immunolocalization of keratan sulphate in the human...
TRANSCRIPT

J. Cell Sci. 80, 181-191 (1986) 181Printed in Great Britain © The Company of Biologists Limited 1986
IMMUNOLOCALIZATION OF KERATAN SULPHATEIN THE HUMAN EMBRYONIC CORNEA AND OTHERHUMAN FOETAL ORGANS
LOUISE HYLDAHL12, RICHARD ASPINALL2 AND FIONA M. WATT3
1 Department of Ophthalmology, Karolinska Hospital, S-104 01 Stockholm, Stueden2Department of Zoology, University of Oxford, South Parks Road, Oxford 0X1 3PS, U.K.3 Molecular Cell Biology Laboratory, The Mathilda and Terence Kennedy Institute ofRheumatology, 6 Bute Gardens, Hammersmith, London W6 7DW, U.K.
SUMMARYAn antibody raised against keratan sulphate was used to localize keratan sulphate in the human
embryonic cornea and other human foetal organs. It was found that keratan sulphate was present inthe corneal endothelium, as well as in the corneal stroma. In contrast, no keratan sulphate could bedemonstrated in the corneal epithelium or in any posterior parts of the human foetal eye. Humancorneal stromal cells grown in monolayers in tissue culture lost their capacity to bind this antibody.This suggests that explantatiorw'n vitro decreases or alters the expression of keratan sulphate in thiscell type. Keratan sulphate was found to be present in abundant quantities in human embryoniccartilage and to a lesser extent in placenta. In contrast, keratan sulphate could not be detected in thefoetal intestine, liver, suprarenal glands, the umbilical cord or in the foetal yolk sack.
INTRODUCTION
The human cornea consists of three different cellular layers: an anteriorepithelium, an intermediate stroma and an endothelial monolayer that lines theposterior surface of the cornea (Hogan, Alvarado & Weddell, 1971). The stroma isthe thickest section of the cornea and contains a fibroblastic cell type, the stromalcell, which is derived from the neural crest during embryogenesis (Hay, 1981). Thematrix of the stroma consists of highly ordered orthogonal lamellae composed of typeI collagen fibrils (Trelsted & Couloumbre, 1971) and of proteoglycans that containkeratan sulphate and chondroitin sulphate (Antonopoulos, Axelsson, Heinegard &Gardell, 1974; Axelsson & Heinegard, 1975; Muthiah, Stuhlsatz & Greiling, 1974).The organization of the collagen and proteoglycans in the stroma as well as themacromolecular characteristics of the proteoglycans are thought to be responsible forthe structure and transparency of the cornea. Although the biochemical charac-teristics of corneal proteoglycans have been extensively studied (Anseth, 1961;Axelsson & Heinegard, 1978, 1980; Coster, Cintron, Damle & Gregory, 1983; Dahl,Johnsen, Anseth & Prydz, 1974; Dahl & Laurent, 1982; Faltz et al. 1979; Gregory,Coster & Damle, 1982; Handley & Phelps, 1972; Hart, 1976, 1978; Hart & Lennarz,1978; Hassel, Newsome & Hascall, 1979; Klintworth & Smith, 1983; Matthews &Cifonelli, 1965; Meier & Hay, 1974; Meyer, Linker, Davidson & Weismann, 1953;
Key words: keratan sulphate, cornea, stroma, endothelium.

182 L. Hyldahl, R. Aspinall and F. M. Watt
Toole & Trelstad, 1971; Trelstad, Hayashi & Toole, 1974), less is known about thesites of their production and how keratan sulphate is distributed in situ.
The present study was undertaken to examine the distribution of keratan sulphatein the human embryonic cornea as well as in other human foetal organs. For thispurpose we took advantage of a recently developed monospecific antibody, whichwas raised against keratan sulphate (Zanetti, Ratcliffe & Watt, 1985). We report herethat this antibody can be used to trace keratan sulphate in human foetal eyes as well asin other parts of the human embryo.
MATERIALS AND METHODS
Primary materialThe primary material used in this study was received from 10- to 12-week-old human embryos
obtained by a vacuum extraction abortion method (Brody, 1980). No apparently misdevelopedmaterial was used, as judged by morphological examination. The foetal age in each case wasestimated according to Shi et al. (1985). All material was processed within 6h after surgery. Thefoetal specimens delivered in collection vessels were initially diluted with an approximately equalvolume of dextrose saline (4% dextrose (w/v) in 0-18% (w/v) aqueous NaCl, pH7-4, purchasedfrom Steriflex pic, Nottingham, U.K.), sieved through a domestic plastic sieve with a hole size of1 mmXI mm; whereafter excess blood and debris were washed through the sieve with two rinseseach of approximately 200 ml of phosphate-buffered saline (PBS) lacking calcium and magnesiumions at pH7-3 (solution A as described by Dulbecco & Vogt (1954) and obtained from Oxoid Ltd,U.K.). The contents of the sieve were then shaken and rinsed into a 37cm X 27cm white traycontaining 700-1000 ml of PBS. Despite several transfers from one vessel to another, eye globes,umbilical cords, placental fragments, yolk sacks and other organs were frequently found.
Tissue fixation and processingFor histology the organs were fixed between 4 and 6h after foetal aspiration. Routine histology
was performed on formol-saline fixed material, which was subsequently embedded in paraffin andcut into 5 /im thick sections. The sections were stained in haematoxylin/eosin and examined andphotographed in a Leitz inverted microscope with an attached camera system. For antigenlocalization foetal organs were snap-frozen in liquid nitrogen and cut with a cryostat into 10 fimthick sections. These sections were attached to Hendley Essex multispot glass slides precoated with1 % (w/v) gelatin and 0-01 % (w/v) chrom alum in Analar water, air dried and stored at —70°Cuntil use.
Establishment of primary cultures from embryonic corneasThoroughly rinsed eye globes were microdissected and the entire cornea was cut out with a pair
of corneal scissors to leave the limbus with the remaining eye bulb. After a thorough rinse in PBSthe corneal button was placed with the endothelial side down (Hyldahl, Auer & Sundelin, 1982) ina 1 cm2 well of a NUNC 24-well plate containing a 1:1 (v/v) mixture of Ham's F12 and Dulbecco'sModified Eagle's Medium (DME) (Morton, 1970) supplemented with 10% foetal calf serum, 50units of penicillin per ml, 50/ig of streptomycin sulphate per ml and 0-5 \i% Fungizone per ml. Allwells had been precoated with fibronectin (10/igml"1 purified according to Engvall & Ruoslahti(1977) for 1 h at 4°C). The cultures were then stored in a humidified 5 % COz/95 % air mixture at37CC for up to 10 days. The primary cultures were either fixed and prepared for antigen localizationor removed for transfer by a trypsinization procedure. The cultures were briefly rinsed inprewarmed PBS and treated with cold (4°C) 0-25% trypsin (1:250, w/v; obtained from DifcoCo., Detroit Mi, U.S.A.) in PBS until all cells had detached from the wells. The cells were thenresuspended in 10% (v/v) foetal calf serum and spun at 200^ for 2-3 min. The supernatant wasdiscarded and the pellet resuspended in 10% (v/v) serum. The suspension was then seeded onto
Keratan sulphate in embryonic cornea 183
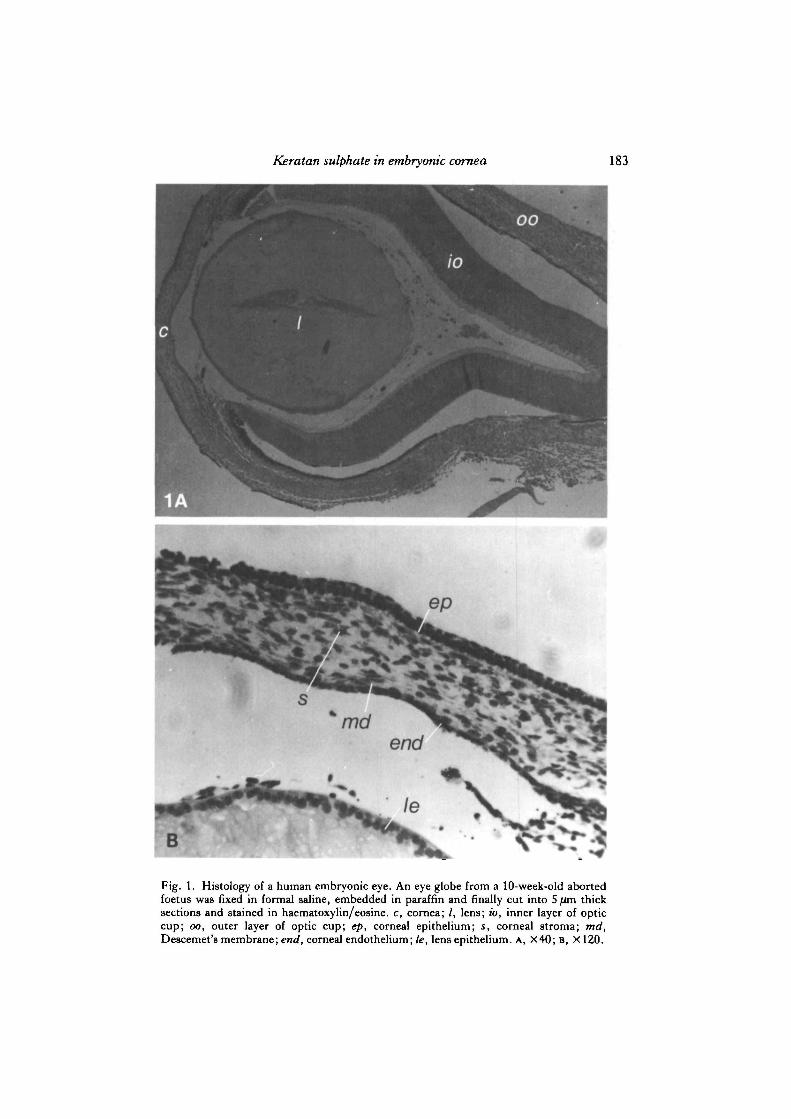
Fig. 1. Histology of a human embryonic eye. An eye globe from a 10-week-old abortedfoetus was fixed in formal saline, embedded in paraffin and finally cut into 5 /an thicksections and stained in haematoxylin/eosine. c, cornea; /, lens; to, inner layer of opticcup; oo, outer layer of optic cup; ep, corneal epithelium; s, corneal stroma; md,Descemet's membrane; end, corneal endothelium; le, lens epithelium. A, X40; B, X120.
184 L. Hyldahl, R. Aspinall and F. M. Watt

Keratan sulphate in embryonic cornea 185
Table 1. Expression of keratan sulphate in cornea! cells in vivo and in vitroCell type MZ15 staining
Corneal endothelium in vivo +
Corneal stroma in vivo +
7- to 10-day-old primary explants from corneas fixed in:(a) 3 -7 % neutral formaldehyde and postfixed in methanol on ice —(b) Air dried and postfixed in cold acetone —(c) Saint-Marie's fixative —(d) 20 % acetic acid/80 % methanol
Passage 10 stromal cells fixed in:(a) 3 -7 % neutral formaldehyde and postfixed in methanol on ice —(b) Air dried and postfixed in cold acetone —(c) Saint-Marie's fixative —(d) 20 % acetic acid/80 % methanol(e) Unfixed and stained live —
gelatinized 50mm tissue culture dishes and split at a routine ratio of 1:5 every 4—7 days. Primarycultures as well as passage 10 cultures were used for antigen localization in this study.
Cell fixation and processingGlass slides with 7- to 10-day-old primary cultures or passage 10 stromal cells (Hyldahl, 1984)
were subjected to antigen analysis and prepared by four different means of fixation. First, cells wereair dried at room temperature and postfixed in acetone for 15 s at 4°C. Secondly, the cells werefixed in 20 % acetic acid/80 °h ethanol (v/v) for 10 min. Thirdly, the cells were fixed in 96 % (v/v)aqueous ethanol/glacial acetic acid, 99:1 (v/v), Engelhardt's modification of Saint-Marie's fixative(Saint-Marie, 1962; Engelhardt, Goussev, Shipova & Abelev, 1971). Fourthly, the cells were fixedin 3-7% formaldehyde in PBS (v/v) for 8 min at room temperature followed by incubation inabsolute methanol on ice for 5 min (Zanetti et al. 1985).
Keratan sulphate antibody MZ15MZ15 is a monoclonal antibody to keratan sulphate, which recognizes an antigenic determinant
that is sensitive to keratanase digestion (Zanetti et al. 1985). It was obtained from a fusion of NS1myeloma cells with spleen cells of a Balb/c mouse that had been immunized with pig laryngealchondrocytes. The antibody has wide species cross-reactivity and recognizes both corneal andskeletal keratan sulphate (Zanetti et al. 1985).
ImmunolocalizationSlides with sections of human foetal organs, primary corneal explants or passage 10 stromal cells
were rinsed briefly in PBS at room temperature. The slides were subsequently incubated in MZ15
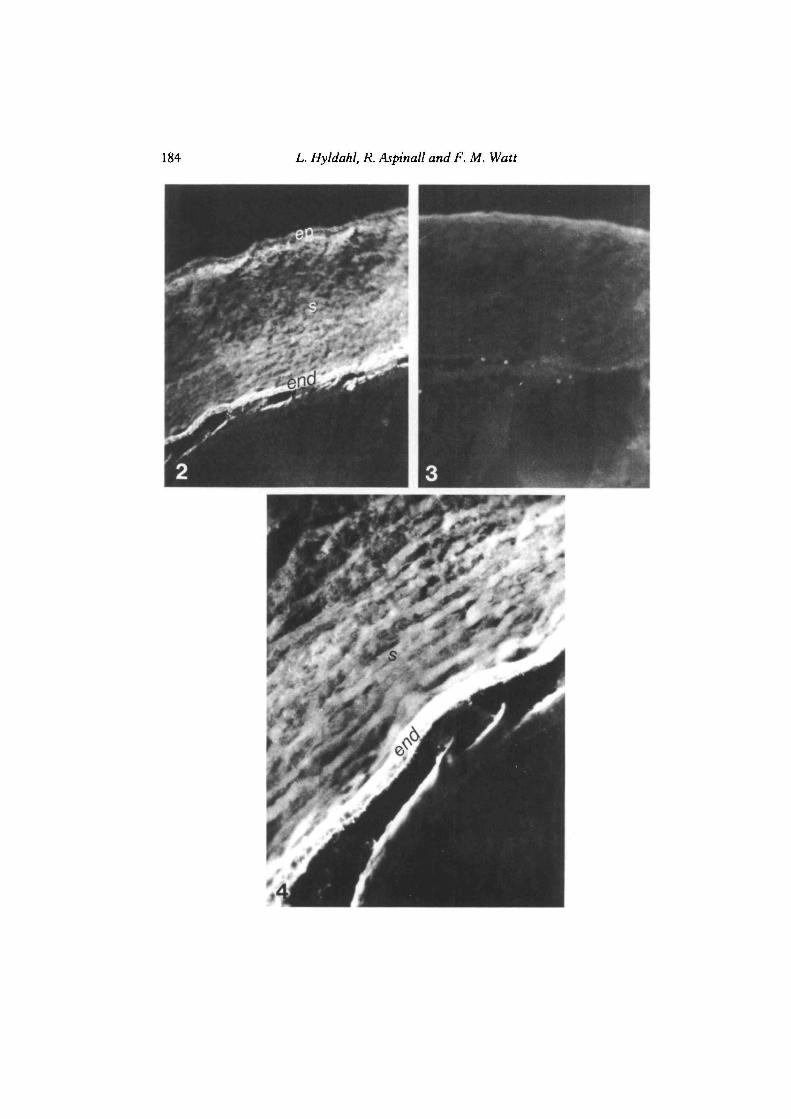
Figs 2—4. Localization of keratan sulphate in the anterior portion of the humanembryonic eye. An eye globe from an 11-week-old human aborted foetus was snap-frozenin liquid nitrogen cut into 5 /tfn saggital sections with a cryostat. ep, corneal epithelium; s,corneal stroma; end, corneal endothelium; le, lens epithelium.
Fig. 2. Incubated with MZ15 antibody at 1/1000 and subsequently with FITC-conjugated rabbit anti-mouse IgG at 1/30 as described in Materials and Methods. X280.
Fig. 3. Reacted with second antibody, i.e. FITC-conjugated rabbit anti-mouse IgG at1/30 only. X280.
Fig. 4. As in Fig. 2 but at X800.

186 L. Hyldahl, R. Aspinall and F. M. Watt
Table 2. Expression of keratan sulphate in human foetal tissues as revealed byMZ15 staining
Placenta (+)Umbilicus -Yolk sackCartilage +Intestine -Liver —Suprarenal glands —Foetal eye:
Cornea +Lens —Inner layer of optic cup —Outer layer of optic cup —Retina -
Organs from 10- to 12-week-old human embryos were snap-frozen in liquid nitrogen, cut with acryostat and subjected to MZ15 binding analysis as described in legend to Fig. 2. +, clearlypositive; (+) weakly positive; —, no detectable binding above background level.
(ascitic fluid diluted at 1/1000 in PBS supplemented with 2% foetal calf serum (v/v) and 0-1 %(w/v) sodium azide) in a humid atmosphere at room temperature for 30 min. After a brief rinse inPBS the specimens were incubated in fluorescein isothiocyanate(FITC)-conjugated rabbit anti-mouse immunoglobulin G (IgG) (Miles, diluted to 1/30) for 30 min at room temperature as above.The reaction was terminated by a rinse in PBS, whereafter the slides were mounted in 30%glycerol/70% PBS (v/v). Slides exposed to second antibody only were used as negative controls.In one case (snap-frozen placenta) the background binding of the second antibody was too high todraw any conclusions about MZ15 reactivity. It was assumed that this was due to the presence of Fcreceptors in placental tissue, and placental sections were therefore preincubated in human plasmadiluted 1:50 in PBS (v/v) for 20 min to block the Fc receptors and then briefly rinsed in PBS beforeMZ1S staining. The slides were examined in a Leitz photomicroscope using epifluorescenceillumination. To produce photographs of fluorescent images, Ilford HP5 400 ASA/27 DIN filmwas used.
RESULTS
Fig. 1 shows histological cross-sections from a 10-week post-fertilization humanembryonic eye, from which it can be seen that the cornea is clearly separated from theanterior surface of the lens (Fig. 1A). Fig. 1B shows that the embryonic cornea, likeits adult counterpart, consists of an outer epithelium, an intermediate stroma and an
Fig. 5. Localization of keratan sulphate in the cartilage end of a human foetal rib. Theventromedial end of the rib was frontally cut in order to obtain sections containing bothcartilage and bone. The section was stained with MZ15 antibody at 1/1000 and FITC-conjugated second antibody as described in Materials and Methods. X280.Fig. 6. Localization of keratan sulphate in the human first trimester placenta. Pieces ofplacental tissue were snap-frozen in liquid nitrogen, cut, preincubated in human plasmaat 1/50 (v/v) in PBS, stained with MZ15 antibody at 1/1000, and FITC-conjugatedsecond antibody as described in Materials and Methods. X280.

Keratan sulphate in embryonic cornea 187

188 L. Hyldahl, R. Aspinall and F. M. Watt
inner endothelial monolayer. Thus, it was possible to search for keratan sulphate inall three layers.
Fig. 2 shows the localization of keratan sulphate in the embryonic cornea asrevealed by antibody MZ15 staining. The staining was most intense in the endo-thelium and around Descemet's membrane, the basal membrane that separates theendothelium from the corneal stroma. The intermediate stroma was also stained withMZ1S, albeit at lower intensity. This difference becomes more evident in Fig. 4,which shows a section of Fig. 2 at higher magnification. In contrast, the cornealepithelium was found to be MZ15-negative. Staining with the second FITC-conjugated antibody revealed a weak background staining evenly distributed over theentire cornea (Fig. 3). It has been previously suggested that corneal cells explantedin vitro alter their expression of proteoglycans (Conrad & Dorfman, 1974; Dahl &Coster, 1978; Gnadinger and Schwager-Hubner, 1975a,b; Klintworth & Smith,1976, 1981; Yue, Baum & Silbert, 1976). To test this hypothesis we have explantedcells from human embryonic corneas and subjected them to 7-10 days' incubation invitro. This interval was chosen because it takes at least 7 days for cells from differentcorneal layers to detach fully from the corneal bulk and attach firmly to the tissue-culture surface (Hyldahl et al. 1982; Hyldahl, 1984, 1985, and unpublished ob-servations). Different fixation procedures were thereafter used, as described inTable 1, but in no case did we detect any MZ 15-positive cells. Nor did we reveal anybinding of this antibody to human corneal stromal cells that had undergone 10passages after explantation in vitro (Table 1). This suggests that corneal cells grownin vitro are phenotypically different from their in vivo counterparts — at least withrespect to MZ15 staining.
In order to study the expression of keratan sulphate in other parts of the humanembryo, different organs from 10- to 12-week-old human embryos were collected,snap-frozen, cut with a cryostat and finally stained with MZ15. The results aresummarized in Table 2. It was found that keratan sulphate could not be detected inthe umbilical cord or in the foetal yolk sack. Nor did MZ15 bind to foetal intestine,liver, suprarenal glands or to the posterior parts of the human foetal eye (i.e. theinner or outer layers of the optic cup or the foetal lens). In contrast, we found thatfoetal cartilage bound this antibody in abundant quantities (Fig. 5). We alsoobserved a traceable binding of MZ15 to sections of human placenta (Fig. 6).
DISCUSSION
This study has taken advantage of the recent development of a monoclonalantibody to keratan sulphate (Zanetti et al. 1985). The antibody MZ15 was used tostudy the distribution of keratan sulphate in the human embryonic cornea. It wasfound that MZ15 bound to human corneal stroma, suggesting the presence of keratansulphate in this tissue. This result is in line with a number of reports showing thatkeratan sulphate is an important constituent of corneal stroma in a variety of species(Anseth, 1961; Axelsson & HeinegSrd, 1975; Dahl, Johnson, Anseth & Prydz, 1974;Faltze* al. 1979; Gregory^ al. 1982; Hart, 1976, 1978; Klintworth & Smith, 1983;

Keratan sulphate in embryonic cornea 189
Meier & Hay, 1974; Toole & Trelsted, 1974). However, we found that the binding ofMZ15 was significantly more intense to the corneal endothelium and Descemet'smembrane. Even though it has been reported that Descemet's membrane frombovine eyes contains keratan sulphate (reviewed by Gospodarowicz & Fuiji, 1981;Hyldahl, 1983) this is the first time that human embryonic corneal endothelial cellshave been shown to contain large quantities of keratan sulphate. Sections from otherhuman foetal organs were used as controls and it was found that foetal cartilagestained intensely with MZ15. Placental sections also stained with MZ15, albeit atlower intensity. In contrast, umbilicus and foetal yolk sack were found to be MZ15-negative. Other differentiated organs of the human embryo such as intestine, liver,the suprarenal glands, the inner and outer layers of the optic cup and the foetal lenswere also negative with respect to the binding of this antibody.
When cells from the human embryonic cornea were explanted and allowed toproliferate into a cell monolayer, they lost their reactivity with the MZ15 antibody.No binding could be detected irrespective of fixation procedure either in primary orlate-passage cultures. This may well represent an adaptation to in vitro conditions,which involves a decrease in the keratan sulphate expression. Even though it has beenshown by Dahl et al. (1974) that rabbit stromal cells in culture synthesize smallquantities of keratan sulphate, it is significant that we have been unable to trace anyMZ15-positive material in either early or late-passage cultures of cells from humanembryonic corneas. A possible but less probable explanation for this apparentinconsistency may be that a putative keratan sulphate molecule produced by stromalcells grown in tissue culture is qualitatively different from the keratan sulphatedetected in situ. It then follows that the MZ15 antibody would fail to detect suchmolecules. This hypothesis would be in line with a recent report by Kenney, Benya,Nimni & Smith (1981), who clearly demonstrated that rabbit corneal endothelialcells dramatically alter their pattern of collagen synthesis after explantation in vitro.It remains to be shown, however, that human embryonic corneal stromal cells grownin culture produce a keratan sulphate molecule that is different from that produced inthe human cornea in vivo.
The development of a monoclonal antibody that recognizes keratan sulphate inhuman corneal tissue has important practical implications. The antibody may beuseful in detecting aberrant keratan sulphate distribution patterns in human corneas.Among the disorders in which such a diagnostic tool would be helpful is the macularcorneal dystrophy — a recessively inherited disease, which is characterized by ageneral diffuse haze throughout the corneal stroma and the accumulation ofirregularly shaped deposits that tend to be anterior in the central region of the stroma(Maumenee, 1981). Recent studies (Hassel, Newsome, Krachmer & Rodriguez,1980) have demonstrated that the cloudiness in corneas from patients with macularcorneal dystrophy results from the failure to synthesize a keratan sulphate or amature keratan sulphate proteoglycan. It can thus be envisaged that a monoclonalantibody that recognizes human keratan sulphate in tissue sections will eventuallybecome useful also in medical practice..

190 L. Hyldahl, R. Aspinail and F. M. Watt
The authors thank Professor Christopher F. Graham FRS for valuable support throughout thisstudy. This work was supported by the Cancer Research Campaign. Louise Hyldahl was onsabbatical leave at the University of Oxford supported by Carmen and Bertil Regner's fund forOphthalmological Research, Axel Hirsch's travel fund for surgeons, The International federationfor University Women (IFUW), the Swedish Society for Medicine and the research funds ofKarolinska Institutet.
REFERENCES
ANSETH, A. (1961). Studies on corneal polysaccharides. III . Topographic and comparativebiochemistry. Expl Eye Res. 1, 106-112.
ANTONOPOULOS, C. A., AXELSSON, I., HEINEGARD, D. & GARDELL. S. (1974). Extraction and
purification of proteoglycans from various types of connective tissue. Biochim. biophys. Acta 338,108-119.
AXELSSON, I. & HEINEGARD, D. (1975). Fractionation of proteoglycans from bovine cornealstroma. Biochem.J. 145, 491-500.
AXELSSON, I. & HEINEGARD, D. (1978). Characterization of the keratan sulphate proteoglycansfrom bovine corneal stroma. Biochem.J. 169, 517-530.
AXELSSON, I. & HEINEGARD, D. (1980). Characterization of chondroitin sulphate rich proteo-glycans from bovine corneal stroma. Expl Eye Res. 30, 57-66.
BRODY, S. (1980). Obstetrics and Gynaecology. Stockholm: Almquist & Wicksell.CONRAD, G. W. & DORFMAN, A. (1974). Synthesis of sulphated mucopolysaccharides by chick
corneal fibroblasts in vitro. Expl Eye Res. 18, 421-434.COSTER, L., CINTRON, C , DAMLE, S. P. & GREGORY, J. D. (1983). Proteoglycans of rabbit
cornea. Labelling in organ culture and in vivo. Expl Eye Res. 36, 517—530.DAHL, I. M. & COSTER, L. (1978). Proteoglycan biosynthesis in cultures of corneas and corneal
stroma cells from adult rabbit. Expl Eye Res. 27, 175-190.DAHL, I. M., JOHNSEN, W., ANSETH, A. & PRYDZ, H. (1974). The synthesis of glycosamino-
glycans by corneal stroma cells in culture. Expl Cell Res. 88, 193—197.DAHL, I. M. & LAURENT, T. C. (1982). Synthesis of glycosaminoglycans in corneal organ cultures.
Expl Eye Res. 34, 83-98.DULBECCO, R. & VOGT, M. (1954). Plaque formation and isolation of pure lines with poliomyelitis
virus. J. exp. Med. 99, 167-182.ENGELHARDT, N., GOUSSEV, A. I., SHIPOVA, L. J. & ABELEV, G. I. (1971). Immunofluorescent
study of an alphafetoprotein in liver and liver rumours. I. Techniques of alphafetoproteinlocalization in tissue sections. Int.jf. Cancer 7, 198—206.
ENGVALL, E. & RuoSLAHTi, E. (1977). Binding of a soluble form of fibroblast cell surface protein -fibronectin - to collagen. Int.jf. Cancer 10, 1-5.
FALTZ, L., CAPUTO, C , KIMURA, J., SCHRODE, J. & HASCALL, V. C. (1979). Structure of the
complex between hyaluronic acid, hyaluronic acid binding region and the link protein ofproteoglycan aggregates from the Swarm rat chondrosarcoma. J. biol. Chem. 254, 1381-1387.
GNADINGER, M. C. & SCHWAGER-HOBNER, M. E. (1975a). Biosynthesis of glycosaminoglycans bymammalian corneal epithelium and fibroblasts in vitro. I. Isolation and fractionation differencesof GAG from the two cell types. Albrecht v Graefes Arch. klin. exp. Ophthal. 196, 9-19.
GNADINGER, M. C. & SCHWAGER-HOBNER, M. E. (19756). Biosynthesis of glycosaminoglycans bymammalian corneal epithelium and fibroblasts in vitro. II . Approach to specify the GAG fromthe two cell types. Albrecht v Graefes Arch. klin. exp. Ophthal. 196, 21-30.
GOSPODAROWICZ, D. & Fura, D. (1981). The extracellular matrix and the control of cellproliferation and differentiation. In Cellular Responses to Molecular Modulators (ed. L. W.Mozes etal.). New York, London: Academic Press.
GREGORY, J. D., COSTER, L. & DAMLE, S. P. (1982). Proteoglycans of rabbit corneal stroma.Isolation and partial characterization, j . biol. Chem. 257, 6965-6970.
HANDLEY, C. J. &PHELPS, C. F. (1972). The biosynthesis in vitro of keratan sulphate in the bovinecornea. Biochem.J. 128, 205-213.
HART, G. W. (1976). Biosynthesis of glycosaminoglycans during corneal development. J. biol.Chem. 251, 6513-6521.

Keratan sulphate in embryonic cornea 191
HART, G. W. (1978). Glycosaminoglycan sulfotransferases of the developing cornea. J. biol. Chem.253, 347-353.
HART, G. W. & LENNARTZ, B. (1978). Effects of tunicamycin on the biosynthesis of glycosamino-glycans by embryonic chick cornea. J. biol. Chem. 253, 5795-5801.
HASSEL, J. R. & NEWSOME, D. A. & HASCALL, V. C. (1979). Characterization and biosynthesis ofproteoglycans of comeal stroma from Rhesus monkey. J . biol. Chem. 254, 12346-12354.
HASSEL, J. R., NEWSOME, D. A., KRACHMER, J. H. & RODRIGUEZ, M. M. (1980). Macular ComealDystrophy — failure to synthesize a mature keratan sulphate proteoglycan. Proc. natn. Acad. Sci.U.SA. 77, 3705-3709.
HAY, E. D. (1981). Cell Biology of Extracellular Matrix. New York: Plenum Press.HOGAN, M. J., ALVARADO, J. A. & WEDDELL, J. E. (1971). Histology of the Human Eye.
Philadelphia: Saunders.HYLDAHL, L. (1983). Permissive effect of the extracellular matrix on attachment, maintenance and
proliferation of normal and malignant cells in vitro. Proc. Int. Conf. Chemother. 235, 37-45.HYLDAHL, L. (1984). Primary cultures from human embryonic corneas. J. Cell Sci. 66, 343-351.HYLDAHL, L. (1985). Proliferation of human embryonic corneal stromal cells in a defined
serumfree medium. Cell. Biol. Int. Rep. 9, 655-662.HYLDAHL, L., AUER, G. & SUNDELIN, B. (1982). A novel method to establish primary cultures of
bovine endothelial cells. Cell Biol. Int. Rep. 6, 523-528.KENNEY, M. C.,"BENYA, P. D., N M N I , M. E. & SMITH, R. E. (1981). Stability of the collagen
phenotype and decreased collagen production in serial subcultures of rabbit comeal endothelialcells. ExplEyeRes. 33, 131-140.
KLINTWORTH, G. K. & SMITH, C. F. (1976). A comparative study of extracellular sulphatedglycosaminoglycans synthesized by rabbit comeal fibroblasts in organ and confluent cultures.Lab. Invest. 35, 258-263.
KLINTWORTH, G. K. & SMITH, C. F. (1981). Difference between the glycosaminoglycanssynthesized by corneal and cutaneous fibroblasts in culture. Lab. Invest. 44, 553—559.
KLINTWORTH, G. K. & SMITH, C. F. (1983). Abnormalities of proteoglycans and glycoproteinssynthesized by comeal organ cultures derived from patients with macular comeal dystrophy.Lab. Invest. 48, 603-612.
MAUMENEE, I. (1978). The cornea in connective tissue diseases. Proc. Am. Acad. Ophthal.Otolaryng. 85, 1014-1017.
MEIER, S. & HAY, E. D. (1974). Stimulation of extracellular matrix synthesis in the developingcornea by glycosaminoglycans. Proc. natn. Acad. Sci. U.SA. 71, 2310—2313.
MEYER, K., LINKER, A., DAVIDSON, E. A. & WEISMANN, B. (1953). The mucopolysaccharides of
bovine cornea..?, biol. Chem. 205, 611-616.MORTON, H. (1970). A survey of commercially available tissue culture media. In Vitro 6, 89-108.MUTHIAH, P., STUHLSATZ, H. W. & GREILING, H. (1974). Composition of comeal proteoglycans.
Density gradient centrifugation and chromatographic studies. Hoppe-Seyler's Z. pkysiol. Chem.355, 924-934.
SAINT-MARIE, G. (1962). A paraffin embedding technique for studies employing immunofluor-escence-7- Histochem. Cytochem. 10, 250-256.
SHI, W. K., HOPKINS, B., THOMPSON, S., HEATH, J. K., LUKE, B. M. & GRAHAM, C. F. (1985).Synthesis of apolipoproteins, alphafetoprotein, albumin and transferrin by the human foetal yolksack and other foetal organs..7. Embryol. exp. Morph. 85, 191-206.
TOOLE, B. & TRELSTAD, R. (1971). Hyaluronate production and removal during comealdevelopment in the chick. Devi Biol. 26, 28—35.
TRELSTAD, R. L. & COULOUMBRE, A. J. (1971). Morphogenesis of the collagenous stroma in thechick cornea. J. Cell Biol. 50, 840-858.
TRELSTAD, R., HAYASHI, K. & TOOLE, B. (1974). Epithelial collagens and glycosaminoglycans inthe embryonic cornea. J . Cell Biol. 62, 815-830.
YUE, B., BAUM, J. L. & SILBERT, J. E. (1976). The synthesis of glycosaminoglycans by cultures ofrabbit corneal endothelial and stromal cells. Biochem.J. 158, 567-573.
ZANETTI, M., RATCUFFE, A. & WATT, F. M. (1985). Subpopulations of differentiated chondro-cytes identified with a monoclonal antibody to keratan sulphate. J. Cell Biol. 101, 53-59.
(Received 13 June 1985 -Accepted 15 July 1985)
