in vitro oocyte maturation, fertilization and culture ... · in vitro oocyte maturation,...
TRANSCRIPT

www.theriojournal.com
Available online at www.sciencedirect.com
Theriogenology 69 (2008) 349–359
In vitro oocyte maturation, fertilization and culture after ovum
pick-up in an endangered gazelle (Gazella dama mhorr)
F. Berlinguer a, R. Gonzalez b, S. Succu a, A. del Olmo b, J.J. Garde c,G. Espeso d, M. Gomendio b, S. Ledda e, E.R.S. Roldan b,*
a Department of Animal Biology, Universita degli Studi di Sassari, 07100 Sassari, Italyb Reproductive Ecology and Biology Group, Museo Nacional de Ciencias Naturales (CSIC), c/Jose Gutierrez Abascal 2,
28006 Madrid, Spainc Instituto de Investigacion en Recursos Cinegeticos (CSIC-UCLM-JCCM), 02071 Albacete, Spain
d Estacion Experimental de Zonas Aridas (CSIC), 04001 Almerıa, Spaine Institute of General Pathology, Pathological Anatomy, and Veterinary Obstetrics – Surgery Clinic,
Universita degli Studi di Sassari, 07100 Sassari, Italy
Received 22 June 2007; received in revised form 1 October 2007; accepted 2 October 2007
Abstract
The recovery of immature oocytes followed by in vitro maturation, fertilization and culture (IVMFC) allows the rescue of
biological material of great genetic value for the establishment of genetic resource banks of endangered species. Studies exist on
sperm cryopreservation of endangered Mohor gazelle (Gazella dama mhorr), but no work has been carried out yet on oocyte
collection, fertilization and culture in this or related species. The purpose of this study was to develop a protocol for ovarian
stimulation for the recovery of oocytes and subsequent IVMFC in the Mohor gazelle using frozen-thawed spermatozoa. Ovum pick-
up was performed after ovarian stimulation with a total dose of 5.28 mg of ovine FSH. A total of 35 oocytes were recovered from 56
punctured follicles (62%) (N = 6 females). Out of 29 cumulus-oocyte complexes matured in vitro, 3% were found at germinal
vesicle stage, 7% at metaphase I, 21% were degenerated, and 69% advanced to metaphase II. Fertilization and cleavage rates of
matured oocytes were 40 and 30%, respectively. Embryos cleaved in vitro up to the 6–8 cell stage but none progressed to the
blastocyst stage, suggesting the existence of a developmental block and the need to improve culture conditions. Although more
studies are needed to improve hormonal stimulation and oocyte harvesting, as well as IVMFC conditions, this study demonstrates
for the first time the feasibility of in vitro fertilization with frozen-thawed semen of in vitro matured oocytes collected by ovum pick-
up from FSH-stimulated endangered gazelles.
# 2008 Elsevier Inc. All rights reserved.
Keywords: Oocyte; Sperm; Fertilization; Embryo; Gazelle
1. Introduction
Habitat preservation is a priority in biodiversity
conservation, but it is often difficult to avoid habitat
* Corresponding author. Tel.: +34 91 411 1328x1245;
fax: +34 91 564 5078.
E-mail address: [email protected] (E.R.S. Roldan).
0093-691X/$ – see front matter # 2008 Elsevier Inc. All rights reserved.
doi:10.1016/j.theriogenology.2007.10.001
destruction and, in some endangered species, other
causes such as over-hunting are responsible for the
declines in population size. Therefore, complementary
strategies for biodiversity conservation, such as captive
breeding, the establishment of genetic resource banks
(GRBs) and the development of assisted reproductive
techniques (ARTs) are needed. Genetic resource banks
preserve current levels of genetic variability, facilitate
genetic flow between natural and captive populations

F. Berlinguer et al. / Theriogenology 69 (2008) 349–359350
[1–4], and could be regarded as a form of ‘‘life
insurance’’ in case wild populations continue their
decline.
The World Conservation Union has recognized the
potential role that captive breeding may play in
conservation [5] and has recommended that taxa with
low numbers of individuals in the wild should be
considered for captive breeding. However, reproductive
processes may be impaired in captivity as a consequence
of behavioral or genetic incompatibilities, inadequate
diet, disease, or due to the risks inherent to small
population size such as inbreeding depression [6].
Reproductive biotechnologies can improve the repro-
duction of wild animals in captivity, allowing for the
recovery and future use of reproductive biomaterials, and
facilitating the exchange of gametes both within and
between populations thus avoiding the negative effects of
inbreeding and overcoming behavioral incompatibilities
between individuals. Various ARTs have been applied
successfully in several species of wild mammals [7–12].
The Parque de Rescate de la Fauna Sahariana of the
Estacion Experimental de Zonas Aridas (EEZA), in
Almerıa, Spain, was created in 1971 with the objective of
avoiding the extinction of the large ungulates of the
Western Sahara. Mohor gazelle (Gazella dama mhorr
Bennet 1833) is a subspecies of dama gazelle for which a
breeding program has been established. The mhorr
subspecies occupied the western strip of the Sahara desert
and became extinct in the wild in 1968 due to excessive
hunting. The Red List of Threatened Species [13]
categorizes G. dama as ‘‘critically endangered’’ and the
species is included in Appendix I of CITES. The captive
population of Mohor gazelles was founded with two
males and nine females [14], and this conservation
program has been managed carefully to minimize
inbreeding effects. At present the captive population is
distributed among several institutions in Europe and
North America, all derived form the original population
bred in Almerıa [14]. At the end of 2003, the total captive
population of Mohor gazelle consisted of 277 indivi-
duals, with 183 individuals (66.5%) within the European
Breeding Program (EEP), and 92 individuals (33.5%) in
the North American Breeding Program (SSP) [14].
The success of captive breeding programs initiated
with a small founding population relies to a great extent
on avoiding the deleterious effects of inbreeding on
reproduction [15]. There is evidence showing the
deleterious effect of inbreeding on reproductive success
both in male and female Mohor gazelles and in other
related endangered North African gazelles [16].
Inbreeding adversely affects ejaculate quality in
gazelles with relatively high coefficients of inbreeding
[17,18]. Reproductive performance is negatively
affected as the coefficient of inbreeding increases in
Cuvier’s (G. cuvieri), Mohor and dorcas (G. dorcas
neglecta) gazelles [19]. At the EEZA, although efforts
have been aimed at minimizing the levels of inbreeding
in the captive populations, there are several limiting
factors: the population is small in size, effective
population size is even smaller because space restric-
tions do not allow all animals the opportunity to breed,
and some individuals die before they can reproduce. In
addition, moving animals between different populations
is risky and costly. These problems can be overcome by
cryopreserving gametes from most of the individuals so
that they can be used to increase their reproductive
opportunities even beyond death, and to facilitate gene
exchange between populations. Previous work has
focused on the management of natural breeding and on
the development of semen freezing protocols for the
three species to initiate a GRB [16,20,21]. No studies
have been carried out on the collection, maturation and
fertilization of oocytes in endangered gazelle.
The development of procedures in endangered
gazelles for in vitro maturation, fertilization and culture
(IVMFC) would allow the rescue of biomaterials from
females with genetic value for establishing GRBs.
Oocyte collection by laparotomy or laparoscopic pick-
up, followed by IVMFC, has been extensively studied in
domestic ruminants [22,23] and several studies have
been performed in non-domestic ruminant species
including red or sika deer [24,25], and mouflons
[26,27]. Several factors affect the success of follicle
growth stimulation [28,29], and ovum pick-up (OPU)
[30], including the possibility of repeated collections
[31]. In addition, conditions for in vitro maturation and
fertilization have been described [22,23].
The aims of the present study on G. dama mhorr
were to develop a protocol for (1) ovarian stimulation
for the recovery of immature oocytes, (2) in vitro oocyte
maturation, fertilization and embryo culture and (3) to
evaluate the ability of cryopreserved spermatozoa to
fertilize in vitro matured oocytes. We regard the
development of these techniques to be a first step
towards the rescue of female reproductive biomaterials
for the preservation of genetic diversity.
2. Materials and methods
2.1. Reagents
All chemicals used in the present study were
purchased from Sigma Chemical Co. (Madrid, Spain),
unless otherwise indicated.

F. Berlinguer et al. / Theriogenology 69 (2008) 349–359 351
2.2. Animals
All animal procedures were performed in accordance
with the Spanish Animal Protection Regulation,
RD223/1988, which conforms to European Union
Regulation 86/609. Procedures also adhered to the
Guiding Principles for Biomedical Research Involving
Animals, as issued by the Council for the International
Organizations of Medical Sciences, and recommended
by the journal.
The study was carried out in December 2004, using
seven healthy mature (aged >2 years) female Mohor
gazelles from the breeding herd maintained and
managed by the Estacion de Zonas Aridas (EEZA) at
the Parque de Rescate de la Fauna Sahariana (PRFS) in
Almerıa, south-eastern Spain. The PRFS is located at
3685001000N and 282704800W and is 100 m above sea
level. The animals were kept in a group in the same
enclosure without the presence of males or young. The
enclosure was visually opened to the exterior and was
provided with shadow areas. Their diet consisted of
commercial pellets, fresh alfalfa hay and barley grain.
Water and mineral salts were available ad libitum.
2.3. Estrous synchronization and stimulation of
follicular growth
Estrous cycles in the gazelle were synchronized by
insertion of controlled progesterone internal drug
release devices (CIDR, type G, 330 mg progesterone;
InterAg Hamilton, New Zealand) for 15 days. CIDRs
were replaced on day 10, and removed during ovum
pick-up (OPU) on day 15. Follicular growth was
stimulated by the administration of a total of 5.28 mg of
ovine FSH (Ovagen, ICPbio, Auckland, New Zealand)
given in four equal doses administered at 8:00 and
18:00 h on days 13 and 14. One female was removed
from the experiment because of stress during handling;
data from this animal were excluded from analyses.
Blood samples were collected for plasma progester-
one and 17b-estradiol quantification on days 0 (before
insertion of CIDR) and 10 (before replacement of
CIDR), at each time of FSH administration (day 13,
8:00 h and 18:00 h; day 14, 8:00 h and 18:00 h) and
before OPU on day 15. Blood samples were centrifuged
at 3500 rpm for 15 min, and plasma was recovered and
frozen at �20 8C and stored at �80 8C until assayed.
Progesterone and 17b-estradiol concentrations were
determined in duplicate by radioimmunoassay at the
Laboratoire de Dosages Hormonaux, Institut Nationale
de la Recherche Agronomique (Tours, France).
2.4. Oocyte recovery
Cumulus-oocyte complexes (COCs) were recovered
using a semi-laparotomy technique. Female donors
were anesthesized with a combination of an intramus-
cular administration of xylazine (0.2 mg/kg body
weight; Rompun, Bayer, Barcelona, Spain) and
intravenous injection of ketamine chlorohydrate
(15 mg/kg body weight; Imalgene 1000, Merial, Lyon,
France) and then secured in a cradle in dorsal
recumbency. When necessary, surgical anesthesia was
maintained using halothane inhalation. The effects of
xylazine were reversed by intravenous administration of
yohimbine chlorohydrate (0.03 mg/kg body weight;
Sigma, Madrid, Spain).
Ovum pick-up was carried out following procedures
described previously [28]. Briefly, an endoscope was
inserted into the abdominal cavity about 10–15 cm
cranial to the udder, and 4–5 cm to one side of the
midline, while atraumatic grasping forceps were
inserted in a contralateral position through a 3–4 cm
long laparotomy incision. After endoscopic visualiza-
tion, the ovaries were fixed at their ligament origin and
exposed through the laparotomy incision. Follicles
present on the ovarian surface were counted and
aspirated with a 2-ml syringe fitted with a 25-gauge
needle carrying 0.5 ml of warm medium, placed in a
15 ml plastic tube (Falcon, Becton Dickinson, Lepont
De Claix, France) and kept in a water bath at 38 8C until
processed. The collection medium was Hepes-buffered
TCM-199 supplemented with antibiotics (50 mg/ml
streptomycin plus 50 IU/ml penicillin), polyvinyl-
alcohol (PVA, 0.1%, w/v) and heparin (15 IU/ml).
After each OPU session, the ovaries were washed with
warm physiologic solution.
2.5. In vitro maturation
The COCs derived from each female donor were
kept separate throughout the in vitro procedures.
Following oocyte collection, the content of each tube
was poured into a 60 mm Petri dish (Falcon #35-1007,
Becton Dickinson) and observed under a stereomicro-
scope (Nikon SMZ-1500). Oocytes that were degener-
ated and/or with expanded cumulus cells were removed.
After rinsing three to four times in Hepes-buffered
TCM-199 with PVA, heparin and antibiotics, the COCs
were placed in four-well culture dishes (Nunclon,
Nalgene, Nunc International, Roskilde, Denmark)
containing the in vitro maturation medium (TCM-199
plus 10% v/v heat-treated estrus sheep serum, 10 mg
ovine FSH/LH/ml, 1 mg estradiol/ml and 0.1 mg

F. Berlinguer et al. / Theriogenology 69 (2008) 349–359352
glutamine/ml), covered with 250–300 ml of mineral oil
and cultured at 38.5 8C under 5% CO2 in air and
maximum humidity.
2.6. In vitro fertilization
After 24 h, oocytes were washed in Hepes-buffered
TCM-199 followed by gentle pipetting with the aid of
an automatic micropipette to dissociate surrounding
cumulus cells. Fertilization medium consisted of
synthetic oviduct fluid (SOF) [32] supplemented with
heat-inactivated 2% (v/v) estrus sheep serum, 1 mg
hypotaurine/ml and 10 mg heparin/ml. Oocytes were
inseminated with cryopreserved spermatozoa. To avoid
confounding effects due to male-to-male variations, the
same male was used to fertilize oocytes from different
females. In preliminary trials, semen samples from
various males were tested for cryosurvival.
Semen was collected by electroejaculation using a
sine-wave stimulator (P.T. Electronics, Boring, OR,
USA) and evaluated for volume, concentration,
motility, viability, membrane integrity, morphology,
and acrosome integrity, as described previously [21]. A
sperm motility index (SMI) was calculated as follows:
[sperm % motility + (quality of motility � 20) � 1/2].
Semen samples were centrifuged at 700 � g for 5 min
and resuspended in a Tes-Tris–glucose diluent with 5%
egg yolk and 6% glycerol [21]. Sperm suspensions
(adjusted to 400 � 106 spermatozoa/ml) were loaded
into 0.25 ml plastic straws, cooled to 5 8C for 1.5 h and
allowed to equilibrate for an additional 2 h. The straws
were frozen in nitrogen vapors for 10 min and then
plunged into liquid nitrogen. Straws were thawed for
30 s at 37 8C in a water bath and the content of each
straw was poured into a conical glass tube and assessed
for sperm motility and acrosome integrity. Spermatozoa
were selected by using a swim-up procedure. Briefly,
spermatozoa in the cryodiluent (0.25 ml) were placed
gently in the bottom of a round-bottom glass tube
(15 mm � 75 mm) which contained 1 ml SOF medium
(as used for IVF) and incubated at 38.5 8C under 5%
CO2 in air. After 30 min the upper layer of 0.3 ml was
removed (aspirating the sperm suspension at a point
located 3 mm below the surface), and sub-samples were
evaluated for sperm motility and acrosome integrity.
Aliquots of 50 ml of sperm suspension (from the upper
layer) were transferred to 1.5-ml tubes and further
incubated at 38.5 8C under 5% CO2 in air for 1 or 24 h.
Sub-samples were taken for assessment of motility and
acrosome integrity.
The final concentration of spermatozoa used during
IVF was 1 � 106 spermatozoa/ml. Gametes were co-
incubated in four-well culture dishes under mineral oil
for 24 h at 38.5 8C under 5% O2, 5% CO2 and 90% N2.
Sperm samples obtained at the end of gamete co-culture
were examined for motility and acrosome integrity.
2.7. In vitro culture
After 24 h of gamete co-culture, presumptive
zygotes were washed in culture medium to remove
spermatozoa and cell debris. They were transferred to
500 ml of SOF medium supplemented with 0.4% BSA
(w/v) and nonessential and essential amino acids at
oviductal concentrations [33] under mineral oil and
cultured in a humidified atmosphere at 38.5 8C under
5% O2, 5% CO2 and 90% N2. Cleaving embryos were
separated from 1-cell presumptive zygotes after 36 h of
culture. Embryo development was monitored for
several days, every 24 h, until it was arrested.
2.8. Evaluation of meiosis progression
The resumption and progression through meiosis
was evaluated retrospectively after in vitro fertilization.
Non cleaved oocytes and embryos were stained with
Hoechst 33342 and propidium iodide (10 mg/ml each)
and visualized in a microscope (Nikon E-400) equipped
with epifluorescence to determine maturation status
and/or fertilization by the presence of �2 pronuclei or
decondensed sperm heads. Fluorescence of Hoechst and
propidium iodide was observed simultaneously using an
Hg excitation beam and a filter set (UV-2A Nikon)
consisting of a UV330-380 excitation filter, a DM400
chromatic beam splitter and a 420 barrier filter.
3. Results
3.1. Oocyte harvesting, culture and fertilization
Of 56 follicles aspirated from six females (9.3 � 1.7
follicles per female), 35 COCs were recovered
(5.8 � 1.5 oocytes per female), resulting in a overall
recovery rate of 62% (mean � S.E.M., 58.6 � 9.2%
recovery per female) (Table 1).
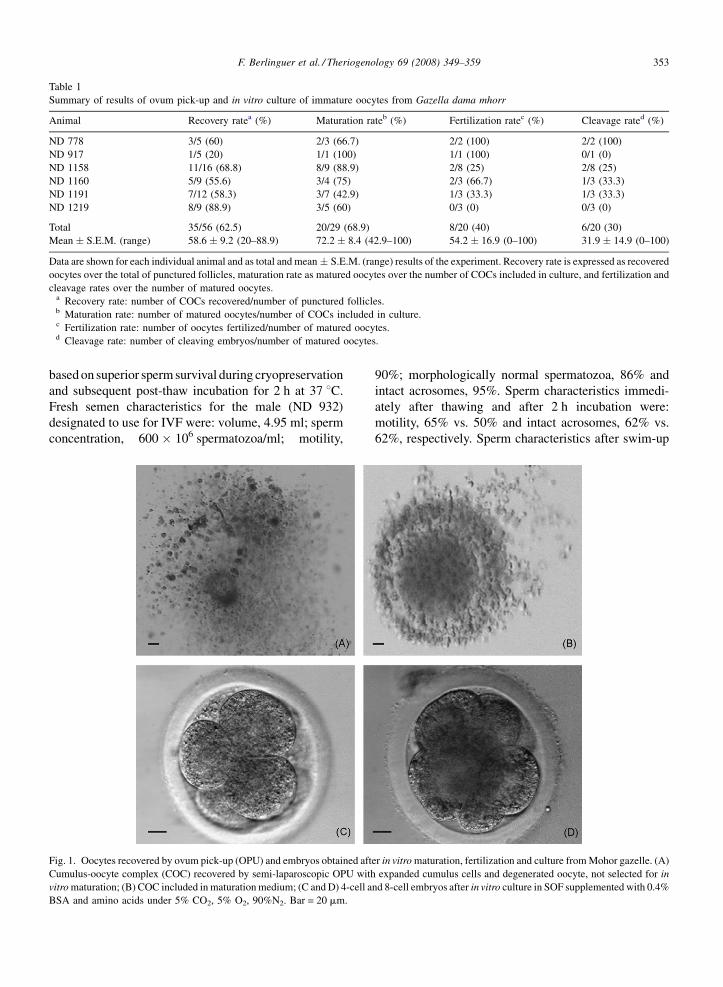
A total of 29 cumulus-surrounded oocytes (Fig. 1)
were selected for in vitro maturation (83% of recovered
oocytes). When examined after culture, 3% (1/29) were
found at the germinal vesicle (GV) stage, 7% (2/29) at
metaphase I (MI), 21% (6/29) were degenerated or not
classified, and 69% (20/29) advanced to metaphase II
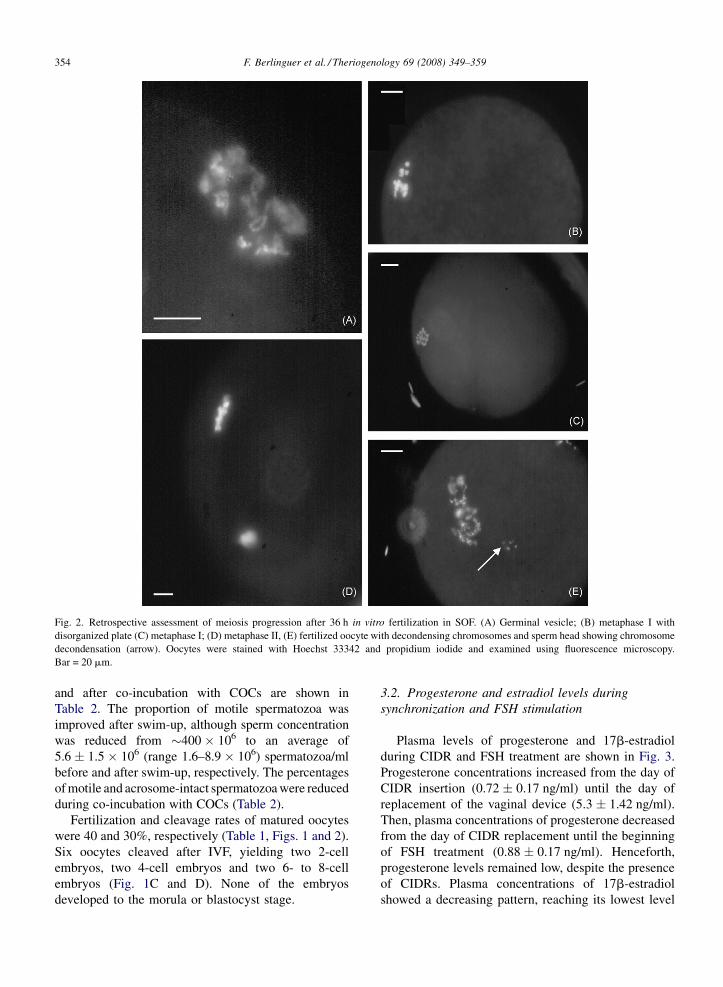
(MII) (Table 1, Figs. 1 and 2).
Semen collected from four males was evaluated pre-
and post-cryopreservation and one was selected for IVF
F. Berlinguer et al. / Theriogenology 69 (2008) 349–359 353
Table 1
Summary of results of ovum pick-up and in vitro culture of immature oocytes from Gazella dama mhorr
Animal Recovery ratea (%) Maturation rateb (%) Fertilization ratec (%) Cleavage rated (%)
ND 778 3/5 (60) 2/3 (66.7) 2/2 (100) 2/2 (100)
ND 917 1/5 (20) 1/1 (100) 1/1 (100) 0/1 (0)
ND 1158 11/16 (68.8) 8/9 (88.9) 2/8 (25) 2/8 (25)
ND 1160 5/9 (55.6) 3/4 (75) 2/3 (66.7) 1/3 (33.3)
ND 1191 7/12 (58.3) 3/7 (42.9) 1/3 (33.3) 1/3 (33.3)
ND 1219 8/9 (88.9) 3/5 (60) 0/3 (0) 0/3 (0)
Total 35/56 (62.5) 20/29 (68.9) 8/20 (40) 6/20 (30)
Mean � S.E.M. (range) 58.6 � 9.2 (20–88.9) 72.2 � 8.4 (42.9–100) 54.2 � 16.9 (0–100) 31.9 � 14.9 (0–100)
Data are shown for each individual animal and as total and mean � S.E.M. (range) results of the experiment. Recovery rate is expressed as recovered
oocytes over the total of punctured follicles, maturation rate as matured oocytes over the number of COCs included in culture, and fertilization and
cleavage rates over the number of matured oocytes.a Recovery rate: number of COCs recovered/number of punctured follicles.b Maturation rate: number of matured oocytes/number of COCs included in culture.c Fertilization rate: number of oocytes fertilized/number of matured oocytes.d Cleavage rate: number of cleaving embryos/number of matured oocytes.
based on superior sperm survival during cryopreservation
and subsequent post-thaw incubation for 2 h at 37 8C.
Fresh semen characteristics for the male (ND 932)
designated to use for IVF were: volume, 4.95 ml; sperm
concentration, 600 � 106 spermatozoa/ml; motility,
Fig. 1. Oocytes recovered by ovum pick-up (OPU) and embryos obtained aft
Cumulus-oocyte complex (COC) recovered by semi-laparoscopic OPU with
vitro maturation; (B) COC included in maturation medium; (C and D) 4-cell a
BSA and amino acids under 5% CO2, 5% O2, 90%N2. Bar = 20 mm.
90%; morphologically normal spermatozoa, 86% and
intact acrosomes, 95%. Sperm characteristics immedi-
ately after thawing and after 2 h incubation were:
motility, 65% vs. 50% and intact acrosomes, 62% vs.
62%, respectively. Sperm characteristics after swim-up
er in vitro maturation, fertilization and culture from Mohor gazelle. (A)
expanded cumulus cells and degenerated oocyte, not selected for in
nd 8-cell embryos after in vitro culture in SOF supplemented with 0.4%
F. Berlinguer et al. / Theriogenology 69 (2008) 349–359354
Fig. 2. Retrospective assessment of meiosis progression after 36 h in vitro fertilization in SOF. (A) Germinal vesicle; (B) metaphase I with
disorganized plate (C) metaphase I; (D) metaphase II, (E) fertilized oocyte with decondensing chromosomes and sperm head showing chromosome
decondensation (arrow). Oocytes were stained with Hoechst 33342 and propidium iodide and examined using fluorescence microscopy.
Bar = 20 mm.
and after co-incubation with COCs are shown in
Table 2. The proportion of motile spermatozoa was
improved after swim-up, although sperm concentration
was reduced from �400 � 106 to an average of
5.6 � 1.5 � 106 (range 1.6–8.9 � 106) spermatozoa/ml
before and after swim-up, respectively. The percentages
of motile and acrosome-intact spermatozoa were reduced
during co-incubation with COCs (Table 2).
Fertilization and cleavage rates of matured oocytes
were 40 and 30%, respectively (Table 1, Figs. 1 and 2).
Six oocytes cleaved after IVF, yielding two 2-cell
embryos, two 4-cell embryos and two 6- to 8-cell
embryos (Fig. 1C and D). None of the embryos
developed to the morula or blastocyst stage.
3.2. Progesterone and estradiol levels during
synchronization and FSH stimulation
Plasma levels of progesterone and 17b-estradiol
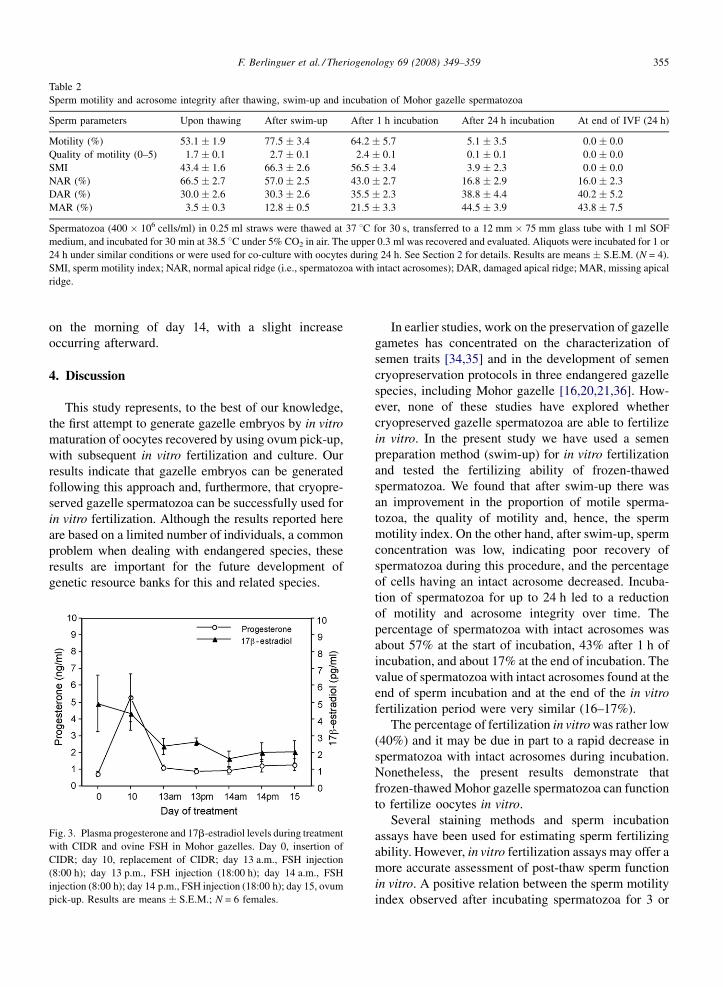
during CIDR and FSH treatment are shown in Fig. 3.
Progesterone concentrations increased from the day of
CIDR insertion (0.72 � 0.17 ng/ml) until the day of
replacement of the vaginal device (5.3 � 1.42 ng/ml).
Then, plasma concentrations of progesterone decreased
from the day of CIDR replacement until the beginning
of FSH treatment (0.88 � 0.17 ng/ml). Henceforth,
progesterone levels remained low, despite the presence
of CIDRs. Plasma concentrations of 17b-estradiol
showed a decreasing pattern, reaching its lowest level
F. Berlinguer et al. / Theriogenology 69 (2008) 349–359 355
Table 2
Sperm motility and acrosome integrity after thawing, swim-up and incubation of Mohor gazelle spermatozoa
Sperm parameters Upon thawing After swim-up After 1 h incubation After 24 h incubation At end of IVF (24 h)
Motility (%) 53.1 � 1.9 77.5 � 3.4 64.2 � 5.7 5.1 � 3.5 0.0 � 0.0
Quality of motility (0–5) 1.7 � 0.1 2.7 � 0.1 2.4 � 0.1 0.1 � 0.1 0.0 � 0.0
SMI 43.4 � 1.6 66.3 � 2.6 56.5 � 3.4 3.9 � 2.3 0.0 � 0.0
NAR (%) 66.5 � 2.7 57.0 � 2.5 43.0 � 2.7 16.8 � 2.9 16.0 � 2.3
DAR (%) 30.0 � 2.6 30.3 � 2.6 35.5 � 2.3 38.8 � 4.4 40.2 � 5.2
MAR (%) 3.5 � 0.3 12.8 � 0.5 21.5 � 3.3 44.5 � 3.9 43.8 � 7.5
Spermatozoa (400 � 106 cells/ml) in 0.25 ml straws were thawed at 37 8C for 30 s, transferred to a 12 mm � 75 mm glass tube with 1 ml SOF
medium, and incubated for 30 min at 38.5 8C under 5% CO2 in air. The upper 0.3 ml was recovered and evaluated. Aliquots were incubated for 1 or
24 h under similar conditions or were used for co-culture with oocytes during 24 h. See Section 2 for details. Results are means � S.E.M. (N = 4).
SMI, sperm motility index; NAR, normal apical ridge (i.e., spermatozoa with intact acrosomes); DAR, damaged apical ridge; MAR, missing apical
ridge.
on the morning of day 14, with a slight increase
occurring afterward.
4. Discussion
This study represents, to the best of our knowledge,
the first attempt to generate gazelle embryos by in vitro
maturation of oocytes recovered by using ovum pick-up,
with subsequent in vitro fertilization and culture. Our
results indicate that gazelle embryos can be generated
following this approach and, furthermore, that cryopre-
served gazelle spermatozoa can be successfully used for
in vitro fertilization. Although the results reported here
are based on a limited number of individuals, a common
problem when dealing with endangered species, these
results are important for the future development of
genetic resource banks for this and related species.
Fig. 3. Plasma progesterone and 17b-estradiol levels during treatment
with CIDR and ovine FSH in Mohor gazelles. Day 0, insertion of
CIDR; day 10, replacement of CIDR; day 13 a.m., FSH injection
(8:00 h); day 13 p.m., FSH injection (18:00 h); day 14 a.m., FSH
injection (8:00 h); day 14 p.m., FSH injection (18:00 h); day 15, ovum
pick-up. Results are means � S.E.M.; N = 6 females.
In earlier studies, work on the preservation of gazelle
gametes has concentrated on the characterization of
semen traits [34,35] and in the development of semen
cryopreservation protocols in three endangered gazelle
species, including Mohor gazelle [16,20,21,36]. How-
ever, none of these studies have explored whether
cryopreserved gazelle spermatozoa are able to fertilize
in vitro. In the present study we have used a semen
preparation method (swim-up) for in vitro fertilization
and tested the fertilizing ability of frozen-thawed
spermatozoa. We found that after swim-up there was
an improvement in the proportion of motile sperma-
tozoa, the quality of motility and, hence, the sperm
motility index. On the other hand, after swim-up, sperm
concentration was low, indicating poor recovery of
spermatozoa during this procedure, and the percentage
of cells having an intact acrosome decreased. Incuba-
tion of spermatozoa for up to 24 h led to a reduction
of motility and acrosome integrity over time. The
percentage of spermatozoa with intact acrosomes was
about 57% at the start of incubation, 43% after 1 h of
incubation, and about 17% at the end of incubation. The
value of spermatozoa with intact acrosomes found at the
end of sperm incubation and at the end of the in vitro
fertilization period were very similar (16–17%).
The percentage of fertilization in vitro was rather low
(40%) and it may be due in part to a rapid decrease in
spermatozoa with intact acrosomes during incubation.
Nonetheless, the present results demonstrate that
frozen-thawed Mohor gazelle spermatozoa can function
to fertilize oocytes in vitro.
Several staining methods and sperm incubation
assays have been used for estimating sperm fertilizing
ability. However, in vitro fertilization assays may offer a
more accurate assessment of post-thaw sperm function
in vitro. A positive relation between the sperm motility
index observed after incubating spermatozoa for 3 or

F. Berlinguer et al. / Theriogenology 69 (2008) 349–359356
6 h and fertilization has been found in the scimitar-
horned oryx [37]. A similar relationship has been
described between sperm longevity and in vitro
fertilization success in the cheetah [38] suggesting that
sperm longevity could be a better predictor of potential
fertility than a single assessment of motility after
thawing. These results should be taken in account for
future studies of semen evaluation in gazelles.
Administration of FSH enhances the number of
oocytes obtained by increasing the number of follicles
that, based on their size and morphological character-
istics, are adequate for aspiration [39]. The response of
female gazelles to treatment with ovine FSH revealed a
rather poor rate of follicular growth. In domestic
ruminants, variation in ovarian response to super-
ovulatory treatment relates to many factors, such as
ovarian status at the beginning of treatment, [40,41],
hormonal profile [42], interval between the last
exogenous gonadotropin stimulation and follicle aspira-
tion [43,44] and donor treatment [45,46]. Among
gazelles, failure to respond could be due either to stress
during animal handling or to insufficient FSH doses.
With regards to the former, improvements could
perhaps be obtained in the future by animal training,
with repeated captures and handling, that may
eventually lead to reduced stress. Another option to
minimize stress could be the use of long-acting
neuroleptics. Use of tranquilizers such as perfenazine
enanthate or haloperidol chlorohydrate in Spanish ibex
(Capra pyrenaica) females has allowed the collection of
transferable embryos after a superovulatory treatment
with ovine FSH [47]. Moreover, administration of the
long-acting neuroleptic flufenazine decanoate prior to
collection of oocytes from European mouflon (Ovis
orientalis musimon) resulted in a better response to the
hormonal treatment, increasing the number of punctu-
red follicles per donor [26]. The only animal that did not
receive neuroleptic treatment had been previously
submitted to regular handling and showed a good
response to superovulatory stimulation [26].
Poor follicular growth could also be due to
insufficient FSH doses. In sheep, successful stimulation
of follicular growth occurs with four doses of FSH (as
used in our study), allowing for a good collection of
healthy oocytes with subsequent fertilization and
development in culture. Constant doses of ovine FSH
seem to produce better results than decreasing ones
[28], although other studies have found no differences
between FSH administration regimes [48,49]. We found
that plasma concentrations of estradiol decreased
during exogenous FSH administration, supporting the
hypothesis that the amount of FSH was insufficient to
adequately stimulate follicular growth. In sheep, FSH
treatment leads to an increase in the number and size of
gonadotropin-responsive follicles and in a subsequent
increase in circulating levels of estradiol [50]. Thus,
estradiol is considered to be a reliable marker of
follicular function [51]. Decreases in follicular secre-
tion of estradiol are related to deficiencies in follicular
health and, moreover, a decrease in the developmental
competence of the oocyte [52]. Therefore, a possible
explanation for the limited ovarian response seen in our
study is that follicles were not responding adequately to
FSH, with stress being an additional factor leading to a
reduced response.
The proportion of oocytes recovered from punctured
follicles (62%) was similar to that obtained using
endoscopic recovery in sheep (55%) [53]. However, it
was lower than that obtained in sheep using laparotomy
(80%) [54] or in goats employing laparoscopy (90%)
[55]. In non-domestic ungulates, oocyte recovery rates
ranged from 36 to 46% in red deer (Cervus elaphus)
[24] and were 57% in sika deer (Cervus nippon) [25],
which are closer to the values obtained in our study.
More studies are needed to improve results of oocyte
harvesting and to explore the possibility of repeated
oocyte collection from the same females.
The size of the follicles found on the ovarian surface
of gazelles was small (2–3 mm). It is known that an
oocyte is unable to resume and progress through
meiosis until it reaches a given size. In sheep, a direct
relationship exists between follicular size and the ability
of oocytes to progress through meiosis [56]. Thus, the
capacity to advance to metaphase II increases with
follicle and oocyte sizes in lambs and adult sheep [56].
It is not clear whether this relationship exists in gazelles,
but it is possible that oocytes contained in small follicles
have a reduced maturation capability.
The overall maturation rate obtained at the end of the
in vitro incubation period was about 70%, with
considerable variation between females, and this result
is similar to data reported in other wild ruminants. In red
deer, a maturation rate of about 55% was found for
oocytes harvested by laparoscopy [24] and a range of
14–85% for oocytes recovered from slaughterhouse
ovaries [25]. In sika deer, 14–76% of oocytes aspirated
by laparoscopy matured in vitro [25]. In domestic sheep,
oocytes recovered by OPU showed a 80–90% matura-
tion rate when conditions similar to those used for
gazelles were employed [28]. Clearly, the fertilization
rate we obtained with gazelle oocytes in the present
study was rather low (40%) as compared to the �80%
fertilization rate achieved in domestic sheep using
similar methodology [28].

F. Berlinguer et al. / Theriogenology 69 (2008) 349–359 357
When gazelle presumptive zygotes were cultured in
vitro, we found that although there was cleavage up to
the 6- to 8-cell stage, none of the embryos progressed to
the morula or blastocyst stage, suggesting the existence
of a developmental block. In sheep, about 15–20% of
embryos develop to the blastocytst stage when a similar
methodology is used [28]. One possible explanation for
our results is that embryo culture conditions were not
adequate for gazelle embryos. In vitro embryo culture
procedures for wild ungulates are currently limited to
protocols developed for domestic animals. Using such
protocols, blastocysts have been obtained after oocyte
in vitro maturation, fertilization and culture in the axis
deer, (Axis axis) [57], red deer [25,58], sika deer [25],
and European mouflon [26]. Adjustments are therefore
needed for improvements in gazelle in vitro embryo
culture, but progress in this area will be slow due to the
scarcity of gametes and embryos from this endangered
species. Additional factors to consider are that
differences exist, even between closely related species,
in the requirements of embryos for in vitro development
[8]. An alternative to overcome the difficulties of in
vitro embryo culture could be an early transfer to
recipient females. In deer, in vitro matured and fertilized
oocytes were capable of establishing pregnancies when
embryos were transferred to recipient hinds at the 2- to
4-cell stage, with this transfer being more efficient than
that of 8-cell embryos [59]. In future gazelle work we
will explore the feasibility of early embryo transfer, i.e.,
oviductal transfer of 2- to 4-cell embryos.
Another likely explanation for poor embryo develop-
ment in culture relates to inadequate oocyte quality and
limited developmental competence. Studies on ewes
treated with progestagen/gonadotropin for in vitro
embryo production have indicated that oocyte quality
(i.e., the ability to develop into a viable embryo after in
vitro maturation, fertilization and culture) seems to
depend on both the reproductive status of the female and
the hormonal treatment employed [28,29]. Since,
gazelles showed a limited response to FSH treatment,
as revealed by poor follicular growth, it is possible that
oocytes had a poor degree of cytoplasmic maturation and
this had an important bearing on post-fertilization deve-
lopment. The fundamental processes involved in the
acquisition of oocyte developmental competence remain
unknown, but there is increasing evidence that such
competence is acquired while still in the follicle [43].
Oocyte pre-maturation and final maturation occurring
during follicular growth are considered crucial steps in
determining the outcome of embryo production [60].
Taken together, our results of in vitro maturation,
fertilization and culture in the Mohor gazelle, achieved
using protocols developed originally for sheep, demon-
strate, for the first time, the production of embryos in
vitro from this endangered species. Although more
studies are needed to improve in vitro conditions, these
results reveal the feasibility of achieving in vitro
fertilization, with frozen-thawed semen, of in vitro
matured oocytes collected from FSH-stimulated endan-
gered gazelles. Aspects that need improvement include
oocyte harvesting conditions, stimulation for the
development of healthier oocytes, and in vitro culture
of embryos to overcome a developmental block that
prevents progression to the blastocysts stage. These
improvements would allow the generation of a
sufficient number of oocytes and embryos for cryo-
banking or transfer.
Acknowledgements
The authors are grateful to Eulalia Moreno for
permission to study the gazelles, Jesus Benzal for
laboratory support, and the staff of the Parque de Rescate
de la Fauna Sahariana, EEZA, CSIC (Juan Belzunces,
Alfonso Lopez, Oscar Salinas, Luis Pozo, Gema Morales,
Leire Correa) for their help in handling the animals. This
study was supported by the Spanish Ministry of
Education and Science (REN 2003-01587, CGL2006-
13340/BOS) and Acciones Integradas (HI20030336).
References
[1] Holt WV, Pickard AR. Role of reproductive technologies and
genetic resource banks in animal conservation. Rev Reprod
1999;4:143–50.
[2] Wildt DE, Wemmer C. Sex and wildlife: the role of reproductive
science in conservation. Biodiv Conserv 1999;8:965–76.
[3] Bennett PM. Establishing animal germplasm resource banks for
wildlife conservation: genetic, population and evolutionary
aspects. In: Watson PF, Holt WV, editors. Cryobanking the
genetic resource. Wildlife conservation for the future?. London:
Taylor and Francis; 2001. p. 47–67.
[4] Wildt DE, Ellis S, Howard JG. Linkage of reproductive sciences:
from ‘‘quick fix’’ to ‘‘integrated’’ conservation. J Reprod Fertil
Suppl 2001;57:295–307.
[5] IUCN. IUCN technical guidelines on the management of ex-situ
populations for conservation. IUCN Publications Service; 2002.
[6] Wildt DE. Potential applications of IVF technology for species
conservation. In: Bavister BD, Cummins J, Roldan ERS, editors.
Fertilization in Mammals. Newton, MA: Serono Symposia,
USA; 1992. pp. 349–364.
[7] Wildt DE, Monfort SL, Donoghue AM, Johnston LA, Howard J.
Embryogenesis in conservation biology—or, how to make an
endangered species embryo. Theriogenology 1992;37:161–84.
[8] Loskutoff NM, Bartels P, Meintjes M, Godke RA, Schiewe MC.
Assisted reproductive technology in nondomestic ungulates. A
model approach to preserving and managing genetic diversity.
Theriogenology 1995;43:3–12.

F. Berlinguer et al. / Theriogenology 69 (2008) 349–359358
[9] Farstad W. Current state in biotechnology in canine and feline
reproduction. Anim Reprod Sci 2000;60:375–87.
[10] Pope CE. Embryo technology in conservation efforts for endan-
gered felids. Theriogenology 2000;53:163–74.
[11] Pukazhenthi BS, Wildt DE. Which reproductive technologies are
most relevant to studying, managing and conserving wildlife?
Reprod Fertil Dev 2004;16:33–46.
[12] Pukazhenthi B, Comizzoli P, Travis AJ, Wildt DE. Applications
of emerging technologies to the study of conservation of threa-
tened and endangered species. Reprod Fertil Dev 2006;18:77–
90.
[13] IUCN, IUCN Red List of threatened species. 2006; [http://
www.iucnredlist.org/, downloaded on April 08, 2007].
[14] Barbosa A, Espeso G. International Studbook of Gazella dama
mhorr. Estacion Experimental de Zonas Aridas. Almeria: CSIC;
2005.
[15] Bainbridge DRJ, Jabbour HN. Potential of assisted breeding
techniques for the conservation of endangered mammalian
species in captivity: a review. Vet Rec 1998;143:159–68.
[16] Roldan ERS, Gomendio M, Garde JJ, Espeso G, Ledda S,
Berlinguer F, et al. Inbreeding and reproduction in endangered
ungulates: preservation of genetic variation through the organi-
zation of genetic resource banks. Reprod Domest Anim
2006;41(Suppl. 2):82–92.
[17] Roldan ERS, Cassinello J, Abaigar T, Gomendio M. Inbreeding,
fluctuating asymmetry, and ejaculate quality in an endangered
ungulate. Proc Roy Soc Lond B 1998;265:243–8.
[18] Gomendio M, Cassinello J, Roldan ERS. A comparative study of
ejaculate traits in three endangered ungulates with different
levels of inbreeding: fluctuating asymmetry as an indicator of
reproductive and genetic stress. Proc Roy Soc Lond B
2000;267:875–82.
[19] Alados CL, Escos J. Phenotypic and genetic characteristics
affecting lifetime reproductive success in female Cuvier, dama
and dorcas gazelles (Gazella cuvieri, G. dama and G. dorcas). J
Zool 1991;223:307–21.
[20] Holt WV, Abaigar T, Jabbour HN. Oestrous synchronization,
semen preservation and artificial insemination in the Mohor
gazelle (Gazella dama mhorr) for the establishment of a genome
resource bank program. Reprod Fertil Dev 1996;8:1215–22.
[21] Garde JJ, Soler AJ, Cassinello J, Crespo C, Malo AF, Espeso G,
et al. Sperm cryopreservation in three species of endangered
gazelles (Gazella cuvieri, G. dama mhorr and G. dorcas
neglecta). Biol Reprod 2003;69:602–11.
[22] Galli C, Crotti G, Notari C, Turini P, Duchi R, Lazzari G.
Embryo production by ovum pick up from live donors. Ther-
iogenology 2001;55:1341–57.
[23] Cognie Y, Poulin N, Locatelli Y, Mermillod P. State-of-the-art
production, conservation and transfer of in-vitro-produced
embryos in small ruminants. Reprod Fertil Dev 2004;16:437–45.
[24] Bainbridge DRJ, Catt SL, Evans G, Jabbour HN. Successful in
vitro fertilization of in vivo matured oocytes aspirated laparosco-
pically from red deer hinds. Theriogenology 1999;51:891–8.
[25] Comizzoli P, Mermillod P, Cognie Y, Chai N, Legendre X,
Mauget R. Successful in vitro production of embryos in the
red deer and the sika deer. Theriogenology 2001;55:649–59.
[26] Ptak G, Clinton M, Barboni B, Muzzeddu M, Cappai P, Tischner
M, et al. Preservation of the wild European mouflon: the first
example of genetic management using a complete program of
reproductive biotechnologies. Biol Reprod 2002;66:796–801.
[27] Berlinguer F, Leoni GG, Bogliolo L, Bebbere D, Succu S, Rosati
I, et al. In vivo and in vitro fertilizing capacity of cryopreserved
European mouflon (Ovis gmelini musimon) spermatozoa used to
restore genetically rare and isolated populations. Theriogenol-
ogy 2005;63:902–11.
[28] Berlinguer F, Leoni G, Bogliolo L, Pintus PP, Rosati I, Ledda S,
et al. FSH different regimes affect the developmental capacity
and cryotolerance of embryos derived from oocytes collected by
ovum pick-up in donor sheep. Theriogenology 2004;61:1477–
86.
[29] Berlinguer F, Gonzalez-Bulnes A, Succu S, Leoni GG, Veiga-
Lopez A, Mossa F, et al. GnRH antagonist enhance follicular
growth in FSH-treated sheep but affect developmental compe-
tence of oocytes collected by ovum pick-up. Theriogenology
2006;65:1099–109.
[30] Rodriguez C, Anel L, Alvarez M, Anel E, Boixo JC, Chamorro
CA, et al. Ovum pick-up in sheep: a comparison between
different aspiration devices for optimal oocyte retrieval. Reprod
Domest Anim 2006;41:106–13.
[31] Morton KM, de Graaf SP, Campbell A, Tomkins LM, Maxwell
WMC, Evans G. Repeat ovum pick-up and in vitro embryo
production from adult ewes with and without FSH treatment.
Reprod Domest Anim 2005;40:422–8.
[32] Tervit HR, Whittingham DG, Rowson LEA. Successful culture
in vitro of sheep and cattle ova. J Reprod Fertil 1972;30:487–93.
[33] Walker SK, Hill JL, Kleemann DO, Nancarrow CD. Develop-
ment of ovine embryos in synthetic oviductal fluid containing
amino acids at oviductal fluid concentrations. Biol Reprod
1996;55:703–8.
[34] Cassinello J, Abaigar T, Gomendio M, Roldan ERS. Character-
istics of the semen of three endangered species of gazelles
(Gazella dama mhorr, G. dorcas neglecta and G. cuvieri). J
Reprod Fertil 1998;113:35–45.
[35] Abaigar T, Cano M, Pickard AR, Holt WV. Use of computer-
assisted sperm motility assessment and multivariate pattern
analysis to characterize ejaculate quality in Mohor gazelles
(Gazella dama mhorr): effects of body weight, electroejacula-
tion technique and short-term semen storage. Reproduction
2001;122:265–73.
[36] Howard J, Pursel VG, Wildt D, Chakraborty P, Bush M. Com-
parative of various extenders for freeze-preservation of semen
from selective captive wild ungulates. J Am Vet Med Assoc
1981;179:1157–61.
[37] Roth TL, Bush ML, Wildt DE, Weiss RB. Scimitar-horned oryx
(Oryx dammah) spermatozoa are functionally competent in a
heterologous bovine in vitro fertilization system after cryopre-
servation on dry ice, in a dry shipper or over liquid nitrogen
vapor. Biol Reprod 1999;60:493–8.
[38] Donoghue AM, Howard JG, Byers AP, Goodrowe KL, Bush M,
Blumer E, et al. Correlation of sperm viability with gamete
interaction and fertilization in vitro in the cheetah (Acynonyx
jubatus). Biol Reprod 1992;46:1047–56.
[39] Hunter MG, Robinson RS, Mann GE, Webb R. Endocrine and
paracrine control of follicular development and ovulation rate in
farm species. Anim Reprod Sci 2004;82:461–77.
[40] Blondin P, Sirard MA. Oocyte and follicular morphology as
determining characteristics for developmental competence in
bovine oocytes. Mol Reprod Dev 1995;41:54–62.
[41] Hagemann LJ, Beaumont SE, Berg M, Donnison MJ, Ledgard A,
Peterson AJ, et al. Development during single IVP of bovine
oocyte from dissected follicle: interactive effects of estrous cycle
stage, follicle size and atresia. Mol Reprod Dev 1999;53:451–8.
[42] Oussaid B, Lonergan P, Khatir H, Guler H, Monniaux D,
Touze JL, et al. Effect of GnRH antagonist-induced prolonged

F. Berlinguer et al. / Theriogenology 69 (2008) 349–359 359
follicular phase on follicular atresia and oocyte developmental
competence in vitro in superovulated heifers. J Reprod Fertil
2000;118:137–44.
[43] Blondin P, Coenen K, Guibault LA, Sirard MA. In vitro produc-
tion of bovine embryos: developmental competence is acquired
before maturation. Theriogenology 1997;47:1061–75.
[44] Sirard MA, Picard L, Dery M, Coenen K, Blomdin P. The time
interval between FSH administration and ovarian aspiration
influences the developmental competence of cattle oocytes.
Theriogenology 1999;51:699–708.
[45] Goodhand KL, Staines ME, Hutchinson JSM, Broadbent PJ. In
vivo oocyte recovery and in vitro embryo production from
bovine oocyte donors treated with progestagen, estradiol and
FSH. Anim Reprod Sci 2000;63:145–58.
[46] Baracaldo MI, Martinez MF, Adams GP, Mapletoft RJ. Super-
ovulatory response following transvaginal follicle ablation in
cattle. Theriogenology 2000;53:1239–50.
[47] Fernandez-Arias A, Alabart JL, Echegoyen E, Folch J. Super-
ovulation of tranquilized Spanish ibex (Capra pyrenaica)
females. Theriogenology 2000;53:331 [abstract].
[48] Baldassarre H, Furnus CC, deMatos DG, Pessi H. In vitro
production of sheep embryos using laparoscopic folliculocent-
esis: alternative gonadotropin treatments for stimulation of
oocyte donors. Theriogenology 1996;45:707–17.
[49] Alberio R, Olivera J, Roche A, Alabart J, Folch J. Performance of
a modified ovum pick-up system using three different FSH
stimulation protocols in ewes. Small Rum Res 2002;46:81–7.
[50] Gonzalez-Bulnes A, Veiga-Lopez A, Garcia P, Garcia-Garcia
RM, Ariznavarreta C, Sanchez MA, et al. Effects of progestagens
and prostaglandin analogues on ovarian function and embryo
viability in sheep. Theriogenology 2005;63:2523–34.
[51] Campbell BK, Scaramuzzi RJ, Webb R. Control of antral follicle
development and selection in sheep and cattle. J Reprod Fertil
Suppl 1995;49:335–50.
[52] Oussaid B, Mariana JC, Poulin N, Fontaine J, Lonergan P,
Beckers JF, et al. Reduction of the developmental competence
of sheep oocytes by inhibition of LH pulses during the
follicular phase with a GnRH antagonist. J Reprod Fertil
1999;117:71–7.
[53] Stangl M, Kuhholzer B, Besenfelder U, Brem G. Repeated
endoscopic ovum pick-up in sheep. Theriogenology 1999;52:
709–16.
[54] Earl CR, Irvine BJ, Kelly JM, Rowe JP, Armstrong DT. Ovarian
stimulation protocols for oocyte collection and in vitro embryo
production from 8 to 9 week old lambs. Theriogenology
1995;43:203 [abstract].
[55] Pierson J, Wang B, Neveu N, Sneek L, Cote F, Karatzas CN, et al.
Effects of repetition, interval between treatments and season on
the results from laparoscopic ovum pick-up in goats. Reprod
Fertil Dev 2004;16:795–9.
[56] Ledda S, Bogliolo L, Leoni G, Naitana S. Follicular size affects
the meiotic competence of in vitro matured prepubertal and adult
oocytes in sheep. Reprod Nutr Dev 1999;39:503–8.
[57] Chapman SA, Keller DL, Westhusin ME, Drew ML, Kraemer
DC. In vitro production of axis deer (Axis axis) embryos, a
preliminary study. Theriogenology 1999;51:280 [abstract].
[58] Berg DK, Thompson JG, Pugh PA, Tervit HR, Asher GW.
Successful in-vitro culture of early cleavage stage embryos
recovered from superovulated red deer (Cervus elaphus). Ther-
iogenology 1995;44:247–54.
[59] Berg DK, Asher GW, Pugh PA, Tervit HR, Thompson JG.
Pregnancies following the transfer of in vitro matured and
fertilized red deer (Cervus elaphus) oocytes. Theriogenology
1995;43:166 [abstract].
[60] Merton JS, de Roos APW, Mullaart E, de Ruigh L, Kaal L, Vos
PLAM, et al. Factors affecting oocyte quality and quantity in
commercial application of embryo technologies in the cattle
breeding industry. Theriogenology 2003;59:651–74.