, B, C, A and D (the B and A domains are similar to the
corresponding domains in insulin) (13). These domains contain residues that are important in binding IGF-1R and IR. The IR (and the structurally similar IGF-1R) is a disulfide linked homodimer composed of two β and two α subunits. While a recent structure of a receptor fragment in complex with insulin has been published, the structure of the entire complex has not been solved (21). Various studies and other structural information have led to a model for how ligands bind to these receptors (22,23). Ligand binds to the IR in a 1:1 stoichiometry despite the existence of two binding pockets within each receptor. Each binding pocket comprises a high and a low affinity binding site (site 1 and site 2 respectively) including residues from both monomers. Binding of ligand results in cross linking of the two receptor halves leading to a structural change and subsequent activation of the receptor’s tyrosine kinase domain. The residues on IGF-II important for binding the IR site 1 include V43, F26 and Y27, while residues E12, F19, L53 and E57 contact site 2 (13). In this study, we used modified forms of IGF-II that contained mutations at these sites of interaction. F19A has high affinity for the IR and IGF-1R. V43M on the other hand has significantly reduced affinities for both the IGF-1R and IR. The signaling efficacy of these analogs has yet to be evaluated; therefore, we have interpreted their effects based on the affinities of these analogs to their receptors.
IGF-2R or cation independent mannose-6-phosphate receptor consists of a large extracellular domain and a very small cytoplasmic tail that displays no intrinsic kinase activity (17,24). IGF-II binds to IGF-2R, via an interaction at F19 on IGF-II (25). There are many recent reports implying that IGF-II activates downstream signaling pathways such as MAPK via IGF-2R, however it is more likely that IGF-2R interacts with other ligands and subsequent receptors rather than directly activating these signaling moieties (26). Recently, hippocampal memory retention and fear extinction were reported to be dependent on IGF-II and, in part, on IGF-2R. Agis-Balboa et. al. (2011) found that IGF-II blocking also inhibited contextual fear extinction, which is hippocampal dependent (27). They reported that blocking IGR-1R using the IR antagonist JB1 negated the effects of IGF-II on fear extinction. However, it is important to note that JB1 cross reacts with IR
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
tyrosine kinase; therefore, an effect via IR-A cannot be ruled out in this study (28). Agis-Balboa et al. (2011) did not see a block in extinction when using an antibody against IGF-2R, which is in contradiction to Chen et. al., 2011 (18,27). Chen et. al., 2011 found that IGF-II administration into the hippocampus enhanced memory retention and prevented extinction in inhibitory avoidance training (18). Additionally they found that blocking IGF-2R abolished the memory enhancement effect of IGF-II. Memory formation in the hippocampus is dependent on neurogenesis (Reviewed in Zhao et. al. 2008 (29)). In our hands, IGF-2R stimulation alone, achieved using V43M, failed to support neurosphere growth. While IGF-2R may play a role in modulating memory enhancement, the exact nature of this role remains elusive as evidence that it acts directly as a signaling receptor is lacking.
IGF-II acts via the IR-A in several different cell types. IGF-II via IR-A has been found to regulate glycogen synthesis in fetal hepatocytes (30). In fibroblasts that express only IR-A, IGF-II and insulin display differential signaling (15). IGF-II is a more potent activator of p70S6 kinase than insulin. Additionally IGF-II results in prolonged ERK activation whereas insulin has prolonged AKT activation (15). Furthermore, differential gene expression and a unique set of phosphorylated proteins was found depending on whether IR-A was activated via IGF-II or insulin (31,32). Integrin alpha5, ICAM and Acidic nuclear phosphoprotein 32, a protein involved in embryogenesis and is switched off in differentiated cells, are more regulated by IGF-II than insulin. Endocytic sorting of IR-A differs depending on whether it is activated by insulin or IGF-II. Stimulation with insulin results in the degradation of IR-A and IRS-1 whereas IGF-II protects IR-A and IRS-1 from degradation, which may be responsible for the prolonged activation observed in p70S6 kinase and ERK (33). This evidence supports the role for different ligands, IGF-II or insulin in this case, producing various effects via the same receptor, IR-A. Here we have shown that IGF-II via IR-A promotes NSCs in culture.
Given our findings that F19A, an IGF-II analog, is more potent than IGF-II to support and expand NSCs in vitro, it would be interesting to
determine its effects after injury. There are numerous consequences to the central nervous system when the homeostasis of CSF is disrupted, such as with aging or after injury. The CSF regulates and supports the development, division and migration of cells. In fact, the presence of modulated and flowing CSF is required for stem cell maintenance (34). If CSF flow is disrupted by injury or disease, neurons cannot migrate properly (35) and there is a reduction in the clearance of toxins from the brain and reduced nutrient content (34,36). Certain growth factors, such as IGF-II, are tightly controlled and modulated after brain injuries, like cortical trauma, and are essential for wound healing which helps to bring the environment back to homeostasis (37). Infusions of various growth factors including FGF, EGF, NGF, VEGF, GDNF and BDNF, after different injury models have improved outcome after injury due to either neurogenesis or neuroprotection (as reviewed in Johanson et al 2010 (38)). The CSF has been hypothesized to modulate the environment after injury. We have shown that IGF-II is an integral player in NSC self-renewal; however altered growth factors with restricted interactions, like F19A, are more potent, and therefore, may be a viable option for therapeutics.
In conclusion, this study demonstrates that a genetically engineered form of IGF-II, F19A, stimulates the expansion of NSCs in culture. F19A appeared to be more potent, as it does not bind IGF-2R and only poorly binds the IGFBPs, thus effectively increasing the local concentration compared to IGF-II or F19L. The action of IGF-II and analogs is independent from IGF-2R and IGF-1R, indicating that its effects are via IR-A. The neurosphere assay demonstrated that the analogs produce an increased number of spheres and that a greater percentage of cells in the population are able to give rise to spheres. Flow cytometry analyses revealed an expansion of the NSCs specifically when cultured in IGF-II or F19A, making F19A a useful analog for culturing NSCs. ACKNOWLEDGEMENTS This work was supported by R21 NS076874 awarded to SWL and TLW, and F31NS065607 awarded to ANZ. We thank ImClone Systems, a wholly owned subsidiary of Eli Lilly and Co., for supplying IMC-A12
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
References 1. Ziegler, A. N., Schneider, J. S., Qin, M., Tyler, W. A., Pintar, J. E., Fraidenraich, D.,
Wood, T. L., and Levison, S. W. (2012) IGF-II promotes stemness of neural restricted precursors. Stem Cells 30, 1265-1276
2. Lehtinen, M. K., Zappaterra, M. W., Chen, X., Yang, Y. J., Hill, A. D., Lun, M., Maynard, T., Gonzalez, D., Kim, S., Ye, P., D'Ercole, A. J., Wong, E. T., LaMantia, A. S., and Walsh, C. A. (2011) The cerebrospinal fluid provides a proliferative niche for neural progenitor cells. neuron 69, 893-905
3. Reynolds, B. A., Tetzlaff, W., and Weiss, S. (1992) A multipotent EGF-responsive striatal embryonic progenitor cell produces neurons and astrocytes. J.Neurosci. 12, 4565-4574
4. Morshead, C. M., Reynolds, B. A., Craig, C. G., McBurney, M. W., Staines, W. A., Morassutti, D., Weiss, S., and Van der Kooy, D. (1994) Neural stem cells in the adult mammalian forebrain: A relatively quiescent subpopulation of subependymal cells. Neuron 13, 1071-1082
5. Reynolds, B. A., and Weiss, S. (1996) Clonal and population analyses demonstrate that an EGF-responsive mammalian embryonic CNS precursor is a stem cell. Dev.Biol. 175, 1-13
6. Reynolds, B. A., and Rietze, R. L. (2005) Neural stem cells and neurospheres--re-evaluating the relationship. Nat Methods 2, 333-336
7. Buono, K. D., Vadlamuri, D., Gan, Q., and Levison, S. W. (2012) Leukemia inhibitory factor is essential for subventricular zone neural stem cell and progenitor homeostasis as revealed by a novel flow cytometric analysis. Dev Neurosci 34, 449-462
8. Pastrana, E., Silva-Vargas, V., and Doetsch, F. (2011) Eyes wide open: a critical review of sphere-formation as an assay for stem cells. Cell Stem Cell 8, 486-498
9. Yamaguchi, Y., Flier, J. S., Benecke, H., Ransil, B. J., and Moller, D. E. (1993) Ligand-binding properties of the two isoforms of the human insulin receptor. Endocrinology 132, 1132-1138
10. Denley, A., Bonython, E. R., Booker, G. W., Cosgrove, L. J., Forbes, B. E., Ward, C. W., and Wallace, J. C. (2004) Structural determinants for high-affinity binding of insulin-like growth factor II to insulin receptor (IR)-A, the exon 11 minus isoform of the IR. Mol Endocrinol 18, 2502-2512
11. Jones, J. I., and Clemmons, D. R. (1995) Insulin-like growth factors and their binding proteins: biological actions. Endocr Rev 16, 3-34
12. Delaine, C., Alvino, C. L., McNeil, K. A., Mulhern, T. D., Gauguin, L., De Meyts, P., Jones, E. Y., Brown, J., Wallace, J. C., and Forbes, B. E. (2007) A novel binding site for the human insulin-like growth factor-II (IGF-II)/mannose 6-phosphate receptor on IGF-II. J Biol Chem 282, 18886-18894
13. Alvino, C. L., McNeil, K. A., Ong, S. C., Delaine, C., Booker, G. W., Wallace, J. C., Whittaker, J., and Forbes, B. E. (2009) A novel approach to identify two distinct receptor binding surfaces of insulin-like growth factor II. J Biol Chem 284, 7656-7664
14. Carrick, F. E., Forbes, B. E., and Wallace, J. C. (2001) BIAcore analysis of bovine insulin-like growth factor (IGF)-binding protein-2 identifies major IGF binding site
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
determinants in both the amino- and carboxyl-terminal domains. J Biol Chem 276, 27120-27128
15. Sacco, A., Morcavallo, A., Pandini, G., Vigneri, R., and Belfiore, A. (2009) Differential signaling activation by insulin and insulin-like growth factors I and II upon binding to insulin receptor isoform A. Endocrinology 150, 3594-3602
16. Alagappan, D., Lazzarino, D. A., Felling, R. J., Balan, M., Kotenko, S. V., and Levison, S. W. (2009) Brain injury expands the numbers of neural stem cells and progenitors in the SVZ by enhancing their responsiveness to EGF. ASN NEURO 1, e00009
17. Ghosh, P., Dahms, N. M., and Kornfeld, S. (2003) Mannose 6-phosphate receptors: new twists in the tale. Nature reviews. Molecular cell biology 4, 202-212
18. Chen, D. Y., Stern, S. A., Garcia-Osta, A., Saunier-Rebori, B., Pollonini, G., Bambah-Mukku, D., Blitzer, R. D., and Alberini, C. M. (2011) A critical role for IGF-II in memory consolidation and enhancement. Nature 469, 491-497
19. Bracko, O., Singer, T., Aigner, S., Knobloch, M., Winner, B., Ray, J., Clemenson, G. D., Jr., Suh, H., Couillard-Despres, S., Aigner, L., Gage, F. H., and Jessberger, S. (2012) Gene expression profiling of neural stem cells and their neuronal progeny reveals IGF2 as a regulator of adult hippocampal neurogenesis. J Neurosci 32, 3376-3387
20. Jung, S., Park, R. H., Kim, S., Jeon, Y. J., Ham, D. S., Jung, M. Y., Kim, S. S., Lee, Y. D., Park, C. H., and Suh-Kim, H. (2010) Id proteins facilitate self-renewal and proliferation of neural stem cells. Stem cells and development 19, 831-841
21. Menting, J. G., Whittaker, J., Margetts, M. B., Whittaker, L. J., Kong, G. K., Smith, B. J., Watson, C. J., Zakova, L., Kletvikova, E., Jiracek, J., Chan, S. J., Steiner, D. F., Dodson, G. G., Brzozowski, A. M., Weiss, M. A., Ward, C. W., and Lawrence, M. C. (2013) How insulin engages its primary binding site on the insulin receptor. Nature 493, 241-245
22. Kiselyov, V. V., Versteyhe, S., Gauguin, L., and De Meyts, P. (2009) Harmonic oscillator model of the insulin and IGF1 receptors' allosteric binding and activation. Molecular systems biology 5, 243
23. De Meyts, P., Wallach, B., Christoffersen, C. T., Urso, B., Gronskov, K., Latus, L. J., Yakushiji, F., Ilondo, M. M., and Shymko, R. M. (1994) The insulin-like growth factor-I receptor. Structure, ligand-binding mechanism and signal transduction. Horm Res 42, 152-169
24. Brown, J., Jones, E. Y., and Forbes, B. E. (2009) Interactions of IGF-II with the IGF2R/cation-independent mannose-6-phosphate receptor mechanism and biological outcomes. Vitamins and Hormones 80, 699-719
25. Brown, J., Delaine, C., Zaccheo, O. J., Siebold, C., Gilbert, R. J., van Boxel, G., Denley, A., Wallace, J. C., Hassan, A. B., Forbes, B. E., and Jones, E. Y. (2008) Structure and functional analysis of the IGF-II/IGF2R interaction. EMBO J 27, 265-276
26. Brown, J., Jones, E. Y., and Forbes, B. E. (2009) Keeping IGF-II under control: lessons from the IGF-II-IGF2R crystal structure. Trends in biochemical sciences 34, 612-619
27. Agis-Balboa, R. C., Arcos-Diaz, D., Wittnam, J., Govindarajan, N., Blom, K., Burkhardt, S., Haladyniak, U., Agbemenyah, H. Y., Zovoilis, A., Salinas-Riester, G., Opitz, L., Sananbenesi, F., and Fischer, A. (2011) A hippocampal insulin-growth factor 2 pathway regulates the extinction of fear memories. EMBO J 30, 4071-4083
28. Blum, G., Gazit, A., and Levitzki, A. (2000) Substrate competitive inhibitors of IGF-1 receptor kinase. Biochemistry 39, 15705-15712
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
29. Zhao, C., Deng, W., and Gage, F. H. (2008) Mechanisms and functional implications of adult neurogenesis. Cell 132, 645-660
30. Liang, L., Guo, W. H., Esquiliano, D. R., Asai, M., Rodriguez, S., Giraud, J., Kushner, J. A., White, M. F., and Lopez, M. F. (2010) Insulin-like growth factor 2 and the insulin receptor, but not insulin, regulate fetal hepatic glycogen synthesis. Endocrinology 151, 741-747
31. Pandini, G., Medico, E., Conte, E., Sciacca, L., Vigneri, R., and Belfiore, A. (2003) Differential gene expression induced by insulin and insulin-like growth factor-II through the insulin receptor isoform A. J Biol Chem 278, 42178-42189
32. Morcavallo, A., Gaspari, M., Pandini, G., Palummo, A., Cuda, G., Larsen, M. R., Vigneri, R., and Belfiore, A. (2011) Research resource: New and diverse substrates for the insulin receptor isoform A revealed by quantitative proteomics after stimulation with IGF-II or insulin. Mol Endocrinol 25, 1456-1468
33. Morcavallo, A., Genua, M., Palummo, A., Kletvikova, E., Jiracek, J., Brzozowski, A. M., Iozzo, R. V., Belfiore, A., and Morrione, A. (2012) Insulin and insulin-like growth factor II differentially regulate endocytic sorting and stability of insulin receptor isoform A. J Biol Chem 287, 11422-11436
34. Owen-Lynch, P. J., Draper, C. E., Mashayekhi, F., Bannister, C. M., and Miyan, J. A. (2003) Defective cell cycle control underlies abnormal cortical development in the hydrocephalic Texas rat. Brain 126, 623-631
35. Sawamoto, K., Wichterle, H., Gonzalez-Perez, O., Cholfin, J. A., Yamada, M., Spassky, N., Murcia, N. S., Garcia-Verdugo, J. M., Marin, O., Rubenstein, J. L., Tessier-Lavigne, M., Okano, H., and Alvarez-Buylla, A. (2006) New neurons follow the flow of cerebrospinal fluid in the adult brain. Science 311, 629-632
36. Silverberg, G. D., Heit, G., Huhn, S., Jaffe, R. A., Chang, S. D., Bronte-Stewart, H., Rubenstein, E., Possin, K., and Saul, T. A. (2001) The cerebrospinal fluid production rate is reduced in dementia of the Alzheimer's type. Neurology 57, 1763-1766
37. Walter, H. J., Berry, M., Hill, D. J., Cwyfan-Hughes, S., Holly, J. M., and Logan, A. (1999) Distinct sites of insulin-like growth factor (IGF)-II expression and localization in lesioned rat brain: possible roles of IGF binding proteins (IGFBPs) in the mediation of IGF-II activity. Endocrinology 140, 520-532
38. Johanson, C., Stopa, E., Baird, A., and Sharma, H. (2011) Traumatic brain injury and recovery mechanisms: peptide modulation of periventricular neurogenic regions by the choroid plexus-CSF nexus. Journal of neural transmission 118, 115-133
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
Tables:
Table 1 Binding properties of IGF-II and Analogs
IGF-1R IC50 nM (Rel affinity)
IGF-2R domains 10-13 KD nM (Rel affinity)
IR-A IC50 nM (Rel affinity)
BP2 affinity nM (Rel affinity)
IGF-II 3.4 ±0.2 (1)a,b 1.87 (1)a,b 4 ±0.4 (1)a,b 0.14 (1)
F19A 7.7±1.3 (0.4)a ND 12.9±0.07 (0.3)b 0.76 (0.19)
F19L 4.6±1.5 (0.7)a 0.29 (6.35)a 2.6±0.38 (0.8) 0.51 (0.28)
V43M >100 (<0.01)a,b
2.59 (0.72)a,b ND 0.12 (0.85)
ND = not detected Rel affinity = Relative affinity vs. IGF-II Values less than 1 indicate reduced interaction. a Modified from Delaine et al., 2007(12) b Modified from Alvino et al., 2009 (13) Table 2 Evaluation of neurosphere generation at reduced ligand concentration.
4.4 µM Insulin
2x KD IGF-II
2x KD F19L
2x KD F19A
8x KD V43M
No Insulin
Neurospheres per well Primaries 1660 ± 68 1150 ± 32 1440 ± 152 2110 ± 85 940 ± 32 810 ±37 Secondaries 1055 ± 89 650 ± 32 570 ± 21 1395 ± 53 570 480
% sphere generating cells 34% 20% 16% 46% 17% 15%
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
Table 3 Evaluation of stem cell genes using QPCR
Data represents mean ± SEM from experiments performed 4 separate times. * indicates P<0.05 by ANOVA with Tukeys’ post-hoc test.
Table 4 Flow Cytometry markers used for analysis of neurospheres
Cell surface
markers
Multipotential Precursors
NSC MP1 MP2 MP3 MP4 PFMP
CD133 + - + - + -
LeX + + + - + +
NG2 - - + + + +
CD140a - - - - + +
Growth condition
BMI-1 Hes 5 ID2 Axin-2
4.4μM Insulin 1 1 1 1
4.4nM + IGF-II 1.40 ± 0.36 2.54 ± 0.69 2.75 ± 0.80* 1.52 ± 0.27
4.4nM + F19A 1.11 ± 0.38 1.56 ± 0.15 1.80 ± 0.28 1.69 ± 0.49
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Figures a
Figure 1
and Figure Le
F19L is mor
egends:
e potent thann IGF-II on n
13
neurospherees
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
Figure 1
Primary neurospheres were dissociated and plated into media containing 4.4 µM insulin, or 4.4
nM insulin with either IGF-II at 30 nM, F19L at 15 nM or F19L at 15nM + IMC-A12. A.
Representative 20x images of neurospheres in various conditions. B,C. Neurosphere number and
volume were evaluated at 6 days in vitro. Error bars indicated SEM, a,b,c,d indicate ANOVA
with Tukeys’ post-hoc p<0.05.
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 2
and F19A
Flow cytome
A analog.
etric analysis of stem an
15
d progenitor
r population
s after treatmment with IGGF-II
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

16
Figure 2
Primary neurospheres were passaged and platted into Ctl (4.4 µM insulin) or 4.4 nM insulin with
IGF-II (8x KD) or F19A (2x KD). In 4.4 nM insulin alone spheres are dramatically reduced in size
and number (previously published) and therefore cannot be used for flow cytometry. After 6 days
in vitro neurospheres were collected and processed for labeling and flow cytometry. A. Live cells
were gated using a Live/Dead dye. B. Single cells were selected for based on SSC-A vs FSC-W.
C, C1. Isotype controls for Alexa 700 NG2 vs PE CD140 (C) and APC CD133 vs FITC LeX
(C1). D. Dot plot overlay of 3 treatments for NG2 vs CD140. E. Dot plot overlay of NG2 CD140
double negative cells (Q4 from D) for CD133 vs LeX, NSCs are in Q6. F. Dot plot overlay of
NG2 CD140 double positive cells (Q2 from D) for CD133 vs LeX, PFMPs are in Q5. D1-3, E1-3,
F1-3 Contours for 4.4 µM insulin (orange), 4.4 nM insulin + IGF-II (light blue) and 4.4 nM
insulin + F19A (red). See Table 3 for surface marker list. Values are ± SEM, * indicates p< 0.05
by ANOVA with Tukeys’ post-hoc test.
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 3
Figure 3
(2x KD) F
473 and p
IGF-II and F
Fibroblasts l
F19A or IGF-
phospho s6 we
F19A activat
acking IGF-1
-II. Cell lysa
ere induced u
tion of IR-A
1R and over
ate was collec
upon IGF-II an
17
expressing IR
cted and sign
nd F19-A stim
R-A, were sti
naling assesse
mulation of ce
imulated for 2
ed via Wester
ells.
20 min with
rn blot. AKT
8 nM
T pSer
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from

W. LevisonAmber N. Ziegler, Shravanthi Chidambaram, Briony E. Forbes, Teresa L Wood and Steven
Promote Neural Stem Cell ExpansionIGF-II and IGF-II Analogs with Enhanced Insulin Receptor-A Binding Affinity
published online January 7, 2014J. Biol. Chem.
10.1074/jbc.M113.537597Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from