in vitro and in vivo enzyme studies of polyhemoglobin ... · in vitro and in vivo enzyme studies of...
TRANSCRIPT

In Vitro and In Vivo Enzyme Studiesof Polyhemoglobin-Tyrosinase
B. Yu, T.M.S. Chang
Artificial Cells and Organs Research Center, Faculty of Medicine, McGillUniversity, 3655 Drummond Street, Montreal, Quebec, Canada H3G 1Y6;telephone: 514-398-3512; fax: 514-398-4983; e-mail: [email protected];website: www.artcell.mcgill.ca
Received 29 October 2003; accepted 4 February 2004
Published online 10 May 2004 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/bit.20084
Abstract: Melanoma is now the fifth most common typeof cancer in North America. At present, there is no optimaltreatment for this cancer. However, the lowering of thetyrosine level can inhibit the growth of melanoma. Unfor-tunately, this diet restriction cannot be humanly toleratedand causes vomiting, nausea, and severe body weight loss.To prevent these problems,weare studying anewapproachinvolving the preparation intermolecularly crosslinked he-moglobin and tyrosinase for intravenous injection. In thisarticle we describe the method of preparation andthe structural and functional properties of polyhemoglobin-tyrosinase. We evaluate the effects of varying glutaral-dehyde ratio, crosslinking time, and enzyme concentrationon the enzyme activity of polyhemoglobin-tyrosinase.We also optimize the molecular weight distribution ofpolyhemoglobin-tyrosinase. The stability of polyhemoglo-bin-tyrosinase at 37jC ismuchmore stablewhen comparedto noncrosslinked tyrosinase solution. Animal studies showthat a higher degree of polymerization correlates with alonger circulation time of polyhemoglobin-tyrosinase,and the optimal crosslinking time is 24 hours. One intra-venous injection of polyhemoglobin-tyrosinase lowers theplasma tyrosine to about 10% of its original level withinone hour. B 2004Wiley Periodicals, Inc.
Keywords: melanoma; tyrosine; polyhemoglobin; polyhe-moglobin-tyrosinase; crosslinking
INTRODUCTION
Melanoma, a fatal skin cancer in its many forms is a com-
mon tumor that accounts for an incidence of around 10%
of all malignancies. Melanoma is most commonly found on
the skin, but 10% arise in the eye (Borden, 2002).
The most important characteristic of differentiation in
melanocyte is the presence of melanin pigment. Melanin
pigment is a heterogeneous biopolymer formed from var-
ious intermediate products. Tyrosinase (EC 1.14.18.1) is
the rate-limiting enzyme in melanin synthesis. This enzyme
converts L-tyrosine as its substrate to L-dopa, and then fur-
ther oxidizes L-dopa to L-dopaquinone, and finally to me-
lanin by different metabolic pathways (Jimbow et al., 1993;
Letellier et al., 1999; Potterf and Hearing, 1998). Under
normal conditions, proliferation and differentiation are re-
gulated by specific growth factors and differentiation
factors, respectively. In neoplastic cells, over-accumulation
of proliferation factors inhibits the differentiation program
of the target cells. Thus, malignant melanoma can be con-
sidered as a disorder of cell proliferation (Li et al., 1997).
However, a number of substances, such as retinoic acid,
corticosteroids, prednisolone, some prostaglandins, phorbol
esters, DMSO, and cyclic AMP, have been reported to in-
duce the differentiation program (Frankenberger et al., 2001;
Hartmann et al., 1997; Lotan, 2003). Bhatnagar et al. (1998)
found that citrate stimulated melanin synthesis and retarded
cellular proliferation since citrate is an activator of cresolase
activity of tyrosinase. Other investigators are looking into
the effects of the use of phenylalanine- and tyrosine-
restricted diets to inhibit the growth and metastasis of
B16BL6 murine melanoma—a malignant melanoma re-
quires higher cellular concentration of tyrosine for its growth
than that for normal melanocytes (Fu et al., 1999).
We introduce in this article the concept of chemically
crosslinking tyrosinase with hemoglobin (polyhemoglobin-
tyrosinase) using a dialdehyde agent, glutaraldehyde. This
new approach can function simultaneously in lowering
systemic tyrosine and carries more oxygen to the melanoma
to increase the efficacy of radiation therapy. Our hypothesis
is that in the form of polyhemoglobin-tyrosinase, tyrosinase
is covered by hemoglobin molecules and therefore has less
immunological properties. Intermolecular polymerization
with hemoglobin also solves the problem of short circulation
time of native tyrosinase.
Here we investigate the structural and functional prop-
erties of this novel polyhemoglobin-tyrosinase (PolyHb-
tyrosinase) in vitro and we also test whether this novel
PolyHb-tyrosinase can be given intravenously to lower the
systemic tyrosine level in animals.
MATERIALS AND METHODS
Materials
Purified bovine hemoglobin was purchased from Biopure
Corporation (Boston, MA). Glutaraldehyde (25%) was
B 2004 Wiley Periodicals, Inc.
Correspondence to: Thomas Ming Swi Chang
Contract grant sponsors: Canadian Institutes of Health Research;
Medical Research Council of Canada

obtained from Polysciences (Warrington, PA). Hemoglobin-
assay kit, molecular distribution standard kit, L-lysine
(monohydrochloride, SigmaUltra > 99%), L-tyrosine
(98% TLC), tyrosinase from mushroom (EC 1.14.18.1,
3000 units/mg stated activity) were purchased from Sigma-
Aldrich (Ontario, Canada). All other reagents were of
analytical grade.
Preparation of PolyHb and PolyHb-Tyrosinase
Reaction mixtures were prepared containing hemoglobin
(10 g/dL), tyrosinase (6000 U/mL) in 0.1M potassium
phosphate buffer, pH 7.6. In PolyHb mixtures, an equiva-
lent volume of buffer replaced the enzyme condition. Prior
to the start of crosslinking, 1.3M lysine was added at a mo-
lar ratio of 7:1 lysine/hemoglobin. Crosslinking reaction
was started with the addition of glutaraldehyde (5%) at mo-
lar ratio of 8:1 or 16:1 glutaraldehyde/hemoglobin. Glutaral-
dehyde was added in four equal aliquots over a period of
15 min. After varied hours of crosslinking (dependent upon
the different experiments) with constant stirring under aero-
bic conditions at 4jC, the reaction was stopped with 2.0M
lysine at a molar ratio of 200:1 lysine/hemoglobin. Solu-
tions were dialyzed using molecular porous dialysis mem-
brane (MWCO:12,000–14,000) against a physiological
saline solution overnight and passed through a sterile
0.45 AM filter. Aliquots (500 AL) of the 16:1 crosslinked
preparation were concentrated using 100-KD microcon-
centrators (Amicon, Beverly, MA). Samples were centri-
fuged at 2500g for 55 min at 23jC. Then, retentate was
collected. Hemoglobin concentration was determined at
540 nm by spectrophotometric analysis with the Total He-
moglobin Kit from Sigma-Aldrich.
Determination of Molecular Weight of PolyHb andPolyHb-Tyrosinase
Molecular weight distribution analysis was performed
using size-exclusion gel filtration chromatography on a Se-
phadex G-200 column (1.6 cm � 70 cm, Vtotal = 102 mL)
equilibrated with 0.1M Tris HCl, pH 7.5. Samples (1 mL
of 10 � dilution) were passed through the column at a rate
of 12 mL/h. The elutioning fractions were collected every
15 min for a period of 8 h. The elution profiles were moni-
tored at 280 nm.
Determination of Tyrosinase Activity
Tyrosinase activity was assessed by measuring the forma-
tion of enzymatic products at 300 nm (Duckworth and
Coleman, 1970). All other substances present in the reac-
tion mixture, i.e., tyrosine, tyrosinase, and hemoglobin had
no significant adsorption at this wavelength. All reactions
were carried out at 23jC. The adsorbance at 300 nm was
followed continuously for 8–14 min using Perkin Elmer
Lambda 4B spectrophotometer, and changes in O.D./min
were used to analyze the activity of the enzyme.
Determination of Tyrosine Concentration inRat’s Plasma
Tyrosine concentration in plasma was analyzed by fluoro-
metric method using Perkin-Elmer Luminescence Spectro-
meter LS50B (Waalkes and Udenferiend, 1957).
Animal Studies
Fasted male Sprague-Dawley rats (245–260 g) were ob-
tained from Charles River Canada (St. Constant, QC,
Canada). They were kept in a controlled 12-h light/dark
environment with food and water ad libitum. All rats were
acclimatized for at least 3 days prior to use. All animal
experiments were performed according to the regulations of
McGill University on animal care. Animals were anesthe-
tized with intraperitoneal injection of 65 mg/kg pentobarbi-
tal (Somnotol, Decton Dickinson, NJ). Body temperature
was maintained by a warming blanket. Incisions were care-
fully performed in one side of hindlimbs below the ingui-
nal ligament, and femoral vessels were carefully isolated.
Polyethylene cannulae were inserted and secured distal to
the superficial epigastric branches in the femoral veins
(PE-10, PE-50 Clay Adams). Proper vessel access was tested
with a small volume injection of heparinized saline (50 IU/
mL). Blood samples were taken from each group at the be-
ginning, then samples were injected through the femoral
vein. The femoral artery cannulae was connected to the ve-
nous cannulae for blood to circulate thoroughly for a short
interval. Then, blood samples were taken from the femoral
artery at different time intervals. The plasma in each blood
sample was separated from the blood, placed in a 1.5-mL
plastic tube, and then stored at � 80jC until analyzed.
RESULTS
Molecular Weight Distribution of PolyHb andPolyHb-Tyrosinase
To determine the degree of polymerization, samples were
analyzed by gel filtration chromatography using a Sephadex
G-200 1.6 cm � 70 cm column at different crosslinking
times from 3.5 h to 48 h. Figure 1 and Table I show the
increase in the higher molecular weight peaks with an in-
crease in the time of crosslinking. Our purpose in this anal-
ysis was to prepare samples with minimal amounts of small
molecular weight molecules since a large amount of
small molecular weight products can be toxic. As a result,
our analysis concentrates on the study of the proportion of
small molecular weight molecules compared to the larger
molecules. To allow us to do this more accurately, we
used a size-exclusion column with molecular exclusion
of 700,000 daltons (Da). Thus, molecules larger than
700,000 Da would not be separated from the exclusion
peak. Hence, the exclusion peak only gives the proportion
of the larger molecules but not the distribution of mole-
cules larger than 700,000 Da. The results show that
836 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 86, NO. 7, JUNE 30, 2004

increasing the reaction time to 24 h significantly decreases
the proportion of the smaller molecules. However, reaction
times beyond 24 h (e.g., 48 h) did not have any further sig-
nificant effects. Thus, we arrive at a reaction time of 24 h for
the rest of the in vitro and in vivo studies. There was no
significant difference in the molecular weight distribution
between PolyHb and PolyHb-tyrosinase (Fig. 1). This was
because the ratio of hemoglobin to tyrosinase was 100:2.
Therefore, the added tyrosinase was not expected to sig-
nificantly change the molecular weight distribution after
being crosslinked with hemoglobin. Our analysis allowed us
to select the 24-h crosslinking reaction at a glutaraldehyde
molar ratio of 16:1 for animal studies. There was no tyro-
sinase activity in the PolyHb samples.
The Effect of Glutaraldehyde: Hemoglobin Ratio andCrosslinking Time on Tyrosinase Activity
This experiment was designed to find the effect on tyrosi-
nase activity with the addition of glutaraldehyde at different
molar ratios of 8:1 and 16:1 glutaraldehyde/hemoglobin.
We considered hemoglobin with tyrosinase activity without
the addition of glutaraldehyde as 100% original activity.
After crosslinking, 99% of tyrosinase activity remained in
PolyHb-tyrosinase when crosslinked using a glutaraldehyde
molar ratio of 8:1. Ninety-five percent of tyrosinase activity
remained for PolyHb-tyrosinase crosslinked using a glutar-
aldehyde molar ratio of 16:1. Since there was no significant
difference in enzyme activity when using 8:1 or 16:1 molar
ratio of glutaraldehyde : hemoglobin, we used the glutar-
aldehyde molar ratio of 16:1 in the following studies. In
our screening test, we studied different ratios of glutaralde-
hyde : hemoglobin including 8:1, 12:1, 14:1, 16:1, and 17:1.
The lower ratio of glutaraldehyde, i.e., 8:1, 12:1, and 14:1
yielded too many smaller molecules that would be toxic
to the kidney. However, a larger ratio of 17:1, resulted in
gelling of the solution. The ratio of 16:1 gave the least
small molecular weight products. We therefore use this
ratio to study the effects of different crosslinking time on
enzyme activities. We did not study in detail the enzyme
activity using the other ratios, since the resulting products
cannot be used. In our study, we found that the cross-
linking ratio of 16:1 did not significantly change the tyro-
sinase activity at different crosslinking time (Fig. 2).
To determine the effect of crosslinking time on tyrosinase
activity, we crosslinked hemoglobin with tyrosinase from
3.5 h up to 48 h, and measured tyrosinase activity. We con-
sidered tyrosinase activity before crosslinking as 100% of
original activity, other data are expressed as the percentage
of the original activity. No significant difference in tyro-
sinase activity was found among these periods (Fig. 2). This
result shows that longer crosslinking time does not decrease
enzyme activity significantly. Therefore, the crosslinking
reaction as described in the Method section has no adverse
effect on the tyrosinase activity.
To further investigate the effect of crosslinking reaction
on enzyme activity, we compared enzyme activity before
and after crosslinking. Our results show that there was no
significant change in enzyme activity before or after cross-
linking (Fig. 3). Before crosslinking, tyrosinase decreased
the tyrosine level to 2.63 F 0.11 mg/dL. After cross-
linking the enzyme reduced the tyrosine level to 2.75 F0.35 mg/dL, and for the noncrosslinked group, the tyrosine
level dropped off to 2.65 F 0.38 mg/dL. There was no
significant difference after crosslinking when compared
to the control. The reaction is a covalent crosslinking reac-
tion between the amino groups of enzyme molecules and the
amino groups of the hemoglobin molecules with the
aldehyde groups of glutaraldehyde. If this reaction is not
Figure 1. Typical elution profiles of 3.5-, 24-, 48-h crosslinked PolyHb or
PolyHb-tyrosinase (1 mL sample) were obtained by running on a Sephadex
G-200 1.6 cm � 70 cm column, VT = 102 mL, equilibrated with 0.1M Tris
HCl, pH 7.5, and eluted at 12 mL/h.
Table I. Percentage of area under molecular weight distribution profiles.
Crosslinking
time (hours) Samples
Percentages of molecular weight distribution (KD)
> 400 kDa Between 100 kDa–400 kDa < 100 kDa
3.5 h PolyHb 39% 31% 30%
PolyHb-tyrosinase 39% 31% 30%
24 h PolyHb 74% 14% 12%
PolyHb-tyrosinase 74% 14% 12%
48 h PolyHb 77% 12% 11%
PolyHb-tyrosinase 77% 12% 11%
YU AND CHANG: ENZYME STUDIES OF POLYHEMOGLOBIN-TYROSINASE 837

carried out under optimized conditions, it results in the de-
activation of much of the enzyme activities. This is why we
carried out our reaction very slowly—over 24 hours—using
lysine during the reaction as well. Statistical analysis
confirms there was no inactivation of enzyme activity.
This further confirms that the crosslinking reaction under
optimal reaction conditions described here does not affect
enzyme activity significantly.
The Effect of Enzyme Concentration onTyrosinase Activity
We then tested the enzyme activity at different enzyme
concentrations after a crosslinking of 3.5 h. Crosslinking
hemoglobin added with equal volume of buffer instead of
enzyme solution served as a control. Figure 4 shows the
effects of increasing the concentration of PolyHb-tyrosinase
on the rate of conversion of tyrosine. The higher the con-
centration of tyrosinase, the faster the enzyme conver-
sion rate.
The Effect of Increase in Time of Crosslinking onDegree of Polymerization of Tyrosinase to PolyHb
We also analyzed whether all the tyrosinase crosslinked to
PolyHb.We crosslinked hemoglobinwith tyrosinase at 3.5 h,
24 h, 30 h, and 48 h. Then, we took a 1 mL of PolyHb-
tyrosinase sample at different time intervals, ran each sample
through Sephadex G-200 1.6 cm � 70 cm column, equi-
librated with 0.1M Tris HCl, and eluted at 12 mL/h. Figure 5
shows when crosslinked for only 3.5 h, a significant amount
of the tyrosinase remained in free solution and not cross-
linked. As the molecular weight of tyrosinase is about
128 kDa, in free form its peak would appear at 70 mL of
elution volume. The longer the time we crosslink, the more
tyrosinase would be crosslinked to PolyHb, and the more
activity would be found in the larger molecular weight
area. When the crosslinking time increased to 24 h, the
uncrosslinked fraction significantly decreased. There was
no significant difference in enzyme activity at the cross-
linking time of 24 h, 30 h, and 48 h. Our results therefore
show that 24 h or more would be needed to optimally
crosslink tyrosinase to hemoglobin during the polymeriza-
tion. In the case of PolyHb-tyrosinase—after crosslinking of
24 h— 79% of the tyrosinase activity was in the molecular
Figure 3. Ability to decrease tyrosine concentration in vitro before
crosslinking, after crosslinking, and in noncrosslinking solution. Group 1:
PolyHb: 2.5 mL Hb + 0.1 mL buffer. Group 2: Before crosslinking: 2.5 mL
Hb + 0.1 mL tyrosinase solution (3000 U). Group 3: After crosslinking:
2.5 mL Hb + 0.1 mL tyrosinase solution (3000 U). Group 4: Noncross-
linking: 2.5 mL Hb + 0.1 mL tyrosinase solution (3000 U), buffer instead of
glutaraldehyde for crosslinking.
Figure 4. Ability to decrease tyrosine concentration in vitro using
PolyHb-tyrosinase. (1) Control: PolyHb (2.5 mL Hb crosslinked with
0.1 mL buffer); (2) PolyHb-tyrosinase: 2.5 mL Hb crosslinked with 3000 U
of tyrosinase (3000U/0.1 mL); (3) PolyHb-tyrosinase: 2.5 mL Hb cross-
linked with 6000 U of tyrosinase (6000U/0.1 mL); (4) PolyHb-tyrosinase:
2.5 mL Hb crosslinked with 9000 U of tyrosinase (9000U/0.1 mL); (5)
PolyHb-tyrosinase: 2.5 mL Hb crosslinked with 12000 U of tyrosinase
(12000U/0.1 mL); (6) PolyHb-tyrosinase: 2.5 mL Hb crosslinked with
15000 U of tyrosinase (15000U/0.1 mL).
Figure 2. PolyHb-tyrosinaseactivity after different crosslinking time from
3.5 h to 48 h.
838 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 86, NO. 7, JUNE 30, 2004

fraction of > 400 kDa, with 12% in the 400 kDa fraction,
and 5% in the 100 kDa fraction (Table II).
The Stability of PolyHb-Tyrosinase at 37jC
To determine whether body temperature is one of the major
factors that affect tyrosinase activity, we tested the enzyme
activity at 37jC for up to 6 h using PolyHb-tyrosinase, and
free tyrosinase solution as the control (Fig. 6). Our results
show that enzyme activity in free tyrosinase solution de-
creases faster than PolyHb-tyrosinase at 37jC. At 6 h, 79%
of the enzyme activity remained in PolyHb-tyrosinase solu-
tion. On the other hand, only 60% of the enzyme activity
was found in free tyrosinase solution after 6 h incubation.
Thus, PolyHb-tyrosinase was stable at body temperature up
to 6 h.
Animal Studies on the Effect of PolyHb-Tyrosinase
The next experiment was designed to determine if PolyHb-
tyrosinase could lower tyrosine levels in rat plasma. We first
did a screening test using increasing dosages and found that
the dosage that gave the lowest systemic tyrosine level was
1 mL. Figure 7 details what happened when we increased
the dosage beyond 1 mL.We injected 1 mL, 2 mL, and 3 mL
of PolyHb-tyrosinase per 250 g body weight per rat,
respectively (Fig. 7). Tyrosine concentration was mea-
sured to investigate if PolyHb-tyrosinase was efficient in
removing tyrosine in plasma. PolyHb concentration was
Figure 5. The activity of tyrosinase separated through Sephadex G-200, and the crosslinking times were 3.5 h, 24 h, 30 h, and 48 h, respectively. Curves
represent the mean of three determinations for each sample.
Table II. Molecular weight distribution and tyrosinase activity of
PolyHb and PolyHb-tyrosinase after 24 h crosslinking.
Molecular weight
(MW)
MW component
(%)
Tyrosinase activity
(%)
PolyHb-tyrosinase sample
> 400 kDa 74 79
100–400 kDa 14 12
< 100 kDa 12 5
PolyHb sample
> 400 kDa 74 0
100–400 kDa 14 0
< 100 kDa 12 0 Figure 6. The stability of tyrosinase in free form and as PolyHb-tyrosinase
at 37jC in vitro.
YU AND CHANG: ENZYME STUDIES OF POLYHEMOGLOBIN-TYROSINASE 839
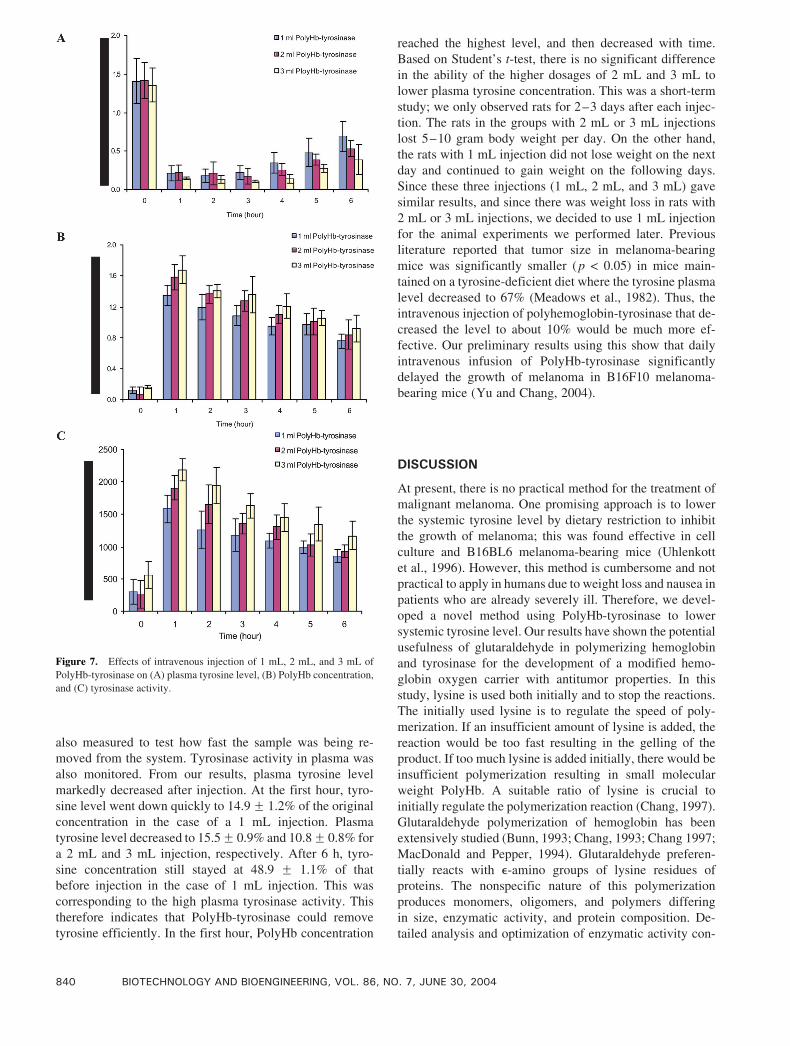
also measured to test how fast the sample was being re-
moved from the system. Tyrosinase activity in plasma was
also monitored. From our results, plasma tyrosine level
markedly decreased after injection. At the first hour, tyro-
sine level went down quickly to 14.9F 1.2% of the original
concentration in the case of a 1 mL injection. Plasma
tyrosine level decreased to 15.5F 0.9% and 10.8F 0.8% for
a 2 mL and 3 mL injection, respectively. After 6 h, tyro-
sine concentration still stayed at 48.9 F 1.1% of that
before injection in the case of 1 mL injection. This was
corresponding to the high plasma tyrosinase activity. This
therefore indicates that PolyHb-tyrosinase could remove
tyrosine efficiently. In the first hour, PolyHb concentration
reached the highest level, and then decreased with time.
Based on Student’s t-test, there is no significant difference
in the ability of the higher dosages of 2 mL and 3 mL to
lower plasma tyrosine concentration. This was a short-term
study; we only observed rats for 2–3 days after each injec-
tion. The rats in the groups with 2 mL or 3 mL injections
lost 5–10 gram body weight per day. On the other hand,
the rats with 1 mL injection did not lose weight on the next
day and continued to gain weight on the following days.
Since these three injections (1 mL, 2 mL, and 3 mL) gave
similar results, and since there was weight loss in rats with
2 mL or 3 mL injections, we decided to use 1 mL injection
for the animal experiments we performed later. Previous
literature reported that tumor size in melanoma-bearing
mice was significantly smaller (p < 0.05) in mice main-
tained on a tyrosine-deficient diet where the tyrosine plasma
level decreased to 67% (Meadows et al., 1982). Thus, the
intravenous injection of polyhemoglobin-tyrosinase that de-
creased the level to about 10% would be much more ef-
fective. Our preliminary results using this show that daily
intravenous infusion of PolyHb-tyrosinase significantly
delayed the growth of melanoma in B16F10 melanoma-
bearing mice (Yu and Chang, 2004).
DISCUSSION
At present, there is no practical method for the treatment of
malignant melanoma. One promising approach is to lower
the systemic tyrosine level by dietary restriction to inhibit
the growth of melanoma; this was found effective in cell
culture and B16BL6 melanoma-bearing mice (Uhlenkott
et al., 1996). However, this method is cumbersome and not
practical to apply in humans due to weight loss and nausea in
patients who are already severely ill. Therefore, we devel-
oped a novel method using PolyHb-tyrosinase to lower
systemic tyrosine level. Our results have shown the potential
usefulness of glutaraldehyde in polymerizing hemoglobin
and tyrosinase for the development of a modified hemo-
globin oxygen carrier with antitumor properties. In this
study, lysine is used both initially and to stop the reactions.
The initially used lysine is to regulate the speed of poly-
merization. If an insufficient amount of lysine is added, the
reaction would be too fast resulting in the gelling of the
product. If too much lysine is added initially, there would be
insufficient polymerization resulting in small molecular
weight PolyHb. A suitable ratio of lysine is crucial to
initially regulate the polymerization reaction (Chang, 1997).
Glutaraldehyde polymerization of hemoglobin has been
extensively studied (Bunn, 1993; Chang, 1993; Chang 1997;
MacDonald and Pepper, 1994). Glutaraldehyde preferen-
tially reacts with e-amino groups of lysine residues of
proteins. The nonspecific nature of this polymerization
produces monomers, oligomers, and polymers differing
in size, enzymatic activity, and protein composition. De-
tailed analysis and optimization of enzymatic activity con-
Figure 7. Effects of intravenous injection of 1 mL, 2 mL, and 3 mL of
PolyHb-tyrosinase on (A) plasma tyrosine level, (B) PolyHb concentration,
and (C) tyrosinase activity.
840 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 86, NO. 7, JUNE 30, 2004

tained in individual molecular weight fractions is also
investigated in the present study.
We first studied the molecular weight distribution of
PolyHb-tyrosinase at different crosslinking hours. We found
that the longer the crosslinking time, the more PolyHb-
tyrosinase we can obtain in high molecular weight. In our
study of properties of PolyHb and PolyHb-tyrosinase, dif-
ferent molar ratios of glutaraldehyde : hemoglobin did not
decrease tyrosinase activity. The crosslinking time from
3.5 h to 48 h had no adverse effect on the tyrosinase activity.
With increased tyrosinase concentration in PolyHb-tyrosi-
nase, higher enzyme activity was observed. To figure out
the enzyme activity of bound and unbound enzyme fraction,
we tested the enzyme activity by passing Sephadex G-200
column and found that longer crosslinking time was
necessary to get high activity of PolyHb-tyrosinase in high
molecular weight. Our results showed that incubation at
37jC, PolyHb-tyrosinase remained at 79% activity after
6 h. This was more stable than free tyrosinase at 37jC. Highconcentrations of hemoglobin will also stabilize enzymes at
body temperature. However, this was not studied because
the high concentration of hemoglobin when injected intra-
venously would become highly diluted in the circulating
blood and thus no longer stabilize the tyrosinase. In a pre-
vious study of the oxygen affinity of free hemoglobin and
PolyHb-tyrosinase, we observed that PolyHb-tyrosinase
possessed oxygen transport characteristics similar to non-
crosslinked hemoglobin. This characteristic of PolyHb is
very important for radiotherapy—as a solution PolyHb-
tyrosinase can easily reach the narrower capillaries of the
melanoma to supply the additional oxygen needed in ra-
diotherapy. Increased circulation half-life of tyrosinase in
the highly polymerized solution suggests that with an in-
crease in time of crosslinking more tyrosinase molecules
crosslink to PolyHb resulting in an increase in retention
time. We speculate that the increased fraction of bound
enzymes have largely retained their activity.
We next did a preliminary animal study of the effect
of an intravenous injection of PolyHb-tyrosinase to lower
the systemic tyrosine level. Higher volumes of PolyHb-
tyrosinase injection yielded the lowest level of systemic
tyrosine, however, the animals dramatically lost body
weight due to severe starvation for tyrosine (Meadows and
Oeser, 1983). We conclude that the optimal preparation
is prepared from a crosslinking time of 24 h using an in-
jection volume of 1 mL of PolyHb-tyrosinase. This way it
decreases the systemic tyrosine level, but also has no ad-
verse effect and can maintain body weight. Therefore, this
preparation of PolyHb-tyrosinase may provide a potential
promising approach to lower systemic tyrosine level in
malignant melanoma.
References
Bhatnagar V, Srirangam A, Abburi R. 1998. In vitro modulation of pro-
liferation and melanization of melanoma cells by citrate. Mol Cell
Biochem 187:57–65.
Borden E. C. (Ed.) 2002. Melanoma: Biologically targeted therapeutics.
New York: Humana Press.
Bunn HF. 1993. The use of hemoglobin as a blood substitute. Am J He-
matol 42:112–117.
Chang TMS. (Ed.) 1993. Blood substitutes and oxygen carriers. New
York: Marcel Dekker.
Chang TMS. 1997. Blood substitutes: Principles, methods, products and
clinical trials, Vol. 1. Basel/New York: Karger Landes Systems.
Duckworth HW, Coleman JE. 1970. Physiochemical and kinetic properties
of mushroom tyrosinase. J Biol Chem 245:1613–1625.
Frankenberger M, Hauck RW, Frankenberger B, Haussinger K, Maier KL,
Heyder J, Ziegler-Heitbrock HW. 2001. All trans-retinoic acid se-
lectively down-regulates matrix metalloproteinase-9 (MMP-9) and up-
regulates tissue inhibitor of metalloproteinase-1 (TIMP-1) in human
bronchoalveolar lavage cells. Mol Med 7:263–270.
Fu YM, Yu ZX, Pelayo BA, Ferrans VJ, Meadows GG. 1999. Focal
adhesion kinase-dependent apoptosis of melanoma induced by tyro-
sine and phenylalanine deficiency. Cancer Res 59:758–765.
Hartmann RR, Rimoldi D, Lejeune FJ, Carrel S. 1997. Cell differentiation
and cell-cycle alterations by tyrosine kinase inhibitors in human me-
lanoma cells. Melanoma Res 7:S27–S33.
Jimbow K, Iwashina T, Alena F, Yamada K, Pankovich J, Umemura T.
1993. Exploitation of pigment biosynthesis pathway as a selective
chemotherapeutic approach for malignant melanoma. J Invest Derma-
tol 100:231S–238S.
Letellier S, Garnier JP, Spy J, Stoitchkov K, Bricon TL, Baccard M, Revol
M, Kerneis Y, Bousquet B. 1999. Development of metastases in ma-
lignant melanoma is associated with an increase in the plasma. Mela-
noma Res 9:389–394.
Li Z, Hromchak R, Bloch A. 1997. Differential expression of proteins
regulating cell cycle progression in growth vs. differentiation. Bio-
chim Biophys Acta 1356:149–159.
Lotan R. 2003. Receptor-independent induction of apoptosis by synthetic
retinoids. J Biol Regul Homeost Agents 17:13–28.
MacDonald SL, Pepper DS. 1994. Hemoglobin polymerization. Methods
Enzymol 231:287–308.
Meadows GG, Pierson HF, Abdallah RM, Desai PR. 1982. Dietary
influence of tyrosine and phenylalanine on the response of B16 mela-
noma to carbiodopa-levodopa methyl ester chemotherapy. Cancer Res
42:3056–3063.
Meadows GG, Oeser DE. 1983. Response of B16 melanoma-bearing mice
to varying dietary levels of phenylalanine and tyrosine. Nutr Rep Int
28:1073–1082.
Potterf BS, Hearing VJ. 1998. Tyrosine transport into melanosomes is
increased following stimulation of melanocyte differentiation. Bio-
chem Biophys Res Commun 248:795–800.
Uhlenkott CE, Huijzer JC, Cardeiro DJ, Elstad CA, Meadows GG. 1996.
Attachment, invasive, chemotaxis, and prteinase expression of B16-
BL6 melanoma cells exhibiting a low metastatic phenotype after ex-
posure to dietary restriction of tyrosine and phenylalanine. Clin Exp
Metastasis 14:125–137.
Waalkes TP, Udenferiend S. 1957. A fluorometric method for the
estimation of tyrosine in plasma and tissues. J Lab Clin Med 11:
733–736.
Yu B, Chang TMS. 2004. In vitro and in vivo effects of polyhemoglobin-
tyrosinase on murine B 16 F 10 melanoma. Melanoma Res (in press).
YU AND CHANG: ENZYME STUDIES OF POLYHEMOGLOBIN-TYROSINASE 841