immunological control of viral and bacterial pathogens
TRANSCRIPT

Immunological Control of Viral andBacterial Pathogens
Josh Slater, BVM&S, MA, PhD, MRCVS
Author’s address: University of Cambridge, Department of Clinical Veterinary Medicine, MadingleyRoad, Cambridge CB3 OES, UK. © 2000 AAEP.
Introduction
The majority of the commercial vaccines availablefor common equine pathogens like equine influenzavirus (EIV), equine herpesvirus-1 (EHV-1) andStreptococcus equi (S. equi) are killed vaccines con-sisting of the inactivated pathogen or its proteinswith an adjuvant, generally alum.1–3 The estab-lished measure of efficacy for these vaccines is theserum antibody (IgM and IgG) response after vacci-nation. Killed vaccines containing alum adjuvantproduce impressive serum antibody titers by stimu-lating type 2 T helper lymphocyte (TH2) responses,inducing B lymphocyte activation and thus efficientantibody production. However, as reviewed by Dr.Lunn earlier, an effective immune response consistsof much more than simply circulating antibody.
Generating a full and effective repertoire of adap-tive immune responses to a particular pathogen isnot straightforward; indeed it is only recently thatthe full sophistication of the immune response hasbecome apparent.4–6 There are also significant tech-nical difficulties in measuring other (non-antibody)aspects of immunity. Assays of the cell-mediatedimmune response, for example cytotoxic T lympho-cyte (CTL) activity, remain technically demanding,are not suitable for batch screening, are not easilyreproducible between laboratories, and are not
available commercially.7–9 In contrast, antibodymeasurement is relatively simple, is highly suitablefor batch screening, is acceptably reproducible be-tween laboratories, and is available commercially.There can be little wonder, then, that serum anti-body titers have been the standard means of assess-ing vaccine efficacy thus far.
However, practicing veterinarians and vaccinolo-gists alike are well aware of the limitations of cur-rent vaccines for several of the major equinepathogens, despite the ability of these vaccines toinduce efficient serum antibody responses. The aimof this paper is to explore the reasons why vaccinesmay fail to control viral and bacterial pathogens andthus induce protective immunity. In order to under-stand the reasons for this we need to consider, foreach pathogen:
1. Its pathogenesis2. The key stages in pathogenesis at which the
immune system may intervene3. The types of immune responses required to
effect control at those key stages4. The type of vaccine that would be likely to
produce such immune responses
Mucosal diseases, especially respiratory diseases,present particular problems for vaccinologists be-
10 2000 / Vol. 46 / AAEP PROCEEDINGS
IN DEPTH: VACCINATION
NOTES
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000
cause the prime requirement for protection is sur-face mucosal immunity rather than circulatingantibody.
Certain respiratory pathogens, for exampleEHV-1 and S. equi, are additionally complex be-cause their pathogenesis involves invasion throughthe respiratory epithelium with potential for sys-temic infection.9,10 Further difficulties are presentedby other respiratory pathogens, for example R. equi,which establishes intracellular infection in macro-phages and is thus shielded from effective immunesurveillance.11
We also need to remember that infection is adynamic process by both host and pathogen. Manypathogens employ sophisticated mechanisms toimpair or circumvent the host’s immune response,processes referred to as immunosuppression. Sev-eral immunosuppressive mechanisms have beenidentified for EHV-1 and S. equi and overcomingthese mechanisms may represent the ultimategoal for vaccine development.12–14
To allow us to understand the reasons why vac-cines may fail to control some equine infections, wewill consider the difficulties that two common respi-ratory pathogens, EHV-1 and S. equi, present. Thefirst step is to understand the pathogenesis of theseinfections.
Pathogenesis of EHV-1 infections
Acquisition of Infection and Colonization of RespiratoryEpitheliumEHV-1 infection is acquired by inhalation of aerosolsor by contact with fomites. The virus attaches to,
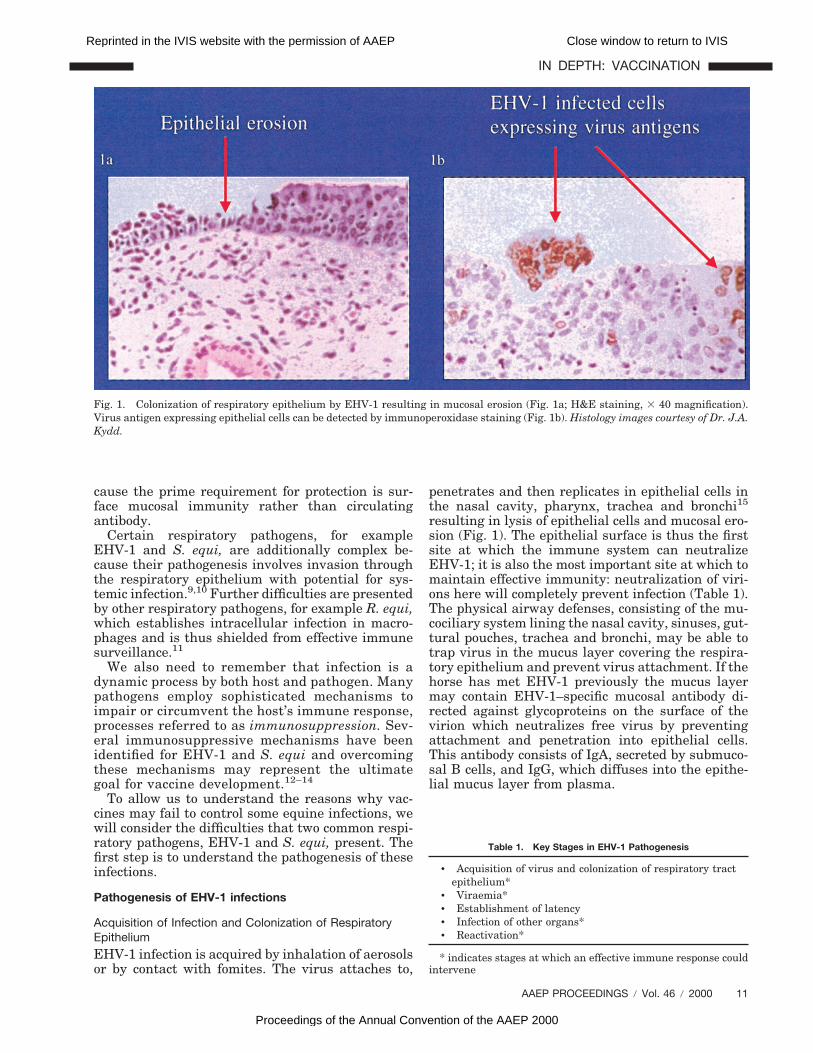
penetrates and then replicates in epithelial cells inthe nasal cavity, pharynx, trachea and bronchi15
resulting in lysis of epithelial cells and mucosal ero-sion (Fig. 1). The epithelial surface is thus the firstsite at which the immune system can neutralizeEHV-1; it is also the most important site at which tomaintain effective immunity: neutralization of viri-ons here will completely prevent infection (Table 1).The physical airway defenses, consisting of the mu-cociliary system lining the nasal cavity, sinuses, gut-tural pouches, trachea and bronchi, may be able totrap virus in the mucus layer covering the respira-tory epithelium and prevent virus attachment. If thehorse has met EHV-1 previously the mucus layermay contain EHV-1–specific mucosal antibody di-rected against glycoproteins on the surface of thevirion which neutralizes free virus by preventingattachment and penetration into epithelial cells.This antibody consists of IgA, secreted by submuco-sal B cells, and IgG, which diffuses into the epithe-lial mucus layer from plasma.
Fig. 1. Colonization of respiratory epithelium by EHV-1 resulting in mucosal erosion (Fig. 1a; H&E staining, 3 40 magnification).Virus antigen expressing epithelial cells can be detected by immunoperoxidase staining (Fig. 1b). Histology images courtesy of Dr. J.A.Kydd.
Table 1. Key Stages in EHV-1 Pathogenesis
• Acquisition of virus and colonization of respiratory tractepithelium*
• Viraemia*• Establishment of latency• Infection of other organs*• Reactivation*
* indicates stages at which an effective immune response couldintervene
AAEP PROCEEDINGS / Vol. 46 / 2000 11
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000

If a virus successfully infects respiratory epithe-lial cells the outcome of infection is determined bythe horse’s immune response. We will concentrateon the adaptive immune response because this aloneis influenced by vaccination. The innate (non-speci-fic) immune response is not influenced by vaccina-tion and will therefore not be considered in detail inthis paper. However, innate responses (consisting ofcomplement; cytokines IL-1, IL-12, IFN-g andTNFa, macrophages, and natural killer cells) arenonetheless extremely important in the initial limi-tation of EHV-1 infection before the adaptive (Band T lymphocyte) immune responses become func-tional.16 We should also remember that cytokinesare also important in the adaptive immune responseand are produced by TH lymphocytes to promoteeither cytotoxic (TH1) or antibody (TH2) responses.The critical role of IL-4, -5, and -6 (produced by TH2)cells in B lymphocyte stimulation is being exploitedin DNA vaccination and IL-6, for example, has beenincluded in experimental EIV DNA vaccines.
Following replication in respiratory epithelium,EHV-1 rapidly breaches the basement membraneto reach the lamina propria. Entry to the laminapropria provides the virus with opportunity for sys-
temic dissemination via the lymphatic and circula-tory systems (Fig. 2). The lamina propria is thesecond site at which the immune system has anopportunity to control infection. Serum neutralizingantibody probably has only a limited protective rolein the lamina propria because it is restricted toneutralizing free (extracellular) virions before theyhave the opportunity to infect cells in the laminapropria. At this point it is important to rememberthat the role of antibody in the removal of EHV-1–infected cells is restricted to opsonization (aidingphagocytosis) and antibody dependent cell mediatedcytotoxicity of infected cells. The protective role ofantibody is therefore likely to be most effective atthe epithelial surface. Once the virus has estab-lished intracellular infection, removal of virus-in-fected cells is achieved by cell-mediated immuneresponses. Virus-infected cells express virus anti-gens via MHC I molecules on their surface whichstimulate TH1 cells and in turn CD81 CTLs totarget and lyse the infected cell. If the immune sys-tem is unable to neutralize EHV-1 in the laminapropria, it infects trafficking CD51/CD81 T lym-phocytes and also endothelial cells, events that ulti-mately result in the development of a T lymphocyte-
Fig. 2. Epithelial invasion by EHV-1 and development of viraemia. Following infection of epithelial cells (Fig. 2a), EHV-1 infectsendothelial cells in the lamina propria (Fig. 2b). Virus-infected mononuclear cells (mainly CD51/CD81) T lymphocytes subsequentlyappear in drainage lymph nodes (Fig. 2c) and are released into the circulation producing viraemia (virus antigens in histology sectionsdemonstrated by immunoperoxidase staining; magnification 3 40). Histology images courtesy of Dr. J.A. Kydd; cartoon courtesy of Prof.G.P. Allen.
12 2000 / Vol. 46 / AAEP PROCEEDINGS
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000

associated viraemia. These are, of course, preciselythe lymphocytes with responsibility for eliminationof virus-infected cells and interference with CTLresponses are one of the immunosuppressive strat-egies employed by EHV-1.
Viraemia
Viraemia is responsible for dissemination of virusthroughout the body and the highly important se-quelae of abortion, neurological disease, and chori-oretinopathy. Prevention of viraemia is thereforethe single most important measure of vaccine effi-cacy. EHV-1 viraemia is strictly cell-associated andthe infected cells are mononuclear leucocytes, prin-cipally CD51/CD81 T lymphocytes. Immunocyto-chemistry studies have shown that T lymphocytesinitially become infected in the lamina propria ofrespiratory epithelium and reach respiratory tractdrainage lymph nodes within 48 h of infection (Fig.2). Release of infected lymphocytes into the circula-tion via efferent lymph from drainage lymph nodesresults in a T lymphocyte-associated viraemia thatis detectable from as early as day 3 post infection16
and may persist up to day 21 post infection, al-though in adult horses, including pregnant mares,the duration of viraemia is usually less than 7 days.Lamina propria vascular endothelium also becomesinfected and it is possible that infected endothelialcells can also contribute to viraemia via infection ofcirculating T lymphocytes by direct contact.
The lymph node and circulation are thus the thirdand fourth stages at which the immune system couldeliminate EHV-1 infection. However, controlling vi-raemia presents major problems for the immunesystem because 1) antibody is not effective againstintracellular virus, and 2) antigen expression oninfected T lymphocytes is transient, limiting theopportunity for generation of CTLs to target virus-infected lymphocytes. However, viraemic lympho-cytes are only briefly targets for CTLs because theexpression of viral glycoproteins on the lymphocytesurface is transient and after the early phase ofviraemia. EHV-1 persists in T lymphocytes in a la-tent, transcriptionally-silent form with no virus an-tigen expression.9 Controlling viraemia clearlypresents major problems for vaccine design. Killedvirus vaccines result in antigen presentation via theexogenous pathway on MHC class II molecules withthe induction of TH2 and antibody responses, notCTL responses.
Establishment of Latency
It seems likely that latency is established in almostall horses following infection with EHV-1. In theinitial stages of nasal and conjunctival epithelialinfection, EHV-1 gains access to neurons of the tri-geminal nerve and reaches the trigeminal ganglionby 48 h post infection where the virus persists in alatent form.18 If local mucosal immunity is unable toneutralize EHV-1 at the epithelial surface and virusgains access to the trigeminal nerve receptors in the
nasal epithelium, the immune system is unable toprevent the transport of virus to the trigeminal gan-glion with subsequent establishment of latency intrigeminal ganglionic neurones. As mentionedabove, EHV-1 also persists in a latent form in circu-lating CD51/CD81 T lymphocytes19 after the virae-mic phase of infection is terminated (Fig. 3).
There is no virus replication in latently infectedcells, whether these are neurons or lymphocytes,and the virus persists as genome only, presumablyin the form of circular DNA as has been described forother herpesviruses. Transcriptional activity of thelatent virus genome is restricted to the region anti-sense to the immediate early gene (ORF 64).20 Othergenes are not transcribed and there is no viral pro-tein translation during latency. This means thatlatently-infected cells do not express virus antigensand are thus antigenically silent, escaping detection,targeting and elimination by the immune system.The elimination of latent infections from the horsepopulation is not a realistic goal for vaccination,although as discussed below, effective vaccines maybe able to limit the consequences of reactivation oflatent virus.
Infection of Other Organs
Viraemia is a prerequisite for the sequelae of abor-tion, neurological disease, and chorioretinopathies,although viraemia does not automatically inducethem. Abortion is the result of ischaemia consequentto vasculitis following endothelial cell infection inuterine vasculature.21,22 This causes thrombo-isch-aemic necrosis of the overlying microcotyledons andintercotyledonary stroma (Fig. 4). If these endome-trial vascular lesions are widespread, the fetus maybe aborted before detectable transplacental spreadof virus has occurred.23 In other cases, disruption ofthe physical integrity of the uteroplacental barrierallows free virus or virus-infected cells to cross theplacenta into the fetal circulation with resultantfetal infection. The mechanisms of virus transferfrom infected circulating lymphocytes to vascularendothelium are unknown, although the virus anti-gens expressed on lymphocytes during the earlystages of viraemia are glycoproteins that mediatefusion with permissive cells, for example epithelialand endothelial cells, thus providing a potentialmeans of virus transfer.
Infection of endothelial cells, with subsequent vas-culitis and ischemia, in the central nervous system(CNS) also appears to be central to the pathogenesisof neurological disease.24 The precise mechanism ofvirus transfer to the CNS is not clear but, as in theuterus, virus antigen-expressing endothelial cellscan be demonstrated in the CNS by immunocyto-chemistry. The pathogenesis of EHV-1 induced cho-rioretinal lesions25 is less well characterized but alsoappears to involve endothelial cell infection withpresumed ischemic injury to the choroid and retina.
Although infected uterine and nervous system en-dothelial cells express EHV-1 antigens and are thus
AAEP PROCEEDINGS / Vol. 46 / 2000 13
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000

subject to detection by the immune system, it seemslikely that circulating antibody or CTLs would havelimited opportunity to eliminate infection and pre-vent ischemic sequelae developing.
ReactivationLatent virus (harbored in both circulating lympho-cytes and trigeminal ganglionic neurons) is knownto undergo periodic reactivation, for example in re-sponse to stressful management events (transport,handling, re-housing, and weaning) and experimen-tally by treatment with corticosteroids.26,27 It istherefore likely that modern, intensive managementpractices, especially in racing and competitionhorses, result in frequent reactivation of latentEHV-1 infections. Reactivation results in asymp-tomatic shedding of infectious virus from the respi-ratory tract which may result in infection of in-contact horses.
The process of reactivation of EHV-1 from neuronsand circulating T lymphocytes is unknown, but mustresult in delivery of virions to nasopharyngeal epi-thelium with subsequent virus replication and shed-ding in nasal secretions. In neurons, reactivatingvirus travels from the cell body to the peripheralepithelial receptor by axonal transport. In lympho-cytes, a lytic replication cycle is probably established
with of fusogenic viral glycoproteins on the lympho-cyte surface and transfer of virus to epithelial cells(Fig. 3).9
The outcome of reactivation after this point isalmost certainly determined by local mucosal immu-nity, consisting of both neutralizing antibody andCTLs. If antibody is present on the epithelial surfaceand antibody and CTLs are present in the laminapropria, reactivating virus is likely to be neutral-ized: nasal shedding of virus does not occur andreactivation is not completed. On the other hand, ifmucosal immunity has not been primed (by previousinfection or vaccination) or is impaired (by stress orcorticosteroid treatment), reactivating virus willsuccessfully infect epithelial cells and establish am-plifying, lytic infection in these cells, thus resultingin shedding of infectious virus in nasal secretions. Inthese circumstances reactivation is completed. Suc-cessful completion of reactivation can also result indevelopment of cell-associated viraemia by repeat-ing the events following initial respiratory tract in-fection. This could result in dissemination ofinfectious virus, once again, to the uterus to produceabortion or, potentially, to the CNS to cause neuro-logical disease. Another source of reactivated EHV-1for causing abortion is from resident lymphocytes
Fig. 3. Establishment of latency by EHV-1. Following infection of respiratory epithelium latent infections are established incirculating T lymphocytes and trigeminal ganglionic neurones (white arrows). Reactivation results in shedding of virus from nasalepithelium and probably uterine infection (yellow arrows). The histology section shows trigeminal ganglionic neurones infected withEHV-1, expressing virus reporter gene product (beta galactosidase) in the cell body and reporter gene DNA in the nucleus (magnifi-cation 3 40).
14 2000 / Vol. 46 / AAEP PROCEEDINGS
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000

within the local environment of the pregnant endo-metrium which may transfer infectious virus di-rectly to the uterine endothelium, thereby initiatingthe cascade of events leading to abortion. Therefore,EHV-1 abortion may occur without the prerequisiteof a lytic respiratory epithelial infection or a detect-able cell-associated viraemia. This may explainabortion in single mares within a group28 and alsoabortions that occur many weeks or months aftertermination of the cell-associated viraemia.29,30
As discussed above, vaccination will not eliminatepre-existing latent EHV-1 infections. However, bystimulating and, most importantly, maintaining ef-fective mucosal immunity, vaccines are likely tolimit the frequency of nasal shedding of infectiousEHV-1 following reactivation and thus limit the im-pact of latency in the epidemiology of this virus.
Pathogenesis of Streptococcus equi Infections
Although the clinical disease and associated grosspathology caused by S. equi infections have beenthoroughly documented, in comparison to EHV-1,the detailed events in pathogenesis are less wellcharacterized. However, it is likely that the se-quence of events in pathogenesis involves the follow-ing stages (Table 2).
Acquisition of Bacteria and Colonization of RespiratoryEpitheliumS. equi is acquired directly by inhalation of ingestionof fomites from infected horses and in some cases
indirectly via fomites in feed and water troughs. Incontrast to other Group C streptococci found inhorses, e.g., S. zooepidemicus, S. equi is not a com-mensal of the equine respiratory tract31 and its pres-ence appears to be associated with either acute orchronic disease syndromes. In view of the highlycontagious nature of the disease it is likely thatacquisition is generally followed by successful colo-nization of the upper respiratory tract, particularlythe nasopharynx and guttural pouches, with subse-quent induction of pathology (Fig. 5).32
The physical barrier afforded by the mucociliaryapparatus and the population of the nasopharynx bycommensal bacteria provides an initial obstacle tocolonization. Mucosal antibody (mainly IgA but alsoIgG) in nasopharyngeal secretions aids in prevent-ing colonization by binding to bacterial surface pro-teins (adhesins) required for attachment to epi-thelial cells and can thus provide a measure of
Fig. 4. Infection of other organs by EHV-1. Abortion results from ischemia following virus infection (demonstrated by immunoper-oxidase staining) of uterine endothelium causing vasculitis and thrombosis (magnification 3 40). Histology images courtesy of Dr. K.C.Smith.
Table 2. Key Stages in S. equi Pathogenesis
• Acquisition of bacteria and colonization of respiratory tractepithelium*
• Invasion of the lamina propria*• Evasion of phagocytosis*• Spread to drainage lymph nodes and the circulation*• Abscess formation in lymph nodes and other organs*
* indicates stages at which an effective immune response couldintervene
AAEP PROCEEDINGS / Vol. 46 / 2000 15
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000

protection against disease.33 As for EHV-1, the epi-thelium is the first site at which the immune re-sponse can eliminate S. equi infection.
If the immune system is unable to prevent adhe-sion of bacteria to respiratory epithelium, coloniza-tion probably progresses as follows. Adhesion torespiratory epithelium is followed by bacterial rep-lication and production of a range of secreted toxins(for example hemolysin) which assist epithelial col-onization by causing cytopathology in ciliated epithe-lium and impairing ciliary clearance of bacteria.34
Invasion of the Lamina PropriaFollowing replication on the epithelium, bacteria in-vade the deeper epithelial layers and breach thebasement membrane to penetrate the lamina pro-pria. If bacteria are not eliminated in the laminapropria by the host immune response, they may gainaccess to lymphatics and reach drainage lymphnodes.
The lamina propria is thus the second site atwhich the immune system can eliminate bacteria.The immune response to S. equi infections differsfrom the immune response to EHV-1. Although ini-tial attachment of bacteria to the epithelium may beprevented by antibody, the elimination of bacteria isachieved primarily by innate immune responses as-sisted by adaptive immune responses, mainly anti-body. The key cells in elimination of bacteria are
phagocytes (neutrophils and macrophages) assistedby B lymphocytes, not CTLs. S. equi also activatesthe alternative complement pathway which resultsin opsonization of bacteria via the binding of C3b tothe bacterial surface thus facilitating phagocytosis(the process of binding of antibody or complement tobacteria resulting in enhanced phagocytosis is calledopsonization). The binding of C3b to bacteria canalso result in complement-mediated bacterial lysisalthough this is probably inefficient in Gram posi-tive bacteria like S. equi.
Bacterial phagocytosis is greatly assisted by anti-body which opsonizes bacteria and increases theirsusceptibility to phagocytosis. Serum antibody andmucosal antibodies are directed principally at themajor bacterial antigen, the 56 kDa S. equi M-likeprotein SeM.35 The binding of antibody to S. equisurface antigens also activates the classic comple-ment pathway, opsonizing bacteria for phagocytosisand also promoting complement-mediated bacteriallysis. In comparison to EHV-1, however, CTL re-sponses play a minor role in bacterial elimination asS. equi is an extracellular pathogen.
Thus, in contrast to EHV-1 infections, the im-mune response to S. equi relies heavily on innateimmunity (phagocytes and complement) augmentedby adaptive immunity (antibody). Although vaccina-tion does not influence phagocytes (in contrast to
Fig. 5. S. equi pathogenesis. S. equi multiplies on respiratory epithelium causing epithelial pathology (Fig. 5a; H&E staining,magnification 3 40). Bacteria reach drainage lymph nodes and persist by evading phagocytosis resulting lymph node abscessation (Fig.5a; Gram staining, magnification 3 100).
16 2000 / Vol. 46 / AAEP PROCEEDINGS
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000

lymphocytes they are non-specific, do not have mem-ory and do not exhibit anamnestic responses), stim-ulation of B lymphocyte responses is extremelyimportant in recovery from infection and preventionof re-infection. The generation of mucosal antibody(from plasma cells in submucosal lymph follicles anddrainage lymph nodes) and, to a lesser extent, cir-culating antibody, provides protection from re-infec-tion by 1) binding to adhesins on the bacterialsurface and blocking bacterial attachment to epithe-lium, and 2) opsonizing bacteria on the epitheliumand in the lamina propria, thus greatly enhancingtheir removal by phagocytes. As discussed in thenext section, S. equi inhibits phagocytosis and effi-cient removal of virulent strains of S. equi abso-lutely requires opsonizing antibody.
Evasion of Phagocytosis
Large numbers of phagocytes migrate to sites of S.equi infection from the circulation. S. equi isstrongly chemotactic for neutrophils and the neutro-phil appears to be the principal cell responsible forbacterial clearance. Neutrophils are recruited inlarge numbers to respiratory tract epithelium anddrainage lymph nodes. Neutrophils have a short lifespan (less than 72 h) in tissues and therefore mi-grate continuously to sites of infection until infectionis eliminated. To achieve bacterial phagocytosis,neutrophils firstly adhere to S. equi and then inter-nalize the bacteria in a structure called the phago-some. Neutrophils possess surface Fc receptors andbacteria which have antibody bound to their surfaceand therefore adhere more efficiently to neutrophils.Complement (C3b) bound to the bacterial surfacealso assists adherence to neutrophils. Antibodies areparticularly effective opsonins and phagocytosis issignificantly enhanced in the presence of opsonizingantibody. The generation of opsonizing antibody istherefore a key requirement for S. equi vaccines.Once internalized in the phagosome, phagocytosedbacteria are killed by a variety of mechanisms whichcan be divided into two groups: oxidative and non-oxidative. The oxidative mechanisms kill bacteria bythe release of reactive oxygen intermediates (e.g.,superoxides) and reactive nitrogen intermediates(e.g., nitric oxide). The cytoplasm of neutrophils con-tains lysosomes which fuse with the phagosome andkill bacteria non-oxidatively by release of potentlyantibacterial hydrolytic enzymes (e.g., lysosyme)and peptides (e.g., azurocidin).
A key feature in pathogenesis, and one which is acentral problem for vaccines, is that S. equi pos-sesses mechanisms that allow it to resist phagocy-tosis36 and persist despite the migration of largenumbers of neutrophils to the infection site. Thebest characterized of these is SeM, mentioned ear-lier as the immunodominant antigen of S. equi. SeMhas potent antiphagocytic activity, allowing bacte-rial survival when incubated with neutrophils invitro. SeM exerts its antiphagocytic action by inhib-iting the deposition of complement activation prod-
ucts (C3b) on the bacterial surface by bindingfibrinogen.37 The hyaluronic acid capsule of S. equiis a further mechanism which confers resistance tophagocytosis. The importance of phagocytosis eva-sion for S. equi virulence has been demonstrated invitro and in field and experimental infections wherebacteria deficient in SeM or hyaluronic acid capsulehave reduced pathogenicity. S. equi also producessecreted protein toxins which inhibit neutrophilfunction. Thus far a peripheral blood mononuclearcell mitogen and a neutrophil cytotoxin38,39 havebeen identified.
Spread to Drainage Lymph Nodes and the CirculationThe ability of S. equi to resist phagocytosis allowsbacteria to gain entry to lymphatics in the laminapropria and thus reach respiratory tract drainagelymph nodes. Bacteria remain viable and multiplydespite the continuing mobilization of large num-bers of neutrophils into infected lymph nodes. Thepersistence of viable bacteria in the node stimulatescontinuing activation of the complement cascadewith continuing release of inflammatory mediatorsand chemotaxis of neutrophils into the node. Thisresults in formation of the lymph node abscessesseen in classical strangles. Clearance of bacteriafrom the node requires bacterial opsonization byantibody to facilitate efficient phagocytosis and bac-terial killing.
Infection of Other OrgansIf the immune response is unable to restrict S. equito respiratory tract drainage lymph nodes, bacteriamay be released into the lymphatic system and intothe circulation to produce a range of sequelae includ-ing metastatic abscesses (bastard strangles) in thethorax, abdomen, CNS, and myocardium.40
The immune control of bacterial spread to respi-ratory tract drainage lymph nodes and beyond relieson efficient opsonization of bacteria by antibody withsubsequent phagocytosis and elimination of bacteriaby neutrophils.
How Do We Make More Effective RespiratoryVaccines?
Having considered pathogenesis carefully, the nextstep is to identify the components of the immuneresponse that effective vaccines against EHV-1 andS. equi would need to generate in order to controlpathogenesis. For each of these respiratory patho-gens, as we follow their pathogenesis from initialinfection of the respiratory epithelium into the lam-ina propria and beyond into drainage lymph nodesinto the circulation, the opportunity for the immunesystem to control infection diminishes. The goldenopportunity for control of infection is at the epithe-lial surface. Stimulation of effective mucosal immu-nity is therefore the primary goal of vaccines againstrespiratory pathogens. With present technologiesthe most efficient means of inducing mucosal immu-nity is by local (intranasal) inoculation of vaccine
AAEP PROCEEDINGS / Vol. 46 / 2000 17
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000

into the respiratory tract. However, the vaccinemust induce the appropriate humoral and cellularimmune responses, which means that alternativesto inactivated vaccines (modified live, protein, orDNA vaccines) must be sought. Although modifiedlive vaccines stimulate efficient humoral and cellu-lar immune responses, safety is a major concern.One further consideration is that modified live vac-cines that retain immunosuppressive mechanismsof the wild-type pathogen may limit the immuneresponse generated. Protein or DNA vaccines mayprove safer alternatives to modified live vaccines,but their efficacy may be less than modified livevaccines.
However, no matter how efficiently immunogenicfuture vaccines become, vaccination alone will not beable to control these diseases. Effective diseasecontrol will always rely on a combination of goodmanagement and vaccination. The principles ofmanagemental control are similar for each of thesepathogens and are based on:
● Isolation and testing of new arrivals to ensurethey are not incubating disease or are carriers
● Segregation of high-risk animals from low-riskanimals, e.g., pregnant mares from youngstock
● Management of horses in small groups andavoidance of mixing groups reduction, wherepossible, of management stresses
● Isolation and investigation of clinical cases
In the United Kingdom, where S. equi vaccines arenot available and only a minority of horses are vac-cinated against EHV-1, these management princi-ples have been formalized into Codes of Practicewhich have reduced the incidence of EHV-1 and S.equi infections.
Limitations of Current EHV-1 VaccinesTen commercial EHV-1 vaccines have been devel-oped and used around the world, seven of which areinactivated whole virus vaccines and three whichare modified live vaccines. In adult horses, at least,these vaccines do stimulate high titers of serumantibody against EHV-1.41 However, the impact ofthese vaccines on the incidence of EHV-1 has beendisappointing, with EHV-1 remaining one of themost common equine pathogens. However, itwould be misleading to suggest that current vac-cines have been totally ineffective: the incidence ofabortion in Kentucky declined by 75% over a 20-year period after the introduction of intensive vac-cination program.42
When the required immune response to EHV-1 isconsidered, the shortcomings of currently availablevaccines become readily apparent. Systemically ad-ministered, inactivated virus vaccines stimulate ef-ficient circulating antibody responses but notmucosal antibody and not CTL responses. Systemi-cally administered modified live vaccines also stim-
ulate circulating antibody and also CTL responsesbut do not stimulate efficient mucosal immunity.
Requirements of an Effective EHV-1 VaccineAn effective EHV-1 vaccine should reduce the inci-dence or severity of disease, contain virus spreadduring outbreaks, and limit virus spread followingreactivation of latent infections by:
● Providing protection from initial respiratoryepithelial infection
● Preventing viraemia● Controlling reactivation of latent virus by pre-
venting replication of reactivating virus in re-spiratory epithelium
To achieve these goals EHV-1 vaccines must induceand maintain effective mucosal as well as systemicimmune responses. Mucosal immunity is most im-portant since disease will not progress if virus isneutralized at the epithelium. However, if we acceptthat vaccination is unlikely to provide more efficientimmunity than natural infection, it is important toremember that although natural infection does pro-duce a robust protective immunity, this is transientwith a duration of between 3–6 months only.
Convalescent horses with protective immunityagainst re-infection have the following immune re-sponses:
● Virus neutralizing antibody in both serum andnasopharyngeal mucus
● Viral antigen driven, CD41 T-lymphocyte pro-liferative activity
● Virus-specific, CD81 cytotoxic T-lymphocyteprecursors (CTLp)
● Herpesvirus reactive natural killer (NK) lym-phocyte activity
● Virus specific antibody-dependent, cell-medi-ated cytotoxic (ADCC) activity
These provide useful guidance for desirable vac-cine-generated immune responses (Table 3). Thebalance between the protective effects of humoraland cellular immunity is not clear, however, sinceno correlation with current assays has been found
Table 3. Immune Requirements for EHV-1 Vaccines
Site Immune Response
Respiratory epithelial surface1. Mucosal neutralizing antibodyEpithelial lamina propria 1. Systemic neutralizing
antibody2. Cytotoxic T lymphocytes
Lymph and lymph nodes 1. Systemic neutralizingantibody2. Cytotoxic T lymphocytes
Circulation 1. Systemic neutralizingantibody2. Cytotoxic T lymphocytes
18 2000 / Vol. 46 / AAEP PROCEEDINGS
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000

between serological or cellular responses and pro-tective immunity.
Nevertheless, it is reasonable to assume that avaccine must generate and maintain mucosal anti-body, systemic antibody, and CTLs.
Future Prospects for EHV-1 Vaccines
The envelope of EHV-1 contains at least 11 glyco-proteins which are the targets of the horse’s hu-moral, and presumably cellular, immune responses.Glycoproteins B, C, and D are the immunodominantantigens responsible for generating virus-neutraliz-ing antibodies and are obvious targets for study asvaccine candidates.43,44 Vaccine technologies thatcould utilize glycoproteins B, C, and D include re-combinant vector vaccines expressing these glycop-roteins, protein vaccines consisting of purifiedglycoproteins, and DNA vaccines comprising plas-mids that contain the genes for these glycoproteins.Each of these applications has received researchinterest, but none of the experimental approachestaken has, as yet, resulted in a commercial vaccine.Problems have included levels of protein expressionin vector systems, delivery methods and prepara-tions for protein vaccines, and a lack of informationabout the immunological interplay between the gly-coproteins and other components of the immune re-sponse.
Five EHV-1 glycoproteins (gB, gD, gH, gL, andgK) are essential for replication of the virus whilethe other six (gC, gE, gG, gI, gM, and gp300) are notrequired for viral growth in cell culture. The deletionof non-essential glycoprotein genes decreases thevirulence of EHV-1, making these genes potentialcandidates for modified live vaccines. The virus con-tains five unique genes (ORFs 1, 2, 67, 71, and 75)that have no structural homologues in any of theherpesviruses sequenced to date.45 These have alsoreceived research interest as candidates for deletionto generate modified live vaccines.
DNA vaccination may provide an alternative, ef-fective strategy for EHV-1 and research is currentlyin progress to identify suitable genes to incorporateinto vaccines.
While EHV-1 places stringent demands on vac-cines, the virus does not appear to exhibit significantantigenic variation suggesting that the virus is, incontrast to EIV, antigenically stable and relativelyhomogeneous. Having reviewed the difficulties thisvirus presents, this final piece of information offersat least a crumb of comfort: an effective EHV-1 vac-cine is likely to be effective against almost all strainsof the virus and is likely to remain effective.
Requirements for an Effective S. equi Vaccine
An effective S. equi vaccine should reduce the inci-dence or severity of disease, reduce bacterial spreadduring outbreaks, and prevent the establishment ofpersistent infections and carrier states by:
● Stimulating respiratory mucosal neutralizingIgA antibody to block attachment of bacteria toepithelial cells
● Stimulating systemic opsonizing IgG antibodyto facilitate phagocytosis
● Generating a population of long-lived memoryB cells in submucosal lymph follicles anddrainage lymph nodes to produce further IgAand IgG when bacteria are subsequently en-countered.
These requirements are summarized in Table 4. Asdiscussed above, systemic vaccination is unlikely toproduce effective immunity against S. equi becauseit promotes circulating, but not mucosal, immuneresponses. For example, the immunodominant S.equi antigen is SeM, making it an attractive vaccinetarget. However, the serum antibody responses gen-erated by systemically-administered SeM proteinvaccines are not protective.
Intranasal vaccination is more likely to inducelocal protective immunity. Intranasally adminis-tered, modified-live bacterial vaccines induce effi-cient local immunity but, as for EHV-1, safety is anissue. However, a commercial modified live S. equi(Pinnacle, Fort Dodge Animal Health) has been suc-cessfully launched in the United States, but is notavailable in Europe. Intranasal protein or DNA vac-cines are potential future alternatives, but thesemust induce activation of the TH2 subset of CD41 Tlymphocytes in order to promote B cell activation.
References1. Mumford JA, Wood JM, Scott AM, et al. Studies with inacti-
vated equine influenza vaccine: 2. Protection against experi-mental infection with influenza virus A/equine/Newmarket/79(H3N8). J Hygiene 1983;90:385–395.
2. Hoffman AM, Staempfli HR, Prescott JF, et al. Field evalua-tion of a commercial M-protein vaccine against Streptococcusequi infection in foals. Am J Vet Res 1991;52:589–592.
3. Ostlund EN. The equine herpesviruses. Vet Clin North Am[Equine Pract] 1993;9:283–294.
4. Zinkernakel RM, Doherty PC. The discovery of MHC restric-tion. Immunol Today 1997;18:14.
5. Doherty PC, Topham DJ, Tripp RA, et al. Effector CD41 andCD81 mechanisms in the control of respiratory virus infec-tions. Immunol Rev 1997;159:105.
6. Doherty PC. Cytotoxic T cell effector and memory function inviral immunity. Curr Top Microbiol Immunol 1996;206:1.
7. O’Neill T, Kydd JH, Allen GP, et al. Determination of equidherpesvirus 1-specific, CD81, cytotoxic T lymphocyte precur-sor frequencies in ponies. Vet Immunol Immunopathol 1999;70(1-2):43–54.
8. Allen GP, Yeargan MR, Costa LR, et al. Major histocompat-ibility complex class I-restricted cytotoxic T-lymphocyte re-
Table 4. Immune Requirements for S. equi Vaccines
Site Immune Response
Respiratory epithelial surface mucosal neutralizing antibodyEpithelial lamina propria systemic opsonizing antibodyLymph and lymph nodes systemic opsonizing antibodyCirculation systemic opsonizing antibody
AAEP PROCEEDINGS / Vol. 46 / 2000 19
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000

sponses in horses infected with equine herpesvirus 1. J Virol1995;69:606–612.
9. Allen GP, Kydd JH, Slater JD, et al. Recent advances inunderstanding the pathogenesis, epidemiology and immunol-ogy of equine herpesvirus-1 abortion, in Proceedings. 8thInternatl Conf Equine Infect Dis, 1998;129–146.
10. Sweeney CR. Strangles: Streptococcus equi infection inhorses. Equine Vet Ed 1996;8:317–322.
11. Yelle MT. Clinical aspects of Streptococcus equi infection.Equine Vet J 1987;19:158–162.
12. Charan S, Palmer K, Chesters P, et al. Transforming growthfactor-beta induced by live or ultraviolet-inactivated equidherpes virus type-1 mediates immunosuppression in thehorse. Immunology 1997;90:586.
13. Hannant D, O’Neil T, Jessett DM, et al. Evidence for non-specific immunosuppression during development of immuneresponses to equid herpesvirus-1. Equine Vet J 1991;(Supp12):10.
14. Boschwitz JS, Timoney JF. Inhibition of C3 deposition onStreptococcus equi subsp. equi by M protein: a mechanism forsurvival in equine blood. Infect Immun 1994;62:3515–3520.
15. Kydd JH, Smith KC, Hannant D, et al. Distribution of equidherpesvirus-1 (EHV-1) in the respiratory tract of ponies: im-plications for vaccination strategies. Equine Vet J 1994;26:466–469.
16. Slater JD, Hannant DA. Immunity to virus infections. VetClin North Am [Equine Pract] 2000;16:49–68.
17. Kydd JH, Smith KC, Hannant D, et al. Distribution of equidherpesvirus-1 in the respiratory tract-associated lymphoidtissue: implications for cellular immunity. Equine Vet J 1994;26:470–473.
18. Slater JD, Borchers K, Thackray AM, et al. The trigeminalganglion is a location for equine herpesvirus 1 latency andreactivation in the horse. J Gen Virol 1994;75:2007–2016.
19. Smith DJ, Iqbal J, Purewal A, et al. In vitro reactivation oflatent equid herpesvirus-1 from CD51/CD81 leukocytes in-directly by IL-2 or chorionic gonadotropin. J Gen Virol 1998;79:2997–3004.
20. Chesters PM, Allsop R, Purewal A, et al. Detection of latency-associated transcripts of equid herpesvirus 1 in equine leu-kocytes but not in trigeminal ganglia. J Virol 1997;71:3437–3443.
21. Smith KC, Mumford JA, Lakhani K. A comparison of equidherpesvirus-1 (EHV-1) vascular lesions in the early versuslate pregnant equine uterus. J Comp Path 1996;114:231–247.
22. Smith KC, Whitwell KE, Mumford JA, et al. An immunohis-tological study of the uterus of mares following experimentalinfection by equid herpesvirus 1. Equine Vet J 1993;25:36–40.
23. Smith KC, Whitwell KE, Binns MM, et al. Abortion of viro-logically negative foetuses following experimental challengeof pregnant pony mares with equid herpesvirus 1 [see com-ments]. Equine Vet J 1992;24:256–259.
24. Edington N, Bridges CG, Patel JR. Endothelial cell infectionand thrombosis in paralysis caused by equid herpesvirus-1:equine stroke. Arch Virol 1986;90:111–124.
25. Slater JD, Gibson JS, Barnett KC, et al. Chorioretinopathyassociated with neuropathology following infection withequine herpesvirus-1. Vet Rec 1992;131:237–239.
26. Burrows R, Goodridge D. Studies of persistent and latentequid herpesvirus-1 and herpesvirus-3 infections in the Pir-bright pony herd. In: Wittmann G, Gaskell RM, Rziha H-J,
eds. Latent herpesvirus infections in veterinary medicine. TheHague: Martinus Nijhoff, 1984;307–319.
27. Edington N, Bridges CG, Huckle A. Experimental reactiva-tion of equid herpesvirus 1 (EHV-1) following the adminis-tration of corticosteroids. Equine Vet J 1985;17:369–372.
28. Mumford JA. The epidemiology of equid herpesvirus abor-tion: a tantalizing mystery [editorial; comment]. Equine Vet J1991;23:77–78.
29. Mumford JA, Hannant D, Jessett DM, et al. Abortigenic andneurological disease caused by experimental infection withequid herpesvirus-1. in Proceedings. 7th Internatl ConfEquine Infect Dis 1994;261–275.
30. Gleeson LJ, Coggins L. Response of pregnant mares to equineherpesvirus 1 (EHV1). Cornell Vet 1980;70:391–400.
31. Woolcock JB. Epidemiology of equine streptococci. Res Vet Sci1975;18:113–114.
32. Timoney JF. Strangles. Vet Clin N Am [Equine Pract] 1993;9(2):365–74.
33. Galan JE, Timoney JF. Mucosal nasopharyngeal immuneresponses of horses to protein antigens of Streptococcus equi.Infect Immun 1985;47:623–628.
34. Flanagan J, Collin N, Timoney JF, et al. Characterisation ofthe haemolytic activity of Streptococcus equi. Microb Pathog1998;24:211–221.
35. Timoney JF, Artiushin SC, Boschwitz JS. Comparison of thesequences and functions of Streptococcus equi M-like pro-teins SeM and SzPSe. Infect Immun 1997;65:3600–3605.
36. Anzai T, Timoney JF, Kuwamoto Y. In vivo pathogenicity andresistance to phagocytosis of Streptococcus equi strains withdifferent levels of capsule expression. Vet Micro 1999;67:277–286.
37. Boschwitz JS, Timoney JF. Characterization of the antiph-agocytic activity of equine fibrinogen for Streptococcus equisubsp. equi. Microb Pathog 1994;17:121–129.
38. Anzai T, Sheoran AS, Kuwamoto Y, et al. Streptococcus equibut not Streptococcus zooepidemicus produces potent mito-genic responses from equine peripheral blood mononuclearcells. Vet Immunol Immunopathol 1999;61:1–12.
39. Muhktar MM, Timoney JF. Chemotactic response of equinepolymorphonuclear leucocytes to Streptococcus equi. Res VetSci 1988;45:225–229.
40. Sweeney CR, Whitlock RH, Meirs DA, et al. Complicationsassociated with Streptococcus equi infection on a horse farm.J Am Vet Med Assoc 1987;191:1446–1448.
41. Ellis JA, Bogdan JR, Kanara EW. Cellular and antibodyresponses to equine herpesviruses 1 and 4 following vaccina-tion of horses with modified-live and inactivated viruses [seecomments]. J Am Vet Med Assoc 1995;206:823–832.
42. Ostlund EN. The equine herpesviruses. Vet Clin N Am[Equine Pract] 1993;9:283–294.
43. Bridges CG, Edington N. The proteins of equid herpesvirus 1(EHV 1) recognised by equine antisera and their ability topromote antibody-dependent cell-mediated cytotoxicity. Tier-arztl Prax 1987;(Suppl 2):47–49.
44. Bridges CG, Ledger N, Edington N. The characterization ofequine herpes virus-1-infected cell polypeptides recognizedby equine lymphocytes. Immunology 1988;63:193–198.
45. Telford EA, Watson MS, McBride K, et al. The DNA sequenceof equine herpesvirus-1. Virology 1992;189:304–316.
46. Lunn DP. Immunological basis of equine vaccination, in Pro-ceedings, 46th Annu Conv Am Assoc Equine Practnr 2000,1–9.
20 2000 / Vol. 46 / AAEP PROCEEDINGS
IN DEPTH: VACCINATION
Reprinted in the IVIS website with the permission of AAEP Close window to return to IVIS
Proceedings of the Annual Convention of the AAEP 2000