immobilized pineapple stem bromelain activity in a wine-like medium: effect of inhibitors
TRANSCRIPT

ARTICLE IN PRESSFBP-460; No. of Pages 6
Iw
Ma
0b
1
Tafbt(
aoltEsp
0h
food and bioproducts processing x x x ( 2 0 1 3 ) xxx–xxx
Contents lists available at ScienceDirect
Food and Bioproducts Processing
j ourna l ho me page: www.elsev ier .com/ locate / fbp
mmobilized pineapple stem bromelain activity in aine-like medium: Effect of inhibitors
arco Estia, Ilaria Benuccia,∗, Katia Liburdia, Anna Maria Vittoria Garzillob
Department for Innovation in Biological, Agro-Food and Forest Systems, University of Tuscia, via S. Camillo de Lellis,1100 Viterbo, ItalyDepartment of Ecological and Biological Sciences, University of Tuscia, via S. Camillo de Lellis, 01100 Viterbo, Italy
a b s t r a c t
The catalytic activity of stem bromelain, covalently immobilized on chitosan beads, was characterized towards a
synthetic substrate (Bz-Phe-Val-Arg-pNA), in a wine-like acidic medium containing wine inhibitors (ethanol, sulphur
dioxide, grape skin and seed tannins), at their average concentration range.
For all tested substances, the inhibition constant (Ki) values of immobilized bromelain were significantly higher
than the corresponding values obtained in a previous work for free enzyme, thus indicating that direct covalent
immobilization on chitosan beads makes protease more resistant to the inhibition effect.
Immobilized protease was affected by ethanol inhibition (competitive type) only if present at abnormal concentra-
tion for real wine. Grape skin and seed tannins exerted uncompetitive inhibition to the same extent, with a Ki value
close to 1 g l−1gallic acid eq. The effect of free sulphur dioxide on immobilized bromelain changed respect to free enzyme,
both becoming uncompetitive inhibitor and dramatically lowering its strength.
These results show that immobilized stem bromelain could have productive biotechnological applications in wine-
making, even though further studies will be necessary to test its proteolytic activity towards wine proteins in real
matrices.
© 2013 The Institution of Chemical Engineers. Published by Elsevier B.V. All rights reserved.
Keywords: Pineapple stem bromelain; Covalent immobilization; Wine inhibitors; Wine-like medium
Moreover, immobilization by covalent linkage could induce
. Introduction
he addition of adsorbent materials, such as bentonite, is non-specific common practice, applied in wine industryor the prevention of protein haze. Despite its effectiveness,entonite fining adversely affect the quality of treated wine,hrough the removal of colour, flavour and texture compoundsWaters et al., 2005).
The possibility of stabilizing white wine by acid proteases,ble to hydrolyze specifically proteins and inert respect tother wine compounds, appears of great interest. Neverthe-
ess, currently, enzyme application, in free form, is limited byhe increased request of additive-free food, prevailing in someuropean countries (Feijoo-Siota and Villa, 2011). For this rea-on, growing attention has been given to the application of
Please cite this article in press as: Esti, M., et al., Immobilized pineapple
Food Bioprod Process (2013), http://dx.doi.org/10.1016/j.fbp.2013.12.001
roteases immobilized on solid support, which could be apply
∗ Corresponding author. Tel.: +39 0761357426; fax: +39 0761357498.E-mail address: [email protected] (I. Benucci).Received 14 February 2013; Received in revised form 4 November 2013
960-3085/$ – see front matter © 2013 The Institution of Chemical Engittp://dx.doi.org/10.1016/j.fbp.2013.12.001
for wine protein stabilization, by a continuous treatment,in different bioreactor configurations with no enzyme resid-ual contamination. Stem bromelain, a cysteine proteinaseextracted from the stem of pineapple plant, has been immo-bilized on various supports (organic and inorganic) and bydifferent methods (Seo et al., 1998; Tan et al., 2008; Shi etal., 2007; Krajewska, 2004; Mahmood and Saleemuddin, 2007;Gupta et al., 2007).
Enzyme immobilization, achieved by fixing a biocatalyst toor within solid supports, is of great interest for applications inthe food industry. Compared with their free disposable forms,immobilized enzymes avoids product contamination in thefood reaction mixture, allowing their reusability (Yodoya et al.,2003).
stem bromelain activity in a wine-like medium: Effect of inhibitors.
; Accepted 2 December 2013
enzyme structure modification to overcome the inhibitory
neers. Published by Elsevier B.V. All rights reserved.

ARTICLE IN PRESSFBP-460; No. of Pages 6
2 food and bioproducts processing x x x ( 2 0 1 3 ) xxx–xxx
effect of different chemical compounds as well as to limit pro-tease self-digestion phenomena (Kim, 2005). Recently, stembromelain has been covalently and directly linked to chitosanbeads without glutaraldehyde, and this food-safe biocatalystappeared to be suitable for application in a wine-like medium(Benucci et al., 2012). Furthermore, the influence of expectedinhibitors on free bromelain activity that is usually present inwine has been investigated, showing that all tested substances(ethanol, sulphur dioxide, grape skin, grape seed, ellagic andgallic tannins) were reversible inhibitors for the protease infree form (Esti et al., 2011). The strongest mixed-type inhibitorwas found to be free sulphur dioxide, which is added to wine,especially white wine, during the winemaking process to pre-vent undesirable microbial growth and oxidation processes(Segundo and Rangel, 2001; Toniolo et al., 2010).
The present work, together with the above citedmanuscripts, completes our preliminary evaluation phase,carried out using a wine-like acidic medium, of protease feasi-bility as a sustainable practice for white wine selective proteinstabilization, and an alternative to bentonite fining. The aimof the study was to characterize the effect of inhibitors,which are usually present in wine (over their average range ofconcentration), on the activity of stem bromelain, covalentlyimmobilized on chitosan beads.
Considering that protein insolubilization and wine turbid-ity phenomena remain unclear and the triggering event is notcaused solely by the protein molecules themselves (Hsu andHeatherbell, 1987; Vincenzi et al., 2011; Batista et al., 2009),bromelain activity has been evaluated towards a syntheticsubstrate.
2. Materials and methods
2.1. The enzyme and chemicals
Stem bromelain (EC 3.4.22.32) from Sigma–Aldrich (Milan,Italy) was immobilized on chitosan beads (Chitopearl BCW-3010, Wako Chemicals GmbH, Neuss, Germany). The syntheticpeptide substrate Bz-Phe-Val-Arg-p-nitroaniline (pNA) waspurchased from Bachem, Germany. Grape skin and seed tan-nins, as preparations intended for enological use, were kindlysupplied by EVERINTEC (Venice, Italy). All other reagents wereobtained from Sigma–Aldrich (Milan, Italy).
2.2. Immobilization procedure
Stem bromelain was covalently immobilized on ChitopearlBCW-3010 by a direct mechanism, which probably involves theprotease carboxyl groups of Asp or Glu residues and the aminogroups of the support.
One ml of enzyme preparation (5 mg ml−1), previously sol-ubilized in tartaric buffer (tartaric acid/sodium tartrate 0.03 M,pH 3.2) was added to 100 mg of chitosan beads, and then incu-bated at room temperature for 12 h (Benucci et al., 2012).
2.3. Determination of immobilization yield
At the end of the immobilization time, the supports werewashed three times with 2 M NaCl solution in order to removeall non-covalently bound proteins. The biocatalysts wererecovered by centrifugation. After collection of the super-
Please cite this article in press as: Esti, M., et al., Immobilized pineapple
Food Bioprod Process (2013), http://dx.doi.org/10.1016/j.fbp.2013.12.001
natant, the beads were resuspended in tartaric buffer andcentrifuged. All supernatants were collected and diluted with
the above buffer solution to a constant final volume in orderto determine the bound protein.
IY (%) was determined by Bradford’s method (Bradford,1976), using Coomassie brilliant blue reagent and measuringabsorbance at 595 nm. BSA was used as standard. The amountof bound protein was then determined indirectly from the dif-ference between the amount of protein in solution before andafter immobilization. All measurements were made in tripli-cate and the standard deviations were reported.
2.4. Enzymatic activity assay
Immobilized bromelain activity was assayed using Bz-Phe-Val-Arg-pNA as a substrate, at 25 ◦C in a wine-like acidicmedium (tartaric acid/sodium tartrate 0.03 M, pH 3.2). Sev-eral concentrations of Bz-Phe-Val-ArgpNA, ranging from 0 to200 �M, were tested in the presence of immobilized biocatalyst(100 mg).
Cleavage of the substrate results in release of free pNA,which was detected colorimetrically at 410 nm. Immobilizedbromelain activity was determined measuring the change inabsorbance vs. time for 30 min using a Shimadzu UV 2450(Milan, Italy). The specific activity of immobilized bromelain,calculated in I.U. of pNA produced (εmM = 8.480 mM−1 cm−1 forpNA), was expressed as I.U. mg−1 of immobilized protein (Haleet al., 2005; Benucci et al., 2011). A blank correction was madeusing a sample that did not contain enzymes. All measure-ments were made in triplicate, and the standard deviationswere reported.
2.5. Kinetic study and determination of kineticparameters
A kinetic study was carried out by varying the substrate con-centration (0–200 �M) at 25 ◦C in the presence of immobilizedstem bromelain in a wine-like acidic medium, which alsocontained one of the following potential inhibitors (I) at dif-ferent concentrations: ethanol (0, 12, 18%, v/v), free SO2 (0, 10,25 mg l−1), skin tannin preparation (0, 0.5, 2 g l−1 correspond-ing to 0, 0.25, 0.99 g l−1
gallic acid eq) and seed tannin preparation
(0, 0.5, 2 g l−1 corresponding to 0, 0.30, 1.20 g l−1gallic acid eq).
Kinetic curves were obtained measuring proteolytic activ-ity at different substrate concentrations (7–8 points), makingthree turns of measurements for each concentration. Thekinetic parameters (kcat, Km, Ka) of stem bromelain were deter-mined according to the Michaelis–Menten equation using anonlinear regression procedure (GraphPad Prism 5.0, Graph-Pad software, Inc.).
The KM value reflects the enzyme substrate complex for-mation, whereas kcat measures the number of substratemolecules turned over per enzyme per minute. Moreover, kcat
is indicative of the product release velocity, and representsthe maximum number of moles of substrate converted to theproduct per number of moles of catalyst per unit time. Thisparameter can be obtained from the equation kcat = Vmax/[E]tot,where [E]tot is the enzyme molar concentration. In addition,the Ka, being the ratio kcat/KM, indicates the affinity of theenzyme towards the substrate. It is indicative of both reactionsteps and expresses the overall catalytic efficiency.
2.6. Inhibition study
stem bromelain activity in a wine-like medium: Effect of inhibitors.
For each tested compound, the effect on immobilized brome-lain activity was evaluated, identifying the inhibition type

ARTICLE IN PRESSFBP-460; No. of Pages 6
food and bioproducts processing x x x ( 2 0 1 3 ) xxx–xxx 3
Table 1 – Kinetic parameters of free (F) (Esti et al., 2011) and immobilized (I) stem bromelain towards Bz-Phe-Val-Arg-pNAsubstrate in tartaric buffer (pH 3.2), containing different ethanol amount: 0%, v/v, 12%, v/v and 18%, v/v.
Ethanol (%, v/v) Enzyme Vmax(app) (I.U. mg−1) KM(app) (�M) kcat(app) (min−1) Ka (min−1 �M−1)
0 F 0.44 ± 0.02 118 ± 10 1166.20 ± 0.02 9.89 ± 1.08I 0.247 ± 0.004 39.19 ± 2.09 27.644 ± 0.004 0.71 ± 0.04
12 F 0.46 ± 0.02 250 ± 27 1201.90 ± 0.02 4.81 ± 0.52I 0.243 ± 0.009 49.5 ± 5.5 17.359 ± 0.009 0.35 ± 0.04
18 F 0.48 ± 0.02 305 ± 20 1287.05 ± 0.02 4.22 ± 0.28I 0.214 ± 0.009 57.31 ± 6.48 23.945 ± 0.009 0.42 ± 0.05
Table 2 – Kinetic parameters of free (F) (Esti et al., 2011) and immobilized (I) stem bromelain towards Bz-Phe-Val-Arg-pNAsubstrate in wine-like acidic medium (pH 3.2, ethanol 12%, v/v) containing different concentration of skin and seedtannins.
Tannins Concentration(g l−1
gallic acid eq)Enzyme Vmax(app) (I.U. mg−1) KM(app) (�M) kcat(app) (min−1) Ka (min−1 �M−1)
0 F 0.46 ± 0.02 250 ± 27 1201.90 ± 0.02 4.81 ± 0.52I 0.243 ± 0.009 49.5 ± 5.5 17.359 ± 0.009 0.35 ± 0.04
Skin 0.25 F 0.191 ± 0.008 125.2 ± 15.8 506.147 ± 0.008 4.04 ± 0.51I 0.137 ± 0.003 30 ± 2 9.187 ± 0.003 0.30 ± 0.02
0.99 F 0.143 ± 0.003 79.14 ± 6.52 378.42 ± 0.003 4.78 ± 0.39I 0.095 ± 0.002 20.7 ± 2.1 6.403 ± 0.002 0.31 ± 0.03
Seed 0.30 F 0.149 ± 0.006 95.28 ± 13.02 393.229 ± 0.006 4.13 ± 0.56I 0.091 ± 0.003 25 ± 3 6.099 ± 0.003 0.25 ± 0.03
1.20 F 0.098 ± 0.002 52.64 ± 3.92 259.235 ± 0.002 4.92 ± 0.37I 0.065 ± 0.003 17.4 ± 2.8 4.428 ± 0.003 0.25 ± 0.04
((ircb
3
Bt
spKAiaaliiokiT
3b
Tc
protease (Esti et al., 2011).
0 50 100 150 2000.00
0.05
0.10
0.15
0.20
0.25
S ( M)Spec
ific
activ
ity (I
.U. m
g-1im
m p
rote
in)
Fig. 1 – Specific activity (I.U. mg−1imm protein) of immobilized
protein towards Bz-Phe-Val-Arg-pNA substrate in tartaricbuffer (pH 3.2) in absence (control, �) and in presence of
competitive, uncompetitive and mixed-type), and Ki valueinhibition constant), which reflects the concentration of annhibitor that decreases the rate of an enzyme-catalyzedeaction by 50%. Esti et al. (2011) extensively described the pro-edure applied to identify the inhibition model, as suggestedy Segel (1975).
. Results and discussion
romelain from pineapple stem was covalently linked on chi-osan beads by a direct mechanism, with an IY of 22%.
The inhibitory study was carried out using thistem bromelain-based biocatalyst, whose kineticarameters (KM = 49.49 �M; Vmax = 0.243 I.U. mg−1 and
a = 0.35 min−1 �M−1) were characterized towards Bz-Phe-Val-rg-pNA in a previous work (Benucci et al., 2012). The expected
nhibitors tested were substances usually present in whitend red wines, such as ethanol, grape skin and seed tannins,nd sulphur dioxide. One by one, they were added to a wine-ike acidic medium, over their average range concentration,n order to characterize the individual inhibitory effect onmmobilized protease activity. In order to compare the effectf wine inhibitors on free and immobilized stem bromelain,inetic parameters obtained for the free form, and reported
n a published paper (Esti et al., 2011), are summarized inables 1–3.
.1. Inhibitory effect of ethanol on immobilizedromelain activity
Please cite this article in press as: Esti, M., et al., Immobilized pineapple
Food Bioprod Process (2013), http://dx.doi.org/10.1016/j.fbp.2013.12.001
he kinetic curves obtained in presence of different ethanoloncentrations (0, 12, 18%, v/v) are shown in Fig. 1.
Kinetic parameters of Brim (Table 1) show that, if alco-hol amount stays within 12%, v/v (average concentrationwidely found in wine), Vmax(app) does not change, as previouslyobserved for Brf whereas its effect becomes significant only atabnormal concentration (18%, v/v).
In both free and immobilized protease, KM(app) significantlyincreased with raising alcohol level, but at larger extent for freeform (11 �M for each ethanol degree) respect to the immobi-lized one.
These results indicate that ethanol, up to an average con-centration of 12%, v/v, could be considered a competitiveinhibitor for immobilized stem bromelain, as revealed for free
stem bromelain activity in a wine-like medium: Effect of inhibitors.
different ethanol concentration: 12%, v/v (©) and 18%, v/v(�).
ARTICLE IN PRESSFBP-460; No. of Pages 6
4 food and bioproducts processing x x x ( 2 0 1 3 ) xxx–xxx
Table 3 – Kinetic parameters of free (F) (Esti et al., 2011) and immobilized (I) stem bromelain towards Bz-Phe-Val-Arg-pNAsubstrate in wine-like acidic medium (pH 3.2, ethanol 12%, v/v) containing different free SO2amount: 0 mg l−1, 10 mg l−1
and 25 mg l−1.
SO2 (mg l−1) Enzyme Vmax(app) (I.U. mg−1) KM(app) (�M) kcat(app) (min−1) Ka (min−1 �M−1)
0 F 0.46 ± 0.02 250 ± 27 1201.90 ± 0.02 4.81 ± 0.52I 0.243 ± 0.009 49.5 ± 5.5 17.359 ± 0.009 0.35 ± 0.04
10 F 0.0218 ± 0.0006 56.3 ± 6.3 57.7282 ± 0.0006 1.03 ± 0.12I 0.195 ± 0.007 39.9 ± 4.6 15.70 ± 0.007 0.39 ± 0.05
25 F 0.0088 ± 0.0003 40.0 ± 6.5 23.2420 ± 0.0003 0.58 ± 0.09I 0.155 ± 0.005 30.63 ± 3.64 12.44 ± 0.005 0.41 ± 0.05
mechanisms between inhibitors and enzymes in free or
0 50 10 0 15 0 20 00.00
0.05
0.10
0.15
0.20
0.25A
S ( M)
Spec
ific
activ
ity (I
.U. m
g-1im
m p
rote
in)
0.25Bm
pro
tein
)
The Ki value of Brim (39.4 ± 1.5%, v/v), determined by areplot of KM(app) vs ethanol concentration (Fig. 2), was signif-icantly higher than the Ki value obtained for Brf (11.4 ± 1.0%,v/v, Esti et al., 2011), indicating that immobilized protease ismore resistant to the ethanol inhibition effect than the freeform.
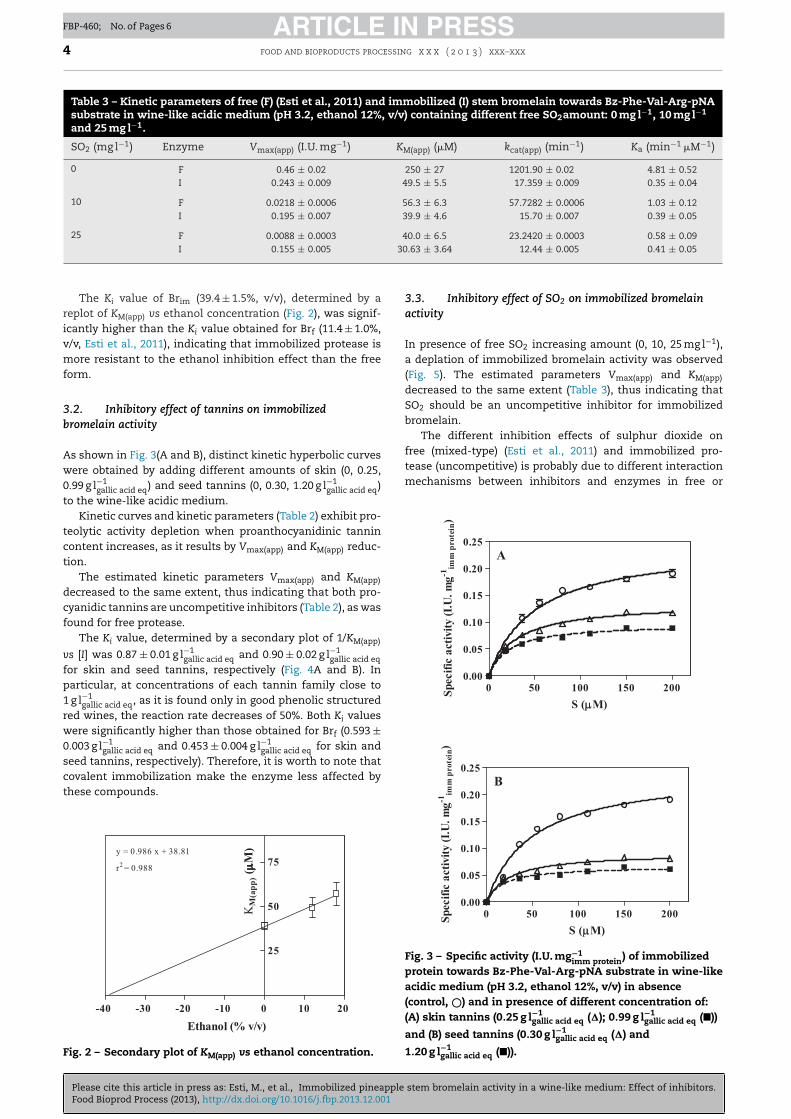
3.2. Inhibitory effect of tannins on immobilizedbromelain activity
As shown in Fig. 3(A and B), distinct kinetic hyperbolic curveswere obtained by adding different amounts of skin (0, 0.25,0.99 g l−1
gallic acid eq) and seed tannins (0, 0.30, 1.20 g l−1gallic acid eq)
to the wine-like acidic medium.Kinetic curves and kinetic parameters (Table 2) exhibit pro-
teolytic activity depletion when proanthocyanidinic tannincontent increases, as it results by Vmax(app) and KM(app) reduc-tion.
The estimated kinetic parameters Vmax(app) and KM(app)
decreased to the same extent, thus indicating that both pro-cyanidic tannins are uncompetitive inhibitors (Table 2), as wasfound for free protease.
The Ki value, determined by a secondary plot of 1/KM(app)
vs [I] was 0.87 ± 0.01 g l−1gallic acid eq and 0.90 ± 0.02 g l−1
gallic acid eqfor skin and seed tannins, respectively (Fig. 4A and B). Inparticular, at concentrations of each tannin family close to1 g l−1
gallic acid eq, as it is found only in good phenolic structuredred wines, the reaction rate decreases of 50%. Both Ki valueswere significantly higher than those obtained for Brf (0.593 ±0.003 g l−1
gallic acid eq and 0.453 ± 0.004 g l−1gallic acid eq for skin and
seed tannins, respectively). Therefore, it is worth to note thatcovalent immobilization make the enzyme less affected by
Please cite this article in press as: Esti, M., et al., Immobilized pineapple
Food Bioprod Process (2013), http://dx.doi.org/10.1016/j.fbp.2013.12.001
these compounds.
-40 -30 -20 -10 0 10 20
25
50
75r2 = 0.98 8
y = 0.986 x + 38.81
Ethanol (% v/v )
K
M(a
pp) (
M)
Fig. 2 – Secondary plot of KM(app) vs ethanol concentration.
3.3. Inhibitory effect of SO2 on immobilized bromelainactivity
In presence of free SO2 increasing amount (0, 10, 25 mg l−1),a deplation of immobilized bromelain activity was observed(Fig. 5). The estimated parameters Vmax(app) and KM(app)
decreased to the same extent (Table 3), thus indicating thatSO2 should be an uncompetitive inhibitor for immobilizedbromelain.
The different inhibition effects of sulphur dioxide onfree (mixed-type) (Esti et al., 2011) and immobilized pro-tease (uncompetitive) is probably due to different interaction
stem bromelain activity in a wine-like medium: Effect of inhibitors.
0 50 10 0 15 0 20 00.00
0.05
0.10
0.15
0.20
S ( M)
Spec
ific
activ
ity (I
.U. m
g-1im
Fig. 3 – Specific activity (I.U. mg−1imm protein) of immobilized
protein towards Bz-Phe-Val-Arg-pNA substrate in wine-likeacidic medium (pH 3.2, ethanol 12%, v/v) in absence(control, ©) and in presence of different concentration of:(A) skin tannins (0.25 g l−1
gallic acid eq (�); 0.99 g l−1gallic acid eq (�))
and (B) seed tannins (0.30 g l−1gallic acid eq (�) and
1.20 g l−1gallic acid eq (�)).

ARTICLE IN PRESSFBP-460; No. of Pages 6
food and bioproducts processing x x x ( 2 0 1 3 ) xxx–xxx 5
-0.9 -0.4 0.1 0.6 1.1
0.02
0.04
0.06r2 = 0.94
y = 0.0264 x + 0.023A
Skin tannins (g l-1gallic acid eq)
1/K
M(a
pp) (
M-1
)
-1.0 -0.5 0.0 0.5 1.0
0.02
0.04
0.06
0.08r2 = 0.89
y = 0.028 x + 0.0252B
Seed tannins (g l-1gallic acid eq)
1/K
M(a
pp) (
M-1
)
Fig. 4 – Secondary plot of 1/KM(app) vs: (A) skin tanninsc
itbot0
lBb
Fpa(1
-40 -30 -20 -10 0 10 20 30
0.01
0.02
0.03
0.04r2 = 0.999
y = 0. 0005 x + 0.02 02
SO2 (mg l-1)
1
/KM
(app
)
Fig. 6 – Secondary plot of: 1/KM(app) vs free SO2
oncentration and (B) seed tannins concentration.
mmobilized form, as a consequence of enzyme conforma-ional changes resulting from the bromelain-support covalentound. For immobilized bromelain, Ki, determined by a replotf 1/KM(app) vs [I], was 40.40 ± 0.54 mg l−1 (Fig. 6), greaterhan Ki and K′
iof free bromelain (4.55 ± 1.07 mg l−1 and
.40 ± 0.09 mg l−1, respectively).These results prove the inhibitory effect of SO2 on brome-
ain activity, even if they clearly show an higher tolerance ofrim respect to Brf. Thus confirms the effectiveness of immo-
Please cite this article in press as: Esti, M., et al., Immobilized pineapple
Food Bioprod Process (2013), http://dx.doi.org/10.1016/j.fbp.2013.12.001
ilized state in reducing enzyme sensitivity to wine inhibitors.
0 50 10 0 15 0 20 00.00
0.05
0.10
0.15
0.20
0.25
S ( M)Spec
ific
acti
vity
(I.U
. mg-1
imm
pro
tein
)
ig. 5 – Specific activity (I.U. mg−1imm protein) of immobilized
rotein towards Bz-Phe-Val-Arg-pNA substrate in wine-likecidic medium (pH 3.2, ethanol 12%, v/v) in absence
control, ©) and in presence of different free SO2 amount:0 mg l−1 (�) and 25 mg l−1 (�).
concentration.
4. Conclusions
For all the wine inhibitors (ethanol, SO2, skin and seed tan-nins) tested in wine-like medium, the Ki values of immobilizedbromelain were significantly higher than the correspondingvalues obtained in a previous work for free enzymes, thus indi-cating that direct covalent immobilization on chitosan beadsmakes protease more resistant to the inhibition effect.
Immobilized protease was affected by ethanol inhibition(competitive type) only if present at abnormal concentration(Ki 39.4%, v/v) for real wine. Grape skin and seed tanninsexerted uncompetitive and equal inhibition, with Ki valuesclose to 1 g l−1
gallic acid eq. The effect of free sulphur dioxide onimmobilized bromelain changed respect to free enzyme, bothbecoming uncompetitive inhibitor and dramatically loweringits strength.
These encouraging results show that immobilized brome-lain could be usefully applied, as an alternative to bentonitefining, for wine protein stabilization by a specific treatment.Further engineering studies will be necessary to design an eas-ily scalable bioreactor, containing the immobilized protease,where wine protein hydrolysis may be carried out.
Notation
GlossaryBSA bovine serum albuminBr bromelain[E]tot total enzyme molar concentrationI inhibitorIY immobilization yield (%)Ka (kcat/KM), affinity constant (min−1 �M−1)kcat (Vmax/[E]tot), turnover number (min−1)Ki inhibition constantK′
iinhibition constant
KM Michaelis–Menten constant (�M)Vmax maximum velocity at which enzyme catalyzes reac-
tion (I.U. mg−1)
Greek symbolsε molar absorptivity (mM−1 cm−1)ε L > subscriptf referred to free enzymeim referred to immobilized enzyme
stem bromelain activity in a wine-like medium: Effect of inhibitors.
(app) apparent kinetic parameter, obtained in presence ofan inhibitor

ARTICLE IN PRESSFBP-460; No. of Pages 6
6 food and bioproducts processing x x x ( 2 0 1 3 ) xxx–xxx
porous copoly(c-methyl-l-glutamate/l-leucine) beads. Eur.
Acknowledgement
This work has been supported by the financial backing of theItalian Ministry of Agricultural, Food and Forestry Policies.
References
Batista, L., Monteiro, S., Loureiro, V.B., Teixeira, A.R., Ferreira, R.B.,2009. The complexity of protein haze formation in wines.Food Chem. 112, 169–177.
Benucci, I., Esti, M., Liburdi, K., Garzillo, A.M.V., 2012. Pineapplestem bromelain immobilized on different supports: catalyticproperties in model wine. Biotechnol. Prog. 28, 1472–1477.
Benucci, I., Liburdi, K., Garzillo, A.M.V., Esti, M., 2011. Bromelainfrom pineapple stem in alcoholic–acidic buffers for wineapplication. Food Chem. 124, 1349–1353.
Bradford, M.M., 1976. A rapid and sensitive method for thequantification of microgram quantities of protein utilizing theprinciple of protein-dye binging. Analyt. Biochem. 72, 248–254.
Esti, M., Benucci, I., Liburdi, K., Garzillo, A.M.V., 2011. Effect ofwine inhibitors on free pineapple stem bromelain activity in amodel wine system. J. Agric. Food Chem. 59, 3391–3397.
Feijoo-Siota, L., Villa, T.G., 2011. Native and biotechnologicallyengineered plant proteases with industrial applications. FoodBioprocess. Technol. 4, 1066–1088.
Gupta, P., Maqbool, T., Saleemuddin, M., 2007. Orientedimmobilization of stem bromelain via the lone histidine on ametal affinity support. J. Mol. Catal. B: Enzym. 45, 78–83.
Hale, L.P., Greer, P.K., Trinh, C.T., James, C.L., 2005. Proteinaseactivity and stability of natural bromelain preparations. Int.Immunopharmacol. 5, 783–793.
Hsu, J.C., Heatherbell, D., 1987. Heat-unstable proteins in wine. I.Characterization and removal by bentonite fining and heattreatment. Am. J. Enol. Vitic. 38, 11–16.
Kim, J.D., 2005. Immobilization of keratinase from aspergillusflavus K-03 for degradation of feather keratin. Mycobiology 33(2), 121–123.
Krajewska, B., 2004. Application of chitin- and chitosan-based
Please cite this article in press as: Esti, M., et al., Immobilized pineapple
Food Bioprod Process (2013), http://dx.doi.org/10.1016/j.fbp.2013.12.001
materials for enzyme immobilization: a review. EnzymeMicrob. Technol. 35, 126–139.
Mahmood, R., Saleemuddin, M., 2007. Additional stabilization ofstem bromelain coupled to a thermosensitive polymer byuniform orientation and using polyclonal antibodies.Biochemistry 72, 307–312.
Segel, I.H., 1975. Enzyme Kinetic. Behaviour and Analysis ofRapid Equilibrium and Steady-State Enzyme Systems. AWiley-Interscience Publication, John Wiley & Sons Inc., NewYork.
Segundo, M.A., Rangel, A.O.S.S., 2001. A gas diffusion sequentialinjection system for the determination of sulphur dioxide inwines. Anal. Chim. Acta 427, 279–286.
Seo, H., Itoyama, K., Morimoto, K., Takagishi, T., Oka, M., Hayashi,T., 1998. Spacer effects on enzymatic activity of bromelainimmobilized onto porous chitosan beads. Eur. Polym. J. 34,917–922.
Shi, W., Wei, M., Jin, L., Li, C., 2007. Calcined layered doublehydroxides as a biomolecular vessel for bromelain:Immobilization, storage and release. J. Mol. Catal. B: Enzym.47, 58–65.
Tan, Y.L., Liu, C.G., Yu, L.J., Chen, X.G., 2008. Effect of linoleic-acidmodified carboxymethyl chitosan on bromelainimmobilization onto self-assembled nanoparticles. Front.Mater. Sci. China 2, 209–213.
Toniolo, R., Pizzariello, A., Susmel, S., Dossi, N., Bontempelli, G.,2010. A sensor based on electrodes supported onion-exchange membranes for the flow-injection monitoring ofsulphur dioxide in wines and grape juices. Talanta 80,1809–1815.
Vincenzi, S., Marangon, M., Tolin, S., Curioni, A., 2011. Proteinevolution during the early stages of white wine making andits relations with wine stability. Aust. J. Grape Wine Res. 17,20–27.
Waters, E.J., Alexander, G., Muhlack, R., Pocock, K.F., Colby, C.,O’neill, B.K., Høj, P.B., Jones, P., 2005. Preventing proteinhaze in bottled white wine. Aust. J. Grape Wine Res. 11,215–225.
Yodoya, S., Takagi, T., Kurotani, M., Hayashi, T., Furuta, M., Oka,M., Hayashi, T., 2003. Immobilization of bromelain onto
stem bromelain activity in a wine-like medium: Effect of inhibitors.
Polym. J. 39, 173–180.