induction of her2 immunity in outbred domestic cats by dna...
TRANSCRIPT

Research Article
Induction of HER2 Immunity in Outbred DomesticCats by DNA ElectrovaccinationHeather M. Gibson1, Jesse J. Veenstra1, Richard Jones1, Ulka Vaishampayan1,Michele Sauerbrey2, Gerold Bepler1, Lawrence Lum1, Joyce Reyes1, Amy Weise1, andWei-Zen Wei1
Abstract
Domestic cats share human living environments and genetictraits. They develop spontaneous feline mammary carcinoma(FMC) with similar histopathology to human breast cancer.HER2 and AKT phosphorylation was demonstrated in primaryFMC by immunoblot analysis, indicating HER2 as a therapeutictarget. FMC lines K12 and K248 expressing HER1, HER2, andHER3 were sensitive to receptor tyrosine kinase (RTK) inhibi-tors gefitinib and lapatinib. To test HER2 vaccine response incats, purpose-bred, healthy cats were electrovaccinated withheterologous (xenogeneic) or point-mutated feline HER2 DNA.T-cell reactivity to feline self-HER2 was detected in 4 of 10 catsthat received bear HER2, human–rat fusion HER2 (E2Neu) ormutant feline HER2 (feHER2-K), which contains a single aminoacid substitution. The variable T-cell responses may resemble
that in the genetically heterogeneous human population. Allimmune sera to heterologous HER2 recognized feline HER2expressed in 3T3 cells (3T3/HER2), but not that in FMC K12 orK248. Immune sera to mutant pfeHER2-K bound 3T3/HER2cells weakly, but they showed better recognition of K12 andK248 cells that also express HER1 and HER3, suggesting distinctHER2 epitopes displayed by FMC that may be simulated byfeHER2-K. In summary, HER2 DNA electroporation overcomesT-cell immune tolerance in approximately 40% of healthy catsand induces antibodies with distinct specificity. Vaccinationstudies in domestic cats can expedite vaccine iteration to guidehuman vaccine design and better predict outcome, with theadded benefit of helping feline mammary tumor patients.Cancer Immunol Res; 3(7); 777–86. �2015 AACR.
IntroductionCancer vaccines hold great promise in disease treatment and
prevention. However, the prolonged and costly process of reg-ulatory approval, GMP production, and clinical trials prohibitsefficient iteration between novel laboratory findings andhuman clinical trials. Test systems that can predict humanimmune responses will expedite vaccine development andcancer immunotherapy. Although murine models have beenenormously successful in revealing basic immunology andproviding proof of concept, they may fall short in predictinghow the heterogeneous human population would respond tocancer vaccines (1–3). Transgenic mice expressing humantumor-associated antigens (TAA) and developing spontaneoustumors have produced more relevant results (2, 4, 5). Weshowed a striking difference in HER2 DNA vaccine responsebetween BALB/c and C57BL/6 HER2 transgenic mice, illustrat-ing the impact of genetic background on vaccine responses (6).Because a syngeneic mouse strain represents a single individual,
each new vaccine should ideally be tested in multiple mousestrains representing diversified genetic backgrounds.
In transgenicmice, exogenous promoters are oftenused todrivethe expression of TAAs, and antigenic peptides are usually pre-sented by mouse rather than human major histocompatibilitycomplex (MHC). T-cell repertoires are selected in the thymus inwhich both mouse and transgenic human antigens are presented,confounding the test system. The short life spans, lack of naturaltumorigenesis, and comparatively small body weights are amongthe incongruities between rodents andhumans. The limitations ofthese models bring clarity to the need for intermediate animalsystems thatmore closely resemble those in humans, have similarcancer etiology, express a comparable repertoire of TAAs, andrespond to the vaccine delivery approaches in a similar mannerto humans. Importantly, immune responses in a geneticallydiverse outbred population would predict more accurately therange of immune responses in humans.
The domestic cat population is estimated at 1 billion world-wide (7) with approximately 95 million cats residing in U.S.households (8). Feline whole-genome sequencing and SNPmapshave been generated (7, 9), revealing homologous genetic dis-eases between humans and cats (10). Pet animals share humandwellings and develop cancers similar to those that develop inhumans.Domestic dogs and cats develop spontaneousmammarytumors. The incidence of mammary tumor is higher in dogs(11, 12), but half of caninemammary tumors are benign, whereas90% of feline mammary tumors are malignant and mostlyadenocarcinomas (13–15). Hormonal regulation of canine andfeline mammary tumors is evident because the incidence isdrastically reduced by ovariohysterectomy (12, 16). About 15%
1KarmanosCancer Institute,WayneStateUniversity,Detroit, Michigan.2Oakland Veterinary Referral Services, Bloomfield Hills, Michigan.
Note: Supplementary data for this article are available at Cancer ImmunologyResearch Online (http://cancerimmunolres.aacrjournals.org/).
Corresponding Author: Wei-Zen Wei, Department of Oncology, KarmanosCancer Institute, 110 East Warren Avenue, Detroit, MI 48201. Phone: 313-578-4651; Fax: 313-578-4658; E-mail: [email protected]
doi: 10.1158/2326-6066.CIR-14-0175
�2015 American Association for Cancer Research.
CancerImmunologyResearch
www.aacrjournals.org 777
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175

of nonspayed domestic cats develop spontaneous mammarycarcinoma with disease progression and histopathology similarto that of human breast cancer, and HER2 expression has beenreported (14, 17–24). Feline mammary carcinoma (FMC)patients are treated with surgery, radiotherapy, and chemo-therapy. Thus, domestic felines represent a particularly suitablemodel for evaluating breast cancer vaccines.
DNA vaccines have advanced more expediently in veterinarymedicine than in humanmedicine. The first approved therapeuticcancer vaccine in any species was a plasmid DNA encodingheterologous (xenogeneic) human tyrosinase for treatment ofcanine oral melanoma (14, 25, 26). Two other DNA vaccinestargeting West Nile virus in horses (27) and hematopoieticnecrosis virus in salmon (28) have also been approved forveterinary use. In this study, intramuscular electrovaccination wasused to evaluate novel feline HER2 DNA vaccines because theprocedure is well tolerated by humans (29, 30), and gene expres-sion is dramatically enhanced by electroporation (29, 31–34).
HER2 DNA vaccines that we developed have been tested inhuman clinical trials, in which a kinase-deficient full-lengthhuman HER2 DNA induced T-cell responses in breast cancerpatients (35–37). We reported another HER2 construct expres-sing the wild-type (WT) human extracellular and transmem-brane domains (36, 37), which was also tested in breast cancerpatients, but without evidence of T-cell responses (38). Ahuman HER2/rat neu hybrid vaccine showed improved efficacyin transgenic mice (4, 39), and a trial is currently open foraccrual. In the present study, heterologous HER2 as well asfeline HER2 with a single amino acid (a.a.) substitution weretested in outbred domestic shorthair cats with an aim toestablish an improved system for predicting cancer vaccineresponse in humans.
Materials and MethodsAnimals and tissues
BALB/c mice were purchased from Charles River Laboratory.Pathogen-free (SPF) purpose-bred domestic shorthair cats ages 6months to 2 years were obtained from Liberty Research, Inc.Animals were housed and maintained in the Department ofLaboratory Animal Resource facility at theWayne State UniversitySchool of Medicine in accordance with Institutional Animal Careand Use Committee guidelines. The experimental cats wereadopted as domestic pets by the caretaker community aftercompletion of the study. Ablack bear legally harvested inOntario,Canada, was the donor of the liver tissue.
FMC samples were obtained frommastectomy tissues of 2 felinepatients treated at Oakland Veterinary Referral Services (OVRS)in Michigan with consent from the cat owners (SupplementaryTable S1). OVRS-1A and OVRS-1B are two independent primarytumors from the same cat. Three additional mammary tumorsamples with paired, uninvolved stromal tissues were purchasedfrom Colorado State University (CSU-133, 418, and 1646).
Cell linesTheK248 cell linewas established fromapulmonarymetastasis
of a Siamese cat that had a mammary carcinoma, and wasprovided by Dr. John Hilkens and the late Dr. Wim Misdorp atthe Netherland Cancer Institute (40). The mammary carcinomacell line K12 was from a 14-year-old cat, and was established byDr. William Hardy Jr, and provided by Dr. Jaime Modiano of
the University of Pennsylvania (40). SKOV3 cells were purchas-ed from the ATCC. MCF7 cells were obtained from Lisa Polinof the Karmanos Cancer Institute. All cells were maintained inDMEM supplemented with fetal bovine sera, penicillin, andstreptomycin. The feline origin of K248 and K12 cells hasbeen authenticated by short tandem repeat analysis of four loci(Supplementary Fig. S1).
Immunohistochemical analysisPathologic diagnoses were performed according to the WHO
classification for tumors in domestic animals. For feline HER2detection, epitopes were retrieved with sodium citrate buffer(pH 6.0) and histologic grade primary antibodies were appliedaccording to the manufacturer's recommendation (HER2, cloneZ4881; Invitrogen) followed by broad-spectrum HRP polymerconjugate (SuperPicTure PolymerDetectionKit; Zymed) andDABsubstrate (Pierce Biotech). Feline mammary tumor cells, K248,were injected s.c. into SCID mice. Tumor explants were used ascontrols.
DNA cloning and constructionAll cloning primer sequences are shown in Supplementary
Table S2. Feline ERBB2 cDNA encoding HER2 was cloned fromcell lines K248 (40) and K12 (41), and from the ovary of adomestic shorthair cat using a Protoscript kit (New EnglandBioLabs), all of which showed identical sequences. The con-firmed full-length feline ERBB2 (feHER2) cDNA sequence hasbeen submitted to Genbank (JN990983). For vaccination, astop codon was introduced after codon 687 to delete theoncogenic intracellular domain, then subcloned into pVax1;the resultant pfeHER2 contains the signal peptide, and theextracellular and transmembrane domains of feline HER2. ThepfeHER2-K contains a single-nucleotide substitution in codon141 (cag ! aag) of the extracellular domain (ECD) I, based onour reported sequence of feline ERBB2 (Genbank JN990983)and was generated by PCR-based methods and verified by DNAsequencing.
Black bear ERBB2 cDNA encoding black bear HER2 was sim-ilarly cloned from the liver tissue of a black bear. The cDNAsequence was submitted to Genbank (JQ040508). DNA vaccinepbearHER2 encoding the signal peptide and the extracellular andtransmembrane domains were constructed by PCR-based meth-ods similar to that described for pfeHER2.
Feline CSF2 cDNA encoding feline GM-CSF was amplifi-ed from a randomly primed cDNA library (Protoscript; NewEngland Biolabs) prepared from ConA-stimulated feline peri-pheral blood mononuclear cells (PBMC) and cloned intopcDNA3.1. The open reading frame sequence was in accordwith the consensus of feline CSF2 cDNAs in Genbank(AY878357, NM_001009846, AF053007, and AY138140).
Generation of recombinant feline HER2 and human Fc fusionprotein—feHER2-Fc
A plasmid encoding the secreted fusion protein of feline HER2ECD fused to human Ig Fc was constructed. Feline ERBB2 cDNAand the human Ig Fc domain (IGHG1, Genbank BC080557)were PCR amplified and fused by overlap-extension priming,giving a 2,680-bp product, which was cloned into the HindIIIand XbaI sites of pVax1. Murine 3T3 cells were transfected withthis construct, and the feHER2-Fc protein in culture supernatant
Gibson et al.
Cancer Immunol Res; 3(7) July 2015 Cancer Immunology Research778
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175

was quantified by ELISA using mouse anti-human HER2 capturemAb (clone TA-1; Calbiochem), which cross-reacts with felineHER2. Rabbit anti-human IgG was the detection antibody (Jack-son ImmunoResearch). huHER2-Fc was purchased from SinoBiologicals.
Electrovaccination of mice and catsMice were injected with an admix of 50 mg each of vaccine
plasmid andplasmid encodingmurineGM-CSF (pmuGM-CSF) in50 mL PBS in the gastrocnemius muscle (42). Conductive gel wasapplied on the skin over the injection sites. Electroporation wasconducted with NEPA21 electroporator (Napagene) using atweezer electrode. Three 50-ms degenerating bipolar pulses of100 V were administered at each site. Cats were injected with1.5 mg each of HER2 vaccine plasmid and pfeGM-CSF in 1.5 mLPBS, divided equally over three injection sites in the biceps femo-ris or quadriceps. Two rounds of electroporation were applied toeach site as described using a 1.5-cm2 caliper electrode (BTX).
Cell proliferation assayCells were plated at 2,000 to 5,000 per well in 96-well plates
and treated with gefitinib or lapatinib in quintuplicate for 48hours. Alamar Blue reagent (Life Technologies) was added andfluorescence measured after 3 to 4 hours. The percentage ofproliferative activity was determined relative to the average ofuntreated samples.
Western blot analysisCells or tissues were lysed in a nonionic detergent lysis buffer
(43) with protease inhibitor cocktail (Roche Diagnostics) imme-diately after the addition of phosphatase inhibitors (NEB). Totalprotein was quantified by BCA assay (Pierce Biotech). Protein(10 mg) was boiled in Laemmli buffer, separated with 8%SDSPAGE gel and transferred onto polyvinylidene difluoridemembrane for overnight incubation with antibody to HER2(42/c-erbB-2; BD Biosciences), phospho-HER2 Y1248 (polyclon-al; Cell Signaling Technology), Akt (polyclonal; Cell SignalingTechnology), phospho-Akt S473 (587F11; Cell Signaling Tech-nology) or b-actin (I-19; Santa Cruz Biotechnology). After wash-ing in TBS-Tween, membranes were incubated with horseradishperoxidase (HRP)–conjugated secondary antibody before wash-ing and development using enhanced chemiluminescent reagents(Thermo Scientific).
Flow cytometric analysisHER2/neu epitopes were detected bymAb TA-1 (Calbiochem),
trastuzumab (Genentech), 7.16.4 (Calbiochem), N12, or N29(hybridoma cell lines were generous gifts of Dr. Yosef Yarden,Weissman Institute, Israel). mAb to human EGFR (528; SantaCruz Biotechnology), HER3 (SGP1; eBioscience) and HLA-ABC(W6/32; eBioscience) were used as indicated. Phycoerythrin (PE)-conjugated goat anti-mouse or anti-human IgGwas the secondaryantibody (Jackson ImmunoResearch). Flow cytometric analysiswas performed using FACS Canto II and data analyzed withFlowJo (Tree Star).
Tomeasure the antibody level in immune sera, mouse or felinesera were incubated with 3T3 cells engineered to express thedesignated antigen and detected by PE-conjugated anti-mouseor feline IgG secondary antibody (Santa Cruz Biotechnology).Mouse antibody concentrations were extrapolated from a stan-
dard curve of HER2 mAb TA-1. Feline antibody titers weredetermined by serial dilution until binding was no longerdetected above that of the isotype control.
Analysis of T-cell response by ELISPOTMouse splenocytes or feline PBMCs isolated byficoll separation
(GE Healthcare) were maintained in RPMI medium supplemen-ted with fetal bovine sera, penicillin–streptomycin. Feline PBMCswere supplemented with 0.5 ng/mL feline IL2 (R&D Systems).Cells were plated at 2 � 105 per well in round bottom 96-wellplates and cultured with 10 mg/mL feHER2-Fc (3T3 supernatant-equivalent as described above), huHER2-Fc, human IgG control(Jackson ImmunoResearch) or control 3T3-conditioned mediumfor 48 (mouse) or 72 (feline) hours. Total well contents were thentransferred to mouse or feline (R&D Systems) IFNg ELISPOTplates and incubated for an additional 48 hours before detectionand enumeration as previously described (44) or per the manu-facturer's protocol. Visualized cytokine spots were enumeratedusing the ImmunoSpot analyzer (CTL) and expressed as thenumber of cytokine-producing cells per 106 splenocytes orPBMCs.
ResultsExpression of HER2 in FMC
Expression of ERBB family receptor tyrosine kinase (RTK) inFMC was measured by flow cytometry. Surface staining of HER1(EGFR), HER2, and HER3 was detected in K12 and K248 cellsusing mAbs to their human homologs (Fig. 1A). MHC I expres-sion was also detected with mAb W6/32 to a constant region of
B
A
K248 OVRS-1A OVRS-1B OVRS-2
H&E
HER2
HER2(TA-1)
EGFR(528)
MHC I(W6/32)
K248(feline)
K12(feline)
SKOV3(human)
HER3(SGP1)
Figure 1.Expression of HER2 in FMC. A, flow cytometric analysis of cell surface EGFR(HER1), HER2, HER3, and MHCI expression (open histograms) in FMC celllines K12 and K248. Human SKOV3 cells are included as controls.Shaded histograms are unstained controls. B, immunohistochemical analysisof 3 primary FMC samples and FMC line K248 outgrowth in SCID miceusing polyclonal Ab to huHER2. H&E is shown in parallel. All images are at�40 magnification.
HER2 DNA Vaccination of Domestic Cats
www.aacrjournals.org Cancer Immunol Res; 3(7) July 2015 779
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175
human MHC I. Control human ovarian cancer cell line SKOV3showed elevatedHER1 andHER2, while lackingHER3 expression(45). Feline HER2 expression in primary FMC clinical samples(Supplementary Table S1) was detected by immunohistochem-ical (IHC) staining (Fig. 1B). Membrane staining of HER2 wasdetected in all 3 primary FMC samples and in K248 explant,consistent with membrane staining of K248 cells by flow cyto-metry (Fig. 1A). Membranous HER2 in FMC would permitrecognitionbybothhumoral and cellular immunity. Cytoplasmicstaining was also detected in OVRS-1A and the K248 explant, andmay indicate accumulation of incompletely or incorrectly pro-cessed HER2. The clinical significance of cytoplasmic HER2remains unclear, but T cells would recognize peptides derivedfrom surface or cytoplasmic HER2.
RTK activity in FMCActivation of RTK signaling in FMC was tested using a human
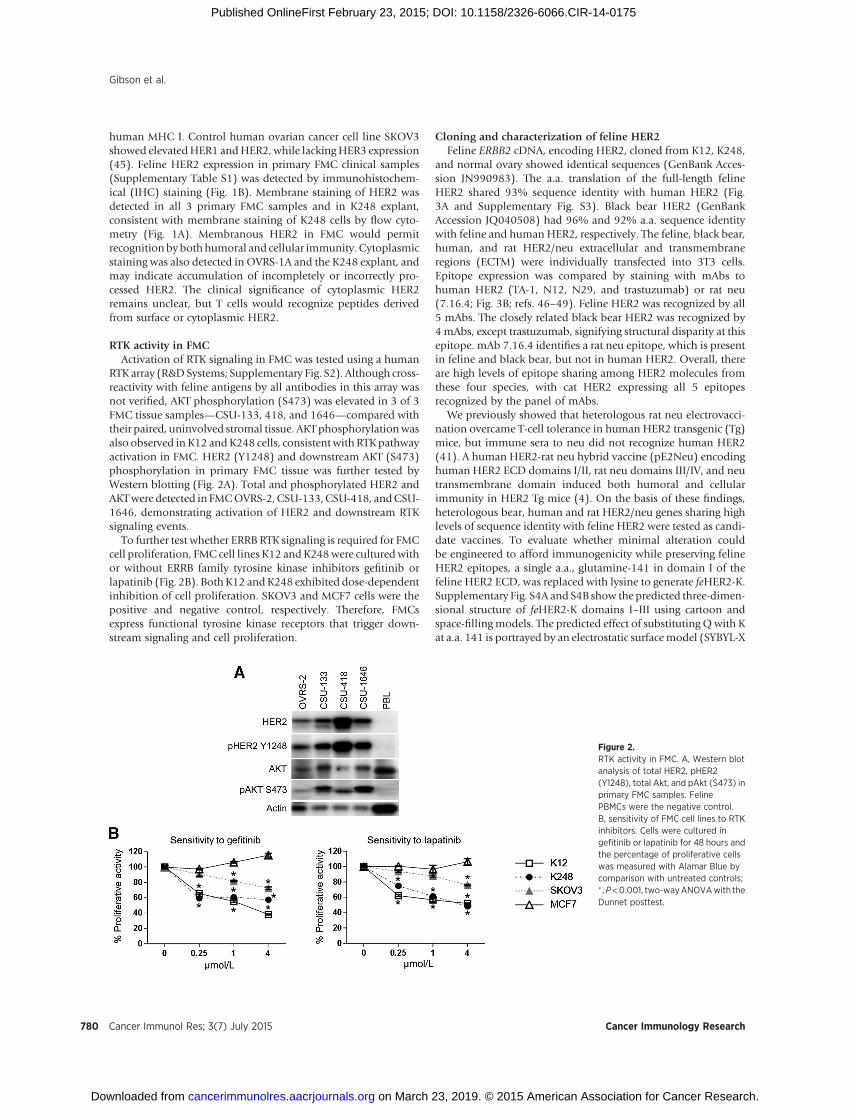
RTK array (R&DSystems; Supplementary Fig. S2). Although cross-reactivity with feline antigens by all antibodies in this array wasnot verified, AKT phosphorylation (S473) was elevated in 3 of 3FMC tissue samples—CSU-133, 418, and 1646—compared withtheir paired, uninvolved stromal tissue. AKTphosphorylationwasalso observed in K12 andK248 cells, consistent with RTKpathwayactivation in FMC. HER2 (Y1248) and downstream AKT (S473)phosphorylation in primary FMC tissue was further tested byWestern blotting (Fig. 2A). Total and phosphorylated HER2 andAKTwere detected in FMCOVRS-2, CSU-133,CSU-418, andCSU-1646, demonstrating activation of HER2 and downstream RTKsignaling events.
To further test whether ERRB RTK signaling is required for FMCcell proliferation, FMC cell lines K12 andK248were culturedwithor without ERRB family tyrosine kinase inhibitors gefitinib orlapatinib (Fig. 2B). BothK12 andK248 exhibited dose-dependentinhibition of cell proliferation. SKOV3 and MCF7 cells were thepositive and negative control, respectively. Therefore, FMCsexpress functional tyrosine kinase receptors that trigger down-stream signaling and cell proliferation.
Cloning and characterization of feline HER2Feline ERBB2 cDNA, encoding HER2, cloned from K12, K248,
and normal ovary showed identical sequences (GenBank Acces-sion JN990983). The a.a. translation of the full-length felineHER2 shared 93% sequence identity with human HER2 (Fig.3A and Supplementary Fig. S3). Black bear HER2 (GenBankAccession JQ040508) had 96% and 92% a.a. sequence identitywith feline and human HER2, respectively. The feline, black bear,human, and rat HER2/neu extracellular and transmembraneregions (ECTM) were individually transfected into 3T3 cells.Epitope expression was compared by staining with mAbs tohuman HER2 (TA-1, N12, N29, and trastuzumab) or rat neu(7.16.4; Fig. 3B; refs. 46–49). Feline HER2 was recognized by all5 mAbs. The closely related black bear HER2 was recognized by4mAbs, except trastuzumab, signifying structural disparity at thisepitope. mAb 7.16.4 identifies a rat neu epitope, which is presentin feline and black bear, but not in human HER2. Overall, thereare high levels of epitope sharing among HER2 molecules fromthese four species, with cat HER2 expressing all 5 epitopesrecognized by the panel of mAbs.
We previously showed that heterologous rat neu electrovacci-nation overcame T-cell tolerance in human HER2 transgenic (Tg)mice, but immune sera to neu did not recognize human HER2(41). A human HER2-rat neu hybrid vaccine (pE2Neu) encodinghuman HER2 ECD domains I/II, rat neu domains III/IV, and neutransmembrane domain induced both humoral and cellularimmunity in HER2 Tg mice (4). On the basis of these findings,heterologous bear, human and rat HER2/neu genes sharing highlevels of sequence identity with feline HER2 were tested as candi-date vaccines. To evaluate whether minimal alteration couldbe engineered to afford immunogenicity while preserving felineHER2 epitopes, a single a.a., glutamine-141 in domain I of thefeline HER2 ECD, was replaced with lysine to generate feHER2-K.Supplementary Fig. S4A and S4B show the predicted three-dimen-sional structure of feHER2-K domains I–III using cartoon andspace-fillingmodels. The predicted effect of substituting Qwith Kat a.a. 141 is portrayed by an electrostatic surfacemodel (SYBYL-X
Figure 2.RTK activity in FMC. A, Western blotanalysis of total HER2, pHER2(Y1248), total Akt, and pAkt (S473) inprimary FMC samples. FelinePBMCs were the negative control.B, sensitivity of FMC cell lines to RTKinhibitors. Cells were cultured ingefitinib or lapatinib for 48 hours andthe percentage of proliferative cellswas measured with Alamar Blue bycomparison with untreated controls;� ,P<0.001, two-wayANOVAwith theDunnet posttest.
Gibson et al.
Cancer Immunol Res; 3(7) July 2015 Cancer Immunology Research780
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175

2.1.1 software; Tripos), inwhich the electropositive side chainofKis indicated by red and Q is shown in blue (Supplementary Fig.S4C).
Immunogenicity of feHER2-KRecombinant feHER2-K was expressed in 3T3 cells and char-
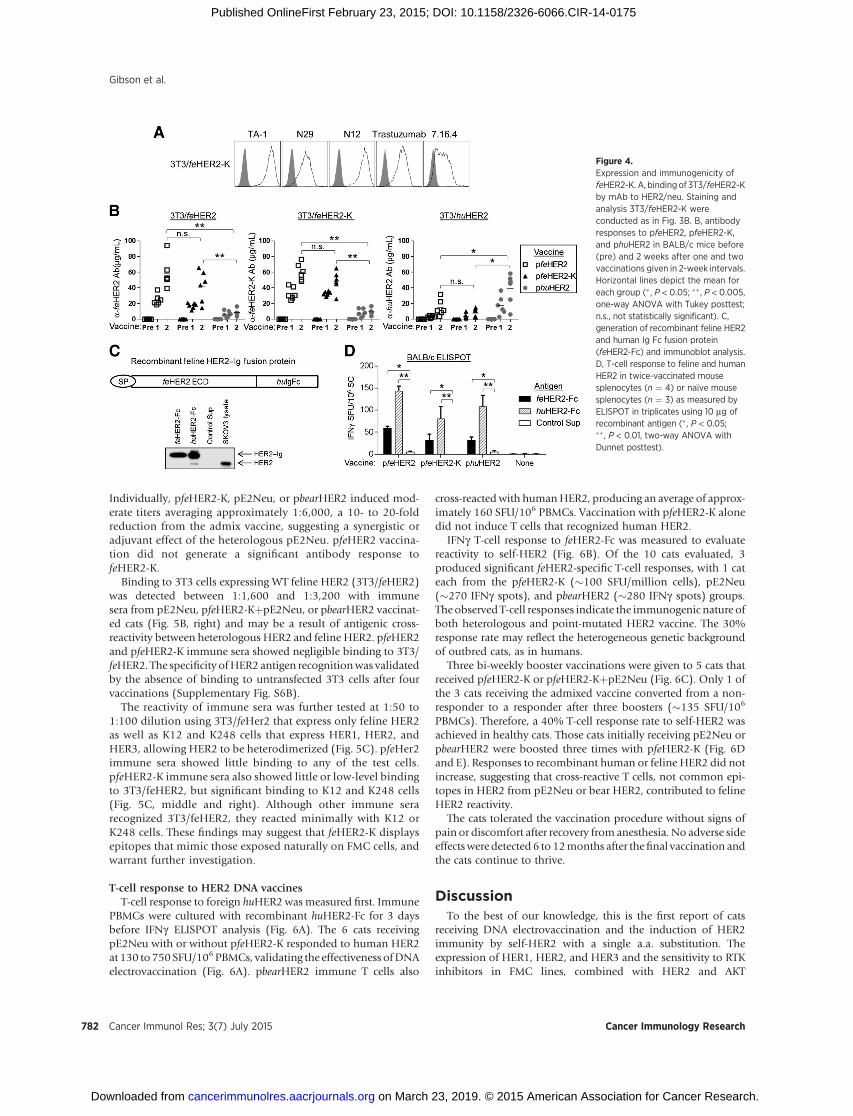
acterized by flow cytometry (Fig. 4A). Positive staining by thepanel of 5 mAbs indicated preservation of 5 HER2/neu epitopesafter the Q! K substitution. Improved binding of feHER2-K byneu-specificmAb 7.16.4 comparedwith its binding toWT feHER2(Fig. 3B) suggests a possible structural alteration of this epitope.
Immunogenicity of feHER2 and feHER2-K was initiallytested in BALB/c mice by electrovaccination with an admix ofpfeHER2, pfeHER2-K, or control phuHER2 (encoding humanHER2 ECTM) together with pmuGM-CSF encoding murineGM-CSF (4, 6, 42, 50). After two immunizations, mice produced59 � 19, 49 � 13, and 39 � 20 mg/mL IgG to their cognateantigens, respectively, as measured with 3T3 cells transfectedwith individual test antigens (Fig. 4B). Expression of feHER2,feHER2-K or huHER2 on individually transfected 3T3 cells wascomparable as verified with mAb TA-1 binding at two differentconcentrations (Supplementary Fig. S5A). pfeHER2 or pfe-HER2-K antibodies are highly cross-reactive, indicating therecognition of dominant foreign epitopes on feline HER2(Fig. 4B, left and middle). The Q141K substitution appears to
create only subtle changes that did not alter the immuneresponse to the dominant foreign epitopes. Modest cross-reac-tivity between feline and human HER2 immune sera was alsoobserved. Neither pre- nor post-vaccination mouse serumbound untransfected 3T3 cells (Supplementary Fig. S5B), sup-porting the HER2 specificity of the immune sera.
To measure T-cell response, a feline HER2 ECD and human IgFc fusionprotein, feHER2-Fc,was generated as the test antigen andverified by Western blot analysis (Fig. 4C). Recombinant humanHER2-Fc (huHER2-Fc) and SKOV3 cell lysate were the controls.Splenocytes from immunized BALB/c mice were incubated withfeHER2-Fc or control huHER2-Fc and IFNg-producing T cells wereenumerated by ELISPOT. T-cell responses to feline HER2 wereinduced by vaccination with pfeHER2, pfeHER2-K, or phuHER2,showing cross-reactivity between feline and humanHER2 antigen(Fig. 4D). This finding also indicates that the Q141K substitutiondoes not alter mouse T-cell response to the WT HER2 protein.Moreprominent responseswere observed after in vitro stimulationwith huHER2-Fc. This result may reflect a more effective stimu-lation with purified huHER2-Fc protein versus feHER2-Fc inculture supernatant.
DNA electrovaccination of catsFor feline vaccinations, GM-CSF cDNA was cloned from feline
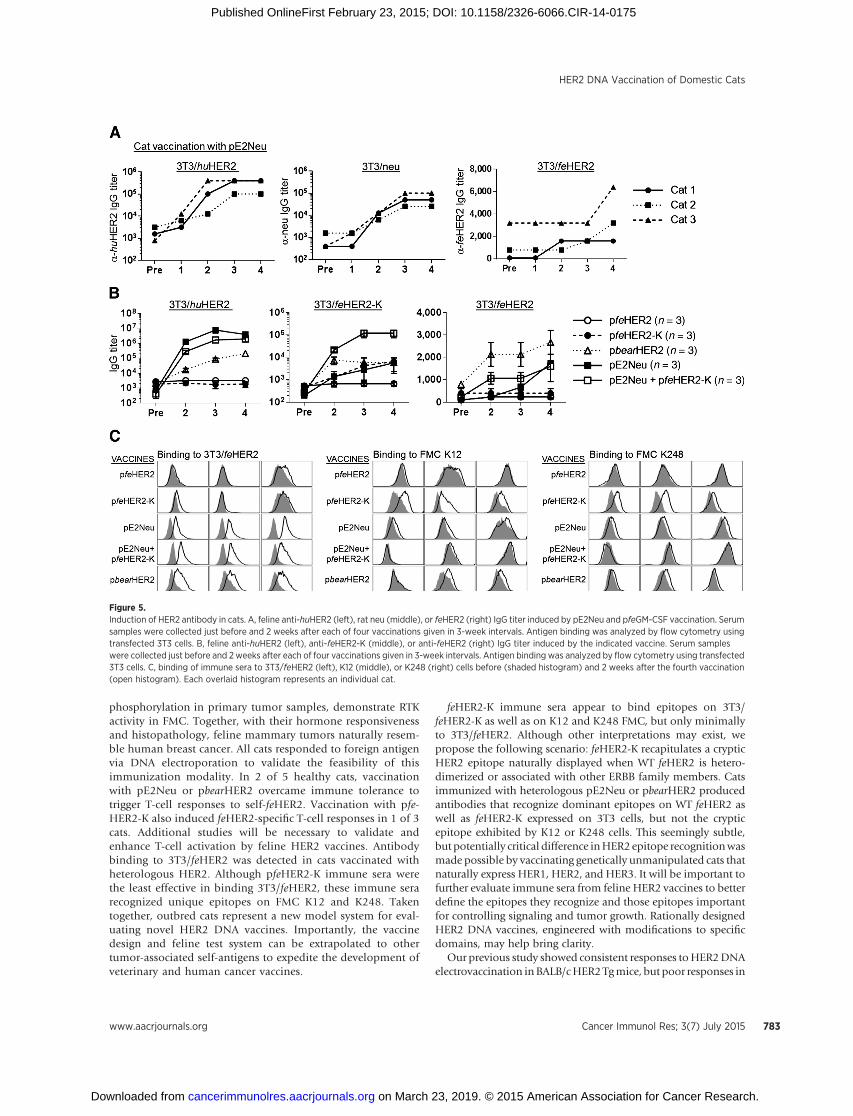
PBMCs (pfeGM-CSF). Generation of recombinant GM-CSF in thesupernatant of transfected 3T3 cells was verified by Western blotanalysis (R&D Systems, Inc.; data not shown). The feasibility ofDNA electrovaccination in cats was initially tested with pE2Neuencoding a fusion protein of HER2 and neu (4). Three healthypurpose-bred, pathogen-free domestic shorthair cats 12 to 24months of agewere injectedwith pE2Neu andpfeGM-CSF in threelegs in the biceps femoralis or quadriceps followed by electropo-ration. Vaccination was administered four times at 3-week inter-vals. Blood was collected through the jugular vein 2 weeks aftereach vaccination. Human HER2 and rat neu binding antibodieswere measured by flow cytometry. Human HER2 binding IgGreached a titer of 1:400,000 in 2 cats and 1:100,000 in the other(Fig. 5A, left). Rat neu binding Ab ranged from 1:25,000 to1:100,000 (Fig. 5A, middle). Vaccination with pE2Neu inducedantibody that also recognized3T3 cells expressingWT felineHER2(Fig. 5A, right), showing reactivity to self-HER2. Neither pre- norpost-fourth vaccination feline serum stained untransfected 3T3cells, illustrating the HER2/neu–specific reactivity of the immunesera (Supplementary Fig. S6A).
Immune response to novel HER2 DNA vaccinesA panel of HER2 vaccines was tested in 15 additional healthy
cats between 5 and 8 months of age. Cats were electrovaccinatedfour times with pfeHER2, pfeHER2-K, pbearHER2, pE2Neu, oran admix vaccine of pfeHER2-K and pE2Neu. As observed pre-viously, the 6 cats that received pE2Neu either alone or incombination with pfeHER2-K developed high levels of IgG anti-body to human HER2 peaking after three vaccinations (Fig. 5B,left). pbearHER2 immune sera cross-reacted with human HER2and the titers increased after each booster immunization. pfe-HER2 or pfeHER2-K immunization did not induce significantantibody response to human HER2.
Recognition of feHER2-K by immune sera was measured bytheir binding to 3T3/feHER2-K (Fig. 5B, middle). A prominentresponse (�1:120,000) was induced by pfeHER2-KþpE2Neu.
TA-17.16.4N29
Full-length HER2 BLAST analysis (%identities/%positives/%gaps)
Cat Human Mouse Rat
Black bear 96 / 98 / 0 92 / 95 / 0 87 / 91 / 0 88 / 91 / 1
Cat 93 / 96 / 0 88 / 92 / 0 88 / 92 / 1
Human 88 / 92 / 0 88 / 92 / 0
Mouse 95 / 97 / 0
A
TrastuzumabN12
TMSP
Rat neu
Feline HER2
Bear HER2
Trastuzumab 7.16.4N12N29TA-1
Human HER2
B
Subdomain I Subdomain II Subdomain III Subdomain IV
Figure 3.HER2 sequences and epitopes. A, BLAST analysis of full-length HER2 a.a.sequence identity. B, schematic of binding by 4 mAbs to human HER2(TA-1, N29, N12, and trastuzumab), and 1 mAb to rat neu (7.16.4). 3T3 cellstransfected to express the indicatedHER2 ECTMwere stainedwith 1mg/mLofthe indicated mAb (open histogram) with detection by PE-conjugatedsecondary antibody. Secondary antibody alone (shaded histogram) was thenegative control.
HER2 DNA Vaccination of Domestic Cats
www.aacrjournals.org Cancer Immunol Res; 3(7) July 2015 781
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175
Individually, pfeHER2-K, pE2Neu, or pbearHER2 induced mod-erate titers averaging approximately 1:6,000, a 10- to 20-foldreduction from the admix vaccine, suggesting a synergistic oradjuvant effect of the heterologous pE2Neu. pfeHER2 vaccina-tion did not generate a significant antibody response tofeHER2-K.
Binding to 3T3 cells expressing WT feline HER2 (3T3/feHER2)was detected between 1:1,600 and 1:3,200 with immunesera from pE2Neu, pfeHER2-KþpE2Neu, or pbearHER2 vaccinat-ed cats (Fig. 5B, right) and may be a result of antigenic cross-reactivity between heterologous HER2 and feline HER2. pfeHER2and pfeHER2-K immune sera showed negligible binding to 3T3/feHER2. The specificity ofHER2 antigen recognitionwas validatedby the absence of binding to untransfected 3T3 cells after fourvaccinations (Supplementary Fig. S6B).
The reactivity of immune sera was further tested at 1:50 to1:100 dilution using 3T3/feHer2 that express only feline HER2as well as K12 and K248 cells that express HER1, HER2, andHER3, allowing HER2 to be heterodimerized (Fig. 5C). pfeHer2immune sera showed little binding to any of the test cells.pfeHER2-K immune sera also showed little or low-level bindingto 3T3/feHER2, but significant binding to K12 and K248 cells(Fig. 5C, middle and right). Although other immune serarecognized 3T3/feHER2, they reacted minimally with K12 orK248 cells. These findings may suggest that feHER2-K displaysepitopes that mimic those exposed naturally on FMC cells, andwarrant further investigation.
T-cell response to HER2 DNA vaccinesT-cell response to foreign huHER2 wasmeasured first. Immune
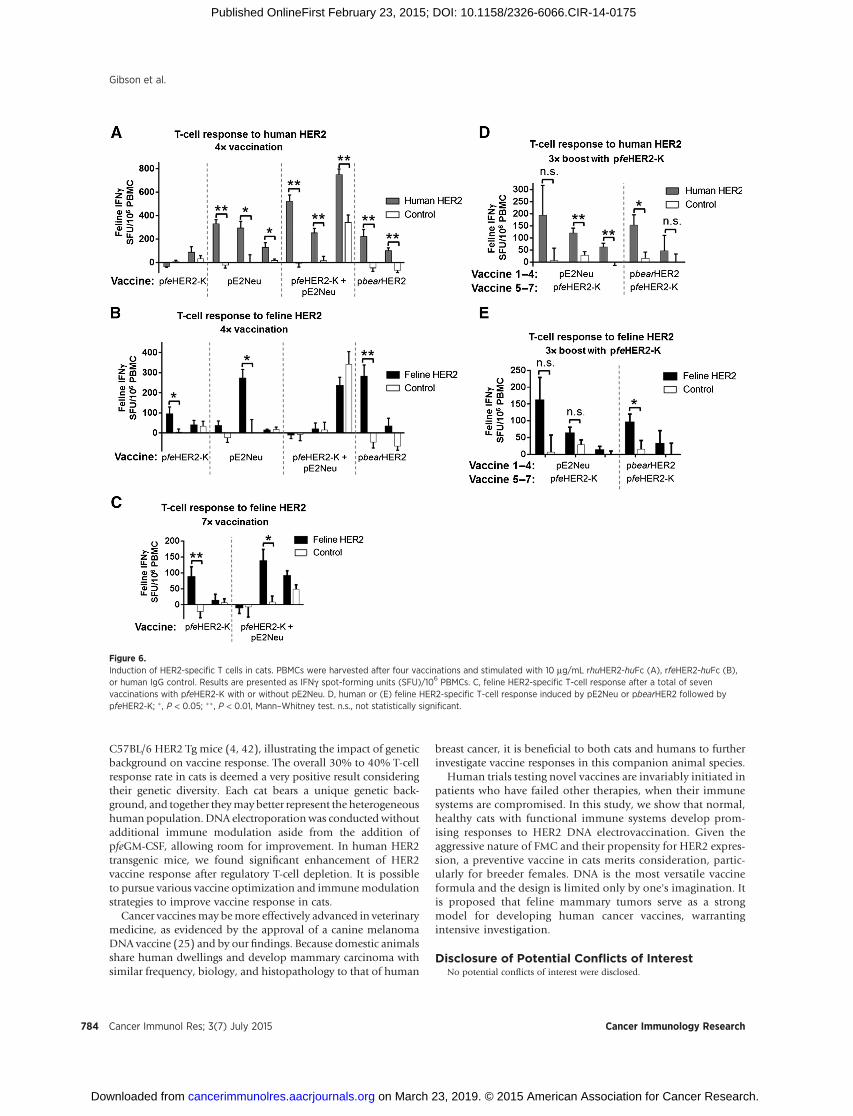
PBMCs were cultured with recombinant huHER2-Fc for 3 daysbefore IFNg ELISPOT analysis (Fig. 6A). The 6 cats receivingpE2Neu with or without pfeHER2-K responded to human HER2at 130 to750 SFU/106PBMCs, validating the effectiveness ofDNAelectrovaccination (Fig. 6A). pbearHER2 immune T cells also
cross-reacted with humanHER2, producing an average of approx-imately 160 SFU/106 PBMCs. Vaccination with pfeHER2-K alonedid not induce T cells that recognized human HER2.
IFNg T-cell response to feHER2-Fc was measured to evaluatereactivity to self-HER2 (Fig. 6B). Of the 10 cats evaluated, 3produced significant feHER2-specific T-cell responses, with 1 cateach from the pfeHER2-K (�100 SFU/million cells), pE2Neu(�270 IFNg spots), and pbearHER2 (�280 IFNg spots) groups.Theobserved T-cell responses indicate the immunogenic nature ofboth heterologous and point-mutated HER2 vaccine. The 30%response rate may reflect the heterogeneous genetic backgroundof outbred cats, as in humans.
Three bi-weekly booster vaccinations were given to 5 cats thatreceived pfeHER2-K or pfeHER2-KþpE2Neu (Fig. 6C). Only 1 ofthe 3 cats receiving the admixed vaccine converted from a non-responder to a responder after three boosters (�135 SFU/106
PBMCs). Therefore, a 40% T-cell response rate to self-HER2 wasachieved in healthy cats. Those cats initially receiving pE2Neu orpbearHER2 were boosted three times with pfeHER2-K (Fig. 6Dand E). Responses to recombinant human or feline HER2 did notincrease, suggesting that cross-reactive T cells, not common epi-topes in HER2 from pE2Neu or bear HER2, contributed to felineHER2 reactivity.
The cats tolerated the vaccination procedure without signs ofpain or discomfort after recovery fromanesthesia. No adverse sideeffectswere detected 6 to 12months after thefinal vaccination andthe cats continue to thrive.
DiscussionTo the best of our knowledge, this is the first report of cats
receiving DNA electrovaccination and the induction of HER2immunity by self-HER2 with a single a.a. substitution. Theexpression of HER1, HER2, and HER3 and the sensitivity to RTKinhibitors in FMC lines, combined with HER2 and AKT
Figure 4.Expression and immunogenicity offeHER2-K. A, binding of 3T3/feHER2-Kby mAb to HER2/neu. Staining andanalysis 3T3/feHER2-K wereconducted as in Fig. 3B. B, antibodyresponses to pfeHER2, pfeHER2-K,and phuHER2 in BALB/c mice before(pre) and 2 weeks after one and twovaccinations given in 2-week intervals.Horizontal lines depict the mean foreach group (� , P < 0.05; �� , P < 0.005,one-way ANOVA with Tukey posttest;n.s., not statistically significant). C,generation of recombinant feline HER2and human Ig Fc fusion protein(feHER2-Fc) and immunoblot analysis.D, T-cell response to feline and humanHER2 in twice-vaccinated mousesplenocytes (n ¼ 4) or na€�ve mousesplenocytes (n ¼ 3) as measured byELISPOT in triplicates using 10 mg ofrecombinant antigen (� , P < 0.05;��, P < 0.01, two-way ANOVA withDunnet posttest).
Gibson et al.
Cancer Immunol Res; 3(7) July 2015 Cancer Immunology Research782
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175
phosphorylation in primary tumor samples, demonstrate RTKactivity in FMC. Together, with their hormone responsivenessand histopathology, feline mammary tumors naturally resem-ble human breast cancer. All cats responded to foreign antigenvia DNA electroporation to validate the feasibility of thisimmunization modality. In 2 of 5 healthy cats, vaccinationwith pE2Neu or pbearHER2 overcame immune tolerance totrigger T-cell responses to self-feHER2. Vaccination with pfe-HER2-K also induced feHER2-specific T-cell responses in 1 of 3cats. Additional studies will be necessary to validate andenhance T-cell activation by feline HER2 vaccines. Antibodybinding to 3T3/feHER2 was detected in cats vaccinated withheterologous HER2. Although pfeHER2-K immune sera werethe least effective in binding 3T3/feHER2, these immune serarecognized unique epitopes on FMC K12 and K248. Takentogether, outbred cats represent a new model system for eval-uating novel HER2 DNA vaccines. Importantly, the vaccinedesign and feline test system can be extrapolated to othertumor-associated self-antigens to expedite the development ofveterinary and human cancer vaccines.
feHER2-K immune sera appear to bind epitopes on 3T3/feHER2-K as well as on K12 and K248 FMC, but only minimallyto 3T3/feHER2. Although other interpretations may exist, wepropose the following scenario: feHER2-K recapitulates a crypticHER2 epitope naturally displayed when WT feHER2 is hetero-dimerized or associated with other ERBB family members. Catsimmunized with heterologous pE2Neu or pbearHER2 producedantibodies that recognize dominant epitopes on WT feHER2 aswell as feHER2-K expressed on 3T3 cells, but not the crypticepitope exhibited by K12 or K248 cells. This seemingly subtle,but potentially critical difference inHER2 epitope recognitionwasmade possible by vaccinating genetically unmanipulated cats thatnaturally express HER1, HER2, and HER3. It will be important tofurther evaluate immune sera from feline HER2 vaccines to betterdefine the epitopes they recognize and those epitopes importantfor controlling signaling and tumor growth. Rationally designedHER2 DNA vaccines, engineered with modifications to specificdomains, may help bring clarity.
Our previous study showed consistent responses toHER2DNAelectrovaccination in BALB/cHER2Tgmice, but poor responses in
Figure 5.Induction of HER2 antibody in cats. A, feline anti-huHER2 (left), rat neu (middle), or feHER2 (right) IgG titer induced by pE2Neu and pfeGM-CSF vaccination. Serumsamples were collected just before and 2 weeks after each of four vaccinations given in 3-week intervals. Antigen binding was analyzed by flow cytometry usingtransfected 3T3 cells. B, feline anti-huHER2 (left), anti-feHER2-K (middle), or anti-feHER2 (right) IgG titer induced by the indicated vaccine. Serum sampleswere collected just before and 2 weeks after each of four vaccinations given in 3-week intervals. Antigen binding was analyzed by flow cytometry using transfected3T3 cells. C, binding of immune sera to 3T3/feHER2 (left), K12 (middle), or K248 (right) cells before (shaded histogram) and 2 weeks after the fourth vaccination(open histogram). Each overlaid histogram represents an individual cat.
HER2 DNA Vaccination of Domestic Cats
www.aacrjournals.org Cancer Immunol Res; 3(7) July 2015 783
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175
C57BL/6 HER2 Tg mice (4, 42), illustrating the impact of geneticbackground on vaccine response. The overall 30% to 40% T-cellresponse rate in cats is deemed a very positive result consideringtheir genetic diversity. Each cat bears a unique genetic back-ground, and together theymay better represent the heterogeneoushuman population. DNA electroporation was conducted withoutadditional immune modulation aside from the addition ofpfeGM-CSF, allowing room for improvement. In human HER2transgenic mice, we found significant enhancement of HER2vaccine response after regulatory T-cell depletion. It is possibleto pursue various vaccine optimization and immunemodulationstrategies to improve vaccine response in cats.
Cancer vaccinesmay bemore effectively advanced in veterinarymedicine, as evidenced by the approval of a canine melanomaDNA vaccine (25) and by our findings. Because domestic animalsshare human dwellings and develop mammary carcinoma withsimilar frequency, biology, and histopathology to that of human
breast cancer, it is beneficial to both cats and humans to furtherinvestigate vaccine responses in this companion animal species.
Human trials testing novel vaccines are invariably initiated inpatients who have failed other therapies, when their immunesystems are compromised. In this study, we show that normal,healthy cats with functional immune systems develop prom-ising responses to HER2 DNA electrovaccination. Given theaggressive nature of FMC and their propensity for HER2 expres-sion, a preventive vaccine in cats merits consideration, partic-ularly for breeder females. DNA is the most versatile vaccineformula and the design is limited only by one's imagination. Itis proposed that feline mammary tumors serve as a strongmodel for developing human cancer vaccines, warrantingintensive investigation.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Figure 6.Induction of HER2-specific T cells in cats. PBMCs were harvested after four vaccinations and stimulated with 10 mg/mL rhuHER2-huFc (A), rfeHER2-huFc (B),or human IgG control. Results are presented as IFNg spot-forming units (SFU)/106 PBMCs. C, feline HER2-specific T-cell response after a total of sevenvaccinations with pfeHER2-K with or without pE2Neu. D, human or (E) feline HER2-specific T-cell response induced by pE2Neu or pbearHER2 followed bypfeHER2-K; � , P < 0.05; �� , P < 0.01, Mann–Whitney test. n.s., not statistically significant.
Gibson et al.
Cancer Immunol Res; 3(7) July 2015 Cancer Immunology Research784
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175

DisclaimerThe content of this article is solely the responsibility of the authors and does
not necessarily represent the official views of theNIHorDepartment ofDefense.
Authors' ContributionsConception and design: H.M. Gibson, R. Jones, L. Lum, J. Reyes, W.-Z. WeiDevelopment of methodology: H.M. Gibson, R. Jones, J. Reyes, W.-Z. WeiAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): H.M. Gibson, J.J. Veenstra, M. Sauerbrey, J. Reyes,W.-Z. WeiAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): H.M. Gibson, R. Jones, U. Vaishampayan, J. Reyes,W.-Z. WeiWriting, review, and/or revision of themanuscript:H.M. Gibson, J.J. Veenstra,R. Jones, U. Vaishampayan, M. Sauerbrey, G. Bepler, A. Weise, W.-Z. WeiAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases):U. Vaishampayan, G. Bepler, J. Reyes, W.-Z. WeiStudy supervision: R. Jones, W.-Z. WeiOther (cloning of feline and Ursus americanus cDNA's used for constructionof vectors and vaccines used in the studydescribed in the article; STRanalysis/authentication of feline cell lines used for construction of these vaccines, andfor analysis of host immune responses to these vaccines): R. Jones
AcknowledgmentsThe authors dedicate this work in memory of Dr. Marie Piechocki, who
devoted her life to cancer research. The authors thank Dr. Elizabeth Dawe andMs. Janine Matthei for their expert handling and care of the cats and Dr. WendyWiesend for her pathology expertise. The authors also thank the MICR, Geno-mics, Multidisciplinary Research, and AMTEC cores of the Karmanos CancerInstitute (P30CA022453). Special thanks to LynnMarla, Colleen Bruning-Fann,Gail Chapman, Nancy Hendrick, Carol Logan, Linda Schroeder, and ColleenSeitz for the black bear harvest.
Grant SupportThis research was supported by NIH RO1 CA76340 (to W.-Z. Wei) and
National Oncogenomics and Molecular Imaging Center W81XWH-10-2-0068(to G. Bepler).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received September 19, 2014; revised January 22, 2015; accepted February14, 2015; published OnlineFirst February 23, 2015.
References1. Davis MM. A prescription for human immunology. Immunity 2008;
29:835–8.2. Ostrand-Rosenberg S. Animal models of tumor immunity, immu-
notherapy, and cancer vaccines. Curr Opin Immunol 2004;16:143–50.
3. Gendler SJ, Mukherjee P. Spontaneous adenocarcinomamousemodels forimmunotherapy. Trends Mol Med 2001;7:471–5.
4. Jacob JB, Quaglino E, Radkevich-Brown O, Jones RF, Piechocki MP, ReyesJD, et al. Combining human and rat sequences in her-2 DNA vaccinesblunts immune tolerance and drives antitumor immunity. Cancer Res2010;70:119–28.
5. Piechocki MP, Ho YS, Pilon S, Wei WZ. Human ErbB-2 (Her-2) transgenicmice: a model system for testing Her-2 based vaccines. J Immunol2003;171:5787–94.
6. Radkevich-BrownO, Jacob J, KershawM,WeiWZ.Genetic regulation of theresponse to Her-2 DNA vaccination in human Her-2 transgenic mice.Cancer Res 2009;69:212–8.
7. Mullikin JC, HansenNF, Shen L, EblingH,DonahueWF, TaoW, et al. Lightwhole genome sequence for SNP discovery across domestic cat breeds.BMC Genomics 2010;11:406.
8. Available from: http://www.humanesociety.org/issues/pet_overpopula-tion/facts/pet_ownership_statistics.html
9. Pontius JU,Mullikin JC, SmithDR, Agencourt Sequencing T, Lindblad-TohK, Gnerre S, et al. Initial sequence and comparative analysis of the catgenome. Genome Res 2007;17:1675–89.
10. O'Brien SJ,Menotti-RaymondM,MurphyWJ, YuhkiN. The FelineGenomeProject. Ann Rev Genetics 2002;36:657–86.
11. Dorn CR, Taylor DO, Schneider R, Hibbard HH, Klauber MR. Survey ofanimal neoplasms in Alameda and Contra Costa Counties, California. II.Cancermorbidity indogs and cats fromAlamedaCounty. JNatl Cancer Inst1968;40:307–18.
12. Schneider R, Dorn CR, Taylor DO. Factors influencing canine mammarycancer development and postsurgical survival. J Natl Cancer Inst1969;43:1249–61.
13. Hayden DW, Nielsen SW. Feline mammary tumours. J Small Anim Pract1971;12:687–98.
14. Munson L,Moresco A. Comparative pathology of mammary gland cancersin domestic and wild animals. Breast Dis 2007;28:7–21.
15. Gimenez F, Hecht S, Craig LE, Legendre AM. Early detection, aggressivetherapy: optimizing the management of feline mammary masses. J FelineMed Surg 2010;12:214–24.
16. Overley B, Shofer FS, Goldschmidt MH, Sherer D, Sorenmo KU. Associ-ation between ovarihysterectomy and feline mammary carcinoma. J VetIntern Med 2005;19:560–3.
17. Soares M, Correia J, Rodrigues P, Simoes M, de Matos A, Ferreira F. FelineHER2 protein expression levels and gene status in feline mammarycarcinoma: optimization of immunohistochemistry (IHC) and in situhybridization (ISH) techniques. Microsc Microanal 2013;19:876–82.
18. De Maria R, Olivero M, Iussich S, Nakaichi M, Murata T, Biolatti B, et al.Spontaneous feline mammary carcinoma is a model of HER2 overexpres-sing poor prognosis human breast cancer. Cancer Res 2005;65:907–12.
19. Ordas J, Millan Y, Dios R, Reymundo C, de Las Mulas JM. Proto-oncogeneHER-2 in normal, dysplastic and tumorous feline mammary glands: animmunohistochemical and chromogenic in situ hybridization study. BMCCancer 2007;7:179.
20. Winston J, Craft DM, Scase TJ, Bergman PJ. Immunohistochemical detec-tion of HER-2/neu expression in spontaneous feline mammary tumours.Vet Comp Oncol 2005;3:8–15.
21. Burrai GP, Mohammed SI, Miller MA, Marras V, Pirino S, Addis MF, et al.Spontaneous felinemammary intraepithelial lesions as amodel for humanestrogen receptor- and progesterone receptor-negative breast lesions. BMCCancer 2010;10:156.
22. Maniscalco L, Iussich S, de LasMulas JM,Millan Y, Biolatti B, SasakiN, et al.Activation of AKT in feline mammary carcinoma: a new prognostic factorfor feline mammary tumours. Vet J 2012;191:65–71.
23. Millanta F, Calandrella M, Citi S, Della Santa D, Poli A. Overexpressionof HER-2 in feline invasive mammary carcinomas: an immunohisto-chemical survey and evaluation of its prognostic potential. Vet Pathol2005;42:30–4.
24. Rasotto R, Caliari D, Castagnaro M, Zanetti R, Zappulli V. An Immuno-histochemical study of HER-2 expression in feline mammary tumours.J Comp Pathol 2011;144:170–9.
25. Grosenbaugh DA, Leard AT, Bergman PJ, Klein MK, Meleo K, Susaneck S,et al. Safety and efficacy of a xenogeneic DNA vaccine encoding for humantyrosinase as adjunctive treatment for oral malignant melanoma in dogsfollowing surgical excision of the primary tumor. Am J Vet Res 2011;72:1631–8.
26. ManleyCA, LeibmanNF,Wolchok JD, Riviere IC, Bartido S, CraftDM, et al.Xenogeneic murine tyrosinase DNA vaccine for malignant melanoma ofthe digit of dogs. J Vet Intern Med 2011;25:94–9.
27. Davis BS, ChangGJ, CroppB, Roehrig JT,MartinDA,Mitchell CJ, et al.WestNile virus recombinant DNA vaccine protects mouse and horse from viruschallenge and expresses in vitro a noninfectious recombinant antigen thatcan be used in enzyme-linked immunosorbent assays. J Virol 2001;75:4040–7.
28. Traxler GS, Anderson E, LaPatra SE, Richard J, Shewmaker B, Kurath G.Naked DNA vaccination of Atlantic salmon Salmo salar against IHNV. DisAquat Organ 1999;38:183–90.
HER2 DNA Vaccination of Domestic Cats
www.aacrjournals.org Cancer Immunol Res; 3(7) July 2015 785
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175

29. Low L, Mander A, McCann K, Dearnaley D, Tjelle T, Mathiesen I, et al.DNA vaccination with electroporation induces increased antibodyresponses in patients with prostate cancer. Hum Gene ther 2009;20:1269–78.
30. Bagarazzi ML, Yan J, Morrow MP, Shen X, Parker RL, Lee JC, et al.Immunotherapy against HPV16/18 generates potent TH1 and cytotoxiccellular immune responses. Sci Transl Med 2012;4:155ra38.
31. Flingai S, Czerwonko M, Goodman J, Kudchodkar SB, Muthumani K,Weiner DB. Synthetic DNA vaccines: improved vaccine potency by elec-troporation and co-delivered genetic adjuvants. Front Immunol 2013;4:354.
32. Senovilla L, Vacchelli E, Garcia P, Eggermont A, FridmanWH,Galon J, et al.Trial watch: DNA vaccines for cancer therapy. Oncoimmunology 2013;2:e23803.
33. Grunwald T, Tenbusch M, Schulte R, Raue K, Wolf H, Hannaman D, et al.Novel vaccine regimen elicits strong airway immune responses and controlof respiratory syncytial virus in non-human primates. J Virol 2014;88:3997–4007.
34. Kutzler MA, Weiner DB. DNA vaccines: ready for prime time? Nat RevGenetics 2008;9:776–88.
35. Norell H, Poschke I, Charo J, Wei WZ, Erskine C, Piechocki MP, et al.Vaccination with a plasmid DNA encoding HER-2/neu together with lowdoses of GM-CSF and IL-2 in patients with metastatic breast carcinoma: apilot clinical trial. J Transl Med 2010;8:53.
36. Wei WZ, Shi WP, Galy A, Lichlyter D, Hernandez S, Groner B, et al.Protection against mammary tumor growth by vaccination with full-length, modified human ErbB-2 DNA. Int J Cancer 1999;81:748–54.
37. Whittington PJ, Piechocki MP, Heng HH, Jacob JB, Jones RF, Back JB, et al.DNA vaccination controls Her-2þ tumors that are refractory to targetedtherapies. Cancer Res 2008;68:7502–11.
38. DiazCM,Chiappori A, Aurisicchio L, Bagchi A, Clark J,Dubey S, et al. Phase1 studies of the safety and immunogenicity of electroporated HER2/CEADNA vaccine followed by adenoviral boost immunization in patients withsolid tumors. J Transl Med 2013;11:62.
39. Quaglino E, Mastini C, Amici A, Marchini C, Iezzi M, Lanzardo S, et al. Abetter immune reaction to Erbb-2 tumors is elicited in mice by DNAvaccines encoding rat/human chimeric proteins. Cancer Res 2010;70:2604–12.
40. Minke JM, Schuuring E, van den Berghe R, Stolwijk JA, Boonstra J,Cornelisse C, et al. Isolation of two distinct epithelial cell lines from asingle feline mammary carcinoma with different tumorigenic potential innude mice and expressing different levels of epidermal growth factorreceptors. Cancer Res 1991;51:4028–37.
41. Modiano JF, Kokai Y, Weiner DB, Pykett MJ, Nowell PC, Lyttle CR.Progesterone augments proliferation induced by epidermal growth factorin a feline mammary adenocarcinoma cell line. J Cell Biochem 1991;45:196–206.
42. Jacob J, RadkevichO, Forni G, Zielinski J, ShimD, Jones RF, et al. Activity ofDNA vaccines encoding self or heterologous Her-2/neu in Her-2 or neutransgenic mice. Cell Immunol 2006;240:96–106.
43. Gibson HM, Mishra A, Chan DV, Hake TS, Porcu P, Wong HK. Impairedproteasome function activates GATA3 in T cells and upregulates CTLA-4:relevance for Sezary syndrome. J Invest Dermatol 2013;133:249–57.
44. Gibson H, Munns S, Freytag S, Barton K, Veenstra J, Bettahi I, et al.Immunotherapeutic intervention with oncolytic adenovirus in mousemammary tumors. Oncoimmunology 2015;4:e984523.
45. Gostring L, Malm M, Hoiden-Guthenberg I, Frejd FY, Stahl S, Lofblom J,et al. Cellular effects of HER3-specific affibody molecules. PloS ONE2012;7:e40023.
46. McKenzie SJ, Marks PJ, Lam T, Morgan J, Panicali DL, Trimpe KL, et al.Generation and characterization of monoclonal antibodies specific for thehuman neu oncogene product, p185. Oncogene 1989;4:543–8.
47. Stancovski I, Hurwitz E, Leitner O, Ullrich A, Yarden Y, Sela M.Mechanistic aspects of the opposing effects of monoclonal antibodiesto the ERBB2 receptor on tumor growth. Proc Natl Acad Sci U S A1991;88:8691–5.
48. Hudziak RM, Lewis GD, Winget M, Fendly BM, Shepard HM, Ullrich A.p185HER2 monoclonal antibody has antiproliferative effects in vitro andsensitizes human breast tumor cells to tumor necrosis factor. Mol Cell Biol1989;9:1165–72.
49. Drebin JA, Stern DF, Link VC, Weinberg RA, Greene MI. Monoclonalantibodies identify a cell-surface antigen associated with an activatedcellular oncogene. Nature 1984;312:545–8.
50. Jacob JB, Kong YC,Meroueh C, Snower DP, David CS, Ho YS, et al. Controlof Her-2 tumor immunity and thyroid autoimmunity by MHC andregulatory T cells. Cancer Res 2007;67:7020–7.
Cancer Immunol Res; 3(7) July 2015 Cancer Immunology Research786
Gibson et al.
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175

2015;3:777-786. Published OnlineFirst February 23, 2015.Cancer Immunol Res Heather M. Gibson, Jesse J. Veenstra, Richard Jones, et al. ElectrovaccinationInduction of HER2 Immunity in Outbred Domestic Cats by DNA
Updated version
10.1158/2326-6066.CIR-14-0175doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerimmunolres.aacrjournals.org/content/suppl/2015/02/21/2326-6066.CIR-14-0175.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerimmunolres.aacrjournals.org/content/3/7/777.full#ref-list-1
This article cites 49 articles, 13 of which you can access for free at:
Citing articles
http://cancerimmunolres.aacrjournals.org/content/3/7/777.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerimmunolres.aacrjournals.org/content/3/7/777To request permission to re-use all or part of this article, use this link
on March 23, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst February 23, 2015; DOI: 10.1158/2326-6066.CIR-14-0175