impact loading and locomotor-respiratory …carrier.biology.utah.edu/dave's pdf/daley human...
TRANSCRIPT

Impact Loading and Locomotor-RespiratoryCoordination Significantly Influence Breathing Dynamicsin Running HumansMonica A. Daley1*, Dennis M. Bramble2, David R. Carrier2
1Department of Comparative Biomedical Sciences, Royal Veterinary College, Hatfield, Hertfordshire, United Kingdom, 2Department of Biology, University of Utah, Salt
Lake City, Utah, United States of America
Abstract
Locomotor-respiratory coupling (LRC), phase-locking between breathing and stepping rhythms, occurs in many vertebrates.When quadrupedal mammals gallop, 1:1 stride per breath coupling is necessitated by pronounced mechanical interactionsbetween locomotion and ventilation. Humans show more flexibility in breathing patterns during locomotion, using LRCratios of 2:1, 2.5:1, 3:1, or 4:1 and sometimes no coupling. Previous studies provide conflicting evidence on the mechanicalsignificance of LRC in running humans. Some studies suggest LRC improves breathing efficiency, but others suggest LRC ismechanically insignificant because ‘step-driven flows’ (ventilatory flows attributable to step-induced forces) contribute anegligible fraction of tidal volume. Yet, although step-driven flows are brief, they cause large fluctuations in ventilatory flow.Here we test the hypothesis that running humans use LRC to minimize antagonistic effects of step-driven flows onbreathing. We measured locomotor-ventilatory dynamics in 14 subjects running at a self-selected speed (2.660.1 ms21) andcompared breathing dynamics in their naturally ‘preferred’ and ‘avoided’ entrainment patterns. Step-driven flows occurredat 1-2X step frequency with peak magnitudes of 0.9760.45 Ls21 (mean 6S.D). Step-driven flows varied depending onventilatory state (high versus low lung volume), suggesting state-dependent changes in compliance and damping ofthoraco-abdominal tissues. Subjects naturally preferred LRC patterns that minimized antagonistic interactions and alignedventilatory transitions with assistive phases of the step. Ventilatory transitions initiated in ‘preferred’ phases within the stepcycle occurred 2x faster than those in ‘avoided’ phases. We hypothesize that humans coordinate breathing and locomotionto minimize antagonistic loading of respiratory muscles, reduce work of breathing and minimize rate of fatigue. Future workcould address the potential consequences of locomotor-ventilatory interactions for elite endurance athletes and individualswho are overweight or obese, populations in which respiratory muscle fatigue can be limiting.
Citation: Daley MA, Bramble DM, Carrier DR (2013) Impact Loading and Locomotor-Respiratory Coordination Significantly Influence Breathing Dynamics inRunning Humans. PLoS ONE 8(8): e70752. doi:10.1371/journal.pone.0070752
Editor: Francois Hug, The University of Queensland, Australia
Received March 4, 2013; Accepted June 28, 2013; Published August 12, 2013
Copyright: � 2013 Daley et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: The authors have no support or funding to report.
Competing Interests: The authors declare that co-author David Carrier is a PLOS ONE Editorial Board member. This does not alter the authors’ adherence to allthe PLOS ONE policies on sharing data and materials.
* E-mail: [email protected]
Introduction
Effective ventilation is essential for sustained animal locomotion.
For many animals, including birds and mammals, this requires
integrating movement and breathing so that inspiration and
expiration occur during mechanically compatible periods of the
locomotor cycle. Several direct mechanical links between locomo-
tion and ventilation necessitate integration [1,2,3,4]. Firstly,
sagittal bending of the trunk assists forward progression during
locomotion in quadrupeds, and creates a ‘bellows’ effect, altering
the pressure and volume of the abdomen and thorax. Additionally,
impact loads induce inertial motions of soft-tissues (viscera,
adipose), creating a ‘visceral piston’ effect, pulling and pushing
on the diaphragm and body wall muscles (abdominals, intercostals)
and altering thoraco-abdominal pressures. Finally, many axial
muscles of terrestrial vertebrates contribute to both breathing and
locomotion [5,6,7,8,9,10]. Indeed, many ‘ventilatory muscles’
cease respiratory action and become entrained with the locomotor
cycle during running in lizards [5,6], birds [7] and dogs [8,9]. As a
result of these factors, active inspiration is most compatible with a
specific and different phase of the locomotor cycle than active
expiration.
Locomotor-respiratory coupling (LRC) refers to phase locking
of running and breathing so that the same number of steps occur
during each breath, and has been observed in numerous
vertebrates, including birds, dogs, hares, horses, wallabies and
humans [2,3,11,12,13]. LRC is a form of entrainment, in which
the two rhythmic activities with different frequencies become
phase-locked due to mechanical and neural interactions. LRC has
been suggested to have a number of important physiological
effects. These include reducing the energy cost of breathing
[14,15,16,17], minimizing conflict in muscles that contribute to
both functions [6,9], body stabilization during motion [11,18], and
enabling trunk bending and inertial movements of soft-tissues to
augment pumping of air in and out of the lungs [2,19].
Whereas most galloping mammals exhibit 1:1 (strides/breath)
coupling, humans demonstrate more flexibility in breathing
patterns during locomotion. Humans frequently use LRC ratios
of 2:1, 2.5:1, 3:1, or 4:1 and sometimes lack entrainment
altogether, using independent breathing and stepping frequencies
PLOS ONE | www.plosone.org 1 August 2013 | Volume 8 | Issue 8 | e70752

[2,20,21,22,23]. Variance in LRC pattern is observed both within
and across individuals [14,21,22,23,24]. This flexibility in human
breathing patterns relative to locomotion most likely relates to an
upright posture and striding bipedal gait. Unlike quadrupeds, the
sagittal bending of the back does not assist forward progression
and the forelimbs are not subjected to direct weight-bearing and
impact loading. The forces transmitted through the thorax,
abdomen and rib cage are consequently smaller, reducing
mechanical interactions between locomotion and ventilation.
Ground birds, which are also bipedal, exhibit similar flexibility
as humans in locomotor-ventilatory coupling patterns [3,17].
Bipedal posture may reduce the mechanical and neuromuscular
conflicts that constrain quadrupeds to a 1:1 LRC pattern [8,9].
Does this greater flexibility suggest that breathing and running
are mechanically independent in humans? The answer remains
controversial. Humans do often exhibit sustained ventilatory
entrainment to locomotor forces [2,21,22]. Furthermore, evidence
suggests coupling improves breathing efficiency in some circum-
stances [2,15,24]. On the other hand, humans also couple
breathing frequency with low impact, non-locomotor movements
such as finger and arm tracking [25,26]. Thus, large mechanical
interactions are not required to induce rhythmic coupling.
Theoretically, human LRC during running could arise entirely
from neural interactions.
The overall ventilatory effect of locomotor-ventilatory interac-
tions likely depends on a number of neural and mechanical factors.
The primary mechanical factors in mammalian locomotor-
ventilatory interactions are thoracic loading and inertial displace-
ment of the soft-tissues associated with body accelerations
[2,12,19,20,27,28]. In humans, impact loading of the body at
footstrike is likely to cause compression of the thorax and
downward displacement of the abdominal viscera. Arm swinging
is also likely to exert a compression load on the thorax during early
to mid-stance [29]. These mechanical actions can alter respiratory
flow by changing thoraco-abdominal volumes and pressures.
Inertial displacement of the abdominal viscera would tend to pull
or push on the respiratory diaphragm through its direct
attachment to the liver. The relative magnitude of the effects on
the rib cage and diaphragm likely depends on the mass of the
abdominal organs, the distribution of other soft-tissues, and the
vertical accelerations of the body. Body wall muscles including the
intercostals and abdominals could act to resist these effects [30,31].
Additionally, the rhythmic activity of intercostal and abdominal
muscles is entrained by afferents from the arms and legs,
facilitating neural coupling [32,33]. The ventilatory outcome of
these neuro-mechanical interactions likely depends on the relative
timing and magnitude of the mechanical factors (thorax compres-
sion and visceral displacement) and the rhythmic activation of
body wall muscles.
To address whether locomotor-ventilatory interactions are
mechanically significant in running humans, Banzett and
colleagues [20] measured ‘step-driven flow’ in humans running
on a treadmill. ‘Step-driven flow’ is ventilatory flow and resulting
volumes attributable to locomotor-induced forces, once the
primary ventilatory pattern is removed (through either ensemble
averaging or filtering – see methods). Banzett and colleagues [20]
found the volumes attributable to step-driven flows to be a
negligible fraction of tidal volume, ,1–2% tidal volume per step.
Consequently, they concluded that locomotion has no meaningful
mechanical influence on breathing in humans [20]. Following
publication of this finding, there has been relatively little research
on the mechanical consequences of locomotor-ventilatory inter-
actions in running humans.
Yet, there are reasons to revisit the potential mechanical
significance of locomotor-ventilatory interactions in humans. In
the study by Banzett and colleagues, the subjects lightly rested
their hands on side rails while jogging on the treadmill [20]. Arm
swinging contributes to compression loads on the thorax during
locomotion, and to control of body and head rotation [29,34]. The
restriction of natural arm motion may have reduced mechanical
interactions between locomotion and ventilation. Furthermore,
although the volumes attributed to step-driven flows amounted to
only 1–2% of tidal volume per step, Banzett and colleagues
measured large transient flows associated with footstrike [20].
These step-driven flows lasted only a small fraction of the
ventilatory cycle, but exhibited peak magnitudes of 1 Ls21.
Although brief, step-driven flows reach a large fraction of
concurrent ventilatory flow, sometimes resulting in a brief mid-
breath reversal in flow (from inspiration to expiration or vice-
versa). We suggest that these transient flows have potential to
influence breathing dynamics and ventilatory muscle loading
without directly driving large ventilatory volumes.
Here we test the hypothesis that humans use LRC to minimize
antagonistic effects of step-driven flows on breathing dynamics.
Due the natural variability in coupling patterns in running humans
[14,21,22,23,24], it is possible to compare breathing dynamics
when individuals use their own ‘preferred’ and ‘avoided’
entrainment patterns. We analyze locomotor-ventilatory interac-
tions in fourteen physically fit subjects running on a treadmill at a
moderate self-selected speed (2.660.1 ms21) with natural arm
motion. Self-selected speed is likely to be close to energetically
optimal speed [35]. Running at speeds above preferred speeds
would increase both impact loads and ventilatory demand,
increasing the potential for antagonistic locomotor-ventilatory
interactions. We therefore consider self-selected speed to provide a
conservative test of the hypothesis that humans use LRC to
minimize antagonistic locomotor-ventilatory dynamics.
Materials and Methods
Ethics StatementThe Institutional Review Board of University of Utah approved
the protocols and informed consent documents, under IRB
#12361, and all subjects gave written informed consent.
Subjects and protocolFourteen adult subjects volunteered for this study (9 females 5
males; age 3662 years, mass 65.162.8 kg, height 1.7260.02 m).
Seven of the subjects were recreational runners who regularly ran
at least 20 miles per week, and seven were physically fit non-
runners (Table 1). This sampling was designed to obtain a broad
representation of human locomotor-ventilatory patterns, not to
statistically compare trained and non-trained runners, which has
been previously studied by others [23,24]. Participants ran at a
steady speed on a treadmill, using natural arm motion. After an
initial practice session, subjects were allowed to select their
preferred speed, with the instructions to run at a comfortable pace
that they could sustain for 30 minutes. They warmed up for
5 minutes and then continued to run for 5 minutes while we
recorded running and breathing patterns.
Data collectionWe tracked the step cycle by measuring acceleration at the top
of the head using an Endevco microtron accelerometer (model
7290A-10). The subjects wore a lightweight facemask equipped
with two-way non-rebreathing valves to measure inspiratory and
expiratory flow separately (Hans Rudoph Inc., Kansas City, MO,
Locomotor-Ventilatory Dynamics in Humans
PLOS ONE | www.plosone.org 2 August 2013 | Volume 8 | Issue 8 | e70752

USA). Each valve was connected by tubing (34.2 mm ID; Hans
Rudolph, Inc.) to screen pneumotachs (model 4813, Hans
Rudolph Inc.), with a calibrated flow range of 0–800 liters per
minute. The pressure ports of the screen pneumotachs were
attached to a differential pressure transducer (Omega Engineering,
Stamford CT, USA). To prevent motion artifact, the tubing and
pneumotachs were secured to a small backpack worn by the
subject, fitted with both waist and shoulder straps. Flow was
calibrated by recording the output voltage from a known volume
of air, using a 3 liter calibration syringe (Hans Rudolph Inc.).
The signals were amplified 10x (model P122, Grass Inc., West
Warwick RI, USA), sampled at 200 Hz and digitally recorded
onto computer using the MP100 data acquisition system with
AcqKnowledge software (BIOPAC Systems Inc., Goleta, CA,
USA.). Before analysis, data were low pass filtered at 30 Hz using
a zero-phase 6th order Butterworth filter in Matlab (butter and
filtfilt functions; Matlab release 13). These filter settings were
selected because they were found to effectively remove noise while
retaining higher frequency signal characteristics such as peaks
associated with footstrike impact.
Calculation of average step-driven flowTo enable comparison to previous findings, we calculate
average step-driven flow using similar analytical methods of
Banzett and colleagues [20]. We calculated an ensemble average
(EA) of ventilatory flow relative to the step cycle for each
individual, including at least 50 steps per subject [20]. The EA
technique samples flow segments distributed equally through the
ventilatory cycle, to quantify step-driven flow independent of
ventilatory phase. We calculated this EA for the 12 out of 14
subjects who exhibited varying phase of entrainment between
running and breathing cycles (Table 1). The resulting EA trace of
step-driven flow was integrated over time to obtain the locomotor
driven volume (LDV). Expiratory (positive) and inspiratory
(negative) volumes were integrated separately to assess the relative
magnitude of each. The LDV was normalized by the total
concurrent ventilatory volume (Vtot) over the same time periods as
the EA flow segments, to obtain a value as a percentage of
concurrent ventilatory flow. Note that our calculation of LDV
differs slightly from that used by Banzett and colleagues, who
reported the fractional contribution to tidal volume per step;
whereas we report the total fractional contribution to ventilation.
Humans typically use coupling ratios of 2:1 and higher (e.g.
[2,20,21,22,23]), taking multiple steps in each breath cycle;
consequently, a per step value does not provide a complete account
of the total magnitude of locomotor-driven ventilation.
A potential source of error in the calculated LDV is flow artifact
caused by motion of the tubes during running. As mentioned
earlier, we minimized this by securing the tubes to a lightweight
backpack worn by the subject. To quantify the error associated
with any remaining motion, we calculated an ‘artifact LDV’ for 5
subjects. We recorded inspiration and expiration in separate
channels, causing each channel to have a baseline of zero during
the opposing ventilatory half-cycle. During this period, the only
flow measured in that channel was small magnitude, high
frequency artifact flow due to tube motion. To quantify this, we
calculated a value of LDV using the same method described
above, except that the flow segments in the ensemble average were
obtained from the baseline of the inactive ventilation channel. This
method resulted in an artifact LDV value of 0.760.1% of Vtot
(mean 6 s.e.m.) during control running trials, with a worst case
artifact magnitude of 1% Vtot.
Testing for variation in step-driven flow with ventilatoryphaseThe visceral piston hypothesis predicts that the effect of
locomotor forces on flow depends on the phase of the breath
cycle in which the step occurs [2] (Fig. 1C). Deceleration of the
body at footstrike likely has two primary mechanical effects: 1)
compression of the thorax, and 2) viscoelastic bounce and rebound
of the guts [20,28]. Contraction of respiratory muscles and
inflation of the lungs differentially alters thoracic and abdominal
Table 1. Subject data.
Subject Gender Age (yrs) Mass (kg) Height (m) Speed (ms) Training Entrainment Coupling Ratio (s)
1 F 39 54.0 1.57 2.4 Casual Coupled, vp,vr* 2:1, 3:1
2 F 41 59.4 1.73 2.4 Nonrunner Coupled 2:1
3 M 35 72.5 1.75 2.9 Nonrunner Coupled, vr* 3:1, 3.5:1
4 M 43 76.2 1.83 2.4 Nonrunner Uncoupled*
5 F 43 60.8 1.73 2.6 Casual Coupled, vp* 2:1
6 F 23 59.0 1.65 2.4 Nonrunner Coupled, vp* 2:1
7 M 26 86.2 1.78 2.4 Nonrunner Coupled, vp,vr* 2:1, 2.5:1, 3:1
8 F 41 51.9 1.67 2.4 Casual Coupled, vp* 2.5:1
9 F 28 64.4 1.70 2.4 Nonrunner Uncoupled*
10 M 49 71.7 1.78 2.8 Trained Coupled, vr * 2:1, 3.5:1
11 F 47 62.6 1.65 2.4 Trained Coupled, vp,vr* 2:1,3:1
12 F 27 59.0 1.75 3.4 Trained Coupled,vp* 2:1
13 F 25 54.5 1.60 3.4 Trained Coupled 2:1
14 M 31 79.4 1.83 2.3 Nonrunner Uncoupled*
mean 36 65.1 1.72 2.6
s.e.m. 2 2.8 0.02 0.1
*Ensemble average calculated for this subject.Coupling abbreviations: vr = variable coupling ratio. vp = variable coupling phase relationship.doi:10.1371/journal.pone.0070752.t001
Locomotor-Ventilatory Dynamics in Humans
PLOS ONE | www.plosone.org 3 August 2013 | Volume 8 | Issue 8 | e70752

stiffness [36]. Locomotor forces are likely to have an enhanced
expiratory effect if they occur when the abdominal muscles are
being recruited for forced expiration, because these muscles resist
the descent of the guts (Fig. 1C).
The ensemble average technique cannot reveal this effect of
ventilatory phase because it averages step-driven flow across all
phases. Therefore, we also compared step-driven ventilation at
different phases of the breathing cycle, in bins of 20% of maximum
tidal volume (Vmax) during inspiration and expiration. From the
ensemble averaged data, we observed that step related oscillations
in flow occurred at step frequency and the 1st and 2nd harmonic of
step frequency. We high-pass filtered the flow signal to remove the
primary breathing pattern, so that only flows associated with the
step frequency or higher remained in the signal. We used a zero-
phase digital Butterworth filter (‘butter’ and ‘filtfilt’ functions in
Matlab version 6.5), with the cutoff frequency and filter order
specified using the Matlab function ‘buttord’ to create a filter that
lost no more than 1% of the signal greater than or equal to step
frequency and attenuated 99% of the signal below half the step
frequency. We then calculated an average 6 s.e.m of step-driven
flow for all steps occurring within each volume bin of the
ventilatory cycle.
Analysis of entrainment patternsWe used phase analysis to examine the locomotor-ventilatory
coupling patterns. The timing of each inspiratory and expiratory
transition was calculated relative to the step cycle. The beginning
of footstrike was defined as zero, and the phase of each ventilatory
transition was expressed in degrees between 0 and 360 or as a
fraction of the step cycle by dividing the phase angle by 360. The
step cycle was divided into 20 bins (18 degrees or 5% of the step
cycle each), and the frequency of inspiratory and expiratory
transitions was calculated for each bin. If breaths and steps occur
randomly with respect to each other, the distribution of ventilatory
transition events should not significantly differ from a uniform
circular distribution. For each subject whose ventilatory transitions
significantly differed from a uniform circular distribution (see
Statistics), we determined the ‘preferred’ phase for inspiratory and
expiratory initiation as the bin in which each of these events
occurred most frequently. Likewise, we determined ‘avoided’
phase as the bin in which each transition event occurred least
frequently.
Ventilatory transition timesTo quantify the time required for transition between ventilatory
half cycles (inspiration and expiration), we measured the time
between 50% peak flows (T50). The expiratory T50 is the time
from 50% peak inspiration to 50% peak expiration. Likewise, the
inspiratory T50 is the same measure between 50% peak expiration
and 50% peak inspiration. We compared T50 values between
‘preferred’ and ‘avoided’ phase bins, described above. If multiple
bins were tied for ‘preferred’ or ‘avoided’, we took the average of
the tied bins.
StatisticsKuiper’s test of circular uniformity was used to test whether
inspiratory and expiratory initiation events were distributed
randomly with respect to the step cycle. For subjects with
ventilatory transitions distributed non-uniformly relative to the
step cycle, a Friedman nonparametric repeated measures ANOVA
test was used to compare ventilatory transition times (T50)
between preferred and avoided phases of step cycle, with posthoc
pairwise comparisons using Dunn’s Multiple Comparisons.
Results
Step-driven oscillations in flowAll subjects exhibited high frequency, step-driven oscillations
in flow (Fig. 1B). An inspiratory pulse occurred immediately after
footstrike, followed by an expiratory pulse as the body
approached peak acceleration (Fig. 2). The associated inspiratory
and expiratory volumes amounted to 212.764.5% (mean 6S.D)
and 10.763.2%, respectively, of the concurrent ventilatory
volume (Fig. 2; Table 2), with peak flow magnitude averaging
0.9760.45 Ls21 across individuals. In some cases, these step-
driven flows were large enough to cause transient reversals in
flow (Fig. 3A).
Locomotor-ventilatory interactions and entrainmentAlthough the step-driven inspiratory and expiratory volumes
were equal on average (Fig. 2), the effect of locomotor acceleration
Figure 1. Locomotor-ventilatory interactions. (A–B) Typical head acceleration (top) and ventilatory flow (bottom: expiration positive,inspiration negative) during quiet standing (A), and moderate speed treadmill running (B). Note the high frequency oscillations in ventilatory flowduring running. (C) Schematic illustration of the ‘visceral-piston’ model for human locomotor-ventilatory interactions. Red arrows indicate muscleactions during inspiration and expiration.doi:10.1371/journal.pone.0070752.g001
Locomotor-Ventilatory Dynamics in Humans
PLOS ONE | www.plosone.org 4 August 2013 | Volume 8 | Issue 8 | e70752

Figure 2. Step-driven ventilatory flows and volumes. (A) Grand mean of step-driven ventilatory flow across subjects (mean6 95% confidenceinterval for 12 subjects, see also individual examples in Fig. 5). Expiration is positive. (B) Mean 6 SD of step-driven volume as a percentage of totalconcurrent ventilatory volume (Vtot), during level moderate speed running (N= 12, black dots show data from individuals). Step-driven flow andvolume data included here are from subjects with variation in the phase locking between steps and breaths (Table 1).doi:10.1371/journal.pone.0070752.g002
Figure 3. Step driven flow depends on ventilatory phase. (A) Variation in the magnitude of step-driven flows is apparent, particularly duringlow frequency breathing, as shown here during 3:1 (strides per breath) coupling. Note the reversal in flow in late expiration (asterisk). Dashed verticallines indicate footstrike events (data from subject 3). (B) Average step-driven flow (means 695%CI) for a representative subjective at four points inthe ventilatory cycle: 1) early expiration, high lung volume (90% Vmax), 2) late expiration, low lung volume (10% Vmax), 3) early inspiration, low lungvolume (10% Vmax) and 4) late inspiration, high lung volume (90% Vmax).doi:10.1371/journal.pone.0070752.g003
Locomotor-Ventilatory Dynamics in Humans
PLOS ONE | www.plosone.org 5 August 2013 | Volume 8 | Issue 8 | e70752

on flow varied considerably with ventilatory phase (Fig. 3). Across
individuals, we observed a significant relationship between tidal
volume at footstrike and the net step-driven volume (Fig. 4). Steps
occurring in late expiration (low lung volume) tend to have a net
inspiratory effect. Similarly, steps occurring in late inspiration
(high lung volume) tend to have a net expiratory effect. Thus, the
net effect of step-driven flow is synergistic early in the ventilatory
half-cycle and antagonistic late in each ventilatory half cycle. Flow
reversals occur most often in late expiration during slow, deep
breaths and coupling with ratios greater than 2.5:1 strides per
breath (Fig. 3A). However, most runners avoided these flow
reversals because they breathed more frequently, closer to a 2:1
stride per breath rhythm.
Runners varied in whether or how tightly they entrained
breathing and stepping rhythm (Table 1). Most (11 of 14) subjects
exhibited periods of fixed-ratio coupling between stepping and
breathing rhythms, with 2:1 strides per breath as the most
common pattern. Two of these subjects exhibited strict phase-
locked coupling with a single coupling ratio, whereas the
remaining 9 subjects exhibited varied phase or switching among
multiple coupling ratios (Table 1). Despite variation in coupling,
13 of 14 subjects had significantly non-uniform distributions of
ventilatory transitions relative to the step cycle (inspiratory,
expiratory or both; Table 3). Preferred transition phases tended
to correspond to regions of the step cycle that mechanically
assisted ventilation, or at least, did not impede it (Fig. 5).
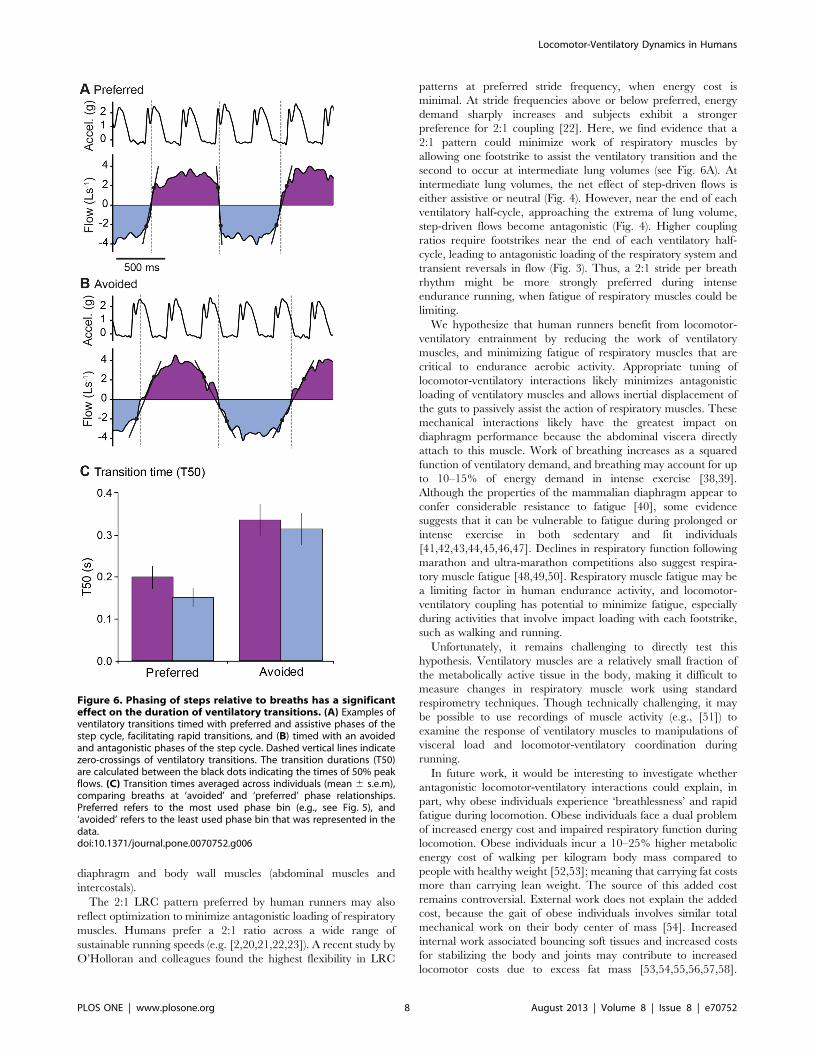
Although step-driven flows are brief, the phasing of transitions
relative to the step cycle does substantially influence breathing
dynamics. Whether coupled or not, most runners avoided reversals
in flow by using a breathing frequency near a 2:1 stride per breath
ratio and timing ventilatory transitions to coincide favorably with
step-driven flow (Fig. 5). Furthermore, the phasing between steps
and breath transitions influenced the time required for ventilatory
transitions. When runners coordinate the transitions to occur in
phases of the step cycle that promote flow, transitions occur 2X
more rapidly (Fig. 6). When compared across all uncoupled or
variably coupled runners (for which ‘avoided’ transition data were
available), ventilatory transitions in ‘preferred’ phase relationships
with the step cycle occurred more rapidly than those in ‘avoided’
phases (Fig. 6). The difference between preferred and avoided
transition T50 values was statistically significant (p = 0.0002 for
Freidman Test, p,0.05 Dunn’s Multiple Comparisons).
Discussion
The goal of this study was to examine whether the mechanical
interaction between running and breathing in humans is large
enough to be physiologically important. We measured the
ventilatory flows and volumes attributable to step-driven flows
during running at a moderate self-selected speed. The timing of
step-driven flows is consistent with the visceral piston hypothesis
for locomotor-ventilatory interactions in humans [2,19,20,27,28].
Table 2. Step-driven ventilatory volume, as a fraction ofconcurrent volume, during inspiration and expiration.
Subject Insp Exp
1 217.2% 15.2%
3 29.8% 9.3%
4 27.7% 7.6%
5 210.5% 7.5%
6 29.6% 9.2%
7 211.5% 8.2%
8 29.9% 8.1%
9 210.1% 9.6%
10 214.7% 14.8%
11 218.8% 14.2%
12 222.2% 15.4%
14 29.9% 8.9%
mean 212.7% 10.7%
s.d. 4.5% 3.2%
doi:10.1371/journal.pone.0070752.t002
Figure 4. The net effect of step-driven ventilation shifts from synergistic to antagonistic in each ventilatory half-cycle. We show netstep-driven volumes (in milliliters per step) as a function of the tidal volume at the time of footstrike during inspiration (left) and expiration (right),averaged across individuals (mean 6 95% CI, N = 12). At low lung volumes, the inspiratory pulse at footstrike is larger, leading to a net inspiratoryeffect. At high lung volumes, a larger expiratory pulse occurs, leading to a net expiratory effect.doi:10.1371/journal.pone.0070752.g004
Locomotor-Ventilatory Dynamics in Humans
PLOS ONE | www.plosone.org 6 August 2013 | Volume 8 | Issue 8 | e70752

Step-driven ventilation averaged around 10–12% of total venti-
latory volume with flow magnitudes around 1 Ls21. We found that
step-driven flow dynamics varied depending on ventilatory state
(high versus low lung volume), suggesting phase-dependent changes
in compliance and damping of thoraco-abdominal tissues. We also
discovered that the timing of impact loading relative to the
ventilatory cycle significantly influenced breathing dynamics.
Ventilatory transitions initiated in preferred (assistive) phases of
the step cycle occurred 2x faster than those in avoided
(antagonistic) phases. These findings suggest a physiologically
significant mechanical interaction exists between locomotion and
ventilation in humans.
The step-driven ventilatory volumes we measured appear to be
larger on average than those reported previously [20]. Here, step-
driven volumes amounted to 10–12% of total ventilation, which
amounts to 2.5–3.0% tidal volume per step during a 2:1 step per
breath rhythm. Banzett and colleagues reported a value of 1–2%
tidal volume per step. Nonetheless, the timing of peak step-driven
flow was consistent with Banzett and colleagues. The difference in
magnitude could be explained by a difference in arm motion
between the two studies. In the earlier study, the subjects lightly
rested their hands on side rails, whereas in the current study the
subjects were allowed to move their arms naturally while running.
Both step-induced thoracic loading and inertial displacement of
abdominal viscera likely contribute to locomotor-ventilatory
interactions while running [2,12,19,20,27]. Minimizing the effects
of arm loading on thoracic compression during running could
reduce both the mechanical and neural interactions between
locomotion and ventilation [29,32,33,37]. Nonetheless, we do find
a similar pattern, and although the total step-driven volumes are
small on a per step basis, they do have a significant mechanical
influence on breathing dynamics.
What is the physiological significance of locomotor-ventilatory entrainment in humans?The frequency of step related flows is too high relative to breath
frequency to allow direct coupling to drive ventilation (Figs. 1, 2).
Yet, our data reveal that runners prefer to time ventilatory
transitions to periods in the step cycle that assist rather than
impede flow (Fig. 5), and this timing significantly reduces the time
required for transitions (Fig. 6). These data suggest that step-driven
flows have potential to influence the work of respiratory muscles,
because they significantly influence breathing dynamics, particu-
larly at ventilatory transitions (expiration to inspiration and vice
versa). Appropriate coordination of stepping and breathing rhythm
may act to minimize antagonistic loading of the respiratory
muscles caused by motions of the abdomen and chest wall.
Although locomotor-ventilatory coupling is more flexible in
humans than in quadrupedal mammals [2,3,11,12,13,22], cou-
pling in humans may reduce conflicting demands placed on the
Figure 5. Subjects prefer to initiate ventilatory transitions atphases that assist rather than imped flow. The distribution ofventilatory transitions relative to the step cycle is non-uniform (Table 3).Here we show the net bias in ventilatory transitions relative to stepcycle (left axis and bars), with transitions to expiration as positive,transition to inspiration as negative. The ‘net bias’ value is the numberof expiratory transitions minus the number of inspiratory transitions ineach phase bin. Step-driven flow is overlaid for reference (right axis andlines). Data from 3 individuals illustrates typical variation betweenstrongly coupled (A), variably coupled (B) and uncoupled (C) subjects.Although variation exists, the timing of transitions is clearly non-random, and exhibits some correspondence to the step-driven flow.doi:10.1371/journal.pone.0070752.g005
Table 3. Mean phase and dispersion (in degrees) ofventilatory transitions relative to the step cycle.
Phase relative to step cycle (degrees)
Subject Inspiration Expiration
N mean disp mean disp
1 57 296* 13.0 121* 6.2
2 72 32* 0.1 352* 0.2
3 48 345* 0.4 285* 0.6
{4 70 286 16.0 282 11.6
5 86 176* 0.9 164* 3.0
6 82 28* 0.5 313* 1.2
7 62 297* 1.1 273* 9.5
8 87 325* 0.8 255* 1.1
9 55 203 99.0 334* 0.9
10 35 296* 0.5 356* 0.5
11 79 153* 6.2 154* 1.8
12 90 282* 1.2 150* 8.2
13 87 308* 0.4 212* 0.4
14 62 324 26.0 17* 13.6
*Indicates significant difference from uniform circular distribution based onKuiper’s test.{Indicates subject with no statistical evidence of transition entrainment ininspiration or expiration.doi:10.1371/journal.pone.0070752.t003
Locomotor-Ventilatory Dynamics in Humans
PLOS ONE | www.plosone.org 7 August 2013 | Volume 8 | Issue 8 | e70752
diaphragm and body wall muscles (abdominal muscles and
intercostals).
The 2:1 LRC pattern preferred by human runners may also
reflect optimization to minimize antagonistic loading of respiratory
muscles. Humans prefer a 2:1 ratio across a wide range of
sustainable running speeds (e.g. [2,20,21,22,23]). A recent study by
O’Holloran and colleagues found the highest flexibility in LRC
patterns at preferred stride frequency, when energy cost is
minimal. At stride frequencies above or below preferred, energy
demand sharply increases and subjects exhibit a stronger
preference for 2:1 coupling [22]. Here, we find evidence that a
2:1 pattern could minimize work of respiratory muscles by
allowing one footstrike to assist the ventilatory transition and the
second to occur at intermediate lung volumes (see Fig. 6A). At
intermediate lung volumes, the net effect of step-driven flows is
either assistive or neutral (Fig. 4). However, near the end of each
ventilatory half-cycle, approaching the extrema of lung volume,
step-driven flows become antagonistic (Fig. 4). Higher coupling
ratios require footstrikes near the end of each ventilatory half-
cycle, leading to antagonistic loading of the respiratory system and
transient reversals in flow (Fig. 3). Thus, a 2:1 stride per breath
rhythm might be more strongly preferred during intense
endurance running, when fatigue of respiratory muscles could be
limiting.
We hypothesize that human runners benefit from locomotor-
ventilatory entrainment by reducing the work of ventilatory
muscles, and minimizing fatigue of respiratory muscles that are
critical to endurance aerobic activity. Appropriate tuning of
locomotor-ventilatory interactions likely minimizes antagonistic
loading of ventilatory muscles and allows inertial displacement of
the guts to passively assist the action of respiratory muscles. These
mechanical interactions likely have the greatest impact on
diaphragm performance because the abdominal viscera directly
attach to this muscle. Work of breathing increases as a squared
function of ventilatory demand, and breathing may account for up
to 10–15% of energy demand in intense exercise [38,39].
Although the properties of the mammalian diaphragm appear to
confer considerable resistance to fatigue [40], some evidence
suggests that it can be vulnerable to fatigue during prolonged or
intense exercise in both sedentary and fit individuals
[41,42,43,44,45,46,47]. Declines in respiratory function following
marathon and ultra-marathon competitions also suggest respira-
tory muscle fatigue [48,49,50]. Respiratory muscle fatigue may be
a limiting factor in human endurance activity, and locomotor-
ventilatory coupling has potential to minimize fatigue, especially
during activities that involve impact loading with each footstrike,
such as walking and running.
Unfortunately, it remains challenging to directly test this
hypothesis. Ventilatory muscles are a relatively small fraction of
the metabolically active tissue in the body, making it difficult to
measure changes in respiratory muscle work using standard
respirometry techniques. Though technically challenging, it may
be possible to use recordings of muscle activity (e.g., [51]) to
examine the response of ventilatory muscles to manipulations of
visceral load and locomotor-ventilatory coordination during
running.
In future work, it would be interesting to investigate whether
antagonistic locomotor-ventilatory interactions could explain, in
part, why obese individuals experience ‘breathlessness’ and rapid
fatigue during locomotion. Obese individuals face a dual problem
of increased energy cost and impaired respiratory function during
locomotion. Obese individuals incur a 10–25% higher metabolic
energy cost of walking per kilogram body mass compared to
people with healthy weight [52,53]; meaning that carrying fat costs
more than carrying lean weight. The source of this added cost
remains controversial. External work does not explain the added
cost, because the gait of obese individuals involves similar total
mechanical work on their body center of mass [54]. Increased
internal work associated bouncing soft tissues and increased costs
for stabilizing the body and joints may contribute to increased
locomotor costs due to excess fat mass [53,54,55,56,57,58].
Figure 6. Phasing of steps relative to breaths has a significanteffect on the duration of ventilatory transitions. (A) Examples ofventilatory transitions timed with preferred and assistive phases of thestep cycle, facilitating rapid transitions, and (B) timed with an avoidedand antagonistic phases of the step cycle. Dashed vertical lines indicatezero-crossings of ventilatory transitions. The transition durations (T50)are calculated between the black dots indicating the times of 50% peakflows. (C) Transition times averaged across individuals (mean 6 s.e.m),comparing breaths at ‘avoided’ and ‘preferred’ phase relationships.Preferred refers to the most used phase bin (e.g., see Fig. 5), and‘avoided’ refers to the least used phase bin that was represented in thedata.doi:10.1371/journal.pone.0070752.g006
Locomotor-Ventilatory Dynamics in Humans
PLOS ONE | www.plosone.org 8 August 2013 | Volume 8 | Issue 8 | e70752

Furthermore, excess fat is associated with reduced operating lung
volumes and decreased respiratory compliance, factors that
increase the work of breathing [59,60]. Soft-tissue bouncing
induced by impact loads during walking and running may further
increase the work of breathing and the rate of respiratory muscle
fatigue in overweight and obese individuals. Whereas in lean
individuals a large fraction of soft-tissue mass is concentrated intra-
abdominally, obesity results in a large fraction of soft-tissue mass
distributed external to the body wall muscles. Thus, high adiposity
will increase soft-tissue bouncing and reduce the ability to actively
tune soft-tissue dynamics through abdominal muscle contraction.
Larger, uncontrolled soft-tissue motions may result in antagonistic
locomotor-ventilatory interactions and higher mechanical work of
breathing during exercise. We predict that the problems of obesity
are exacerbated by antagonistic locomotor-ventilatory interactions
during walking and running.
ConclusionsBiomechanics studies often focus on musculoskeletal and
biomechanical factors as limits to performance. For example, leg
muscle strength is widely thought to limit top running speed
during sprinting [61,62]. However, during endurance locomotion,
respiratory muscles, not leg muscles, may limit maximum exercise
intensity and duration [47]. We suggest that when assessing the
physiological importance of locomotor-respiratory coupling in
humans, it may be short sighted to place too much emphasis on
the comparatively small volumes attributed to it. Entrainment
might serve a number of beneficial physiological functions – i.e.,
reducing the work of ventilatory muscles, preventing respiratory
muscle fatigue, and improving respiratory efficiency through
enhanced gas mixing, transport and exchange. Consequently, the
precise volume driven by locomotion might be less important than
the associated intrapulmonary dynamics. Humans are well
adapted for economic walking and endurance running
[34,63,64,65]. Multiple derived characters suggest aerobic endur-
ance as a key evolutionary pressure in the human lineage,
including skeletal morphology, developed tendon springs and
enhanced heat dissipation [34,63,64,66]. We will not be surprised,
therefore, if the unusual locomotor-respiratory coupling patterns
exhibited by human runners proves to be another example of
evolutionary adaptation in support of the exceptional endurance
running capabilities of humans [63,64].
Acknowledgments
We thank Eric Stakebake for assistance in the experiments, Franz Goller
for loan of equipment and the late Farish Jenkins Jr. for helping to make
the project possible. We also thank Joanne Gordon for her comments on
the manuscript draft.
Author Contributions
Conceived and designed the experiments: MAD DRC DMB. Performed
the experiments: MAD. Analyzed the data: MAD. Wrote the paper: MAD.
Interpreted the data and revised the manuscript: DMB DRC.
References
1. Alexander RM (1989) On the synchronization of breathing with running in
wallabies (Macropus Spp) and horses (Equus-Caballus). Journal of Zoology 218: 69–85.
2. Bramble DM, Carrier DR (1983) Running and breathing in mammals. Science219: 251–256.
3. Boggs DF (2002) Interactions between locomotion and ventilation in tetrapods.Comparative Biochemistry and Physiology A-Molecular and Integrative
Physiology 133: 269–288.
4. Giuliodori MJ, Lujan HL, Briggs WS, DiCarlo SE (2009) A model of locomotor-
respiratory coupling in quadrupeds. Advances in Physiology Education 33: 315–318.
5. Carrier DR (1990) Activity of the hypaxial muscles during walking in the lizard.Journal of Experimental Biology 152: 453–470.
6. Carrier DR (1991) Conflict in the hypaxial musculoskeletal system –
Documenting an evolutionary constraint. American Zoologist 31: 644–654.
7. Codd JR, Boggs DF, Perry SF, Carrier DR (2005) Activity of three muscles
associated with the uncinate processes of the giant canada goose Branta canadensismaximus. Journal of Experimental Biology 208: 849–857.
8. Carrier DR (1996) Function of the intercostal muscles in trotting dogs:Ventilation or locomotion? Journal of Experimental Biology 199: 1455–1465.
9. Deban SM, Carrier DR (2002) Hypaxial muscle activity during running andbreathing in dogs. Journal of Experimental Biology 205: 1953–1967.
10. Hodges PW, Gandevia SC (2000) Changes in intra-abdominal pressure duringpostural and respiratory activation of the human diaphragm. Journal of Applied
Physiology 89: 967–976.
11. Simons RS (1999) Running, breathing and visceral motion in the domestic
rabbit (Oryctolagus cuniculus): Testing visceral displacement hypotheses. Journal ofExperimental Biology 202: 563–577.
12. Baudinette RV, Gannon BJ, Runciman WB, Wells S, Love JB (1987) Docardiorespiratory frequencies show entrainment with hopping in the tammar
wallaby? Journal of Experimental Biology 129: 251–263.
13. Funk GD, Sholomenko GN, Valenzuela IJ, Steeves JD, Milsom WK (1993)
Coordination of wing beat and respiration in canada geese during free flight.Journal of Experimental Biology 175: 317–323.
14. Bernasconi P, Kohl J (1993) Analysis of coordination between breathing and
exercise rhythms in man. Journal of Physiology-London 471: 693–706.
15. Bonsignore MR, Morici G, Abate P, Romano S, Bonsignore G (1998)
Ventilation and entrainment of breathing during cycling and running in
triathletes. Medicine and Science in Sports and Exercise 30: 239–245.
16. Funk GD, Valenzuela IJ, Milsom WK (1997) Energetic consequences ofcoordinating wingbeat and respiratory rhythms in birds. Journal of Experimental
Biology 200: 915–920.
17. Nassar PN, Jackson AC, Carrier DR (2001) Entraining the natural frequencies of
running and breathing in guinea fowl (Numida meleagris). Journal ofExperimental Biology 204: 1641–1651.
18. Simons RS (1996) Lung morphology of cursorial and non-cursorial mammals:
Lagomorphs as a case study for a pneumatic stabilization hypothesis. Journal of
Morphology 230: 299–316.
19. Bramble DM (1989) Axial-appendicular dynamics and the integration of
breathing and gait in mammals. American Zoologist 29: 171–186.
20. Banzett RB, Mead J, Reid MB, Topulos GP (1992) Locomotion in men has no
appreciable mechanical effect on breathing. Journal of Applied Physiology 72:
1922–1926.
21. Persegol L, Jordan M, Viala D (1991) Evidence for the entrainment of breathing
by locomotor pattern in human. Journal De Physiologie 85: 38–43.
22. O’Halloran J, Hamill J, McDermott WJ, Remelius JG, Van Emmerik REA
(2012) Locomotor-respiratory coupling patterns and oxygen consumption during
walking above and below preferred stride frequency. European Journal of
Applied Physiology 112: 929–940.
23. McDermott W, Emmerik RA, Hamill J (2003) Running training and adaptive
strategies of locomotor-respiratory coordination. European Journal of Applied
Physiology 89: 435–444.
24. Bernasconi P, Burki P, Buhrer A, Koller EA, Kohl J (1995) Running training
and coordination between breathing and running rhythms during aerobic and
anaerobic donditions in humans. European Journal of Applied Physiology and
Occupational Physiology 70: 387–393.
25. Ebert D, Rassler B, Hefter H (2000) Coordination between breathing and
forearm movements during sinusoidal tracking. European Journal of Applied
Physiology 81: 288–296.
26. Rassler B, Ebert D, Waurick S, Junghans R (1996) Coordination between
breathing and finger tracking in man. Journal of Motor Behavior 28: 48–56.
27. Bramble DM, Jenkins FA (1993) Mammalian locomotor-respiratory integration
– Implications for diaphragmatic and pulmonary design. Science 262: 235–240.
28. Lee HT, Banzett RB (1997) Mechanical links between locomotion and
breathing: Can you breathe with your legs? News in Physiological Sciences
12: 273–278.
29. Pontzer H, Holloway JH, Raichlen DA, Lieberman DE (2009) Control and
function of arm swing in human walking and running. Journal of Experimental
Biology 212: 523–534.
30. Loring SH (1992) Action of human respiratory muscles inferred from finite
element analysis of rib cage. Journal of Applied Physiology 72: 1461–1465.
31. Loring SH, De Troyer A (1985) Actions of respiratory muscles. In: Macklem PT,
editor. The Thorax. New York: M. Dekker. 327–349.
32. Giraudin A, Cabirol-Pol M-J, Simmers J, Morin D (2008) Intercostal and
abdominal respiratory motoneurons in the neonatal rat spinal cord: Spatiotem-
poral organization and responses to limb afferent stimulation. Journal of
Neurophysiology 99: 2626–2640.
33. Giraudin A, Le Bon-Jego M, Cabirol M-J, Simmers J, Morin D (2012) Spinal
and pontine relay pathways mediating respiratory rhythm entrainment by limb
Locomotor-Ventilatory Dynamics in Humans
PLOS ONE | www.plosone.org 9 August 2013 | Volume 8 | Issue 8 | e70752

proprioceptive inputs in the neonatal rat. Journal of Neuroscience 32: 11841–
11853.
34. Pontzer H, Raichlen D, Sockol M (2009) The metabolic cost of walking in
humans, chimpanzees, and early hominins. Journal of Human Evolution 56: 43–
54.
35. Steudel-Numbers KL, Wall-Scheffler CM (2009) Optimal running speed and the
evolution of hominin hunting strategies. Journal of Human Evolution 56: 355–
360.
36. DeTroyer A (1991) Respiratory Muscles. In: West JB, editor. The Lung:
Scientific Foundations. New York: Raven Press, Ltd. 869–883.
37. Fabre N, Perrey S, Arbez L, Rouillon J-D (2007) Neuro-mechanical and
chemical influences on locomotor respiratory coupling in humans. Respiratory
Physiology & Neurobiology 155: 128–136.
38. Aaron EA, Seow KC, Johnson BD, Dempsey JA (1992) Oxygen cost of exercise
hyperpnea: implications for performance. Journal of Applied Physiology 72:
1818–1825.
39. Dominelli PB, Sheel AW (2012) Experimental approaches to the study of the
mechanics of breathing during exercise. Respiratory Physiology & Neurobiology
180: 147–161.
40. Dempsey JA, Adams L, Ainsworth DM, Fregori RF, Gallagher CG, et al. (1996)
Airway, lung, and respiratory muscle function during exercise. In: Shepard JT,
editor. Handbook of Physiology, Sect 12, Exercise: Regulation and Integration
of Multiple Systems: Oxford Univ. Press. 448–514.
41. Johnson BD, Aaron EA, Babcock MA, Dempsey JA (1996) Respiratory muscle
fatigue during exercise: Implications for performance. Medicine and Science in
Sports and Exercise 28: 1129–1137.
42. Babcock MA, Pegelow DF, Johnson BD, Dempsey JA (1996) Aerobic fitness
effects on exercise-induced low-frequency diaphragm fatigue. Journal of Applied
Physiology 81: 2156–2164.
43. Mador MJ, Magalang UJ, Rodis A, Kufel TJ (1993) Diaphragmatic fatigue after
exercise in healthy-human subjects. American Review of Respiratory Disease
148: 1571–1575.
44. Johnson BD, Babcock MA, Suman OE, Dempsey JA (1993) Exercise-induced
diaphragmatic fatigue in healthy humans. Journal of Physiology-London 460:
385–405.
45. Babcock MA, Pegelow DF, Harms CA, Dempsey JA (2002) Effects of respiratory
muscle unloading on exercise-induced diaphragm fatigue. Journal of Applied
Physiology 93: 201–206.
46. Romer LM, McConnell AK, Jones DA (2002) Inspiratory muscle fatigue in
trained cyclists: Effects of inspiratory muscle training. Medicine & Science in
Sports & Exercise 34: 785–792.
47. Romer LM, Polkey MI (2008) Exercise-induced respiratory muscle fatigue:
Implications for performance. Journal of Applied Physiology 104: 879–888.
48. Warren G, Cureton KJ, Sparling PB (1989) Does lung function limit
performance in a 24-hour ultramarathon? Respiration Physiology 78: 253–2264.
49. Mahler DA, Loke JD (1981) Lung function after marathon running at warm and
cold ambient temperatures. American Review Respiratory Disease 124: 154–157.
50. Loke JD, Virgulto JA (1982) Respiratory muscle fatigue after marathon running.
Journal Applied Physiology 52: 821–824.51. Saunders SW, Rath D, Hodges PW (2004) Postural and respiratory activation of
the trunk muscles changes with mode and speed of locomotion. Gait & Posture20: 280–290.
52. Browning RC, Baker EA, Herron JA, Kram R (2006) Effects of obesity and sex
on the energetic cost and preferred speed of walking. Journal of AppliedPhysiology 100: 390–398.
53. Peyrot N, Thivel D, Isacco L, Morin J-B, Duche P, et al. (2009) Do mechanicalgait parameters explain the higher metabolic cost of walking in obese
adolescents? Journal of Applied Physiology 106: 1763–1770.54. Browning RC, McGowan CP, Kram R (2009) Obesity does not increase
external mechanical work per kilogram body mass during walking. Journal of
Biomechanics 42: 2273–2278.55. Zelik KE, Kuo AD (2010) Human walking isn’t all hard work: Evidence of soft
tissue contributions to energy dissipation and return. Journal of ExperimentalBiology 213: 4257–4264.
56. DeVita P, Helseth J, Hortobagyi T (2007) Muscles do more positive than
negative work in human locomotion. Journal of Experimental Biology 210:3361–3373.
57. Daley MA, Usherwood JR (2010) Two explanations for the compliant runningparadox: reduced work of bouncing viscera and increased stability in uneven
terrain. Biology Letters 6: 418–421.58. Soo CH, Donelan JM (2010) Mechanics and energetics of step-to-step transitions
isolated from human walking. Journal of Experimental Biology 213: 4265–4271.
59. Dreher M, Kabitz H-J (2012) Impact of obesity on exercise performance andpulmonary rehabilitation. Respirology 17: 899–907.
60. Villiot-Danger J-C, Villiot-Danger E, Borel J-C, Pepin J-L, Wuyam B, et al.(2011) Respiratory muscle endurance training in obese patients. International
Journal of Obesity 35: 692–699.
61. Usherwood JR, Wilson AM (2006) Accounting for elite indoor 200 m sprintresults. Biology Letters 2: 47–50.
62. Weyand PG, Sternlight DB, Bellizzi MJ, Wright S (2000) Faster top runningspeeds are achieved with greater ground forces not more rapid leg movements.
Journal of Applied Physiology 89: 1991–1999.63. Carrier DR (1984) The energetic paradox of human running and Hominid
evolution. Current Anthropology 25: 483–495.
64. Bramble DM, Lieberman DE (2004) Endurance running and the evolution ofHomo. Nature 432: 345–352.
65. Liebenberg L (2008) The relevance of persistence hunting to human evolution.Journal of Human Evolution 55: 1156–1159.
66. Raichlen DA, Armstrong H, Lieberman DE (2011) Calcaneus length determines
running economy: implications for endurance running performance in modernhumans and Neandertals. Journal of Human Evolution 60: 299–308.
Locomotor-Ventilatory Dynamics in Humans
PLOS ONE | www.plosone.org 10 August 2013 | Volume 8 | Issue 8 | e70752