importancia de las playas arenosas en el reciclaje de la
TRANSCRIPT

Importancia de las playas arenosas en el reciclaje de la materia orgánica
Begoña Araújo de Miguel
TESIS DOCTORAL
2017


Escola Internacional de Doutoramento
Begoña Araújo de Miguel
TESE DE DOUTORAMENTO
Importancia de las playas arenosas en el reciclaje de la materia orgánica
Dirixida polos doutores:
Mariano Lastra Valdor
Jesús López Pérez
Ano:
2017

Contenido CAPÍTULO 1. INTRODUCCIÓN GENERAL…………..………...4
ANTECEDENTES EN EL ESTUDIO DE LAS PLAYAS ARENOSAS .... 6
DEFINICIÓN DE PLAYA .................................................................. 7
DESCRIPCIÓN DE LA PLAYA ARENOSA ........................................ 7
Ambiente físico ................................................................................................. 8
Morfología y características del oleaje ................................................................. 8
Morfodinamismo ................................................................................................. 10
Grado de exposición al oleaje .............................................................................. 12
Efecto de la marea ............................................................................................... 13
ECOLOGÍA DE LA PLAYA ARENOSA ............................................ 14
Intercambios con otros ecosistemas ............................................................. 15
Cadenas tróficas .............................................................................................. 17
Cadena trófica macroscópica ............................................................................... 17
Cadena trófica intersticial .................................................................................... 17
Cadena trófica microbiana .................................................................................. 18
Flujos de energía ............................................................................................ 19
Macrofauna ..................................................................................................... 20
Teorías ecológicas ................................................................................................ 21
WRACK ......................................................................................... 23
¿Qué es? .......................................................................................................... 23
Rol e importancia ........................................................................................... 23
PLAYAS EN PELIGRO ................................................................... 26
Calentamiento global y subida del nivel del mar .......................................... 26
OBJETIVOS ................................................................................... 27
BIBLIOGRAFÍA ............................................................................. 28
CAPÍTULO 2 VARIACIÓN ESTACIONAL DE LA ACTIVIDAD
METABÓLICA DE LA PLAYA…………………………………………..38
INTRODUCCIÓN ........................................................................... 36
MATERIAL Y MÉTODOS ............................................................... 38
Área de estudio ........................................................................................... 38
Caracterización de las playas ................................................................. 40
Diseño del muestreo ................................................................................. 40
Toma de las muestras..................................................................................... 42
Análisis de las muestras ................................................................................. 43
Análisis estadístico ......................................................................................... 43

RESULTADOS ............................................................................... 44
Caracterización de las playas ......................................................................... 44
Parámetros físicos .......................................................................................... 47
Macrofauna ..................................................................................................... 48
Efecto de la biomasa algal ................................................................................... 52
Biomasa algal .................................................................................................. 52
Flujo de CO2 .................................................................................................... 55
Efecto de los parámetros físicos y biológicos ..................................................... 57
Nutrientes: TNI y fosfato ............................................................................... 60
Nivel inferior de marea (N1) ............................................................................... 60
Niveles superiores de marea (N2 y N3) .............................................................. 60
Efecto de los parámetros físicos y biológicos ..................................................... 65
Carbono biopolimérico ............................................................................ 67
Efecto de los parámetros físicos y biológicos ...................................................... 71
DISCUSIÓN ................................................................................... 73
Efecto de la biomasa algal sobre la macrofauna .............................. 73
Flujo de CO2 y nutrientes ........................................................................ 74
Efectos de los parámetros físicos ........................................................................ 74
Efecto de la biomasa algal ................................................................................... 76
Efecto de la macrofauna ...................................................................................... 77
Carbono biopolimérico .................................................................................. 78
CONCLUSIONES ........................................................................... 78
BIBLIOGRAFÍA ............................................................................. 80
CAPÍTULO 3 VARIACIÓN ESPACIAL DE LA ACTIVIDAD
METABÓLICA DE LA PLAYA
PARTE I VARIACIÓN DIARIA…………………………………………….88
INTRODUCCIÓN ........................................................................... 88
MATERIAL Y MÉTODOS ............................................................... 89
Área de estudio ............................................................................................... 89
Diseño del muestreo ....................................................................................... 90
Toma de las muestras..................................................................................... 90
Análisis de las muestras ................................................................................. 91
Análisis estadístico ......................................................................................... 92
RESULTADOS ............................................................................... 92
Variación diaria del CO2 ................................................................................. 92
Efecto de la temperatura ..................................................................................... 92
Efecto de la biomasa algal y el tipo alga ............................................................. 95

Efecto de la macrofauna ...................................................................................... 95
“Edad” de la materia orgánica ....................................................... 96
Efecto de la biomasa del parche de algas ........................................................... 96
Liberación de nitrógeno inorgánico .............................................................. 97
Efecto de la biomasa algal y tipo de alga ............................................................ 97
Calidad de la materia orgánica .....................................................................101
DISCUSIÓN ................................................................................. 102
Variación diaria del CO2 ............................................................................... 102
Efecto de la temperatura ................................................................................... 102
Efectos del tipo de alga, biomasa del parche de algas y macrofauna ............. 103
Tipo de alga y calidad de la materia orgánica ............................................ 103
Efecto de la biomasa del parche de algas ......................................................... 103
Liberación de nutrientes .............................................................................. 105
Efecto del tipo de alga y biomasa del parche ................................................... 105
CONCLUSIONES ......................................................................... 107
PARTE II VARIACIÓN ESPACIAL………………………………………115
BIBLIOGRAFÍA ........................................................................... 107
INTRODUCCIÓN .......................................................................... 115
MATERIAL Y MÉTODOS .............................................................. 116
Área de estudio .............................................................................................. 116
Diseño del muestreo ................................................................................ 119
Toma de las muestras................................................................................... 120
Análisis de las muestras ............................................................................... 120
Análisis estadístico ........................................................................................ 121
RESULTADOS .............................................................................. 121
Caracterización de las playas ........................................................................ 121
Parámetros físico-ambientales y biológicos ..................................................... 122
Macrofauna ..................................................................................................... 126
Flujo de CO2 .................................................................................................. 126
Efecto de la biomasa algal y el tipo alga ........................................................... 126
Efecto de la macrofauna .................................................................................... 130
Liberación de nitrógeno inorgánico y fósforo ............................................. 132
Efecto de la macrofauna .................................................................................... 135
Carbono biopolimérico y ―edad‖ del alga .................................................... 136
Efectos sobre la macrofauna ............................................................................. 138
DISCUSIÓN ................................................................................. 142
Parámetros físico-ambientales y biológicos................................................ 142
Efectos de la biomasa algal ............................................................................... 143
Efectos de la macrofauna ........................................................................... 143

Flujo de CO2 .................................................................................................. 145
Liberación de nitrógeno inorgánico y fósforo ............................................. 145
Carbono biopolimérico ................................................................................ 146
CONCLUSIONES GENERALES ..................................................... 147
BIBLIOGRAFÍA ........................................................................... 148


6
El objetivo de este capítulo es proporcionar una visión general de los principales temas, incluyendo
el conocimiento actual y la importancia (como lo conocemos) de los depósitos de macroalgas (wrack) en las
playas arenosas y en los ecosistemas costeros. Por lo tanto, tiene como objetivo proporcionar una
introducción al wrack y su papel en el ecosistema de la playa arenosa, antecedentes del estudio y el motivo
de esta investigación. También se identifican las preguntas claves de la investigación.
ANTECEDENTES EN EL ESTUDIO DE LAS PLAYAS
ARENOSAS
El fuerte hidrodinamismo de estos hábitats hace que los sedimentos de las playas arenosas no estén
consolidados y frecuentemente son perturbados por las olas, haciéndolos inadecuados para la fijación de
macrófitas (macroalgas y pastos marinos). Por lo tanto, la productividad primaria in situ es baja (Brown &
McLachlan, 2002) lo que determina su estructura ecológica. Precisamente este rasgo característico es el
responsable de que la comunidad científica haya ido retrasando el interés por su estudio debido a la falsa
creencia de que las playas fuesen desiertos de arena por la aparente ausencia de fauna. Comparando con
otros sistemas marinos y costeros, la ecología de playas es una disciplina muy reciente (Schlacher et al.,
2007). Los primeros escritos sobre las playas datan de comienzos del siglo pasado; unos pocos visionarios
escribieron los primeros documentos como Bruce (1928). A partir de este momento, se comienzan a
estudiar las playas: Stephen (Inglaterra, 1931), Remane (Escocia, 1933) en el Mar del Norte y Pearse et al.,
(1942) en los Estados Unidos que representó el primer intento de aproximación cualitativa del ecosistema
playa. En los años 50 comienzan los primeros trabajos descriptivos sobre la ecología de las playas de arena
como los de Dhal (1952) y Hedgpeth (1957). En los años 70, dos investigaciones demostraron que las playas
no eran grandes desiertos como había creído hasta el momento ya que registraron valores de biomasa de
macrofauna de 25753 g peso seco m-1 en Perú (Penchaszadeh, 1971) y 6621 g peso seco m-1 en el Cabo del
Este (McLachlan, 1977). No fue hasta el primer simposio internacional celebrado en Sudáfrica en 1983
(McLachlan & Erasmus, 1983) donde la ecología de playas surge por primera vez como un nuevo campo de
la ciencia costera. En ese momento, ambos autores llevaron a cabo una revisión sobre la ecología de playas
arenosas. Este hecho fue el que impulsó la investigación ecológica en las playas. En la década de los 90,
Brown & McLachlan (1990) revisan todos los trabajos realizados hasta momento. A partir de aquí,
numerosas investigaciones se han llevado a cabo demostrando que a pesar de su apariencia estéril, las

7
playas albergan una comunidad que parece invisible pero lo cierto es que está repleta de vida microscópica
y macroscópica. Todos estos componentes interactúan en una red trófica para crear el ecosistema de la
playa de arena, que intercambia materiales con el mar abierto y la tierra. Con el tiempo se ha ido
demostrando que las playas son ecosistemas interesantes y a menudo productivos. Y así, afortunadamente,
su simplicidad relativa proporciona un escenario propicio para la investigación.
DEFINICIÓN DE PLAYA
Las zonas costeras incluyen una amplia variedad de sistemas como playas rocosas, acantilados,
humedales y playas arenosas (Burke et al., 2001; Martínez et al., 2007). Existen varias definiciones
aceptadas; de acuerdo con Brown & McLachlan (1990): ―Áreas litorales arenosas expuestas al mar que
abarcan la zona de asomeramiento, la zona de rompiente y la zona de disipación del oleaje en la zona de
batida‖ (Figura 1). Short (1999) las define como la acumulación de sedimentos depositados por las olas
entre la base de la ola (profundidad máxima en la cual las olas pueden transportar material sedimentario
hacia tierra) y el límite de swash (límite hacia tierra de la acción subaérea de la ola y el transporte de los
sedimentos). Componen dos tercios de las costas libres del hielo del mundo (1.634.701 km) (Short, 1999).
Se encuentran en todas las costas sedimentarias expuestas al oleaje, en todas las latitudes y climas (Short,
1999) predominando en las regiones tropicales y templadas (Davis & Fitzgerald, 2004). En la formación de
las playas debe existir una mayor proporción entre la acumulación de sedimento que aquel que se retira de
la costa y donde la forma de ésta ha permitido la estabilización de dicha acumulación y la predominancia de
los eventos de acreción sobre los erosivos a largo plazo (Seco-Hernández, 2004).
DESCRIPCIÓN DE LA PLAYA ARENOSA
La zona costera es una zona de transición entre los componentes marino y continental del planeta.
En ellas el mar se encuentra con la tierra disipando sobre ésta su energía. Estas zonas combinan
características propias del ambiente marino y terrestre y el balance entre los procesos de cada ambiente
origina heterogeneidad en la estructura física y ecológica. A su vez, sostienen su función ecosistémica y los
ciclos biogeoquímicos en la zona costera. Es ampliamente reconocida como uno de los elementos más
importantes de la biosfera con una amplia diversidad de ambientes y recursos.
Las playas de arena son uno de los tipos de costa dinámica más resistentes debido a su capacidad
para absorber la energía de las olas. Esta energía, se disipa sobre la zona de surf (rompiente) transportando
el sedimento desde alta mar durante las tormentas hacia tierra durante la calma. La playa se caracteriza por
el continuo transporte de sedimentos en la costa por acción del oleaje y en las dunas por la acción del

8
viento. La mayoría de las playas están respaldadas por dunas e interactúan con ellas ya sea suministrando o
recibiendo arena. En muchas costas, este transporte representa grandes volúmenes de arena. Por lo tanto,
la playa de arena es un entorno hostil para la vida (perfectamente adaptada a la severidad que impera) y
extremadamente dinámico, donde arena y agua están en continuo movimiento.
Ambiente físico
Morfología y características del oleaje
En la formación de una playa intervienen conjuntamente tres componentes básicos: tamaño de
grano, energía de la ola y el rango mareal (Brown & McLachlan, 1990; Short, 1999). Estos tres componentes
determinan la morfología, morfodinámica, las comunidades infaunales y la cantidad y calidad de la materia
orgánica de las playas. La forma del perfil de una playa es cóncavo generalmente debida a procesos que
operan a lo largo de un plano vertical en la dirección del avance de las olas, mientras que la forma del plano
de la playa es debido a procesos que operan de forma paralela a la costa (Short, 1999). En las playas se
diferencian tres zonas en función de su morfología y las características del oleaje (Figura 1a).
Zona de transición o asomeramiento (submareal): Esta zona también es conocida como
―playa exterior‖. Abarca desde la base de la ola hasta la zona de rompiente. Aquí la ola siente el fondo y por
lo tanto el sedimento se resuspende y es transportado por la corriente hacia la orilla. La profundidad de este
límite depende de la altura y periodo de ola de forma proporcional directa. Puede alcanzar los 30 m. La
forma del fondo es cóncava debido a la erosión del oleaje.
Zona de rompiente o surf (submareal): Se extiende desde el punto de rotura de las olas
hasta la orilla. La rotura de las olas se produce cuando su altura es 1.5 veces mayor que la profundidad. En
este punto, la ola para compensar la reducción de la profundidad, aumenta su altura hasta que su cresta
colapsa y se rompe. Debido a la alta energía que se genera en esta zona, ésta es la que más varía de las tres.
En consecuencia se forman corrientes de resaca (perpendiculares a la costa) y corrientes paralelas a la costa
produciendo un importante transporte de sedimentos a lo largo de la playa. Estas corrientes favorecen la
formación de estructuras subacuáticas como barras, canales y ripples.
Zona de barrido o swash (intermareal): Es la zona más estrecha que comprende desde el
punto de rotura de las olas y el límite superior de barrido (hasta donde llegan las mareas vivas en pleamar).
En bajamar es la zona que se queda expuesta al aire. Al romper la ola, esta asciende en forma de lámina
hasta que por percolación pierde volumen y por gravedad desciende el volumen restante. Por lo tanto, en
esta zona se produce un movimiento bidireccional, swash y backswash (ascendente y descendente). Este
movimiento está sujeto a factores como: altura y tipo de ola, rango y estado de marea, morfología de la
playa, tamaño de grano, etc. En su zona más baja se puede formar un escalón de tamaño de grano más
grueso que el de las zonas adyacentes. Si existe gran aporte de sedimentos, se forman barras y surcos, y en
la zona superior, la berma. Esta estructura marca el límite entre el intermareal y el supramareal.

9
Zona de spray (supramareal): se sitúa entre el límite superior del swash y la base de las dunas
en el caso de que haya. Si no, será hasta alguna estructura construida por el hombre. Esta zona tiene usos
recreativos para el hombre. En periodos de tormenta o mareas vivas el supramareal es salpicado por las
gotas procedentes del oleaje. Aquí el agente trasportador es el viento y en las playas donde existen dunas,
constituyen un gran reservorio de sedimentos para la playa (Figura 1b).
Figura 1a. Morfología de la playa y sus distintas zonas dinámicas. Esquema modificado de Short (1999).
Figura 1b. Esquema de la ubicación de la zona supramareal y sus componentes principales.

10
Morfodinamismo
En las playas, el tipo de sedimento, el oleaje y régimen mareal produce un rango de tipos
morfodinámicos continuos desde playas micromareales reflectivas (angostas e inclinadas) hasta playas
macromareales disipativas (amplias y planas bajo condiciones de mareas amplias) con variaciones
extremas entre los dos extremos (Short, 1999). Wright & Short (1984) clasificaron las playas según su
estado morfodinámico usando el parámetro de Dean (Ω) (Gourlay, 1968; Dean, 1973) en reflectivas,
intermedias y disipativas (Figuras 2 y 3).
Reflectivas: Se forman donde las olas son pequeñas (0.5 m de altura aproximadamente). Las
playas están formadas por arena gruesa y pendientes empinadas (5-10º). La ola impacta directamente
sobre la playa por lo que el lavado (swash) es fuerte. En mareas bajas se acumulan sedimentos gruesos
formando un escalón. Este tamaño de sedimento permite un alto drenaje (Short, 1999).
Intermedias: La altura de las olas oscilan entre los 0.5 y 2 m y los sedimentos arenosos van
desde finos a medios. Se caracterizan por tener una zona donde se disipa del oleaje (surf) que se incrementa
con la altura de éste. Pueden formarse barras de arena, canales y corrientes de retorno. La forma de la playa
depende de la altura de la ola. En general todos los tipos de playas intermedias permiten la vida en ellas, ya
que aunque la energía del oleaje es mayor que en las reflectivas, las amplias zonas de disipación reducen el
efecto de la energía del oleaje sobre la playa (Short, 1999).
Disipativas: Estas playas se forman donde la altura de la ola está comprendida entre los 2 y 3
m, el sedimento es fino (0.2 mm de diámetro) y por lo tanto el drenaje es lento. Presentan zonas de
disipación del oleaje que pueden llegar a superar los 500 m de amplitud donde rompen olas en forma de
derrame (spilling breakers). Debido a que toda su energía se disipa en esta zona, llegan débiles a la cara de
la playa (Short, 1999). Son los tipos de playa con mayor productividad por presentar altas biomasas de
fitobentos adaptado a vivir en la zona de barrido; esto a su vez permite el desarrollo de altas biomasas de
invertebrados filtradores (Defeo & McLachlan, 2005).
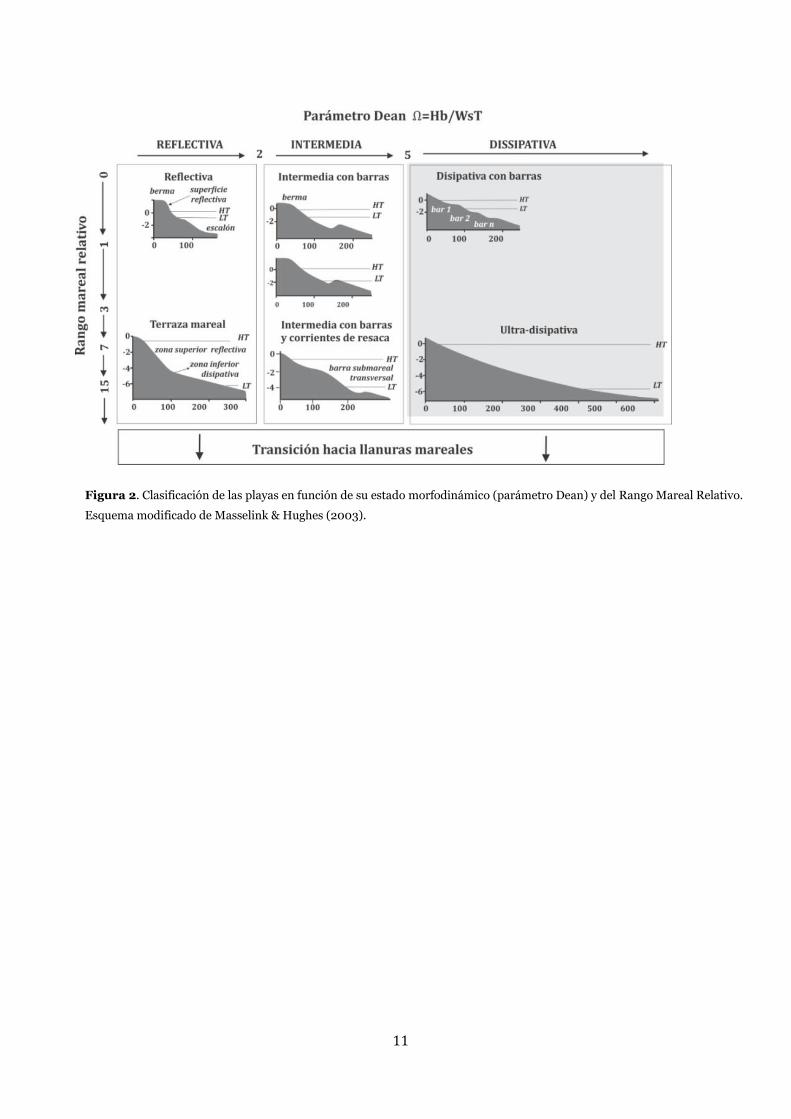
11
Figura 2. Clasificación de las playas en función de su estado morfodinámico (parámetro Dean) y del Rango Mareal Relativo.
Esquema modificado de Masselink & Hughes (2003).

12
Grado de exposición al oleaje
La pendiente de la playa depende de los procesos de swash y backswash (McLachlan & Brown,
2006). El swash es la película de agua que asciende por la zona de barrido una vez la ola ha colapsado. Esta
película transporta sedimentos hacia la parte alta de la playa causando acreción por lo que se pronuncia la
pendiente de la playa. Por el contrario, el backswash es el descenso de la lámina de agua (el resto después

13
de la percolación) por la zona de barrido arrastrando el sedimento hacia el mar. En este caso, la cara de la
playa se erosiona y la pendiente se suaviza. Por eso las playas de arena fina tienen pendientes poco
pronunciadas; retienen agua entre sus granos debido a su baja permeabilidad, de tal forma que cada swash
es seguido por un backswash total que aplana la playa. En arena gruesa, el percolado es mayor
disminuyendo el backswash por lo que la fina lámina de agua (si la hay) no puede arrastrar el sedimento.
Así pues, aunque las olas de gran altura resultan en grandes swash, el tamaño del grano será el que
determine la fuerza del backswash (Short, 1999). La pendiente de la playa es por lo tanto, el resultado de
los procesos erosivos o acrecionales. Así, el grado de exposición, dará cuenta de los procesos físicos que
tienen lugar en el ambiente de una playa.
La exposición de la playa al oleaje es también un factor sustancial en el ámbito de la ecología
marina ya que la exposición determina la estructura y funcionamiento de la playa. Explica los patrones en
la riqueza y abundancia de la macrofauna que habita en dichos sistemas. El oleaje además de influir en el
transporte del sedimento también influye en los gradientes químico y biológico (vertical y horizontal) y en
el intercambio de nutrientes en el sistema (McLachlan, 1989; de la Huz, 2008). Por tal razón, McLachlan
(1980) propuso un simple y efectivo sistema de clasificación de las playas arenosas según su grado de
exposición, basándose para ello en las características que resultan de las fuerzas hidrodinámicas ejercidas
sobre el intermareal. McLachlan (1980a) creó un sistema de clasificación para estimar la fuerza
hidrodinámica que soporta el intermareal. Según esta clasificación las playas pueden ser: muy protegidas,
protegidas, expuestas y muy expuestas (Tabla 1).
Tabla 1. Clasificación de playas según su exposición al oleaje de acuerdo a McLachlan (1980).
Efecto de la marea
Las características físicas del intermareal de una playa se basan en el tipo de sedimento, exposición
al oleaje y al régimen mareal. Mientras que el oleaje determina el estado morfodinámico predominante, el
rango mareal afecta a los procesos que se dan lugar en la zona de barrido y de rompiente (Masselink &
Short, 1993). El papel de la marea es alterar la exposición y sumergir parte de la playa y la zona de
rompiente interna cíclicamente. El resultado de este movimiento es el de retardar el transporte del

14
sedimento y por lo tanto, la morfología de la playa (Davis, 1985). Para poder caracterizar las playas en
función de su rango mareal y su estado morfodinámico, se creó el Índice del Estado de la Playa (BSI)
obteniéndose las siguientes categorías: intermedias-disipativas con alto oleaje, totalmente disipativas y
playas macromareales ultradidipativas.
ECOLOGÍA DE LA PLAYA ARENOSA
La ecología de la playa trata de comprender las fuerzas críticas que estructuran las comunidades
biológicas y, de este modo, lograr una comprensión completa de los procesos del ecosistema de la playa. Las
playas arenosas expuestas son ambientes muy dinámicos debido a las fuerzan que convergen en la zona de
rompiente del oleaje. Además, las duras condiciones que imperan debido al fuerte hidrodinamismo, la
movilidad del sedimento, los periodos de emersión durante la bajamar, la desecación y los cambios de
temperatura hacen de ellas ambientes muy hostiles para la vida. Pero muchos organismos han sido capaces
de adaptarse de una forma sorprendente a este caótico ambiente. Su distribución y abundancia está
controlada principalmente por los factores físicos mientras que las interacciones biológicas quedan en un
segundo plano (McLachlan, 1983; McLachlan et al., 1993). Por ello, esta zona alberga especies únicas y
diversas que no se encuentran en ningún otro hábitat marino (McLachlan & Brown, 2006; Schlacher et al.,
2008; Defeo et al., 2009). Las playas son un ambiente único ocupado por especies de invertebrados que se
han adaptado al movimiento constante del sedimento y a la acción del oleaje (McLachlan & Brown, 2006;
Schlacher et al., 2007) y además proporcionan hábitats para muchas especies (aves, anfibios, reptiles o
peces) que anidan, crían y se alimentan en las dunas costeras o en la playa expuesta (Dugan et al., 2003;
Schlacher et al., 2007).
Desde el punto de vista ecológico, como interface que son las playas arenosas, están constituidas
por dos componentes (McGwyne & McLachlan, 1992):
Ecosistema marino: Está controlado por la acción del sedimento, oleaje, marea y es habitado
por la biota marina.
Ecosistema terrestre. Está controlado por la acción del viento, la lluvia y es habitado por la
biota terrestre.
Ambos componentes interactúan en la misma unidad geomorfológica, en la Zona Litoral Activa (ZLA)
(Figura 4). Esta ZLA es una interface entre el mar y la tierra en equilibrio dinámico, donde los sedimentos
se mueven constantemente (Tinley, 1985; McGwynne & McLachlan, 1992). La ZLA se divide en tres zonas:
ZLA INFRA: Se extiende desde profundidades donde el oleaje ejerce su fuerza hasta el ―swash‖.
ZLA MESO: Abarca desde el swash hasta la base de las dunas, donde interaccionan la energía
eólica y la del oleaje.

15
ZLA SUPRA: Se localiza desde las dunas hacia el continente, donde finaliza el transporte eólico
de arena.
Figura 4. Zona Litoral Activa (ZLA) (Tinley, 1985; McGwynne & McLachlan, 1992). Modificada de Álava (2007).
Intercambios con otros ecosistemas
El sistema surf-playa-duna forma una sola unidad funcional intercambiando organismos,
sedimento, materia orgánica y nutrientes. Las playas son el nexo entre el mar y la tierra que permite los
intercambios de materia orgánica (fitoplancton, plantas de las dunas y desechos orgánicos marinos).
Constituye uno de los procesos más importantes para mantener el flujo de energía entre los sistemas
terrestre y marino (McLachlan & Brown, 2006, 2011). El material orgánico que llega a la playa es asimilado
por una amplia gama de consumidores a través de varios niveles tróficos (macro, meio y microfauna y
depredadores terrestres superiores) que interactúan en una red alimentaria compleja (Dugan et al., 2000;
Dugan et al., 2003; Catenazzi & Donnelly, 2007; Lastra et al., 2008; Spiller et al., 2010; Colombini et al.,
2011a).
Las cadenas tróficas comienzan y terminan principalmente en el mar, la tierra desempeña un papel
relativamente menor. Las playas son ambientes caracterizadas por una baja productividad por eso sus
fuentes de alimento son variadas:
Microflora bentónica (diatomeas bentónicas): Son los productores primarios residentes
en el sedimento de la playa. Son una fuente limitada de alimento debido al oleaje. Por lo que su producción
aumenta desde playas reflectivas a disipativas. Están disponible para la macrofauna y la meiofauna
bentónica.
Diatomeas de la zona de surf: Se encuentran en el agua de la zona de surf. Las tasas de
producción primaria de la zona de surf son menos conocidas pero se sabe que son más variables. En
ausencia de dichos parches de diatomeas, las tasas de producción primaria en las aguas de surf son mucho
más bajas en general. Las tasas más altas ocurren en zonas de surf disipativas con parches consistentes de
fitoplancton y las tasas más bajas en las zonas intermedias de bajo consumo de energía. Es un recurso
alimentario más consistente que otros insumos marinos. No obstante, la producción primaria puede ser
ampliamente variable a lo largo del día.

16
Detritus (material orgánico en partículas): Es una fuente de alimento relativamente constante
de origen marino. Puede proceder de la descomposición de plantas y animales, heces, pastoreo o
agregaciones.
Macrófitas: Pueden proceder de marismas, pastos submareales de playas protegidas o de
orillas rocosas. La mayoría de las playas reciben una pequeña cantidad de insumos, pero en algunas
situaciones (por cercanía a la fuente de alimento) la entrada puede ser sustancial, especialmente después
de las tormentas de invierno o debido a la senescencia al final del verano. Pueden cubrir grandes
extensiones de la playa y se distribuyen sobre ella dependiendo de su morfodinámica (Griffiths et al., 1983;
McLachlan & McGwynne, 1986; Dugan et al., 2003) dominando totalmente las cadenas alimentarias de las
playas de arena (Colombini y Chelazzi, 2003). Las macrófitas de la línea de deriva alimentan a anfípodos
talitrídos, isópodos e insectos. También la microfauna (bacterias y hongos) se ven favorecidos por estos
inputs. Es el nivel trófico más importante ya que son los principales responsables de la descomposición de
macrófitas, pueden representar más de un 90% (Koop y Lucas, 1983; Griffiths et al., 1983; Jedrzejczak,
2003). Después de la descomposición, grandes cantidades de material disuelto y particulado pueden
lixiviar en el sedimento y quedar disponibles para la fauna intersticial. Sin embargo, estas macrófitas
sueltas pueden no terminar en la playa, y cantidades significativas pueden lavarse en la zona de surf, donde
proporcionan alimento y refugio de los depredadores (Robertson & Lenanton, 1984).
Carroña: Es de origen marino y común en todas las playas. Representa un suministro muy
variable. Procede de animales muertos pero pierde importancia cuando hay fitoplancton de la zona de surf
o macrófitas varadas. Sin embargo, en ausencia de otros insumos importantes, la carroña puede ser de gran
importancia (Colombini & Chelazzi, 2003). En playas de reflectivas sin zonas de surf, donde la vida de los
filtradores es dura debido a la pronunciada pendiente los depredadores dominan la fauna. Suelen ser
supramareales pero descienden ya que la carroña es su principal alimento.
Materia orgánica disuelta: se producen en alta concentración en todos los lugares del mar y
tienen varios orígenes: exudados fitoplanctónicos, excreción de la fauna, lixiviados de plantas y animales
dañados y material generado por la alimentación de la macrofauna. Este material puede ser concentrado
por las olas en una espuma amarilla que puede acumularse en las olas o en la playa. Es de gran importancia
para la microbiota de la columna de agua y para la fauna intersticial. Cuando se deposita sobre la superficie
de la arena, es consumida por la macrofauna.
Detritus de insectos o de plantas: Proceden de tierra. No suelen ser de gran importancia
cuantitativa pero se suelen encontrar en la playa y en las aguas de surf. Son potencialmente importantes
cuando el viento sopla hacia el mar. Tiene valor alimenticio para pájaros, peces e invertebrados.
Dependiendo de las cantidades relativas de los diferentes recursos disponibles en la playa, existirá
gran variedad de rutas tróficas diferentes.

17
Cadenas tróficas
Cadena trófica macroscópica
Los componentes de estas cadenas comprenden animales con un tamaño superior a 1 mm como el
bentos (filtradores, herbívoros y depredadores), zooplancton, peces y aves (Figura 5).
En playas protegidas los depositívoros son los dominantes ya que el sedimento fino y estable
favorece la acumulación de grandes cantidades de detrito. También pueden alimentarse de la microflora
bentónica y de la meiofauna. En las playas expuestas, sin embargo, este componente es ausente o
insignificante ya que el contenido orgánico en el sedimento es significativamente menor.
En playas donde las condiciones del swash son favorables para la alimentación, los filtradores
son generalmente dominantes y a menudo muy abundantes. Al alimentarse del fitoplancton de la resaca
y/o con los detritos particulados, pueden representar la principal vía de flujo de energía a través del bentos,
representando hasta el 90%. Cuanto más disipativa sea una playa, mayor será la tendencia a desarrollar
acumulaciones de fitoplancton de surf.
En playas con grandes aportes de macrófitas, los carroñeros y depredadores (anfípodos
supramareales, isópodos e insectos) que rara vez alcanzan grandes abundancias, en estos ambientes
pueden llegar a dominar. Aquí, la biomasa bentónica se concentra alrededor de la línea de deriva y
disminuye hacia abajo, una tendencia opuesta a la que se encuentra en la mayoría de las playas. Esta
macrofauna puede ser presa de las aves.
Existe un gradiente de los depredadores desde tierra hacia el mar. En la zona supramareal, los
depredadores terrestres (aves, cangrejos e insectos) son dominantes y los depredadores marinos (peces,
cangrejos y gasterópodos) en la zona inter y submareal. El grupo de depredadores más importante depende
de la ubicación de la biomasa bentónica. Como generalmente se encuentra en el intermareal inferior o en el
submareal, en la mayoría de las playas los depredadores marinos tienen el mayor impacto sobre las redes
tróficas macrófitas. Los peces, los invertebrados carnívoros y las aves suelen ocupar el último eslabón de las
cadenas tróficas macroscópicas de las playas arenosas (Brown & McLachlan, 2010).
Así pues, la macrofauna bentónica al alimentarse de fuentes primarias y ser presa de los
depredadores superiores, ocupa una posición clave en el centro de las cadenas tróficas de las playas
arenosas
Cadena trófica intersticial
Es la cadena trófica que se localiza en el interior del sedimento de la playa. Está formada por
bacterias, protozoos y meiofauna y no tiene vínculos tróficos significativos con la macrofauna. Se alimenta
de materia orgánica disuelta arrojada en la arena por acción del oleaje y la marea. El sistema intersticial

18
puede ser visto como un gran filtro natural, limpiando y purificando las aguas del surf (Pearse et al., 1942).
Mineraliza los materiales orgánicos que recibe en el sedimento y los devuelve en forma de nutrientes al
mar. Por tanto, la importancia del sistema intersticial en el procesamiento de la materia orgánica viene
determinada por cuatro factores:
El volumen de agua filtrado por la arena.
El tiempo de residencia de esta agua en la arena.
El contenido orgánico de esta agua.
La temperatura del agua que controla la tasa metabólica de la biota intersticial.
Cadena trófica microbiana
La comunidad microbiana tiene menos relevancia que otras cadenas alimenticias. Sin embargo,
existen dos posibles conexiones tróficas que permiten el flujo de energía otras redes alimenticias: los
flagelados pueden ser consumidos por los filtradores bivalvos y filtradores zooplanctónicos. Los flagelados
son un componente alimenticio abundante para los filtradores bivalvos. Las densidades del
microzooplancton son demasiado bajas para representar una importante fuente de alimento para el
zooplancton más grande. Por eso se cree que la energía que llega al último eslabón de la cadena es muy
baja. Por lo tanto, podemos seguir viendo el bucle microbiano como un sistema trófico distinto, con
intercambios limitados sólo con las cadenas alimentarias macroscópicas. Se puede concluir que a pesar de
su pequeño tamaño, los organismos que constituyen el bucle microbiano representan un sistema trófico
extremadamente importante en las aguas de surf.
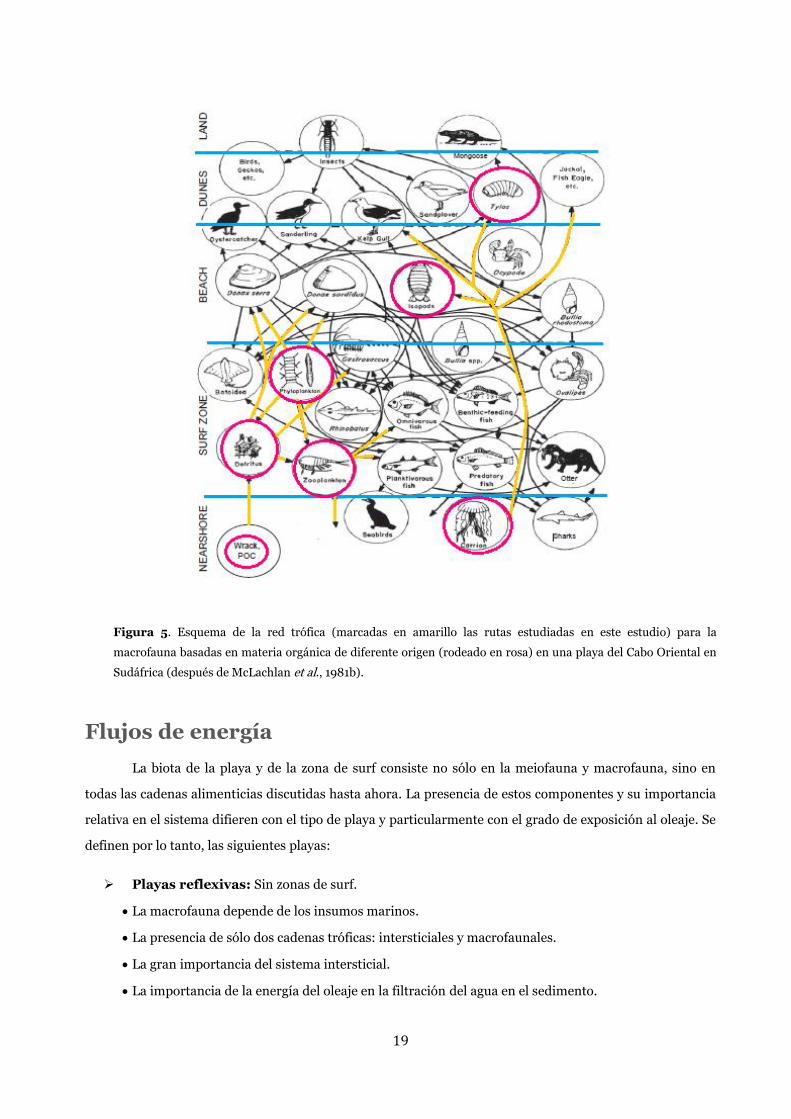
19
Figura 5. Esquema de la red trófica (marcadas en amarillo las rutas estudiadas en este estudio) para la
macrofauna basadas en materia orgánica de diferente origen (rodeado en rosa) en una playa del Cabo Oriental en
Sudáfrica (después de McLachlan et al., 1981b).
Flujos de energía
La biota de la playa y de la zona de surf consiste no sólo en la meiofauna y macrofauna, sino en
todas las cadenas alimenticias discutidas hasta ahora. La presencia de estos componentes y su importancia
relativa en el sistema difieren con el tipo de playa y particularmente con el grado de exposición al oleaje. Se
definen por lo tanto, las siguientes playas:
Playas reflexivas: Sin zonas de surf.
La macrofauna depende de los insumos marinos.
La presencia de sólo dos cadenas tróficas: intersticiales y macrofaunales.
La gran importancia del sistema intersticial.
La importancia de la energía del oleaje en la filtración del agua en el sedimento.

20
Insumos de grandes cantidades de macrófitas (en algunos casos).
Baja biomasa macrofaunal en la mayoría de los casos.
Exportación de macrofauna a depredadores marinos y terrestres.
Playas disipativas: Tienen zonas de surf bien desarrolladas con patrones de circulación del agua.
En estas zonas de surf, existe una rica producción primaria basada en diatomeas. Las células de
circulación tienden a retener este y otros materiales orgánicos, de modo que el ecosistema no
depende de los insumos del mar.
Una circulación de oleaje accionada por olas que retiene material orgánico.
La presencia de no menos de cuatro sistemas bióticos (fitoplancton, bucle microbiano, sistema
intersticial y macrofauna).
Alta producción primaria.
Gran importancia del bucle microbiano.
Importancia de la energía de las olas en el control de la producción primaria, del sistema intersticial
y exportación de la producción excedente.
Altas biomasas.
Playas intermedias: Tienen una zona de surf moderadamente desarrollada y su producción
primaria es insignificante y transitoria, formada por zooplancton y alguna actividad microbiana en
el agua.
Macrofauna
En función del tamaño, los organismos que viven en las playas arenosas expuestas se pueden
dividir en tres clases: microfauna, meiofauna y macrofauna (Schwinghammer, 1981; Warwick, 1984). La
macrofauna invertebrada tiene un tamaño mayor de 1 mm (Bishop & Hartley, 1986; Lastra, 1991; Hacking,
1997; de la Huz, 2008). Su distribución a lo largo y ancho de las playas es irregular como resultado del
efecto del oleaje, de las mareas, de la disponibilidad de alimento, de la capacidad de penetración en el
sedimento, etc. Brown y McLachlan (1983) definieron 2 zonas dentro de la playa en base a dos límites
naturales: la zona de resurgencia y la línea de varamiento o drift-line. En función de estos límites se definen
dos zonas:
La parte superior: zona donde vive la fauna de respiración aérea.
La parte inferior: viven los organismos con respiración subacuática.
En las playas de alta energía, las grandes fuerzas hidrodinámicas y el estrés ambiental limitan el
número de especies (McLachlan et al., 1996) influyendo en la riqueza específica y en la abundancia

21
observándose un incremento desde playas reflectivas hacia playas disipativas (McLachlan et al., 1981; Defeo
et al., 1992).
Teorías ecológicas
McLachlan (2001) identificó 3 paradigmas que definen a gran escala los patrones comunitarios en
las playas arenosas de alta energía:
el control físico en ambientes severos.
Modelos de morfodinámica de la playa.
Incremento de la riqueza específica, abundancia y biomasa desde playas micromareales reflectivas
hacia macromareales disipativas (Defeo & McLachlan, 2005) (Figura 6 y 7).
Hipótesis autoecológica (HA)
Se basa en el control de los factores físicos sobre las comunidades de la playa expuesta resultando
en la ausencia de una estructura biogénica (AH, Autecological Hypothesis: Noy-Meir, 1979; Defeo &
McLachlan, 2005). Las comunidades están estructuradas en respuesta al ambiente físico, siendo mínimas
las interacciones biológicas. Esta hipótesis predice el incremento de las comunidades desde playas
reflectivas hacia playas disipativas (McLachlan et al., 1981, 1993, 1996; McLachlan, 1990; Jaramillo et al.,
1995; Defeo & McLachlan, 2005).
Hipótesis de exclusión del swash (SEH)
La idea de la AH fue refinada con la Hipótesis de Exclusión del Swash (SEH, Swash Exclusion
Hypothesis: McLachlan et al., 1993) sólo es válida para el intermareal. Se basa en el clima del swash que
experimenta la macrofauna (McArdle & McLachlan 1991, 1992). Predice que el ambiente físico regula la
vida de las comunidades macrofaunísticas debido a la rigurosidad del ambiente intermareal y al tamaño del
grano. Por eso, la riqueza específica y su abundancia disminuyen desde playas disipativas (condiciones más
favorables del swash y tamaño fino del grano) a reflectivas (rigurosidad del clima de swash y tamaño de
grano grueso) (McLachlan, 1990; Defeo et al., 1992; McLachlan et al., 1993; Jaramillo et al., 2000; Rodil et
al., 2003; McLachlan et al., 2005; Lercari et al., 2003).
Hipótesis de severidad ambiental (enfoque poblacional) (HHH)
(HHH, HabitatHarshness Hypothesis: Defeo et al., 2001, 2003) postula que la rigurosidad
ambiental de las playas reflectivas obliga a los organismos a invertir más energía en su mantenimiento. Por
lo tanto, la fecundidad, crecimiento, masa corporal y sobrevivencia de las poblaciones aumenta de playas
reflectivas hacia disipativas.
Hipótesis de seguridad de hábitat (enfoque poblacional) (HSH)
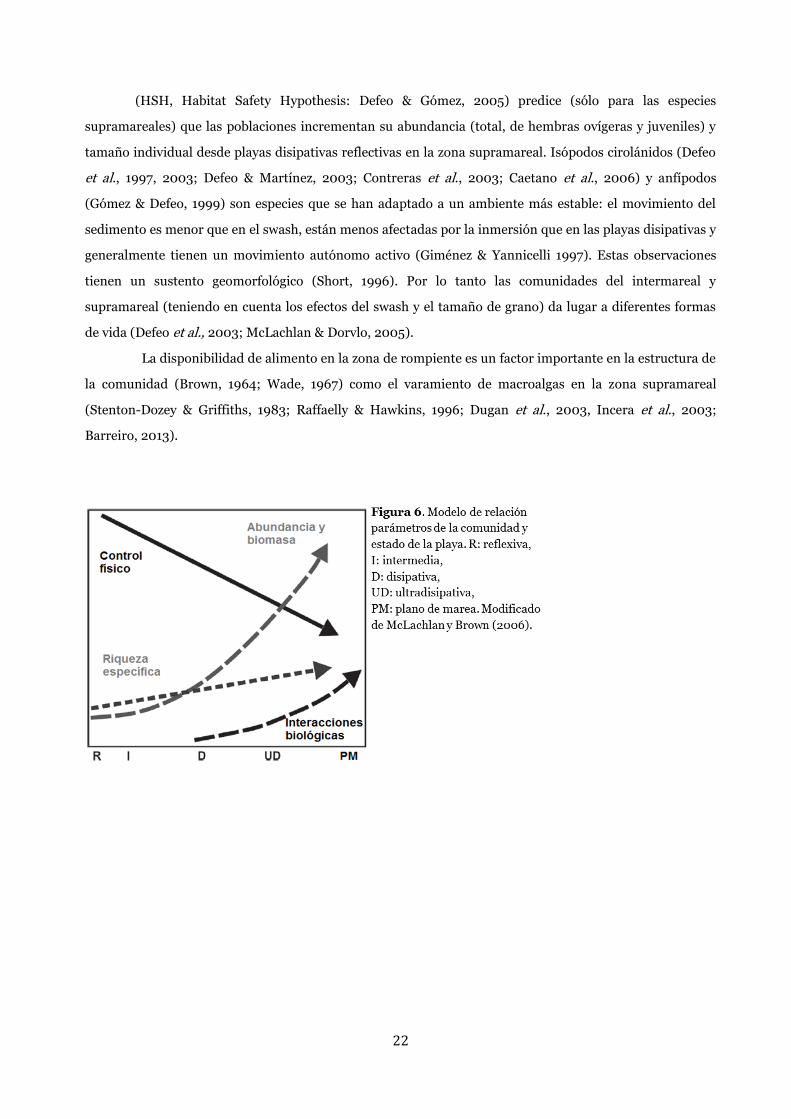
22
(HSH, Habitat Safety Hypothesis: Defeo & Gómez, 2005) predice (sólo para las especies
supramareales) que las poblaciones incrementan su abundancia (total, de hembras ovígeras y juveniles) y
tamaño individual desde playas disipativas reflectivas en la zona supramareal. Isópodos cirolánidos (Defeo
et al., 1997, 2003; Defeo & Martínez, 2003; Contreras et al., 2003; Caetano et al., 2006) y anfípodos
(Gómez & Defeo, 1999) son especies que se han adaptado a un ambiente más estable: el movimiento del
sedimento es menor que en el swash, están menos afectadas por la inmersión que en las playas disipativas y
generalmente tienen un movimiento autónomo activo (Giménez & Yannicelli 1997). Estas observaciones
tienen un sustento geomorfológico (Short, 1996). Por lo tanto las comunidades del intermareal y
supramareal (teniendo en cuenta los efectos del swash y el tamaño de grano) da lugar a diferentes formas
de vida (Defeo et al., 2003; McLachlan & Dorvlo, 2005).
La disponibilidad de alimento en la zona de rompiente es un factor importante en la estructura de
la comunidad (Brown, 1964; Wade, 1967) como el varamiento de macroalgas en la zona supramareal
(Stenton-Dozey & Griffiths, 1983; Raffaelly & Hawkins, 1996; Dugan et al., 2003, Incera et al., 2003;
Barreiro, 2013).

23
WRACK
¿Qué es?
La palabra "wrack" en inglés no tiene una traducción exacta en español. La Comisión Costera de
California ha definido "wrack" como "materia orgánica compuesta principalmente de algas y plantas
marinas que son arrojadas a la playa por el oleaje, las mareas y el viento‖. Después de la pleamar más
reciente, el material varado en la playa se distribuye por ella en bandas mediante la acción del viento y la
acción del oleaje. Estas bandas se denominan líneas de deriva o ―drift line‖ paralelas a la orilla (Marsden,
1991a; Ochieng & Erftemeijer, 1999). A veces se acumula sobre la playa en forma de parches (cúmulos). Su
distribución sobre la playa varía dependiendo de la estación del año (climatología). Su composición y
tamaño también es muy heterogéneo en el espacio y en el tiempo (Ochieng & Erftemeijer, 1999; Wells,
2002).
Rol e importancia
Las algas varadas son una fuente importante de nutrientes y centros de alta diversidad de
organismos de todos los tamaños. Los depósitos de wrack pueden desempeñar diversos papeles
importantes en los ecosistemas costeros:
Pueden modificar el perfil de la playa por acreción debido a su función como barrera protectora
contra la fuerza del viento, oleaje y mareas (Nordstrom et al., 2006).
Pueden actuar como microhábitats, muchas especies encuentran en ellos refugio o alimento.

24
Pueden propiciar la formación de dunas de arena costeras.
Son puntos de regeneración de nutrientes a través de su descomposición. Por lo que
proporciona una fuente importante de nutrientes y energía para el ecosistema de la playa (McLachlan,
1981; Alongi, 1998).
De las cuatro funciones anteriormente citadas, la que genera mayor interés es la función del material
de arribazón como punto de regeneración de nutrientes.
La línea de deriva o drift line es una zona muy dinámica en la cantidad de materia orgánica
(fundamentalmente macroalgas) pero cuando los insumos son sustanciales, la actividad metabólica
aumenta considerablemente al producirse la degradación de la materia orgánica varada (Griffiths et al.,
1983; Heck et al., 2008; Zobell, 1971). En comparación con otros hábitats, los procesos biogeoquímicos que
ocurren en las playas arenosas expuestas han recibido significativamente menos atención debido
probablemente a su relativo bajo contenido orgánico y a la percepción de que estos sistemas tienen menos
impacto en las aguas costeras (Rocha, 2008). Sin embargo, estas creencias han sido cuestionadas por varios
estudios (Anschutz et al., 2009, Boudreau et al., 2001; Huettel & Rusch, 2000; Jahnke et al., 2005; Rocha,
2008; Rusch et al., 2006). Observaciones recientes sugieren dejar de ignorar los flujos biogeoquímicos de
los sedimentos arenosos por su importancia ecológica (Rusch et al., 2006).
Estas interfaces han sido identificadas como ―puntos calientes‖ biogeoquímicos de la biosfera (McClain et
al., 2003). Este nuevo concepto del intermareal es el resultado de numerosos estudios que sitúan a las
playas como grandes reactores de la materia orgánica en comparación con las áreas circundantes (McClain
et al., 2003; Coupland et al., 2007). Estos ―puntos calientes‖ han sido considerados como prioridad de
estudio y un gran desafío para la ciencia (McClain et al., 2003). Muchas investigaciones se han centrado
principalmente en estudiar los procesos biogeoquímicos de los sedimentos debido a sus rápidas tasas de
remineralización así como su potencial para la acumulación y preservación de la materia orgánica (Avery et
al.,2008). Su variabilidad temporal, espacial y la biomasa de estos aportes afectan tanto a los procesos
metabólicos del ecosistema (ciclo del C, dinámica de nutrientes, flujos de CO2, etc.) como a la composición
de la comunidad microfaunística (Rieper-Kirchner, 1989), meiofaunística (Urban-Malinga & Burska, 2009)
y macrofaunísitica (Dugan et al., 2003; Jaramillo et al., 2006), produciéndose un efecto cascada sobre los
niveles superiores de la cadena trófica hasta llegar a aves y mamíferos, entre otros (Dugan et al., 2003).
En el momento que los subsidios alóctonos (sobre todo macroalgas y plantas vasculares) se
depositan en la línea de varamiento comienzan a sufrir una serie de procesos (lixiviación, catabolismo y
trituración o fragmentación) donde cada grupo (macro, meio y microfauna) comienza una desenfrenada
actividad en el proceso correspondiente de degradación. Su actividad tiene como objeto la supervivencia.
Algunos lo hacen usando este wrack como refugio, lugar de cría o como recurso alimenticio pero entre
todos, están llevando a cabo la descomposición de la materia orgánica. Esta cadena involucra a multitud de

25
organismos los cuales a su vez se convierten en base importante de la pirámide trófica de los ecosistemas
costeros. Por eso, es un proceso muy dinámico y complejo ya que interviene una comunidad entera. A nivel
ecológico el drift line es rico en nutrientes y concentra una alta biodiversidad en comparación con el resto
de la playa.
Los procesos de remineralización dan como resultado componentes geoquímicamente importantes
incluyendo nutrientes, compuestos orgánicos, dióxido de carbono (Capone & Kiene, 1988). La clave puede
estar en el tamaño arena; permite el flujo advectivo en el interior del sedimento mejorando la circulación
del oxígeno disuelto en los poros favoreciendo la descomposición. Además, la arena tiene la capacidad de
filtrar grandes volúmenes de agua de mar o fluviales (McLachlan et al., 1985) facilitando la descomposición
y la remineralización de la materia orgánica. La zona intermareal, por lo tanto, representa un entorno clave
en la liberación de nutrientes (Billerbeck et al., 2006; Deborde et al., 2008). Las playas son por lo tanto,
fuente de nutrientes para los ecosistemas vecinos como para sí mismas (Dugan et al., 2011) favoreciendo a
toda la pirámide trófica costera. Su estudio aporta información sobre la productividad de la playa y eficacia
de ésta para reciclar la materia orgánica. Además, con la información obtenida se puede caracterizar el tipo
de playa según su naturaleza autotrófica o heterotrófica (Migne et al., 2005). Las playas representan
aproximadamente un 70% de las costas abiertas del mundo convirtiéndose inevitablemente en foco de
estudio (Bascom, 1980). Cada vez se reconoce que cuantificar la función del ecosistema de las playas en el
ciclo de los nutrientes es de suma importancia para evaluar el procesamiento de nutrientes costeros y la
remineralización de la materia orgánica (Anschutz et al., 2009; Rauch & Denis, 2008). Estudiar los
procesos asociados a la degradación del wrack y los factores que la controlan, son, por lo tanto, esenciales
para determinar el destino de estos materiales y evaluar su importancia en el ciclo de la materia y la energía
en la interfaz tierra-mar. También ha de tenerse en cuenta la materia orgánica procedente de las descargas
fluviales y de las aguas subterráneas por representar un 40% de las aguas que llegan a la costa (Moore,
1996). Las playas arenosas se han convertido en "grandes sistemas de digestión e incubación" (Pearse et al.,
1942) por su rol en la mineralización de la materia orgánica y reciclaje de nutrientes.
El CO2 es un componente muy importante en el ciclo global del carbono y está directamente
afectado por el calentamiento global (Xu & Qi, 2001). La respiración (del sedimento y de la fauna) es
proporcional a la Productividad Primaria Neta (PPN) del ecosistema (Wang & Polglase, 1995) y después de
la Productividad Primaria Bruta (PPB), es el segundo flujo de carbono más importante en el balance global
del carbono (Schimel, 1995). Por este motivo, el propósito de este estudio es conocer y cuantificar los
procesos degradativos y la mineralización de la materia orgánica. Estudiar cómo los factores físico-
ambientales (temperatura, humedad, tamaño de grano, etc.) y los biológicos (biomasa algal y macrofauna)
afectan al metabolismo de la playa. Para ello se utiliza el flujo de CO2 (liberado durante la mineralización)
como indicador de la actividad metabólica. Como resultado del aumento de la relevancia del cambio
climático, así como el incremento de la importancia de los suelos en su papel como sumideros de carbono,

26
este trabajo aportará un mayor conocimiento sobre el metabolismo de las playas arenosas y su importancia
como interfaces. Con esta información se podrá predecir y/o gestionar el litoral ante el escenario de cambio
climático que se está viviendo.
PLAYAS EN PELIGRO
Varios procesos del cambio global antropogénico en los ecosistemas costeros tienen el potencial de
afectar a la diversidad, el metabolismo y la estructura de las comunidades de las playas arenosas. Estos
procesos incluyen el calentamiento de la superficie del mar y del aire, el aumento de la radiación UV, la
acidificación y los cambios en la disponibilidad de macronutrientes y metales traza a través de los inputs
que recibe la playa (Finkel et al., 2010; Winder & Sommer, 2012). Dada la imposibilidad práctica de
abordar simultáneamente los efectos interactivos de múltiples factores de estrés, se ha optado por estudiar
los factores que se espera sean los más relevantes.
Calentamiento global y subida del nivel del mar
El cambio climático representa una amenaza importante para los ecosistemas costeros y las
comunidades de organismos que viven en ellos. En muchas áreas, las amenazas proyectadas combinadas de
aumento del nivel del mar, aumento de las temperaturas y reducción de las lluvias, crearán una situación
sin precedentes en las especies, los ecosistemas y los asentamientos humanos. Teniendo en cuenta que
incluso las más estrictas políticas de mitigación del cambio climático son poco propensas a detener o
reducir la amenaza del cambio climático, es necesario prever cómo las especies, los ecosistemas y las
comunidades humanas podrían adaptarse a los cambios previstos. Aunque no se han realizado estudios
directos sobre el cambio climático en los ecosistemas de playa (Defeo et al., 2009), se considera que las
playas de arena son extremadamente vulnerables, particularmente a las amenazas asociadas con el
aumento del nivel del mar (Schlacher et al., 2008). Los impactos del cambio climático variarán
geográficamente aunque en general estarán más expuestas al riesgo de aumento de las tormentas y ciclones
(Castelle et al., 2008; Lucrezi et al., 2010). En algunas áreas, el cambio climático puede inducir un lento
retroceso de la costa con pocos efectos sobre los ecosistemas de las playas (Elliot et al, 2006), mientras que
la pérdida total del hábitat de la playa arenosa podría ocurrir en escenarios de cambio climático más
extremos.
A escala local, la interfaz duna - playa es a menudo la más afectada por los eventos de tormenta y la
erosión y se considera el área más sensible a los impactos del cambio climático (Brown & McLachlan,
2002). Las temperaturas crecientes tendrán mayores efectos negativos sobre las especies adaptadas a
condiciones más frías. Correrán el riesgo de verse mermada su abundancia o incluso verse abocadas a la
extinción local (Jones et al., 2008). El aumento de las temperaturas afectará a procesos como la

27
fotosíntesis, la descomposición y el ciclo de los nutrientes provocando cambios en la productividad y en la
estructura de la red trófica. Un posible cambio consistirá en la desintegración acelerada de la playa que
puede desoxigenar la arena subyacente con efectos deletéreos para muchas especies de playas de arena
(Jones et al., 2008). Los cambios en la productividad y en la estructura trófica se verán alterados por la
acción de las olas, las mareas de tormenta y las corrientes oceánicas, así como los efectos de la acidificación
de los océanos. La conectividad entre ecosistemas costeros se verán en peligro ya que las contribuciones
marinas a las redes tróficas de la playa, juegan un papel decisivo en su productividad (Jones et al., 2004).
Una de las consecuencias del cambio climático más catastróficas en la costa, será su erosión debido
al aumento del nivel del mar y por lo tanto, la pérdida del hábitat de las playas. La microfauna de la playa se
considera relativamente resistente a las oleadas de tormenta y a las perturbaciones asociadas, pero sus
respuestas a largo plazo de cómo la frecuencia e intensidad de los eventos catastróficos son desconocidas.
Los efectos sobre los ecosistemas del incremento del CO2 atmosférico, deposición de nitrógeno,
modificaciones climáticas y, en general, procesos de cambio global generados por la acción humana, no
pueden ser analizados sin considerar convenientemente la descomposición de materia orgánica,
especialmente al componente microbiano asociado a la misma, debido a su carácter clave en el
funcionamiento de los sistemas. El proceso de descomposición puede actuar como sumidero o fuente de
nutrientes del sistema, dependiendo de cómo varíen las condiciones ambientales y de una serie de
umbrales no conocidos en su totalidad. Los efectos que el cambio climático tendrá sobre las playas, y su
capacidad para almacenar o liberar carbono son muy complejos y difíciles de predecir. El aumento de las
temperaturas y la concentración de CO2 atmosférico aumentarán la producción primaria y, por tanto, el
almacenamiento de carbono. Esto también contribuirá especialmente en climas tradicionalmente fríos, al
descenso de los niveles hídricos, y al aumento de la actividad biológica y, por lo tanto a la descomposición y,
consecuentemente a la liberación de carbono (Gitay et al., 2001; Cao et al., 1998; Lal et al., 2001). Debido a
todas estas incertidumbres y a la necesidad de establecer modelos sólidos de cara a escenarios futuros, es
necesario ahondar en el análisis y comprensión de manera integral del proceso de descomposición, y de los
factores que lo controlan, especialmente a nivel microbiano, y desde un punto de vista funcional (Sahagian
& Melack, 1998; Larsen et al., 2002).
OBJETIVOS
El objetivo general de este proyecto consiste en obtener la información necesaria que permita
conocer el efecto del calentamiento global sobre los procesos degradativos de los varamientos de
macroalgas sobre los ecosistemas intermareales arenosos. Muchos de esos efectos todavía son
impredecibles por el gran desconocimiento que hay sobre este tema. Dada la elevada velocidad del
crecimiento y expansión de las algas (muchas de ellas invasoras), debido probablemente a cambios globales

28
(contaminación, cambio climático…), dichos efectos serán de gran relevancia, especialmente sobre las
comunidades de los ecosistemas intermareales sedimentarios y sobre los ciclos biogeoquímicos.
La hipótesis general está basada en que el calentamiento global provocará alteraciones en el ciclo y
composición de la materia orgánica, en la frecuencia y cantidad del wrack, en la riqueza específica,
abundancia, biomasa y organización trófica de las comunidades faunísticas y cambios en los ciclos
biogeoquímicos. Todo ello provocará un incremento de la liberación de nutrientes y del metabolismo
general de estos ecosistemas.
BIBLIOGRAFÍA
Alongi, D. M. (1998). Coastal ecosystem processes. CRC Pres, Boca Raton, Florida.
Anschutz, P., Smith, T., Mouret, A., Deborde, J., Bujan, S., Poirier, D., & Lecroart, P. (2009). Tidal
sands as biogeochemical reactors. Estuarine, Coastal and Shelf Science, 84(1), 84-90.
Avery, L. M., Williams, A. P., Killham, K., & Jones, D. L. (2008). Survival of Escherichia coli O157: H7 in
waters from lakes, rivers, puddles and animal-drinking troughs. Science of the total environment,
389(2), 378-385.
Barreiro F. (2013) Ecology of wrack macroalgae on exposed sandy beaches. In: Departamento de Ecología y
Biología Animal. Universidad de Vigo
Bascom, W. (1980) Waves and beaches: the dynamics of the ocean surface. Anchor Press, Garden City, New
York.

29
Billerbeck, M., Werner, U., Polerecky, L., Walpersdorf, E., DeBeer, D., & Huettel, M. (2006). Surficial and
deep pore water circulation governs spatial and temporal scales of nutrient recycling in intertidal
sand flat sediment. Marine Ecology Progress Series, 326, 61-76.
Bishop, J. D. D., & Hartley, J. P. (1986). A comparison of the fauna retained on 0· 5 mm and 1· 0 mm
meshes from benthic samples taken in the Beatrice Oilfield, Moray Firth, Scotland. Proceedings of
the Royal Society of Edinburgh, Section B: Biological Sciences, 91, 247-262.
Boudreau, B. P., Huettel, M., Forster, S., Jahnke, R. A., McLachlan, A., Middelburg, J. J., ... & Webster, I.
(2001). Permeable marine sediments: overturning an old paradigm. EOS, Transactions American
Geophysical Union, 82(11), 133-136.
Burke, L., Reytar, K., Spalding, M., & Perry, A. (2011). Reefs at risk revisited.
Brown, A. C. (1964). Food relationships on the intertidal sandy beaches of the Cape Peninsula. South
African Journal of Science, 60(2), 35-41.
Brown, A. C., and McLachlan, A. (1990). Ecology of sandy shores. Amsterdam: Elsevier.
Brown, A. C., & McLachlan, A. (2010). The ecology of sandy shores. Academic Press.
Caetano, C. H., Cardoso, R. S., Veloso, V. G., & Silva, E. S. (2006). Population biology and secondary
production of Excirolana braziliensis (Isopoda: Cirolanidae) in two sandy beaches of southeastern
Brazil. Journal of coastal research, 825-835.
Cao, M., & Woodward, F. I. (1998). Dynamic responses of terrestrial ecosystem carbon cycling to global
climate change. Nature, 393(6682), 249-252.
Castelle, B., Y. Le Corre and R. Tomlinson (2008). Can the Gold Coast beaches withstand extreme events?
Geo-Marine Letters 28(1): 23-30.
Colombini, I., Chelazzi, L., Gibson, R. N., & Atkinson, R. J. A. (2003). Influence of marine allochthonous
input on sandy beach communities. Oceanography and Marine Biology: An Annual Review, 41, 115-
159.
Contreras, H., Jaramillo, E., Duarte, C., and McLachlan, A. (2003). Population abundances, growth and
natural mortality of the crustacean macroinfauna at two sandy beach morphodynamic types in
southern Chile. Rev. Chilena Hist. Nat. 76:543–561.
Coupland, G. T., C. M. Duarte & D. I. Walker, 2007. High metabolic rates in beach cast communities.
Ecosystems 10:1341–1350
Davis Jr, R. A. (1985). Beach and nearshore zone. In Coastal sedimentary environments (pp. 379-444).
Springer New York.
Davis R., Fitzgerald D., 2004. Beach and nearshore environment, In: Davis R., Fitzgerald D., Beaches and
coasts, Blackwell Science Ltd, United Kingdom, pp. 115-129
Dahl, E. (1952). Some aspects of the ecology and zonation of the fauna of sandy beaches. Oikos
4:1–27.

30
de la Huz R. (2008) Ecología de la macrofauna de las playas expuestas de Galicia. In: Departamento de
Ecología y Biología Animal. Universidad de Vigo
Dean, R. G. (1973). Heuristic models of sand transport in the surf zone. In First Australian Conference on
Coastal Engineering, 1973: Engineering Dynamics of the Coastal Zone (p. 215). Institution of
Engineers, Australia.
Deborde, M., & Von Gunten, U. R. S. (2008). Reactions of chlorine with inorganic and organic compounds
during water treatment—kinetics and mechanisms: a critical review. Water research, 42(1), 13-51.
Defeo, O., Ortiz, E., & Castilla, J. C. (1992). Growth, mortality and recruitment of the yellow clam
Mesodesma mactroides on Uruguayan beaches. Marine Biology, 114(3), 429-437.
Defeo, O., Brazeiro, A., De Alava, A., & Riestra, G. (1997). Is sandy beach macrofauna only physically
controlled? Role of substrate and competition in isopods. Estuarine, Coastal and Shelf Science,
45(4), 453-462.
Defeo, O., Gomez, J., and Lercai, D. (2001). Testing the swash exclusion hypothesis in sandy beach
populations: The mole crab Emerita brasiliensis in Uruguay. Mar. Ecol. Prog. Ser. 212:159–170.
Defeo, O. (2003). Marine invertebrate fisheries in sandy beaches: An overview. J. Coast. Res. S. I. 35:56–
65.
Defeo, O., and Gomez, J. (2005). Morphodynamics and habitat safety in sandy beaches: Life history
adaptations in a supralittoral amphipod. Mar. Ecol. Prog. Ser. 293:143–153.
Defeo, O., and McLachlan, A. (2005). Patterns, processes and regulatory mechanisms in sandy
beach macrofauna: A multi-scale analysis. Mar. Ecol. Prog. Ser. 295:1–20.
Defeo, O., McLachlan, A., Schoeman, D. S., Schlacher, T. A., Dugan, J., Jones, A. & Scapini, F. (2009).
Threats to sandy beach ecosystems: a review. Estuarine, Coastal and Shelf Science, 81(1), 1-12.
Dugan J, Hubbard DM, McCrary MD, Pierson MO (2003) The response of macrofauna communities and
shorebirds to macrophyte wrack subsidies on exposed sandy beach of southern California. Estuar
Coast Shelf S 58:25-40
Dugan, J. E., Hubbard, D. M., Page, H. M., & Schimel, J. P. (2011). Marine macrophyte wrack inputs and
dissolved nutrients in beach sands. Estuaries and Coasts, 34(4), 839-850.
Elliot, M. J., A. Travers and I. Eliot (2006). Morphology of a Low-Energy Beach, Como Beach, Western
Australia. Journal of Coastal Research 22(1): 63-77.
Gitay, H., Brown, S., Easterling, W., Jallow, B., Antle, J., Apps, M. J., ... & Laine, J. (2001). Ecosystems and
their goods and services.
Giménez, L., & Yannicelli, B. (1997). Variability of zonation patterns in temperate microtidal Uruguayan
beaches with different morphodynamic types. Marine Ecology Progress Series, 197-207.
Gomez, J., & Defeo, O. (1999). Life history of the sandhopper Pseudorchestoidea brasiliensis (Amphipoda)
in sandy beaches with contrasting morphodynamics. Marine Ecology Progress Series, 209-220.
Gourlay, M. R., & Meulen, T. (1968). Beach and dune erosion tests (I). Deltares (WL).

31
Griffiths, C. L., Stenton-Dozey, J. M. E., and Koop, K. (1983). Kelp wrack and energy flow through a sandy
beach. In: Sandy beaches as ecosytems, A. McLachlan and T. Erasmus (eds.), pp 547–556, The
Hague: Dr W. Junk.
Hacking, N. (1997). Sandy beach macrofauna of eastern Australia: A geographical comparison. Ph.D. thesis,
University of New England, Australia.
Heck, K. L., Carruthers, T. J., Duarte, C. M., Hughes, A. R., Kendrick, G., Orth, R. J., & Williams, S. W.
(2008). Trophic transfers from seagrass meadows subsidize diverse marine and terrestrial
consumers. Ecosystems, 11(7), 1198-1210.
Huettel, M., Røy, H., Precht, E., & Ehrenhauss, S. (2003). Hydrodynamical impact on biogeochemical
processes in aquatic sediments. In The Interactions between Sediments and Water (pp. 231-236).
Springer Netherlands.
Incera, M., Cividanes, S. P., Lastra, M., & López, J. (2003). Temporal and spatial variability of sedimentary
organic matter in sandy beaches on the northwest coast of the Iberian Peninsula. Estuarine, Coastal
and Shelf Science, 58, 55-61.
Jahnke, R., Richards, M., Nelson, J., Robertson, C., Rao, A., & Jahnke, D. (2005). Organic matter
remineralization and porewater exchange rates in permeable South Atlantic Bight continental shelf
sediments. Continental Shelf Research, 25(12), 1433-1452.
Jaramillo, E., McLachlan, A., and Dugan, J. E. (1995). Total sample area and estimates of species richness
in exposed sandy beaches. Mar. Ecol. Prog. Ser. 119:311–314.
Jaramillo, E., Dugan, J., and Contreras, H. (2000). Abundance, population structure, tidal movement and
burrowing rate of Emerita analoga (Stimpson 1857) (Anomura: Hippidae) at a dissipative and a
reflective sandy beach in south central Chile. PZNI Mar. Ecol. 21:113–127.
Jaramillo, E., De la Huz, R., Duarte, C., & Contreras, H. (2006). Algal wrack deposits and macroinfaunal
arthropods on sandy beaches of the Chilean coast. Revista Chilena de Historia Natural, 79(3).
Jędrzejczak M.F. (2002) Stranded Zostera marina L. vs wrack fauna community interactions on a Baltic
sandy beach (Hel, Poland): a short-term pilot study. Part II. Driftline effects of succession changes
and colonization of beach fauna. . Oceanologia, 44, 367-387
Jones, A. R., W. Gladstone and N. J. Hacking (2008). Australian sandy-beach ecosystems and climate
change: ecology and management. Zoologist 34: 190-202
Koop, K., & Lucas, M. I. (1983). Carbon flow and nutrient regeneration from the decomposition of
macrophyte debris in a sandy beach microcosm. In Sandy beaches as ecosystems (pp. 249-262).
Springer Netherlands.
Lal, R., Kimble, J., Follett, R. F., & Stewart, B. A. (2001). Assessment methods for soil carbon (No. 631.41
A846a). Boca Raton, US: CRC Press.
Lastra, M., and McLachlan, A. (1996). Spatial and temporal variations in recruitment of Donax

32
serra Roding (Bivalvia: Donacidae) on an exposed sandy beach of South Africa. Rev. Chilena Hist.
Nat. 69:631–639.
Lercari, D., and Defeo, O. (2003). Variation of a sandy beach macrobenthic community along a
human-induced environmental gradient. Estuar. Coast. Shelf Sci. 58S:17–24.
Lucrezi, S., T. A. Schlacher and W. Robinson (2010). Can storms and shore armouring exert additive
effectson sandy-beach habitats and biota? Marine and Freshwater Research 61(9): 951-962.
Masselink G. & Hughes M.G. (2003) Introduction to coastal processes and geomorphology. Oxford
University, New York.
Masselink, G., & Short, A. D. (1993). The effect of tide range on beach morphodynamics and morphology: a
conceptual beach model. Journal of Coastal Research, 785-800.
Marsden, I. D. (1991) Kelp sandhopper interactions on a sand beach in New Zealand 1. Drift composition
and distribution, Journal of Experimental Marine Biology and Ecology 152, 61-74.
Martinez-Ribes, L., Basterretxea, G., Palmer, M., & Tintoré, J. (2007). Origin and abundance of beach
debris in the Balearic Islands. Scientia Marina, 71(2), 305-314.
McArdle, S., and McLachlan, A. (1991). Dynamics of the swash zone and effluent line on sandy
beachs. Mar. Ecol. Prog. Ser. 76:91–99.
McArdle, S., and McLachlan, A. (1992). Sand beach ecology: Swash features relevant to the macrofauna.J.
Coast. Res. 8:398–407.
McClain, M. E., Boyer, E. W., Dent, C. L., Gergel, S. E., Grimm, N. B., Groffman, P. M., ... & McDowell, W.
H. (2003). Biogeochemical hot spots and hot moments at the interface of terrestrial and aquatic
ecosystems. Ecosystems, 6(4), 301-312.
McLachlan, A., Erasmus, T., and Furstenberg, J. P. (1977). Migrations of sandy beach meiofauna. Zool. Afr.
12:257–277.
McLachlan A. (1980) The definition of sandy beaches in relation to exposure: a simple rating system. South
African Journal of Science, 76, 137-138
McLachlan, A., and Harty, B. (1981). Effects of oil on water filtration by exposed sandy beaches.
Mar. Pollut. Bull. 12:374–378.
McLachlan, A. (1982). A model for the estimation of water filtration and nutrient regeneration by exposed
sandy beaches. Marine Environmental Research, 6(1), 37-47.
McLachlan A. (1983) Sansy beach ecology – a review. In: Sandy beaches as ecosystems. McLachlan A, T
(ed), pp.321-380. Junk, The Hague.
McLachlan, A. (1985). The biomass of macro- and interstitial fauna on clean and wrack-covered
beaches in Western Australia. Estuar. Coast. Shelf Sci. 21:587–599.
McLachlan A. (1988) Dynamics of an exposed beach/dune coast, Algoa Bay, southeast Africa. Journal of
Coastal Research, 3, 91-95

33
McLachlan, A., Dugan, J. E., Defeo, O., Ansell, A. D., Hubbard, D. M., Jaramillo, E., and Penchaszadeh, P.
(1996a). Beach clam fisheries. Oceanogr. Mar. Biol. Ann. Rev. 34:163–232.
Mclachlan, A., Kerley, G., and Rickard, C. (1996b). Ecology and energetics of slacks in the Alexandria
coastal dunefield. Landscape Urban Plan. 34:267–276.
McLachlan, A., and Dorvlo, A. (2005). Global patterns in sandy beach macrofauna communities. J. Coast.
Res. 21:674–687.
McLachlan A. & Brown A.C. (2006) The ecology of sandy shores. Second edn. Elsevier, Amsterdam
McGwynne, L., McLachlan, A., & Rust, I. C. (1992). Ecology and management of sandy coasts (No. 30).
Institute for Coastal Research UPE.
Moore, W. S. (1996). Large groundwater inputs to coastal waters revealed by 226Ra enrichments. Nature,
380(6575), 612.
Noy-Meir, I. (1979). Structure and function of desert ecosystems. Israel Journal of Botany, 28(1), 1-19.
Nordstrom, K. R., Jackson, N. L., Klein, A. H. F., Sherman, D. J. & Hesp, P. A. (2006) Offshore aeolian
transport across a low foredune on a developed barrier island, Journal of Coastal Research 22,
1260-1267.
Ochieng, C. A. & Erftemeijer, P. L. A. (1999) Accumulation of seagrass beach cast along the Kenyan coast: a
quantitative assessment, Aquatic Botany 65, 221-238.
Pearse, A. S., Humm, H. J., and Wharton, G. W. (1942). Ecology of sandy beaches at Beaufort,
North Carolina. Ecol. Monogr. 12:135–190.
Raffaelli, D., & Hawkins, S. J. (2012). Intertidal ecology. Springer Science & Business Media.
Rauch, M., Denis, L., & Dauvin, J. C. (2008). The effects of Phaeocystis globosa bloom on the dynamics of
the mineralization processes in intertidal permeable sediment in the Eastern English Channel
(Wimereux, France). Marine pollution bulletin, 56(7), 1284-1293.
Rieper-Kirchner, M. (1989). Microbial degradation of North Sea macroalgae: field and laboratory studies.
Botanica Marina, 32(3), 241-252.
Robertson, A. I., & Lenanton, R. C. J. (1984). Fish community structure and food chain dynamics in the
surf-zone of sandy beaches: the role of detached macrophyte detritus. Journal of Experimental Marine
Biology and Ecology, 84(3), 265-283.
Rocha, C. (2008). Sandy sediments as active biogeochemical reactors: compound cycling in the fast lane.
Aquatic Microbial Ecology, 53(1), 119-127.
Rodil, I. F., and Lastra, M. (2003). Environmental factors affecting benthic macrofauna along a
gradient of intermediate sandy beaches in northern Spain. Estuar. Coast. Shelf Sci. 61:37–44.
Rusch, A., Huettel, M., Wild, C., & Reimers, C. E. (2006). Benthic oxygen consumption and organic matter
turnover in organic-poor, permeable shelf sands. Aquatic Geochemistry, 12(1), 1-19.
Sahagian, D. & J. Melack. 1998. Global wetlands distribution and functional characterization.
Trace changes and hydrologic cycle. Report from joint GAIM, BAHC, IGBP-DIS, IGAC and

34
LUCC workshop, Santa Barbara, CA USA. IGBP report 46.
Seco-Hernández, R. 2004. Geomorfología. Universidad de la Habana. Cuba.
Schlacher TA, Dugan J, Schoeman DS, Lastra M, Jones A, Scapini F, McLachlan A, Defeo O (2007) Sandy
beaches at the brink. Divers Distrib 13:556-560
Schlacher, T. A., Schoeman, D. S., Dugan, J., Lastra, M., Jones, A., Scapini, F., & McLachlan, A. (2008).
Sandy beach ecosystems: key features, sampling issues, management challenges and climate
change impacts. Marine ecology, 29(s1), 70-90.
Stenton-Dozey, J. M. E., & Griffiths, C. L. (1983). The fauna associated with kelp stranded on a sandy
beach. In Sandy beaches as ecosystems (pp. 557-568). Springer Netherlands.
Short A.D. (1999) Handbook of beach and shore morphodynamics. Wiley and Sons, London.
Schwinghamer, P. (1981). Characteristic size distributions of integral benthic communities. Can.
J. Fish. Aquatic Sci. 38:1255–1263.
Schimel, D. S. (1995). Terrestrial ecosystems and the carbon cycle. Global change biology, 1(1), 77-91.
Spilmont, N., Migné, A., Lefebvre, A., Artigas, L. F., Rauch, M., & Davoult, D. (2005). Temporal variability
of intertidal benthic metabolism under emersed conditions in an exposed sandy beach (Wimereux,
eastern English Channel, France). Journal of Sea Research, 53(3), 161-167.
Tinley, K. L. (1985). Coastal dunes of South Africa. National Scientific Programmes Unit: CSIR.
Urban-Malinga, B., & Burska, D. (2009). The colonization of macroalgal wrack by the meiofauna in the
Arctic intertidal. Estuarine, Coastal and Shelf Science, 85(4), 666-670.
Wade, B. A. (1967). Studies on the biology of the West Indian beach clam, Donax denticulatus Linné. 1.
Ecology. Bulletin of Marine Science, 17(1), 149-174.
Wang, Y. P., & Polglase, P. J. (1995). Carbon balance in the tundra, boreal forest and humid tropical forest
during climate change: scaling up from leaf physiology and soil carbon dynamics. Plant, Cell &
Environment, 18(10), 1226-1244.
Warwick, R. W. (1984). Species size distribution in marine benthic communities. Oceologia 61:32–41.
Wells, J. C., & Cole, T. J. (2002). Birth weight and environmental heat load: A between‐population analysis.
American journal of physical anthropology, 119(3), 276-282.
Wright, L. D., & Short, A. D. (1984). Morphodynamic variability of surf zones and beaches: a synthesis.
Marine geology, 56(1-4), 93-118.
Xu, M., & Qi, Y. (2001). Spatial and seasonal variations of Q 10 determined by soil respiration
measurements at a Sierra Nevadan forest. Global Biogeochemical Cycles, 15(3), 687-696.
ZoBell, C. E. (1971) Drift seaweeds on San Diego County beaches, In: The biology of giant kelp beds
(Macrocystis) in California, North, W.J. (ed), Nova Hedwigia 32, 269-314.

35

36
INTRODUCCIÓN
La actividad metabólica informa sobre los ciclos de materia y flujos de energía a través de los
ecosistemas, por lo que proporciona una de las visiones más integradoras de su funcionamiento (Enquist et
al., 2003). Comprender estos flujos a través de los ecosistemas es fundamental para resolver muchas
preguntas sobre el efecto del cambio global sobre la biología y la ecología.
Las playas tienen un papel fundamental en los ciclos de carbono y nitrógeno tanto a nivel local
como global a través de su papel como recicladoras de la materia orgánica. Sin embargo, existen pocos
estudios destinados a comprender sus aspectos funcionales (Spilmont et al., 2005; Williams & del Giorgio,
2005). Con el fin de obtener mayor conocimiento sobre los ciclos de carbono y nutrientes en la zona de
transición tierra-océano, se necesita información sobre el metabolismo neto de las playas (Jickells & Rae
1997; Anschutz et al., 2009). Una de las principales características ecológicas de las playas es su reducida
producción primaria in situ en comparación con los ecosistemas vecinos (McLachlan & Brown, 2006). La
cadena trófica de las playas se basa en subsidios alóctonos denominados wrack. Este término se refiere a la
materia orgánica que llega a la playa en forma, sobre todo, de varamientos de macroalgas y plantas
vasculares. La procedencia de estos inputs son el océano abierto, la costa rocosa, las marismas, etc.
(Griffiths et al., 1983; Colombini et al., 2000; Dugan et al., 2003; Lastra et al., 2008).
Una vez el wrack llega a la playa, es sometido a una variedad de procesos: consumo in situ de los
herbívoros (macro, meio y microfauna), degradación, desecación y exportación por las mareas y corrientes
(Griffiths & Stenton-Dozey 1981; Griffiths et al., 1983; Inglis, 1989; Jedrzejczak, 2002; Orr et al., 2005;
Lastra et al., 2006). Durante el proceso de degradación se produce la fragmentación de partículas de gran
tamaño y el catabolismo de los compuestos orgánicos (Satchell, 1974; Álvarez, 2005). Posteriormente, las
bacterias, oxidan estos compuestos obteniendo compuestos inorgánicos simples (CO2, H2S, NH4+, PO4
-3,
H2O). El CO2, el nitrógeno inorgánico y los fosfatos (NH4++NOx
ˉ y PO4
-3) son buenos indicadores de la
actividad biológica (micro, meio y macrofauna) e intensidad de descomposición de materia orgánica. El
CO2 describe el ciclo de la materia orgánica en los ecosistemas, dado que integra todas las posibles fuentes
de materia orgánica así como la variabilidad temporal en las tasas de aporte. Los nutrientes a su vez,
aportan información acerca de la actividad biogeoquímica de la playa.
Todos los cambios que se producen durante el proceso de degradación, convierten a los parches de
algas en ―hot spots‖. Caracterizados por una elevada actividad metabólica en comparación con su entorno
(Coupland et al., 2007; McClain et al., 2003). Esto implica una aceleración en los procesos metabólicos de
la playa, lo que se traduce en un incremento de la actividad de las comunidades micro, meio y
macrofaunística. Actualmente se están destinando muchos esfuerzos a mejorar las técnicas de estimación

37
de la respiración del suelo y a aumentar el conocimiento de las interacciones entre la degradación del wrack
con las variables ambientales (Martínez-García et al., 2013).
La variabilidad en las emisiones de CO2 está influenciada por factores bióticos y abióticos
(Buchmann, 2000; Curiel-Yuste et al., 2003; Concilio et al., 2006; Irvine et al., 2007; Almagro et al.,
2009). Los factores abióticos más influyentes son la temperatura y la humedad (Raich y Schlesinger, 1992;
Raich & Potter 1995; Davidson et al., 1998; Pei-Li Shiet et al., 2006). La temperatura determina la
velocidad de metabolización de la materia orgánica ya que regula las velocidades enzimáticas mediadas por
la comunidad microbiana hasta alcanzar el óptimo de temperatura donde ocurre la actividad máxima. La
relación directa entre el flujo de CO2 y los nutrientes con la temperatura está bien documentada (Raich &
Schlesinger, 1992; Hanisak, 1993; Lloyd & Taylor, 1994; Fang & Moncrieff, 2001). Dentro de los rangos
térmicos favorables, existe un aumento positivo, a menudo exponencial, de la tasa metabólica a medida que
aumenta la temperatura (Gillooly et al., 2001). Cabe señalar que cada especie microbiana y cepa tiene su
propia temperatura óptima y ésta puede variar dependiendo de la interacción entre factores (temperatura,
carga hidráulica, porosidad y naturaleza de la materia orgánica) (Kadlec & Reddy, 2001). La comunidad
microbiana es la responsable de la mayor parte de la mineralización de la materia orgánica (McLachlan et
al., 1981; Koop et al., 1982a,b; Koop & Griffiths, 1982; Newell et al., 1982; Stenton-Dozey & Griffiths, 1983;
McLachlan, 1985; Colombini & Chelazzi, 2003). Así pues, es evidente que aquellos factores ambientales que
afectan al crecimiento y al metabolismo de los organismos, modificarán a su vez la velocidad de
transformación de la materia orgánica (Davidson et al., 1998; Subke et al., 2003; Pei-Li Shi et al., 2006).
Por lo tanto, el objetivo de este trabajo es investigar la relación existente entre la temperatura y la
velocidad de degradación de los arribazones de macroalgas, y conocer cómo afectan a los ciclos
biogeoquímicos que se producen en playas expuestas de la costa de Galicia. Para ello se llevó a cabo un
seguimiento anual de tres playas con distinto estado morfodinámico y distinto grado de subsidio algal,
donde se analizaron diferentes factores físicos (ej. temperatura, humedad) así como las concentraciones de
nutrientes (nitrógeno inorgánico y fosfatos) y los flujos de CO2. Para alcanzar el objetivo del trabajo se
plantearon las siguientes HIPÓTESIS: (1) la temperatura y la humedad junto con la biomasa algal,
influyen directamente sobre la liberación de nutrientes y de CO2, (2) esta relación varía en el espacio y en el
tiempo.

38
MATERIAL Y MÉTODOS
Área de estudio
El estudio de la actividad metabólica se llevó a cabo durante un año en tres playas expuestas de la
costa oeste de Galicia (NO España) (Figura 1). Esta costa se caracteriza por estar formada por rías. Son
consideradas como uno de los ecosistemas marinos de mayor diversidad biológica del planeta debido a su
gran fertilidad y por sus especiales características (situación geográfica, orientación y dinámica). El
responsable de esta fertilidad es el fenómeno de afloramiento de aguas profundas (Blanton et al. 1984), que
típicamente fertiliza la costa de abril a septiembre. Las playas estudiadas, son playas arenosas expuestas a
la acción del oleaje. Las condiciones mareales fueron meso-macromareal con un rango de 3.5 a 4 m en
mareas vivas. Abra está ubicada en la Ensenada de Carreira en el margen suroeste de la Ría de Vigo (42º
9´47‖ N; 8º49´43.12‖ W). Pertenece al municipio de Nigrán (Pontevedra). Es una playa urbana sin paseo
marítimo y su estado morfológico es intermedia-reflectiva con arena gruesa. Es una playa ventosa con
oleaje moderado. Ladeira y Balieiros pertenecen al municipio de Ribeira (A Coruña) (LAD: 42º 34´33‖ N;
9º 3´16‖ W y BAL: 42º 34´39.28‖ N; 9º 4´45.21‖ W). Ladeira es una playa semiurbana sin paseo. Es
disipativa con tamaño de grano constituido por arena media. El estado habitual de la mar es oleaje
moderado. Está integrada en el Parque Natural del Complejo Dunar de Corrubedo. Balieiros es una playa
aislada sin paseo marítimo. Es reflectiva, con arena gruesa, ventosa y el estado habitual de la mar es oleaje
fuerte. Se encuentra en el límite de los términos municipales de Porto do Son y Ribeira. Las condiciones
mareales de las tres playas fueron meso-macromareal, mareas vivas de 3.5 a 4 m (Figuras 1 y 2).

39

40
Caracterización de las playas
Las características físicas y morfodinámicas fueron medidas como se detalla a continuación y así
poder clasificarlas según los parámetros que se muestran en la Tabla 1.
Las dimensiones de la playa se obtuvieron de la base de datos del Ministerio de Agricultura y
Pesca, Alimentación y Medio Ambiente (www.mapama.gob.es). La anchura se midió desde la base de la
duna hasta el nivel de la bajamar en mareas vivas.
El grado de exposición al oleaje se determinó mediante la clasificación de McLachlan (1980).
Reflectiva, intermedia y disipativa.
El estado morfodinámico se determinó mediante la clasificación de Short (1999b). Para ello fue
necesario medir el periodo y altura de ola. El periodo fue medido con un cronómetro el intervalo de
tiempo entre las rompientes de las olas (n=30). La altura de la ola fue medida con un estadal (Figura 5).
Las medidas se realizaron desde el punto más bajo de la playa haciendo coincidir la cresta de la ola en el
momento de rotura con el horizonte (n=30).
La pendiente y el perfil de la playa se determinaron mediante el método Emery con dos
estadales (Emery 1961). Se expresa como su inversa (1/pte). Los valores altos representan pendientes
suaves y los valores bajos, pendientes pronunciadas (Figura 3).
El tamaño medio de grano (T.M.G) (mm) se midió con un LS 13 320 MW, analizador de
tamaño de partícula de difracción láser.
La cobertura algal se midió todos los meses sobre un transecto perpendicular a la orilla usando
el método interceptor lineal; los valores se dan en porcentaje sobre el área total de la playa (Dugan et al.,
2003; Barreiro et al., 2013).
Diseño del muestreo
Los muestreos se llevaron a cabo mensualmente desde marzo de 2012 hasta marzo de 2013 durante
la bajamar de mareas vivas (abril carece de datos debido a las adversas condiciones meteorológicas). En
cada playa se establecieron seis transectos perpendiculares a la línea de costa distribuidos aleatoriamente a
lo largo de los 100 m centrales con una separación mínima de al menos 5 m. En cada transecto se
diferenciaron tres zonas según los niveles de marea: el nivel superior (N3) corresponde con la línea de
marea de la última marea viva alta del mes anterior, el nivel medio (N2) es la línea de la última marea del
mismo mes de estudio y el nivel inferior (N1) es el límite superior de la zona de saturación. La intersección
ente los transectos y los niveles de marea fueron los puntos de muestreo (n=18) (Figura 4).

41
Figura 3. A) Cálculo del grado de exposición al oleaje y esquema de la pendiente, B) medida del ancho de la playa, se
muestra también el drift line y el transecto sobre el que se mide la cobertura algal, C) cálculo de la pendiente mediante
el método Emery y D) el método interceptor lineal para el cálculo de la cobertura.
Figura 4. Esquema del diseño de muestreo para cada playa. Las letras indican los transectos
perpendiculares a la línea de costa. Los puntos de corte con los niveles de marea, son los puntos de
estudio (n=18).

42
Toma de las muestras
En cada mes, en cada punto de estudio, se tomaron medidas in situ del flujo de CO2 (µmol C m-2 s-
1); para ello, se utilizó un medidor de flujo difuso con un detector LI-COR de CO2 (con un volumen de la
campana de acumulación de 2.76 10-3 m3 y un área en su base de 3.14 10-2 m-2). Una vez medido el flujo de
CO2, las algas se metieron en bolsas de plástico, en el caso de que las hubiese. A continuación, se llenaron
dos botes de 30 cm3 de sedimento para determinar el porcentaje de humedad del suelo y el tamaño medio
de grano (mensualmente, n= 12) y la concentración total de nutrientes (Nitrógeno inorgánico total (TNI) y
PO4-3 µM g-1) (estacionalmente, n= 5) (en N2 y N3). En el nivel inferior (N1), se tomaron muestras del agua
de resurgencia en botellas de 30 cm3 para analizar las concentraciones de nutrientes (TNI y PO4-3 µM)
(estacionalmente, n= 5). La temperatura del sedimento se midió introduciendo un termómetro a 5 cm de
profundidad en cada punto de estudio. Las muestras de macrofauna se tomaron estacionalmente; se
introdujo en el sedimento un corer de 10 cm de diámetro a una profundidad de 10 cm (157 cm3) de forma
aleatoria alrededor del punto de muestreo (n=5). Las muestras fueron tamizadas in situ a través de una
malla de 1 mm (Figura 5). Posteriormente se transportaron al laboratorio donde fueron conservadas en una
solución de agua de mar y formol al 4%. Los individuos se determinaron al nivel taxonómico más bajo
posible. Una vez en el laboratorio, todas las muestras se congelaron y almacenaron a -20ºC hasta su
posterior análisis.
Figura 5. Fotografías del material usado en la recogida de las muestras: ―A‖
medidor de flujo difuso con un detector LI-COR de CO2, ―B‖ corer para la recogida
de sedimento para macrofauna y ―C‖ bolsa de tamizaje de malla para retener la
macrofauna.

43
Análisis de las muestras
Las muestras de algas se descongelaron y se separaron del resto de material varado. Se secaron en
la estufa a 60º C durante 48 horas para obtener la relación Peso fresco: Peso seco. De esta relación se
obtuvo el porcentaje de humedad del alga. A continuación se molieron y se tomó una alícuota de 5g de peso
seco que se calcinó en la mufla durante 4h a 500ºC. De esta forma se obtuvo la biomasa algal (g peso seco
libre de cenizas) (g PSLC). El porcentaje de humedad del sedimento se obtuvo a partir de la pérdida de peso
tras el secado de las muestras durante 72 horas a 60ºC. Los nutrientes (TNI y PO4-3) de N2 y N3, se
extrajeron de 5 g de sedimento según la normativa española (UNE 77306, 1999). Después de la extracción,
las muestras acuosas se congelaron en tubos Falcon junto con las botellas de agua de resurgencia de N1.
Posteriormente fueron llevadas a la ―Unidad de Análisis Elemental‖ de la Universidad de Vigo para su
análisis. Las muestras se analizaron con un Autoanalizador de Flujo continuo Segmentado. La
determinación se realiza de forma directa frente a patrones acuosos.
Análisis estadístico
Para determinar la variación asociada del CO2 (µmol C m-2 s-1) al tipo de playa, mes y nivel de marea
se realizó un análisis de varianza (ANOVA) ortogonal de tres vías con ―playa‖ (3 niveles), ―mes‖ (12 niveles)
y ―nivel de marea‖ (3 niveles) como factores fijos del análisis (paquete estadístico Win GMAV5; EICC,
Universidad de Sidney). Los seis transectos de cada playa fueron usados como réplicas (n=6). De igual
forma se procedió para los nutrientes (TNI y PO4-3) siendo en este caso ―mes‖ (5 niveles). El análisis de N1
se realizó por separado de N2 y N3 ya que los nutrientes de N1 fueron analizados en muestras de agua de
resurgencia mientras que N2 y N3 de sedimento (µM y µM g-1 sed.). Previamente los datos fueron
transformados para cumplir las asunciones de normalidad y homogeneidad de varianzas, evaluadas con los
test de Sapiro-Wilk y Cochran respectivamente. A posteriori, se realizó un análisis de comparaciones
múltiples mediante el test de Student-Newman-Keul (SNK) (α= 0.05) cuando hubo diferencias
significativas entre los factores.
Para la estimación de la las variables (CO2, nutrientes y Carbono biopolimérico) en función de los
factores físicos y biológicos (temperatura, humedad del sedimento, humedad del alga y biomasa algal) se
realizaron regresiones múltiples (paquete estadístico R 3.1.1).

44
RESULTADOS
Caracterización de las playas
Las medidas que se realizaron sobre cada playa permitieron definir sus características físicas como muestra
la tabla 1. El arenal con mayor superficie fue Ladeira y el más pequeño, Abra. Aunque son todas playas
expuestas, la que recibe mayor energía del oleaje e impacto sobre la orilla es Balieiros ya que es reflectiva y
por lo tanto es la que tiene mayor pendiente. Por el contrario, Ladeira, es casi el doble de ancha que
Balieiros por lo que el oleaje se disipa a lo largo del swash y a lo largo de sus 5 km de costa. Así pues, la
pendiente es muy suave. Esta morfodinámica es la que determina el tamaño de arena grueso de Balieiros y
la arena fina de Ladeira. En un punto intermedio se encuentra la playa de Abra. Es la más reducida pero su
grado de exposición más bajo que las otras dos anteriores, le permite tener una pendiente intermedia
(Figura 5) y un tamaño de grano grueso aunque no tanto como Balieiros. En la figura 5 se puede observar el
resultado de la interacción de todos estos elementos, el perfil de las playas.
Las características morfodinámicas determinan las biológicas, concretamente en la cantidad de material de
arribazón que llega a cada una de ellas. Abra es en términos generales la que recibe mayores cantidades de
biomasa algal y por lo tanto, tiene mayor cobertura. Ladeira y Balieiros son playas con bajo aporte de
material de arribazón aunque los valores pueden variar a lo largo del año.
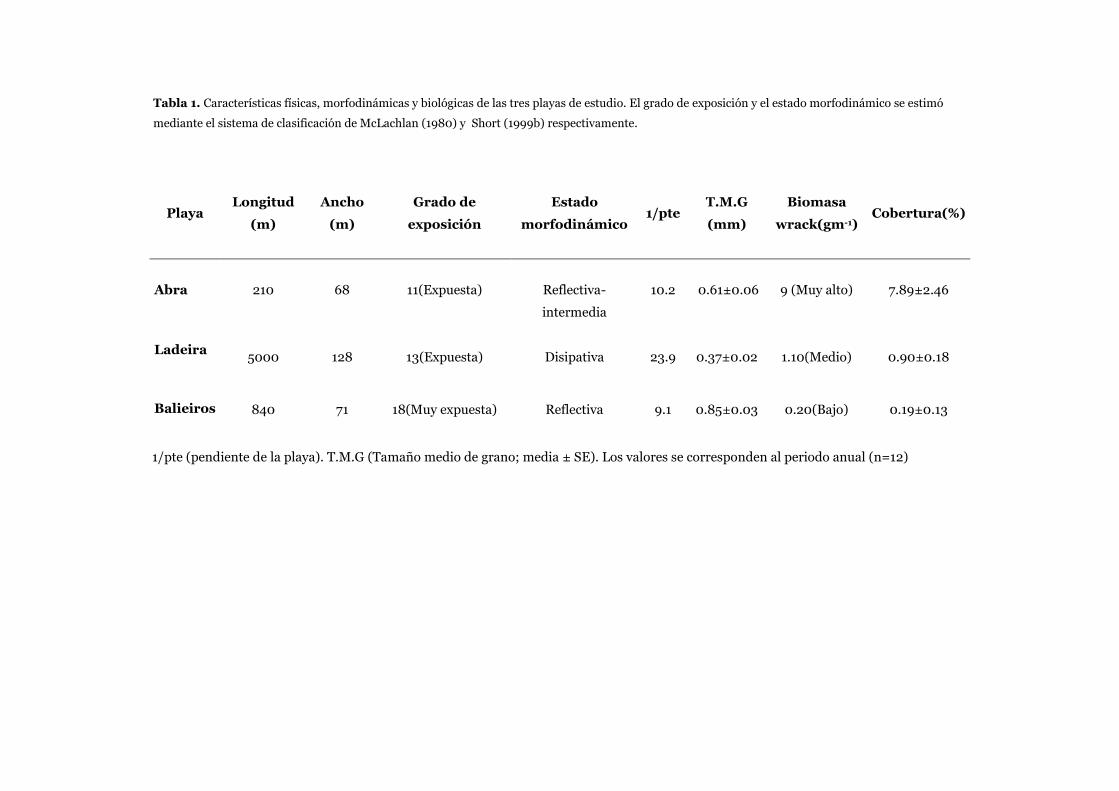
1/pte (pendiente de la playa). T.M.G (Tamaño medio de grano; media ± SE). Los valores se corresponden al periodo anual (n=12)
Playa Longitud
(m)
Ancho
(m)
Grado de
exposición
Estado
morfodinámico 1/pte
T.M.G
(mm)
Biomasa
wrack(gm-1) Cobertura(%)
Abra
210
68
11(Expuesta)
Reflectiva-
intermedia
10.2
0.61±0.06
9 (Muy alto)
7.89±2.46
Ladeira
5000
128
13(Expuesta)
Disipativa
23.9
0.37±0.02
1.10(Medio)
0.90±0.18
Balieiros
840
71
18(Muy expuesta)
Reflectiva
9.1
0.85±0.03
0.20(Bajo)
0.19±0.13
Tabla 1. Características físicas, morfodinámicas y biológicas de las tres playas de estudio. El grado de exposición y el estado morfodinámico se estimó
mediante el sistema de clasificación de McLachlan (1980) y Short (1999b) respectivamente.
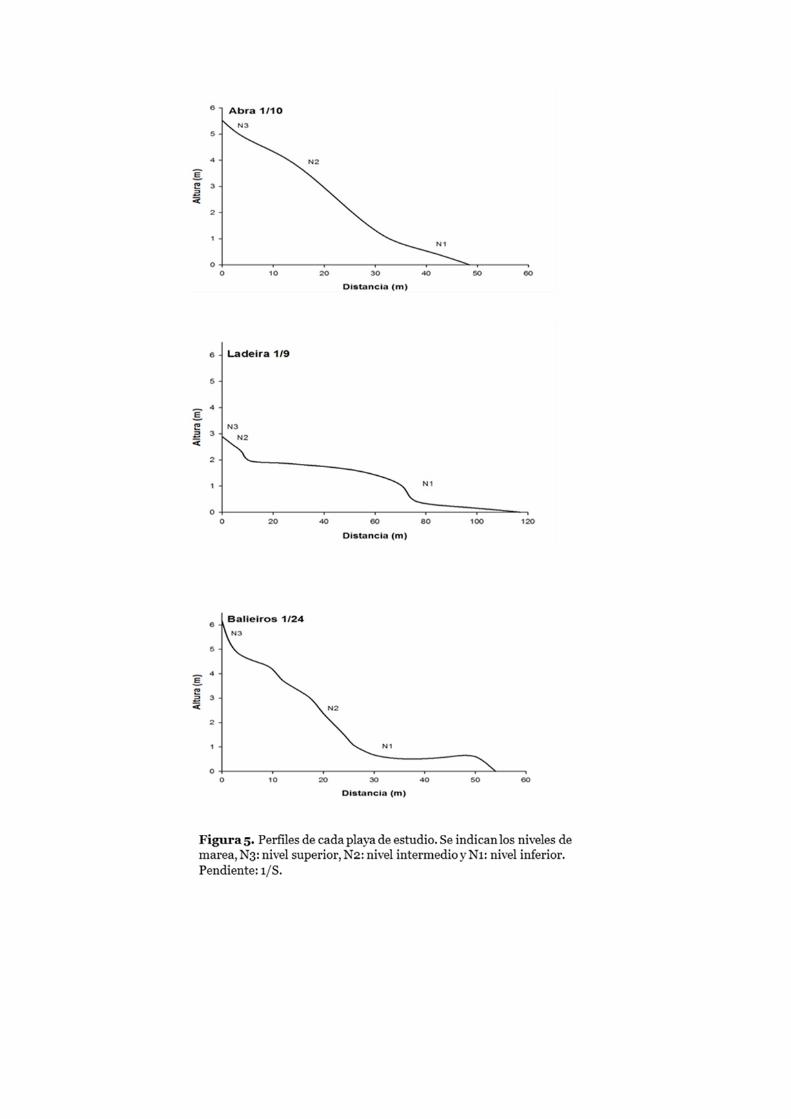
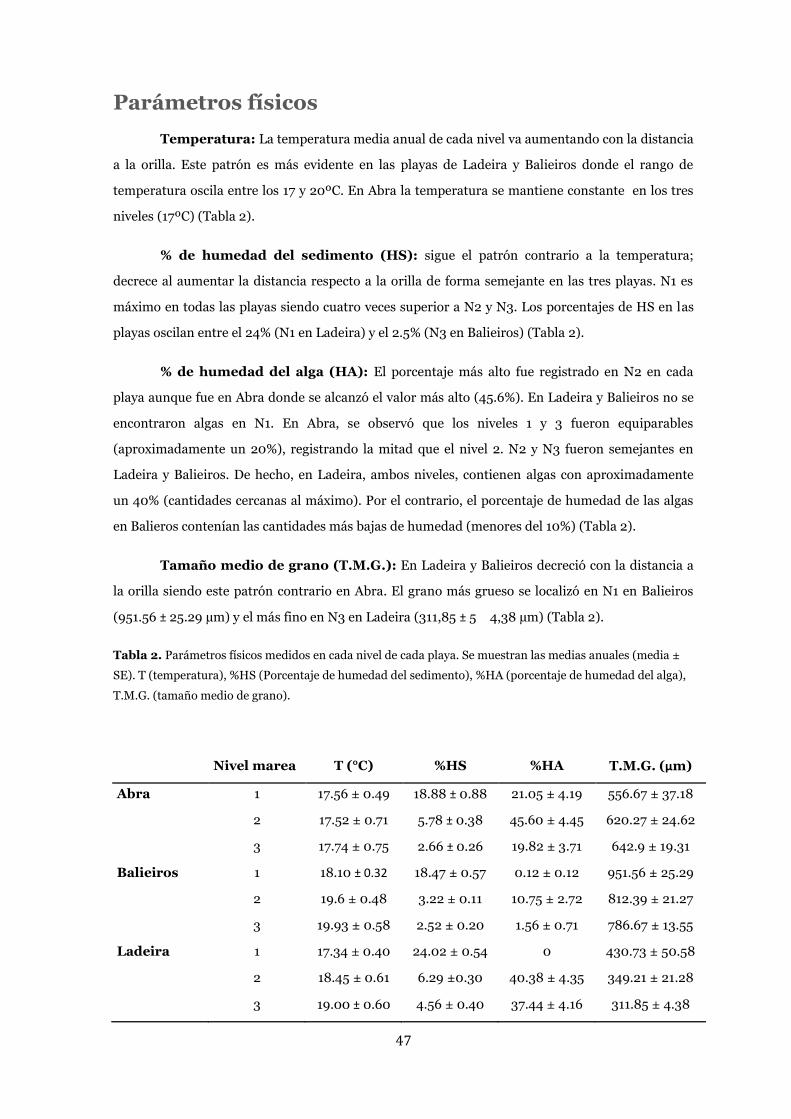
47
Parámetros físicos
Temperatura: La temperatura media anual de cada nivel va aumentando con la distancia
a la orilla. Este patrón es más evidente en las playas de Ladeira y Balieiros donde el rango de
temperatura oscila entre los 17 y 20ºC. En Abra la temperatura se mantiene constante en los tres
niveles (17ºC) (Tabla 2).
% de humedad del sedimento (HS): sigue el patrón contrario a la temperatura;
decrece al aumentar la distancia respecto a la orilla de forma semejante en las tres playas. N1 es
máximo en todas las playas siendo cuatro veces superior a N2 y N3. Los porcentajes de HS en las
playas oscilan entre el 24% (N1 en Ladeira) y el 2.5% (N3 en Balieiros) (Tabla 2).
% de humedad del alga (HA): El porcentaje más alto fue registrado en N2 en cada
playa aunque fue en Abra donde se alcanzó el valor más alto (45.6%). En Ladeira y Balieiros no se
encontraron algas en N1. En Abra, se observó que los niveles 1 y 3 fueron equiparables
(aproximadamente un 20%), registrando la mitad que el nivel 2. N2 y N3 fueron semejantes en
Ladeira y Balieiros. De hecho, en Ladeira, ambos niveles, contienen algas con aproximadamente
un 40% (cantidades cercanas al máximo). Por el contrario, el porcentaje de humedad de las algas
en Balieros contenían las cantidades más bajas de humedad (menores del 10%) (Tabla 2).
Tamaño medio de grano (T.M.G.): En Ladeira y Balieiros decreció con la distancia a
la orilla siendo este patrón contrario en Abra. El grano más grueso se localizó en N1 en Balieiros
(951.56 ± 25.29 µm) y el más fino en N3 en Ladeira (311,85 ± 5 4,38 µm) (Tabla 2).
Tabla 2. Parámetros físicos medidos en cada nivel de cada playa. Se muestran las medias anuales (media ±
SE). T (temperatura), %HS (Porcentaje de humedad del sedimento), %HA (porcentaje de humedad del alga),
T.M.G. (tamaño medio de grano).
Nivel marea T (°C) %HS %HA T.M.G. (µm)
Abra 1 17.56 ± 0.49 18.88 ± 0.88 21.05 ± 4.19 556.67 ± 37.18
2 17.52 ± 0.71 5.78 ± 0.38 45.60 ± 4.45 620.27 ± 24.62
3 17.74 ± 0.75 2.66 ± 0.26 19.82 ± 3.71 642.9 ± 19.31
Balieiros 1 18.10 ± 0.32 18.47 ± 0.57 0.12 ± 0.12 951.56 ± 25.29
2 19.6 ± 0.48 3.22 ± 0.11 10.75 ± 2.72 812.39 ± 21.27
3 19.93 ± 0.58 2.52 ± 0.20 1.56 ± 0.71 786.67 ± 13.55
Ladeira 1 17.34 ± 0.40 24.02 ± 0.54 0 430.73 ± 50.58
2 18.45 ± 0.61 6.29 ±0.30 40.38 ± 4.35 349.21 ± 21.28
3 19.00 ± 0.60 4.56 ± 0.40 37.44 ± 4.16 311.85 ± 4.38

48
Macrofauna
La macrofauna se midió en términos de abundancia (nº individuos m-2). En la tabla 3 se
puede observar la cantidad de individuos existentes por cada grupo en cada mes estudiado. El
grupo más abundante fue el de los anfípodos en junio con 2030 individuos m-2 superando con
creces al de poliquetos e isópodos (727 y 280 individuos m-2 respectivamente). En general, a lo
largo del año, se detectó una alta biodiversidad pero con bajas abundancias exceptuando los
valores anteriormente comentados.
Tabla 3. Clasificación de los grupos taxonómicos en cada mes de estudio. Los valores se dan en abundancia
(nº individuos m-2).
Grupo Marzo Junio Septiembre Diciembre Marzo´13
Anfípodos 3 2030 3 3 2
Isópodos 0 280 0 73 0
Coleópteros 0 8 4 1 0
Dípteros 5 4 5 2 1
Misidáceos 0 9 0 0 2
Poliquetos 72 4 727 9 25
Nemertinos 17 0 3 1 2
Artrópodos 0 0 0 0 1
Los valores resaltados corresponden a las abundancias más altas.
El nivel 2 registró las mayores abundancias en todas las playas con valores que variaron
desde los 14 (Ladeira) a 389 isópodos m-2 (Abra) y de los 16 a 2696 de anfípodos m-2 (Abra). Por
otro lado, N1 se caracterizó por poseer la mayor abundancia de poliquetos (Tabla 4). Estas
tendencias se vieron afectadas por variaciones espacio-temporales (ANOVA interacción: playa x
nivel x mes, F16, 225= 1.91; p < 0.05). El nivel 2 de Abra fue significativamente superior a N3 y N1
(N2: 62.5 ± 18.31 ind. m-2). Los meses más cálidos (junio, septiembre y marzo de 2013) registraron
las abundancias más altas (256.58 ± 22.40 ind. m-2; n=3). Por el contrario, marzo y diciembre no
superaron los 10 ind. m-2 (SNK test) (Figura 7).
49
Tabla 4. Clasificación de los grupos taxonómicos más abundantes en cada nivel de marea a lo largo del
periodo de estudio en las tres playas.
Anfípodos Isópodos Poliquetos Nemertinos
Marzo 22 4 0 17
Junio 0 0 4 0
NIVEL 1 Septiembre 1 3 718 2
Diciembre 0 0 0 1
Marzo´13 0 0 25 2
Marzo 3 0 0 0
Junio 1801 218 0 0
Abra NIVEL 2 Septiembre 2696 389 0 0
Diciembre 54 15 0 0
Marzo´13 2077 283 0 0
Marzo 124 141 0 0
Junio 218 8 0 0
NIVEL 3 Septiembre 16 1 0 0
Diciembre 285 46 0 0
Marzo´13 40 92 0 0
Marzo 9 1 0 0
Junio 0 2 0 0
NIVEL 1 Septiembre 0 56 0 0
Diciembre 7 1 0 0
Marzo´13 0 3 0 0
Marzo 35 0 0 0
Junio 8 0 0 0
Balieiros NIVEL 2 Septiembre 446 42 0 0
Diciembre 3 0 0 0
Marzo´13 7 0 0 0
Marzo 4 0 0 0
Junio 0 2 0 0
NIVEL 3 Septiembre 17 11 0 0
Diciembre 0 0 0 0
Marzo´13 0 4 0 0
50
Marzo 8 0 72 0
Junio 0 12 0 0
NIVEL 1 Septiembre 5 4 9 1
Diciembre 0 10 9 0
Marzo´13 0 1 0 0
Marzo 29 0 0 0
Junio 3 0 0 0
Ladeira NIVEL 2 Septiembre 3 14 0 0
Diciembre 2 1 0 0
Marzo´13 64 41 0 0
Marzo 3 0 0 0
Junio 0 0 0 0
NIVEL 3 Septiembre 0 0 0 0
Diciembre 15 0 0 0
Marzo´13 0 0 0 0
TOTAL 15992 2810 1674 46
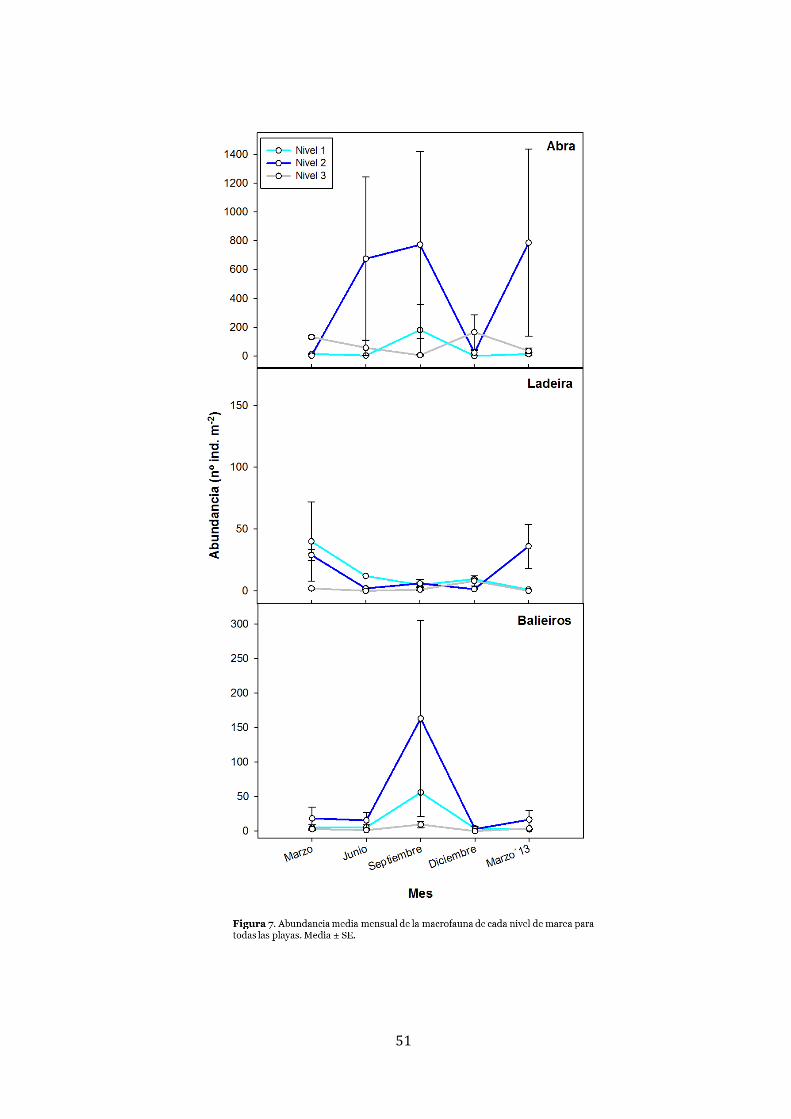
51
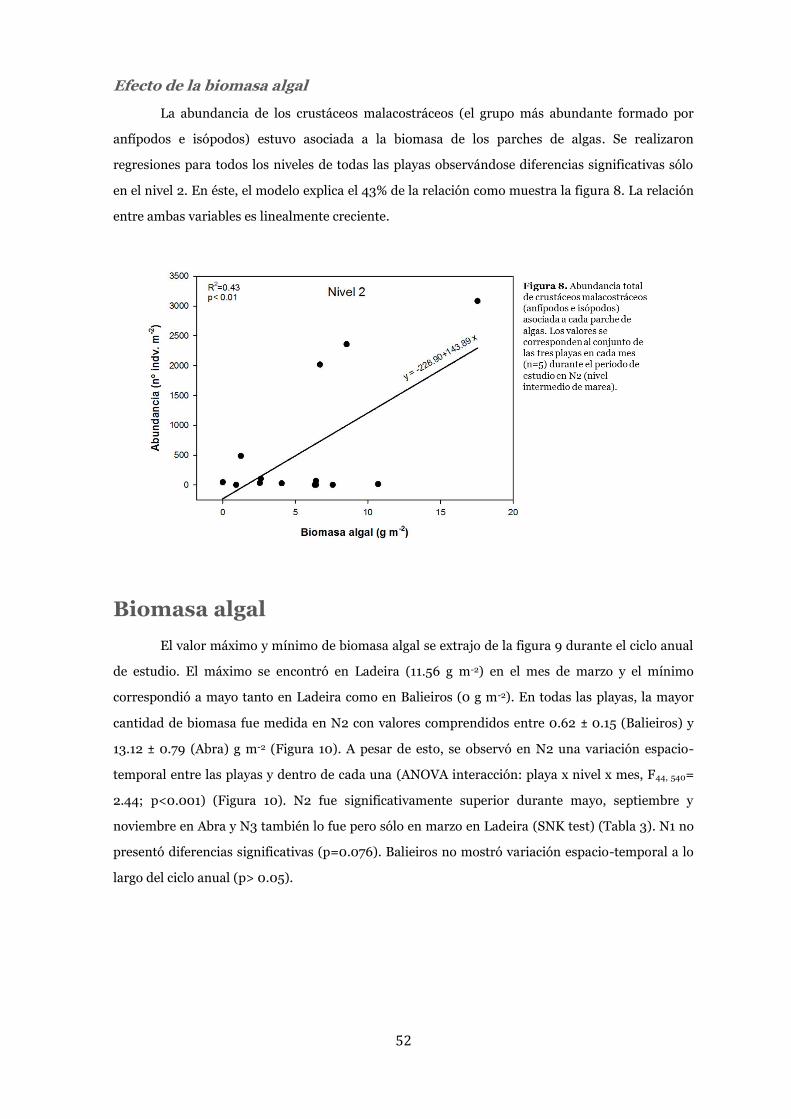
52
Efecto de la biomasa algal
La abundancia de los crustáceos malacostráceos (el grupo más abundante formado por
anfípodos e isópodos) estuvo asociada a la biomasa de los parches de algas. Se realizaron
regresiones para todos los niveles de todas las playas observándose diferencias significativas sólo
en el nivel 2. En éste, el modelo explica el 43% de la relación como muestra la figura 8. La relación
entre ambas variables es linealmente creciente.
Biomasa algal
El valor máximo y mínimo de biomasa algal se extrajo de la figura 9 durante el ciclo anual
de estudio. El máximo se encontró en Ladeira (11.56 g m-2) en el mes de marzo y el mínimo
correspondió a mayo tanto en Ladeira como en Balieiros (0 g m-2). En todas las playas, la mayor
cantidad de biomasa fue medida en N2 con valores comprendidos entre 0.62 ± 0.15 (Balieiros) y
13.12 ± 0.79 (Abra) g m-2 (Figura 10). A pesar de esto, se observó en N2 una variación espacio-
temporal entre las playas y dentro de cada una (ANOVA interacción: playa x nivel x mes, F44, 540=
2.44; p<0.001) (Figura 10). N2 fue significativamente superior durante mayo, septiembre y
noviembre en Abra y N3 también lo fue pero sólo en marzo en Ladeira (SNK test) (Tabla 3). N1 no
presentó diferencias significativas (p=0.076). Balieiros no mostró variación espacio-temporal a lo
largo del ciclo anual (p> 0.05).
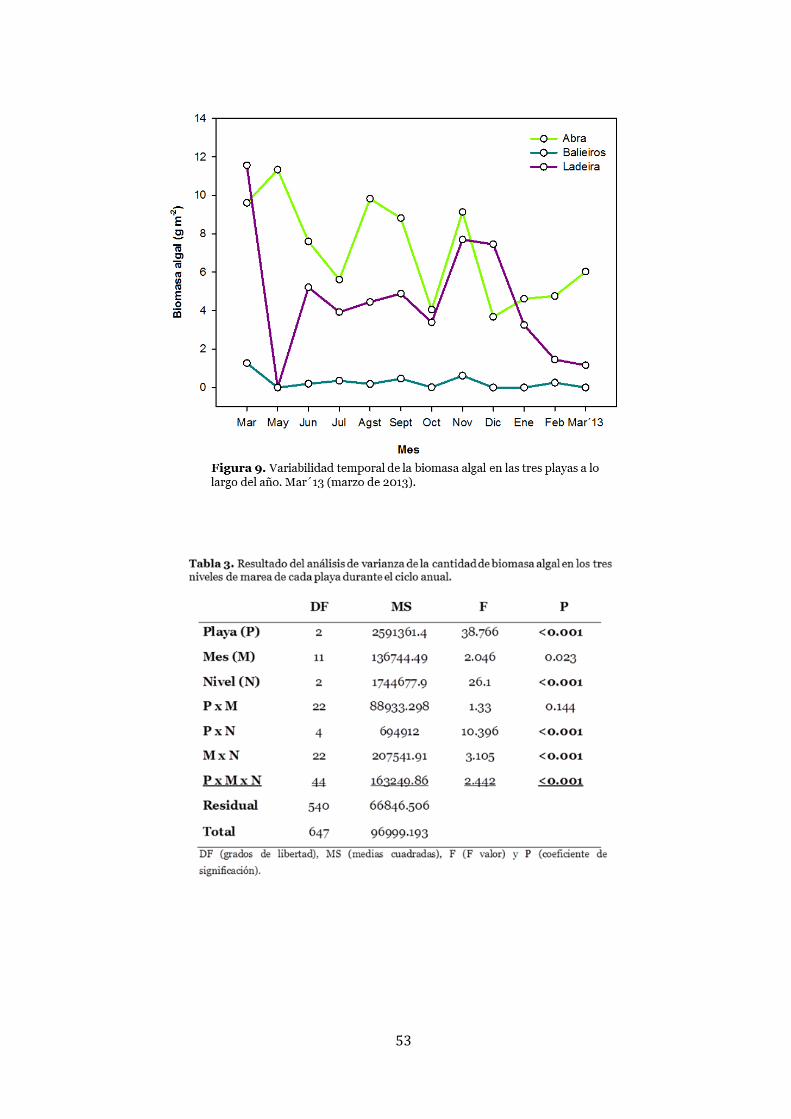
53
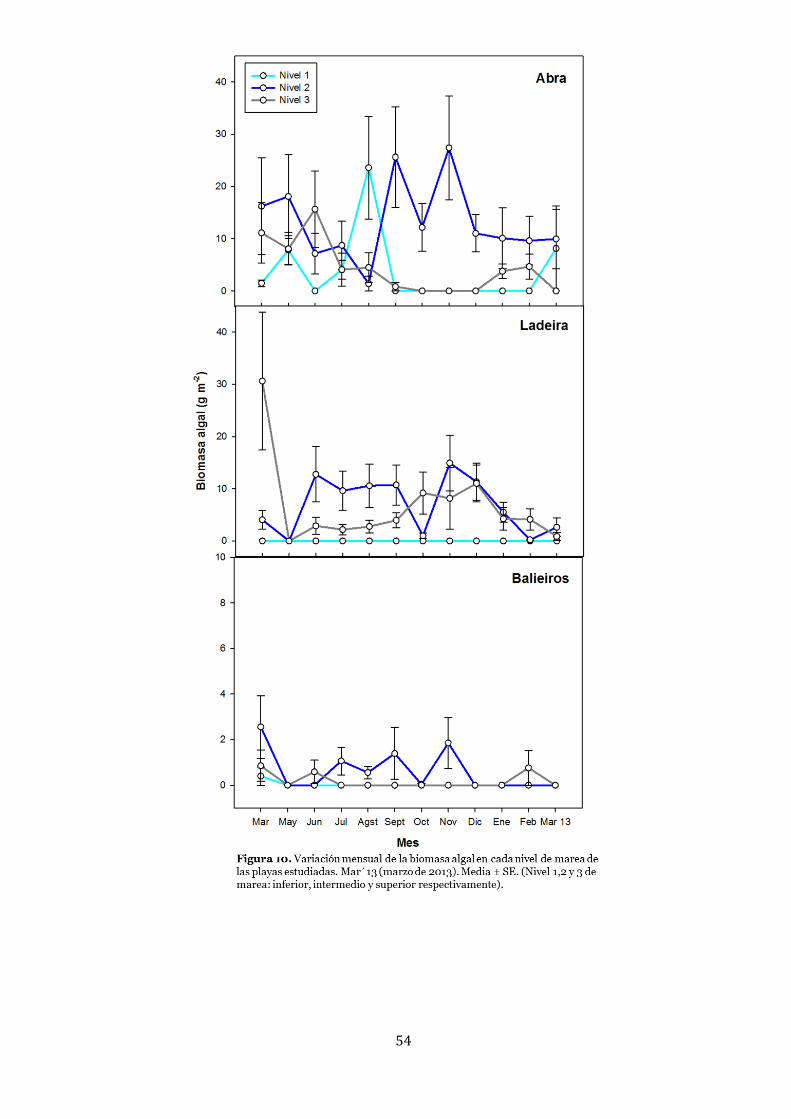
54
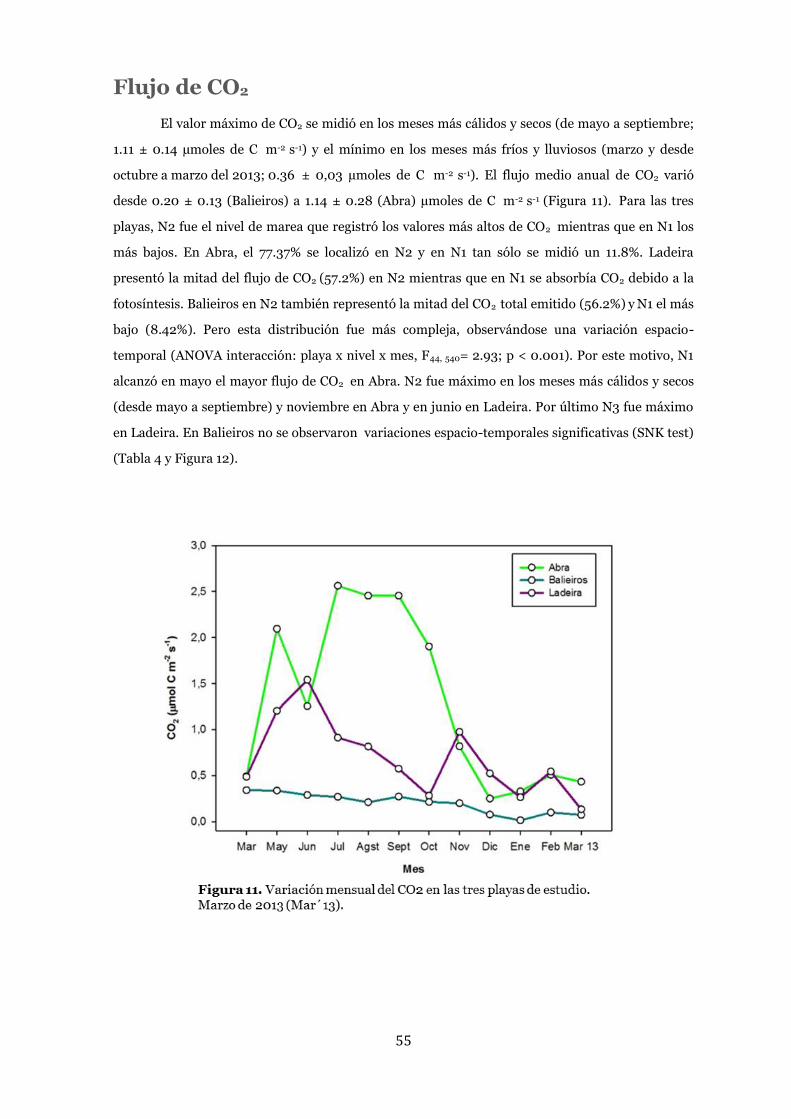
55
Flujo de CO2
El valor máximo de CO2 se midió en los meses más cálidos y secos (de mayo a septiembre;
1.11 ± 0.14 µmoles de C m-2 s-1) y el mínimo en los meses más fríos y lluviosos (marzo y desde
octubre a marzo del 2013; 0.36 ± 0,03 µmoles de C m-2 s-1). El flujo medio anual de CO2 varió
desde 0.20 ± 0.13 (Balieiros) a 1.14 ± 0.28 (Abra) µmoles de C m-2 s-1 (Figura 11). Para las tres
playas, N2 fue el nivel de marea que registró los valores más altos de CO2 mientras que en N1 los
más bajos. En Abra, el 77.37% se localizó en N2 y en N1 tan sólo se midió un 11.8%. Ladeira
presentó la mitad del flujo de CO2 (57.2%) en N2 mientras que en N1 se absorbía CO2 debido a la
fotosíntesis. Balieiros en N2 también representó la mitad del CO2 total emitido (56.2%) y N1 el más
bajo (8.42%). Pero esta distribución fue más compleja, observándose una variación espacio-
temporal (ANOVA interacción: playa x nivel x mes, F44, 540= 2.93; p < 0.001). Por este motivo, N1
alcanzó en mayo el mayor flujo de CO2 en Abra. N2 fue máximo en los meses más cálidos y secos
(desde mayo a septiembre) y noviembre en Abra y en junio en Ladeira. Por último N3 fue máximo
en Ladeira. En Balieiros no se observaron variaciones espacio-temporales significativas (SNK test)
(Tabla 4 y Figura 12).
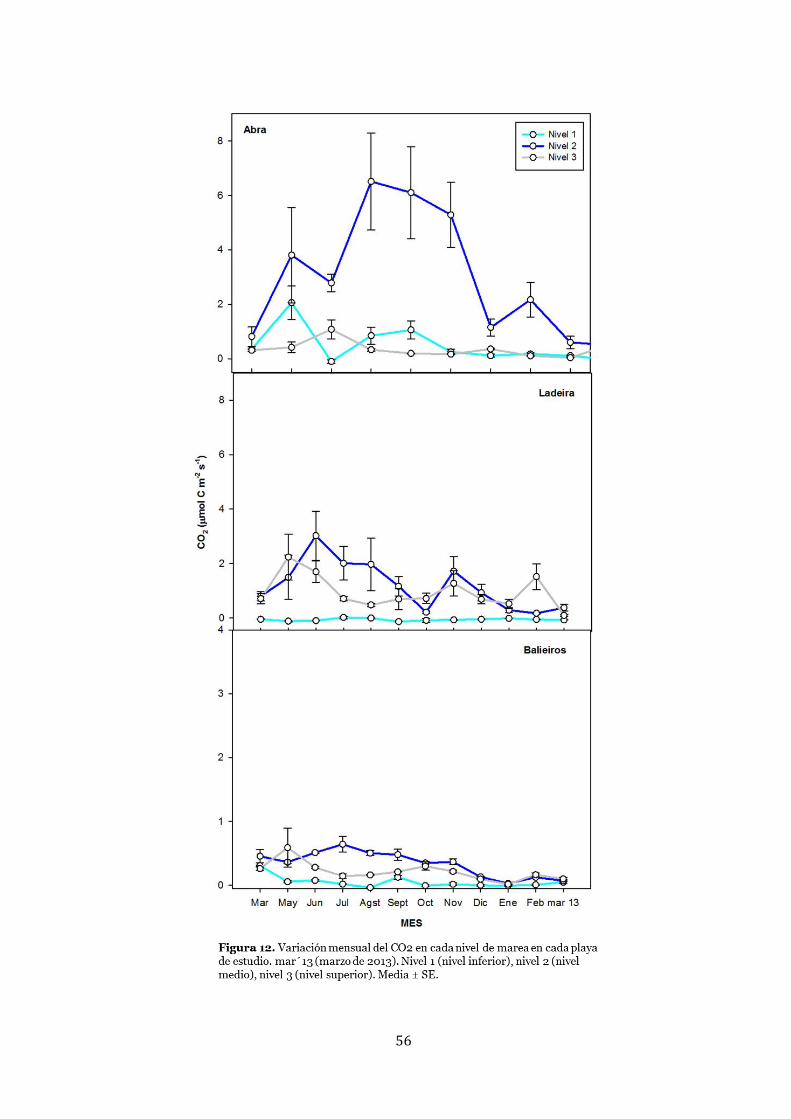
56
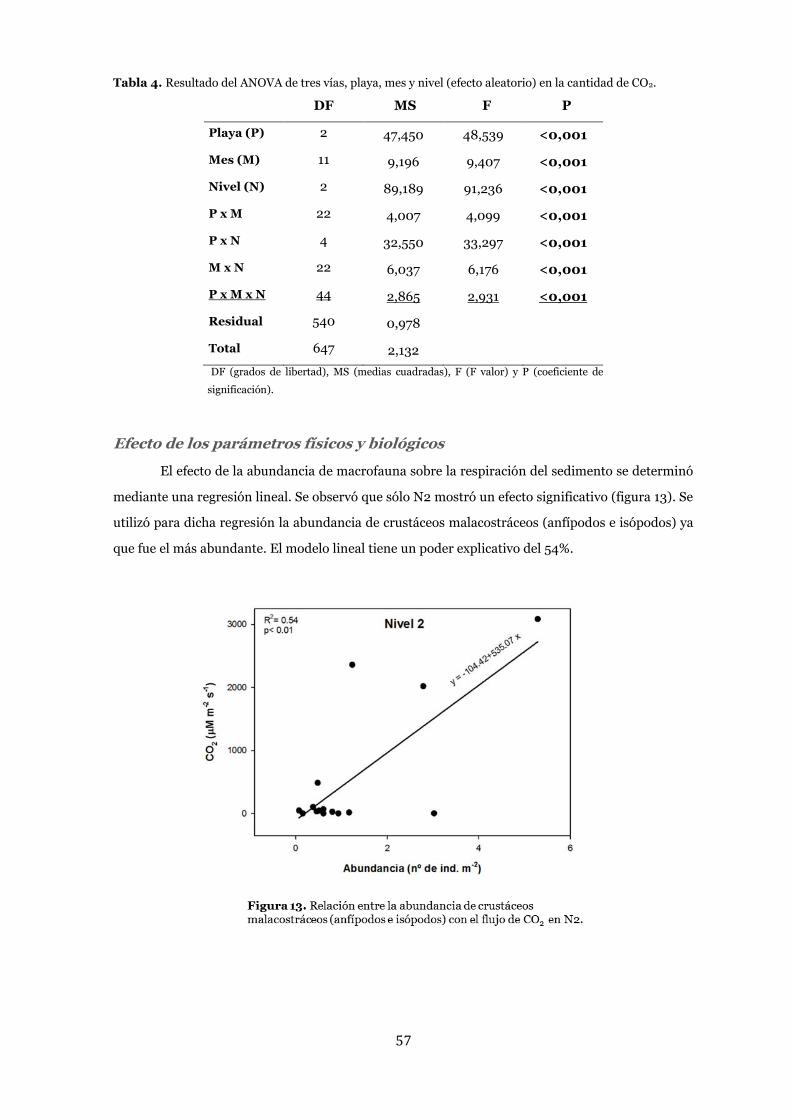
57
Tabla 4. Resultado del ANOVA de tres vías, playa, mes y nivel (efecto aleatorio) en la cantidad de CO2.
DF MS F P
Playa (P) 2 47,450 48,539 <0,001
Mes (M) 11 9,196 9,407 <0,001
Nivel (N) 2 89,189 91,236 <0,001
P x M 22 4,007 4,099 <0,001
P x N 4 32,550 33,297 <0,001
M x N 22 6,037 6,176 <0,001
P x M x N 44 2,865 2,931 <0,001
Residual 540 0,978
Total 647 2,132
DF (grados de libertad), MS (medias cuadradas), F (F valor) y P (coeficiente de
significación).
Efecto de los parámetros físicos y biológicos
El efecto de la abundancia de macrofauna sobre la respiración del sedimento se determinó
mediante una regresión lineal. Se observó que sólo N2 mostró un efecto significativo (figura 13). Se
utilizó para dicha regresión la abundancia de crustáceos malacostráceos (anfípodos e isópodos) ya
que fue el más abundante. El modelo lineal tiene un poder explicativo del 54%.
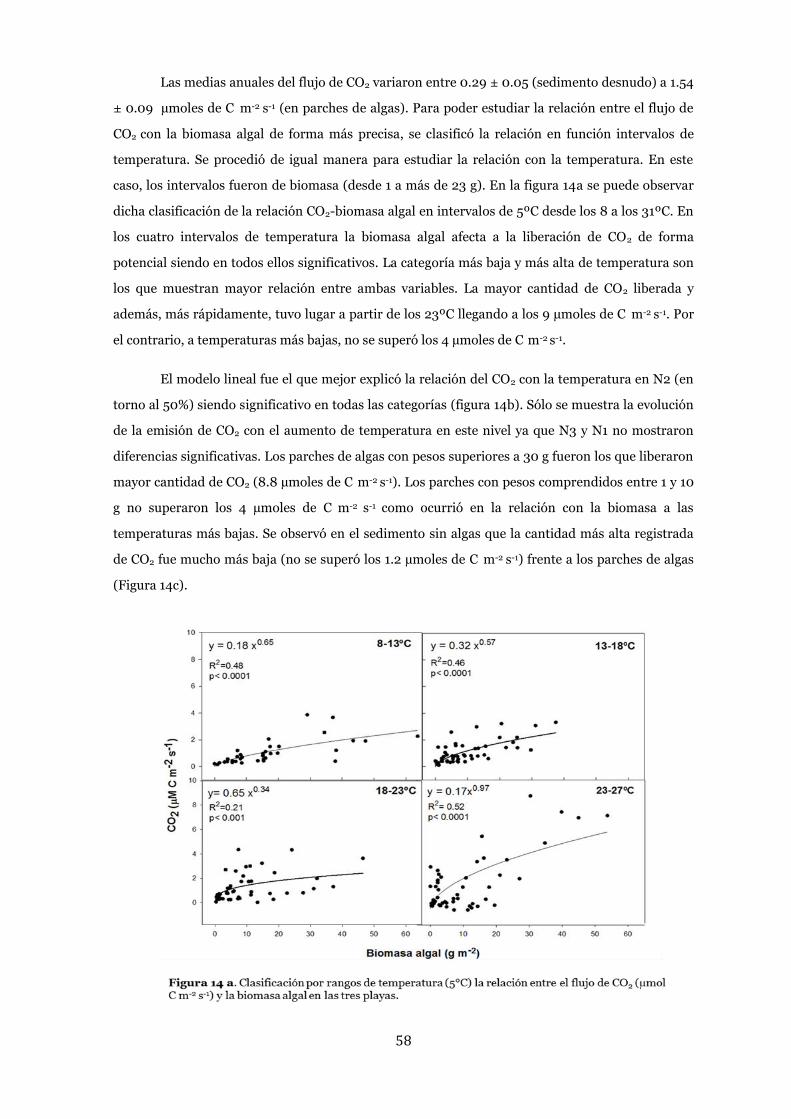
58
Las medias anuales del flujo de CO2 variaron entre 0.29 ± 0.05 (sedimento desnudo) a 1.54
± 0.09 µmoles de C m-2 s-1 (en parches de algas). Para poder estudiar la relación entre el flujo de
CO2 con la biomasa algal de forma más precisa, se clasificó la relación en función intervalos de
temperatura. Se procedió de igual manera para estudiar la relación con la temperatura. En este
caso, los intervalos fueron de biomasa (desde 1 a más de 23 g). En la figura 14a se puede observar
dicha clasificación de la relación CO2-biomasa algal en intervalos de 5ºC desde los 8 a los 31ºC. En
los cuatro intervalos de temperatura la biomasa algal afecta a la liberación de CO2 de forma
potencial siendo en todos ellos significativos. La categoría más baja y más alta de temperatura son
los que muestran mayor relación entre ambas variables. La mayor cantidad de CO2 liberada y
además, más rápidamente, tuvo lugar a partir de los 23ºC llegando a los 9 µmoles de C m-2 s-1. Por
el contrario, a temperaturas más bajas, no se superó los 4 µmoles de C m-2 s-1.
El modelo lineal fue el que mejor explicó la relación del CO2 con la temperatura en N2 (en
torno al 50%) siendo significativo en todas las categorías (figura 14b). Sólo se muestra la evolución
de la emisión de CO2 con el aumento de temperatura en este nivel ya que N3 y N1 no mostraron
diferencias significativas. Los parches de algas con pesos superiores a 30 g fueron los que liberaron
mayor cantidad de CO2 (8.8 µmoles de C m-2 s-1). Los parches con pesos comprendidos entre 1 y 10
g no superaron los 4 µmoles de C m-2 s-1 como ocurrió en la relación con la biomasa a las
temperaturas más bajas. Se observó en el sedimento sin algas que la cantidad más alta registrada
de CO2 fue mucho más baja (no se superó los 1.2 µmoles de C m-2 s-1) frente a los parches de algas
(Figura 14c).
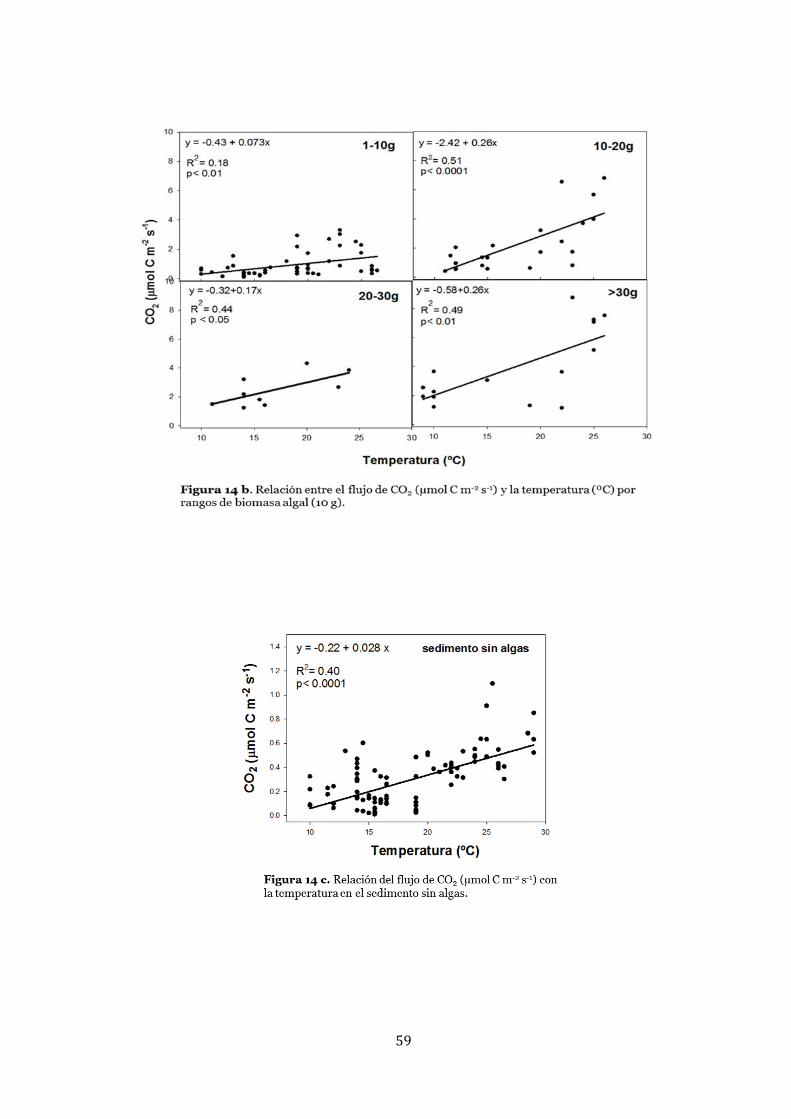
59

60
Nutrientes: TNI y fosfato
Nivel inferior de marea (N1)
Las concentraciones más bajas de nutrientes se encontraron en Balieiros y las más altas en
Abra (Figura 15). Esta playa destacó por registrar la concentración de nitrito y nitrato más alta y
Ladeira la de amonio y fosfatos (Tabla 6).
Las concentraciones anuales medias de TNI (nitrógeno inorgánico total) variaron desde
28.74 ± 2.70 µM en Balieiros y 57.27 ± 5.62 µM en Abra. El mes con mayor cantidad de TNI anual
fue marzo de 2013 pero esto varió espacialmente (ANOVA interacción: playa x mes, F8,74= 8.20;
p<0.001). Así pues, septiembre y marzo de 2013 fueron significativamente superiores en Abra y
junio mostró diferencias significativamente mayores que el resto de meses en Balieiros. Por último,
en Ladeira no se detectaron diferencias significativas entre meses (SNK test) (Tabla 7).
La concentración de fosfato más alta se midió en septiembre en todas las playas. Estas
concentraciones fluctuaron entre los 3.03 ± 0.23 µM de Balieiros y 5.00 ± 1.26 µM de Ladeira.
Mediante el análisis ANOVA (interacción: playa x mes, F8,74= 34.62; p< 0.001) se observó una
ligera variación temporal. Así pues, en Ladeira, también junio mostró diferencias significativas
(SNK test) (Tabla 7).
Niveles superiores de marea (N2 y N3)
En Abra se midieron las concentraciones anuales más altas de nutrientes (Figura 16). El
nitrógeno inorgánico de cada una de las especies fue máximo en N2 y los fosfatos en N3 (Tabla 6).
Las concentraciones de TNI entre meses variaron desde los 12.58 ± 1.03 (diciembre) y
59.50 ± 16.81 µM g-1 sed. (junio). Entre playas varió desde los 9.62 ± 0.89 (Balieiros) y 36.89 ± 7.05
µM g-1 sed. (Abra). Aunque hubo una interacción significativa entre playa x nivel (Tabla 7). Así
pues, TNI difirió significativamente entre playas en N2 y N3 (SNK test). Abra registró en N2 la
concentración máxima y Ladeira en N3.
La concentración de fosfato aumentó desde los 5.26 ± 0.45 (Balieiros) hasta 22.91 ± 3.18
µM g-1 sed. (Abra) (Figura 18). Abra registró la concentración máxima de fosfato aunque se detectó
un efecto temporal significativo (Tabla 7). Así pues, en diciembre se registraron las
concentraciones máximas en Abra y Ladeira (SNK test). Balieiros no mostró diferencias
significativas.
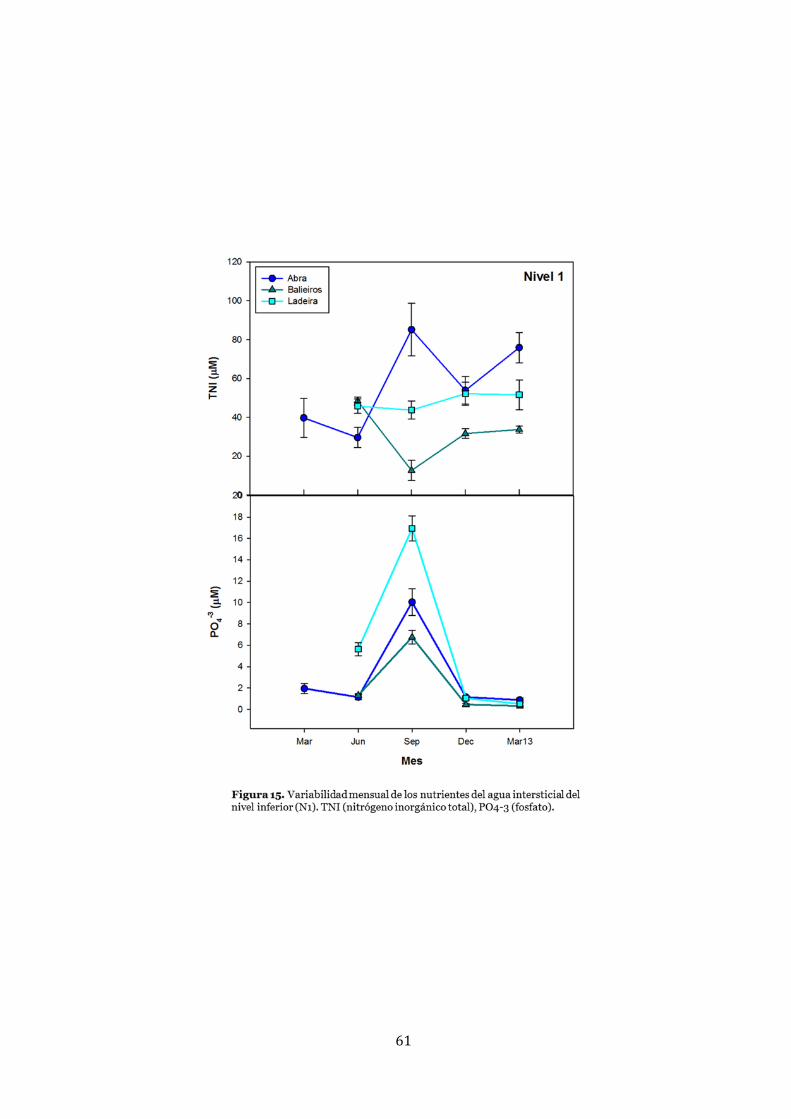
61
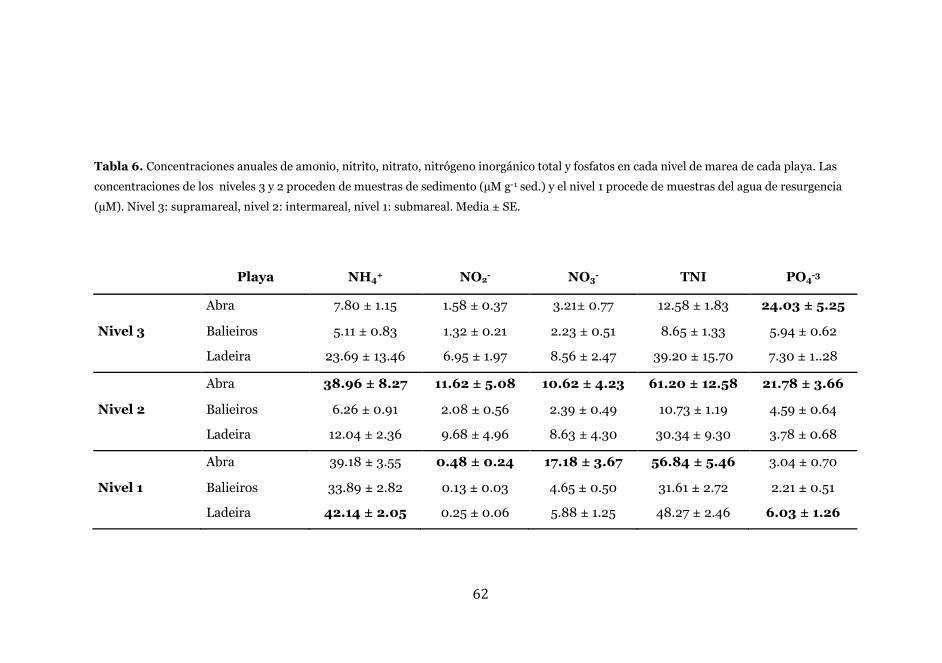
62
Tabla 6. Concentraciones anuales de amonio, nitrito, nitrato, nitrógeno inorgánico total y fosfatos en cada nivel de marea de cada playa. Las
concentraciones de los niveles 3 y 2 proceden de muestras de sedimento (µM g-1 sed.) y el nivel 1 procede de muestras del agua de resurgencia
(µM). Nivel 3: supramareal, nivel 2: intermareal, nivel 1: submareal. Media ± SE.
Playa NH4+ NO2
- NO3- TNI PO4
-3
Abra 7.80 ± 1.15 1.58 ± 0.37 3.21± 0.77 12.58 ± 1.83 24.03 ± 5.25
Nivel 3 Balieiros 5.11 ± 0.83 1.32 ± 0.21 2.23 ± 0.51 8.65 ± 1.33 5.94 ± 0.62
Ladeira 23.69 ± 13.46 6.95 ± 1.97 8.56 ± 2.47 39.20 ± 15.70 7.30 ± 1..28
Abra 38.96 ± 8.27 11.62 ± 5.08 10.62 ± 4.23 61.20 ± 12.58 21.78 ± 3.66
Nivel 2 Balieiros 6.26 ± 0.91 2.08 ± 0.56 2.39 ± 0.49 10.73 ± 1.19 4.59 ± 0.64
Ladeira 12.04 ± 2.36 9.68 ± 4.96 8.63 ± 4.30 30.34 ± 9.30 3.78 ± 0.68
Abra 39.18 ± 3.55 0.48 ± 0.24 17.18 ± 3.67 56.84 ± 5.46 3.04 ± 0.70
Nivel 1 Balieiros 33.89 ± 2.82 0.13 ± 0.03 4.65 ± 0.50 31.61 ± 2.72 2.21 ± 0.51
Ladeira 42.14 ± 2.05 0.25 ± 0.06 5.88 ± 1.25 48.27 ± 2.46 6.03 ± 1.26
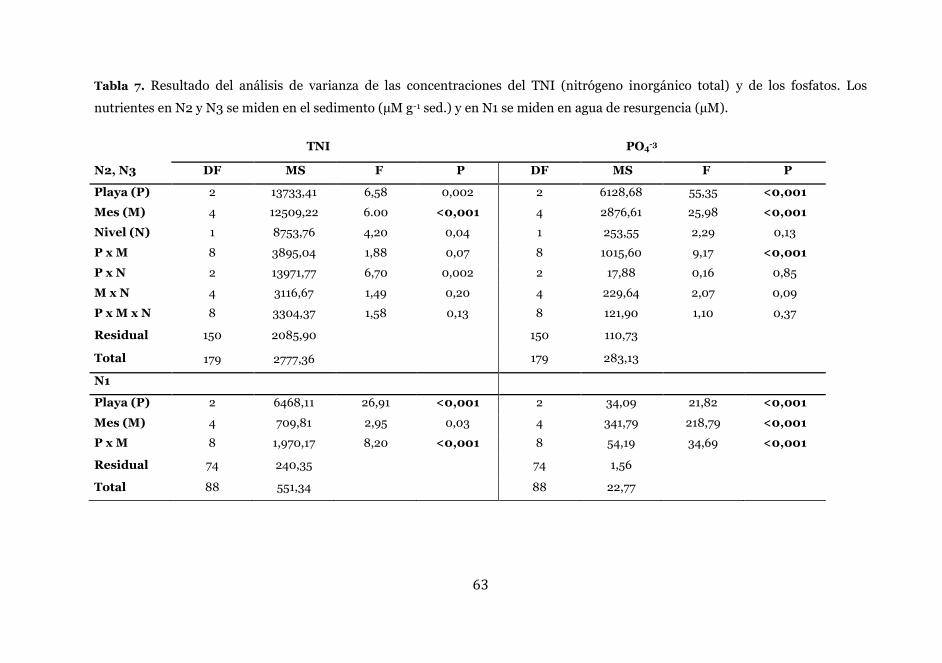
63
Tabla 7. Resultado del análisis de varianza de las concentraciones del TNI (nitrógeno inorgánico total) y de los fosfatos. Los
nutrientes en N2 y N3 se miden en el sedimento (µM g-1 sed.) y en N1 se miden en agua de resurgencia (µM).
TNI
PO4-3
N2, N3 DF MS F P DF MS F P
Playa (P) 2 13733,41 6,58 0,002 2 6128,68 55,35 <0,001
Mes (M) 4 12509,22 6.00 <0,001 4 2876,61 25,98 <0,001
Nivel (N) 1 8753,76 4,20 0,04 1 253,55 2,29 0,13
P x M 8 3895,04 1,88 0,07 8 1015,60 9,17 <0,001
P x N 2 13971,77 6,70 0,002 2 17,88 0,16 0,85
M x N 4 3116,67 1,49 0,20 4 229,64 2,07 0,09
P x M x N 8 3304,37 1,58 0,13 8 121,90 1,10 0,37
Residual 150 2085,90
150 110,73
Total 179 2777,36 179 283,13
N1
Playa (P) 2 6468,11 26,91 <0,001 2 34,09 21,82 <0,001
Mes (M) 4 709,81 2,95 0,03 4 341,79 218,79 <0,001
P x M 8 1,970,17 8,20 <0,001 8 54,19 34,69 <0,001
Residual 74 240,35
74 1,56
Total 88 551,34 88 22,77
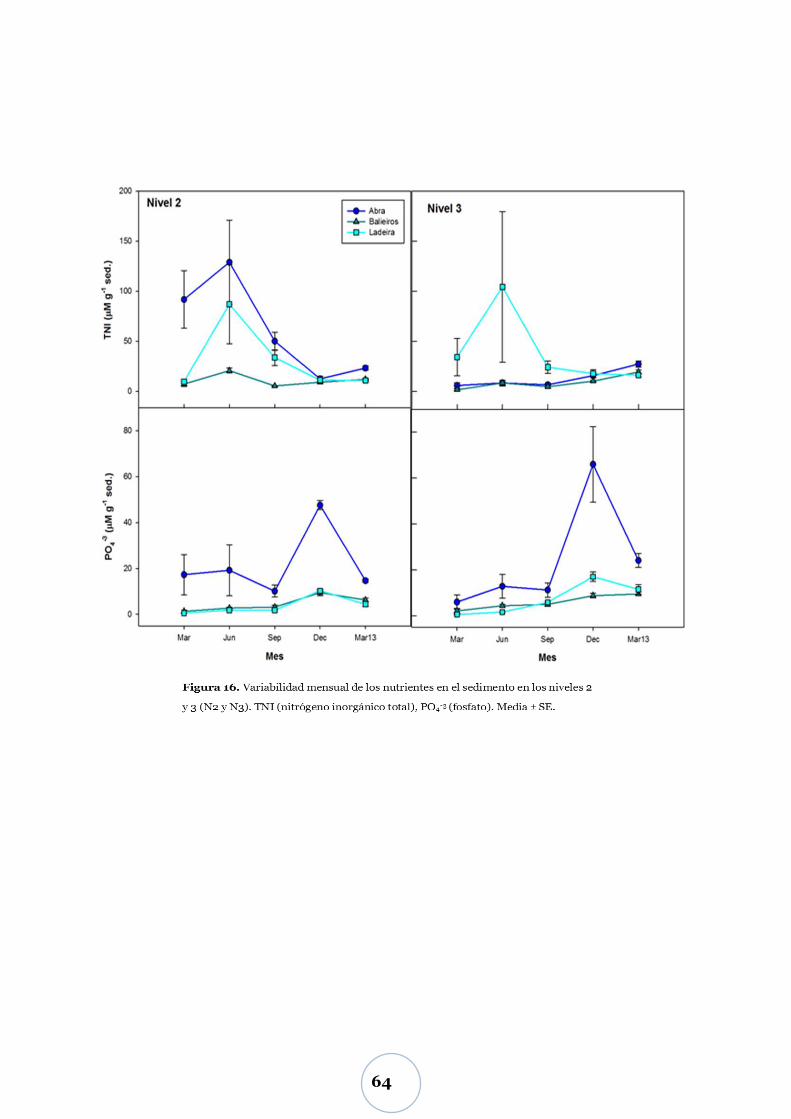
64
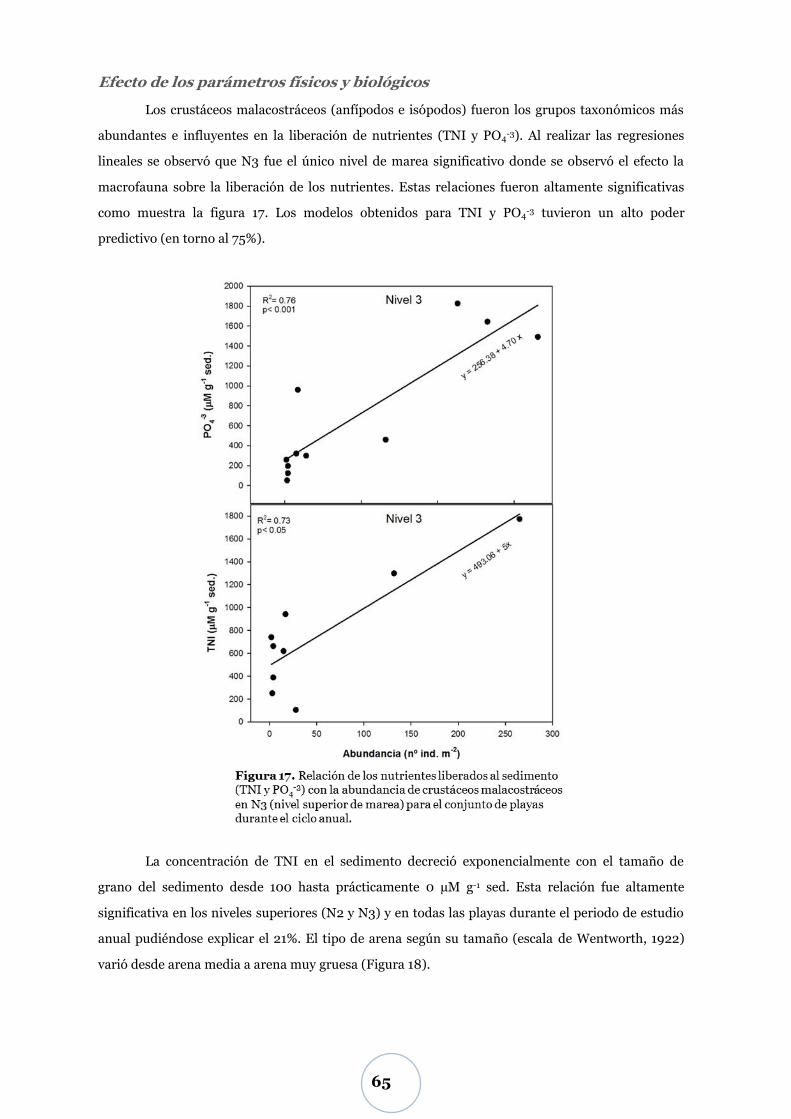
65
Efecto de los parámetros físicos y biológicos
Los crustáceos malacostráceos (anfípodos e isópodos) fueron los grupos taxonómicos más
abundantes e influyentes en la liberación de nutrientes (TNI y PO4-3). Al realizar las regresiones
lineales se observó que N3 fue el único nivel de marea significativo donde se observó el efecto la
macrofauna sobre la liberación de los nutrientes. Estas relaciones fueron altamente significativas
como muestra la figura 17. Los modelos obtenidos para TNI y PO4-3 tuvieron un alto poder
predictivo (en torno al 75%).
La concentración de TNI en el sedimento decreció exponencialmente con el tamaño de
grano del sedimento desde 100 hasta prácticamente 0 µM g-1 sed. Esta relación fue altamente
significativa en los niveles superiores (N2 y N3) y en todas las playas durante el periodo de estudio
anual pudiéndose explicar el 21%. El tipo de arena según su tamaño (escala de Wentworth, 1922)
varió desde arena media a arena muy gruesa (Figura 18).
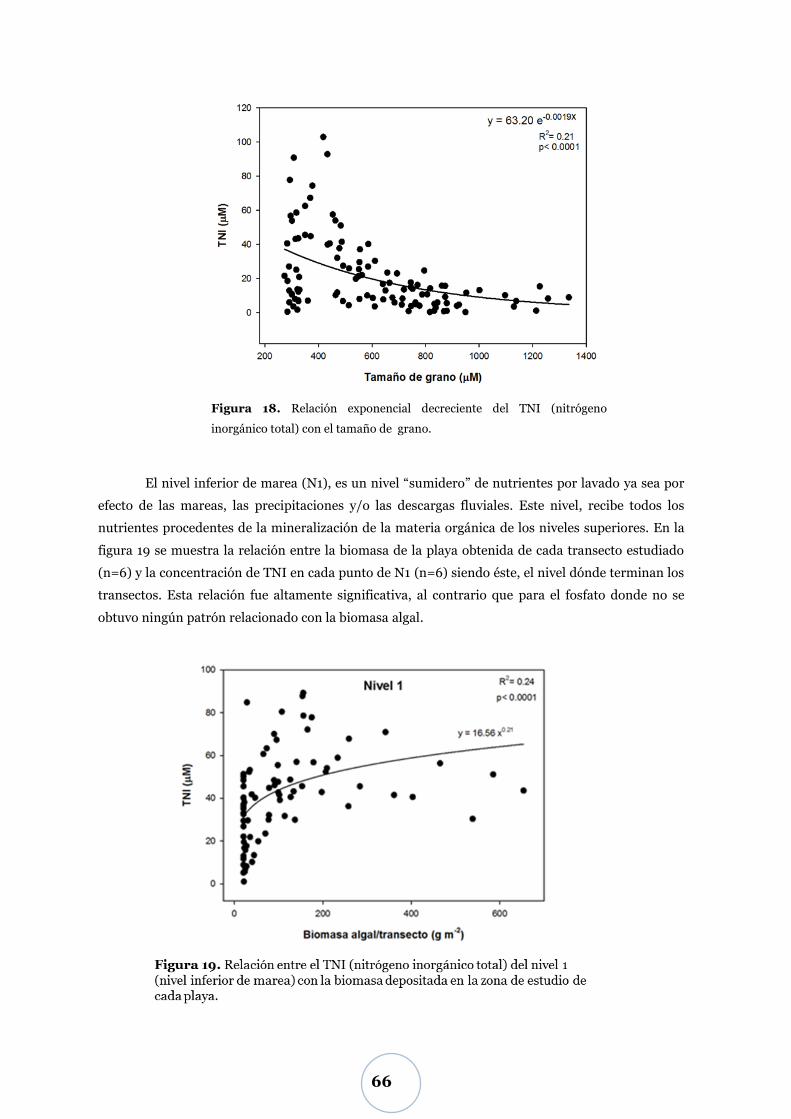
66
Figura 18. Relación exponencial decreciente del TNI (nitrógeno
inorgánico total) con el tamaño de grano.
El nivel inferior de marea (N1), es un nivel ―sumidero‖ de nutrientes por lavado ya sea por
efecto de las mareas, las precipitaciones y/o las descargas fluviales. Este nivel, recibe todos los
nutrientes procedentes de la mineralización de la materia orgánica de los niveles superiores. En la
figura 19 se muestra la relación entre la biomasa de la playa obtenida de cada transecto estudiado
(n=6) y la concentración de TNI en cada punto de N1 (n=6) siendo éste, el nivel dónde terminan los
transectos. Esta relación fue altamente significativa, al contrario que para el fosfato donde no se
obtuvo ningún patrón relacionado con la biomasa algal.
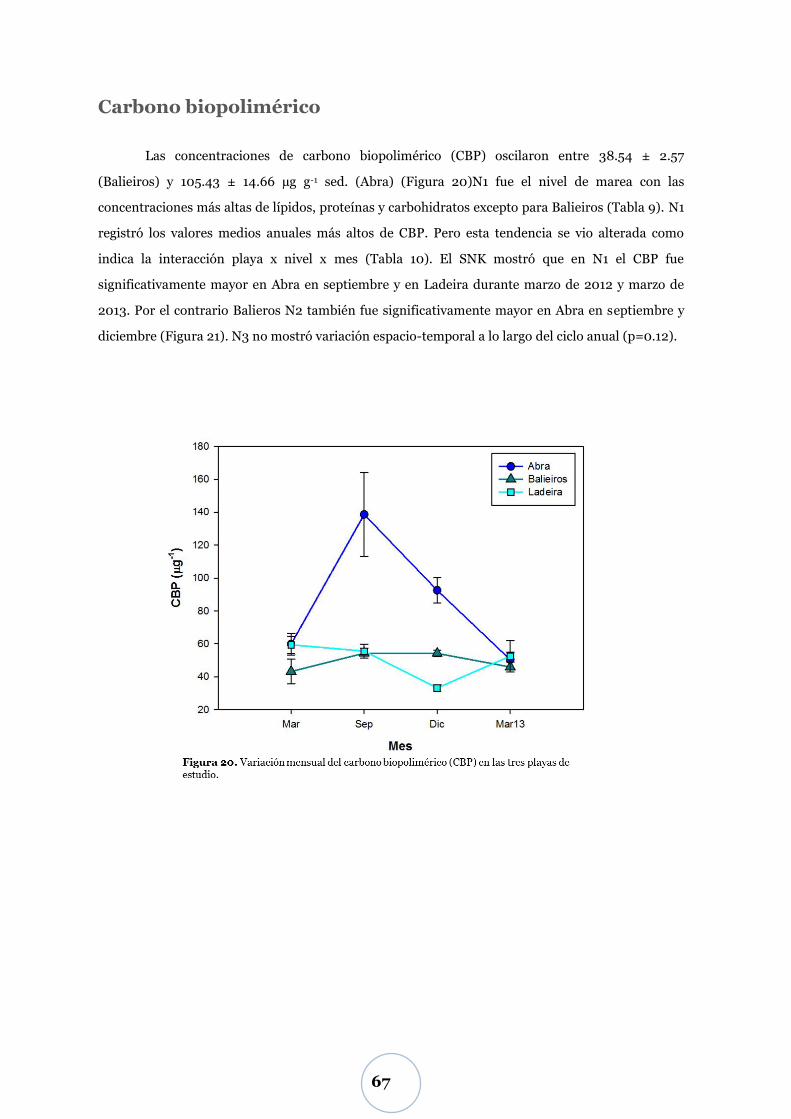
67
Carbono biopolimérico
Las concentraciones de carbono biopolimérico (CBP) oscilaron entre 38.54 ± 2.57
(Balieiros) y 105.43 ± 14.66 µg g-1 sed. (Abra) (Figura 20)N1 fue el nivel de marea con las
concentraciones más altas de lípidos, proteínas y carbohidratos excepto para Balieiros (Tabla 9). N1
registró los valores medios anuales más altos de CBP. Pero esta tendencia se vio alterada como
indica la interacción playa x nivel x mes (Tabla 10). El SNK mostró que en N1 el CBP fue
significativamente mayor en Abra en septiembre y en Ladeira durante marzo de 2012 y marzo de
2013. Por el contrario Balieros N2 también fue significativamente mayor en Abra en septiembre y
diciembre (Figura 21). N3 no mostró variación espacio-temporal a lo largo del ciclo anual (p=0.12).
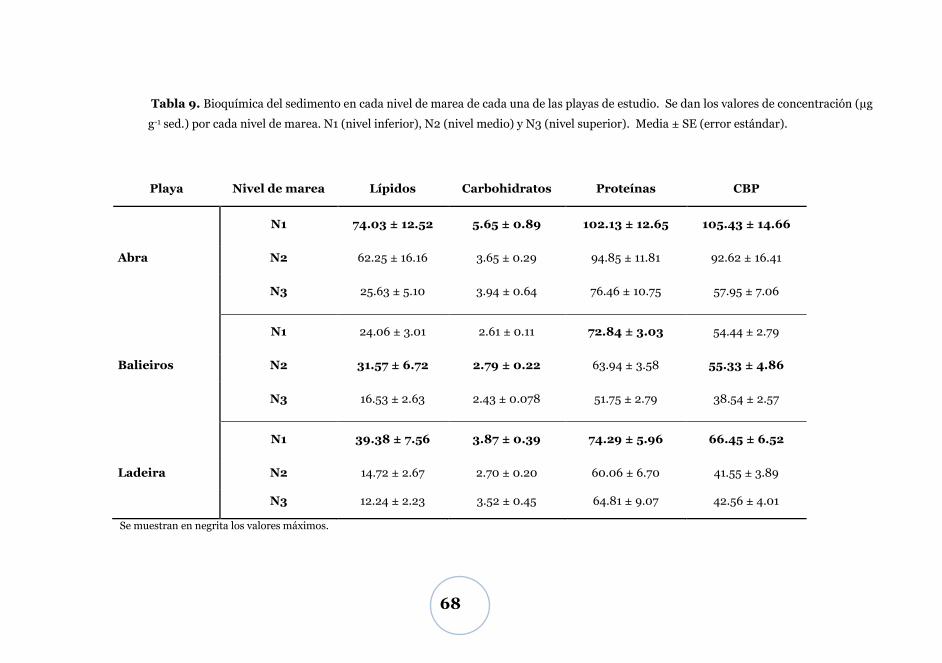
68
Tabla 9. Bioquímica del sedimento en cada nivel de marea de cada una de las playas de estudio. Se dan los valores de concentración (µg
g-1 sed.) por cada nivel de marea. N1 (nivel inferior), N2 (nivel medio) y N3 (nivel superior). Media ± SE (error estándar).
Playa Nivel de marea Lípidos Carbohidratos Proteínas CBP
N1 74.03 ± 12.52 5.65 ± 0.89 102.13 ± 12.65 105.43 ± 14.66
Abra N2 62.25 ± 16.16 3.65 ± 0.29 94.85 ± 11.81 92.62 ± 16.41
N3 25.63 ± 5.10 3.94 ± 0.64 76.46 ± 10.75 57.95 ± 7.06
N1 24.06 ± 3.01 2.61 ± 0.11 72.84 ± 3.03 54.44 ± 2.79
Balieiros N2 31.57 ± 6.72 2.79 ± 0.22 63.94 ± 3.58 55.33 ± 4.86
N3 16.53 ± 2.63 2.43 ± 0.078 51.75 ± 2.79 38.54 ± 2.57
N1 39.38 ± 7.56 3.87 ± 0.39 74.29 ± 5.96 66.45 ± 6.52
Ladeira N2 14.72 ± 2.67 2.70 ± 0.20 60.06 ± 6.70 41.55 ± 3.89
N3 12.24 ± 2.23 3.52 ± 0.45 64.81 ± 9.07 42.56 ± 4.01
Se muestran en negrita los valores máximos.
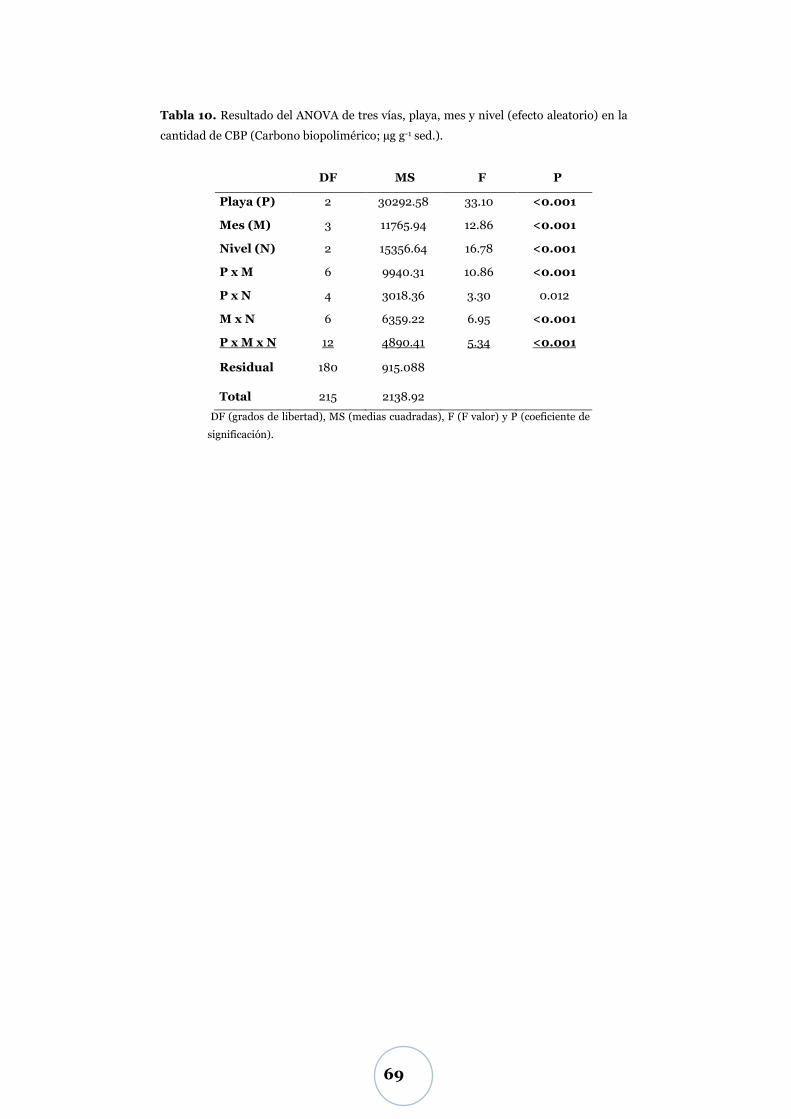
69
Tabla 10. Resultado del ANOVA de tres vías, playa, mes y nivel (efecto aleatorio) en la
cantidad de CBP (Carbono biopolimérico; µg g-1 sed.).
DF MS F P
Playa (P) 2 30292.58 33.10 <0.001
Mes (M) 3 11765.94 12.86 <0.001
Nivel (N) 2 15356.64 16.78 <0.001
P x M 6 9940.31 10.86 <0.001
P x N 4 3018.36 3.30 0.012
M x N 6 6359.22 6.95 <0.001
P x M x N 12 4890.41 5.34 <0.001
Residual 180 915.088
Total 215 2138.92
DF (grados de libertad), MS (medias cuadradas), F (F valor) y P (coeficiente de
significación).
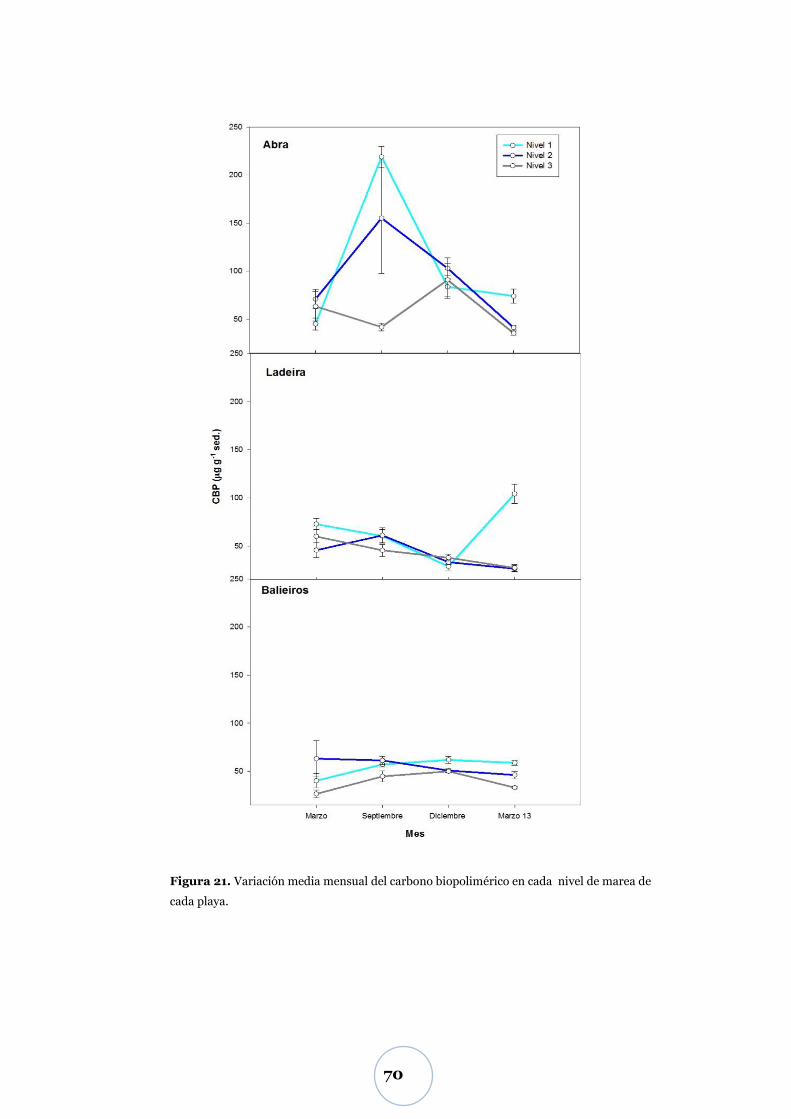
70
Figura 21. Variación media mensual del carbono biopolimérico en cada nivel de marea de
cada playa.
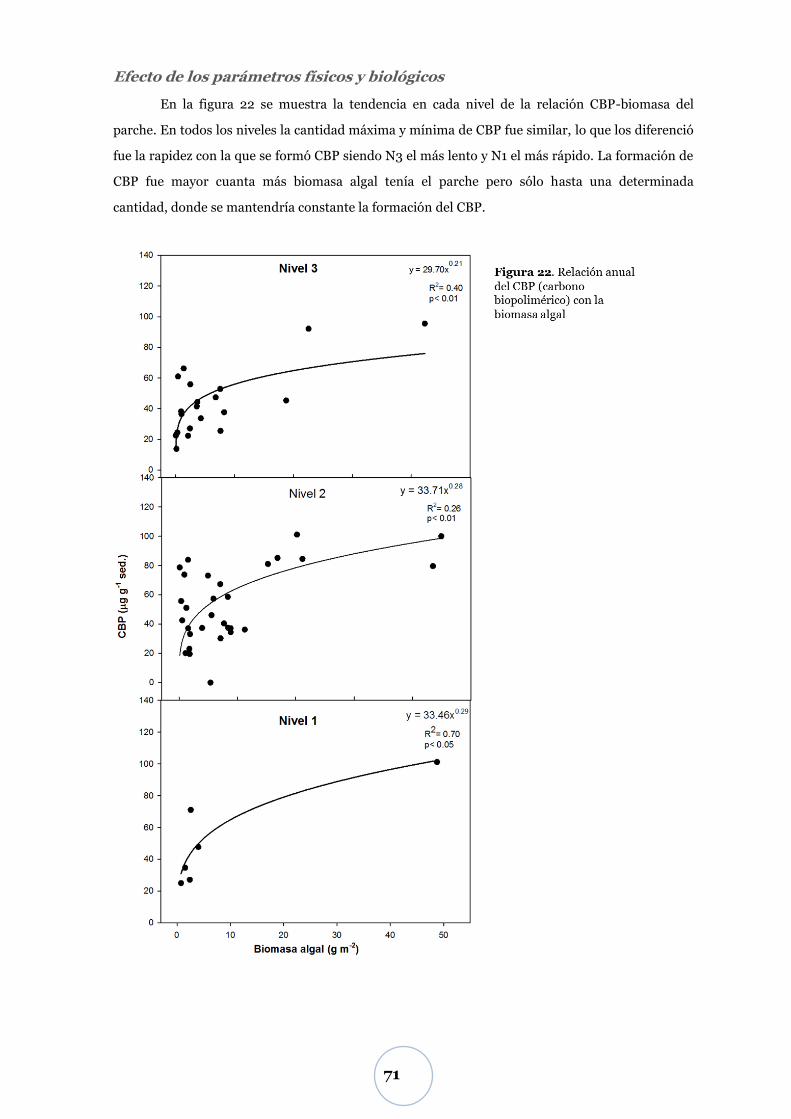
71
Efecto de los parámetros físicos y biológicos
En la figura 22 se muestra la tendencia en cada nivel de la relación CBP-biomasa del
parche. En todos los niveles la cantidad máxima y mínima de CBP fue similar, lo que los diferenció
fue la rapidez con la que se formó CBP siendo N3 el más lento y N1 el más rápido. La formación de
CBP fue mayor cuanta más biomasa algal tenía el parche pero sólo hasta una determinada
cantidad, donde se mantendría constante la formación del CBP.
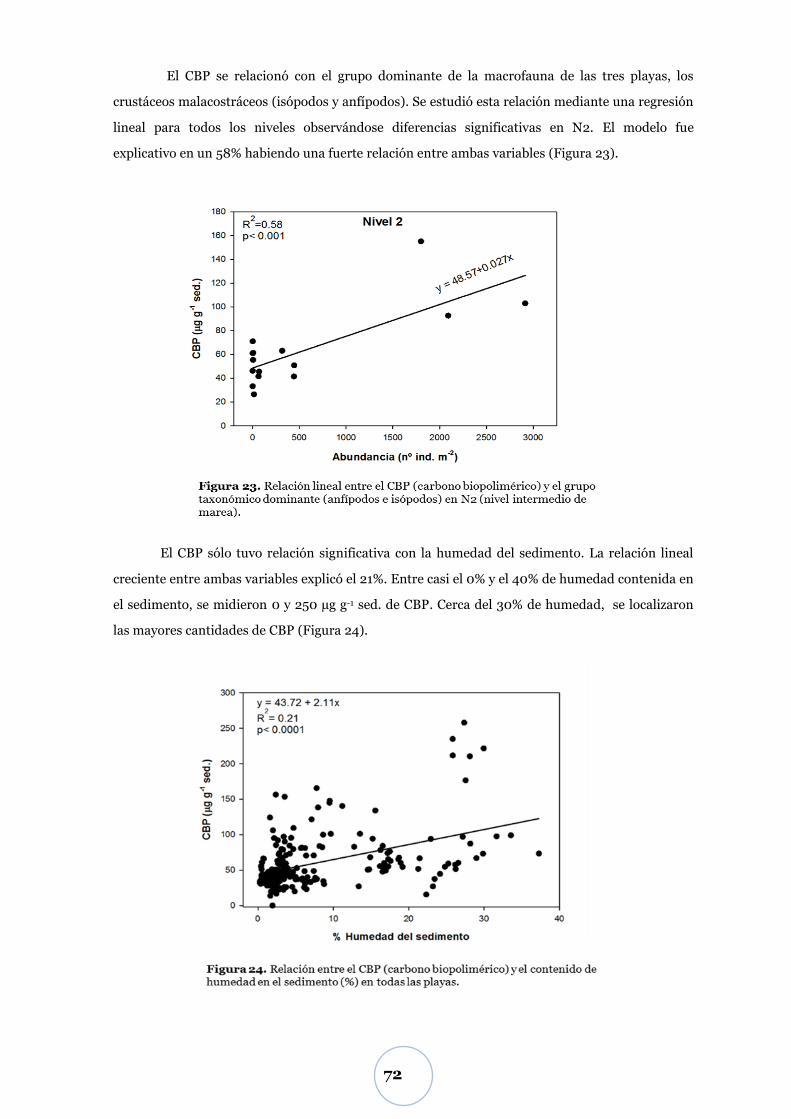
72
El CBP se relacionó con el grupo dominante de la macrofauna de las tres playas, los
crustáceos malacostráceos (isópodos y anfípodos). Se estudió esta relación mediante una regresión
lineal para todos los niveles observándose diferencias significativas en N2. El modelo fue
explicativo en un 58% habiendo una fuerte relación entre ambas variables (Figura 23).
El CBP sólo tuvo relación significativa con la humedad del sedimento. La relación lineal
creciente entre ambas variables explicó el 21%. Entre casi el 0% y el 40% de humedad contenida en
el sedimento, se midieron 0 y 250 µg g-1 sed. de CBP. Cerca del 30% de humedad, se localizaron
las mayores cantidades de CBP (Figura 24).

73
DISCUSIÓN
Efecto de la biomasa algal sobre la macrofauna
La dinámica observada en los varamientos macroalgales durante este estudio concuerda
con estudios anteriores, en los que muestran que existe una estacionalidad en dichos depósitos.
Durante el ciclo anual, se detectó que las biomasas algales más altas correspondieron a meses de
otoño y primavera (noviembre y marzo). La climatología de la costa gallega durante estos meses
se caracteriza por la entrada de borrascas acompañadas de fuerte oleaje. Estos factores físicos son
la principal causa de los desprendimientos de las macroalgas de los fondos rocosos (Duarte, 1974;
Tegner & Dayton, 1987; Dayton et al., 1992; Rodríguez, 1999; Hobday, 2000; Pennings et al.,
2000). Por otro lado, la senescencia natural, al igual que el fuerte oleaje, tiene un patrón
estacional en el suministro de detritus a las playas (Zobell, 1971). El ciclo de vida de la mayoría de
las macroalgas de la costa gallega finaliza en agosto y septiembre, acumulándose grandes
cantidades de algas a lo largo y ancho de las playas. Por eso, en estos meses se registraron en las
playas cantidades importantes de wrack aunque menores que en noviembre y marzo. Estos
resultados son apoyados por estudios realizados por McKechnie y Fairweather (2003);
observaron en tres regiones del sur de Australia que los mayores inputs se producían en la
estación cálida. Así pues, la combinación de estos factores físicos (meteorología) y biológicos
(ciclos de vida) pueden resultar en una gran variabilidad espacial y temporal en la biomasa algal.
La morfología de la playa también influye en la cantidad de macroalgas que se depositan
en el intermareal. Barreiro et al., 2013, demostró que en las playas con una elevada relación
perímetro : área (L:A) (playas pequeñas o de bolsillo) los aportes de algas son mayores por unidad
de superficie ya que tienen una mayor superficie de contacto con el ecosistema donante (costa
rocosa). Las playas encajadas entre promontorios rocosos, como es Abra, reciben mayores
cantidades de inputs. Este hecho puede explicarse por una reducción en el flujo de agua que
ocurre alrededor de los promontorios de la playa. La baja energía de las olas en el frente de la playa
provoca una reducción en el flujo, lo que crea condiciones más favorables para la deposición del
wrack (Komar, 1998; Witman et al., 2004). Por este motivo, las diferencias en la biomasa algal
registrada en Abra fueron significativamente superiores a Ladeira y Balieiros. Estas dos playas
tienen grados de exposición al oleaje superior a Abra. Así pues, la deposición del wrack puede ser
más dinámico y sensible a otros factores, como fuertes vientos u olas. En Abra, debido a la
morfología de su perfil, la mayor cantidad de macroalgas se acumularon en N2, en cambio, en
Ladeira, debido a la suave pendiente, las macroalgas alcanzaron N3 donde se registraron las
máximas biomasas de toda la playa. En Balieiros no se observó diferencias entre los niveles siendo
muy escasa la cantidad de wrack que llega a la playa durante todo el año; estos resultados
coincidieron con estudios anteriores en la misma playa (Barreiro et al., 2011).
La abundancia y las tasas demográficas de los consumidores están fuertemente
influenciadas por la variabilidad espacial y temporal de la acumulación del wrack (Dugan et al.,
2003; Olabarria et al., 2007). Por este motivo las mayores abundancias estuvieron en consonancia
con las de la biomasa algal detectándose en verano y en marzo los máximos. Así pues, fue en junio
en Abra donde se contabilizó el mayor número de individuos y en marzo en Ladeira. La

74
macrofauna puede influir en el procesamiento de los aportes de algas, lo cual ha sido probado en
latitudes templadas (Dugan et al., 2003; Lastra et al., Urban - Malinga & Burska, 2009). Los
estudios de varios autores apoyan los resultados de este estudio al considerar que los anfípodos
son los principales colonizadores de los depósitos de algas varadas en playas de todo el mundo y
por lo tanto los principales responsables del reciclaje de la materia orgánica (Griffiths & Stenton-
Dozey, 1981; Inglis, 1989; Colombini et al., 2000).
Flujo de CO2 y nutrientes
La descomposición es un proceso ecosistémico, resultado de la actividad de la comunidad
saprófita, pero también es un proceso biogeoquímico complejo que integra actividades enzimáticas
y reacciones abióticas (Álvarez, 2005). El 90% de la descomposición es ejecutada por la comunidad
bacteriana liberándose durante el proceso, nutrientes inorgánicos (NO3- y PO4
-3) y CO2 (Álvarez,
2005). Debido a esto, la cantidad de nitrógeno total mineralizado (TNI) estuvo relacionada con la
respiración microbiana del suelo, con la biomasa de ésta y por lo tanto con la liberación de CO2. Así
pues, cualquier variación en el metabolismo bacteriano provocado por cualquier factor ambiental
es fácilmente detectable.
Efectos de los parámetros físicos
La temperatura y la humedad son los factores que más varían a lo largo de las estaciones y
por lo tanto, son los que mejor explican las variaciones estacionales de los flujos de CO2 y las
concentraciones de nutrientes observadas en las playas (Nicolardot et al., 1994; Xu & Qi, 2001; Rey
et al., 2002, Yuste et al., 2003; Davidson et al., 2006; Balogh et al., 2011,). Los valores más altos de
ambos se registraron en los meses más cálidos y secos (de junio a septiembre) que en los meses
más fríos y lluviosos (de octubre a marzo). Esta tendencia estacional en la respiración del suelo fue
observada en estudios previos donde la diferencia entre verano e invierno fue 4 veces superior en
la estación estival (Epron et al., 1999; Subke, 2013). La liberación de nutrientes, concretamente el
TNI, fue apoyada por las investigaciones realizadas por Avery et al., (2008), donde también se
observó que en verano la concentración de TNI era cuatro veces superior en verano. Avery afirmó
que esta tendencia estacional sugiere una mayor producción primaria y/o remineralización. Las
concentraciones máximas del fósfato en sedimentos registradas durante el período más cálido,
pueden explicarse debido al aumento de las tasas de mineralización de la materia orgánica a altas
temperaturas (Chapelle, 1995; Asmus et al., 2000; Serpa et al., 2007).
El efecto de la temperatura sobre la tasa de descomposición, reside en la velocidad de las
reacciones enzimáticas de los microorganismos saprófitos. Generalmente, la tasa de
mineralización aumenta de 2 a 3 veces por cada 10ºC que se eleve la temperatura en el intervalo de
10 a 40ºC, lo que muestra que el proceso es muy sensible a las variaciones de temperatura
(Miranda et al., 2014). Según la teoría cinética de Arrhenius (1889), la energía cinética depende
únicamente de la temperatura absoluta. De esta manera la velocidad de una reacción enzimática se

75
incrementa al aumentar la temperatura dentro de un determinado rango, alcanzando un valor
máximo a la denominada temperatura óptima. Dentro de este rango se produce una rápida
mineralización del nitrógeno orgánico en el sedimento (Álvarez, 2005). A valores superiores, la
actividad disminuye debido a que el enzima, como cualquier otra proteína, sufre procesos de
desnaturalización y, por lo tanto, de inactivación. Así pues, tal como afirma Hanisak (1993) en uno
de sus estudios, la temperatura es importante para determinar la velocidad de descomposición de
las algas y la liberación de nitrógeno. Por eso, las temperaturas más cálidas fueron las más
favorables para la descomposición. Por el contrario, temperaturas por debajo del óptimo, las
velocidades de reacción serán más lentas y se producirá menor cantidad de CO2 y de nutrientes
inorgánicos. Esto se observó a partir del mes de octubre, donde las cantidades de CO2 y nutrientes
fueron disminuyendo hasta alcanzar los mínimos en diciembre.
Aunque la temperatura del suelo suele representar una gran fracción de la variación
estacional de los flujos de CO2 y liberación de nutrientes, se sabe que otros factores como el
contenido de agua en el sedimento, también interviene en la descomposición de la materia
orgánica (Linn & Doran 1984; Trumbore et al., 1995). Ambos factores covarían en el campo siendo
la temperatura modulada por la humedad del suelo (Hol et al., 1990; O’Connell, 1990; Lloyd &
Taylor, 1994; Davidson et al., 1998; Janssens et al., 1998; Kelliher et al., 1999; Xu & Qi, 2001;
Almagro et al., 2009). Los modelos obtenidos para el flujo de CO2 como para los nutrientes
mostraron su dependencia a la humedad (junto con otros factores). El efecto de la humedad se
basa en la interferencia de las moléculas de agua en el movimiento del aire reduciendo el
suministro de oxígeno a través del sedimento hasta los centros de actividad microbiana (Paul &
Clark 1989; Castillo et al., 2010). Pero un alto contenido de agua puede impedir la difusión de CO2
en el suelo (Linn & Doran 1984; Doran et al., 1990; Skopp et al., 1990). Por el contrario, un bajo
contenido de agua en el suelo (humedad del suelo en su punto de marchitamiento) puede inhibir la
actividad microbiana por falta de moléculas de agua que difundan el oxígeno hasta los
microorganismos aerobios (Davidson et al., 1998; Epron et al., 1999; Kelliher et al., 1999, Xu & Qi,
2001; Rastogi, 2002; Curiel Yuste et al., 2003, Almagro et al., 2009). Aunque también puede
continuar ocurriendo, lo que sugiere que puede haber acumulación de nitrógeno inorgánico en el
sedimento durante periodos secos; asimismo, el humedecimiento del suelo seco parece estimular
la mineralización y provocar la liberación de nitrógeno disponible (Foth & Ellis, 1997). Esto es lo
que pudo haber pasado en Ladeira en los meses de marzo y diciembre y en marzo de 2013 en
Balieiros en N1. En este nivel se registró el máximo de TNI, posiblemente arrastrado por las
precipitaciones típicas de los meses de invierno desde los niveles superiores. En cada playa se
observaron diferencias significativas entre los tres niveles de marea. En N2 las tasas de
mineralización fueron las más altas, ya que al ser el nivel intermedio el porcentaje de humedad fue
el óptimo. Además, en este nivel se retuvo más humedad que en N3 porque es donde se acumulan
las cantidades más altas de macroalgas y éstas contienen agua en su interior, coincidiendo con las

76
observaciones de Coupland et al., (2007). Los resultados de este estudio prueban el efecto
significativo de la temperatura y la humedad. En estudios realizados por Long (2003) se observó
que las diferencias entre las concentraciones de TNI en el sedimento de las playas arenosas y en el
agua subterránea de N1 fueron de más de un orden de magnitud. Esto sugiere que el reciclaje del
nitrógeno en los sedimentos de las playas, es una fuente significativa de nutrientes puesto que se
producen localmente elevadas concentraciones que podrían desempeñar un papel importante en la
productividad primaria neta a lo largo de la costa. Además, debido a que las precipitaciones son
episódicas, son menos relevantes que la acción de las mareas que diariamente lava el intermareal.
En consecuencia, la alta variabilidad espacial y temporal observada durante el período de estudio,
las concentraciones de los nutrientes, estuvieron en consonancia con los procesos biogeoquímicos
(complejos y dinámicos) que tienen lugar en los entornos intersticiales de las orillas sedimentarias
(Jickells & Rae, 1997).
Efecto de la biomasa algal
La materia orgánica que se mineraliza en las playas puede proceder de las macroalgas que
quedan varadas en ellas, ser importada por las mareas o ser de origen terrestre. En cualquier caso,
su mineralización libera al medio, nutrientes y CO2. Por eso, el papel de las playas en el reciclaje de
la materia orgánica es de suma importancia, porque le devuelve al medio los nutrientes necesarios
para que el ciclo de la materia comience de nuevo (Boudreau et al., 2001, Ehrenhauss & Huettel,
2004; Rocha, 2008). Por esta razón, las playas se caracterizan por ser reactores biogeoquímicos
activos (Anschutz et al., 2009). Los resultados de este estudio muestran que existe relación entre el
wrack y los nutrientes y CO2 ya que se observó en Abra y en Ladeira. En los meses donde hubo
mayor biomasa algal, hubo mayor mineralización. Por el contrario Balieiros no mostró esta
dependencia y aunque las cantidades de nutrientes y de CO2 fuesen bajas, existió mineralización.
En este tipo de playas, con escaso aporte algal, pronunciada pendiente y tamaño de arena gruesa,
se suman los efectos de dichos factores favoreciendo el lavado del sedimento y eliminado de él lo
poco que ha generado durante la mineralización. Los resultados de estudio son comparables al
estudio realizado en las mismas playas que Barreiro et al., (2011). Con todo esto, se muestra que
actividad metabólica de una playa como Balieiros será más baja que otra como Abra. Pero en
playas con aportes de wrack similares no tienen por qué registrar las mismas cantidades de
nutrientes y CO2. Se comparó Abra (playa con alto aporte algal) con otras playas de California o
Sudáfrica (Koop & Lucas 1983; Dugan et al., 2003) que reciben aportes semejantes o superiores
pero con distinta circulación del agua intersticial. Esto resultó en una disminución en la
concentración de nutrientes en comparación con Abra. Los flujos de agua dependen de múltiples
factores como el tamaño de grano, la pendiente y el ciclo de las mareas (Yin & Harrison, 2000;
McLachlan & Brown 2006; Wilson et al., 2008). Así que, al igual que en Balieiros, el lavado de los
nutrientes hizo que playas con grandes aportes de wrack no acumulasen nutrientes. Por otro lado,

77
debido a este lavado, estas playas proporcionan grandes cantidades de nutrientes a las aguas
costeras convirtiéndose en lugares de gran importancia para la producción primaria. En
consecuencia, las concentraciones más altas de nutrientes generalmente se detectan en el
sedimento bajo los parches de macroalagas (Rossi & Underwood, 2002; Corzo et al., 2009). De
hecho, en varias playas, el mayor contenido de nutrientes se detectó cerca de la línea de deriva o
drift line (Swarzenski & Izbicki, 2009; Dugan et al., 2011). Al igual que en estudios de Coupland
(2007), no sorprende que en todas las playas exista mayor actividad metabólica en N2 (drift line).
Las dimensiones de los parches en este nivel permiten, por su capacidad aislante, mantener en su
interior condiciones de temperatura y humedad óptimas para que la actividad metabólica sea
máxima. El oxígeno necesario es suministrado por los intersticios para el metabolismo aeróbico
(Coupland et al., 2007). Por el contrario, N1 por exceso en el contenido de agua y N3 por defecto,
impiden la actividad microbiana (Coupland et al., 2007). Por lo tanto, en este estudio, N3 y N1
registraron las menores actividades metabólicas a lo largo del año exceptuando Ladeira en N3. La
suave pendiente de Ladeira permite que los varamientos macroalgales alcancen y se acumulen en
el los niveles superiores (N2 y N3). Así pues, este estudio pone en valor el papel de las playas
arenosas en el ciclo biogeoquímico y ha demostrado la importancia de las algas en descomposición
como una importante fuente de nitrógeno para las aguas costeras.
Efecto de la macrofauna
Los anfípodos en el nivel 2 de marea, influyeron en las emisiones de CO2 bien por su
contribución por su propia respiración ya que las altas biomasas de macroalgas atraen a gran
número de individuos o por sus actividades facilitando el proceso de descomposición del wrack. La
liberación de los nutrientes estuvo influenciada sólo en N3. En el caso de TNI, tanto los anfípodos
como los isópodos favorecieron su liberación. Para los fosfatos, sólo los anfípodos tuvieron un
papel relevante. En los parches de algas encuentran alimento, lugar de cría y refugio (Coupland et
al., 2007; Gómez, 2015) y pese a la baja contribución directa de la macrofauna (20%) en el proceso
de descomposición, su participación es activa y tiene dos importantes papeles. El primero es
favorecer la respiración microbiana mediante la fragmentación y trituración de residuos vegetales
(Harrison, 1982; Bonkowski et al., 2000; Chaves et al., 2014; Gómez, 2015). Con la fragmentación,
se aumenta la superficie de contacto con el aire y la superficie disponible para la acción
microbiana. Así pues, se ve incrementada la velocidad de degradación y por consiguiente, la
actividad metabólica. El segundo, es que gracias a su actividad trituradora forma orificios dentro
del parche; esto favorece la penetración y la difusión de oxígeno hacia el interior de los acúmulos
de algas. De esta forma el suministro de oxígeno para la comunidad microbiana está garantizado.
El metabolismo de la macrofauna, al igual que el de la microbiota, está influenciado por los
factores climáticos. En invierno, ve reducida su actividad locomotora afectando a su

78
comportamiento alimenticio y un bajo contenido en humedad, le provoca estrés metabólico por
deshidratación (Lastra et al., 2014).
Carbono biopolimérico
El CBP se considera una estimación fiable de la fracción lábil de materia orgánica total
fácilmente asimilable para los organismos e indica la calidad y cantidad del alimento disponible
para ellos (Faviano et al., 1995; Pusceddu et al., 2007). Las concentraciones de CBP estuvieron
fuertemente relacionadas con la biomasa algal y la abundancia de macrofauna. Estos resultados se
vieron apoyados por investigaciones de Pusceddu et al. (2007) donde se afirma que los aportes de
materia orgánica en forma de macroalgas están vinculados con las cantidades de CBP. También la
relación con la abundancia de individuos estuvo respaldada por estudios de Incera et al. (2003).
Debido a esto, las mayores cantidades de CBP se registraron en Abra (playa con mayores aportes
algales) en septiembre (debido a la senescencia se acumulan grandes cantidades de wrack a lo
largo y ancho de Abra) y en N1 y N2 (el sedimento está formado por arena fina). Estas
características, a su vez, determinan la distribución y abundancia de la macrofauna. La
granulometría está en gran parte determinada por la hidrodinámica del ambiente. Así pues, en
ambientes poco hidrodinámicos como es Abra, el sedimento es mucho más fino y tiende a
acumular mayores cantidades de materia orgánica que llega a la playa que en ambientes con mayor
hidrodinamismo (Incera et al., 2003). De forma contraria, Balieiros fue la que registró las
cantidades más bajas de CBP ya que es la playa con mayor exposición al oleaje y por lo tanto la que
tiene mayor pendiente y tamaño de grano. Por eso, el CBP que se puede fomar, es lavado con las
mareas diariamente. Ladeira por sus características físicas (tiene el tamaño de grano más fino de
las tres playas y la pendiente más suave) cabría esperar que acumulase las cantidades más altas de
CBP ya que el lavado sería mínimo pero los bajos aportes macroalgales son determinantes en
términos de alimenticios y Ladeira recibe menos aportes que Abra, por lo que no es la que acumula
más CBP. Esto sugiere que los cambios en la cantidad de CBP podría responder a cambios y/ o
diferencias en la productividad de los sistemas (Fabiano et al., 1995).
CONCLUSIONES
El presente estudio ha permitido demostrar que la cantidad de CO2 y de nutrientes
inorgánicos varía en el espacio y en el tiempo. Las variaciones en el espacio dependen
fundamentalmente de los aportes de algas, su ―edad‖ y su distribución a lo largo del intermareal;
éstos a su vez dependen de la morfología de la playa. Las variaciones temporales son de carácter
estacional. Se diferencian las tasas metabólicas entre los meses cálidos y fríos. Con la llegada de la
subida de las temperaturas, todas las reacciones químicas se ven aceleradas, incrementándose así,
la velocidad de descomposición. La cara inversa del proceso se observó durante los meses más

79
fríos. Se caracteriza por una ralentización a nivel de comunidad involucrado en el proceso. Pero
para conocer cómo afectan a los ciclos biogeoquímicos se deberían realizar estudios teniendo en
cuenta otros factores bióticos, como la actividad microbiana. Hay que tener en cuenta que la
contribución de los factores bióticos puede estar eclipsada por los abióticos debido a la
dependencia de las variaciones los bióticos.

80
BIBLIOGRAFÍA
Álvarez, S. (2005). La descomposición de materia orgánica en humedales: la importancia del
componente microbiano. Revista Ecosistemas, 14(2).
Barreiro F., Gómez M., López J., Lastra M. & la Huz R. (2013) Coupling between macroalgal inputs
and nutrients outcrop in exposed sandy beaches. Hydrobiologia, 700, 73-84.
Boone, R.D., Nadelhoffer, K.J., Canary, J.D., Kaye, J.P., 1998. Roots exert a strong influence on the
temperature sensitivity of soil respiration. Nature 396, 570-572.
Cauwet, G., 1978. Organic chemistry of sea water particulates: concepts and developments.
Oceanol. Acta 1: 99–105.
Chapman aRO, Craigie JS (1977) Seasonal growth in Laminaria longicruris: Relations with
dissolved inorganic nutrients and internal reserves of nitrogen. Mar Biol 40:197-205.
Cividanes, F. J., & Souza, V. P. (2003). Exigências térmicas e tabelas de vida de fertilidade de
Myzus persicae (Sulzer)(Hemiptera: Aphididae) em laboratório. Neotropical Entomology,
32(3), 413-419.
Colombo J C, Silverberg N, Gearing J N (1996) Biogeochemistry of organic matter in the
Laurentian trough, II. Bulk composition of the sediments and relative reactivity of major
components during early diagenesis; Marine Chemistry. 51:295-314.
Coupland, G.T., Duarte, C.M. & Walker, D.I., 2007a. High metabolic rates in beach cast
communities. Ecosystems, 10(8), pp.1341–1350.
Cronin G. & Hay M.E. (1996) Susceptibility to herbivores depends on recent history of both the
plant and the animal. Ecology, 77, 1531-1543
Curiel, Yuste, J., Baldocchi, D.D., Gershenson, A., Goldstein, A., 2007. Microbial soil respiration
and its dependence on carbon inputs, soil temperature and moisture. Global Change Biology
13, 1-18.
Davidson, E.A., Belk, E., Boone, R.D., 1998. Soil water content and temperature as independent or
confounded factors controlling soil respiration in a temperate mixed harwood forest. Global
Change Biology 4, 217-227.
Danovaro, R., M. Fabiano & N. Della Croce, 1993. Labile organic matter and microbial biomasses
in deep-sea sediments (Eastern Mediterranean Sea). Deep-Sea Res. 5: 953–965.

81
Dell’Anno A, Mei M L, Pusceddu A, Danovaro R (2002) Assessing the trophic state and
eutrophication of coastal marine systems. A new approach based on the biochemical
composition of sediment organic matter; Marine Pollution Bulletin. 44:611-622.
Dodds W K, Cole J J (2007) Expanding the concept of trophic state in aquatic ecosystems:It’s not
just the autotrophs; Aquatic Sciences. 69:427-439.
Dubois, M., Gilles, K. A., Hamilton, S. K., & Rebers, P. A. (1956). Colorimetric method for
determination of sugars and related substances. Analytical Chemistry 28, 350–356.
Duong, Huynh Lien Stephanie. 2008. Investigating the ecological implications of wrack removal
on South Australian sandy beaches.
Findlay S, Pace M L, Lints D, Howe K (1992) Bacterial metabolism of organic carbon in tidal
freshwater Hudson estuary; Marine Ecology Progress Series. 89:147-153.
Frankenberger Jr., W.T., Tabatabai, M.A., 1991. L-Glutaminase activity of soils. Soil Biology and
Biochemistry 23, 869-874.
García-Robledo E, Corzo A, García de Lomas J, van Bergeijk S (2008) Biogeochemical effects of
macroalgal decomposition on intertidal microbenthos: a microcosm experiment. Mar Ecol
Prog Ser 356:139 151.
Gómez Rodríguez, Marina (2015). Ecología de los varamientos de macroalgas en playas estuáricas
de la costa de Galicia.
Griffiths, C. L., Stenton-Dozey, J. M. E., & Koop, K. (1983). Kelp wrack and the flow of energy
through a sandy beach ecosystem. In Sandy beaches as ecosystems (pp. 547-556). Springer
Netherlands.
Grundmann, G.L., Renault, P., Rosso, L., Bardin, R., 1995. Differential effects of soil water content
and temperature on nitrification and aeration. Soil Science Society of America Journal 59,
1342-1349.
Guntiñas, M. E. (2009). Influencia de la temperatura y de la humedad en la dinámica de la materia
orgánica de los suelos de Galicia y su relación con el cambio climático. Universidad de
Santiago de Compostela. Santiago de Compostela.
Hanisak M (1993) Nitrogen release from decomposing seaweeds: species andtemperature effects. J
Appl Phycol 5:175-181.
Hobson, L. A., 1967. The seasonal and vertical distribution of suspended particulate matter in an
area of the Northeast Pacific Ocean. Limnol. 12: 642–649.

82
Ince, R., Hyndes, G. A., Lavery, P. S., & Vanderklift, M. A. (2007). Marine macrophytes directly
enhance abundances of sandy beach fauna through provision of food and habitat. Estuarine,
Coastal and Shelf Science, 74(1), 77-86.
Incera, M., Cividanes, S. P., López, J., & Costas, R. (2003). Role of hydrodynamic conditions on
quantity and biochemical composition of sediment organic matter in sandy intertidal
sediments (NW Atlantic coast, Iberian Peninsula). Hydrobiologia, 497(1), 39-51.
Jędrzejczak M.F. (2002a) Stranded Zostera marina L. vs wrack fauna community interactions on a
Baltic sandy beach (Hel , Poland ): a short-term pilot study . Part I. Driftline effects of
fragmented detritivory , leaching and decay rates. Oceanology, 44, 273-286.
Joergensen, R. G., & Brookes, P. C. (1990). Ninhydrin-reactive nitrogen measurements of
microbial biomass in 0.5 M K2SO4 soil extracts. Soil Biology and Biochemistry, 22(8), 1023-
1027.
Kirkman H, Kendrick GA. 1997. Ecological significance and commercial harvesting of drifting and
beach-cast macro-algae and seagrass in Australia: a review. J Appl Phycol 9:311–326.
Kirschbaum, M.U.F., 2004. Soil respiration under prolonged soil warming: are rate reductions
caused by acclimation or substrate loss? Global Change Biology 10, 1870-1877.
Knorr, W., Prentice, I.C., House, J.I., Holland, E.A., 2005. On the available evidence for the
temperature dependence of soil organic carbon. Biogeosciences Discussions 2, 749-755.
Kowalenko, C.G., Cameron, D.R., 1976. Nitrogen transformations in an incubated soil as affected
by combinations of moisture content and temperature and sorption-fixation of ammonium.
Canadian Journal of Soil Science 56, 63-70.
Leirós, M.C., Trasar-Cepeda, C., Seoane, S., Gil-Sotres, F., 1999. Dependence of mineralization of
soil organic matter on temperature and moisture. Soil Biology and Biochemistry 31, 327-335.
Linn, D.M., Doran, J.W., 1984. Effect of water-filled pore space on carbon dioxide and nitrous
oxide production in tilled and nontilled soils. Soil Science Society of America Journal 48,
1667-1672.
Lomander, A., Kätterer, T., Andrén, O., 1998. Carbon dioxide evolution from top- and subsoil as
affected by moisture and constant and fluctuating temperature. Soil Biology and Biochemistry
30, 2017-2022.
Lowry, O. H., & Rosenbrough, N. J. (1951). Protein measurement with the folin phenol reagent.
Journal of Biology and Chemistry 193, 265–275.

83
Markwell, M. A. K., Haas, S. M., Bieber, L. L., & Tolbert, M. E. (1978). A modification of the Lowry
procedure to simplify protein determination in membrane and lipoprotein samples. Annals of
Biochemistry 87, 206–210.
McClain, M.E., Boyer, E.W., Dent, C.L., Gergel, S.E., Grimm, N.B., Groffman, P.M., Hart, S.C.,
Harvey, J.W., Johnston, C.A., Mayorga, E., McDowell, W.H., Pinay, G., 2003. Biogeochemical
hot spots and hot moments at the interface of terrestrial and aquatic ecosystems. Ecosystems
6, 301–312.
Miranda, R., Alavi, G., Aliaga, S., & Caballero, A. (2013). Mineralización del nitrógeno y generación
de CO2 por descomposición del estiércol ovino. Altiplano central, Bolivia. Venesuelos, 21(2).
Moorhead, D. L., Sinsabaugh, R. L., Linkins, A. E. y Reynolds, J. F., 1996. Decomposition
processes: modelling approaches and applications. The Science of the Total Environment 183:
137-149.Waksman, S.A. y Tenney, F.G. 1928. Composition of natural organic materials and
their decomposition in the soil: III. The influence of nature of plant upon the rapidity of its
decomposition. Soil Science 26:155-171.
Moreira, F. M. y J. O. Siqueira. 2006. Microbiologia y Bioquímica do solo. 2da. Edic. Editorial
Lavras, Universidade Federal de Lavras. 626 p.
Newell, R. C. & J. G. Field, 1983. The contribution of bacteria and detritus to carbon and nitrogen
flow in a benthic community.Mar. Biol. Lett. 4: 23–36.
Nicorladot, B., Fauvet, G., Cheneby, D., 1994. Carbon and nitrogen cycling through soil microbial
biomass at various temperatures. Soil Biology and Biochemistry 26, 253- 261.
Olabarria C., Lastra M. & Garrido J. (2007) Succession of macrofauna on macroalgal wrack of an
exposed sandy beach: effects of patch size and site. Marine Environmental Research, 63, 19-
40.
Pearson, T. H., & Rosenberg, R. (1978). Macrobenthic succession in relation to organic enrichment
and pollution of the marine environment. Oceanogr. Mar. Biol. Ann. Rev, 16, 229-311.
Pöhhacker, R., Zech, W., 1995. Influence of temperature on CO2 evolution, microbial biomass C
and metabolic quotient during decomposition of two humic forest horizons. Biology and
Fertility of Soils 19, 239-245.
Polis GA, Hurd SD. 1996. Linking marine and terrestrial food webs: allochthonous input from the
ocean supports high secondary productivity on small islands and coastal land communities.
Am Nat 147:396– 423.

84
Poore A.G.B. & Hill N.A. (2006) Sources of variation in herbivore preference: among-individual
and past diet effects on amphipod host choice. Marine Biology, 149, 1403-1410
Pusceddu, A., Dell’Anno, A., Fabiano, M., & Danovaro, R. (2009). Quantity and bioavailability of
sediment organic matter as signatures of benthic trophic status. Marine Ecology Progress
Series, 375, 41-52.
Pusceddu A, Bianchelli S, Gambi C, Danovaro R (2011) Assessment of benthic trophic status of
marine coastal ecosystems: Significance of meiofaunal rare taxa; Estuarine, Coastal and Shelf
Science. 93:420-430.
Rey, M., 2000. Desarrollo de una metodología analítica para la determinación de gases
invernadero. Aplicación al estudio de la influencia del cambio climático global sobre la
mineralización de la materia orgánica del suelo, Tesis Doctoral, Universidad de Santiago de
Compostela, 251 pp.
Rey, A., Jarvis, P., 2006. Modelling the effect of temperature on carbon mineralization rates across
a network of European forest sites (FORCAST). Global Change Biology 12, 1894-1908.
Rossi F. & Underwood A. (2002) Small-scale disturbance and increased nutrients as influences on
intertidal macrobenthic assemblages: experimental burial of wrack in different intertidal
environments. Marine Ecology Progress Series, 242, 29-39.
Sinsabaugh, R.L. y Moorhead, D.L. 1997. Synthesis of litter quality and enzyme approaches to
decomposition modelling. En Driven by Nature: Plant litter quality and Decomposition.
Cadish, G y K.E. Giller (Eds)., CAB International. Wallinford, Oxon,UK.
Sinsabaugh, R.L, Carreiro, M.M. y Álvarez, S. 2002. Enzyme and Microbial Dynamics of Litter
Decomposition (Chapter 9). Enzymes in the Environment. Burns, R.G, R.P. Dick. (Eds.),
Marcel Dekker, New York. pp 249-266.
Skujins, J.J., 1967. Enzymes in soil. En: McLaren, A.D., Peterson, G.H. (Eds.) Soil Biochemistry,
vol 1, Marcel Dekker, New York, pp 371-414.
Smith V H, (2003) Eutrophication of freshwater and coastal marine ecosystems. A global problem;
Environmental Science and Pollution Research.10:126-139.
Tenore KR, Hanson RB (1980) Availability of detritus of different types and ages to apolychaete
macroconsumer, Capitella capitata. Limnol Oceanogr 25:553-558.
Tisdale, S. L., W. L. Nelson y J. D. Beaton. 1985. Soil fertility and fertilizers. 4ta ed. Macmillan:
Publishing Company; p. 754.

85
Tremblay L, Benner R (2006) Microbial contributions to N-immobilization and organic matter
preservation in decaying plant detritus; Geochimica et Cosmochimica Acta. 70:133-146.
Van Alstyne K.L., McCarthy III J.J., Hustead C.L. & Duggins D.O. (1999) Geographic variation in
polyphenolic levels of Northeastern Pacific kelps and rockweeds. Marine Biology, 133, 371-379
Venturini N, Pita A, Brugnoli E, García-Rodriguez F, Burone L, Kandratavicius N, Hutton M,
Muniz P M (2012) Benthic trophic status of sediments in a metropolitan area (Rio de la Plata
estuary): Linkages with natural and human pressures; Estuarine, Coastal and Shelf Science.
112:139-152.
Visser S., Parkinson D. (1992): Soil biological criteria as indicators of soil quality: soil
microorganisms. American Journal of Alternative Agriculture, 7: 33–37.
Wardle, D.A., Lavelle, P., 1997. Linkages between soil biota, plant litter quality and decomposition.
En: Cadish, G., Giller, K.E. (Eds.) Driven by Nature, Plant litter.
Wentworth, C. K. (1922). A scale of grade and class terms for clastic sediments. The Journal of
Geology, 30(5), 377-392.
Williams, P. M. & A. F. Carlucci, 1976. Bacterial utilisation of organic matter in the deep-sea.
Nature 262: 810–811.
Williams, S (1984) Decomposition of the tropical macroalga Caulerpa cupressoides (West) C.
Agardh: Field and laboratory studies. J Exp Mar Biol Ecol 80:109-124.
Williams PB, del Giorgio PA (2005) Respiration in aquatic systems: history and background. The
global significance of respiration in aquatic systems: from single cells to the biosphere. En:
del Giorgio PA, Williams PjleB (eds). Respiration in aquatic ecosystems. Oxford University
Press, Oxford, UK, pp 1–17.
Zöllner, N., & Kirsch, K. (1962). Über die quantitative Bestimmung von Lipoiden (Mikromethode)
mittels der vielen natürlichen Lipoiden (allen bekannten Plasmalipoiden) gemeinsamen
Sulfophosphovanillin-Reaktion. Research in Experimental Medicine, 135(6), 545-561.

86

87

88
INTRODUCCIÓN
La descomposición de materia orgánica constituye un proceso clave y limitante en los ciclos
de macronutrientes por lo que desde muy antiguo se ha intentado comprender los mecanismos que
la controlan (Waksman & Tenney, 1928). Se han desarrollado muchos modelos que relacionan la
descomposición con multitud de variables (climáticas, edáficas, cantidad y calidad del detrito y
microbiológicas) (Moorhead et al., 1996; Álvarez, 2005). El problema que entraña este proceso, es
su modelización ya que no es simplemente el proceso inverso a la producción primaria. La
fotosíntesis es un proceso que, en última instancia, reside y se desarrolla en cada organismo
productor de forma individual, mientras que la descomposición se manifiesta a nivel de la
comunidad. Es un proceso compuesto, constituyendo probablemente el proceso ecológico más
complejo de la biosfera (Sinsabaugh & Moorhead, 1997; Sinsabaugh et al., 2002). Su complejidad e
importancia van de la mano, por lo que muchos esfuerzos están siendo destinados a esclarecer
cuáles son los compartimentos involucrados y sus interacciones para así poder encajar todas las
piezas.
La interface tierra-mar es una franja que soporta una desproporcionada actividad
biogeoquímica respecto a los ecosistemas circundantes debido a la convergencia de recursos de
ambos ecosistemas. A lo largo de esta interface, se acumulan en las playas, grandes cantidades de
detritos marino arrastrado por las mareas y el oleaje. Dicho detrito, también conocido como wrack,
suele estar formado fundamentalmente por macroalgas y/o fanerógamas. Esta materia orgánica es
integrada en los procesos metabólicos de la red alimentaria. El material varado, durante el tiempo
que está depositado en la arena, puede experimentar períodos de mojado al ser arrastrado por las
mareas y ser redistribuido localmente por la acción de las olas y el viento. Esto sugiere que la
dinámica de los depósitos puede variar con la distancia al borde del agua. Además, el espesor de
los depósitos afecta a la temperatura y a la humedad de su interior, lo que a su vez, puede afectar a
las tasas metabólicas y al procesamiento del carbono y de los nutrientes (Kirkman & Kendrick
1997; Coupland et al., 2007).
Los aportes de materia orgánica en intermareales arenosos se definen como puntos
calientes o ―Hot Spots‖ (McClain et al., 2003; Coupland et al., 2007). Estos puntos, evidencian la
importancia de la degradación de los parches de algas que llegan a las playas (McClain et al., 2003)
por ser claves en la conexión de redes alimentarias marinas y terrestres, especialmente cuando los
ecosistemas marinos altamente productivos se yuxtaponen a la baja productividad de los
ambientes terrestres (Polis & Hurd, 1996; Coupland et al., 2007). A pesar de su importancia, estas
interfaces no han recibido mucha atención hasta la fecha. El estudio de los procesos
biogeoquímicos asociados al wrack es esencial para determinar el papel de estos subsidios y el
metabolismo de las playas. Un proceso clave para este estudio, es la respiración de la comunidad
involucrada en la degradación del material de arribazón, ya que proporciona el mejor índice del
flujo de materia orgánica en los ecosistemas (Williams & Del Giorgo, 2005).

89
El proceso de descomposición está influenciado por un amplio grupo de propiedades que
operan a escalas muy diferentes (espaciales y temporales) (Wardle & Lavelle, 1997). Los estudios
de campo que se han realizado para investigar dicho proceso, relacionan la variación diaria de la
respiración (emisión de CO2 y liberación de nutrientes) con las variaciones de la temperatura
(Boone et al., 1998; Davidson et al., 1998; Leirós et al., 1998; Rey et al., 2002). Al efecto de la
temperatura se pueden sumar otra variables medioambientales, tales como la humedad del suelo y
las poblaciones de los microorganismos edáficos (Rey et al., 2002). La edad de la materia orgánica
también es punto de discusión; de hecho se desconoce la sensibilidad de la edad de la materia
orgánica depositada en los sedimentos a los cambios de temperatura; lo que hace que exista un
gran interés en dilucidar este aspecto (Rey & Jarvis, 2006; Curiel Yuste et al., 2007).
Debido a esto, el objetivo principal del presente trabajo fue determinar la hora a la que la
temperatura era máxima y por lo tanto la actividad metabólica; dar a conocer el efecto de la
temperatura sobre el proceso de degradación de los depósitos intermareales de macroalgas
depositadas en las playas a través del análisis de las emisiones del CO2. Además, se analizará el
efecto de otros factores tales como la humedad del sedimento, la edad (distancia del parche a la
línea de costa) y biomasa del parche de algas y la abundancia de macrofauna, sobre el metabolismo
de la playa. Para ello, se realizó un experimento en el drift line, en la zona central de la playa.
Consistió en medir la temperatura y el flujo de CO2 durante 12 horas a intervalos de dos horas
sobre los mismos puntos de estudio.
Con el fin de cumplir dicho objetivo, se plantearon las siguientes HIPÓTESIS: (1) En las
horas centrales del día, coincidiendo con las temperaturas máximas diarias, se liberan mayores
cantidades de CO2. (2) Las algas más frescas tendrán mayor calidad nutricional afectando
positivamente a la liberación de nutrientes y de CO2. (3) El metabolismo de las playas está
supeditado a los factores climáticos (temperatura y humedad) y a la calidad del alimento.
MATERIAL Y MÉTODOS
Área de estudio
Se realizó un estudio en la playa de Abra, perteneciente al municipio de Nigrán
(Pontevedra). Abra está ubicada en la Ensenada de Carreira en el margen suroeste de la Ría de
Vigo (42º 9´N; 8º 49´ 43.1 W). Es una playa urbana sin paseo marítimo. Su estado morfológico es
reflectiva con arena gruesa. Es ventosa con oleaje moderado (Figura 1).

90
Figura 1. Fotografía de satélite de la playa de estudio y de la playa in situ.
Diseño del muestreo
Para analizar la variación diaria del flujo de CO2, la liberación de nutrientes, la abundancia
de la macrofauna, el tipo de alga y la calidad de la materia orgánica, se eligió una franja paralela a
la línea de costa comprendida entre el nivel superior e inferior del drift line. Según la distancia a la
línea de costa se clasificaron tres tipos de algas: Las secas (nivel de marea más antiguo), las
húmedas (nivel de marea intermedio) y las frescas (nivel de marea del mismo día). Dentro de esta
franja se eligieron 9 parches de algas. Igualmente, se escogieron de cada nivel de marea, 3 puntos
sin parches de algas que fueron utilizados como puntos de control.
Toma de las muestras
Se realizaron mediciones con una frecuencia temporal de 2 horas, durante 12 horas (de 8 a
20 h) utilizando un medidor de flujo en continuo con un detector LI-COR de CO2 (µmol C m-2 s-1)
(volumen de la campana de acumulación de 2.756 10-3 m3 y un área en su base de 3.140 10-2 m-2).
Las temperaturas del aire y del sedimento, se midieron con termómetro de campo; para el
sedimento, el termómetro se introdujo 5 cm en el sustrato bajo el parche de algas. Una vez
finalizado el ciclo de 12 horas (20:00), los parches de algas de cada punto de estudio se metieron
en bolsas de plástico y fueron transportados al laboratorio (Figura 2). En cada uno de los puntos se
recogieron muestras del sedimento en botes de plástico estériles para el análisis de nutrientes en
forma de nitrógeno inorgánico (NH4+, NO2
- y NO3- µM g-1 sed.), la determinación del carbono
biopolimérico (CBP) (µg g-1 sed.), el cálculo la relación proteína-carbohidrato (PRT:CHO) y el
porcentaje de humedad del sedimento. Las muestras de macrofauna se recogieron con un corer de
10 cm de diámetro hasta una profundidad del sedimento de 10 cm (área= 0.16 m-2) de forma
aleatoria alrededor del punto de muestreo (n=3). Las muestras fueron tamizadas in situ a través de

91
una malla de 1 mm y se guardaron en bolsas plásticas. Una vez en el laboratorio, a cada una se le
añadió una solución de agua de mar y formol al 4%. Los individuos se determinaron al nivel
taxonómico más bajo posible. Una vez en el laboratorio, todas las muestras se congelaron y
almacenaron a -20ºC hasta su posterior análisis.
Análisis de las muestras
% Humedad del alga y biomasa: Se descongelaron las algas y se separaron del resto
de material varado. Se secaron en la estufa a 60º C durante 48 horas para obtener la relación Peso
fresco: Peso seco. De esta relación se obtuvo el porcentaje de humedad del alga. A continuación se
molieron y se tomó una alícuota de 5g de peso seco que se calcinó en la mufla durante 4h a 500ºC.
Este método permite eliminar por combustión la materia orgánica quedando la inorgánica, que por
diferencia, permite conocer la cantidad y el porcentaje de materia orgánica contenida en los 5 g de
muestra. De esta forma se obtuvo la biomasa algal (g peso seco libre de cenizas m-2) en el parche de
algas mediante el producto de este porcentaje y el peso total del parche.
%Humedad del sedimento: El porcentaje de humedad del sedimento se obtuvo a partir
de la pérdida de peso tras el secado de las muestras durante 72 horas a 60ºC.
Nitrógeno inorgánico (amonio, nitrito y nitrato): Se extrajeron del sedimento
utilizando una disolución de cloruro de calcio como disolución extractora. Una vez obtenida, se
congelaron las muestras en tubos Falcon. Posteriormente fueron llevadas a la ―Unidad de Análisis
Elemental‖ de la Universidad de Vigo para su análisis. Las muestras se analizaron con un
Autoanalizador de Flujo continuo Segmentado. La determinación se realiza de forma directa frente
a patrones acuosos.
Análisis bioquímicos: Se realizaron sobre muestras de sedimento previamente
liofilizadas. La cantidad de sedimento que se tomó para cada análisis fue de 2.5 g. Los datos se
expresaron en µg g-1 de peso seco. Los lípidos totales se extrajeron directamente del sedimento por

92
elución directa con una disolución cloroformo:metanol (2:1, v/v). Posteriormente se analizó por el
método de Zöllner y Kirsch (1962). Para analizar los carbohidratos totales, se tuvo que extraer
previamente con ácido tricloroacético (TCA) al 5% mediante el método descrito por Dubois et al.,
(1956). Por último, las proteínas totales también se tuvieron que extraer previamente a su análisis
mediante el método de Lowry & Rosebrough (1951) modificado por Markwell et al., (1978). Las
concentraciones fueron expresadas como equivalentes de seroalbúmina bovina (BSA).
Calidad de la materia orgánica: carbono biopolimérico (CBP): A partir de las
concentraciones de los lípidos, carbohidratos y proteínas se determinó el valor nutricional del
sedimento (CBP). Para eso se tuvo que transformar cada concentración en equivalente de carbono
asumiendo los factores de conversión específicos para cada compuesto (0.45 carbohidratos, 0.5
proteínas, 0.7 lípidos) (Fabiano et al., 1995). La suma del carbono de lípidos, proteínas y
carbohidratos resultó ser el CBP.
“Edad del alga”: Es la relación proteínas:carbohidratos (Cauwet, 1978; Fabiano et al.,
1997). Mediante este cálculo, se corroboró la clasificación del tipo de alga en función de la distancia
a la línea de costa.
Análisis estadístico
Para determinar la variación del CO2 (µmol C m-2 s-1) asociada a la antigüedad del alga, se
realizó un análisis de varianza (ANOVA) ortogonal de dos vías con ―tiempo‖ (6 niveles) y ―edad del
alga‖ (3 niveles) como factores fijos del análisis (paquete estadístico WinGMAV5; EICC,
Universidad de Sidney). Los tres tipos de alga fueron usados como réplicas (n=3). Previamente los
datos fueron transformados para cumplir las asunciones de normalidad y homogeneidad de
varianzas, evaluadas con los test de Sapiro-Wilk y Cochran respectivamente. A posteriori, se realizó
un análisis de comparaciones múltiples mediante el test de Student-Newman-Keul (SNK) (α=
0.05) cuando hubo diferencias significativas entre los factores. La relación entre las variables (CO2,
nutrientes, CBP y PRT:CHO) y los factores ambientales (temperatura, % humedad del sedimento,
biomasa algal y abundancia de individuos) se analizó mediante regresiones múltiples. Se
obtuvieron modelos que ajustan la variación de las variables estudio a dichos factores.
RESULTADOS
Variación diaria del CO2
Efecto de la temperatura
Durante las 12 horas que duró el experimento se observó una clara relación entre la hora
del día, la temperatura y la cantidad de CO2 emitida. La temperatura aumentó linealmente desde
las 8 h hasta las 16 h; a partir de esta hora, descendió ligeramente. Los mayores incrementos de
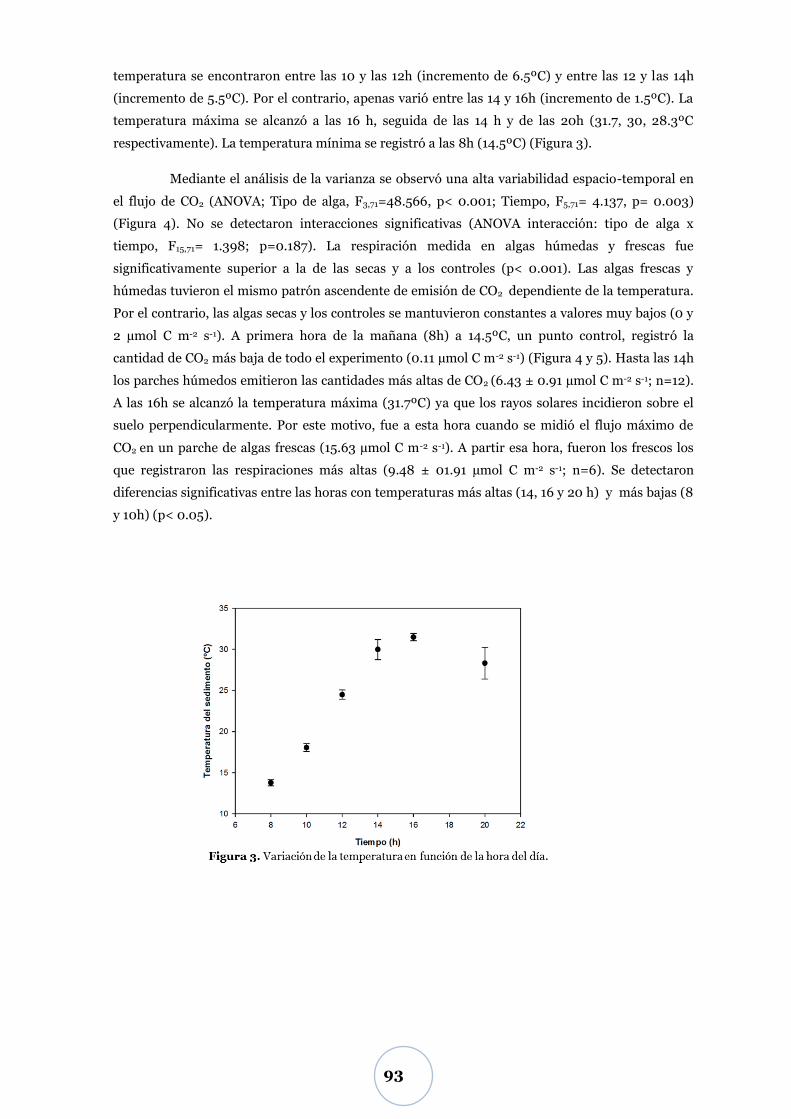
93
temperatura se encontraron entre las 10 y las 12h (incremento de 6.5ºC) y entre las 12 y las 14h
(incremento de 5.5ºC). Por el contrario, apenas varió entre las 14 y 16h (incremento de 1.5ºC). La
temperatura máxima se alcanzó a las 16 h, seguida de las 14 h y de las 20h (31.7, 30, 28.3ºC
respectivamente). La temperatura mínima se registró a las 8h (14.5ºC) (Figura 3).
Mediante el análisis de la varianza se observó una alta variabilidad espacio-temporal en
el flujo de CO2 (ANOVA; Tipo de alga, F3,71=48.566, p< 0.001; Tiempo, F5,71= 4.137, p= 0.003)
(Figura 4). No se detectaron interacciones significativas (ANOVA interacción: tipo de alga x
tiempo, F15,71= 1.398; p=0.187). La respiración medida en algas húmedas y frescas fue
significativamente superior a la de las secas y a los controles (p< 0.001). Las algas frescas y
húmedas tuvieron el mismo patrón ascendente de emisión de CO2 dependiente de la temperatura.
Por el contrario, las algas secas y los controles se mantuvieron constantes a valores muy bajos (0 y
2 µmol C m-2 s-1). A primera hora de la mañana (8h) a 14.5ºC, un punto control, registró la
cantidad de CO2 más baja de todo el experimento (0.11 µmol C m-2 s-1) (Figura 4 y 5). Hasta las 14h
los parches húmedos emitieron las cantidades más altas de CO2 (6.43 ± 0.91 µmol C m-2 s-1; n=12).
A las 16h se alcanzó la temperatura máxima (31.7ºC) ya que los rayos solares incidieron sobre el
suelo perpendicularmente. Por este motivo, fue a esta hora cuando se midió el flujo máximo de
CO2 en un parche de algas frescas (15.63 µmol C m-2 s-1). A partir esa hora, fueron los frescos los
que registraron las respiraciones más altas (9.48 ± 01.91 µmol C m-2 s-1; n=6). Se detectaron
diferencias significativas entre las horas con temperaturas más altas (14, 16 y 20 h) y más bajas (8
y 10h) (p< 0.05).
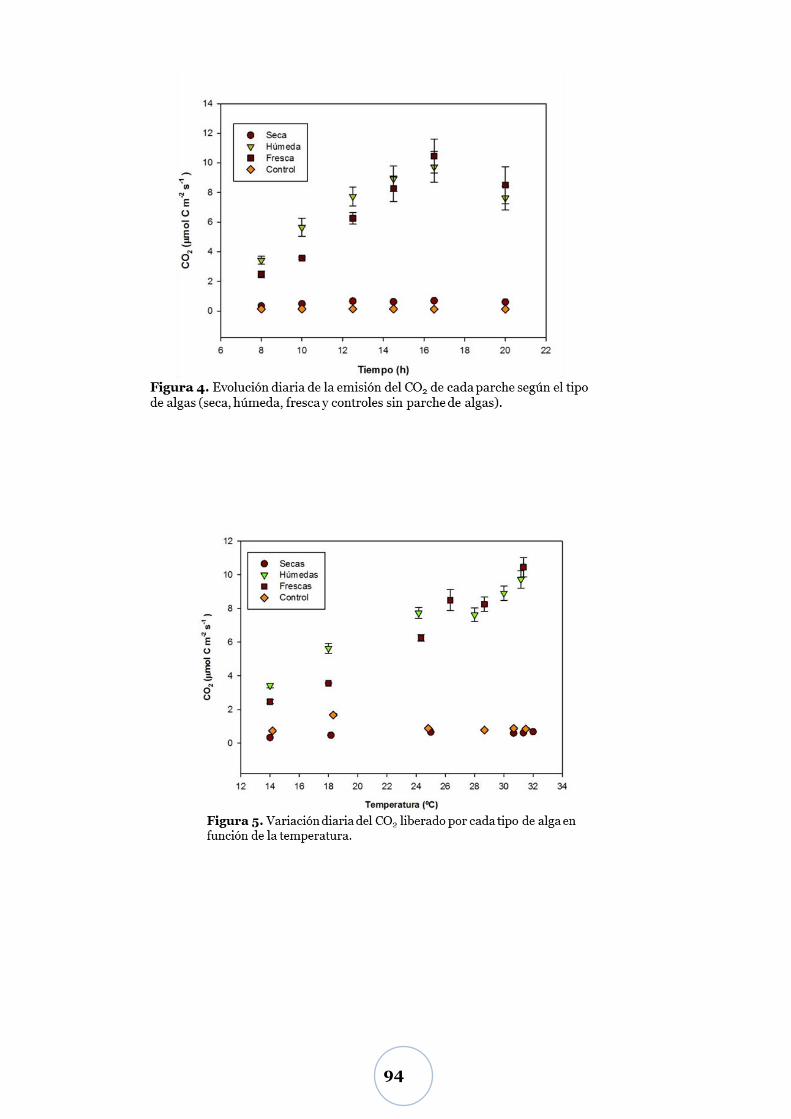
94
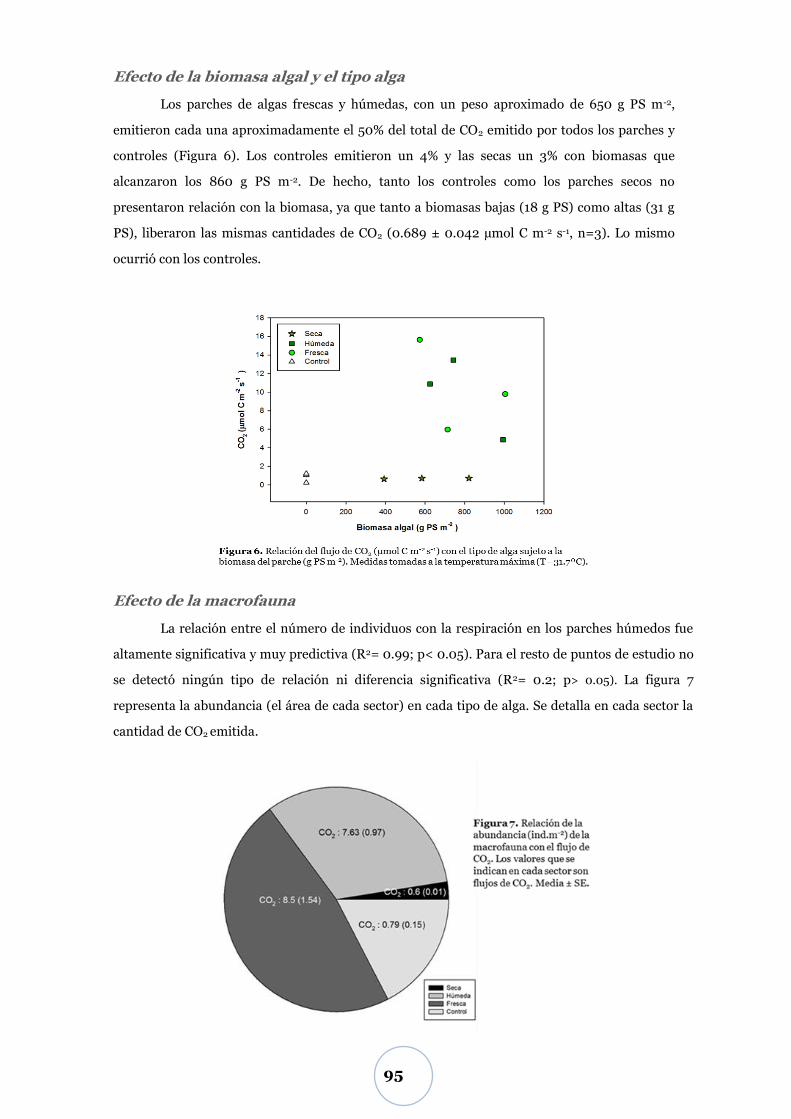
95
Efecto de la biomasa algal y el tipo alga
Los parches de algas frescas y húmedas, con un peso aproximado de 650 g PS m-2,
emitieron cada una aproximadamente el 50% del total de CO2 emitido por todos los parches y
controles (Figura 6). Los controles emitieron un 4% y las secas un 3% con biomasas que
alcanzaron los 860 g PS m-2. De hecho, tanto los controles como los parches secos no
presentaron relación con la biomasa, ya que tanto a biomasas bajas (18 g PS) como altas (31 g
PS), liberaron las mismas cantidades de CO2 (0.689 ± 0.042 µmol C m-2 s-1, n=3). Lo mismo
ocurrió con los controles.
Efecto de la macrofauna
La relación entre el número de individuos con la respiración en los parches húmedos fue
altamente significativa y muy predictiva (R2= 0.99; p< 0.05). Para el resto de puntos de estudio no
se detectó ningún tipo de relación ni diferencia significativa (R2= 0.2; p> 0.05). La figura 7
representa la abundancia (el área de cada sector) en cada tipo de alga. Se detalla en cada sector la
cantidad de CO2 emitida.
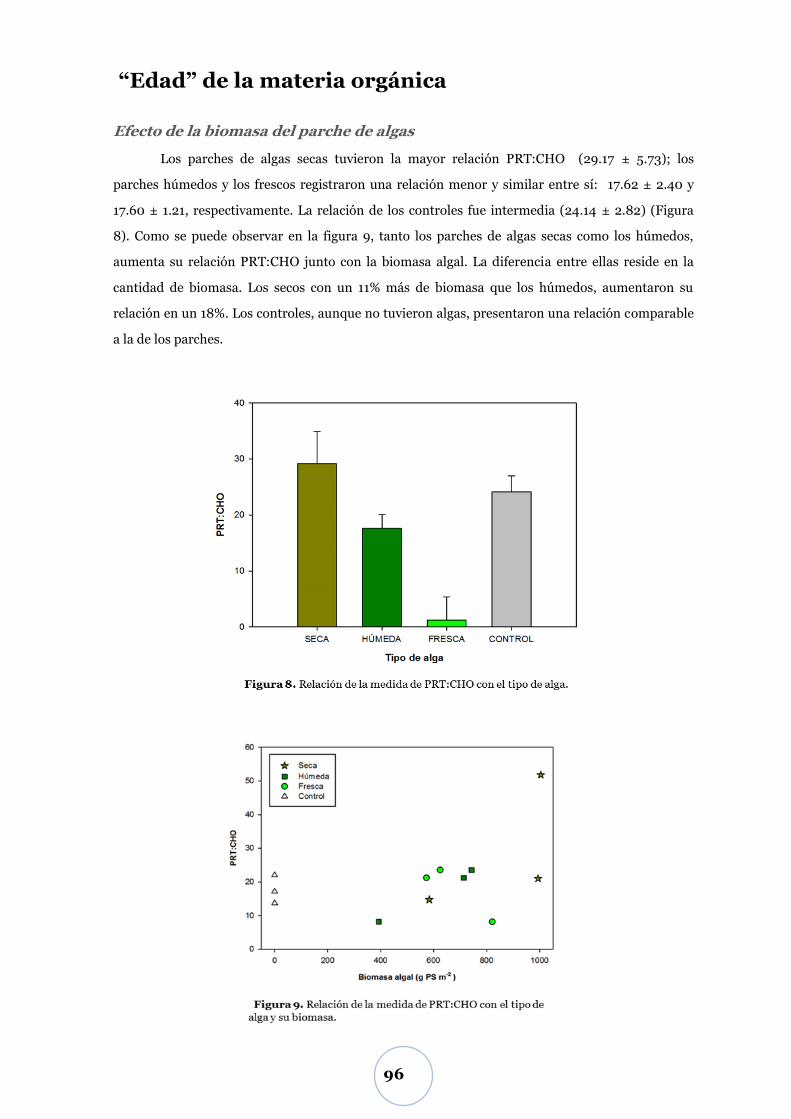
96
“Edad” de la materia orgánica
Efecto de la biomasa del parche de algas
Los parches de algas secas tuvieron la mayor relación PRT:CHO (29.17 ± 5.73); los
parches húmedos y los frescos registraron una relación menor y similar entre sí: 17.62 ± 2.40 y
17.60 ± 1.21, respectivamente. La relación de los controles fue intermedia (24.14 ± 2.82) (Figura
8). Como se puede observar en la figura 9, tanto los parches de algas secas como los húmedos,
aumenta su relación PRT:CHO junto con la biomasa algal. La diferencia entre ellas reside en la
cantidad de biomasa. Los secos con un 11% más de biomasa que los húmedos, aumentaron su
relación en un 18%. Los controles, aunque no tuvieron algas, presentaron una relación comparable
a la de los parches.

97
Liberación de nitrógeno inorgánico
Efecto de la biomasa algal y tipo de alga
La concentración de amonio predominó en todos los puntos de estudio aunque fue en los
parches húmedos donde se midió la concentración más alta (857.56 ± 104.35 µM g-1 sed.). De
hecho, duplicó el valor obtenido a los secos y a los frescos, y cuadriplicó a los controles. Los
parches húmedos también destacaron por contener las concentraciones más bajas de nitritos y
nitratos (32.22 ± 2.32 y 102.56 ± 17.62 µM g-1 sed. respectivamente). Los parches secos registraron
la máxima concentración de nitrito y la mínima de nitrato. En los parches frescos, las
concentraciones de nitritos y nitratos fueron similares a éstos. Por último, los controles siguieron
una tendencia in crescendo (en cada fase de mineralización) en las concentraciones a lo largo de la
mineralización partiendo desde el inicio con la concentración de amonio más baja de todos los
puntos de estudio (Figura 10).
La cantidad de biomasa algal del parche fue determinante a la hora de su degradación y
posterior liberación de nutrientes. La relación de la biomasa algal con el amonio fue positiva, en
cambio para el nitrito y nitrato fue negativa en todos los parches. Las proporciones entre las
distintas especies tuvieron un carácter inverso entre el amonio con los nitritos y nitratos.
Aproximadamente el 70% del nitrógeno inorgánico de los puntos de estudio está en forma de
amonio, excepto para los parches húmedos donde el amonio alcanzó el 92%. Para todos, las
diferencias entre nitritos y nitratos fue de 1.5%, siendo superior la concentración de nitrato. En los
parches secos, la cantidad de ambas especies de nitrógeno fue la misma (Figura 11).
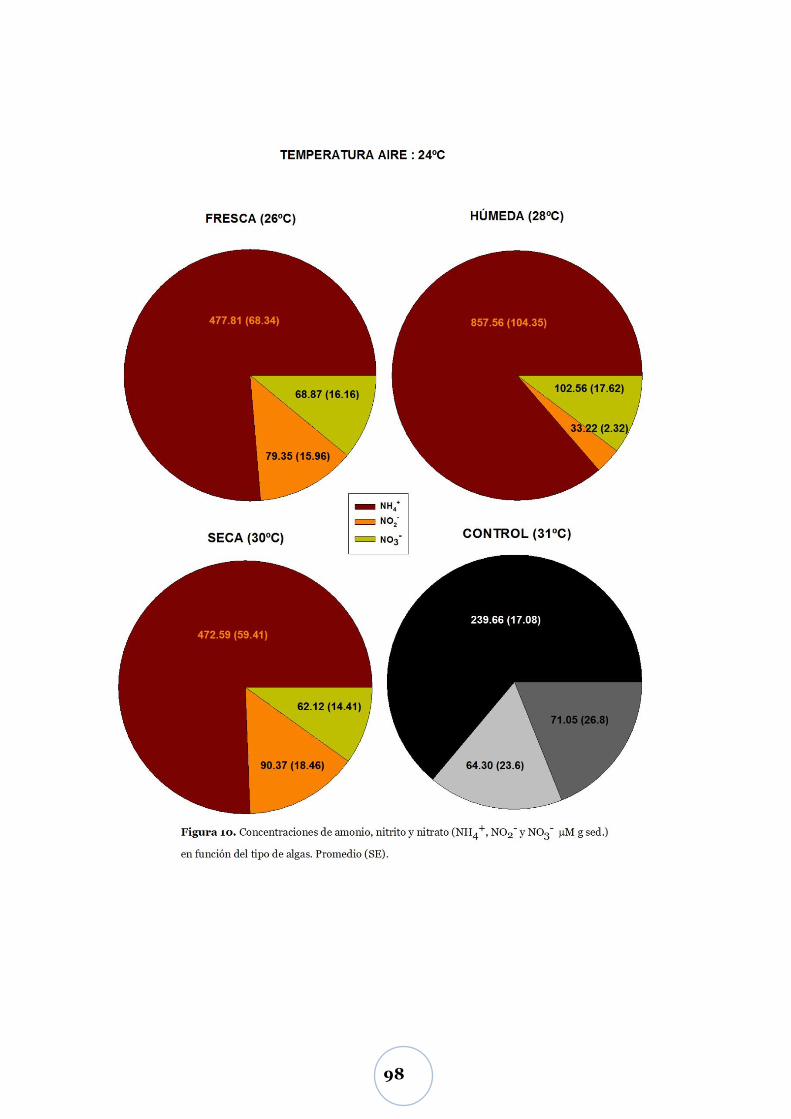
98
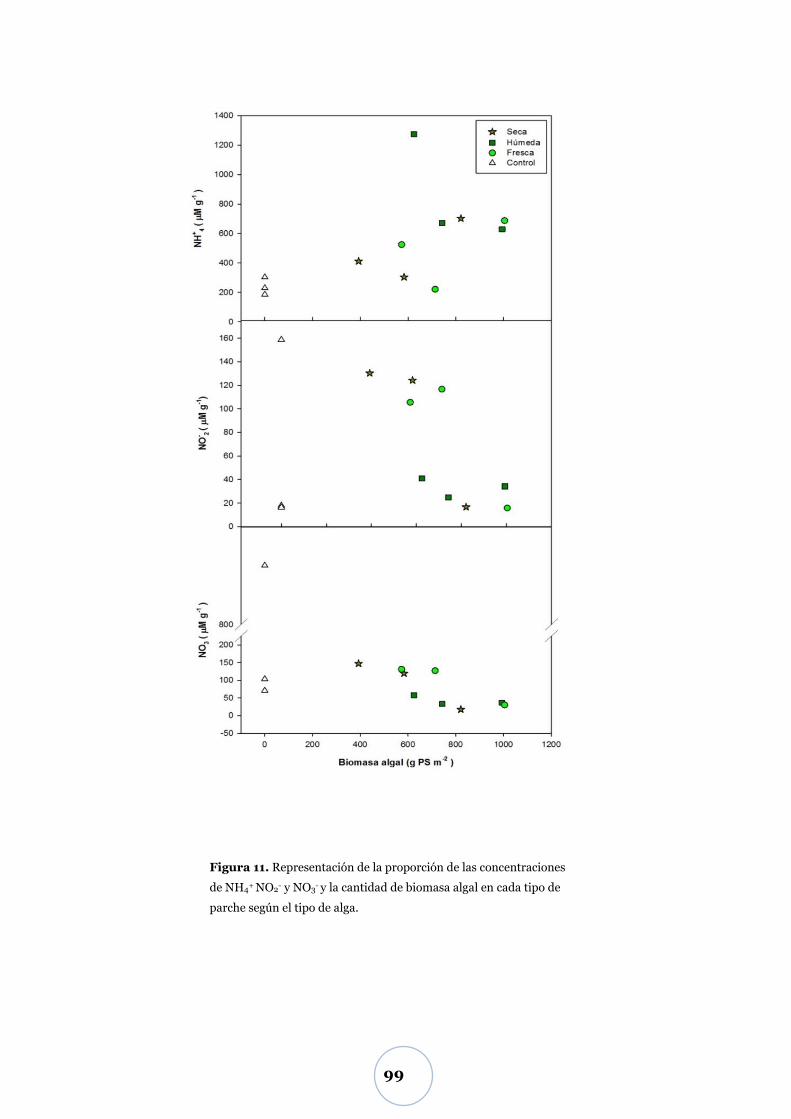
99
Figura 11. Representación de la proporción de las concentraciones
de NH4+ NO2
- y NO3- y la cantidad de biomasa algal en cada tipo de
parche según el tipo de alga.
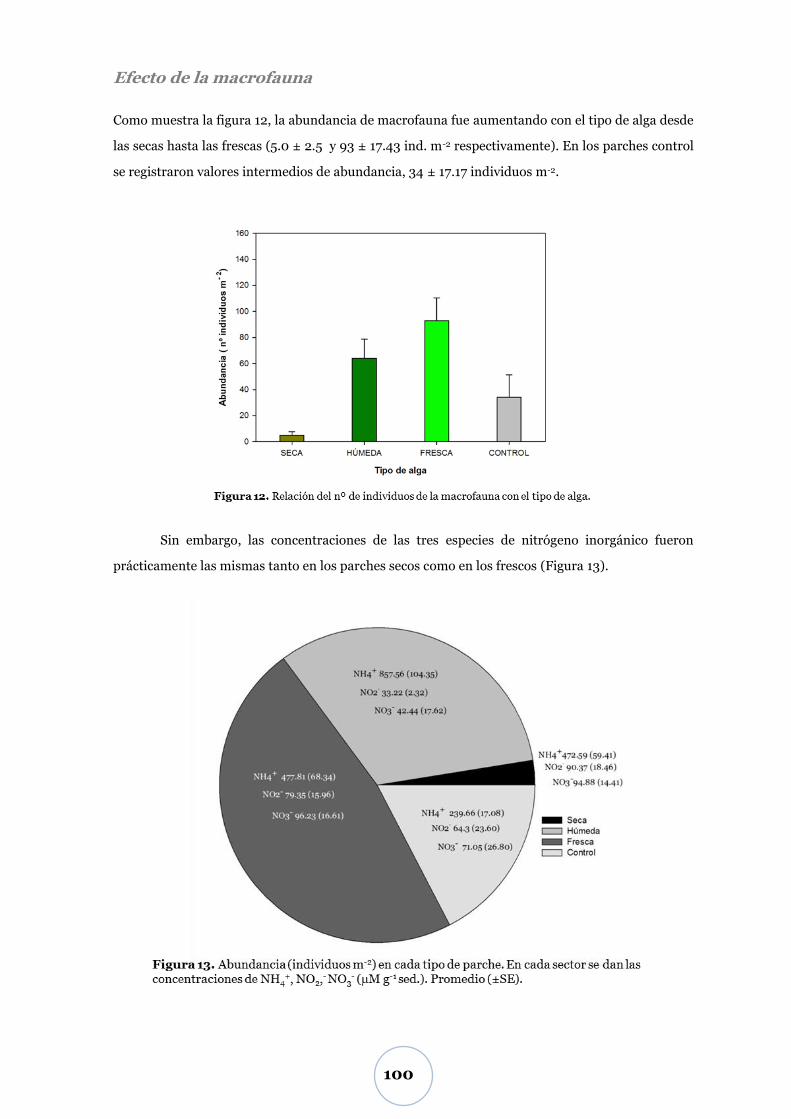
100
Efecto de la macrofauna
Como muestra la figura 12, la abundancia de macrofauna fue aumentando con el tipo de alga desde
las secas hasta las frescas (5.0 ± 2.5 y 93 ± 17.43 ind. m-2 respectivamente). En los parches control
se registraron valores intermedios de abundancia, 34 ± 17.17 individuos m-2.
Sin embargo, las concentraciones de las tres especies de nitrógeno inorgánico fueron
prácticamente las mismas tanto en los parches secos como en los frescos (Figura 13).
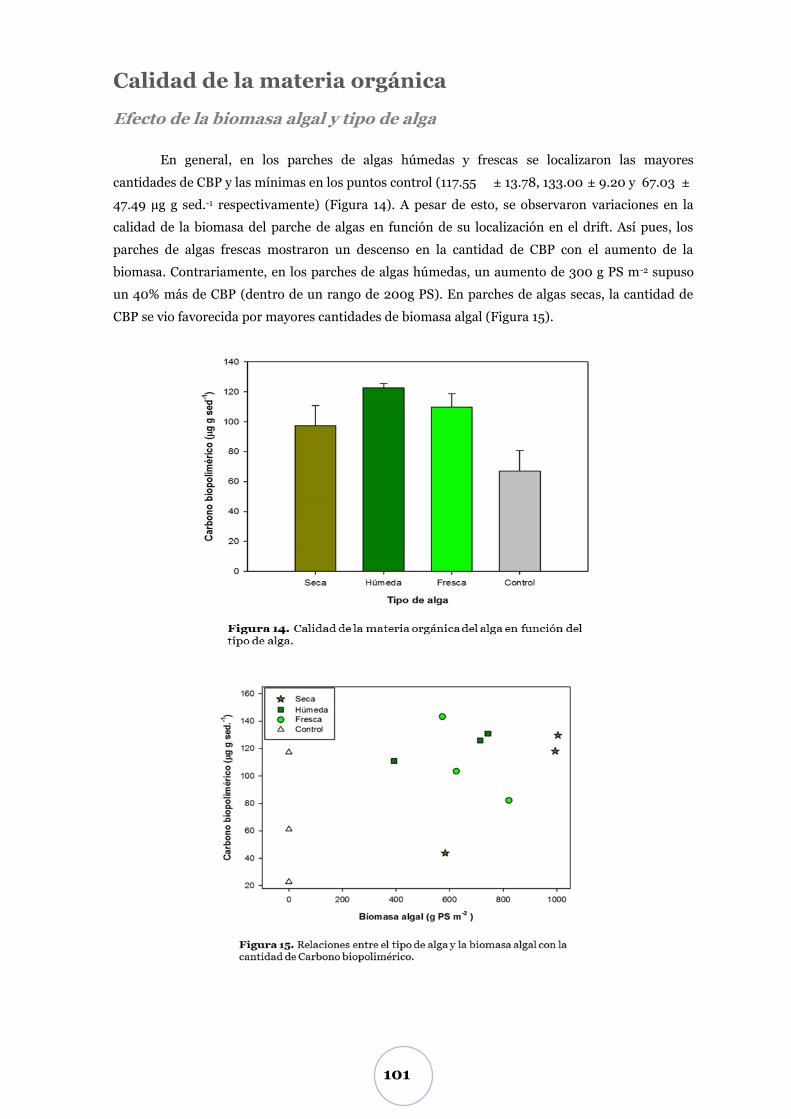
101
Calidad de la materia orgánica
Efecto de la biomasa algal y tipo de alga
En general, en los parches de algas húmedas y frescas se localizaron las mayores
cantidades de CBP y las mínimas en los puntos control (117.55 ± 13.78, 133.00 ± 9.20 y 67.03 ±
47.49 µg g sed.-1 respectivamente) (Figura 14). A pesar de esto, se observaron variaciones en la
calidad de la biomasa del parche de algas en función de su localización en el drift. Así pues, los
parches de algas frescas mostraron un descenso en la cantidad de CBP con el aumento de la
biomasa. Contrariamente, en los parches de algas húmedas, un aumento de 300 g PS m-2 supuso
un 40% más de CBP (dentro de un rango de 200g PS). En parches de algas secas, la cantidad de
CBP se vio favorecida por mayores cantidades de biomasa algal (Figura 15).

102
DISCUSIÓN
Variación diaria del CO2
Efecto de la temperatura
Todos los procesos involucrados en la degradación de la materia orgánica están catalizados
por enzimas que intervienen en las reacciones bioquímicas de los organismos (Kirschbaum, 2004;
Knorr et al., 2005). De este modo, los factores que afectan a las enzimas, afectarán al metabolismo
microbiano y modificarán la velocidad de la mineralización. La actividad enzimática depende,
fundamentalmente, de la temperatura y de la humedad, por lo tanto, determina la actividad
biológica y la cantidad de CO2 que se emite como producto de las reacciones (Visser & Parkinson,
1992). Cualquier enzima tiene una temperatura óptima de actuación a la cual se observa un valor
máximo de actividad. El valor de temperatura será característico para cada enzima (Skujins, 1967;
Frankenberger & Tabatabai, 1991). Por lo tanto, Davidson et al. (2006), consideraron que la
descomposición de la materia orgánica se podría describir haciendo uso de la teoría cinética y las
restricciones ambientales (temperatura y humedad).
Los resultados obtenidos en el experimento concuerdan con lo anteriormente explicado. Al
igual que en los estudios de Xu, M. et al. (2001) y Shi, P. L. et al. (2006), desde la hora de comienzo
del experimento (a las 8:00 h), la temperatura va en aumento alcanzando el máximo a las 16:00 h
(31,7ºC) y de forma paralela, la emisión de CO2. Este comportamiento resulta lógico si se tiene en
cuenta que la temperatura óptima de crecimiento para la mayoría de los microorganismos se
encuentra entre 25 y 35ºC (Guntiñas, 2009). Este patrón fue el mismo para los tres tipos de
parches de algas, pero los niveles de emisión de cada uno fueron distintos. Esto es debido a la
distinta sensibilidad de cada tipo de parche según su edad, la cual viene determinada por la calidad
del sustrato orgánico derivado de la actividad microbiana y su contenido en humedad (Guntiñas,
2009). Según estudios de Rey & Jarvis (2006), la sensibilidad de la fracción recalcitrante es menor
que la de las fracciones más lábiles; por lo tanto, esta última se mineraliza más rápidamente a
medida que la temperatura aumenta. La sensibilidad del carbono orgánico remanente en el
sedimento será menor aun cuando la temperatura siga incrementándose. Según los resultados,
hasta las 14:00 h, los parches húmedos fueron los que más CO2 emitieron; es a partir de esta hora,
cuando los parches frescos empiezan a superar los niveles de los húmedos. Por otro lado, los
niveles de emisión de los parches secos y de los controles permanecen prácticamente constantes y
muy inferiores. Esto pone de manifiesto que desde el nivel inferior de humedad hasta la capacidad
de campo (punto de saturación del suelo), la respiración del sedimento aumenta progresivamente
independientemente del tipo de sedimento y para todas las temperaturas (Lomander et al., 1998;
Davidson et al., 2000). La explicación a esto reside en la estructura del sedimento. Los
microorganismos demandan oxígeno y nutrientes solubles. En condiciones cercanas a la capacidad

103
de campo, los macroporos están llenos de aire facilitando la difusión del oxígeno (Linn et al., 1984)
y los microporos llenos de agua facilitando la difusión de los sustratos solubles. Hay que tener en
cuenta que el contenido en agua es de tipo cúbico y cualquier cambio en la cantidad de agua
provoca fuertes restricciones en la difusión de solutos, máxime si entre ambos existe dependencia
ya que provocará una disminución en la actividad microbiana. . Por eso, los parches frescos, hasta
las 16:00 h, tuvieron un porcentaje elevado de humedad del sedimento, lo cual impedía emitir
mayores cantidades que las húmedas a pesar de su alta calidad por ser materia orgánica muy
reciente. Pero durante toda la mañana, el contenido en agua del sedimento se fue evaporando y fue
las 16:00 h cuando se alcanzó la humedad cercana a la capacidad de campo (relacionada con la
máxima actividad microbiana) (Guntiñas, 2009). En los controles se observó que el efecto de la
temperatura fue negativo ya que el sedimento alcanzó estrés hídrico. A medida que la temperatura
ascendía, los flujos de CO2, iban descendiendo, con lo que los microorganismos encargados de la
descomposición, se vieron inhibidos por falta de agua en el sedimento. Así pues, se puede concluir
que los parches de algas actúan como aislantes térmicos evitando la evaporación (Duong, 2008)
Efectos del tipo de alga, biomasa del parche de algas y macrofauna
Los parches con menor biomasa fueron los más recientes aunque emitieron mayores
cantidades de CO2 que los parches secos con mayor biomasa. Esto se puede atribuir a que toda la
fracción lábil se descompone rápidamente, por eso un alga recién varada emitirá mayor cantidad
de CO2 que otra que lleve semanas depositada (Jędrzejczak, 2002a; Rossi & Underwood, 2002;
Olabarria et al., 2007). Además, el proceso de secado es más rápido en parches con menor biomasa
algal, acelerando la pérdida total de la materia orgánica del parche (Ochieng & Erftemeijer, 1999;
Olabarria et al., 2007). La edad del detrito, el contenido en lignina y el grado de desintegración del
substrato influye en la actividad de los organismos. De tal forma que el tipo de alga y su biomasa
están relacionadas con la abundancia de individuos de la macrofauna. El patrón de colonización
varía entre parches de diferentes tamaños. Los parches con biomasas pequeñas albergan menor
número de individuos que los de mayor biomasa (Olabarria et al., 2007) y además los organismos
migran hacia desechos recién varados para alimentarse (Jędrzejczak, 2002a).
No obstante, la comparación de los datos obtenidos con los bibliográficos es simplemente
orientativa y debe tomarse con cautela, ya que pueden existir diferencias en la metodología
utilizada.
Tipo de alga y calidad de la materia orgánica
Efecto de la biomasa del parche de algas
El uso de la relación de las concentraciones de proteínas y carbohidratos (PRT:CHO) se
ha venido usando para determinar la ―edad‖ de la materia orgánica (Hobson, 1967; Cawet, 1978).
En las primeras fases de descomposición, las bacterias consumen más rápidamente las proteínas

104
debido a su facilidad de transformación que los carbohidratos (Williams & Carlucci, 1976; Newell
& Field, 1983). Estas proteínas son asimiladas y se transforman en biomasa bacteriana. Por este
motivo, las relaciones PRT:CHO más altas se encontraron bajo parches secos; estas proteínas
formaban parte de la biomasa bacteriana confirmando que el detritus era de nueva generación
(Donovaro et al., 1993). Además, las altas biomasas de los parches secos, propiciaron un
incremento de la actividad microbiana (descomposición y asimilación de la materia orgánica). Por
el contrario, al comienzo de la degradación existe una baja relación PRT:CHO ya que no ha
transcurrido el tiempo suficiente para que la microbiota haya descompuesto la suficiente cantidad
de materia orgánica. Por eso, los parches húmedos y frescos que tuvieron la misma relación
PRT:CHO, fueron los que registraron las relaciones más bajas. En este estudio se registraron
valores medios de 22.13 ± 11.42 siendo muy superiores a los registrados en otros trabajos llevados
a cabo en playas de Galicia (playas expuestas= 12.27 y protegidas= 5.24) (Incera et al., 2003). Esto,
informa de que la materia orgánica que se estaba reciclando había sido depositada recientemente.
En el tipo de playas expuestas con arena gruesa (como ocurre en Abra), no se acumula el detritus
ya que gracias al drenaje y a la gran aireación, los microorganismos reciclan la materia orgánica
con mayor rapidez y el metabolismo de las playas es muy alto.
La materia orgánica lábil se puede calcular mediante el carbono biopolimérico (CBP)
(sumatorio de las concentraciones de todas las proteínas, carbohidratos y lípidos de la materia
orgánica) (Danovaro et al., 1993; Cividanes & Souza, 2003; Pusceddu et al., 2009). La fracción
lábil de la materia orgánica, se denota como la porción fácilmente asimilable por los organismos
(Tenore & Hanson, 1980; Donovaro et al., 1993; Incera et al., 2003). El CBP se ha establecido
como una metodología fiable para la evaluación de la calidad de la materia orgánica sedimentaria y
a través de esta evaluación, se puede obtener una mejor comprensión del estado trófico del
ecosistema. El CBP también aporta información sobre el origen de la materia orgánica (Colombo et
al., 1996; Dell 'Anno et al., 2002; Pusceddu et al., 2009, 2011; Venturni et al., 2012). Los procesos
biogeoquímicos asociados con la mineralización de la materia orgánica en los sedimentos están
fuertemente influenciados por los microorganismos heterotróficos (Findlay et al., 1992; Tremblay
& Benner, 2006) ya que su actividad depende en gran medida de la calidad de la materia orgánica.
Así pues, la comunidad biótica depende de los recursos de carbono para sostener las redes
alimentarias y mantener los organismos que habitan en ellas. La evaluación del estado trófico en
los ambientes costeros es crucial para entender los vínculos de la red alimentaria, así como las
características biogeoquímicas (Smith, 2003). En la presente investigación, la evaluación del
estado trófico se realizó a partir de la cantidad de CBP y la contribución de la biomasa algal. Los
parches más frescos tuvieron mayor cantidad de CBP ya que éstos al tener un bajo porcentaje de
materia degradada, mantienen altas concentraciones de proteínas, lípidos y carbohidratos,
haciéndolos más apetecibles para los organismos. Por el contrario, en los parches secos y en los
controles, gran parte de la materia orgánica está degradada y las concentraciones de proteínas,

105
lípidos y carbohidratos son más bajas. Como cabría esperar, en los parches de mayor biomasa, el
contenido en CBP también aumentó excepto en los parches frescos. Esto pudo deberse al alto
contenido en agua, inhibiendo la actividad microbiana o al alto contenido en fenoles impidiendo su
degradación. Con estos resultados obtenidos en este experimento se pudo clasificar la playa según
su estado trófico. Para ellos se utilizó la clasificación trófica de Pusceddu et al. (2011) basada en las
concentraciones de BPC y a la contribución de las algas al BPC. La clasificación general es la
siguiente: oligotrófico (improductivo: CBP<1 mg C g-1, fracción de alga> 25% de CBP), mesotrófico
(productividad intermedia: BPC= 1-3 mg C g-1, fracción de algas= 12-25% de CBP) y eutrófico
(altamente productivo: BPC> 3 mg C g-1, fracción de alga <12% BPC). En nuestro estudio la media
de CBP fue de 0.099 ± 0.038 mg C g-1, por lo tanto, el estado trófico de la playa de estudio, Abra,
fue claramente oligotrófico. Según Dodds & Cole (2007), la naturaleza del estado trófico puede
estar influenciada por factores como la luz, la fuente externa de carbono, los nutrientes, la
hidrología o la estructura de la red alimentaria. En el caso de Abra, su carácter oligotrófico fue el
esperado, dado que las playas expuestas arenosas son ambientes pobres en nutrientes debido a las
condiciones morfodinámicas que imperan. El tamaño grueso de la arena, la pronunciada pendiente
y las fuertes condiciones hidrodinámicas, no permiten la acumulación de materia orgánica en el
sedimento.
Liberación de nutrientes
Efecto del tipo de alga y biomasa del parche
Las macroalgas, al tener una baja complejidad estructural y un alto potencial para
almacenar grandes cantidades de nutrientes, su descomposición provoca una rápida lixiviación de
compuestos orgánicos y nutrientes inorgánicos (Chapman & Craigie, 1977; Tenore & Hanson,
Williams, 1984; Hanisak, 1993). Las concentraciones más altas de nutrientes generalmente se
localizan bajo los parches de macroalgas (Barreiro et al., 2013). El nitrógeno inorgánico,
especialmente amonio (NH4+) y nitrato (NO3
-) son las formas absorbidas por los autótrofos.
Provienen principalmente de la mineralización de la materia orgánica del suelo vía amonificación y
nitrificación (Tisdale et al., 1985; Miranda et al., 2013). El amonio en el sedimento puede seguir
dos vías, ser consumido por las bacterias heterotróficas (García-Robledo et al., 2008) o puede ser
convertido a nitrato gracias a los microorganismos por medio de la nitrificación. Este proceso es
realizado en dos fases por diferentes bacterias: en la primera los nitrosomonas convierten el
amonio en nitrito (NO2-) y, en la segunda, nitrobacter, oxida el nitrito en nitrato. Este proceso se ve
completado entre dos a cuatro semanas y está dominado básicamente por las condiciones de
temperatura y humedad.
Un factor importante en la cinética de la mineralización es la calidad del material que se
mineraliza (Nicolardot et al., 1994); por lo tanto, la edad del parche del alga va a ser determinante
en la velocidad de liberación de los nutrientes. El detritus de los puntos de estudio fue de nueva

106
generación, por lo que las bacterias consumieron rápidamente las proteínas obteniendo la energía
suficiente para llevar a cabo la mineralización (Incera et al., 2003). El amonio fue la forma
predominante en todos los parches y controles porque la amonificación es menos dependiente de
la humedad del suelo al ser realizada por diversas clases de microorganismos (aeróbicos y
anaeróbicos). Por el contrario, la nitrificación ocurre en un rango más estrecho (Guntiñas, 2009).
El rápido agotamiento de las formas orgánicas más lábiles fue explicado por la acción de las
temperaturas relativamente elevadas (24ºC). Éstas produjeron una despolimerización de la
materia orgánica facilitando el ataque microbiano (Joergensen, 1990). Incluso se ha argüido que la
despolimerización anterior sería a su vez realzada por el incremento de actividad de los
microorganismos, como reacción al estrés generado por la temperatura (Pohhacker & Zech, 1995).
A pesar de que el amonio predominó en todos los puntos de estudio, hubo variaciones
dependientes de la edad del parche de algas. Así pues, en los controles, donde ya no había algas, se
registró un patrón creciente de las concentraciones desde el amonio hasta el nitrato (siendo la
concentración de amonio la más baja de todos los puntos). Los parches húmedos registraron los
máximos de amonio y los mínimos de nitrito y nitrato; esto indica que la mineralización se
encontraba en una primera fase porque las diferencias entre el amonio y los nitritos y nitratos son
muy elevadas (todavía no ha dado tiempo a que se produzca la nitrificación). Posiblemente las
condiciones óptimas de humedad y temperatura propiciaron una mayor actividad metabólica
produciendo estos productos. Por el contrario, en los parches secos, la nitrificación se vio inhibida
por el déficit de humedad, por eso en estos parches la concentración de nitrito es máxima (la
mineralización se ve interrumpida). En los parches de algas frescas se registró la concentración
más alta de nitrato de todos los puntos; esto sugiere que se están formando nitratos a pesar de
tratarse de algas frescas, lo cual indica que el metabolismo se está produciendo a gran velocidad,
favorecido por condiciones óptimas de temperatura y humedad. La biomasa algal no pareció
afectar al proceso de mineralización; las tasas más altas se correspondieron con los parches secos,
y no por ello se liberó más cantidad de amonio que en los húmedos.
Efecto de la macrofauna
En las regiones templadas, la fauna supralitoral en playas arenosas con una entrada de
macrófitos moderada, a menudo están dominadas por anfípodos talítridos (Colombini et al., 2000,
Griffiths & Stenton-Dozey, 1981; Inglis, 1989; Jędrzejczak, 2002); por lo tanto, son considerados
los colonizadores primarios del wrack más reciente (Behbehani & Croker, 1982; Colombini et al.,
2000; Griffiths & Stenton-Dozey, 1981; Inglis, 1989; Marsden, 1991; Stenton-Dozey & Griffiths,
1983). Se sabe que la abundancia y composición específica de la macrofauna asociada a los
depósitos de algas afecta a su descomposición. Estos trabajos apoyan a los resultados obtenidos en
el presente estudio, ya que en los parches frescos se contabilizó casi un 50% de la abundancia de

107
todos los puntos de estudio. En el lado contrario están los parches más secos y los controles.
Además del contenido en humedad, pudo ser la composición bioquímica la que determinó la
elección del alimento y las tasas de consumo de los organismos detritívoros (Cronin & Hay, 1996;
Van Alstyne et al., 1999; Poore & Hill 2006; Gómez, 2015). Los resultados indican que las
abundancias de macrofauna fueron mayores en los parches con algas que en los controles, lo que
se corresponde con resultados de varios estudios de degradación de algas (Olabarria et al., 2007;
Rodil, 2008; Gómez, 2015). Al no tener datos de periodos más dilatados, es difícil obtener
conclusiones claras; la abundancia de fauna en los puntos de estudio depende de la sucesión de las
poblaciones a lo largo del tiempo, y en este estudio sólo se midieron en un momento concreto. Tan
sólo parece evidente que el material más fresco es el más favorable para la presencia de fauna, ya
sea para el consumo, refugio o lugar de cría.
CONCLUSIONES
Los resultados obtenidos han demostrado ciertos patrones de comportamiento de las
variables estudiadas a lo largo del proceso de descomposición de las macroalgas. Se comprobó que
a medida la temperatura aumenta, también aumenta el CO2 siendo máximo en las horas centrales
del día cuando la temperatura es más alta. Cada tipo de alga dependiendo de su estado de
descomposición, responderá de diferente manera ante las variaciones térmicas en cuanto a las
cantidades de CO2 y nutrientes. Por eso, las algas frescas y las húmedas, liberaron más
rápidamente CO2 que las algas secas y que los controles. La distribución de la macrofauna depende
de la cantidad de CBP por eso predominan en los parches de algas frescas que son las que tienen
mayor valor nutricional. Por lo tanto, se demostró que el metabolismo de las playas depende en
última instancia de los factores climáticos como la temperatura y el porcentaje de humedad.

108
BIBLIOGRAFÍA
Álvarez, S. (2005). La descomposición de materia orgánica en humedales: la importancia del
componente microbiano. Revista Ecosistemas, 14(2).
Barreiro F., Gómez M., López J., Lustra M. & la Huz R. (2013) Coupling between macroalgal inputs
and nutrients outcrop in exposed sandy beaches. Hydrobiologia, 700, 73-84.
Boone, R.D., Nadelhoffer, K.J., Canary, J.D., Kaye, J.P., 1998. Roots exert a strong influence on the
temperature sensitivity of soil respiration. Nature 396, 570-572.
Cauwet, G., 1978. Organic chemistry of sea water particulates: concepts and developments.
Oceanol. Acta 1: 99–105.
Chapman aRO, Craigie JS (1977) Seasonal growth in Laminaria longicruris: Relations with
dissolved inorganic nutrients and internal reserves of nitrogen. Mar Biol 40:197-205.
Cividanes, F. J., & Souza, V. P. (2003). Exigências térmicas e tabelas de vida de fertilidade de
Myzus persicae (Sulzer)(Hemiptera: Aphididae) em laboratório. Neotropical Entomology,
32(3), 413-419.
Colombo J C, Silverberg N, Gearing J N (1996) Biogeochemistry of organic matter in the
Laurentian trough, II. Bulk composition of the sediments and relative reactivity of major
components during early diagenesis; Marine Chemistry. 51:295-314.
Coupland, G.T., Duarte, C.M. & Walker, D.I., 2007a. High metabolic rates in beach cast
communities. Ecosystems, 10(8), pp.1341–1350.
Cronin G. & Hay M.E. (1996) Susceptibility to herbivores depends on recent history of both the
plant and the animal. Ecology, 77, 1531-1543
Curiel, Yuste, J., Baldocchi, D.D., Gershenson, A., Goldstein, A., 2007. Microbial soil respiration
and its dependence on carbon inputs, soil temperature and moisture. Global Change Biology
13, 1-18.
Davidson, E.A., Belk, E., Boone, R.D., 1998. Soil water content and temperature as independent or
confounded factors controlling soil respiration in a temperate mixed harwood forest. Global
Change Biology 4, 217-227.
Danovaro, R., M. Fabiano & N. Della Croce, 1993. Labile organic matter and microbial biomasses
in deep-sea sediments (Eastern Mediterranean Sea). Deep-Sea Res. 5: 953–965.

109
Dell’Anno A, Mei M L, Pusceddu A, Danovaro R (2002) Assessing the trophic state and
eutrophication of coastal marine systems. A new approach based on the biochemical
composition of sediment organic matter; Marine Pollution Bulletin. 44:611-622.
Dodds W K, Cole J J (2007) Expanding the concept of trophic state in aquatic ecosystems:It’s not
just the autotrophs; Aquatic Sciences. 69:427-439.
Dubois, M., Gilles, K. A., Hamilton, S. K., & Rebers, P. A. (1956). Colorimetric method for
determination of sugars and related substances. Analytical Chemistry 28, 350–356.
Duong, Huynh Lien Stephanie. 2008. Investigating the ecological implications of wrack removal
on South Australian sandy beaches.
Findlay S, Pace M L, Lints D, Howe K (1992) Bacterial metabolism of organic carbon in tidal
freshwater Hudson estuary; Marine Ecology Progress Series. 89:147-153.
Frankenberger Jr., W.T., Tabatabai, M.A., 1991. L-Glutaminase activity of soils. Soil Biology and
Biochemistry 23, 869-874.
García-Robledo E, Corzo A, García de Lomas J, van Bergeijk S (2008) Biogeochemical effects of
macroalgal decomposition on intertidal microbenthos: a microcosm experiment. Mar Ecol
Prog Ser 356:139 151.
Gómez Rodríguez, Marina (2015). Ecología de los varamientos de macroalgas en playas estuáricas
de la costa de Galicia.
Griffiths, C. L., Stenton-Dozey, J. M. E., & Koop, K. (1983). Kelp wrack and the flow of energy
through a sandy beach ecosystem. In Sandy beaches as ecosystems (pp. 547-556). Springer
Netherlands.
Grundmann, G.L., Renault, P., Rosso, L., Bardin, R., 1995. Differential effects of soil water content
and temperature on nitrification and aeration. Soil Science Society of America Journal 59,
1342-1349.
Guntiñas, M. E. (2009). Influencia de la temperatura y de la humedad en la dinámica de la materia
orgánica de los suelos de Galicia y su relación con el cambio climático. Universidad de
Santiago de Compostela. Santiago de Compostela.
Hanisak M (1993) Nitrogen release from decomposing seaweeds: species andtemperature effects. J
Appl Phycol 5:175-181.
Hobson, L. A., 1967. The seasonal and vertical distribution of suspended particulate matter in an
area of the Northeast Pacific Ocean. Limnol. 12: 642–649.

110
Ince, R., Hyndes, G. A., Lavery, P. S., & Vanderklift, M. A. (2007). Marine macrophytes directly
enhance abundances of sandy beach fauna through provision of food and habitat. Estuarine,
Coastal and Shelf Science, 74(1), 77-86.
Incera, M., Cividanes, S. P., López, J., & Costas, R. (2003). Role of hydrodynamic conditions on
quantity and biochemical composition of sediment organic matter in sandy intertidal
sediments (NW Atlantic coast, Iberian Peninsula). Hydrobiologia, 497(1), 39-51.
Jędrzejczak M.F. (2002a) Stranded Zostera marina L. vs wrack fauna community interactions on a
Baltic sandy beach (Hel , Poland ): a short-term pilot study . Part I. Driftline effects of
fragmented detritivory , leaching and decay rates. Oceanology, 44, 273-286.
Joergensen, R. G., & Brookes, P. C. (1990). Ninhydrin-reactive nitrogen measurements of
microbial biomass in 0.5 M K2SO4 soil extracts. Soil Biology and Biochemistry, 22(8), 1023-
1027.
Kirkman H, Kendrick GA. 1997. Ecological significance and commercial harvesting of drifting and
beach-cast macro-algae and seagrass in Australia: a review. J Appl Phycol 9:311–326.
Kirschbaum, M.U.F., 2004. Soil respiration under prolonged soil warming: are rate reductions
caused by acclimation or substrate loss? Global Change Biology 10, 1870-1877.
Knorr, W., Prentice, I.C., House, J.I., Holland, E.A., 2005. On the available evidence for the
temperature dependence of soil organic carbon. Biogeosciences Discussions 2, 749-755.
Kowalenko, C.G., Cameron, D.R., 1976. Nitrogen transformations in an incubated soil as affected
by combinations of moisture content and temperature and sorption-fixation of ammonium.
Canadian Journal of Soil Science 56, 63-70.
Leirós, M.C., Trasar-Cepeda, C., Seoane, S., Gil-Sotres, F., 1999. Dependence of mineralization of
soil organic matter on temperature and moisture. Soil Biology and Biochemistry 31, 327-335.
Linn, D.M., Doran, J.W., 1984. Effect of water-filled pore space on carbon dioxide and nitrous
oxide production in tilled and nontilled soils. Soil Science Society of America Journal 48,
1667-1672.
Lomander, A., Kätterer, T., Andrén, O., 1998. Carbon dioxide evolution from top- and subsoil as
affected by moisture and constant and fluctuating temperature. Soil Biology and Biochemistry
30, 2017-2022.
Lowry, O. H., & Rosenbrough, N. J. (1951). Protein measurement with the folin phenol reagent.
Journal of Biology and Chemistry 193, 265–275.

111
Markwell, M. A. K., Haas, S. M., Bieber, L. L., & Tolbert, M. E. (1978). A modification of the Lowry
procedure to simplify protein determination in membrane and lipoprotein samples. Annals of
Biochemistry 87, 206–210.
McClain, M.E., Boyer, E.W., Dent, C.L., Gergel, S.E., Grimm, N.B., Groffman, P.M., Hart, S.C.,
Harvey, J.W., Johnston, C.A., Mayorga, E., McDowell, W.H., Pinay, G., 2003. Biogeochemical
hot spots and hot moments at the interface of terrestrial and aquatic ecosystems. Ecosystems
6, 301–312.
Miranda, R., Alavi, G., Aliaga, S., & Caballero, A. (2013). Mineralización del nitrógeno y generación
de CO2 por descomposición del estiércol ovino. Altiplano central, Bolivia. Venesuelos, 21(2).
Moorhead, D. L., Sinsabaugh, R. L., Linkins, A. E. y Reynolds, J. F., 1996. Decomposition
processes: modelling approaches and applications. The Science of the Total Environment 183:
137-149.Waksman, S.A. y Tenney, F.G. 1928. Composition of natural organic materials and
their decomposition in the soil: III. The influence of nature of plant upon the rapidity of its
decomposition. Soil Science 26:155-171.
Moreira, F. M. y J. O. Siqueira. 2006. Microbiologia y Bioquímica do solo. 2da. Edic. Editorial
Lavras, Universidade Federal de Lavras. 626 p.
Newell, R. C. & J. G. Field, 1983. The contribution of bacteria and detritus to carbon and nitrogen
flow in a benthic community.Mar. Biol. Lett. 4: 23–36.
Nicorladot, B., Fauvet, G., Cheneby, D., 1994. Carbon and nitrogen cycling through soil microbial
biomass at various temperatures. Soil Biology and Biochemistry 26, 253- 261.
Olabarria C., Lastra M. & Garrido J. (2007) Succession of macrofauna on macroalgal wrack of an
exposed sandy beach: effects of patch size and site. Marine Environmental Research, 63, 19-
40.
Pearson, T. H., & Rosenberg, R. (1978). Macrobenthic succession in relation to organic enrichment
and pollution of the marine environment. Oceanogr. Mar. Biol. Ann. Rev, 16, 229-311.
Pöhhacker, R., Zech, W., 1995. Influence of temperature on CO2 evolution, microbial biomass C
and metabolic quotient during decomposition of two humic forest horizons. Biology and
Fertility of Soils 19, 239-245.
Polis GA, Hurd SD. 1996. Linking marine and terrestrial food webs: allochthonous input from the
ocean supports high secondary productivity on small islands and coastal land communities.
Am Nat 147:396– 423.

112
Poore A.G.B. & Hill N.A. (2006) Sources of variation in herbivore preference: among-individual
and past diet effects on amphipod host choice. Marine Biology, 149, 1403-1410
Pusceddu, A., Dell’Anno, A., Fabiano, M., & Danovaro, R. (2009). Quantity and bioavailability of
sediment organic matter as signatures of benthic trophic status. Marine Ecology Progress
Series, 375, 41-52.
Pusceddu A, Bianchelli S, Gambi C, Danovaro R (2011) Assessment of benthic trophic status of
marine coastal ecosystems: Significance of meiofaunal rare taxa; Estuarine, Coastal and Shelf
Science. 93:420-430.
Rey, M., 2000. Desarrollo de una metodología analítica para la determinación de gases
invernadero. Aplicación al estudio de la influencia del cambio climático global sobre la
mineralización de la materia orgánica del suelo, Tesis Doctoral, Universidad de Santiago de
Compostela, 251 pp.
Rey, A., Jarvis, P., 2006. Modelling the effect of temperature on carbon mineralization rates across
a network of European forest sites (FORCAST). Global Change Biology 12, 1894-1908.
Rossi F. & Underwood A. (2002) Small-scale disturbance and increased nutrients as influences on
intertidal macrobenthic assemblages: experimental burial of wrack in different intertidal
environments. Marine Ecology Progress Series, 242, 29-39.
Sinsabaugh, R.L. y Moorhead, D.L. 1997. Synthesis of litter quality and enzyme approaches to
decomposition modelling. En Driven by Nature: Plant litter quality and Decomposition.
Cadish, G y K.E. Giller (Eds)., CAB International. Wallinford, Oxon,UK.
Sinsabaugh, R.L, Carreiro, M.M. y Álvarez, S. 2002. Enzyme and Microbial Dynamics of Litter
Decomposition (Chapter 9). Enzymes in the Environment. Burns, R.G, R.P. Dick. (Eds.),
Marcel Dekker, New York. pp 249-266.
Skujins, J.J., 1967. Enzymes in soil. En: McLaren, A.D., Peterson, G.H. (Eds.) Soil Biochemistry,
vol 1, Marcel Dekker, New York, pp 371-414.
Smith V H, (2003) Eutrophication of freshwater and coastal marine ecosystems. A global problem;
Environmental Science and Pollution Research.10:126-139.
Tenore KR, Hanson RB (1980) Availability of detritus of different types and ages to apolychaete
macroconsumer, Capitella capitata. Limnol Oceanogr 25:553-558.
Tisdale, S. L., W. L. Nelson y J. D. Beaton. 1985. Soil fertility and fertilizers. 4ta ed. Macmillan:
Publishing Company; p. 754.

113
Tremblay L, Benner R (2006) Microbial contributions to N-immobilization and organic matter
preservation in decaying plant detritus; Geochimica et Cosmochimica Acta. 70:133-146.
Van Alstyne K.L., McCarthy III J.J., Hustead C.L. & Duggins D.O. (1999) Geographic variation in
polyphenolic levels of Northeastern Pacific kelps and rockweeds. Marine Biology, 133, 371-379
Venturini N, Pita A, Brugnoli E, García-Rodriguez F, Burone L, Kandratavicius N, Hutton M,
Muniz P M (2012) Benthic trophic status of sediments in a metropolitan area (Rio de la Plata
estuary): Linkages with natural and human pressures; Estuarine, Coastal and Shelf Science.
112:139-152.
Visser S., Parkinson D. (1992): Soil biological criteria as indicators of soil quality: soil
microorganisms. American Journal of Alternative Agriculture, 7: 33–37.
Wardle, D.A., Lavelle, P., 1997. Linkages between soil biota, plant litter quality and decomposition.
En: Cadish, G., Giller, K.E. (Eds.) Driven by Nature, Plant litter.
Williams, P. M. & A. F. Carlucci, 1976. Bacterial utilisation of organic matter in the deep-sea.
Nature 262: 810–811.
Williams, S (1984) Decomposition of the tropical macroalga Caulerpa cupressoides (West) C.
Agardh: Field and laboratory studies. J Exp Mar Biol Ecol 80:109-124.
Williams PB, del Giorgio PA (2005) Respiration in aquatic systems: history and background. The
global significance of respiration in aquatic systems: from single cells to the biosphere. En:
del Giorgio PA, Williams PjleB (eds). Respiration in aquatic ecosystems. Oxford University
Press, Oxford, UK, pp 1–17.
Zöllner, N., & Kirsch, K. (1962). Über die quantitative Bestimmung von Lipoiden (Mikromethode)
mittels der vielen natürlichen Lipoiden (allen bekannten Plasmalipoiden) gemeinsamen
Sulfophosphovanillin-Reaktion. Research in Experimental Medicine, 135(6), 545-561.

114

115
INTRODUCCIÓN
En la playa al combinarse los ecosistemas marinos y terrestres, cada uno aporta los
factores ―clave‖ para que se den tasas de descomposición superiores a otros ecosistemas muy
productivos como las selvas tropicales (Coupland et al., 2007). Estos factores ―clave‖ aportados
por el mar son la materia orgánica y la humedad contenida en el interior de los parches de algas.
La tierra, a su vez, aporta el oxígeno necesario para que los organismos descomponedores puedan
realizar sus actividades. Por este motivo, Pearse et al. (1942) definió a las playas como grandes
digestores de materia orgánica ya que se dan las condiciones necesarias para que la
descomposición transcurra a su máxima capacidad.
La materia orgánica vegetal que llega a las playas lo hace en forma de macroalgas y/o
fanerógamas. El tamaño de estos arribazones puede oscilar desde pequeños fragmentos hasta la
macroalga completa. Este material puede procede de las praderas de fanerógamas del submareal
y/o intermareal o de costas rocosas. Cuando se deposita en la playa, actúa como lugar de
regeneración de nutrientes a través de su descomposición (Ochieng & Erftemeijer, 1999). También
puede tener un papel estructural proporcionando un hábitat o puede ser utilizado como alimento
para los herbívoros de la meio y macrofauna. Aunque su papel más importante en estos
ecosistemas tan dinámicos y con tan baja producción primaria, es sin duda la de proveer a las
playas de nutrientes inorgánicos y energía para los autótrofos. De esta manera, se construye una
red alimentaria basada y dependiente de la presencia de macroalgas (McLachlan, 1981; Alongi,
1998).
Durante el tiempo que esté la macroalga en la playa, pasará por varias etapas que la irán
reduciendo (―envejeciendo‖) hasta hacerla desaparecer. Este proceso de ―envejecimiento‖ no es
otro que el de la descomposión. Consta de 3 etapas principales: desintegración saprofítica,
fragmentación y lixiviación (Robertson & Mann, 1980; Harrison, 1982; Jedrzejczak, 2002a). La
primera etapa es la descomposición saprofítica. Se caracteriza por un rápido aumento de la
actividad microbiana mientras consumen los componentes fácilmente asimilables (Robertson &
Hansen, 1982; Harrison, 1982; Rieper-Kirchner, 1990). La siguiente etapa es la fragmentación
mecánica y biológica (Robertson & Mann, 1980; Alongi, 1998). La macrofauna en esta estapa tiene
un papel de suma importancia al triturar el alga. En este proceso la va reduciendo hasta pequeños
fragmentos que serán fácilmente atacados por los microorganismos al aumentar la superficie de
contacto (Harrison, 1982). Por último, la lixiviación es el proceso abiótico por el cual la materia
soluble del alga es eliminada por el lavado del agua. En este proceso se produce una pérdida de
peso del alga importante y cambios en su composición química (Ochieng & Erftemeijer, 1999). La
velocidad a la que ocurren estos procesos depende del ambiente físico y biótico local (Alongi,
1998), de la composición química del alga (Jenrzejczak, 2002b) y de su ―edad‖. Este factor es clave

116
porque la capacidad de reacción de los materiales depositados con el paso del tiempo va
disminuyendo.
El objetivo de este capítulo es proporcionar una visión espacial de las variaciones en la
actividad metabólica de los distintos tipos de alga. Para ello es necesario conocer cómo varían las
cantidades de los productos de la descomposición (CO2, nitrógeno inorgánico, fósforo y
carbonobiopolimérico) dependiendo de la ―edad‖ del alga y de su biomasa. Como se comentó
anteriormente, la macrofauna tiene un papel muy importante en las tasas de descomposición del
alga pero dependiendo del estado en el que se encuentre ésta, el número de individuos variará. Así
pues, también es necesario conocer la distribución y abundancia de la macrofauna en la playa en
función de la ―edad‖ de las algas varadas. Para cumplir el objetivo de este trabajo se plantearon las
siguientes HIPÓTESIS: (1) Un ―envejecimiento‖ del alga provocará un desdenso de la actividad
metabólica, (2) el aumento de la biomasa algal podrá compensar este descenso de actividad y (3)
habrá mayor abundancia de macrofauna en los parches de algas recientemente depositdos.
MATERIAL Y MÉTODOS
Área de estudio
Las ocho playas elegidas para este estudio se localizan en las Rías Baixas de Galicia (NO
España). Su elección se basó por ser playas morfodinámicamente parecidas. La mayoría están
situadas en ensenadas, están influenciadas por un régimen meso-macromareal con un rango de
3.5 a 4 m en mareas vivas haciendo que el tipo de arena oscile entre arena media a gruesa. Ninguna
es una playa urbana pero la mayoría suelen ser playas muy frecuentadas en la época estival. En la
Figura 1 se muestran las imágenes de satélite de cada una y su localización en la costa Gallega. Se
puede apreciar que todas tienen la misma forma de media luna. Las características más específicas
de cada una se detallan en la tabla 1.

117
Figura 1. Se muestra una imagen de satélite en planta para cada playa con sus coordenadas y a
continuación se sitúan en el mapa de Galicia.
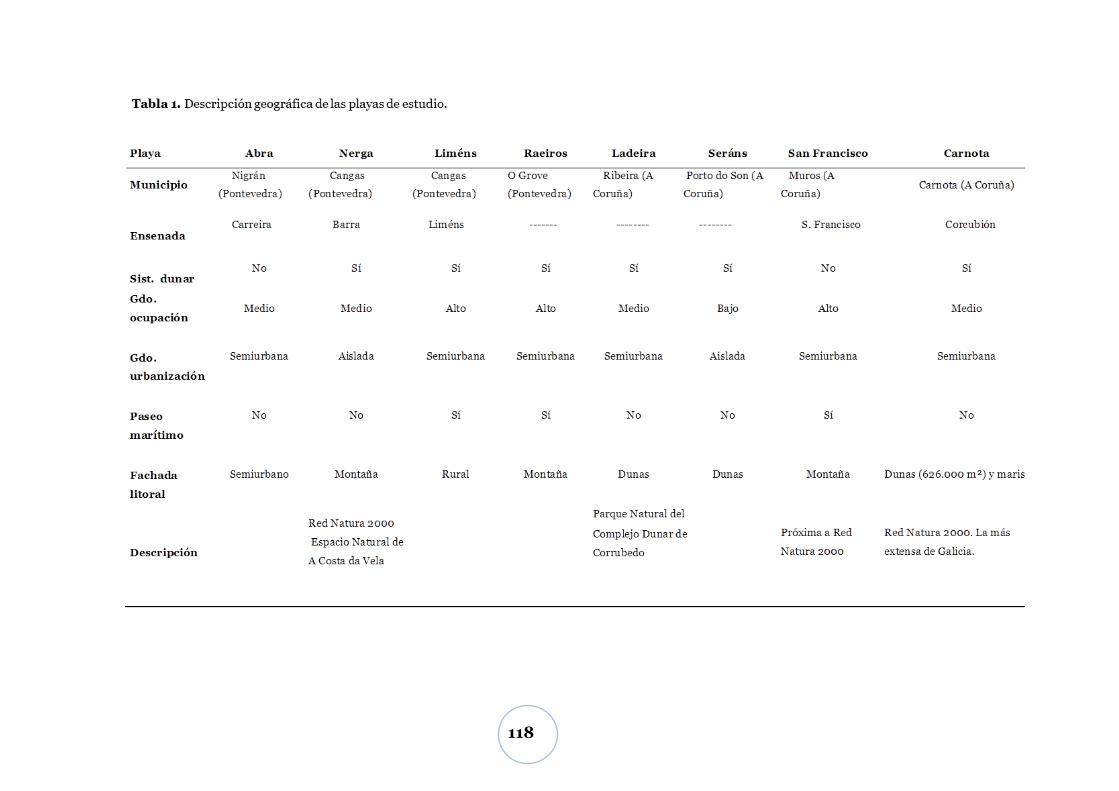
118

119
Diseño del muestreo
El diseño de este muestreo se basó en los resultados obtenidos en la parte I del presente
capítulo. A partir de éstos se pudo conocer la franja horaria con mayor actividad metabólica. Esto
permitió diseñar este muestreo eliminando la variabilidad debida a la temperatura pudiéndose
observar el efecto de la biomasa algal en función de su ―edad‖, es decir, del tiempo que lleva varada
en la playa o de su estado degradativo. Basándose en esto, se clasificaron las algas en tres tipos: las
más recientes en la playa son las frescas y las que llevan mucho tiempo son las secas. Las que llevan
varadas en la playa durante un tiempo intermedio entre unas y otras, son las húmedas.
La zona de muestreo escogida en todas las playas fue la misma. La zona de estudio fue la
franja paralela a la costa comprendida entre el nivel superior e inferior del drift line. En esta zona se
escogieron 20 puntos de muestro al azar siendo cinco de ellos los puntos control (sin parches de
algas) (Figura 2). Cada uno de ellos se clasificó en parche fresco, húmedo o seco dependiendo de su
distancia a la línea de costa. Esta clasificación permite estimar la ―edad‖ de cada parche ya que esta
característica determinará la emisión de CO2 y la liberación de nutrientes procedente de la
mineralización de la materia orgánica. Como es de esperar, cada tipo de alga en función de su ―edad‖
liberará diferentes cantidades de cada producto. También el tipo de alga repercutirá en la
abundancia de la macrofauna que se encuentre en el parche debido a su calidad nutritiva.
Figura 2. Esquema general de la zona de estudio. Se muestran los 15 puntos con
parches de algas y los 5 controles (C).

120
Toma de las muestras
El muestreo se llevó a cabo durante un mes (desde el 9 de septiembre al 9 de octubre de
2013). El CO2 liberado de cada punto de estudio, se cuantificó con un medidor de flujo en continuo
con un detector LI-COR de CO2 (µmol C m-2 s-1) (volumen de la campana de acumulación de 2.756
10-3 m3 y un área en su base de 3.140 10-2 m-2). Las temperaturas del aire y del sedimento, se
midieron con termómetro de campo; para el sedimento, el termómetro se introdujo 5 cm en el
sustrato bajo el parche de algas. Una vez se tomó la medida, las algas del parche se metieron en
bolsas de plástico para ser transportadas al laboratorio. En cada uno de los puntos se recogieron
muestras del sedimento en botes de plástico estériles para el análisis de nutrientes en forma de
nitrógeno inorgánico (TNI= NH4+ + NO2
- + NO3- µM g-1 sed.), fósforo en forma de fosfatos (PO4
-3 µM
g-1 sed.), carbono biopolimérico (CBP) (µg g-1 sed.), el cálculo la relación proteína-carbohidrato
(PRT:CHO) y el porcentaje de humedad del sedimento (% HS). Las muestras de macrofauna se
recogieron con un corer de 10 cm de diámetro hasta una profundidad del sedimento de 10 cm (área=
0.16 m-2) de forma aleatoria alrededor del punto de muestreo (n=3). Las muestras fueron tamizadas
in situ a través de una malla de 1 mm y se guardaron en bolsas plásticas. Una vez en el laboratorio, a
cada una se le añadió una solución de agua de mar y formol al 4%. Los individuos se determinaron al
nivel taxonómico más bajo posible. Una vez en el laboratorio, todas las muestras se congelaron y
almacenaron a -20ºC hasta su posterior análisis.
Análisis de las muestras
Se descongelaron las algas y se separaron del resto de material varado en una bandeja con
agua. Se secaron en la estufa a 60º C durante 48 horas para obtener la relación Peso fresco: Peso
seco. De esta relación se obtuvo el porcentaje de humedad del alga. A continuación se molieron y se
tomó una alícuota de 5g de peso seco que se calcinó en la mufla durante 4h a 500ºC. De esta forma
se obtuvo la biomasa algal (g peso seco libre de cenizas). El porcentaje de humedad del sedimento se
obtuvo a partir de la pérdida de peso tras el secado de las muestras durante 72 horas a 60ºC. El
nitrógeno inorgánico total (TNI= NH4+ + NO2
- + NO3-) y los fosfatos (PO4
-3) se extrajeron de 5 g de
sedimento según la normativa española (UNE 77306, 1999). Una vez obtenida, se congelaron las
muestras en tubos falcon. Posteriormente fueron llevadas a la ―Unidad de Análisis Elemental‖ de la
Universidad de Vigo para su análisis. Las muestras se analizaron con un Autoanalizador de Flujo
continuo Segmentado. La determinación se realiza de forma directa frente a patrones acuosos.
121
Análisis estadístico
Las playas estudiadas fueron seleccionadas por su similar morfodinámica, por ello, en la
mayoría de los análisis, se utilizan como réplicas (n=8). El CO2, los nutrientes (NH4+, NO3
- y PO4-3) y
el CBP fueron evaluados mediante ANOVA. Las réplicas de cada tipo fueron desiguales por lo que se
utilizó Kruskal-Wallis, análisis de varianza de una vía en rangos paquete estadístico R 3.1.1. El
modelo para la variable se construyó con ―playa-tipo alga‖ (n= 32); Previamente los datos fueron
transformados para cumplir las asunciones de normalidad y homogeneidad de varianzas, evaluadas
con los test de Sapiro-Wilk y Cochran respectivamente.
Para estudiar la relación entre las variables (CO2, NH4+, NO3
- y PO4-3) y los factores físicos y
biológicos (temperatura, humedad del sedimento, tamaño de grano, humedad y biomasa algal y
abundancia de la macrofauna) se realizaron regresiones con paquete estadístico R 3.1.1.
RESULTADOS
Caracterización de las playas
Cada playa fue caracterizada de la misma forma que la descrita en el capítulo 2. Todas las
playas están formadas por arena (media o gruesa). La playa con el tamaño de grano más pequeño
fue San Francisco, formada por arena media y la playa con mayor tamaño de grano fue Raeiros con
la arena gruesa (Tabla 2). La pendiente se expresa como su inversa (1/pte), de este modo, las playas
más extensas y con pendientes más suaves fueron Carnota y Ladeira (29.69 y 17.44 respectivamente)
y la más pequeña y con una pendiente muy pronunciada fue Seráns (5.85). En la figura 3 se
muestran los perfiles de cada playa donde se pueden comparar las pendientes.
Tabla 2. Características físicas y morfodinámicas de las playas. Ordenadas de menor a mayor pendiente.
Playa Long. (m) Ancho(m) Área (m²) 1/pte T.M.G
(mm)
Ladeira 5000 128 6.4*105 29.69 0.47
Carnota 6500 500 3.25*106 17.44 0.53
Nerga 750 30 22500 16.49 0.49
Raeiros 670 70 46900 9.38 0.95
Abra 210 68 14280 9.36 0.46
San Francisco 760 35 26600 8.45 0.33
Liméns 310 25 7750 8.44 0.79
Seráns 200 30 6000 5.85 0.59
Longitud (Long), inversa de la pendiente (1/pte), tamaño medio de grano (T.M.G)
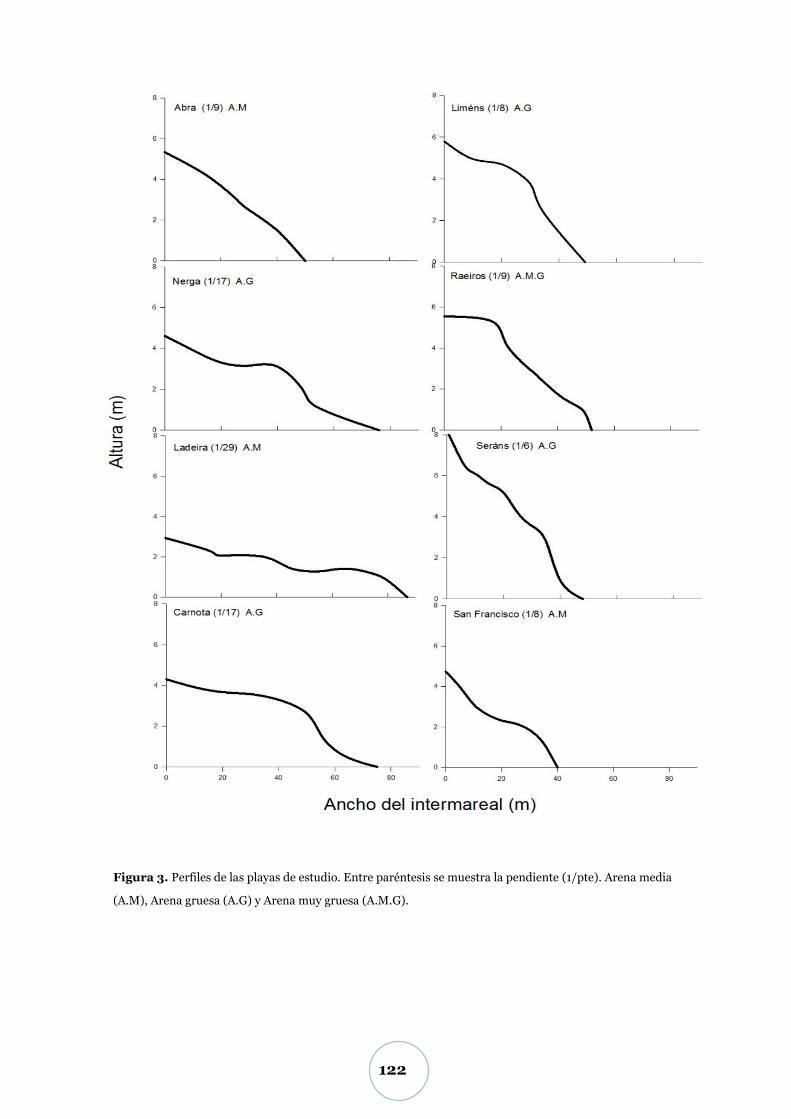
122
Figura 3. Perfiles de las playas de estudio. Entre paréntesis se muestra la pendiente (1/pte). Arena media
(A.M), Arena gruesa (A.G) y Arena muy gruesa (A.M.G).

123
Parámetros físico-ambientales y biológicos
Como muestra la tabla 3, el rango de temperatura entre las ocho playas fue pequeño. La
mayor diferencia de temperatura fue de 4.5ºC; entre Raeiros (mayor temperatura) y Nerga (la
temperatura más baja). Raeiros, como consecuencia de las altas temperaturas, registró el porcentaje
de humedad más bajo (1.69 ± 0.14%) aunque el máximo de humedad no correspondió con Nerga, si
no con Carnota (6.17 ± 0.83%).
El porcentaje cubierto por material de arribazón osciló entre 0.004% en Carnota y 0.21% en
Abra (Tabla 4). La biomasa algal más alta se detectó en Seráns, acompañada de las abundancias de
macrofauna más altas y la biomasa algal más baja se localizó en San Francisco pero fue en Carnota
donde se encontró el menor número de individuos de la macrofauna.
Tabla 3. Parámetros físico-ambientales
Playa T (°C) % HS
Abra 20.38 ± 0.17 1.77 ± 0.33
Carnota 22.08 ± 0.15 6.17 ± 0.83
Ladeira 21.075 ± 0.20 5.09 ± 0.46
Liméns 24.33 ± 0.08 2.2 ± 0.22
Nerga 19.90 ± 0.20 3.56 ± 0.64
Raeiros 24.45 ± 0.13 1.69 ± 0.14
San Francisco 20.53 ± 0.25 5.57 ± 0.33
Seráns 22.35 ± 0.17 2.26 ± 0.43
Temperatura (T), % de humedad del sedimento (% HS). Media ± SE.
La clasificación de las algas de los puntos de estudio se basó en su estado de degradación
que a su vez, depende del porcentaje de humedad del alga. Cada tipo de alga se localizó en su
correspondiente línea de varamiento (drift line). Cada uno de ellos registró un porcentaje de
humedad algal proporcional al tiempo de depósito. Las algas que llevaban más tiempo varadas en la
playa (secas) tuvieron los porcentajes más bajos de humedad que las más recientemente varadas
(frescas) difiriendo en un 12% (Tabla 5). En cambio, la diferencia entre las algas secas y húmedas fue
baja (siendo mayor el de las húmedas). La media de la biomasa algal recogida en cada parche de
algas varió entre un 7 (frescas) y un 32% (Húmedas) aproximadamente.
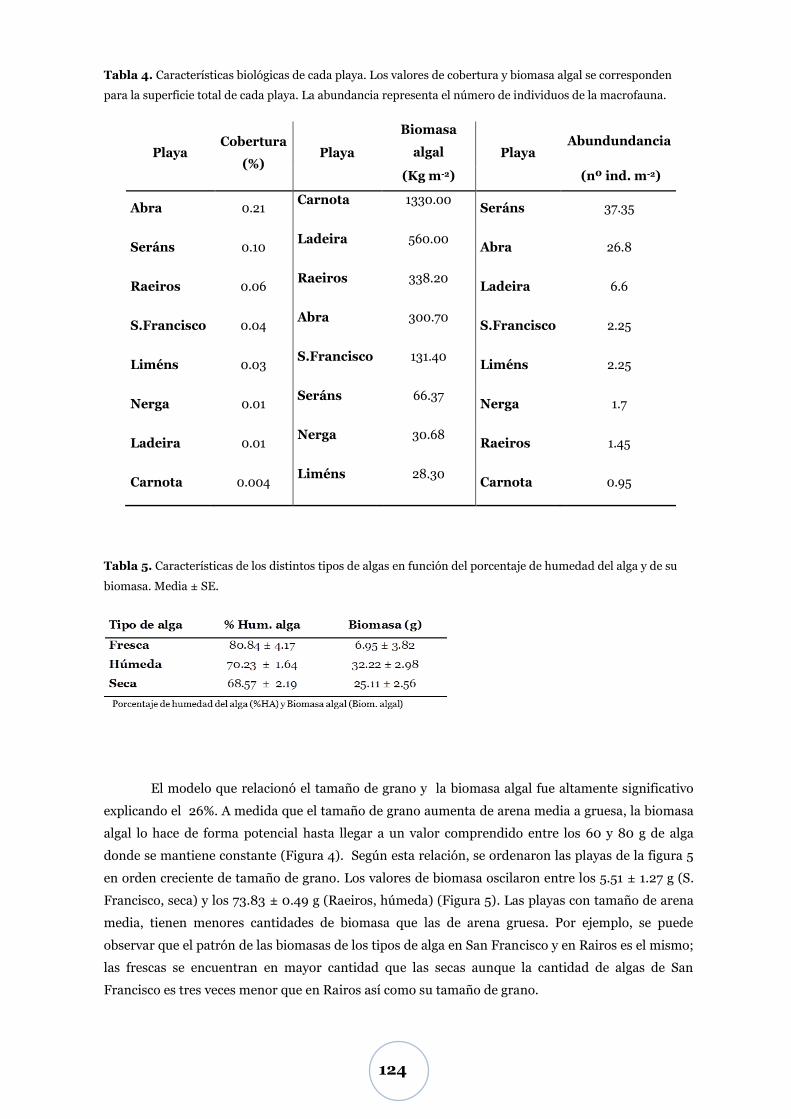
124
Tabla 4. Características biológicas de cada playa. Los valores de cobertura y biomasa algal se corresponden
para la superficie total de cada playa. La abundancia representa el número de individuos de la macrofauna.
Tabla 5. Características de los distintos tipos de algas en función del porcentaje de humedad del alga y de su
biomasa. Media ± SE.
El modelo que relacionó el tamaño de grano y la biomasa algal fue altamente significativo
explicando el 26%. A medida que el tamaño de grano aumenta de arena media a gruesa, la biomasa
algal lo hace de forma potencial hasta llegar a un valor comprendido entre los 60 y 80 g de alga
donde se mantiene constante (Figura 4). Según esta relación, se ordenaron las playas de la figura 5
en orden creciente de tamaño de grano. Los valores de biomasa oscilaron entre los 5.51 ± 1.27 g (S.
Francisco, seca) y los 73.83 ± 0.49 g (Raeiros, húmeda) (Figura 5). Las playas con tamaño de arena
media, tienen menores cantidades de biomasa que las de arena gruesa. Por ejemplo, se puede
observar que el patrón de las biomasas de los tipos de alga en San Francisco y en Rairos es el mismo;
las frescas se encuentran en mayor cantidad que las secas aunque la cantidad de algas de San
Francisco es tres veces menor que en Rairos así como su tamaño de grano.
Playa Cobertura
(%) Playa
Biomasa
algal Playa Abundundancia
(Kg m-2) (nº ind. m-2)
Abra 0.21 Carnota 1330.00
Seráns 37.35
Seráns 0.10 Ladeira 560.00
Abra 26.8
Raeiros 0.06 Raeiros 338.20
Ladeira 6.6
S.Francisco 0.04 Abra 300.70
S.Francisco 2.25
Liméns 0.03 S.Francisco 131.40
Liméns 2.25
Nerga 0.01 Seráns 66.37
Nerga 1.7
Ladeira 0.01 Nerga 30.68
Raeiros 1.45
Carnota 0.004 Liméns 28.30
Carnota 0.95
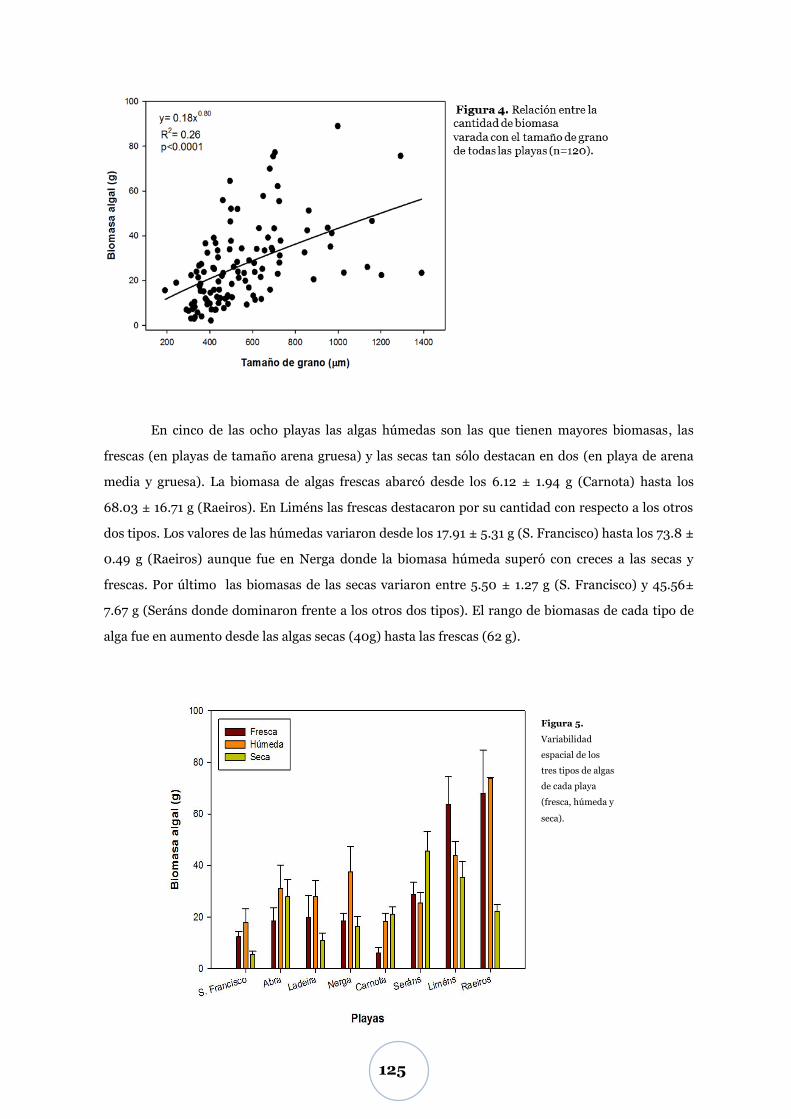
125
Figura 5.
Variabilidad
espacial de los
tres tipos de algas
de cada playa
(fresca, húmeda y
seca).
En cinco de las ocho playas las algas húmedas son las que tienen mayores biomasas, las
frescas (en playas de tamaño arena gruesa) y las secas tan sólo destacan en dos (en playa de arena
media y gruesa). La biomasa de algas frescas abarcó desde los 6.12 ± 1.94 g (Carnota) hasta los
68.03 ± 16.71 g (Raeiros). En Liméns las frescas destacaron por su cantidad con respecto a los otros
dos tipos. Los valores de las húmedas variaron desde los 17.91 ± 5.31 g (S. Francisco) hasta los 73.8 ±
0.49 g (Raeiros) aunque fue en Nerga donde la biomasa húmeda superó con creces a las secas y
frescas. Por último las biomasas de las secas variaron entre 5.50 ± 1.27 g (S. Francisco) y 45.56±
7.67 g (Seráns donde dominaron frente a los otros dos tipos). El rango de biomasas de cada tipo de
alga fue en aumento desde las algas secas (40g) hasta las frescas (62 g).

126
Macrofauna
Como se vió en el apartado anterior, Seráns y Abra fueron las playas con mayor abundancia
de macrofauna (32.08 ± 5.28 ind. m-2). El resto de playas no superaron los 7 ind.m-2, siendo
Carnota la menos abundante (0.95 ind. m-2). La tabla 5 refleja que los anfípodos son el grupo más
abundante en Seráns y Abra (490.5 ± 52.5 nº ind. m-2) seguido por los dípteros también encontrados
en Seráns (188 ind. m-2). Abra también destacó por tener el número más alto de isópodos (97 ind. m-
2). Cabe destacar que el único grupo que se encontró en todas las playas con una media de 39 ± 8.31
ind. m-2 (sin tener en cuenta sus máximos en Seráns y en Abra) fueron los anfípodos. En general el
resto de playas no albergaron a penas otros grupos.
Tabla 5. Abundancia de cada grupo taxonómico en cada playa.
Playa Anfípodos Isópodos Coleópteros Dípteros
Abra 438 97 0 1
Carnota 15 12 4 0
Ladeira 75 5 52 0
Liméns 41 0 0 3
Nerga 33 0 1 1
Raeiros 27 0 0 2
San Francisco 43 0 1 1
Seráns 543 0 13 188
Los valores destacados en negro se corresponden con los más altos de cada grupo.
Flujo de CO2
Efecto de la biomasa algal y el tipo alga
Los valores medios del flujo de CO2 emitidos por cada tipo de alga y por los controles en
todas las playas se muestran en la tabla 6. El máximo corresponde a las algas frescas y el mínimo a
los controles. El valor máximo de CO2 se midió en un parche de alga frescas de Liméns (10.58
µmoles de C m-2 s-1) y el mínimo en un control de Raeiros (0.05 µmoles de C m-2 s-1). Se observó una
variación espacial en cada playa (ANOVA en rangos, Kruskal-Wallis; p< 0.001) entre los controles y
las algas frescas. Se detectaron diferencias significativas entre las algas frescas y húmedas de Liméns
con los controles del resto de las playas (6.55 ± 1.18, 3.88 ± 0.70 y 0.22 ± 0.02 µmol C m-2 s-1
respectivamente). También las algas frescas de Raeiros (5.24 ± 1.70 µmol C m-2 s-1), Abra (3.92 ±

127
0.48 µmol C m-2 s-1) y Nerga (3.27 ± 0.42 µmol C m-2 s-1) mostraron diferencias significativas con los
controles de Carnota, Liméns, Nerga y Raeiros (0.14 ± 0.01 µmol C m-2 s-1).
El modelo explica la influencia de la cantidad de material varado en la playa sobre las
emisiones de CO2 en un 57% (figura 6). Este efecto fue exponencial positivo y altamente significativo,
lo cual indica que con el aumento de la biomasa, la cantidad de CO2 no aumenta de forma
proporcional. En la figura 6 se representa esta relación diferenciándola por playas. Así, se puede ver
cómo la mayor parte de las playas se localizan entre valores de biomasa que no superan los 20g y los
4 µmol C m-2 s-1 de CO2. Nerga, Liméns y Raeiros destacaron por emitir las cantidades de CO2 más
altas en biomasas de algas mayores. Aunque el modelo se ajusta correctamente a dicha relación, se
tuvo en cuenta la variabilidad existente debido a los distintos tipos de algas (frescas, húmedas y
secas). Por lo tanto, se construyó para cada tipo de alga el mismo modelo (Figura 7).
Tabla 6. Cantidad media de CO2 para cada tipo de alga y controles
Tipo de alga CO2 (µmol C m-2 s-1)
Fresca 3.30 ± 0.33
Húmeda 2.31 ± 0.21
Seca 0.93 ± 0.08
Control (sin
alga) 0.21 ± 0.02
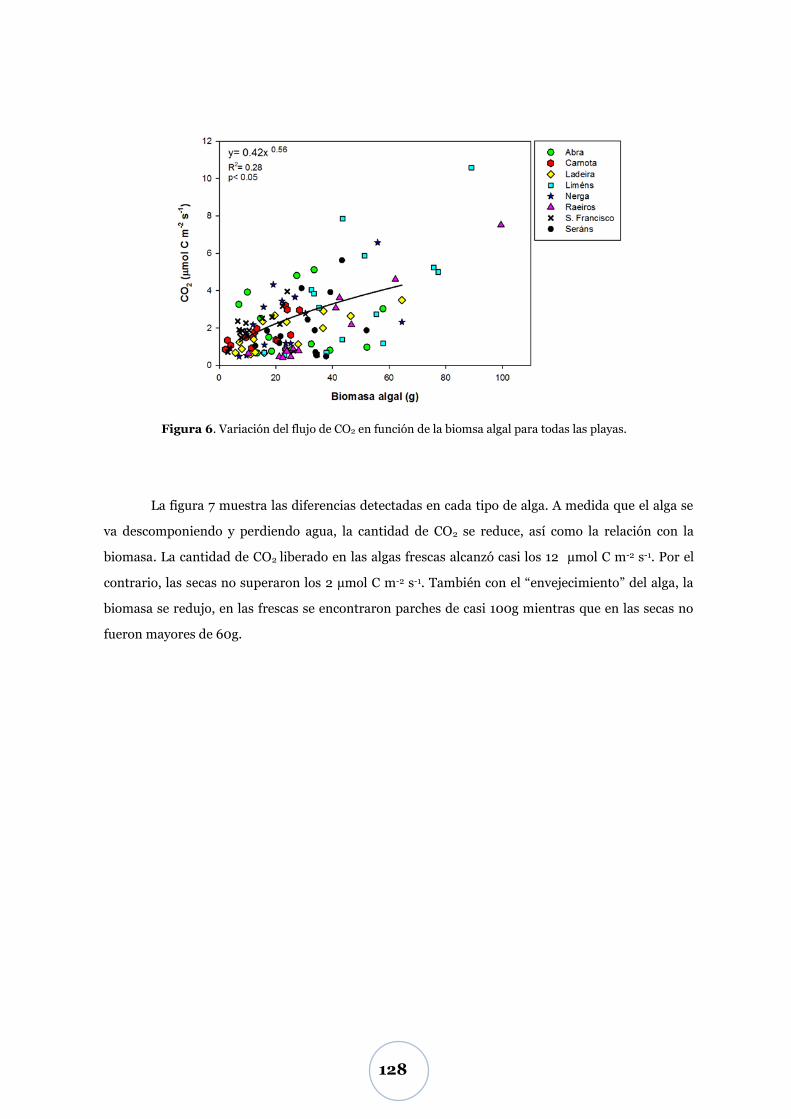
128
Figura 6. Variación del flujo de CO2 en función de la biomsa algal para todas las playas.
La figura 7 muestra las diferencias detectadas en cada tipo de alga. A medida que el alga se
va descomponiendo y perdiendo agua, la cantidad de CO2 se reduce, así como la relación con la
biomasa. La cantidad de CO2 liberado en las algas frescas alcanzó casi los 12 µmol C m-2 s-1. Por el
contrario, las secas no superaron los 2 µmol C m-2 s-1. También con el ―envejecimiento‖ del alga, la
biomasa se redujo, en las frescas se encontraron parches de casi 100g mientras que en las secas no
fueron mayores de 60g.
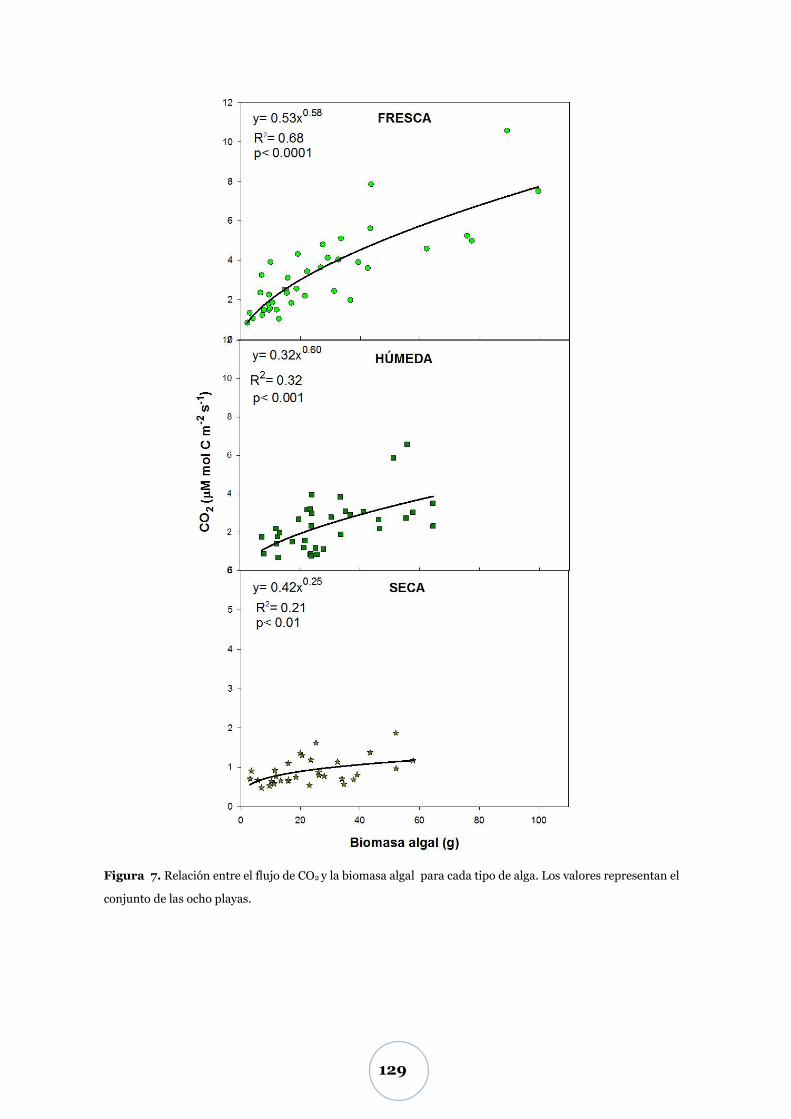
129
Figura 7. Relación entre el flujo de CO2 y la biomasa algal para cada tipo de alga. Los valores representan el
conjunto de las ocho playas.
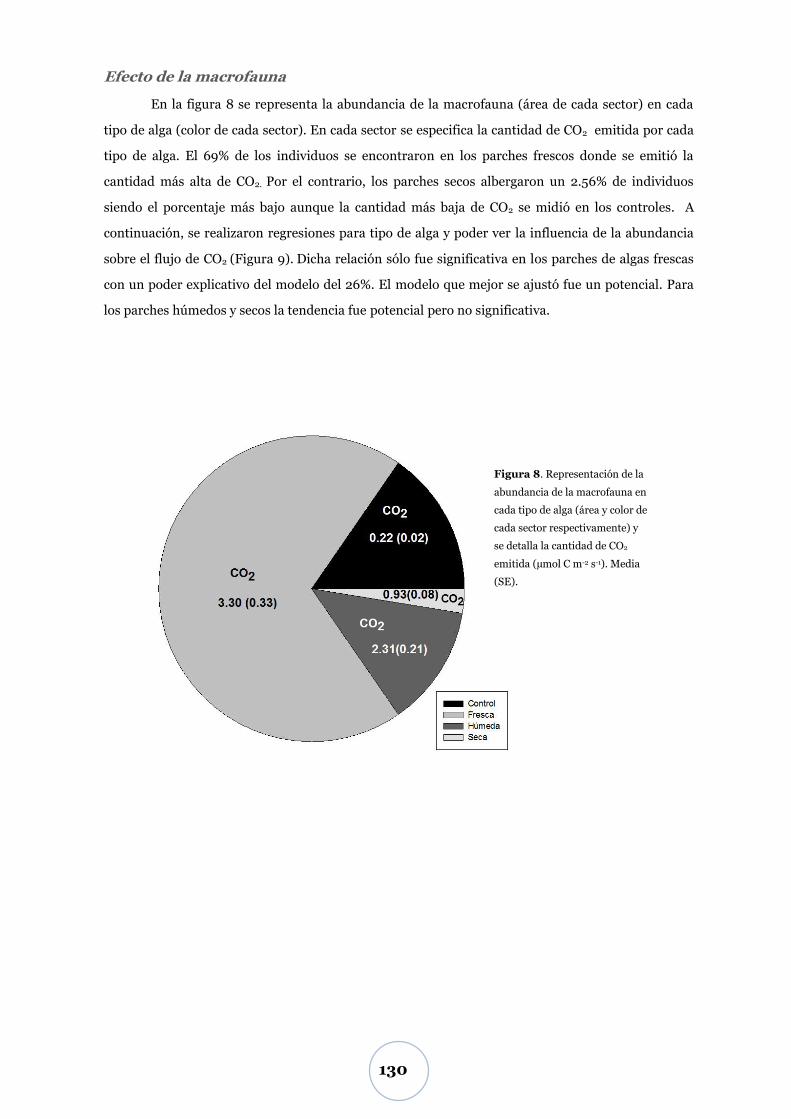
130
Efecto de la macrofauna
En la figura 8 se representa la abundancia de la macrofauna (área de cada sector) en cada
tipo de alga (color de cada sector). En cada sector se especifica la cantidad de CO2 emitida por cada
tipo de alga. El 69% de los individuos se encontraron en los parches frescos donde se emitió la
cantidad más alta de CO2. Por el contrario, los parches secos albergaron un 2.56% de individuos
siendo el porcentaje más bajo aunque la cantidad más baja de CO2 se midió en los controles. A
continuación, se realizaron regresiones para tipo de alga y poder ver la influencia de la abundancia
sobre el flujo de CO2 (Figura 9). Dicha relación sólo fue significativa en los parches de algas frescas
con un poder explicativo del modelo del 26%. El modelo que mejor se ajustó fue un potencial. Para
los parches húmedos y secos la tendencia fue potencial pero no significativa.
Figura 8. Representación de la
abundancia de la macrofauna en
cada tipo de alga (área y color de
cada sector respectivamente) y
se detalla la cantidad de CO2
emitida (µmol C m-2 s-1). Media
(SE).
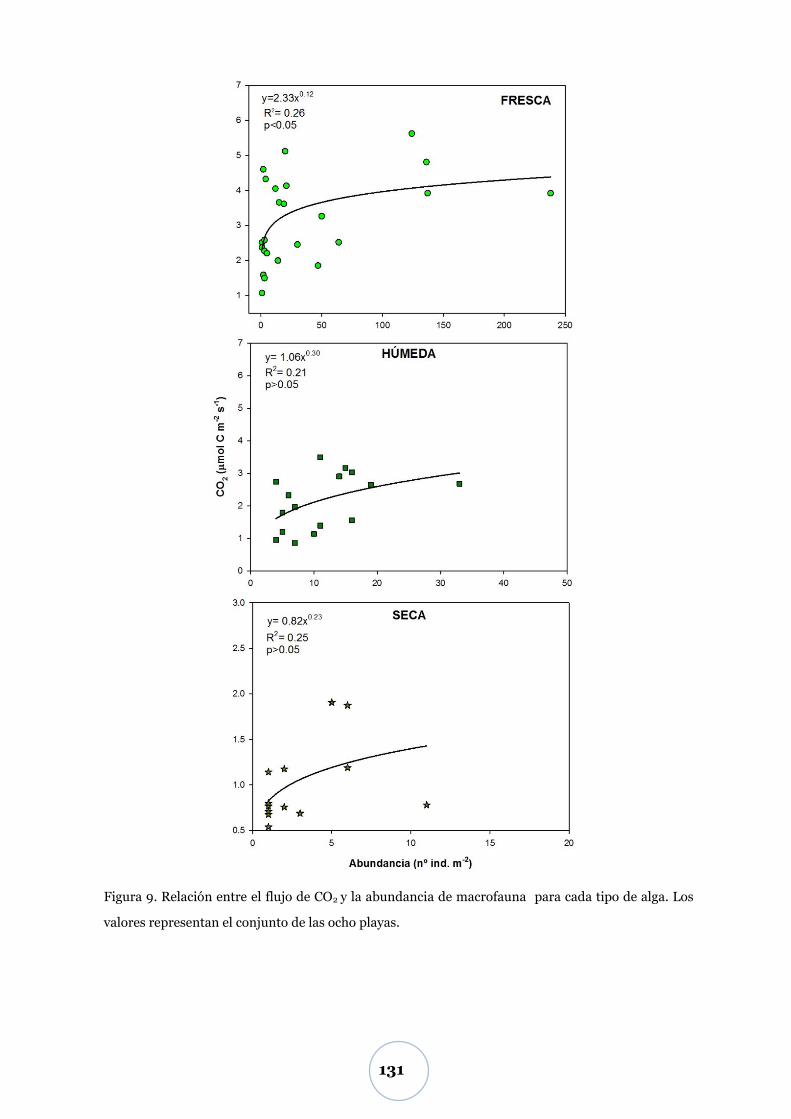
131
Figura 9. Relación entre el flujo de CO2 y la abundancia de macrofauna para cada tipo de alga. Los
valores representan el conjunto de las ocho playas.

132
En base a los resultados obtenidos, las relaciones de la biomasa algal, abundancia de
macofauna y el porcentaje de humedad del sedimento con el CO2 se pueden resumir mediante un
modelo de regresión múltiple. Las tres variables son altamente significativas y además tienen la
capacidad de explicar las variaciones del flujo de CO2 en un 45% (Tabla 7):
Tabla 7. Regresión múltiple del CO2 con la biomasa algal, la abundancia de la macrofauna y el porcentaje de
humedad del sedimento.
CO2 = -0.504 + (0.0550 * BIOM) + (0.0124 * ABUND) + (0.277 * HS)
R2 P P
0.45 <0.0001 Biomasa <0.001
Abundancia 0.001
%Hum.sed. <0.001
Biomasa algal (BIOM), abundancia de macrofauna (ABUND), porcentaje
de humedad en el sedimento (HS). P (coeficiente de significación) y R2
(coeficiente de correlación).
Liberación de nitrógeno inorgánico y fósforo
Efecto de la biomasa algal y tipo de alga
Las concentraciones de los nutrientes no mostraron variación espacial en las playas según el
tipo de alga (ANOVA en rangos, Kruskal-Wallis; p> 0.05). En todos los tipos de alga y control, la
concentración de amonio fue tres veces superior (en la mayoría de los tipos) a la de nitrito. Las algas
frescas fueron las que registraron las concentraciones más altas de amonio, nitrito y nitrato. No
ocurrió lo mismo para el fosfato que su valor más alto se detectó en las algas frescas. Por el
contrario, las algas secas fueron las que registraron las concentraciones más bajas aunque la de
fosfato se encontró en los controles. Al igual que se hizo para el CO2, se relacionaron las biomasas de
los parches de algas con la liberación de los nutrientes al sedimento (Tabla 8).
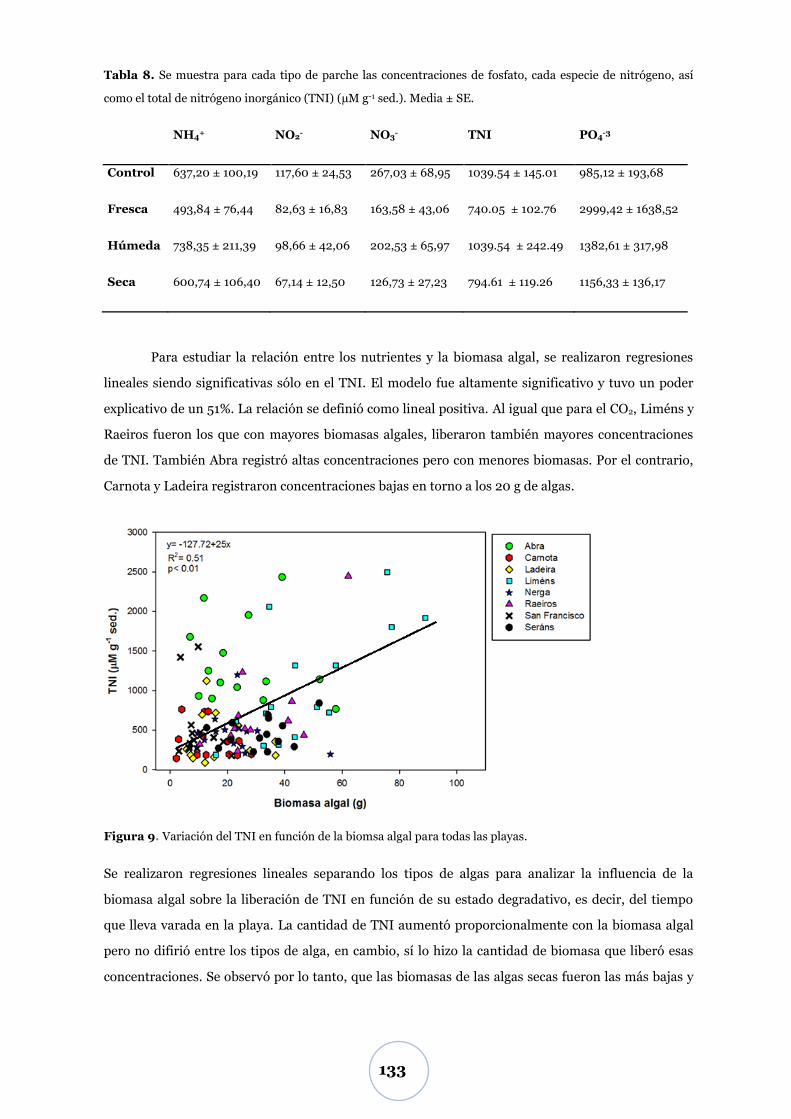
133
Tabla 8. Se muestra para cada tipo de parche las concentraciones de fosfato, cada especie de nitrógeno, así
como el total de nitrógeno inorgánico (TNI) (µM g-1 sed.). Media ± SE.
NH4+ NO2
- NO3- TNI PO4
-3
Control 637,20 ± 100,19 117,60 ± 24,53 267,03 ± 68,95 1039.54 ± 145.01 985,12 ± 193,68
Fresca 493,84 ± 76,44 82,63 ± 16,83 163,58 ± 43,06 740.05 ± 102.76 2999,42 ± 1638,52
Húmeda 738,35 ± 211,39 98,66 ± 42,06 202,53 ± 65,97 1039.54 ± 242.49 1382,61 ± 317,98
Seca 600,74 ± 106,40 67,14 ± 12,50 126,73 ± 27,23 794.61 ± 119.26 1156,33 ± 136,17
Para estudiar la relación entre los nutrientes y la biomasa algal, se realizaron regresiones
lineales siendo significativas sólo en el TNI. El modelo fue altamente significativo y tuvo un poder
explicativo de un 51%. La relación se definió como lineal positiva. Al igual que para el CO2, Liméns y
Raeiros fueron los que con mayores biomasas algales, liberaron también mayores concentraciones
de TNI. También Abra registró altas concentraciones pero con menores biomasas. Por el contrario,
Carnota y Ladeira registraron concentraciones bajas en torno a los 20 g de algas.
Figura 9. Variación del TNI en función de la biomsa algal para todas las playas.
Se realizaron regresiones lineales separando los tipos de algas para analizar la influencia de la
biomasa algal sobre la liberación de TNI en función de su estado degradativo, es decir, del tiempo
que lleva varada en la playa. La cantidad de TNI aumentó proporcionalmente con la biomasa algal
pero no difirió entre los tipos de alga, en cambio, sí lo hizo la cantidad de biomasa que liberó esas
concentraciones. Se observó por lo tanto, que las biomasas de las algas secas fueron las más bajas y
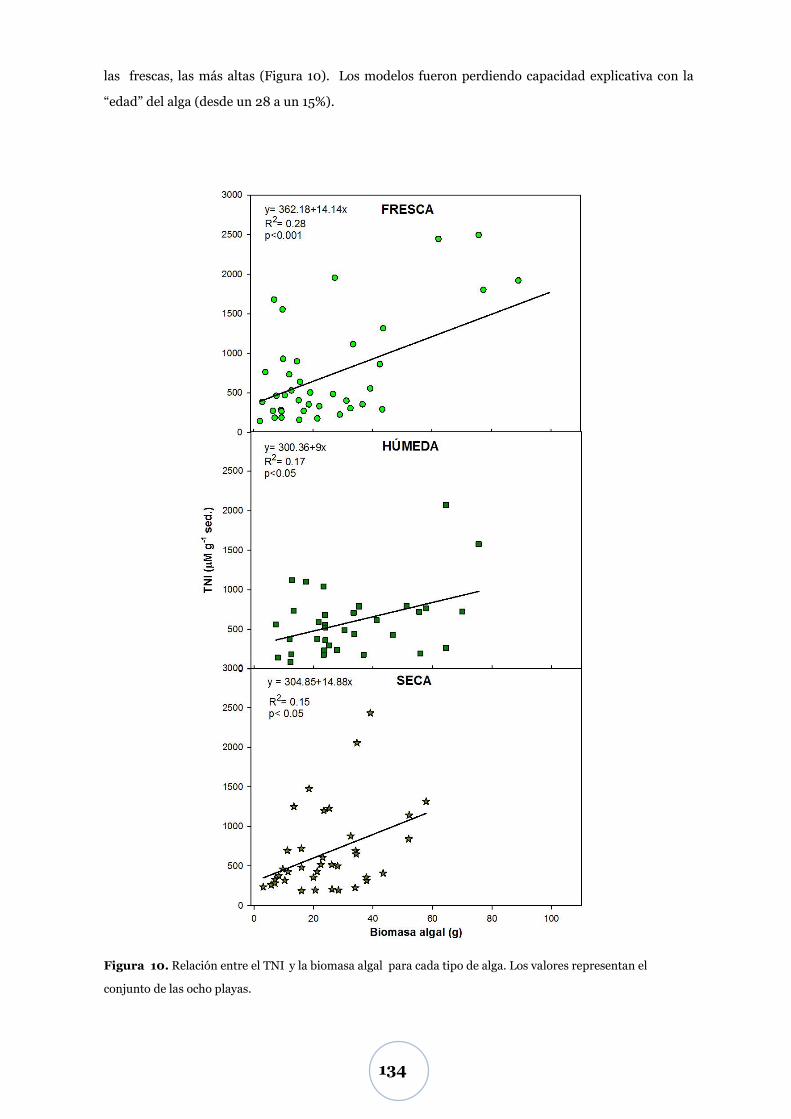
134
las frescas, las más altas (Figura 10). Los modelos fueron perdiendo capacidad explicativa con la
―edad‖ del alga (desde un 28 a un 15%).
Figura 10. Relación entre el TNI y la biomasa algal para cada tipo de alga. Los valores representan el
conjunto de las ocho playas.

135
Efecto de la macrofauna
Se realizaron regresiones para estudiar la influencia que podría tener el número de
individuos de la macrofauna sobre la liberación de nutrientes al sedimento pero no se encontró (p>
0.05). En la figura 11 se representa la cantidad de individuos (área de cada sector) existentes en cada
tipo de parche de algas (color de cada sector). En cada sector se detallan las concentraciones
medidas de TNI y PO4-3 en cada tipo de alga. En las algas frescas se encontró el mayor número de
individuos de macrofauna (69%) y en las secas el menor (2.56%) pero no por ello se registró la
concentración de TNI más baja. De hecho, se encontró en las frescas aun teniendo el mayor número
de individuos. La concentración más alta se registró en las algas húmedas. El patrón que siguió el
fosfato fue distinto. La concentración más baja se registró en los controles mientras que la más alta,
en las frescas (Figura 11).
Figura 11. Representación de la abundancia de la macrofauna en cada tipo de alga (área y color de cada sector
respectivamente). En cada sector se indican las concentraciones medias de cada tipo de alga y de los controles.
Media (SE).
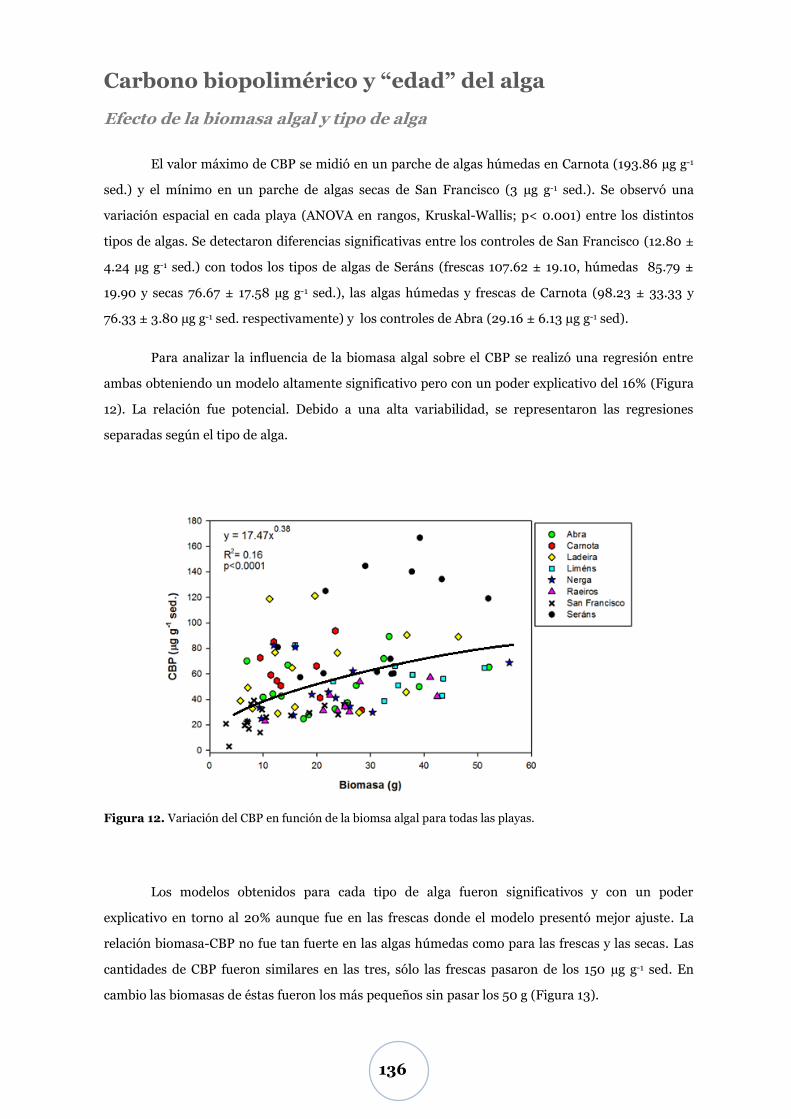
136
Carbono biopolimérico y “edad” del alga
Efecto de la biomasa algal y tipo de alga
El valor máximo de CBP se midió en un parche de algas húmedas en Carnota (193.86 µg g-1
sed.) y el mínimo en un parche de algas secas de San Francisco (3 µg g-1 sed.). Se observó una
variación espacial en cada playa (ANOVA en rangos, Kruskal-Wallis; p< 0.001) entre los distintos
tipos de algas. Se detectaron diferencias significativas entre los controles de San Francisco (12.80 ±
4.24 µg g-1 sed.) con todos los tipos de algas de Seráns (frescas 107.62 ± 19.10, húmedas 85.79 ±
19.90 y secas 76.67 ± 17.58 µg g-1 sed.), las algas húmedas y frescas de Carnota (98.23 ± 33.33 y
76.33 ± 3.80 µg g-1 sed. respectivamente) y los controles de Abra (29.16 ± 6.13 µg g-1 sed).
Para analizar la influencia de la biomasa algal sobre el CBP se realizó una regresión entre
ambas obteniendo un modelo altamente significativo pero con un poder explicativo del 16% (Figura
12). La relación fue potencial. Debido a una alta variabilidad, se representaron las regresiones
separadas según el tipo de alga.
Figura 12. Variación del CBP en función de la biomsa algal para todas las playas.
Los modelos obtenidos para cada tipo de alga fueron significativos y con un poder
explicativo en torno al 20% aunque fue en las frescas donde el modelo presentó mejor ajuste. La
relación biomasa-CBP no fue tan fuerte en las algas húmedas como para las frescas y las secas. Las
cantidades de CBP fueron similares en las tres, sólo las frescas pasaron de los 150 µg g-1 sed. En
cambio las biomasas de éstas fueron los más pequeños sin pasar los 50 g (Figura 13).
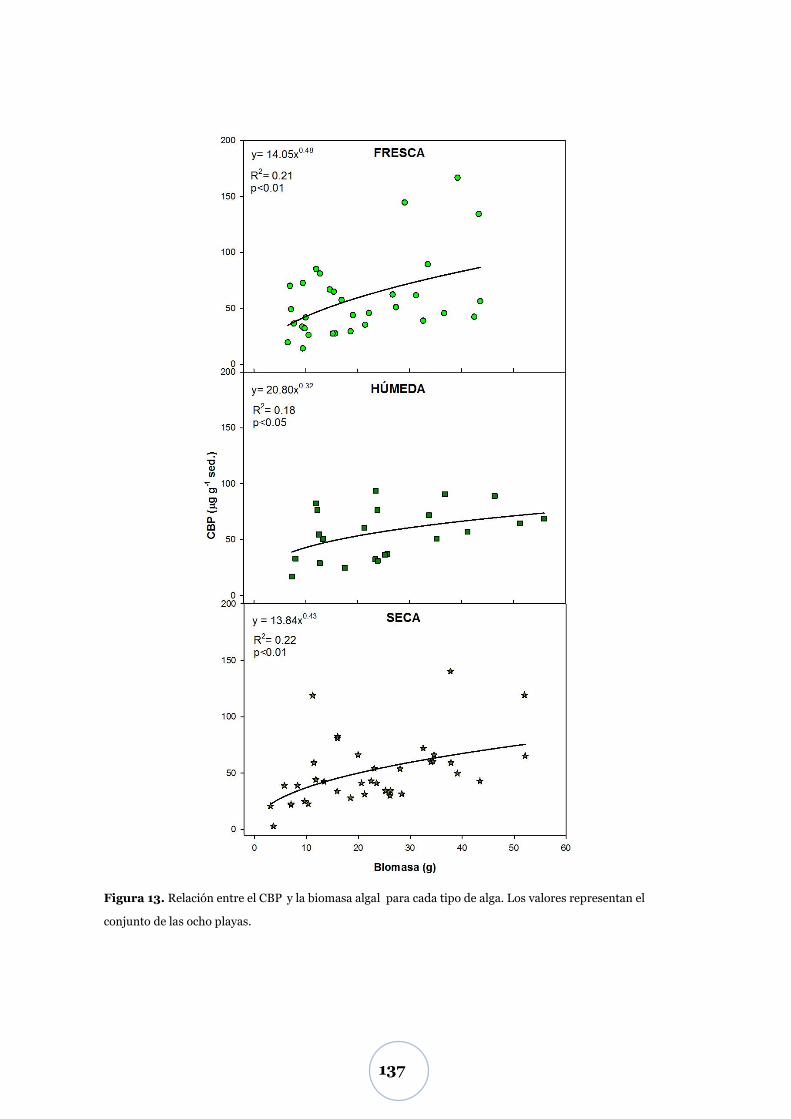
137
Figura 13. Relación entre el CBP y la biomasa algal para cada tipo de alga. Los valores representan el
conjunto de las ocho playas.
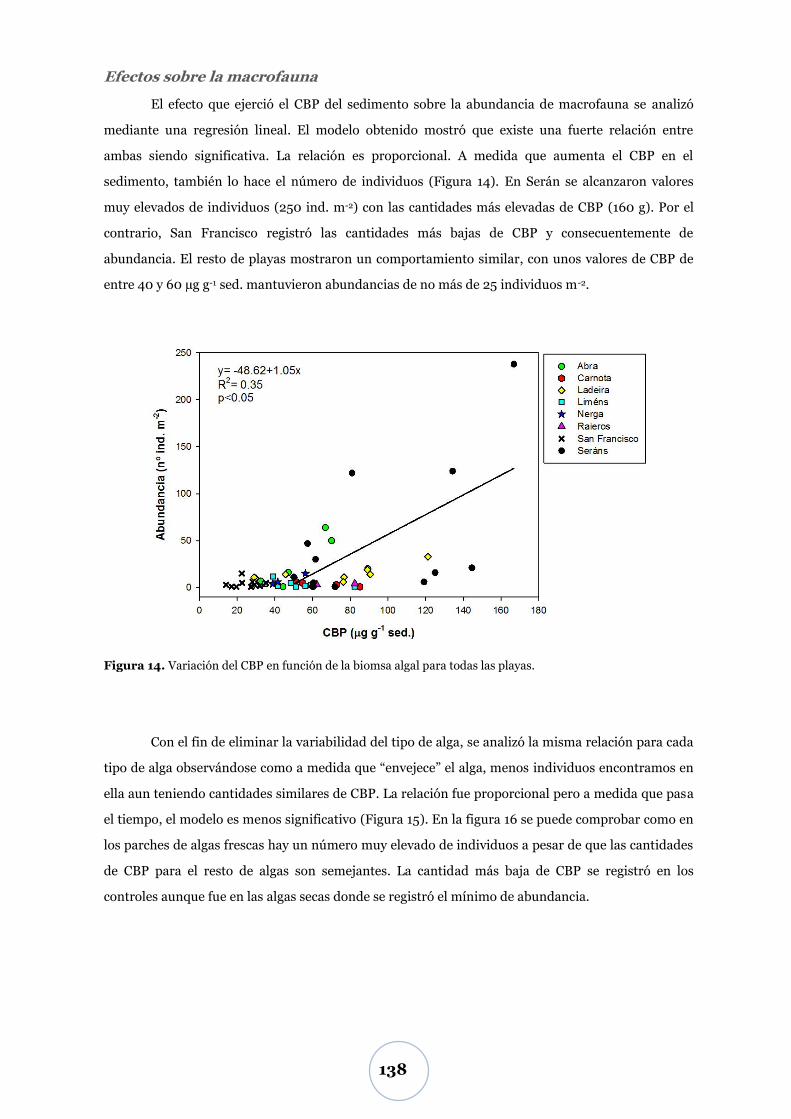
138
Efectos sobre la macrofauna
El efecto que ejerció el CBP del sedimento sobre la abundancia de macrofauna se analizó
mediante una regresión lineal. El modelo obtenido mostró que existe una fuerte relación entre
ambas siendo significativa. La relación es proporcional. A medida que aumenta el CBP en el
sedimento, también lo hace el número de individuos (Figura 14). En Serán se alcanzaron valores
muy elevados de individuos (250 ind. m-2) con las cantidades más elevadas de CBP (160 g). Por el
contrario, San Francisco registró las cantidades más bajas de CBP y consecuentemente de
abundancia. El resto de playas mostraron un comportamiento similar, con unos valores de CBP de
entre 40 y 60 µg g-1 sed. mantuvieron abundancias de no más de 25 individuos m-2.
Figura 14. Variación del CBP en función de la biomsa algal para todas las playas.
Con el fin de eliminar la variabilidad del tipo de alga, se analizó la misma relación para cada
tipo de alga observándose como a medida que ―envejece‖ el alga, menos individuos encontramos en
ella aun teniendo cantidades similares de CBP. La relación fue proporcional pero a medida que pasa
el tiempo, el modelo es menos significativo (Figura 15). En la figura 16 se puede comprobar como en
los parches de algas frescas hay un número muy elevado de individuos a pesar de que las cantidades
de CBP para el resto de algas son semejantes. La cantidad más baja de CBP se registró en los
controles aunque fue en las algas secas donde se registró el mínimo de abundancia.
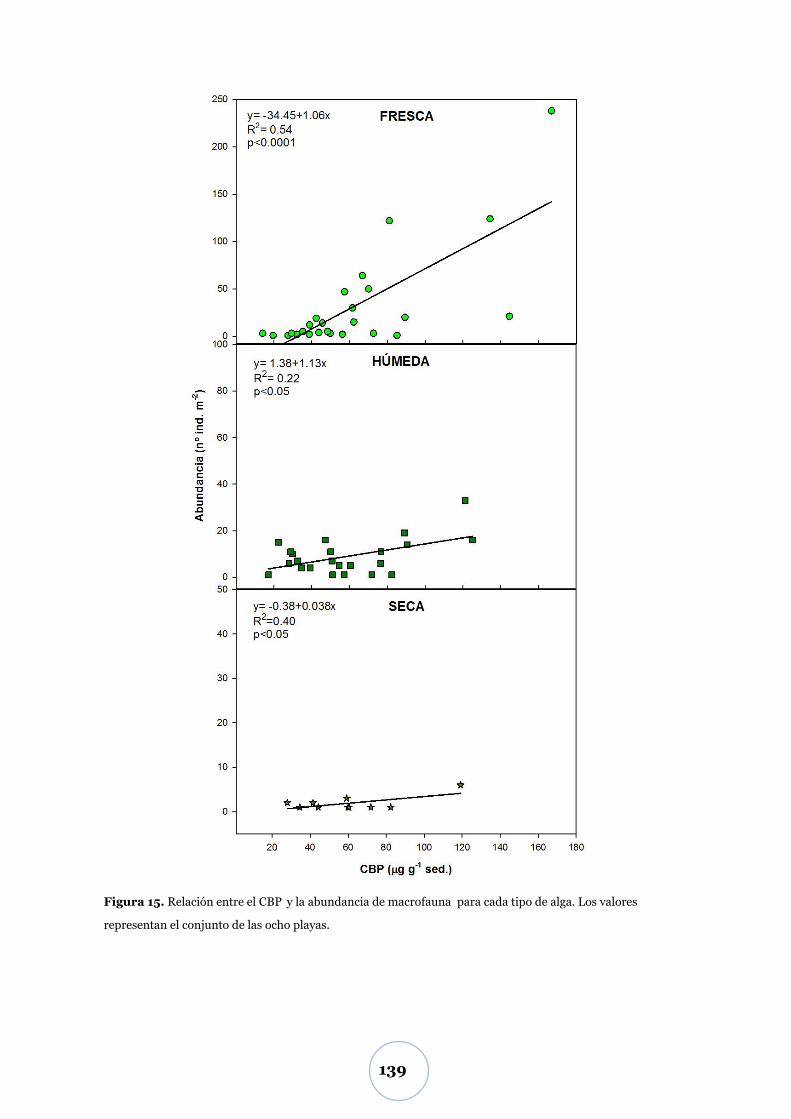
139
Figura 15. Relación entre el CBP y la abundancia de macrofauna para cada tipo de alga. Los valores
representan el conjunto de las ocho playas.
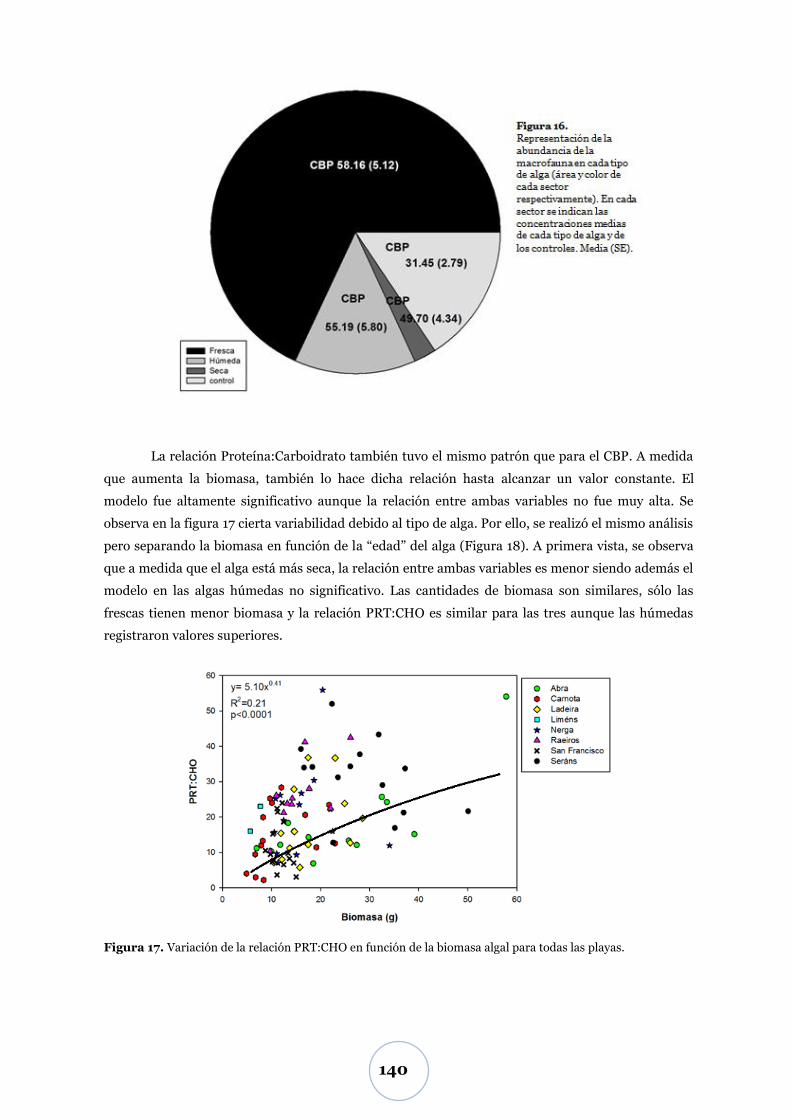
140
La relación Proteína:Carboidrato también tuvo el mismo patrón que para el CBP. A medida
que aumenta la biomasa, también lo hace dicha relación hasta alcanzar un valor constante. El
modelo fue altamente significativo aunque la relación entre ambas variables no fue muy alta. Se
observa en la figura 17 cierta variabilidad debido al tipo de alga. Por ello, se realizó el mismo análisis
pero separando la biomasa en función de la ―edad‖ del alga (Figura 18). A primera vista, se observa
que a medida que el alga está más seca, la relación entre ambas variables es menor siendo además el
modelo en las algas húmedas no significativo. Las cantidades de biomasa son similares, sólo las
frescas tienen menor biomasa y la relación PRT:CHO es similar para las tres aunque las húmedas
registraron valores superiores.
Figura 17. Variación de la relación PRT:CHO en función de la biomasa algal para todas las playas.
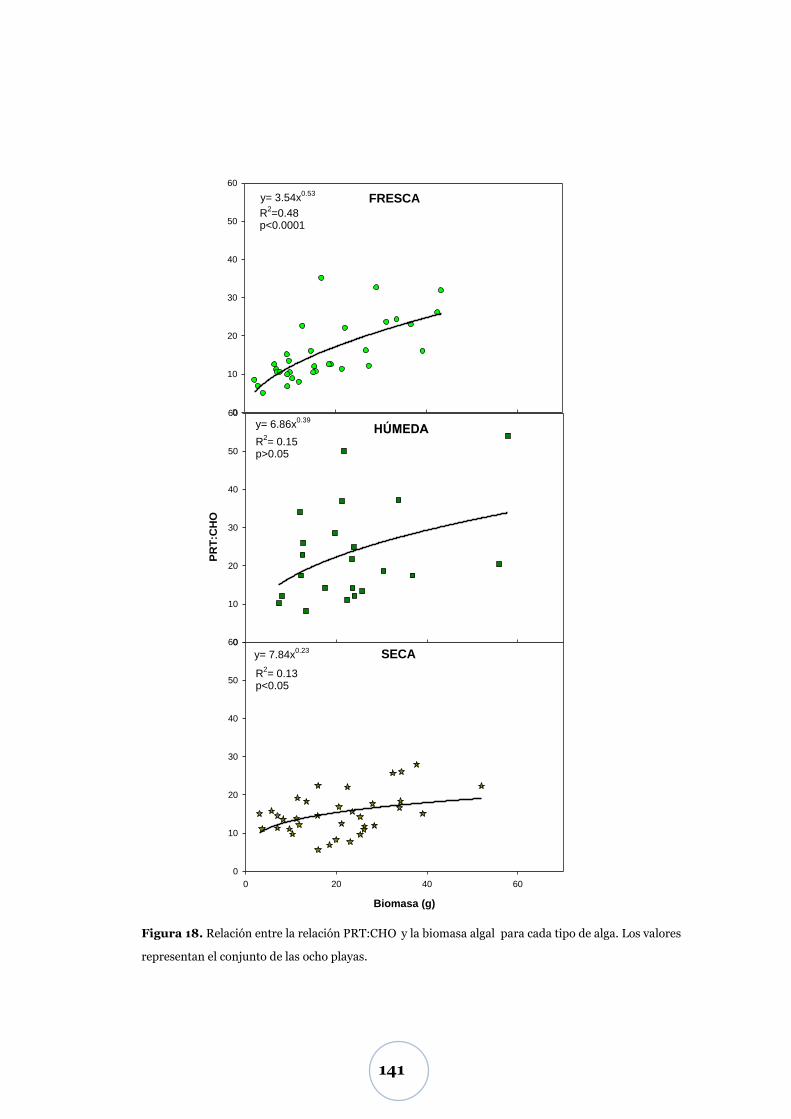
141
FRESCAP
RT
:CH
O
0
10
20
30
40
50
60
y= 3.54x0.53
R2=0.48
p<0.0001
HÚMEDA
0
10
20
30
40
50
60
y= 6.86x0.39
R2= 0.15
p>0.05
SECA
Biomasa (g)
0 20 40 60
0
10
20
30
40
50
60
y= 7.84x0.23
R2= 0.13
p<0.05
Figura 18. Relación entre la relación PRT:CHO y la biomasa algal para cada tipo de alga. Los valores
representan el conjunto de las ocho playas.

142
La relación PRT:CHO osciló entre 18.43 ± 1.17 (fresca) y 13.17 ± 1.13 (control) (Figura 19). La
relación con la abundancia sólo coincidió en el parche de algas frescas ya que éstas fueron las que
albergaron mayor número de individuos. En el caso contrario no hubo coincidencias ya que el
número más bajo de individuos no estuvo en el parche con menor relación , si no, en el parche de las
algas secas.
DISCUSIÓN
Parámetros físico-ambientales y biológicos
Cuando el wrak queda varado a lo largo de la playa, éste se ve sometido a numerosos
procesos físicos y biológicos como la fragmentación, descomposición y mineralización por los
organismos descomponedores. Este proceso de ―envejecimiento‖ que va sufriendo el wrack es clave
para la degradación de la materia orgánica por parte de los descomponedores (Krumhansl &
Scheibling, 2012). Los factores que determinan la velocidad de degradación de la materia orgánica
son numerosos. Por un lado están los factores externos o físico-ambientales (temperatura, humedad,
oxígeno, composición y tamaño del sedimento, etc) y por el otro los factores biológicos (la
composición química de la materia orgánica y la eficiencia de los organismos descomponedores).
Ambos parámetros han de estar en consonancia para que se dé la máxima velocidad de
descomposición posible (Colombi et al., 2003; Coupland et al., 2007).
Figura 19. Representación de la
abundancia de la macrofauna en cada
tipo de alga (área y color de cada sector
respectivamente). En cada sector se
indican las relaciones PRT:CHO medias
de cada tipo de alga y de los controles.
Media (SE).

143
Efectos de la biomasa algal
La actividad metabólica depende en gran medida de la cantidad de biomasa algal
acumulada. Los resultados obtenidos de las regresiones con el CO2, el TNI, CBP y relación
PRT:CHO, son apoyados por anteriores estudios (Cividades et al., 2002; Mateo et al., 2006; Dugan
et al., 2011; Barreiro et al.,2012; Colombini et al., 2003). La biomasa algal influyó de igual manera
para las variables estudiadas. El aumento de la biomasa algal provocó la formación de CO2, el TNI y
CBP (productos de la descomposición). La relación PRT:CHO fue utilizada como indicador de la
―edad‖ del alga varada (Hobson, 1967; Cauwet, 1978). Su uso ha facilitado el entendimiento de las
cantidades emitidas de CO2, TNI y CBP ya que dependiendo del estado de descomposición del alga,
los resultados varían. Por lo tanto, en las algas frescas, la actividad metabólica fue superior que en
las secas. En las primeras fases de descomposición existe una rápida formación de CO2, el TNI y
CBP debido al acelerado crecimiento de la comunidad microbiana que son las que los producen. En
el caso del CO2 y CBP, las relaciones con la biomasa para las algas frescas y secas fueron muy
parecidas. Puede deberse a que muchas algas poseen polifenoles que las protegen del ataque
microbiano impidiendo su consumo (Goecke et al., 2010; Nagayama, 2002; Sosik & Simenstad,
2013; Gómez, 2015). Por eso existen pocas diferencias entre ambos estados degradativos. En estos
casos, el paso del tiempo permitirá que los factores físico-ambientales vayan degradando la pared
celular para permitir el consumo de los organismos (Duong, 2008). La relación entre la biomasa
algas con la relación PRT:CHO mostró también una relación proporcional, indicando que a mayor
biomasa, la relación PRT:CHO aumentaba. Esto puede ser debido a que los parches de menor
tamaño, tienen mayor número de organismos y por lo tanto, mayor actividad metabólica (debido a
una mayor oxigenación). Así pues, la elevada cantidad de proteína medida, indica que el alimento es
de origen animal posiblemente procedente de los organismos que se encuentran en los parches de
algas descomponiéndolas (Figura 20).
Efectos de la macrofauna
Entre los grupos más abundantes destacaron los anfípodos, isópodos, dípteros y
coleópteros. Estos dos últimos son típicos del ambiente terrestre, de la zona supramareal (de la Huz,
2008). Ladeira y Seráns destacaron por albergar el mayor número de estos insectos. Éstas son
playas con complejos dunares (se encuentran en las inmediaciones del complejo dunar de
Corrubedo) y la mayor parte de la biomasa algal se concentra en la base de las dunas. Por el otro
lado, los anfípodos e isópodos característicos del ambiente marino (Dahl, 1952), fueron más
abundantes en Abra. Esta playa se caracteriza por los altos aportes de wrack y por lo tanto soporta
extensas poblaciones de estos crustáceos (como se vio en los capítulos anteriores).
Al igual que los depósitos de macroalgas, la macrofauna asociada son variables también
varía en el espacio pero lo hace en función de la distribución del alimento. Los anfípodos, isópodos,

144
dípteros y coleópteros colonizan rápidamente los depósitos de algas en sólo uno o dos días (Griffiths
& Stenton-Dozey, 1981; Koop & Griffiths, 1982, 1989; Jedrzejczak, 2002a; Colombini & Chelazzi,
2003). Dependiendo del tipo de alga según su ―edad‖ puede haber un cambio de la macrofauna
hacia los taxones marinos o terrestres (Lavoie, 1985).
Está demostrado que la disponibilidad de alimento en las playas es uno de los principales
factores que determinan la presencia de los organismos y por lo tanto, la variación espacial de éstos
(Witlatch, 1981). Por este motivo, en los parches de mayor tamaño se encontró en el sedimento
subyacente mayor cantidad de CBP y por lo tanto, las abundancias más altas de macrofauna como en
estudios realizados por Egglishaw (1965) o Griffiths y Stenton-Dozey (1981). La macrofauna que
coloniza los depósitos de wrack incluye miembros de múltiples niveles tróficos. La mayoría de los
estudios indican que el consumo de algas marinas por la macrofauna es mínimo (Robertson & Mann
1980; Jedrzejczak, 2002a) pero el consumo de algas por anfípodos y dípteros puede llegar a ser
importante (entre el 60 y el 80%) en la primera quincena (Griffiths & Stenton-Dozey 1981). A sí
pues, la macrofauna tiene el potencial de consumir grandes cantidades de macroalgas. Este hecho
estuvo asociado a las altas actividades metabólicas, traducido en altas cantidades de CO2. El caso
contrario fue lo ocurrido en las algas secas, donde tanto la actividad metabólica como la abundancia
fueron menores debido a la ausencia de valor nutricional. Tan sólo pudieron tener función
estructural como refugio o lugar para la cría (Gómez, 2015). Al igual que en estudios anteriores de
Catenazzi & Donnelly (2007), las algas frescas fueron las que albergaron mayor número de
individuos pero a medida que el alga era más ―vieja‖ (seca) el consumo descendió debido a que sus
tejidos se endurecieron y fueron más complicados de digerir (Dethier et al., 2014). La macrofauna se
encontró siempre en las algas frescas y su relación con la biomasa algal se fue debilitando con la
―edad‖ del alga. Además, las pendientes de los modelos obtenidos son superiores en las algas frescas
que en las secas, por lo que aunque si el parche de algas tuviese una función meramente estructural,
se podrían encontrar más individuos en parches más grandes aunque fuesen más ―viejos‖ pero esto
no sucedió.
Los resultados no mostraron relación entre las concentraciones de nutrientes y la
abundancia de macrofauna. Puede estar relacionado con el consumo, ya que la macrofauna no
consume directamente los nutrientes, si no, a través del herbivorismo (Griffiths & Stenton-Dozey,
1981).

145
Figura 20. Relación entre la biomasa algal con la relación PRT:CHO por gramo de biomasa.
Flujo de CO2
El CO2 es utilizado como indicador de la actividad metabólica (Williams & del Giorgio, 2005;
Gómez, 2015). Del conjunto de playas, las algas frescas presentaron la actividad metabólica más
elevada ya que en los primeros días, aproximadamente 3, la comunidad microbiana se desarrolla
rápidamente (Robertson & Hansen, 1982; Gómez, 2015). En el lado opuesto, los controles emitieron
las cantidades más bajas de CO2 ya que, al no haber algas, no hubo ninguna comunidad
descomponedora asociada. Esto hace que en una misma playa existan zonas próximas entre sí,
donde las diferencias en la actividad metabólica sean muy grandes derivando en una alta
variabilidad espacial como la que se obtuvo en la ANOVA. Estos resultados coincidieron con los
realizados por Coupland et al. (2007) donde el wrack emitió una media de 6.62 ± 0.88 µmol C m-2 s-
1, comparado con el valor medio de una zona de arena sin algas (2.21 ± 1.25 µmol C m-2 s-1). Es lo que
se conoce como ―Hot spots‖ o puntos calientes creado por McClain (2003); son zonas con tasas
metabólicas desproporcionadamente altas en comparación con el ambiente que le rodea (McClain,
2003; Coupland et al., 2007).
Liberación de nitrógeno inorgánico y fósforo
El conjunto de la actividad descomponedora de hongos, bacterias y organismos
pertenecientes a la meio y macrofauna, hacen posible la mineralización de la materia orgánica. Este
es un proceso catabólico donde los compuestos orgánicos son oxidados para obtener compuestos
inorgánicos simples (CO2, H2S, NH4+, PO4
-3 y H2O) (Wetzel y Likens, 1991; Álvarez, 2005). Parte de

146
los materiales orgánicos son incorporados como biomasa en distintos organismos detritívoros
(Maltby, 1996; Álvarez, 2005); estudios anteriores han descubierto que aproximadamente el 94.2 %
del nitrógeno del alga mineralizado por las bacterias. (Colombini et al., 2003). Los resultados del
ANOVA no mostraron diferencias en las concentraciones de nutrientes (TNI y PO4-3) entre las playas
y el tipo de alga o control. En capítulos anteriores se han explicado qué factores físicos son los que
influyen en la actividad microbiana. Puesto que las bacterias son las principales responsables de
estas variaciones, los resultados obtenidos pueden explicar que las condiciones físico-ambientales
(temperatura, oxígeno, % de humedad del alga y del sedimento) estuviesen dentro de los rangos
óptimos en todas las playas (Kirkman & Kendrick 1997; Coupland et al., 2007). Al realizarse este
estudio en la misma época del año durante un corto periodo, en la misma franja horaria y en playas
con características similares, pudo haberse eliminado cierta variabilidad. Las altas concentraciones
de amonio medidas en todos los tipos de algas, revelan que todas ellas se encontraban en las
primeras fases de descomposición. Se sabe que el amonio que se libera puede ser transformado en
NO2- y NO3
- por nitrificación (Miranda et al., 2013) o consumido por bacterias heterotróficas y / o
microalgas bentónicas (García-Robledo et al., 2008; Barreiro et al., 2012). Pero la detección de altas
concentraciones de amonio en todos los tipos de algas, demuestran que se encuentran en las
primeras fases de descomposición.
Las altas concentraciones de fósforo (procedente del fosfato) pueden indicar como se
mencionó anteriormente, que las algas se encuentran en las primeras fases de descomposición.
Durante la descomposición la mayor parte del fósforo orgánico es mineralizado, el resto es
adsorbido por los sedimentos (Sundby et al., 1992). De ahí que se hayan obtenido altas
concentraciones, ya que en este estudio el fósforo se extrajo del sedimento. De lo contrario, el
fósforo habría sido consumido por la comunidad bacteriana y las concentraciones habrían sido
mucho más bajas (García-Robledo et al., 2008; Barreiro et al., 2012).
Carbono biopolimérico
El carbono biopolimérico es el resultado de la suma de las concentraciones de lípidos,
proteínas y carbohidratos que le confieren al sedimento su valor nutricional (Incera et al., 2003). En
el presente estudio se pudo observar variación espacial entre playas y dentro de ellas según su
distancia a la orilla como ha ocurrido en anteriores estudios (Coupland et al., 2007). El patrón
general del CBP es encontrar mayores cantidades en el sedimento subyacente de los parches de algas
frescas y húmedas. Los controles de las playas de San Francisco y Raeiros mostraron diferencias
significativas ya que las cantidades de CBP fueron en muchos casos la mitad de las cantidades
registradas en Seráns o Carnota (sobre todo en algas frescas y húmedas). El CBP de los controles de
S. Francisco y Raeiros fue bajo, seguramente por su consumo o lavado por las mareas (en estos casos

147
se descartaron las descargas fluviales y las precipitaciones ya que fueron inexistentes). Raeiros
recibió menor cantidad de biomasa que Carnota y al tener mayor pendiente y tamaño de grano que
ésta, todo indica a que el lavado del CBP pudo ser la principal causa de la diferencia significativa.
Además Carnota fue la playa con la abundancia más baja de todas por lo que fue menos probable
que el CBP fuese consumido por la macrofauna (Pearson & Rosenberg, 1978, 1987; Whitlaton, 1981;
Incera et al., 2003). La diferencia entre los controles de S. Francisco y Abra se basa principalmente
en la biomasa que reciben siendo Abra casi el triple. Al aumentar la biomasa algal, aumenta el
desarrollo de la comunidad microbiana encargada de catabolizar las reacciones químicas que
producirán el CBP (Daumas et al., 1983; Fichez, 1991; Danovaro et al., 1993). Aunque S. Francisco
reciba mayores biomasas de algas, las diferencias con Seráns parecen ser que se basan en el tamaño
de grano más grueso de Seráns. Se sabe que la oxigenación es imprescindible para que la comunidad
bacteriana pueda descomponer la materia orgánica (Mc Lachlan, 1983). El sedimento de Seráns
es más favorable que el de S. Francisco, que a pesar de recibir mayor cantidad de algas, éstas al
acumularse en parches, no permiten el intercambio gaseoso con el medio reduciéndose en gran
medida la actividad bacteriana y por lo tanto la formación de carbono biodisponible.
CONCLUSIONES
El alga cuanto más tiempo lleva depositada en la playa, su estado degradativo está más
avanzado y su composición química es distinta que al comienzo del proceso de descomposición.
Esto afecta inevitablemente a las comunidades faunísticas que la vayan a colonizar. Por eso, en los
primeros días tras su varamiento, será colonizada rápidamente por la comunidad bacteriana, lo que
se traducirá en altas cantidades de CO2 y nutrientes (TNI y fosfatos). También se le sumará la
macrofauna ya que en el sedimento subyacente al parche de algas existe CBP. Por otro lado, las algas
secas no albergarán más individuos aunque tengan mayor biomasa. Son precisamente los parches
más pequeños los que tienen mayor actividad metabólica según la relación PRT:CHO aunque sobre
esta cuestión, se debería seguir estudiando para poder concretar mejor el papel de la biomasa algal
sobre la ―edad‖ del alga.

148
BIBLIOGRAFÍA
Alongi, D. M. (1998) Beaches and tidal flats, In: Coastal ecosystem processes, CRC Press, New York.
Barreiro, F., Gómez, M., López, J., Lastra, M., & De la Huz, R. (2012). Coupling between macroalgal
inputs and nutrients outcrop in exposed sandy beaches. Hydrobiologia, 1-12.
Catenazzi, A., & Donnelly, M. A. (2007). Role of supratidal invertebrates in the decomposition of
beach-cast green algae Ulva sp. Marine Ecology Progress Series, 349, 33-42.
Cauwet, G., 1978. Organic chemistry of sea water particulates: concepts and developments. Oceanol.
Acta 1: 99–105. Barreiro, F., Gómez, M., López, J., Lastra, M., & De la Huz, R. (2012).
Coupling between macroalgal inputs and nutrients outcrop in exposed sandy beaches.
Hydrobiologia, 1-12.
Cividanes, S., Incera, M., & López, J. (2002). Temporal variability in the biochemical composition of
sedimentary organic matter in an intertidal flat of the Galician coast (NW Spain).
Oceanologica Acta, 25(1), 1-12.
Colombini, I., Chelazzi, L., Gibson, R. N., & Atkinson, R. J. A. (2003). Influence of marine
allochthonous input on sandy beach communities. Oceanography and Marine Biology: An
Annual Review, 41, 115-159.
Coupland, G. T., C. M. Duarte & D. I. Walker, 2007. High metabolic rates in beach cast communities.
Ecosystems 10:1341–1350.
Dahl, E. (1952). Some aspects of the ecology and zonation of the fauna on sandy beaches. Oikos,
4(1), 1-27.
Danovaro, R., M. Fabiano & N. Della Croce, 1993. Labile organicmatter and microbial biomasses in
deep-sea sediments (Eastern Mediterranean Sea). Deep-Sea Res. 5: 953–965.
Daumas, R., D. Sautriot & D. Calmet, 1983. Evolution de constituants labiles de la matière organique
dans les sediments profonds de diverses marges continentales. D’Orgon à Misedor. C.N.R.S.
Paris (ed.), Géochimie organique des sédiments marins: 99–150.
De La Huz R (2008) Ecologia de la macrofauna de las playas expuestas de Galicia. Universidade de
Vigo.

149
Decho, A. W., & Fleeger, J. W. (1988). Microscale dispersion of meiobenthic copepods in response to
food-resource patchiness. Journal of Experimental Marine Biology and Ecology, 118(3), 229-
243.
Dethier, M. N., Brown, A. S., Burgess, S., Eisenlord, M. E., Galloway, A. W. E., Kimber, J., ... &
Duggins, D. O. (2014). Degrading detritus: changes in food quality of aging kelp tissue varies
with species. Journal of Experimental Marine Biology and Ecology, 460, 72-79.
Dugan, J. E., Hubbard, D. M., Page, H. M., & Schimel, J. P. (2011). Marine macrophyte wrack inputs
and dissolved nutrients in beach sands. Estuaries and Coasts, 34(4), 839-850.
Duong, H. L. S., & Stephanie, L. (2008). Investigating the ecological implications of wrack removal
on South Australian sandy beaches. Flinders University, School of Biological Sciences.
Egglishaw, H. J. (1965) Observations on the fauna of wrack beds, Transactions of the Society for
British Entomology 16, 189-216.
Fichez, R., 1991. Composition and fate of organic matter in submarine cave sediments, implications
for the biogeochemical cycle of organic carbon. Oceanol. Acta 14: 369–377.
García-Robledo E, Corzo A, García de Lomas J, van Bergeijk S (2008) Biogeochemical effects of
macroalgal decomposition on intertidal microbenthos: a microcosm experiment. Mar Ecol
Prog Ser 356:139 151.
Goecke, F., Labes, A., Wiese, J., Imhoff, J., 2010. Chemical interactions between marine macroalgae
and bacteria. Mar. Ecol. Prog. Ser. 409, 267–299.
Gómez Rodríguez, M. (2015). Ecología de los varamientos de macroalgas en playas estuáricas de la
costa de Galicia (Doctoral dissertation, Ecoloxía e bioloxía animal).
Griffiths CL, J Stenton-Dozey & K Koop (1983) Kelp wrack and the flow of energy through a sandy
beach ecosystem. En: McLachlan A & T Erasmus (eds) Sandy beaches as ecosystems: 547-
556. Dr. W. Junk Publishers, The Hague, The Netherlands. Hobson, L. A., 1967. The
seasonal and vertical distribution of suspended particulate matter in an area of the
Northeast Pacific Ocean. Limnol. Oceanogr. 12: 642–649.
Harrison, P. G. (1982) Control of microbial growth and of amphipod grazing by water-soluble
compounds from leaves of Zostera marina, Marine Biology 67, 225-230.
Incera, M., Cividanes, S. P., López, J., & Costas, R. (2003). Role of hydrodynamic conditions on
quantity and biochemical composition of sediment organic matter in sandy intertidal
sediments (NW Atlantic coast, Iberian Peninsula). Hydrobiologia, 497(1), 39-51.

150
Jedrzejczak, M. F. (2002a) Spatio-temporal decay 'hot spots' of stranded wrack in a Baltic sandy
coastal system. Part I. Comparative study of the pattern: 1 type of wrack vs 3 beach sites,
Oceanologia 44, 491-512.
Kirkman H, Kendrick GA. 1997. Ecological significance and commercial harvesting of drifting and
beach-cast macro-algae and seagrass in Australia: a review. J Appl Phycol 9:311–326.
Krumhansl, K.A., Scheibling, R.E., 2012. Detrital subsidy from subtidal kelp beds is altered by the
invasive green alga Codium fragile ssp. fragile. Mar. Ecol. Prog. Ser. 456, 73–85.
Maltby, L 1996. Detritus processing. En River Biota: diversity & dynamics. Selected extracts from
the Rivers Handbook. Petts, G., P. Calow, (Eds). Blackwell science. Oxford.145-167.
Mateo, M., Cebrián, J., Dunton, K., & Mutchler, T. (2006). Carbon flux in seagrass ecosystems.
Seagrasses: biology, ecology and conservation, 159-192.
McClain ME, Boyer EW, Dent CL, Gergel SE, Grimm NB, Groffman PM, Hart SC, Harvey JW,
Johnston CA, Mayorga E, McDowell WH, Pinay G. 2003. Biogeochemical hotspots and hot
moments at the interface of terrestrial and aquatic ecosystems. Ecosystems 6:310–312.
McLachlan, A. (1981) Exposed sandy beaches as semi-enclosed ecosystems, Marine Environmental
Research 2, 59-63.
McLachlan A., 1982. A model for the estimation of water filtration and nutrient regeneration by
exposed sandy beaches. Mar. Environ. Res. 6: 37–47.
Miranda, R., Alavi, G., Aliaga, S., & Caballero, A. (2013). Mineralización del nitrógeno y generación
de CO2 por descomposición del estiércol ovino. Altiplano central, Bolivia. Venesuelos, 21(2).
Nagayama, K., 2002. Bactericidal activity of phlorotannins from the brown alga Ecklonia kurome. J.
Antimicrob. Chemother. 50 (6), 889–893.
Ochieng, C. A. & Erftemeijer, P. L. A. (1999) Accumulation of seagrass beach cast along the Kenyan
coast: a quantitative assessment, Aquatic Botany 65, 221-238.
Pearse, A.S., H.J. Humm, and G.W. Wharton. 1942. Ecology of sandbeaches at Beaufort, NC.
Ecological Monographs 12(2): 135–190.
Pearson, T. H. & R. Rosenberg, 1978. Macrobenthic succession in relation to organic enrichment and
pollution of the marine environment. Oceanogr. mar. biol. Ann. Rev. 16: 229–311.

151
Pearson, T. H. & R. Rosenberg, 1987. Feast and famine: Structuring factors in marine benthic
communities. In Gee, J. H. R. & P. S. Giller (eds), Organization of Communities, Past and
Present. Blackwell Sci. Publ., Oxford: 373–395.
Rieper-Kirchner, M. (1990) Macroalgal decomposition: laboratory studies with particular regard to
microorganisms and meiofauna, Helgolander Meeresuntersuchungen 44, 397-410.
Robertson A. & Hansen J.A. (1982) Decomposing seaweed: a nuisance or a vital link in coastal food
chains? In: CSIRO Marine Laboratories Research Report, pp. 75-83. CSIRO Marine
Laboratories, Cronulla, NSW.
Robertson, A. I. & Mann, K. H. (1980) The role of isopods and amphipods in the initial
fragmentation of eelgrass detritus in Nova Scotia, Canada, Marine Biology 59, 63-69.
Sosik, E.A., Simenstad, C.A., 2013. Isotopic evidence and consequences of the role of microbes
inmacroalgae detritus-based food webs. Mar. Ecol. Prog. Ser. 494, 107–119.
Wetzel, R.G. y Likens, G.E, 1991. Limnological analysis. Second Edition. Springer-Verlang. New-
York.
Williams P.J. & del Giorgio P.A. (2005) Respiration in aquatic systems: history and background. The
global significance of respiration in aquatic systems: from single cells to the bio- sphere. In:
Respiration in aquatic ecosystems (eds. del Giorgio PA & Williams PJ), pp. 1-17. Oxford
University Press, Oxford.
Whitlaton, R. B., 1981. J. exp. mar. Biol. Ecol. 53: 31–45.

152

153
La actividad metabólica de las playas varía estacionalmente con la temperatura. Por eso, en
verano las cantidades de CO2 y de nutrientes son máximas ya que las reacciones químicas
responsables de la descomposición se ven aceleradas.
En cada playa existe variabilidad espacial en la actividad metabólica debida principalmente
a la distribución y cantidad de biomasa varada, la ―edad‖ de esta biomasa y al contenido de
humedad tanto del alga como del sedimento. Esta variación depende por lo tanto del nivel
de marea (distancia a la costa).
Los efectos de los factores bióticos dependen de los abióticos por eso es importante tenerlos
en cuenta en cualquier estudio relacionado con la actividad microbiana.
Las respuestas de los distintos tipos de algas según su ―edad‖ dependen de las variaciones
térmicas en cuanto a las cantidades de CO2 y nutrientes siendo mayores en algas depositdas
recientemente.
La distribución de la macrofauna depende de la disponibilidad de alimento. Por lo tanto se
encontrarán en aquellos lugares de la playa donde la cantidad de carbono biopolimérico sea
mayor. Así pues, predominan en los parches de algas recientemente varadas.
La respiración prodedente de la macrofauna representa un porcentaje mucho más bajo que
el procedente de la comunidad microbiana ya que es ésta la responsable directa del proceso
de descomposición.
La contribución de la macrofauna a la respiración es sobretodo de forma indirecta al triturar
el detritus y transformándolo en trozos mucho más pequeños que son más vulnerables al
ataque microbiológico acelerando su consumo.
La materia orgánica que se deposita en las playas por las mareas, es de suma importancia en
el reciclaje de ésta obteniénose nutrientes que quedan a disposición de los autótrofos. Por lo
tanto el material de arribazón o wrack es la base de las cadenas tróficas de los ecosistemas
costeros.

154
La obtención de estudios como el de la presente tesis son la base fundamental para una
adecuada praxis en la gestión de los recursos del litoral permitiendo un adecuado balance
entre economía y ecología.

