in u.s.a. predominant catalase-negative soilaem.asm.org/content/18/3/340.full.pdf · predominant...
TRANSCRIPT

APPuLE MICROBIOLOGY, Sept. 1969, p. 340-349Copyrights 1969 American Society for Microbiology
Vol. 18, No. 3Printed in U.S.A.
Predominant Catalase-negative Soil Bacteria1III. Agromyces, gen. n., Microorganisms Intermediary to
Actinomyces and NocardiaWILLIAM E. GLEDHILL2 AND L. E. CASIDA, JR.
Department of Microbiology, The Pennsylvania State University, University Park, Pennsylvania 16802
Received for publication 18 February 1969
The occurrence of filamentous, branching, catalase-negative bacteria as a nu-merically predominant microflora of various soils was demonstrated by using a dilu-tion frequency isolation procedure. The major characteristics of these organismswere those of the order Actinomycetales. However, they could not be placed in anyof the present genera of this order and, therefore, a new genus, Agromyces, was pro-posed for these organisms. This genus includes catalase-negative, nutritionally-fastidious microorganisms whose cells produce a true branching mycelium thatfragments into coccoid and diphtheroid forms. Also, they have an oxidative metabo-lism, are microaerophilic to aerobic, and contain neither diaminopimelic acid norlysine as major constituents of the cell wall glycopeptide. The type species wouldbe Agromyces ramosus, gen. n., sp. n. The possible importance of these organismsin clarifying certain phylogenetic relationships of the Actinomycetales is discussed.
Reports of the occurrence of actinomycetepopulations in soil are limited mainly to thoseorganisms which have an oxidative metabolism,produce catalase, and contain diaminopimelicacid (DAP) in their cell walls. In contrast, thefermentative organisms of the family Actino-mycetaceae generally have been isolated onlyfrom warm-blooded animals (21, 30), and havebeen observed only rarely from other naturalsources (16, 18).The ease with which the oxidative species are
obtained from soil has allowed the accumulationof an array of forms, several of which possesscharacteristics intermediary to those of Nocardia,Streptomyces, Corynebacterium, and Mycobac-terium (2, 13). However, thiiere have been noreports on the presence of major soil populationsserving as links between the aerobic and faculta-tive-to-anaerobic genera of the order Actino-mycetales. Such intermediary forms could beimportant in the elucidation of certain phyloge-netic relationships within this order.
Perhaps this scarcity of intermediary formscan be attributed to a lack of suitable isolationtechniques. Casida (6) described a modifieddilution frequency procedure for the isolation ofa numerically predominant, but previously un-
]This research was authorized for publication as paper no. 3533in the journal series of the Pennsylvania Agricultural ExperimentStation on 21 January 1969.
2 Present address: Microbiological Research, Tenneco Chemi-cals, Inc., Piscataway, New Jersey 08854.
known, coccoid soil bacterium. As a result ofcontinued use of this procedure, it was realizedthat the technique was not selective for onlythis one population, but that, depending on theparticular soil examined, several other numeri-cally predominant catalase-negative speciescould be obtained. Thus, from many soils wehave routinely encountered various filamentous,branched, catalase-negative, microaerophilic bac-teria at one or two logarithmic units beyond thedilution endpoint for the "conventional" soilbacteria.The purpose of the present study was to define
the morphological, biochemical, and physiologi-cal characteristics of certain of these forms. Asa result, a new genus, Agromyces, and the typespecies A. ramosus are described, and it is shownthat this genus presents certain characteristicsintermediary to both Actinomyces and Nocardia.
MATERIALS AND METHODSIsolation and culture conditions. The soils and
methods of cell isolation were those employed pre-viously (12). Approximately 100 catalase-negativestrains, suspected of being members of the Actino-mycetales, were isolated for this study. Refrigeratedand frozen stock cultures were grown and maintainedin medium no. 1 (heart infusion broth, 2.5%0; agar,0.08%; pH 7.0), or medium no. 2 [tryptone, 0.5%,0;Casamino acids, 0.4%; (NH4)2HPO4, 0.07%7; NaCl,0.5%; agar, 0.2%; pH 7.0]. Bacto (Difco) mediumcomponents were used throughout this study. Allstrains were grown at 30 C, and working cultures were
340
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

CATALASE-NEGATIVE SOIL BACTERIA
stored at 4 C and transferred bimonthly. Nocardiasalmonicolor PSU-strain N-18, Rothia dentocariosa(ATCC 17931), as well as the Actinomyces species de-scribed previously (12), were employed for compara-tive purposes.
Morphological studies. Growth from one-week-oldmedium no. 1 broth cultures was transferred to heartinfusion agar slants and to medium no. 2. At intervals,samples from each medium were prepared as Gramstains and wet mounts, and photographs were taken onKodak high-contrast copy film with a Zeiss photo-microscope. Microcolonies on heart infusion agarplates were observed at 24 and 48 hr and photographedas above. Mature colonies (7 to 10 days) were photo-graphed at low magnification by using Kodak Pana-tomic X film. The ability of isolates to form aerialhyphae was studied by using the various media sug-gested by Gordon and Smith (14).Oxygen requirements. One loopful of a uniformly
mixed 3-day-old medium no. 1 broth culture wasstreaked as a single line from the butt to the top of aheart infusion agar slope. Tubes prepared in this man-ner were incubated at 30 C inside a candle jar and un-der the aerobic and anaerobic seals described by Pineand Howell (28). Oxygen requirements were alsoassessed in shake tubes by uniformly dispersing onedrop of 1-in-10 dilution of the above inoculum intoheart infusion agar.
Biochemical tests. Catalase determinations wereroutinely employed to check for culture purity.Growth from an agar surface was removed with aglass rod, mixed with freshly prepared 3% H202 on amicroscope slide, and observed at 10-fold magnifica-tion. The cultures were considered to be catalase-nega-tive if oxygen evolution was not detected during a 10-min observation. The presence of a cytochrome sys-tem was determined by the oxidase (20) and benzidine(8) tests.
Oxidative versus fermentative utilization of sugarswas based on the method of Hugh and Leifson (15),except that medium no. 2 containing 0.003% bromo-thymol blue served as the basal medium. All carbo-hydrates were filter-sterilized as 10% (w/v) solutionsand were added to the medium at a final concentrationof 1%. Tests were considered positive if the pH valuedecreased by at least one unit during a 3-week incuba-tion period.
For examination of organic acid utilization, me-dium no. 2 was adjusted to pH 6.7 and supplementedwith 0.04% phenol red. Solutions of the organic acidswere adjusted to pH 6.7, filter-sterilized, and addedto the medium at a final concentration of 1%. Theappearance of a bright red color (alkaline reaction)during growth indicated acid utilization.
Nitrate reduction and indole production weredetermined with medium no. 2 supplemented, respec-tively, with 0.1% KNO3 or 2% tryptone. The methylred and Voges-Proskauer tests were conducted inmedium no. 2 containing 1% fructose, and argininedeamination and esculin hydrolysis (12) were studiedin medium no. 2. Lysozyme sensitivity was determinedby adding 5 ml of a 0.1% solution of lysozyme in 0.01N HCl (filter-sterilized; Sigma Chemical Co., St. Louis,Mo.) to 100 ml of medium. Evaluation of peroxide
production employed manganese dioxide and heatedblood O-dianisidine agars (38), except that mediumno. 2 served as the basal medium.The hydrolysis of casein, gelatin, starch, and tri-
butyrin, and the utilization of tyrosine and xanthinewere examined, respectively, by the incorporation of0.5% skim milk, 0.4% gelatin, 0.5% starch, 1.0%tributyrin, 0.5% tryosine, or 0.5% xanthine into heartinfusion agar. H2S production was tested by suspend-ing a lead acetate paper strip over a brain heart in-fusion agar slant containing 0.1% cysteine, and by ob-serving growth and blackening of S I M and TripleSugar Iron Agar (Difco). Urease production wasstudied with agar slants containing 2% urea and 0.04%phenol red (pH 6.8). Hemolysis was detected on BloodAgar Base (Difco) containing 5% defibrinated sheepblood.
Nutritional requirements. The study of nutritionalrequirements was based essentially on the work ofLochhead and Chase (23) and Taylor (35). Basal me-dium B was filter-sterilized; it consisted of K2HPO4,0.04%; (NH4)2HP04, 0.05%; NaCl, 0.05%; MgSO4.7H20, 0.005%; FeCl3, 0.001%; and fructose, 0.2%(pH 7.0). Medium BA was basal medium B plus 0.5%vitamin-free Casamino Acids, and medium BAV wasmedium BA plus 1% of a filter-sterilized solution con-taining the following (per 100 ml): riboflavine, 2 mg;calcium pantothenate, 4 mg; biotin, 100 Lug; uracil,150 mg; adenine sulfate, 150 mg; pyridoxine HCI, 12mg; nicotinic acid, 6 mg; p-aminobenzoic acid, 500,ug; choline chloride, 75 isg; thiamine hydrochloride,4 mg; vitamin B12, 30,g; inositol, 0.5 g; and cysteine,0.5 g. Medium BAVT was medium BAV plus 0.5%tryptone. For medium BAVTS, 25% of the water ofmedium BAVT was replaced by soil extract. Glassbeads (3 mm) were added to tubes of the above mediato provide a surface for cell growth. The optimal tem-perature and pH for growth were established in me-dium BAVT. For all media, the extent of growth at 5days was determined as turbidity at 450 nm.
Inoculum (one drop per tube) for the above mediawas grown for 3 days on agar slants of medium no. 2.The growth was washed from the slants with ba-sal medium B, centrifuged, and washed once with simi-lar medium. The cells were then resuspended in basalmedium, incubated for 1 hr at room temperature,washed, and resuspended in medium B to approximatea MacFarland no. 4 standard (1).
Cell wall analysis. Cells grown as shaken cultures inheart infusion broth were centrifuged and washedwith distilled water at 2 C. The method for quantitativepreparation of cell walls (12) was slightly modified inthat the crude cell walls, after being broken with glassbeads, were treated for 24 hr with 0.5% KOH inethyl alcohol at 37 C. To rapidly assess the cell wallcompositions of a large number of isolates, the extrac-tion procedure of Boone and Pine (4) was employed.Hydrolysis and chromatography (12) included oneadditional solvent system (methanol, water, pyridine,HCl in proportions of 32, 7, 4, 1 respectively) for theresolution of basic amino acids (26). Quantities ofamino acids were measured by the method of Kay etal. (19), as modified by Mandelstam and Rogers (24).
Serology. The serological relationships of our
341VOL. 18, 1969
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
GLEDHILL AND CASIDA
isolates to 15 strains of Actinomyces, anaerobic diph-theroids, Rothia, Ramibacterium, and Bacterionemawere studied by J. M. Slack and M. A. Gerencser(Medical Center, University of West Virginia), whoused the fluorescent antibody approach (33).
Pathogenicity. Washed suspensions of our iso-lates were injected intraperitoneally into 6-week-oldmale CF-i mice (Carworth Farms, Rockland Co.,N.Y.). Pathological findings were interpreted on thebasis of the work of Buchanan and Pine (5).
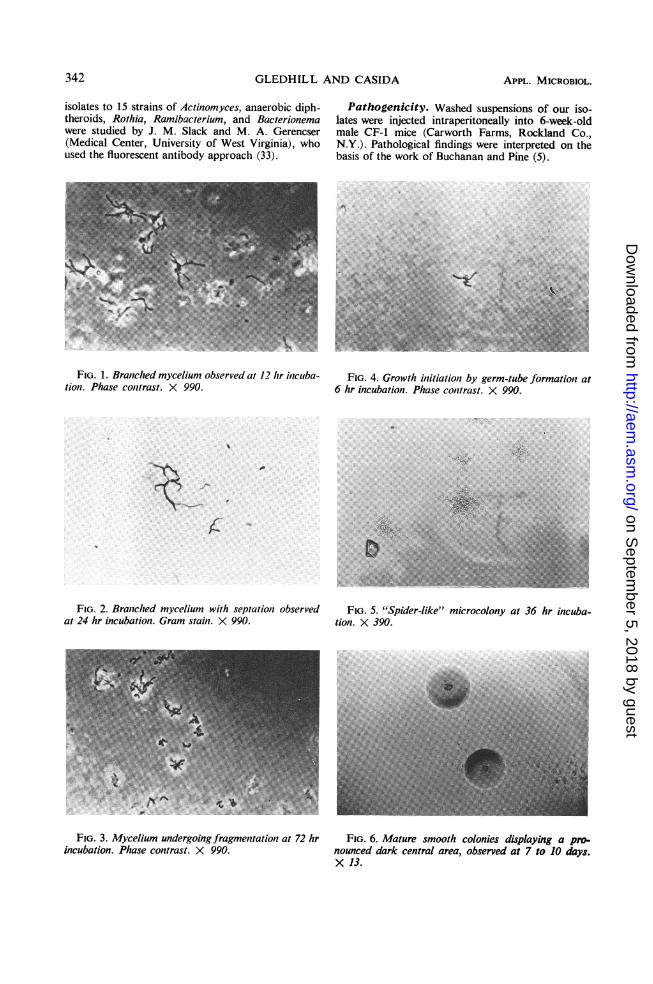
FIG. 1. Branched mycelium observed at 12 hr incuba- FIG. 4. Growth initiation by germ-tube formation attion. Phase contrast. X 990. 6 hr incubation. Phase contrast. X 990.
FIG. 2. Branched mycelium with septation observed FIG. 5. "Spider-like" microcolony at 36 hr incuba-at 24 hr incubation. Gram stain. X 990. tion. X 390.
FIG. 3. Mycelium undergoing fragmentation at 72 hr FIG. 6. Mature smooth colonies displaying a pro-incubation. Phase contrast. X 990. nounced dark central area, observed at 7 to 10 days.
X 13.
342 APPL. MICROBIOL.
s6
ft
7.7 A04.o,.. ...A...w
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
CATALASE-NEGATIVE SOIL BACTERIA
DNA base composition. The DNA base compo-sition (moles per cent guanine plus cytosine of the totalbases) was determined by M. Mandel (M. D. Ander-son Tumor Clinic, University of Texas) by means ofthe CsCl density gradient method (31).
RESULTSOccurrence. Filamentous, catalase-negative
bacteria were isolated regularly from high dilu-tions of 12 out of 26 soils examined. The soilswhich harbored these organisms ranged fromfertile meadow to barren desert soils, withmoisture contents from 0.7 to 14.8%, pH valuesfrom 6.4 to 8.2, and organic matter contentsfrom 2.7 to 6.5 %.The numbers of these orga-misms, per gram of soil were 10- to 100-fold dm'greater than the total of the platable microflora,and their initial appearance in the dilution tubesinvariably required at least 2 weeks.
Cell morphology. These bacteria formed atypical gram-positive, branched mycelium char-acteristic of the family Actinomycetaceae (27)(Fig. 1, 2). This hyphal structure was evident inthe presence of a wide variety of carbon sourcesand both in liquid and on agar media. However,hyphal formation was enhanced by the presenceof fructose, sucrose, or carbohydrates containinga terminal sucrose moiety. After its develop-ment, the mycelium segmented with gram-nega-tive areas visible within the myceliumn (Fig. 2)and then fragmented into coccoid and diph-theroid cells (Fig. 3) which were gram-positive.When cells from older cultures, such as thoseobserved in Fig. 3, were transferred to freshmedia, growth initiation proceeded by the for-mation of a "germ tube" from one or morepoints on the parent cell (Fig. 4). Conidia,spores, and other resting structures were notobserved.
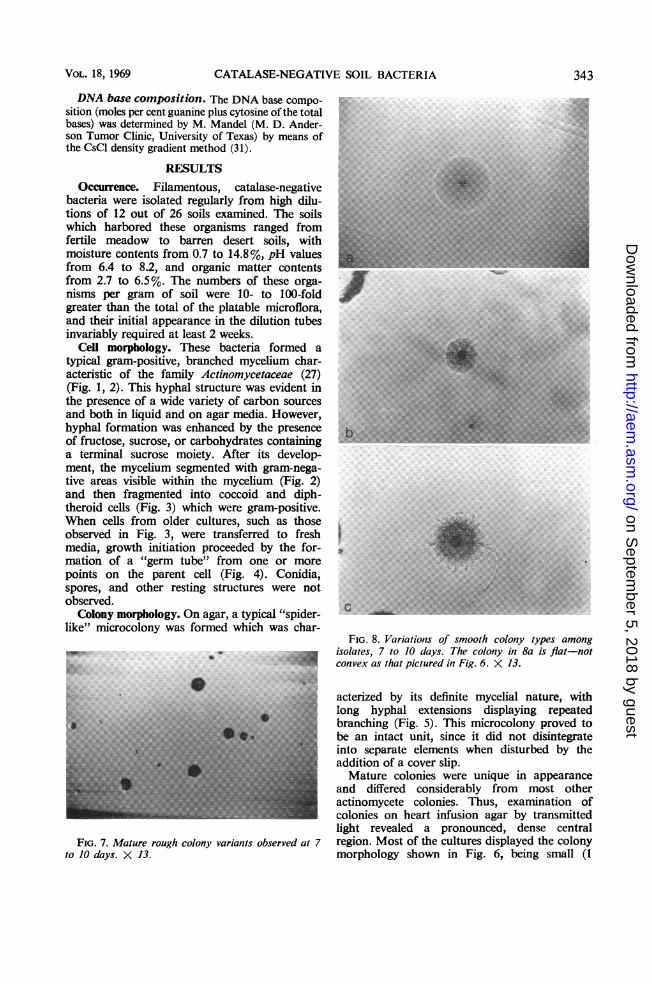
Colony morphology. On agar, a typical "spider-like" microcolony was formed which was char-
FIG. 8. Variations of smooth colony types amongisolates, 7 to J days. The colony iin 8a is flat-not
;appica: convex as that pictured in Fig. 6. X 13.
acterized by its definite mycelial nature, withlong hyphal extensions displaying repeatedbranching (Fig. 5). This microcolony proved tobe an intact unit, since it did not disintegrateinto separate elements when disturbed by theaddition of a cover slip.Mature colonies were unique in appearance
and differed considerably from most otheractinomycete colonies. Thus, examination ofcolonies on heart infusion agar by transmittedlight revealed a pronounced, dense central
FIG. 7. Mature rough colony variants observed at 7 region. Most of the cultures displayed the colonyto 10 days. X 13. morphology shown in Fig. 6, being small (1
VOL. 18, 1969 343
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

GLEDHILL AND CASIDA
mm or less in diameter), smooth, entire, opaque,and convex, and having a dark center which wasusually slightly depressed and which adheredtenaciously to the agar surface. Rough colonyvariants (Fig. 7), often noted among thesecultures, were composed of extensively branchedhyphal elements, and these colonies could beremoved readily from the agar surface as anentire unit. The colony variations noted in Fig. 8also were observed among the isolates.A few strains produced a pale yellow, alcohol-
soluble pigment which was evident only whenquantities of cells were amassed from severalplates. Aerial mycelia, characteristic of the genusNocardia (14), were not observed.
Oxygen requirements and growth character-istics. The isolates were microaerophilic toaerobic. In heart infusion agar shake tubes,growth occurred primarily as a distinct micro-aerophilic band approximately 2.5 mm belowthe agar surface, with comparatively less growthabove and below this region. Increased CO2tension during growth either had no effect orwas slightly inhibitory. Growth in thioglycollatebroth occurred only within the oxidized regionor at the interface between the oxidized andreduced zones.
All strains grew relatively slowly in the mediastudied, requiring from 5 to 7 days to attainmaximal cell density. Growth in liquid media
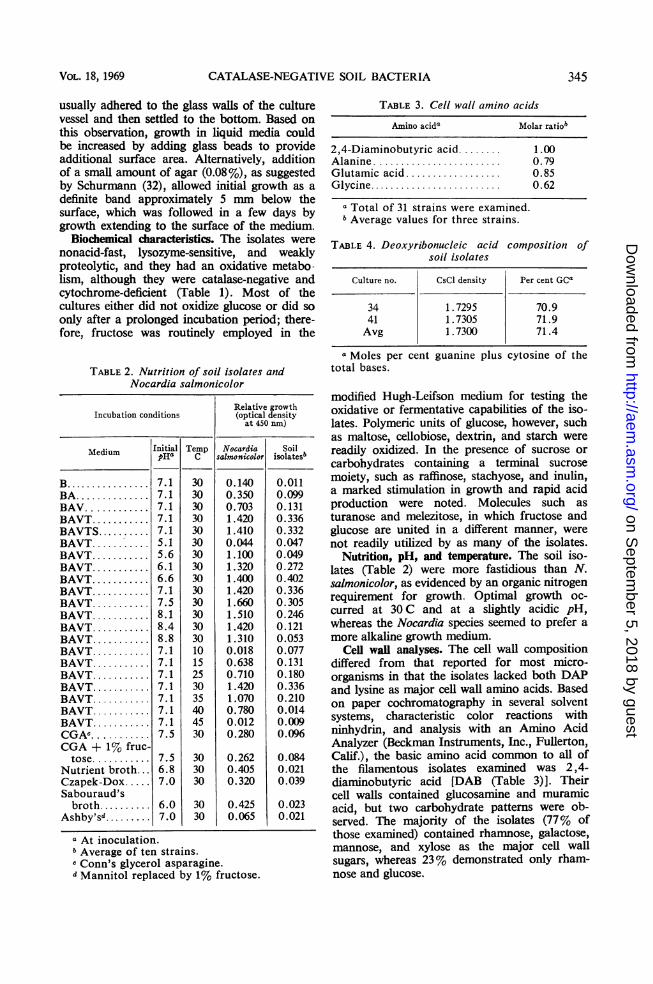
TABLE 1. Biochemical characteristics of 60 soil isolates
Positive PositiveTest strains Test strains
(%) (%)
Catalase activity.......................Benzidine test.........................Oxidase test...........................Nitrate reduction......................Hydrolysis of
Starch...............................Gelatin..............................Tributyrin...........................Casein...............................Esculin..............................
Decomposition ofXanthine............................Tyrosine.............................Urea ................................
Methyl red test........................Voges-Proskauer test..................Greening on blood agar................Lysozyme sensitivity...................Growth at
10 C.................................40 C................................45 C................................
Utilization ofAcetate .............................Propionate ..........................Gluconate...........................Oxalate..............................Citrate ..............................Succinate............................Lactate..............................Fumarate............................Pyruvate ............................a-Ketoglutarate ......................
Litmus milkNo reaction........... ; .
Slight peptonization...................Acid fast..............................Growth in 4% NaCl...................
00015
97180075
0000138598
100180
000005
279798100
4555057
Production ofNH3 from peptone...................NH3 from arginine...................Indole...............................H2S.................................H202 (slight)........................
Sugar fermentation...................Sugar oxidation.......................Acid fromGlycerol ............................Arabinose ...........................Ribose ..............................Xylose..............................Fructose ............................Galactose ...........................Glucose .............................Mannose ............................Rhamnose..........................Salicin ..............................Adonitol ............................Dulcitol.............................Inositol .............................Mannitol ............................Sorbitol .............................Cellobiose..........................Genitobiose .........................Lactose .............................Maltose .............................Melibiose ...........................Sucrose..............................Trehalose ...........................Turanose............................Melezitose..........................Raffinose ............................Stachyose ...........................Dextrin .............................Inulin ...............................Starch...............................
0S0
61120
100
98979895100756877981327S
9010833
109533921228489895958295
aSugar fermentation and oxidation are used in the sense of Hugh and Leifson (15).
344 APPL. MICROBIOL.
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
CATALASE-NEGATIVE SOIL BACTERIA
usually adhered to the glass walls of the culturevessel and then settled to the bottom. Based on
this observation, growth in liquid media couldbe increased by adding glass beads to provideadditional surface area. Alternatively, additionof a small amount of agar (0.08%), as suggestedby Schurmann (32), allowed initial growth as a
definite band approximately 5 mm below thesurface, which was followed in a few days bygrowth extending to the surface of the medium.
Biochemical characteristics. The isolates were
nonacid-fast, lysozyme-sensitive, and weaklyproteolytic, and they had an oxidative metabo-lism, although they were catalase-negative andcytochrome-deficient (Table 1). Most of thecultures either did not oxidize glucose or did so
only after a prolonged incubation period; there-fore, fructose was routinely employed in the
TABLE2. Nutrition of soil isolates andNocardia salmonicolor
Relative growthIncubation conditions (optical density
at 450 nm)
Medium Initial Temp Nocardia SoilpHa C salmonicolor isolatesb
B 7.1 30 0.140 0.011
BA... 7.1 30 0.350 0.099
BAV... 7.1 30 0.703 0.131
BAVT 7.1 30 1.420 0.336
BAVTS. ... 7.1 30 1.410 0.332
BAVT 5.1 30 0.044 0.047
BAVT 5.6 30 1.100 0.049
BAVT 6.1 30 1.320 0.272
BAVT 6.6 30 1.400 0.402
BAVT 7.1 30 1.420 0.336
BAVT 7.5 30 1.660 0.305
BAVT 8.1 30 1.510 0.246
BAVT 8.4 30 1.420 0.121
BAVT 8.8 30 1.310 0.053
BAVT 7.1 10 0.018 0.077
BAVT 7.1 15 0.638 0.131
BAVT 7.1 25 0.710 0.180
BAVT 7.1 30 1.420 0.336
BAVT 7.1 35 1.070 0.210
BAVT 7.1 40 0.780 0.014
BAVT 7.1 45 0.012 0.009
CGAC.. 7.5 30 0.280 0.096
CGA + 1% fruc-tose ........... 7.5 30 0.262 0.084
Nutrient broth... 6.8 30 0.405 0.021Czapek-Dox.. 7.0 30 0.320 0.039Sabouraud'sbroth 6.0 30 0.425 0.023
Ashby'sd......... 7.0 30 0.065 0.021
a At inoculation.b Average of ten strains.c Conn's glycerol asparagine.d Mannitol replaced by 1% fructose.
TABLE 3. Cell wall amino acids
Amino acida Molar ratiob
2,4-Diaminobutyric acid........ 1.00Alanine ........................ 0.79Glutamic acid .................. O.85Glycine......................... 0.62
a Total of 31 strains were examined.bAverage values for three strains.
TABLE 4. Deoxyribonucleic acid compositionsoil isolates
of
Culture no. CsCl density Per cent GCa
34 1.7295 70.941 1.7305 71.9Avg 1.7300 71.4
a Moles per cent guanine plus cytosine of thetotal bases.
modified Hugh-Leifson medium for testing theoxidative or fermentative capabilities of the iso-lates. Polymeric units of glucose, however, suchas maltose, cellobiose, dextrin, and starch werereadily oxidized. In the presence of sucrose orcarbohydrates containing a terminal sucrosemoiety, such as raffinose, stachyose, and inulin,a marked stimulation in growth and rapid acidproduction were noted. Molecules such asturanose and melezitose, in which fructose andglucose are united in a different manner, werenot readily utilized by as many of the isolates.
Nutrition, pH, and temperature. The soil iso-lates (Table 2) were more fastidious than N.salmonicolor, as evidenced by an organic nitrogenrequirement for growth. Optimal growth oc-curred at 30 C and at a slightly acidic pH,whereas the Nocardia species seemed to prefer amore alkaline growth medium.
Cell wail analyses. The cell wall compositiondiffered from that reported for most micro-organisms in that the isolates lacked both DAPand lysine as major cell wall amino acids. Basedon paper cochromatography in several solventsystems, characteristic color reactions withninhydrin, and analysis with an Amino AcidAnalyzer (Beckman Instruments, Inc., Fullerton,Calif.), the basic amino acid common to all ofthe filamentous isolates examined was 2,4-diaminobutyric acid [DAB (Table 3)]. Theircell walls contained glucosamine and muramicacid, but two carbohydrate patterns were ob-served. The majority of the isolates (77% ofthose examined) contained rhamnose, galactose,mannose, and xylose as the major cell wallsugars, whereas 23% demonstrated only rham-nose and glucose.
VOL. 18,1969 345
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

GLEDHILL AND CASIDA
Serology. Twenty-four strains showed noserological cross-reactivity with sera from thegenera Actinomyces, Rothia, Bacterionema, andRamibacterium. Two strains displayed a weakpositive reaction with anaerobic diphtheroidsera, but this was interpreted as being nonspecific.Immunological relationships to the genusNocardia were not studied since serology is notroutinely employed in their taxonomy.
Pathogenicity. The isolates were not patho-genic for mice. All 28 animals appeared healthyafter 3 weeks, and mice sacrificed at this timeshowed no significant pathology. Multiple lesionsand abscesses on internal organs, as have beennoted for certain Actinomyces species (5), werenot observed.
Nucleic acid composition. Based on two repre-sentative strains, the average deoxyribonucleicacid base composition of our isolates was foundto be 71.4% guanine plus cytosine (Table 4).This value agrees well with published reports(17, 25, 36, 37) for various Nocardia and Strepto-myces species.Proposed taxonomy. Based on the character-
istics described in this report, we believe thatour soil isolates should be included in the orderActinomycetales, but that they differ significantlyfrom all other genera within this order. There-fore, we propose that a new genus, Agromyces,be established for these organisms. The char-acterization of this genus would be as follows.Agromyces, gen. n. Gledhill and Casida,
1969. Gr. noun agros, field or soil; Gr. nounmyces, fungus; M. L. noun Agromyces soil-fungus.
Initial growth on agar as microcolonies com-posed of branched, filamentous elements (1,um or less in diameter) which subsequentlyundergo septation and fragmentation to yieldcoccoid and diphtheroid cells. Gram-positive,with segments of the mycelium becoming gram-negative during fragmentation. Aerial myceliumand spores not produced; nonacid-fast, non-motile. Pigmentation not evident during agaror broth growth. Microaerophilic to aerobic,poor or no growth anaerobically. CO2 not re-quired for growth. Catalase-negative, benzidine-negative, oxidase-negative. Organic nitrogenrequired for growth. Weakly proteolytic. Sugarsoxidized without gas production. Cell wallscontain DAB, alanine, glutamic acid, glycine,and rhamnose as major components. Lyso-zyme-sensitive. Optimal growth at 30 C.Agromyces ramosus sp. n. Gledhill and Casida,
1969. L. adj. ramosus, much-branched.Mature colonies small, opaque, entire, smooth,
and convex with a pronounced dense, dark,
central area adhering to the agar surface. Roughcolony variants occur. Growth in broth poor,adhering to glass surfaces and settling to thebottom of the culture vessel. Acid from glycerol,arabinose, ribose, xylose, glucose, fructose,mannose, galactose, mannitol, sucrose, maltose,cellobiose, rhamnose, raffinose, stachyose, inulin,starch, and dextrin. Nitrates not reduced. Cellwall sugars consist of rhamnose, galactose,xylose, and mannose. Hydrogen sulfide produced.Indole not produced. Methyl red and Voges-Proskauer tests negative. Starch and esculinhydrolyzed. Gelatin, casein, and tributyrin nothydrolyzed. Growth in the presence of 4%NaCl. Sodium lactate, succinate, fumarate,pyruvate, and a-ketoglutarate utilized forgrowth. Ammonia not produced from arginine.Xanthine and tyrosine not decomposed. Guanineplus cytosine 71% of the total DNA bases.Habitat: soil. Type species of the genus Agro-myces. The type culture was deposited at theAmerican Type Culture Collection as ATCC25173.
DISCUSSIONThe occurrence of branched, filamentous,
catalase-negative, microaerophilic bacteria as anumerically predominant soil population wasdemonstrated. Based on various major char-acteristics (Table 5), a new genus, Agromyces,in the order Actinomycetales is proposed forthese organisms.They demonstrate several features which are
of diagnostic value in their identification. Theyhave, basically, an oxidative metabolism, al-though they are catalase-negative and cyto-chrome-deficient (benzidine and oxidase tests);they grow in the presence of oxygen without theproduction of hydrogen peroxide. Since in-ducible catalase systems may function in certainorganisms (7) under special conditions (3), at-tempts were made to demonstrate catalase pro-duction by Agromyces isolates grown undervarious aerobic and anaerobic seals and in thepresence of various concentrations of hemin.These procedures, however, failed to inducecatalase formation in Agromyces.Colony morphology is useful for identification
of Agromyces on initial isolation. Althoughtheir small, mycelical microcolonies are typicalof several genera within the Actinomycetales,the "fried-egg" appearance of mature colonies(after 4 days of incubation), which resemblesslightly the colonial appearance of certainActinomyces bovis strains on primary isolation(29), is somewhat unusual. After several monthsof laboratory cultivation, however, the colony
346 APPL. MICROBIOL.
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
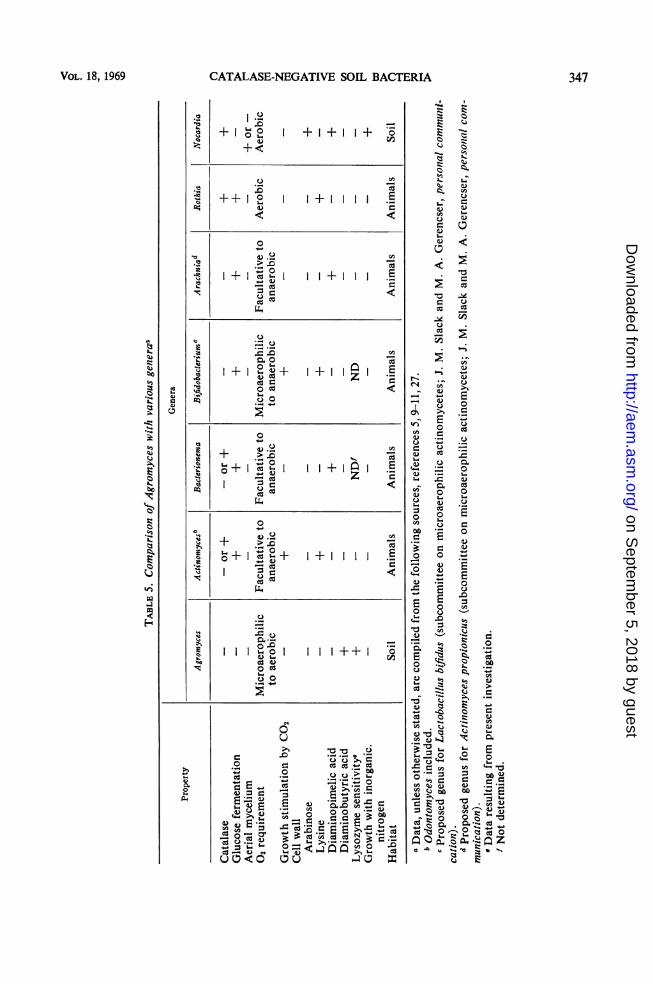
CATALASE-NEGATIVE SOIL BACTERIA
.0+Io o+
V+1+1 1+
"o ++ 2 1+111 E
0
e~ I-+ I
%1 ._5.
U ~~~~~~~~~~cia,+
1e I+ ° + I l .ci 1 1 50+z *
0- o E
0" O ,+IU .
.̂ Y0Cv+~+111
0.
C4)I-0._
I I I o. O Iod w
CZ._
0
co
S =
4-D0
wEQw w 5 .iw (A e
UO<0
I I I ++ I
ac
0
.0. -A
=3
o;:I
347VOL. 18, 1969
5-.
b4
boC15
A.
;Sh
.CqJ-.
~cl
;eIN
5. 5-
s 4)* U
4)
cCci
* 4)
4 )
4)
4)
E .^
-) 0
._4
o> 04) U
- *~
8 O
4)
8
8._
-0
30 0_ *_
0 0
1-~
C4)U
-4)
_,
00
C)C:0w
4)
o;,C.)uS0
U
_o
"0
40)
_
Eo
Cc)
c)
:b
2P4
60
cd,
4_
.4)-
._
4)0E
S
1._
0.
0'-_0 _
r',4)
4 4)4- 4-0
. 5C5
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

GLEDHILL AND CASIDA
morphology of many of the isolates underwent achange. Although originally isolated in thesmooth phase, they became mixtures of bothsmooth and rough forms. Parameters affectingcolony dissociation were not studied.For the genus Agromyces, as well as for other
related genera, cell wall composition is assigneda prime role in taxonomy. The occurrence ofDAB in the cell wall of Agromyces is uniquewithin the order Actinomycetales, and for othermicrobial orders has been reported only for oneplant-pathogenic corynebacterium (26). Thepresence of xylose in most strains of Agromycesis also interesting, since within the Actinomy-cetales this sugar has been reported only in thefamily Actinoplanaceae (34).The genus Agromyces cannot at present be
placed with certainty within a family of theActinomycetales, since rearrangements of thefamilies Actinomycetaceae and Streptomycetaceaeare currently in progress. For example, it is notknown whether the nocardiae will be included in anew family (22) or will be placed within thefamily Streptomycetaceae (27). The familyposition of our isolates presents an additionalproblem, since they would appear to be a transi-tional form possessing various characteristics incommon with both Actinomyces and Nocardia.If the oxidative and fermentative types should beseparated into different families, the genusAgromyces would have to be placed with No-cardia. This, however, would necessitate modifi-cation of the family to include catalase-nega-tive, microaerophilic organisms which lack DAPin their cell walls, characteristics not possessedby any nocardia or streptomycete. Alternatively,placement of Agromyces within the Actino-mycetaceae would present problems similar tothose caused by the inclusion of Nocardia inthis family (22, 27).The above discussion reiterates some of the
taxonomic problems within the Actinomycetales.The proposed genus Agromyces should notlend confusion to the situation; on the contrary,it would aid in clarification, particularly if itis realized that there are forms intermediaryto the nocardiae and streptomycetes (2, 13).Members of the genus Agromyces comprise amajor population of many soils and, hopefully,after this report other workers will devise andemploy suitable isolation techniques to locatefurther soil groups linking Actinomyces andNocardia. We have isolated a few such repre-sentatives (in addition to those described in thepresent report), but for lack of sufficient numbersof strains a detailed study has not yet beenundertaken.
LiTERATURE CITED
1. Bailey, W. R., and E. G. Scott. 1966. Diagnostic microbiology.p. 329, 2nd ed. C. V. Mosby, Co., St. Louis.
2. Becker, B., M. P. Lechevalier, and H. A. Lechevalier. 1965.Chemical composition of cell-wall preparations from strainsof various form-genera of aerobic actinomycetes. Appl.Microbiol. 13:236-243.
3. Biberstein, E. L., and M. Gills. 1961. Catalase activity ofHaemophilus species grown with graded amounts of hemin.J. Bacteriol. 81:380-384.
4. Boone, C. J., and L. Pine. 1968. Rapid method for characteri-zation of actinomycetes by cell wall composition. Appl.Microbiol. 16:279-284.
5. Buchanan, B. B., and L. Pine. 1962. Characterization of apropionic acid producing actinomycete, Actinomycespropionicus, sp. nov. J. Gen. Microbiol. 28:305-323.
6. Casida, L. E., Jr. 1965. Abundant microorganism in soil.Appl. Microbiol. 13:327-334.
7. Clayton, R. K. 1960. Protein synthesis in the induced forma-tion of catalase in Rhodopseudomonas spheroides. J. Biol.Chem. 235:405-407.
8. Deibel, R. H., and J. B. Evans. 1960. Modified benzidine testfor the detection of cytochrome-containing respiratorysystems in microorganisms. J. Bacteriol. 79:356-360.
9. Georg, L. K., and J. M. Brown. 1967. Rothia, Gen. Nov. Anaerobic genus of the family Actlnomycetaceae. Int. J. Syst.Bacteriol. 17:79-88.
10. Gerencser, M. A., and J. M. Slack. 1967. Isolation and char-acterization of Actinomyces propionicus. J. Bacteriol. 94:109-115.
11. Gilmour, M. N., A. Howell, Jr., and B. G. Bibby. 1961. Theclassification of organisms termed Leptotrichia (Leptothrix)buccalis. L. Review of the literature and proposed separationinto Leptotrichia buccalis Trevisan, 1879 and Bacterionemagen. nov., B. matruchott (Mendel, 1919) comb. nov. Bac-teriol. Rev. 25:131-141.
12. Gledhill, W. E., and L. E. Casida, Jr. 1969. Predominantcatalase-negative soil bacteria. I. Streptococcal populationindigenous to soil. Appl. Microbiol. 17:208-213.
13. Gordon, R. E. 1966. Some strains in search of a genus-Corynebacterium, Mycobacterium, Nocardia or what? J.Gen. Microbiol. 43:329-343.
14. Gordon, R. E., and M. M. Smith. 1955. Proposed group ofcharacters for the separation ofStreptomyces and Nocardia.J. Bacteriol. 69:147-150.
15. Hugh, R., and E. Leifson. 1953. The taxonomic significanceof fermentative versus oxidative metabolism of carbohy-drates by various gram negative bacteria. J. Bacteriol. 66:24-26.
16. Hvid-Hansen, N. 1951. Anaerobic actinomycetes (Actino-myces Israelil) in ground-water. Acta Pathol. Microbiol.Scand. 29:335-338.
17. Jones, L. A., and S. G. Bradley. 1964. Phenetic classificationof actinomycetes. Develop. Ind. Microbiol. 5:267-272.
18. Kalakutskii, L. V. 1960. Study of anaerobic proactinomycetes.I. Isolation of pure cultures from nature. Microbiology29:59-63.
19. Kay, R. E., D. C. Harris, and C. Entenman. 1956. Quantifica-tion of the ninhydrin color reaction as applied to paperchromatography. Arch. Biochem. Biophys. 63:14-25.
20. Kovacs, N. 1956. Identification of Pseudomonas pyocyaneaby the oxidase reaction. Nature 178:703.
21. Lechevalier, H. 1964. The Actinomycetes, p. 230-252. InH. Heukelekian and N. C. Dondero (ed.), Principles andapplications in aquatic microbiology. John Wiley & Sons,Inc., New York.
22. Lechevalier, H., and M. P. Lechevalier. 1965. Classificationdes actinomycetes aerobies basee sur leur morphologle etleur composition chimique. Ann. Inst. Pasteur 108:662-673.
23. Lochhead, A. G., and F. B. Chase. 1943. Quantitative studies
348 APPL. MICROBIOL.
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

CATALASE-NEGATIVE SOIL BACTERIA
of soil microorganisms: V. Nutritional requirements of thepredominant bacterial flora. Soil ScL 55:185-195.
24. Mandelstam, J., and H. J. Rogers. 1959. The incorporation ofamino acids into the cell-wall mucopeptide of staphylococciand the effect of antibiotics on the process. Biochem. J. 72:654-662.
25. Marmur, J., S. Falkow, and M. MandeL 1963. New appro-
aches to bacterial taxonomy. Ann. Rev. Microbiol. 17:329-372.
26. Perkins, H. R., and C. S. Cummins. 1964. Chemical structureof bacterial cell walls. Ornithine and 2,4-diaminobutyricacid as components of the cell walls of plant pathogeniccorynebacteria. Nature 201:1105-1107.
27. Pine, L., and L Georg. 1965. The classification and phylo-genetic relationships of the Actinomycetakes. Int. Bull.Bacteriol. Nomenc. Taxon. 15:143-163.
28. Pine, L., and A. Howell, Jr. 1956. Comparison ofphysiologicaland biochemical characters of Actinomyces spp. with thoseof Lactobacillus befdui. J. Gen. MicrobioL 15:428-445.
29. Pine, L., A. Howell, Jr., and S. J. Watson. 1960. Studies ofthe morphological, physiological, and biochemical char-acters of Acilnomyces boyls. J. Gen. Microbiol. 23:403-424.
30. Rosebury, T. 1944. The parasitic actinomycetes and otherfilamentous microorganisms of the mouth. A review of theircharacteristics and relationships, of the bacteriology of
actinomycosis, and of salivary calculus in man. Bacteriol.Rev. 8:189-223.
31. Schildkraut, C L., J. Marmur, and P. Doty. 1962. Determina-tion of the base composition of deoxyribonucleic acid fromits buoyant density in CsCl. J. Mol. Biol. 4:430-443.
32. Schurmann, C. 1967. Growth of myxococci in suspension inliquid media. Appl. Microbiol. 15:971-974.
33. Slack, J. M., A. Winger, and D. W. Moore, Jr. 1961. Sero-logical grouping of Actinomyces by means of fluorescentantibodies. J. Bacteriol. 82:54-65.
34. Szaniszlo, P. J., and H. Gooder. 1967. Cell wall compositionin relation to the taxonomy of some Actinoplanaceae. J.Bacteriol. 94.2037-2047.
35. Taylor, C. B. 1951. The nutritional requirements of the pre-dominant bacterial flora of the soil. Proc. Soc. Appl. Bac-teriol. 14:101-111.
36. Tewfik, M., and S. G. Bradley. 1967. Characterization ofdeoxyribonucleic acids from streptomycetes and nocardiae.J. Bacteriol. 94:1994-2000.
37. Tewfik, E., S. G. Bradley, S. Kuroda, and R. Y. Wu. 1968.Studies on the deoxyribonucleic acids from streptomycetesand nocardiae. Develop. Ind. Microbiol. 9:242-249.
38. Whittenbury, R. 1964. Hydrogen peroxide formation andcatalase activity in lactic acid bacteria. J. Gen. Microbiol35:13-26.
VOL. 18, 1969 349
on Septem
ber 5, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from