impairments of the medial olivocochlear system increase the risk
TRANSCRIPT

Impairments of the Medial Olivocochlear SystemIncrease the Risk of Noise-Induced Auditory
Neuropathy in Laboratory Mice
*Bradford J. May, *Amanda M. Lauer, and †Matthew J. Roos
Departments of *OtolaryngologyYHead and Neck Surgery, and ÞNeuroscience, Johns Hopkins University,Baltimore, Maryland, U.S.A.
Hypothesis: Impairments of the medial olivocochlear system(MOCS) increase the risk of environmentally induced auditoryneuropathy spectrum disorder (ANSD).Background: ANSD is a problem in the neural transmissionof auditory information that accounts for 10% to 15% of thecases of pediatric hearing loss. The underlying mechanisms ofthe disorder remain poorly understood, but noise exposure isan important risk factor. The goal of this study was to identifyenvironmental conditions and genetic predispositions that leadto ANSD. Our approach was based on the assumption that noiseinduces ANSD by impeding the functional maturation of thebrain’s sound coding pathways. Because the MOCS adjusts thesensitivity of the inner ear to noise, impairments of this feed-back are predicted to increase the disruptive effects of environ-mental exposures.Methods: An animal model of ANSD was created by rearingmice in noise. MOCS protection was assessed by comparingthe incidence of noise-induced ANSD among knockout micelacking feedback and wild-type (WT) controls. The mice werescreened for ANSD with distortion product otoacoustic emis-
sions, auditory brainstem responses, and behavioral measuresof gap detection. Single-unit recording procedures were usedto link these deficits to impaired synaptic transmission in theventral cochlear nucleus.Results: ANSD manifested in noise-reared mice as intact dis-tortion product otoacoustic emissions, abnormal auditory brain-stem responses, and impaired gap detection. The phenotype wasnot observed among quiet-reared WT mice but was occasionallynoted among noise-reared WT mice. The incidence of ANSDsignificantly increased among knockout mice, especially whenthey were reared in noise.Conclusion: Noise promotes ANSD by altering the functionalmaturation of the brain’s temporal pathways. Noise-inducedimpairments are reduced by the sound-attenuating effects of theMOCS. Noise levels do not need to be unnaturally loud toconstitute significant risk in MOCS-compromised individuals.KeyWords: Background noiseVDevelopmental impairmentVEfferent protectionVTemporal processing deficit.
Otol Neurotol 32:1568Y1578, 2011.
Auditory neuropathy spectrum disorder (ANSD) de-scribes a prevalent hearing disorder that is characterizedby normal cochlear outer hair cell (OHC) function andabnormal auditory nerve function (1). Minimum diag-nostic criteria include normal otoacoustic emissions andmissing or aberrant auditory brainstem responses (ABRs).Changes in the ABR are assumed to arise from a dys-synchronization of sound-driven activity in the auditorynerve. The loss of temporal processing can impair speech,sound localization, and listening in noise.
The impact of ANSD on speech has encouraged neo-natal screening programs for early diagnosis of the dis-order. These programs have revealed a higher incidenceof ANSD among infants who have been treated in neo-natal intensive care units (NICUs) (2,3). Although thehealth concerns that necessitate early hospitalization arevaried, a constant factor in NICU treatments is the noiseexposure that is generated by equipment, procedures, andstaff (4,5). Several surveys have reported NICU noiselevels consistently above the recommendations of theAmerican Academy of Pediatrics for both sustained (Leq)and maximum transient exposures (Lmax) (6).
At present, significant ambiguities remain on whatconstitutes an acceptable level of noise exposure in hu-man infants. Although it is clear that a single loud exposureat a young age can accelerate hearing loss and promotespiral ganglion degeneration (7), the effects of persis-tent moderate exposures are difficult to estimate because
Address correspondence and reprint requests to Bradford J. May,Ph.D., Traylor Research Building, Room 521, 720 Rutland Avenue,Baltimore, MD 21205; E-mail: [email protected] of support: This research was funded by grants DC009353,
DC010095, and DC005211 from the National Institute on Deafness andother Communication Disorders.
Otology & Neurotology32:1568Y1578 � 2011, Otology & Neurotology, Inc.
1568
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.

environmental noise exerts pervasive effects on the de-veloping auditory system. The deficits often manifest asperceptual disorders that are independent of overt hearingloss (8,9).
Physiologic studies of the cochlea and auditory nervehave provided detailed functional analyses of the medialolivocochlear system (MOCS). In healthy individuals, therelease of acetylcholine by the MOCS hyperpolarizes themembrane potentials of OHCs (10). The resulting de-crease in cochlear mechanical sensitivity protects the earfrom the damaging levels of noise (11) and enhancesauditory processing in distracting levels of noise (12,13).Recent clinical evidence suggests that ANSD is moreprevalent among MOCS-compromised children (14).One explanation for this increased risk is that synapticdevelopment of the central auditory pathways requiresregulated sound-driven activity in the ascending inputsof the auditory nerve (15Y17). When young individualsare exposed to environmental noise, MOCS feedback maypreserve healthy synaptic development by regulating po-tentially disruptive levels of noise-driven activity.
To investigate this novel developmental role for theMOCS, the feedback system was silenced in laboratorymice by a genetic modification of cholinergic signaling(18). The mice were reared in background noise and thenscreened for ANSD using established diagnostic criteria.The audiologic implications of the observed processingdeficits were further delineated with a behavioral assayof temporal acuity. The refinement of this animal modelallowed us to relate common clinical observations to di-rect electrophysiologic recordings from the postsynaptictargets of the auditory nerve in the ventral cochlear nucleus.These recordings revealed significant functional impair-ments of the synaptic specializations that dictate the qualityof temporal coding in the auditory brainstem.
METHODS
Interactions between the potential environmental and geneticprecursors of ANSD were evaluated with a 2 � 2 factorialdesign. Environmental factors were investigated by rearing micein quiet or noisy environments. Genetic factors were assessedby comparing noise effects in wild-type (WT) and knockout(KO) mice.Breeding pairs of KO mice were obtained from Jackson Labs
(CBACaJ;129S-Chrna9tm1Bedv/J, stock no. 005696). This strainwas generated with a mixed CBA/CaJ and 129S6/SvEvTacbackground to control for the early onset of age-related hearingloss. The mice were born and housed in the test environmentsfor the duration of the study. They were periodically removedfor 1-hour behavioral sessions and concurrent distortion productotoacoustic emission (DPOAE) and ABR recordings. All testingwas completed by 8 weeks of age. DPOAE input-output func-tions, conventional ABR thresholds, and cochlear histologyshowed no indications of hearing loss among KO or WT mice.It has been previously established that a null mutation of
the >9 nicotinic acetylcholine receptor subunit in the KOstrain eliminates MOC-mediated control of cochlear sensitivity(18,19). MOC projections remain intact, but their release ofacetylcholine has no effect on OHCs in the absence of functional
>9 receptors. Homozygous WT mice from the same backgroundstrain maintain a functional MOCS. A total of 34 KO mice and31 WT mice were tested. Both male and female mice were used.Environmental noise levels were manipulated by rearing
mice in institutional vivaria with different patterns of investi-gator access (20). Daily noise levels in the 2 facilities are shownin Figure 1. The heavy-traffic room provided high-densityhousing for a large number of investigators. Sustained increasesin ambient noise levels were produced by high-volume air ex-change systems. High-energy transients were associated withbursts of investigator activity. The low-traffic room was quieterand tended to have fewer high level transients. Investigatoractivity decreased during nonworking hours in both facilities(20), but air-exchange systems remained louder in the heavy-traffic room. For comparison, the time scale of the noise recordsis expanded in Figure 1C to match published noise levels in aNICU (4). Although the acoustic environment of the heavy-traffic vivarium was complex, it provided a conservative esti-mate of the NICU exposure.
Assessments of OHC Survival PatternsFunctional assessments of the integrity of OHCs were based
on published measures for relating sensorineural hearing lossto DPOAEs in patient populations (21). General methods forrecording DPOAEs in mice are described by Parham et al.(22). Because ANSD is attributed to the abnormal transmis-sion properties of the auditory nerve (23), DPOAEs and OHCsurvival patterns were expected to remain normal in all treat-ment groups (24).DPOAEs were measured using an ear insert consisting of
independent conduction tubes for 2 remote speakers and a mi-crophone (Etymotic ER-10C). F2 primary tones with a fre-quency of 16 kHz were presented from one speaker. F1 primarytones with an F2/F1 ratio of 1.2 were presented simultaneouslyfrom the second speaker. The level of F1 was varied from 5 to65 dB SPL to produce input-output functions relating DPOAEmagnitude to stimulus level. The level of F2 was always 10 dBlower than the level of F1. Responses were averaged over 50repetitions.Eight cochleae were processed (4 quiet-reared KO and 4
noise-reared KO) for direct visualization of OHC survival pat-terns in the apical, middle, and basal turns of the organ of Corti.The cochleae were perfused with 4% buffered paraformalde-hyde, post-fixed at 4-C for 3 hours, and rinsed in phosphatebuffer. Individual turns of the organ of Corti were separatedfrom the tectorial membrane and removed for processing aswhole mounts. Hair cells were stained with antibodies directedagainst myosin 7a (25). Olivocochlear terminations were labeledwith antibodies targeting synaptic vesicle protein 2 (26).
The Auditory Brainstem ResponseANSD patients show missing or abnormal ABRs, presum-
ably because the conduction of sound-evoked activity throughthe auditory nerve is abnormally dys-synchronized (27). Thisobservation was replicated by comparing the effects of rear-ing environment and MOCS status on the magnitude of ABRwaveforms.ABRs were recorded in a sound isolation chamber under light
ketamine anesthesia. Sound-driven activity was evoked by free-field presentations of 8-kHz tone bursts with a duration of 20 msand rise/fall times of 0.5 ms. The interstimulus interval (ISI)ranged from 1 to 15 ms. This protocol was designed to assessthe quality of the ABR over the same timing conditions that
1569AUDITORY NEUROPATHY IN MICE
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.

produced the most robust indications of ANSD during behavioraltesting.Sound-evoked potentials were recorded with a triad of plati-
num needle electrodes that were attached to the vertex (non-inverting electrode), to the skin covering the bulla (invertingelectrode), and to a hind limb (ground electrode). Electrical ac-tivity was amplified (300,000�), band-pass filtered (0.3Y3 kHz),and digitally sampled (30-ms recording at a 10-kHz sampling
rate). The averaged waveform was based on 300 stimulusrepetitions.
Single-Unit Recording in the VentralCochlear Nucleus
The analysis of ABR waveforms was supplemented withdirect electrophysiologic recordings of single-unit activity in theventral cochlear nucleus. Equipment for single-unit recording
FIG. 1. Daily noise levels in the 2 rearing environments. A, Bursts of investigator-generated noise are louder and more frequent inthe heavy-traffic vivarium than in the low-traffic vivarium. B, Long-term spectra also indicate higher sustained noise levels in the heavy-traffic vivarium. C, The time scale has been expanded to facilitate comparisons with published noise levels in a typical NICU (adapted fromBusch-Vishniac et al., 2005). Gray lines representing the 80-dB noise floor in the NICU are superimposed on vivarium recordings.
1570 B. J. MAY ET AL.
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.

in the ventral cochlear nucleus has been described in previouspublications (28,29). Physiologic protocols were implementedon programmable real-time processors (Tucker-Davis Tech-nologies, RX6) using custom MATLAB software. Auditorystimuli were generated at a 200-kHz sampling rate, attenuated tocontrol presentation levels (Tucker-Davis Technologies, PA5),and amplified (Crown D-75) before being passed to a calibratedfree-field speaker (Tannoy, Arena). Single-unit activity was am-plified (10,000�), low-pass filtered (5 kHz), and digitized witha voltage level discriminator.Electrophysiologic methods were based on a surgical expo-
sure of the dorsal surface of the ventral cochlear nucleus inanesthetized mice (xylazine and ketamine). A tungsten micro-electrode was placed on the surface of the nucleus and slowlyadvanced with a hydraulic micromanipulator. When the repeatedpresentations of search tones revealed a well-isolated auditoryunit, its best frequency (BF, the most sensitive frequency) andthreshold were determined by manually sweeping the frequencyand level of the tone bursts.The early components of the ABR have been previously linked
to the synchronized activity of globular bushy cells (GBCs),which receive end-bulb synapses from the auditory nerve (30).These excitatory synapses are among the largest in the centralnervous system (31) and presumably are specialized to ensurethe secure transmission of temporal information. The objectiveof single-unit recording was to identify GBCs and then mea-sure how their ability to encode temporal information wasaffected by early noise exposure and the elimination of MOCSfeedback.
GBCs were identified during single-unit recording by their re-sponses to short BF tone bursts (50-ms on, 200-ms off ) (32Y34).These responses were used to construct a peristimulus time his-togram (PSTH) plotting the timing of action potentials relativeto stimulus onset (32,34,35). GBCs produce a primary-notchPSTH that has a precisely timed onset response followed bya brief period of inactivity. Spherical bushy cells integrate asmaller number of end-bulb synapses and produce a primary-like PSTH with less onset precision and no notch. Multipolarcells receive auditory nerve inputs through conventional bou-ton synapses and produce a PSTH with a regularly timed‘‘chopped’’ appearance.Dys-synchronization of the auditory nerve synapse was char-
acterized by measuring how well action potentials followed dy-namic changes in the amplitude envelope of a BF tone (36,37).The envelope was sinusoidally amplitude modulated in discretefrequency steps from 50 to 1,000 Hz. The envelope-followingresponse was quantified by the synchronization coefficient (SC)(38). An SC of 1 indicates that all spike times coincided with thesame phase component of the periodic envelope. An SC of 0indicates a random distribution of spike times.
Behavioral PerformanceANSD typically manifests in adults as a difficulty in speech
comprehension (39). Because language-based testing is pro-blematic in animal subjects, our behavioral assay used gap de-tection to estimate the acuity of temporal processing. Wherea healthy listener might hear 2 sounds separated by a brief gap,a temporally impaired listener hears 1 sound. Previous studies
FIG. 2. Effects of early noise exposure on OHC function and structure. A, Frequency spectrum of a DPOAE in a noise-reared KO mouse.The emission shows a prominent cubic distortion product (DP). The signal level and noise level at the DP frequency was 28.4 andY24.4 dB SPL, respectively. B, Input-output functions relating the magnitude of DP to F2 sound levels. The average DP magnitudes (TSE)of KO mice are compared with the normal baselines of quiet-reared WT mice (bold gray line ). Numerical labels indicate the number ofsubjects in each treatment. C, Immunolabeling showing intact hair cells and olivocochlear terminals in a noise-reared KO mouse.
1571AUDITORY NEUROPATHY IN MICE
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.
confirm that ANSD patients show elevated gap detectionthresholds (40,41).Gap inhibition of the acoustic startle reflex (GIASR) is a
variant of prepulse inhibition (42), in which the offset of back-ground noise before the onset of a startle-eliciting stimulus de-creases startle magnitude. The magnitude of the inhibitory effectindicates the detectability of the silent gap between the 2 stimuli.GIASR tests were implemented with real-time digital pro-
cessors (Tucker-Davis Technologies, RP2) that monitored thesubject’s activity, presented acoustic stimuli, and recorded themagnitude of startle responses. The testing procedure was auto-mated in a MATLAB programming environment running cus-tom software (17).Behavioral testing was conducted in 70 dBA background
noise inside a sound-isolation box. The mouse was held at acalibrated sound location by a tube-shaped cage that was coupledto a movement sensor. Trials were presented at random time in-tervals, when automated monitoring of the movement sensorindicated that the mouse was sitting quietly.During gap trials, the noise was turned off just before the
presentation of the startle-eliciting stimulus. During referencetrials, the noise was not turned off. Gap inhibition was quanti-fied as the relative magnitude of the startle response on gapversus reference trials. A relative magnitude of less than 1 in-dicated the mouse startled less on gap trials, which was inter-preted as gap detection. Subjects showing statistically less gapdetection than quiet-reared WT mice were considered to testpositively for ANSD. Tests were conducted with gap durationsof 2 and 10 ms to determine the consistency of ANSD screen-ing with different degrees of task difficulty.
RESULTS
The hypothesized protective role of the MOCS wastested by rearing KO and WT mice in different levels ofbackground noise. The heavy-traffic vivarium representeda high-risk noise exposure. The low-traffic vivarium wasconsidered a low-risk quiet exposure.
As predicted, noise-reared KO mice replicated the ex-isting diagnostic criteria for ANSD. The presence of nor-mal OHC function, as verified by intact DPOAEs, was thenextended to a direct histologic assessment of hair cell sur-vival patterns. The dys-synchronization of auditory nervetransmission was demonstrated with conventional ABRprocedures and then expanded to single-unit recordings inthe ventral cochlear nucleus. Temporal coding deficits werespecific to neurons that displayed the physiologic proper-ties of GBCs. Behavioral screening procedures revealedsignificantly impaired temporal acuity among noise-rearedKO mice. The expression of the perceptual deficit washighly individualized within each treatment group.
OHC Function and StructureAll treatment groups met the diagnostic criteria for
ANSD by producing normal DPOAEs. Representativemeasures are shown for a noise-reared KO mouse inFigure 2A. A cubic distortion product is clearly evidentin this example at 10.6 kHz (2F1YF2).
Input-output functions relating the magnitude of thecubic distortion product to the level of the primary tonesare shown in Figure 2B. Results are presented for WT
mice reared in quiet and KO mice reared in quiet or noise.A statistical analysis of the effects of F2 level and treat-ment group was performed using a 2-way analysis ofvariance (ANOVA; implemented with the anova2 func-tion in MATLAB). The effect of F2 level was significant( p G 0.05), but the effect of treatment group was not.
A typical example of cochlear histology is shown inFigure 2C. The tissue was collected from the middle turnof the cochlea in a noise-reared KO mouse. Immunola-beling indicated a full complement of both inner andOHCs (myosin 7a) and MOCS terminals (synaptic vesicleprotein 2).
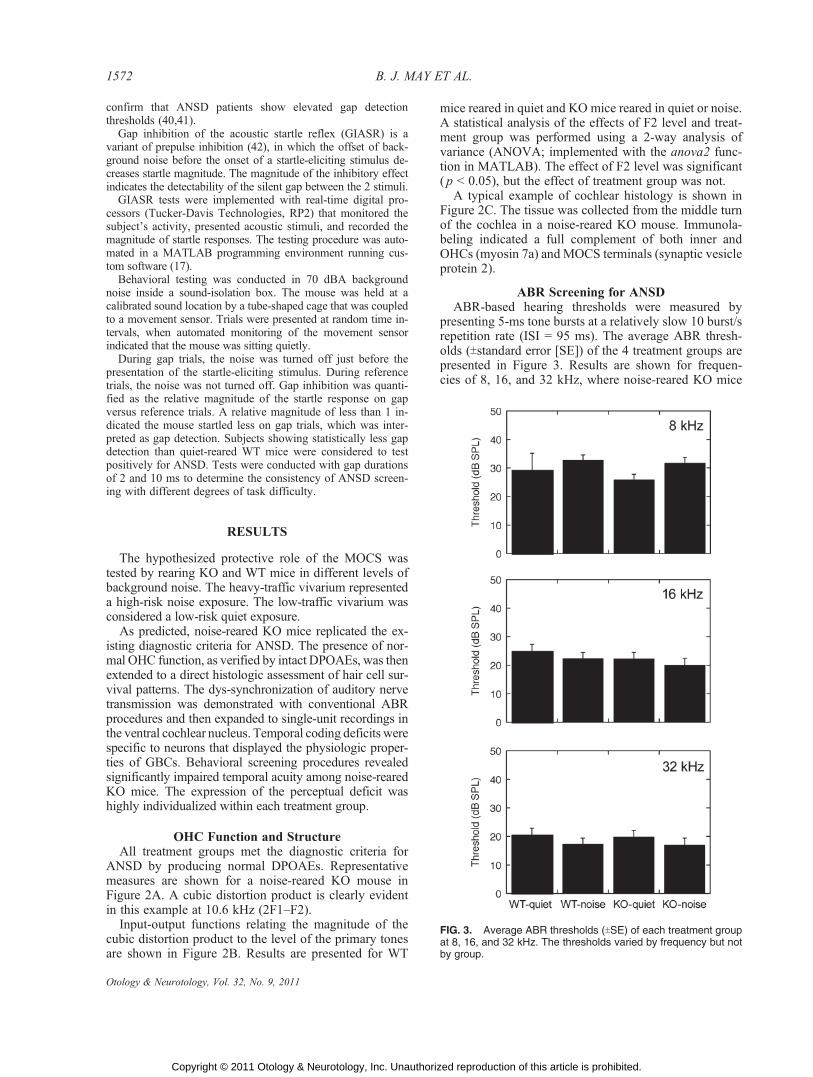
ABR Screening for ANSDABR-based hearing thresholds were measured by
presenting 5-ms tone bursts at a relatively slow 10 burst/srepetition rate (ISI = 95 ms). The average ABR thresh-olds (Tstandard error [SE]) of the 4 treatment groups arepresented in Figure 3. Results are shown for frequen-cies of 8, 16, and 32 kHz, where noise-reared KO mice
FIG. 3. Average ABR thresholds (TSE) of each treatment groupat 8, 16, and 32 kHz. The thresholds varied by frequency but notby group.
1572 B. J. MAY ET AL.
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.
showed deficits in gap detection. A statistical analysis(2-way ANOVA) indicated a significant effect of fre-quency ( p G 0.05) but no threshold difference betweentreatment groups.
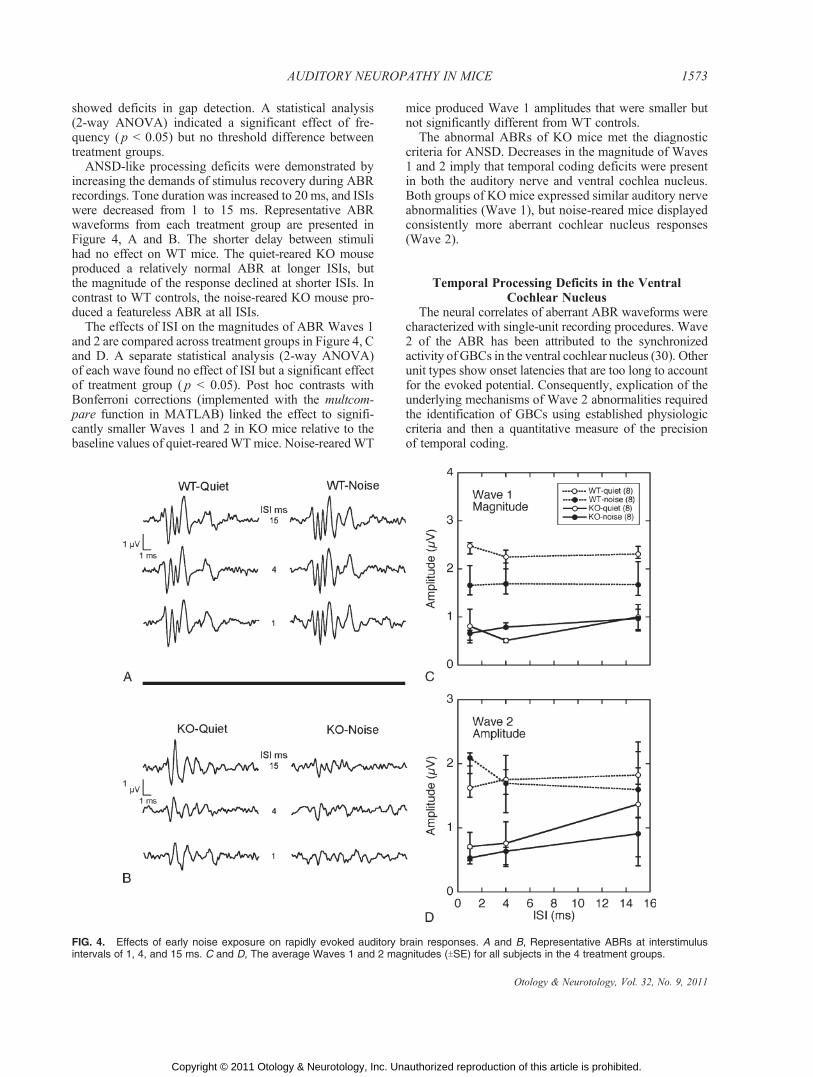
ANSD-like processing deficits were demonstrated byincreasing the demands of stimulus recovery during ABRrecordings. Tone duration was increased to 20 ms, and ISIswere decreased from 1 to 15 ms. Representative ABRwaveforms from each treatment group are presented inFigure 4, A and B. The shorter delay between stimulihad no effect on WT mice. The quiet-reared KO mouseproduced a relatively normal ABR at longer ISIs, butthe magnitude of the response declined at shorter ISIs. Incontrast to WT controls, the noise-reared KO mouse pro-duced a featureless ABR at all ISIs.
The effects of ISI on the magnitudes of ABR Waves 1and 2 are compared across treatment groups in Figure 4, Cand D. A separate statistical analysis (2-way ANOVA)of each wave found no effect of ISI but a significant effectof treatment group ( p G 0.05). Post hoc contrasts withBonferroni corrections (implemented with the multcom-pare function in MATLAB) linked the effect to signifi-cantly smaller Waves 1 and 2 in KO mice relative to thebaseline values of quiet-rearedWTmice. Noise-rearedWT
mice produced Wave 1 amplitudes that were smaller butnot significantly different from WT controls.
The abnormal ABRs of KO mice met the diagnosticcriteria for ANSD. Decreases in the magnitude of Waves1 and 2 imply that temporal coding deficits were presentin both the auditory nerve and ventral cochlea nucleus.Both groups of KO mice expressed similar auditory nerveabnormalities (Wave 1), but noise-reared mice displayedconsistently more aberrant cochlear nucleus responses(Wave 2).
Temporal Processing Deficits in the VentralCochlear Nucleus
The neural correlates of aberrant ABR waveforms werecharacterized with single-unit recording procedures. Wave2 of the ABR has been attributed to the synchronizedactivity of GBCs in the ventral cochlear nucleus (30). Otherunit types show onset latencies that are too long to accountfor the evoked potential. Consequently, explication of theunderlying mechanisms of Wave 2 abnormalities requiredthe identification of GBCs using established physiologiccriteria and then a quantitative measure of the precisionof temporal coding.
FIG. 4. Effects of early noise exposure on rapidly evoked auditory brain responses. A and B, Representative ABRs at interstimulusintervals of 1, 4, and 15 ms. C and D, The average Waves 1 and 2 magnitudes (TSE) for all subjects in the 4 treatment groups.
1573AUDITORY NEUROPATHY IN MICE
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.

Figure 5A provides examples of the major PSTH re-sponse types that were encountered in the mouse. Primary-notch neurons showed a fast first spike latency, a preciselytimed onset, and a brief refractory period (notch). Primary-like neurons produced a similar PSTH but did notdisplay a notch because their onset responses were lessprecisely timed. Chopper neurons fired a train of actionpotentials with uniform intervals between spikes, produc-ing a PSTH with a ‘‘chopped’’ appearance. Intracellularlabeling of physiologically characterized neurons has as-signed the primary-notch PSTH to GBCs, the primary-likePSTH to spherical bushy cells, and the chopper PSTH tomultipolar cells (33).
Figure 5B compares the average SC scores of KO micewith the normal baselines of WT controls. Data from 70neurons have been separated according to the physiologicresponse types in Figure 5A. Each panel indicates howwell the unit type remained synchronized to a broad rangeof envelope frequencies. The 3 functions within eachpanel describe how the quality of temporal coding wasinfluenced by noise exposure and the functional statusof the MOCS.
As in previous studies of temporal coding in the ven-tral cochlear nucleus, primary-notch and primary-likeneurons produced low-pass temporal modulation func-
tions that extended to high envelope frequencies, whereaschopper neurons produced band-pass functions that peakedat intermediate frequencies.
A statistical analysis (2-way ANOVA) of the primary-notch data set indicated a significant effect of treatmentgroup ( p G 0.05) but not modulation frequency. Post hoccontrasts noted a significant decrease in the SC scores ofnoise-reared KOmice. These results establish a direct linkbetween noise-induced ANSD and the defective end-bulb transmission of GBCs. Primary-like neurons inquiet-reared KO mice did not show the deficit. Sphericalbushy cells (primary-like PSTHs) and multipolar cells(chopper PSTHs) showed normal coding regardless oftreatment group.
Behavioral Consequences of ANSDPrevious clinical studies of ANSD have reported a
decline in gap detection (41). This finding was replicatedwith behavioral measures of GIASR in noise-reared KOmice. The distribution of relative inhibition is describedby the box plots in Figure 6, A and B. Each box showsthe interquartile range (middle 50%) of inhibition for the4 treatment groups. The line bisecting each box indi-cates the median score. Whiskers extend to the most ex-treme data points falling within T1.5 interquartile ranges.
FIG. 5. Effects of early noise exposure on single-unit activity in the ventral cochlear nucleus. A, Representative PSTHs for the 3 majorphysiologic response types. B, Temporal modulation functions showing the SC in relation to envelope frequency. Each panel indicates theaverage SC (TSE) for all neurons with the PSTH shown on the left. Results from KO mice are compared with the normal baselines of quiet-reared WT mice (bold gray line ).
1574 B. J. MAY ET AL.
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.

Data falling outside these limits are plotted as individualoutliers.
Quiet-reared WT mice showed the lowest relativestartle magnitudes (best gap detection) at both gap dura-tions. The scores of these controls also were less variable.At the opposite extreme, noise-reared KO mice showedthe highest startle magnitudes (worst gap detection) andlargest intersubject variability. Because the distributionswere highly skewed by outliers, the startle magnitudeswere ranked and analyzed with a nonparametric Kruskal-Wallis test (implemented with the kruskalwallis functionin MATLAB). A significant effect of treatment groupwas noted at both durations (p G 0.05). Post hoc contrastsrevealed a statistically significant increase in the startlemagnitudes of noise-reared KO mice relative to baselinemeasures in quiet-reared WT mice. No other contrastsattained significance.
The skewed distributions of startle responses in noise-reared KO mice suggest a heterogeneous sample, withsome subjects showing normal behavior, whereas othersmanifest highly impaired gap detection. An importantcaveat from these comparisons is the potential to under-estimate the perceptual or physiologic consequences ofANSD by mixing the results of healthy and impaired sub-jects in the same treatment group.
To refine the behavioral criteria for distinguishinghealthy and impaired mice, a new screening procedurewas derived from the limits of gap detection in quiet-reared WT controls. This approach is illustrated by thescatter plot in Figure 6C, which compares each subject’srelative startle magnitude for the 2- and 10-ms gaps. Thelargest startle score of the quiet-reared WT mice definedthe worse case scenario for normal performance alongeach dimension. Scores that exceeded these limits alongboth dimensions indicated a positive test for ANSD.Scores that failed one criterion and passed the other wereconsidered an intermediate test result.
When these diagnostic criteria were applied to the re-sults of the GIASR tests, 53% (9/17) of noise-reared KOmice tested positively for ANSD. An additional 29%(5/17) of the mice displayed intermediate deficits. Inaddition to the highest incidence of ANSD, this treat-ment group produced the largest deviations from normalbaselines. Quiet-reared KO mice yielded 35% (6/17) po-sitive test results and 35% (6/17) intermediate results.Deficits in this group were consistently smaller than thoseobserved in noise-reared KO mice. None of the noise-rearedWTmice demonstrated a convincing departure fromnormal baselines.
CONCLUSION
An animal model of ANSD was created by exposingyoung mice to environmental noise. Individual suscep-tibility to the noise exposure varied within treatmentgroups but was significantly increased among mice thatlacked a functional MOCS. The successful induction ofANSD was confirmed with the diagnostic criteria ofnormal DPOAEs and aberrant ABR waveforms that are
FIG. 6. Effects of early noise exposure on gap detection. A andB, Box plots summarizing GIASR at gap durations of 2 and 10 ms.Lower startle magnitudes indicate better gap detection (arrows).See text for a description of plotting conventions. C, Scatter plotof the same data. Behavioral criteria for ANSD are defined as adecrease in gap detection relative to the normal range of quiet-reared WT mice.
1575AUDITORY NEUROPATHY IN MICE
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.

typically used to screen human infants (2,27). Mice thattested positively for ANSD demonstrated a significantimpairment of gap detection, which is a perceptual deficitthat has been reported in clinical studies of adult ANSDpatients (41).
Single-unit recordings revealed selective defects in thetransmission of temporal information from the auditorynerve to the specialized end-bulb synapses of GBCs inthe ventral cochlear nucleus. Although these transmis-sion errors are implied by abnormalities of the ABRwaveform (43), they have not been previously describedat the neuronal level. Because GBCs provide critical in-puts to the temporal pathways of the binaural brainstem,their disordered transmission properties are likely to havea pervasive impact on speech recognition, masking re-lease, pitch perception, and sound localization (44). Thesedimensions of auditory processing are essential for theperceptual segregation of competing sound sources, whichmay explain why ANSD patients are uniquely challengedby complex listening environments (45,46).
Environmental Risk FactorsIt is well established that the functional status of the
auditory nerve must be preserved to maintain synapticdevelopment in the central auditory system. Others haveshown that deafness impedes the maturation of end-bulbsynapses in the ventral cochlear nucleus (15,47) and theirtargets in the medial nucleus of the trapezoid body (16).Our results indicate that these same developmental pro-cesses are altered when the auditory nerve is driven tounnaturally high levels of activity by background noise.In defining noise levels for the prevention of ANSD, itis important to distinguish the immediate damaging ef-fects of very loud exposures from the insidious long-termdevelopmental consequences of moderate exposures.
The complex acoustic environment of the heavy-trafficvivarium represents a conservative estimate of risk fac-tors that are posed by NICU exposures (4,5). Althoughthe suitability of this experimental manipulation was ulti-mately confirmed by its ability to induce ANSD in a highpercentage of young mice, this approach did not isolatewhich environmental factors represented the most sig-nificant risk. For example, sustained noise levels in thevivarium exceeded the Leq exposures that are recom-mended by the American Academy of Pediatrics (6). Theambient environment also contained high levels of tran-sient energy that exceeded Lmax recommendations. Atpresent, it is not possible to determine whether either typeof noise was necessary or sufficient for inducing the de-velopmental impairments.
The mice in the present study were reared from birth innoise and screened for ANSD as soon as they reached anadult age. Activity-dependent development of the cen-tral auditory system is known to begin with the onsetof hearing. Consequently, our findings can only restrictthe induction of ANSD to a period that begins as earlyas birth and persists as late as early adulthood. It is notknown whether starting the environmental exposure ata later age would interrupt ongoing developmental pro-
cesses or whether stopping the exposure before matura-tion would restore normal development. These unansweredquestions are highly relevant for the prevention and treat-ment of ANSD. The present mouse model of ANSD isan ideal experimental context for future investigations ofcritical periods for induction and recovery.
Individual SusceptibilityRelative to quiet-reared WT mice, the incidence of
ANSD was increased in both groups of KO mice. Somemice with compromised feedback maintained normalbehavior when reared in noise, whereas others developeddeficits when reared under quiet conditions. Althoughthese patterns of susceptibility suggest distinct influencesof MOCS feedback and noise exposure, they also pointout how the risk factors are amplified by a powerfulinteraction between rearing environment and geneticpredisposition.
Although the MOCS is found in all general mammalianspecies and many non-mammalian vertebrates, there is ahigh degree of anatomic variation within a species. As aresult, individuals may show substantial differences inthe strength of MOCS feedback. Presumably, the noise-reared WT mice that tested positively for ANSD devel-oped the disorder because their naturally weak MOCSfailed to provide sufficient protection. These findingsencourage the early screening of MOCS function inhuman infants to identify individuals with an increasedrisk for noise-induced ANSD. Contralateral suppressionof DPOAEs is a noninvasive method for the rapid assess-ment of efferent function that can be efficiently coordi-nated with existing infant screening protocols (48).
Underlying Mechanisms and TheirPerceptual Consequences
ANSD has been attributed to the dys-synchronizedtransmission of temporal information by the auditorynerve (49). This interpretation is largely inferred fromthe unusual pattern of DPOAE and ABR measurementsin patient populations. The introduction of adequate ani-mal models makes it possible to explore the underlyingmechanisms of the disorder with direct anatomic andphysiologic methods. Recent studies in mice have usedthis approach to confirm structural abnormalities of thespiral ganglion (24). The present study extends thoseobservations to deficiencies in central coding and theirperceptual consequences.
Abnormalities of ABR Waves 1 and 2 were demon-strated in KO mice regardless of rearing environment.However, Wave 2 showed larger deficits and less re-covery at longer ISIs when it was recorded in noise-rearedKO mice. Because this treatment group showed thehighest incidence of impaired temporal processing duringbehavioral testing, Wave 2 seems to provide a betterprediction of the perceptual consequences of ANSD. Oneinterpretation is a progressive loss of coding that firstaffects the auditory nerve (Wave 1) and then progressesto the cochlear nucleus (Wave 2). From this perspective,
1576 B. J. MAY ET AL.
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.

noise-reared KO mice exhibited larger deficits becauseANSD had reached a more advanced state.
Electrophysiologic recordings indicated that early noiseexposure distorted the neural coding of rapid envelopemodulations. Noise-induced deficits were limited to theGBCs of MOCS-compromised mice (i.e., neurons witha primary-notch PSTH). GBCs are unique because theyintegrate a large number of auditory nerve inputs viapowerful end-bulb synapses (50). Multipolar cells receivemore inputs, but they are communicated by weaker bou-ton synapses. Spherical bushy cells receive larger endbulbs, but they are fewer in number. Consequently, GBCsdisplay exceptional fidelity for the coding of stimulusonsets and envelope modulations (51,52). The selectivityof the deficit among physiologic response types impliesthat ANSD has less effect on nontemporal aspects ofauditory coding such as threshold, loudness, and frequencyselectivity.
Artificial rearing environments are known to altersound processing throughout the central auditory system.For example, an immature state persists in the lateralsuperior olive when 1 ear is deafened (16). The abnor-malities can be traced back to defective inputs from themedial nucleus of the trapezoid body and, presumably, theimpaired end bulbs of GBCs. These findings suggest thatANSD reflects a cascade of synaptic changes that beginswith the auditory nerve and eventually manifests through-out the auditory pathways. The resulting deficits may bemost apparent for the coding of temporal informationbecause small errors in the timing of action potentialsare magnified by large errors in the perceptual identity ofauditory signals.
REFERENCES
1. Dowley AC, Whitehouse WP, Mason SM, Cope Y, Grant J, GibbinKP. Auditory neuropathy: unexpectedly common in a screenednewborn population. Dev Med Child Neurol 2009;51:642Y6.
2. Berg AL, Spitzer JB, Towers HM, Bartosiewicz C, Diamond BE.Newborn hearing screening in the NICU: profile of failed auditorybrainstem response/passed otoacoustic emission. Pediatrics 2005;116:933Y8.
3. D’Agostino JA, Austin L. Auditory neuropathy: a potentially under-recognized neonatal intensive care unit sequela. Adv Neonatal Care2004;4:344Y53.
4. Busch-Vishniac IJ, West JE, Barnhill C, Hunter T, Orellana D,Chivukula R. Noise levels in Johns Hopkins Hospital. J Acoust SocAm 2005;118:3629Y45.
5. Williams AL, van Drongelen W, Lasky RE. Noise in contemporaryneonatal intensive care. J Acoust Soc Am 2007;121:2681Y90.
6. American Academy of Pediatrics. Noise: a hazard for the fetus andnewborn. Pediatrics 1997;100:724Y7.
7. Kujawa SG, Liberman MC. Acceleration of age-related hearing lossby early noise exposure: evidence of a misspent youth. J Neurosci2006;26:2115Y23.
8. Moore DR. Auditory processing disorders: acquisition and treat-ment. J Commun Disord 2007;40:295Y304.
9. Balen SA, Bretzke L, Mottecy CM, Liebel G, Boeno MR, GondimLM. Temporal resolution in children: comparing normal hearing,conductive hearing loss and auditory processing disorder. BrazJ Otorhinolaryngol 2009;75:123Y9.
10. Fuchs PA, Murrow BW. Cholinergic inhibition of short (outer) haircells of the chick’s cochlea. J Neurosci 1992;12:800Y9.
11. Liberman MC. The olivocochlear efferent bundle and suscep-tibility of the inner ear to acoustic injury. J Neurophysiol 1991;65:123Y32.
12. May BJ, McQuone SJ. Effects of bilateral olivocochlear lesionson pure-tone intensity discrimination in cats. Aud Neurosci 1995;1:385Y400.
13. Winslow RL, Sachs MB. Effect of electrical stimulation of thecrossed olivocochlear bundle on auditory nerve response to tonesin noise. J Neurophysiol 1987;57:1002Y21.
14. Hood LJ, Berlin CI, Bordelon J, Rose K. Patients with auditoryneuropathy/dys-synchrony lack efferent suppression of transientevoked otoacoustic emissions. J Am Acad Audiol 2003;14:302Y13.
15. Saada AA, Niparko JK, Ryugo DK. Morphological changes in thecochlear nucleus of congenitally deaf white cats. Brain Res 1996;736:315Y28.
16. Sanes DH. The development of synaptic function and integrationin the central auditory system. J Neurosci 1993;13:2627Y37.
17. Lauer AM, May BJ. The medial olivocochlear system attenuatesthe developmental impact of early noise exposure. J Assoc ResOtolaryngol 2011;12:329Y43.
18. Vetter DE, Liberman MC, Mann J, et al. Role of alpha9 nicotinicACh receptor subunits in the development and function of cochlearefferent innervation. Neuron 1999;23:93Y103.
19. Elgoyhen AB, Johnson DS, Boulter J, Vetter DE, Heinemann S.Alpha 9: an acetylcholine receptor with novel pharmacologicalproperties expressed in rat cochlear hair cells. Cell 1994;79:705Y15.
20. Lauer AM, May BJ, Hao ZJ, Watson J. Analysis of environmentalsound levels in modern rodent housing rooms. Lab Anim (NY) 2009;38:154Y60.
21. Di Nardo W, Ghirlanda G, Paludetti G, et al. Distortion-productotoacoustic emissions and selective sensorineural loss in IDDM.Diabetes Care 1998;21:1317Y21.
22. Parham K, Sun XM, Kim DO. Distortion product otoacousticemissions in the CBA/J mouse model of presbycusis. Hear Res1999;134:29Y38.
23. Boo NY. Current understanding of auditory neuropathy. Ann AcadMed Singapore 2008;37:60Y3.
24. Kujawa SG, Liberman MC. Adding insult to injury: cochlearnerve degeneration after ‘‘temporary’’ noise-induced hearing loss.J Neurosci 2009;29:14077Y85.
25. Si F, Brodie H, Gillespie PG, Vazquez AE, Yamoah EN. Devel-opmental assembly of transduction apparatus in chick basilarpapilla. J Neurosci 2003;23:10815Y26.
26. Simmons DD, Moulding HD, Zee D. Olivocochlear innervation ofinner and outer hair cells during postnatal maturation: an immu-nocytochemical study. Brain Res Dev Brain Res 1996;95:213Y26.
27. Berlin CI, Morlet T, Hood LJ. Auditory neuropathy/dyssynchrony:its diagnosis and management. Pediatr Clin North Am 2003;50:331Y40, vii-viii.
28. May BJ, Aleszczyk CM, Sachs MB. Single-unit recording in theventral cochlear nucleus of behaving cats. J Neurosci Methods 1991;40:155Y69.
29. May BJ, Prell GS, Sachs MB. Vowel representations in the ventralcochlear nucleus of the cat: effects of level, background noise, andbehavioral state. J Neurophysiol 1998;79:1755Y67.
30. Melcher JR, Kiang NY. Generators of the brainstem auditoryevoked potential in cat. III: Identified cell populations. Hear Res1996;93:52Y71.
31. Ryugo DK, Fekete DM. Morphology of primary axosomatic end-ings in the anteroventral cochlear nucleus of the cat: a study of theendbulbs of Held. J Comp Neurol 1982;210:239Y57.
32. Blackburn CC, Sachs MB. Classification of unit types in the ante-roventral cochlear nucleus: PST histograms and regularity analysis.J Neurophysiol 1989;62:1303Y29.
33. Rhode WS, Oertel D, Smith PH. Physiological response propertiesof cells labeled intracellularly with horseradish peroxidase in catventral cochlear nucleus. J Comp Neurol 1983;213:448Y63.
34. Pfeiffer RR. Anteroventral cochlear nucleus: wave forms of extra-cellularly recorded spike potentials. Science 1966;154:667Y8.
35. Winter IM, Palmer AR. Responses of single units in the ante-roventral cochlear nucleus of the guinea pig. Hear Res 1990;44:161Y78.
1577AUDITORY NEUROPATHY IN MICE
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.

36. Palmer AR. Encoding of rapid amplitude fluctuations by cochlear-nerve fibres in the guinea-pig. Arch Otorhinolaryngol 1982;236:197Y202.
37. Rhode WS, Greenberg S. Encoding of amplitude modulationin the cochlear nucleus of the cat. J Neurophysiol 1994;71:1797Y825.
38. Goldberg JM, Brown PB. Response of binaural neurons of dogsuperior olivary complex to dichotic tonal stimuli: some physiolo-gical mechanisms of sound localization. J Neurophysiol 1969;32:613Y36.
39. Zeng FG, Oba S, Garde S, Sininger Y, Starr A. Temporal and speechprocessing deficits in auditory neuropathy. Neuroreport 1999;10:3429Y35.
40. Michalewski HJ, Starr A, Nguyen TT, Kong YY, Zeng FG. Audi-tory temporal processes in normal-hearing individuals and inpatients with auditory neuropathy. Clin Neurophysiol 2005;116:669Y80.
41. Yalcinkaya F, Muluk NB, Atas A, Keith RW. Random gap detec-tion test and random gap detection test-expanded results in childrenwith auditory neuropathy. Int J Pediatr Otorhinolaryngol 2009;73:1558Y63.
42. Hoffman HS, Fleshler M. Startle reaction: modification by back-ground acoustic stimulation. Science 1963;141:928Y30.
43. El-Badry MM, McFadden SL. Evaluation of inner hair cell andnerve fiber loss as sufficient pathologies underlying auditory neu-ropathy. Hear Res 2009;255:84Y90.
44. Zeng FG, Kong YY, Michalewski HJ, Starr A. Perceptual con-sequences of disrupted auditory nerve activity. J Neurophysiol2005;93:3050Y63.
45. Kraus N, Bradlow AR, Cheatham MA, et al. Consequences ofneural asynchrony: a case of auditory neuropathy. J Assoc ResOtolaryngol 2000;1:33Y45.
46. Narne VK, Vanaja CS. Perception of envelope-enhanced speechin the presence of noise by individuals with auditory neuropathy.Ear Hear 2009;30:136Y42.
47. Redd EE, Pongstaporn T, Ryugo DK. The effects of congenitaldeafness on auditory nerve synapses and globular bushy cells incats. Hear Res 2000;147:160Y74.
48. James AL. The assessment of olivocochlear function in neonateswith real-time distortion product otoacoustic emissions. Laryngo-scope 2011;121:202Y13.
49. Hood LJ. A review of objective methods of evaluating auditoryneural pathways. Laryngoscope 1999;109:1745Y8.
50. Ostapoff EM, Morest DK. Synaptic organization of globular bushycells in the ventral cochlear nucleus of the cat: a quantitative study.J Comp Neurol 1991;314:598Y613.
51. Joris PX, Carney LH, Smith PH, Yin TC. Enhancement of neuralsynchronization in the anteroventral cochlear nucleus. I. Responsesto tones at the characteristic frequency. J Neurophysiol 1994;71:1022Y36.
52. Rhode WS. Response patterns to sound associated with labeledglobular/bushy cells in cat. Neuroscience 2008;154:87Y98.
1578 B. J. MAY ET AL.
Otology & Neurotology, Vol. 32, No. 9, 2011
Copyright © 2011 Otology & Neurotology, Inc. Unauthorized reproduction of this article is prohibited.