importance of plant resistance to insect and mite...
TRANSCRIPT

Importance of Plant Resistance to Insect and Mite Vectors in Controlling Virus Diseases of Plants: Resistance
to the Wheat Curl Mite (Acari: Eriophyidae)l, 2
T, L. Harvey, T. J. Martin, and D. L. Seifers
Kansas State University Hays, KS 67601 USA
J. Agric. Entomol. 11(3): 271-277 (July 1994) ABSTRACT The importance of host plant resistance to insect and mite vectors in controlling virus diseases of plants is not well established, but 13 examples of resistance to aphids, leafhoppers, planthoppers, and onc mite species that have reduced the incidence of virus diseases in the field are cited. In Kansas, an estimated 97% of the acreage of the five major crops (wheat, sorghum, corn, soybeans, alfalfa) is resistant to one or more diseases compared with 31% resistant to insects and 14% resistant to a mite vector. Resistance to the wheat curl mite, Eriophyes tulipae Keifer, has been reported from wheat relatives in the genera Thinopyrum (Agropyron), Aegilops, and Secale and most recently in common wheat, Triticum aesliuum L. So far, only the resistance from Secale (rye) is being utilized commercially. The rye source of resistance is derived from the IRJIA translocation line Amigo. This resistance is a major component of an integrated program to control wheat streak mosaic, which was the most important plant disease in Kansas from 1987 to 1991. Resistance to the wheat curl mite reduced the incidence of wheat streak mosaic in wheat fields by an average of 76% from 1979 to 1988. Although resistance to the wheat curl mite illustrates the potential for using resistance to vectors to control virus diseases, it may not be typical of most viruses that are spread by aphids and leafhoppers. More research is needed to clarify the importance of the resistance already available for many insect species that are vectors of plant viruses.
KEY WORDS Acari, Eriophyidae, Eriophyes lulipae, wheat curl mite, resistance, virus, vector.
The value of plant resistance to insect and mite vectors to control plant viruses is not well established and more research is needed to confirm its importance. Kennedy (1976) and Maramorosch (1980) reviewed the subject from both the theoretical and applied standpoints. Kennedy (1976) found relatively few documented instances where resistance to an insect vector has contributed to disease control. Gibson and Plumb (1977) stated that little information exists about
I Contribution No. 93-400-J from the Kansas Agricultural Station. 2 Received for publication 23 June 1993; accepted 15 February 1994.
271

272 J. Agl·ic. Entomol. Vol. 11, No.3 (1994)
the effects of aphid resistance on the spread of virus diseases. Heinrichs (1979) reviewed the effects of resistance to leafhoppers and planthoppers on virus transmission and concluded that very few data are available on the value of host plant resistance to a vector in the absence of virus resistance in rice.
Plant breeders understandably prefer the morc direct approach of developing cultivars resistant to viruses rather than to vectors. Virus control has seldom, if ever, been the aim of a breeding program for aphid resistance (Maramorosch 1980), but breeding for mile resistance to control a virus disease of wheat (Martin et a1. 1983) and breeding for resistance to planthoppers to control virus diseases of rice (Heinrichs 1986) have been attempted with some success. However, when resistance was obtained, it was a byproduct of a program aimed at the direct damage caused by an insect which was also a disease vector. The percentages of the acreage or cultivars of the five major crops grown in Kansas during 1991-1992 that are resistant to diseases, insects, and a mite vector of a plant virus are shown in Table L Nearly all of the acreage is resistant to one 01'
more diseases, and about one third of the acreage is resistant to insects. The insects controlled by resistance in sorghum (greenbug, Schizaphis graminum [Rondani)) and alfalfa (pea aphid, Acyrthosipho/l. pisllm [Harris)) are vectors of plant viruses, but the resistance was developed to control the direct damage caused by the insects. The effect of resistance on transmission of maize dwarf mosaic virus to sorghum and alfalfa mosaic virus to alfalfa is unknown. Resistance in wheat (23% of the hectarage in 1991) to wheat curl mite, Eriophyes tulipae Keifer, is effective in limiting the spread of wheat streak mosaic virus (WSMV). However, resistance to the wheat curl mite was not a breeding objective during the development of resistant cultivars. For example, the cultivar 'TAM 107' was developed for resistance to biotype C of the gl'cenbug (Porter et aL 1987) utilizing resistance from 'Amigo' which has the TRiIA tl'anslocation from rye (Sebesta and Wood 1978), which incidentally
Table 1. Resistance of Kansas crops to diseases, insects, and a virus vector.
Hectares Percent of hectares or cultivars resistant Crops
Millions Diseases Insects Vector
Wheal 4.9 98 20 23 Sorghum 1.2 90 65 a
Corn 0.8 99 50 0 Soybean 0.8 95 0 0 Alfalfa 0.4 99 93 a
All Crops" 8.1 97 31 14
" Most of the sorghum and alfalfa hectaragc is rcsistnnt to aphids that arc virus vClCLors, but the effect of this resistnncc on control of the viruses transmitted j~ unknown.
II Percent ofhectarcs resistant based on resistant hectares divided by total hectares.

273 HARVEY et al.: Resistance to Vectors for Virus Control
provided resistance to the wheat curl mite. Resistance to the wheat curl mite and its effectiveness in controlling the spread of WSMV will be discussed in more detail later.
Resistance to insect and mite vectors has reduced the incidence of plant virus diseases in the field in at least 13 cases (Table 2). All references ploior to 1976 were cited by Kennedy (1976). This list of 13 examples is not impressive, considering that hundreds of species of insects, mostly aphids and leaOlOppers, transmit hundreds of different viruses and that plant resistance to aphids and leaOloppers is more common than to other groups of insects. Nevertheless, these examples provide encouragement for more research in this area. The incentive to use resistance to insect and mite vectors to control virus diseases of plants is strongest when an important disease affects a major crop and resistance is more readily available for the vector than for the virus.
Table 2. Examples of vector resistance that reduced disease incidence in the field.
Crops Vectors Viruses References
Raspberry Aphid Raspberry Mosaic Harris 1935
Peanut Aphid Rosette Evans 1954
Red Clover Aphid Clover Wilcoxon & Peterson 1960
Sugar Beet Aphid Beet Yellows Hills et al. 1969
Cranberry Leafhopper False-blossom Wilcox & Beckwith 1933
Lettuce Leafhopper Aster Yellows Linn 1940
Tomato Leafhopper Curly Top Thomas & Martin 1971
Rice Leafhopper Tungro Pathak 1970
Rice Planthopper Hoja Blanca Jennings & Pineda 1970
Rice Planthopper Grassy Stunt Pathak 1970
Rice Planthopper Ragged Stunt Heinrichs 1986
Rice Planthopper Wilted Stunt Heinrichs 1986
Wheat Mite Wheat Streak Mosaic Martin et al. 1984
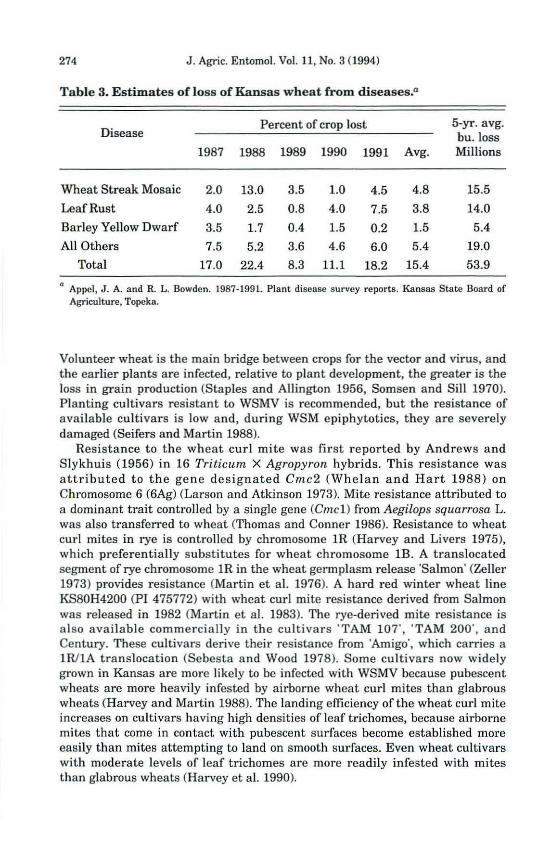
Resistance to the wheat curl mite is an example of "esistance to a vector that contributes to the control of a plant virus. The wheat curl mite is the only vector of WSMV (Slykhuis 1955), which caused the most important disease of wheat in Kansas for the 5-yr period 1987-91 (Table 3). The average annual loss to WSMV during that time was 5.5 million hectoliters 05.5 million bushels). An epiphytotic in 1987-88 resulted in a 13% crop loss, estimated at $150 million, indicating a need for improvement in \VSMV control methods. Control is primarily cultural, elimination of volunteer wheat and delay in planting.
274 J. Agric. Entomol. Vol. 11. No.3 (l994)
Table 3. Estimates of loss of Kansas wheat from diseases.a
Percent of crop lost 5-yr. avg. Disease bu. loss
1987 1988 1989 1990 1991 Avg. Millions
Wheat Streak Mosaic 2.0 13.0 3.5 1.0 4.5 4.8 15.5
Leaf Rust 4.0 2.5 0.8 4.0 7.5 3.8 14.0
Barley Yellow Dwarf 3.5 1.7 0.4 1.5 0.2 1.5 5.4
All Others 7.5 5.2 3.6 4.6 6.0 5.4 19.0
Total 17.0 22.4 8.3 11.1 18.2 15.4 53.9
<I Appel. J. A. and R. L. Bowden. 1987-1991. Plant disease survey reports. Kansas State Board of Agriculture, Topeka.
Volunteer wheat is the main bridge between crops for the vector and virus, and the earlier plants are infected, relative to plant development. the greater is the loss in grain production (Staples and Allington 1956, Somsen and Sill 1970). Planting cultivars resistant to W8MV is recommended, but the resistance of available cultivars is low and, during WSM epiphytotics, they are severely damaged (Seifers and Martin 1988).
Resistance to the wheat curl mite was first reported by Andrews and Slykhuis (1956) in 16 Triticum X Agropyron hybrids. This resistance was attributed to the gene designated Cmc2 (Whelan and Hart 1988) on Chromosome 6 (6Ag) (Larson and Atkinson 1973). Mite resistance attributed to a dominant trait controlled by a single gene (Cm.c!) from Aegilops squarrosa L. was also transferred to wheat (Thomas and Conner 1986). Resistance to wheat curl mites in rye is controlled by chromosome lR (Harvey and Livers 1975), which preferentially substitutes for wheat chromosome lB. A translocated segment of rye chromosome lR in the wheat germplasm release 'Salmon' (Zeller 1973) provides resistance (Martin et aL 1976). A hard red winter wheat line KS80H4200 (PI 475772) with wheat curl mite resistance derived from Salmon was released in 1982 (Martin et aL 1983). The rye-derived mite resistance is also available commercially in the cultivars 'TAlvI 107', 'TAM 200', and Century. These cultivars derive their resistance [rom 'Amigo', which carries a lR/lA translocation (Sebesta and Wood 1978). Some cultivars now widely grown in Kansas are more likely to be infected with WSMV because pubescent wheats are more heavily infested by airborne wheat curl mites than glabrous wheats (Harvey and Martin 1988). The landing efficiency of the wheat curl mite increases on cultivars having high densities of leaf trichomes, because airborne mites that come in contact with pubescent surfaces become established more easily than mites attempting to land on smooth surfaces. Even wheat cultivars with moderate levels of leaf trichomes are more readily infested with mites than glabrous wheats (Harvey et al. 1990).

275 HARVEY et al.: Resistance to Vectors for Virus Control
Resistance to wheat curl mites in common wheat, other than that associated with glabrous leaves, was first discovered in eight introductions (PIs 221699, 222651, 222655, 222661, 222679, 222680, 222682, and Cl9355) that were previously selected for resistance to Russian wheat aphid, Diuraphis noxia (Mordvilko) (Harvey and Martin 1992). The effectiveness of those new sources of resistance from common wheat in reducing the incidence of WSM in the field has not been determined. However, resistance to the wheat curl mite from distant relatives of wheat (Secale and Aegilops) effectively reduced the incidence of WSM in the field (Martin et al. 1984, Harvey and Martin 1988, Harvey et al. 1990, Conner et al. 1991).
The first evidence that resistance to wheat curl mite reduced WSM in the field was obtained in 1979 (Table 4). During 1979, 1981, and 1982, the rye SaUTee of mite resistance from 'Salmon' had WSM infection rates of 9, S, and 21%, respectively, compared with 32, 14, and 52% for the susceptible cultivar 'Sage'. From 1986 to 1988, the rye source of resistance in 'TAM 107' had infection rates of 4, 12, and 4% compared with 25, 33, and 70% for the susceptible cultivars 'TAM lOS' and 'Larned'. During that 6-yr period, mite resistant cultivars had a 76% lower rate ofWSM infection than mite susceptible cultivars (Table 4). The most recent evidence of WSM control by resistance to wheat curl mite was reported from Canada. Following a light natural infestation of wheat curl mite, lines utilizing a source of resistance (CmcI) from Aegilops squarrosa L. had 40% fewer WSMV-infected plants than lines without mite resistance (Conner et al. 1991).
Wheat curl mite resistance provides an outstanding example of the importance of resistance to a vector in controlling a plant virus. However, only three viruses are known to be vectored by eriophyid mites (Slykhuis 1980). The
Table 4. Control of wheat streak mosaic (WSM) by resistance to tbe wheat curl mite (WCM).
Incidence of WSM (% of plants infected) Type of wheat 1979 1981 1982 1986 1987 1988 Avg.
Resistanta 9 5 21 4 12 4 9.2
Susceptibleb 32 14 52 25 33 70 37.7
a 19790; 'Salmon', 1980, 1981 '" KSBOH4200, 1986-1988", 'TAM 107'.
b 1979 _ 1982 = 'Sage', 1986 = 'TMI 105', 1987-1988 ='Lamed'.

276 J. Agric. Entomol. Vol. 11, No.3 (1994)
effectiveness of resistance to the wheat curl mite in control of WSMV may not be comparable to the control from resistance to insects, which vector most of the plant viruses. Resistance to the mite vector is a major tactic in the integrated control of some diseases such as WSM, but more research is needed to evaluate the importance of resistance to vectors to control the vast majority of virus diseases of plants that are vectored by aphids and leafhoppers.
References Cited
Andrews, J. E. & J. T. Slykhuis. 1956. Reaction of winter wheat varieties and Triticum X Agropyron hybrids when inoculated with streak mosaic virus by the mite vector, Aceria tulipae. Plant Dis. Rep. 40: 513·516.
Conner, R. L., J. B. Thomas & E. D. P. Whelan. 1991. Comparison of mile resistance for control of wheat streak mosaic. Crop Sci. 31: 315-318.
Evans, A C. 1954. Rosette disease of groundnuts. Nature (Lond.) 173: 1242. Gibson, R. W. & R. T. Plumb. 1977. Aphid resistance and the spread of viruses, pp. 488~
491. In K. F. Harris and K Mararnorosch [cds.J, Aphids as virus vectors. Academic Press, New York, 559 pp.
Harris, R. V. 1935. Some observations on the raspberry disease situation in North America. East Malling Res. Stn. Rept.. 17: 156-164.
Harvey, T. L. & R. W. Livers. 1975. Resistance to wheat curl mite, Aceria tulipae Keifer, in rye and wheat-rye addition lines. Environ. Entomol. 4: 523-526.
Harvey, T. L. & T. J. Martin. 1988. Sticky-tape method to measure cultivar effect on wheat curl mite (Acari: Eriophyidae) populations in wheat spikes. J. Econ. Entomol. 81: 731-734.
1992. Resistance to the wheat curl mite (Acari: Eriophyidael in common wheat. Cereal Res. Commun. 20: 63-66.
Harvey, T. L., T. J. Martin & D. L. Seifers. 1990. Wheat curl mite and wheat streak mosaic in moderate trichome density wheat cuHivars. Crop Sci. 30: 534~536.
Heinrichs, E. A. 1979. Control of leafhopper and planthopper vectors of rice viruses, pp. 545·551. In K Maramorosch and K. F. Harris [cds.], Leafhopper veclors and plant disease agents. Academic Press, New York, 654 pp.
1986. Perspectives and directions for the continued development of insect~resistantrice varieties. Agric. Ecosyst. Environ. 18: 9~36.
Hills, F. J., W. H. Lange & J. Kisbiyama. 1969. Varietal resistance to yellows. Vector control and planting dale as factors in the suppression of yellows mosaic of sugar beet. Phytopathology 59: 1728~ 1731.
Jennings, P. R. & A. T. Pineda. 1970. Screening rice for resistance to the planthopper, Sogatodes orizicola (Muir). Crop Sci. 10; 687-689.
Kennedy, G . G. 1976. Host plant resistance and the spread of plant viruses. Environ. Entornol. 5: 827-832.
Larson, R. 1. & T. G. Atkinson. 1973. Wheat~Ag,.opyronchromosome substitution lines as sources of resistance to wheat streak mosaic virus and its vector Aceria tulipae, pp. 173-177. III Proceedings, 4th International Wheat Genetics Symposium, Aug. 6-11, 1973, University of Missouri, Columbia, 955 pp.
Linn, M. B. 1940. The yellows disease of let.tuce and endive. New York State Agricultural Experiment Station (Bulletin 742), Ithaca, 34 pp.
Maramoro6cb, K. 1980. Breeding for resistance to vectors, pp. 146·153. III F. G. Maxwell, and P. R. Jennings [eds.) Breeding plants resistant to insects. John Wiley and Sons, New York, 683 pp.
Martin, T. J., T. K. Harvey & R. W. Livers. 1976. Resistance to wheat streak mosaic virus and its vector, Aceria tulipae. Phytopathology 66: 346-349.

277 HARVEY et al.: Resistance to Vectors for Virus Control
Martin, T. J., T. L. Harvey, C. G. Bender, D. L. Seifers & J. H. Hatchett. 1983. Wheat curl mite resistant wheat germplasm. Crop Sci. 23: 809.
Martin, T. J., T. L. Harvey, C. G. Bender & D. L. Seifers. 1984. Control of wheat streak mosaic virus with vector resistance in wheat. Phylopathology 74: 963-964.
Pathak, M. D. 1970. Genetics of plants in pest management, pp. 138·157. In R. L. Rabb and F. E. Guthrie [eds.1 Concepts of pest management. North Carolina State University, Raleigh, 472 pp.
Porter, K. B., W. D. Worran, J. H. Gardenhire, E. C. Gilmore, M. E. McDaniel & N. A. TuUeen.1987. Registration of 'TAM lOT wheat. Crop Sci. 27: 818·819.
Sebesta, E. E. & E. A. Wood, Jr_ 1978. Transfer of greenbug resistance from rye to wheat with x-rays, pp. 61-62. /n Agronomy abstracts. American Society of Agronomy, Madison, WI, 205 pp.
Seifers, D. L. & T. J_ Martin. 1988. Correlation of low level wheat streak mosaic virus resistance in Triumph 64 wheat with low virus titer. Phytopathogy 78: 703-707.
Slykhuis, J. T. 1955. Aceria lulipae (Acarina: Eriophyidae) in relation to the spread of wheat streak mosaic. Phytopathology 45: 116-128
1980. Mites, pp. 325-356. /11 F. G. Maxwell, and P. R. Jennings (eds.J Breeding plants resistant to insects. John Wiley and Sons, New York, 693 pp.
Somsen, H. W. & W. R. Sill, Jr. 1970. The wheat curl mite, Aceria tulipae Keifer, in relation to epidemiology and control of wheat streak mosaic. Kans. Agric. Exp. Stn. Res. Pub!. 162: 1-24.
Staples, R. & W. B. Allington, 1956. Streak mosaic of wheat in Nebraska and its control. Univ. of Nebraska College of Agricultural Research Bulletin 178, Lincoln, 40 pp.
Thomas, J. B. & R. L. Conner. 1986. Resistance to colonization by the wheat curl mite in Aegilops squarrosa and its inhm'ilance after transfer to common wheat. Crop Sci. 26: 527-530.
Thomus, P. E., and M. W. Murtin. 1971. Vector preference, a factor of resistance to curly top virus in certain tomato cultivars. Phytopatholob'Y 61: 1257·1260.
Whelan, E. D. P., and G. E. Hart. 1988. A spontaneous translocation that transfers wheat curl mite resistance rrom decaploid Agropyron ehmgal,wn to common wheat. Genome 30: 289-292.
Wilcox, R. B., and C. S. Beckwith. 1933. A factor in the varietal resistance of" cranberries to the false-bloom disease. J. Agric. Res. 47: 583-590.
Wilcoxon, R. D. and A. G. Peterson. 1960. Resistance in Dollard red clover to the pea aphid Mac:ro.'liphum. pisi. J. Econ. Entomo!. 53: 863-865.
Zeller, F. J. 1973. IB/lR wheat-rye chromosome substitutions and translocations, pp. 209-222. /11 Proceedings, 4th International Wheat Genetics Symposium, Aug. 6·11, L973, University ofl\'1issouri, Columbia, 955 pp.