informe tÉcnico final - sappi.ipn.mx · se encontró una débil correlación de tallas...
TRANSCRIPT

INSTITUTO POLITÉCNICO NACIONAL
CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
INFORME TÉCNICO FINAL Proyecto SIP 20070360
Ecología trófica de Octopus bimaculatus Verril, 1883 en
Bahía de Los Ángeles, B.C., México
La Paz, B.C.S. diciembre de 2007
1

RESUMEN
Los cefalópodos desempeñan un papel importante en los ambientes marinos, como presas y como predadores de una amplia variedad de presas. Entender y cuantificar sus relaciones tróficas es clave para comprender la estructura y funcionamiento de los ecosistemas marinos. Sin embargo, la investigación sobre su alimentación se dificulta debido a que las presas se encuentran muy fragmentadas por la acción mecánica del pico. Debido a esto y al hábito de descarte de restos duros de las presas, la alimentación de los pulpos se ha estudiado también por medio del análisis de acumulaciones de restos de presas que dejan en sus refugios y por la observación directa de sus hábitos. En este contexto durante un ciclo anual se realizaron recolectas mensuales de Octopus bimaculatus de julio de 2006 a junio de 2007 y de los cúmulos de descartes de presas cercanos a sus refugios con la finalidad de estudiar sus hábitos alimentarios en la bahía de Los Ángeles, B.C. Se identificaron las presas al mínimo taxón posible y se aplicaron los métodos cuantitativos tradicionales (numérico, frecuencia de aparición y gravimétrico) y el índice de importancia relativa de Pinkas et al. (1971). Se estimo la amplitud de la dieta (estandarizado de Levin) y el traslapo trófico (Morisita-Horn) entre sexos y tallas. Asimismo, se realizaron pruebas de regresión entre las tallas de las conchas y la de los pulpos para determinar si existe algún tipo de relación. Se analizó un total de 288 individuos, 222 de los cuales presentaron alimento. Las presas se encontraron en avanzado estado de digestión y el Índice de Peso-Llenado más alto se presentó en Primavera. Se encontraron 85 categorías de presas en ocho phyla distintos. Los cangrejos xanthoideos fueron la presa más importante, seguida de moluscos bivalvos y gusanos equiuros. Consumieron principalmente bivalvos durante el invierno y más cangrejos xanthoideos durante la primavera. O. bimaculatus es un consumidor especialista. Presentaron una alta sobreposición en las dietas entre sexos y tallas por madurez gonádica. Se encontró una débil correlación de tallas concha/pulpo en las especies de bivalvos.
2

INTRODUCCIÓN
Las pesquerías de cefalópodos, que incluyen calamares, pulpos y
sepias, se han incrementado a nivel mundial de manera importante en
las últimas cinco décadas, alcanzando su máximo en 2004 con una
captura total de más de 3.4 millones de toneladas (www.fao.org/figis).
En México las pesquerías de cefalópodos más importantes son la del
calamar gigante, Dosidicus gigas en el Golfo de California (Morales-
Bojórquez et.al., 2000) y la conjunta de pulpo rojo Octopus maya y
pulpo común O. vulgaris en el Mar Caribe y Golfo de México (Solís-
Ramírez, 1987), la cual se caracteriza por su elevado valor comercial
(Hernández-Flores et al., 2000; www.sagarpa.gob.mx). Otras especies
de pulpos como O. macrocopus, O. bimaculatus (Roper et. al., 1995) O.
micropyrsus, O. fitchi, O. veligero, O. alecto, O. digueti, O. rubescens, O.
hubbsorum (Villarreal et al., 1997) son capturadas en menor escala en
los litorales del Océano Pacífico Mexicano y Golfo de California, aunque
se reportan indistintamente como recurso “pulpo”
(www.sagarpa.gob.mx). Los campos pesqueros más importantes del
Océano Pacífico, se encuentran en Jalisco y Baja California
(www.sagarpa.gob.mx). En Bahía de Los Ángeles, B.C., se pesca
principalmente Octopus bimaculatus durante casi todo el año, con un
descanso de agosto a octubre en los que se captura O. hubbsorum
(Valdez et al., 2006).
Esta última zona, junto con los canales Salsipuedes y de
Ballenas, ha sido reconocida como una de las áreas marinas de mayor
importancia para la conservación de la biodiversidad en México
(CONABIO, 1996; Enríquez-Andrade y Danemann, 1998). Se considera
además, una de las zonas más productivas del Golfo de California, con
pesquerías y actividad turística regionalmente importantes (Daneman y
Peynador, 2004). La fauna de invertebrados marinos intermareales y
submareales es de gran diversidad y presenta especies poco comunes y
poco estudiadas (Millen y Bertsch, 2000). Debido a la importancia
ecológica de esta bahía, es necesaria la realización de estudios
3

biológicos y ecológicos sobre las especies que se distribuyen en esta
zona que permitan apoyar su decreto como área natural protegida
(Daneman y Peynador, 2004).
Por otro lado, los cefalópodos, dentro de los ecosistemas marinos,
constituyen un componente importante de biomasa marina (Rathjen,
1991), ocupando generalmente el rol de depredadores subdominantes.
Los cefalópodos son consumidores voraces con una alta eficiencia
metabólica que mantienen elevadas tasas de crecimiento (Boucher-
Rodoni et al., 1987; Rodhouse y Nigmatullin, 2000) además al ser
oportunistas, son capaces de alimentarse de un amplio rango de presas,
principalmente crustáceos, peces y moluscos (Nixon, 1987). La gran
relevancia de los cefalópodos en las redes tróficas marinas puede ser
evaluada desde la importancia relativa en sus depredadores (Clarke,
1987), siendo importantes presas de gran número de especies de peces
(Smale, 1996; Segura-Zaragoza, et al., 1997; Aguiar-dos-santos y
Haimovici, 2002; Bachok et al, 2004; Markaida, 2005), aves (Croxall y
Prince, 1996; Aguiar-dos-santos y Haimovici, 2002) y mamíferos
marinos (Clarke, 1996; Aguiar-dos-santos y Haimovici, 2002). En
particular los pulpos son presas de aves acuáticas, odontocetos
(Guerra, 1992), de un gran número de especies de peces tales como
espáridos, balístidos, morénidos, tetradóntidos (Hernández-López,
2000), lutjánidos y serránidos (Leyva-Villarreal et al., 1987) así como de
sepias y otros pulpos (Hernández-López, 2000).
Las actividades relacionadas a la alimentación son probablemente
las actividades voluntarias más frecuentes en la mayoría de los
animales durante todo su ciclo vital. El escape de los depredadores, los
eventos ligados a la reproducción, migración y otras actividades pueden
ser ocasionales o periódicas, pero la alimentación es normalmente parte
de la rutina diaria en la vida de cualquier animal (Ridley, 1995). Es así
que la morfología, anatomía, hábitat, distribución y otros posibles
aspectos están de alguna manera ligados a la dieta (Ridley, 1995;
Castro y Hernández-García, 1995). La composición de la dieta de una
especie no es constante a lo largo de su vida, ya que existen
4

variaciones con el crecimiento, grado de madurez, estación del año, etc.
(Castro, 1993). La alimentación y los procesos digestivos generalmente
cambian durante el ciclo de vida de los cefalópodos. La cantidad y
calidad del alimento varía en relación a las variantes de cambio de
hábitat durante su ontogenia (Boucher-Rodoni et al., 1987) así como a
los cambios morfológicos que acompañan al crecimiento (Castro y
Hernández-García, 1995). El reconocimiento de estos cambios
ontogenéticos en la dieta resulta esencial para el entendimiento de la
ecología del individuo (Guerra-Sierra y Sánchez-Lizaso, 1998). Durante
las etapas tempranas en pulpos (paralarvas y juveniles) se ha
determinado que las limitantes en el desarrollo son la cantidad y
calidad del alimento disponible (Villanueva et al., 2000); por lo que al
igual que en peces, la inanición es la causa principal de mortalidad en
este periodo de vida (Ferron y Leggett, 1994).
A pesar de la importancia ecológica y comercial de las especies de
pulpo en México, los estudios sobre su biología y ecología son escasos e
incluso inexistentes para la mayoría de las especies. Este es el caso de
O. bimaculatus, cuya distribución abarca desde Santa Bárbara
California hasta el sur de la Península de California, y en el Golfo de
California desde Puerto Peñasco, Son. y San Felipe, B.C. en el golfo
hasta Bahía de La Paz (Hochberg y Fileds, 1980). Se le encuentra en
agujeros y hendiduras de sustrato rocoso desde la zona intermareal
hasta los 50 metros de profundidad (Hochberg, 1980). Existen
concentraciones importantes en Bahía de los Ángeles e Isla Ángel de la
Guarda (Valdez et al., 2006; Vázquez-Solórzano et al., 2006).
Se propone el estudio de la ecología trófica de O. bimaculatus con
el fin de conocer las especies con las que interacciona en Bahía de Los
Ángeles, B.C. Este estudio también puede contribuir al conocimiento de
la fauna de la zona, lo cual aportará bases que permitan decretarla
como área natural protegida, también puede proporcionar elementos
para la futura elaboración de dietas de pulpos de esta especie en
cultivo.
ANTECEDENTES
5

Sistemática, diagnosis y hábitos alimenticios de la especie.
Phyllum Mollusca
Clase Cephalopoda
Subclase Oegopsida
Superorden Octobrachia
Orden Octopodida
Familia Octopodidae
Octopus bimaculatus Verril, 1883.
Sistemática de la especie según Doyle et al., (1994).
O. bimaculatus es una especie de la familia Octopodidae que
presenta un cuerpo grande y musculoso, manto piriforme y una
longitud de manto que oscila entre 7 y 18 cm y de 30 a 120 cm de
longitud total en adultos. De color variable; generalmente gris oscuro,
café, rojo u olivo moteado. Presenta dos ocelos conspicuos (manchas en
forma de ojos) situados uno debajo de cada ojo, entre el segundo y
tercer par de brazos (Hochberg, 1980).
Esta especie tiene un periodo de vida de 12 a 18 meses
aproximadamente. Al igual que todos los cefalópodos, O. bimaculatus es
carnívoro durante todo su ciclo de vida, alimentándose de presas
invertebradas vivas del bentos. Se considera un depredador generalista
y macrófago (Ambrose, 1984). O. bimaculatus acostumbra desechar las
partes duras de sus presas, tales como exoesqueletos de crustáceos y
conchas de moluscos en las proximidades de sus refugios, formando
acumulaciones que es posible identificar y cuantificar para estudios de
hábitos alimenticios (Ambrose, 1982; Ambrose, 1987; Nixon, 1987;
Mather, 1991; Dodge y Scheel, 1999; Smith, 2003).
Se ha encontrado que varias especies de octópodos realizan
perforaciones en la conchas y exoesqueletos de sus presas por medio de
la acción combinada de la rádula y la saliva (Tucker and Mapes, 1978;
6

Nixon and Boyle, 1982; Guerra and Nixon, 1987; Saunders et al., 1991;
Nixon and Maconnachie, 1988; Mather and Nixon, 1990).
Asímismo, se ha encontrado que a una mayor talla relativa de la
concha (longitud de la concha) a su depredador hay un mayor
porcentaje de perforación (Cortez et al., 1993; Steer y Semmens, 2003),
lo cual puede estar influido por la especie de la presa.
Ambrose (1984) basado en el análisis de las acumulaciones y en
observaciones directas en campo, menciona que los juveniles se
alimentan de crustáceos principalmente y los adultos principalmente de
gasterópodos (75%): Tegula aureotincta 37%, Norrisia norrisi 12%,
Tegula eiseni 11%, Astrea undosa 11%; un quitón (Stenoplax conspicua
11%), bivalvos (5%) y cangrejos-camarones (5%). Midiendo la
abundancia relativa de las presas, menciona que la especies más
abundantes son T. eiseni, T. aureotincta y A. undosa, respectivamente.
En el mismo estudio, el autor mide preferencia por las presas,
dando mayor importancia a los cangrejos, en segundo lugar a bivalvos y
quitones y en tercero a gasterópodos. La diferencia entre dieta y
preferencias puede estar principalmente en el carácter oportunista del
pulpo al preferir presas abundantes de su hábitat y que le impliquen el
menor esfuerzo de captura. Se observa que la dieta refleja una
combinación de ambos factores, principalmente de oportunismo sobre
presas abundantes pero también de cierta preferencia hacia algunas
especies (Ambrose, 1984).
Hábitos alimenticios con otras metodologías y en otras
especies.
Para la determinación de dietas también es posible el análisis
tradicional de contenidos del tracto digestivo, sobretodo para las
especies que consumen presas con exoesqueletos o conchas menores al
diámetro del pico o cuando las presas son principalmente crustáceos y
peces (Quetglas et al., 1998; Cardoso et al., 2004, Quetglas et al., 2005).
Sin embargo, existen algunas limitantes al respecto, tales como que las
presas se encuentran muy fragmentadas por la acción mecánica del
7

pico y la alta digestión en la que suelen encontrarse las mismas (Nixon,
1987; Rodhouse y Nigmatullin, 1996). Se ha mencionado que este tipo
de estudio tiende a subestimar la abundancia de moluscos en la dieta,
aunque la metodología que utiliza las acumulaciones de los refugios
tiende a sobreestimarlas (Nixon, 1987; Quetglas et al., 1998).
Hernández-López (2000) analiza estómagos de O. vulgaris en las costas
de Gran Canaria, España, obteniendo una mayor cantidad de peces
(Scorpaenidae, Sparidae, Mullidae, Gobiidae) y crustáceos (cangrejos,
isópodos y anfípodos), presentando un decremento de crustáceos de
acuerdo a la talla, al igual que en otros cefalópodos (Rodhouse y
Nigmatullin, 1996), así como un incremento de peces en la dieta de
acuerdo a la talla. Las hembras juveniles presentaron mayor porcentaje
de canibalismo. También observó cambios estacionales en la
composición específica de la dieta.
Con esta misma metodología Villanueva (1993) describe las
principales presas de O. magnificus en el sudeste del Atlántico por su
frecuencia de aparición: 67% del cangrejo Bathynectes piperitus, 23%
del estomatópodo Pontophilus gracilis y 20% del cangrejo ermitaño
Parapagurus sp.”A”, también encontró importante la presencia de peces
bentopelágicos (58%). Cabe mencionar que ambos estudios no utilizan
el Índice de Importancia Relativa (IIR) propuesto por Pinkas (1971). En
otro estudio para O. vulgaris en el Mar Mediterráneo, se menciona que
la alimentación está basada en crustáceos decápodos y en segundo
grado de peces, ocasionalmente de gasterópodos y cefalópodos
(Quetglas et al., 1998).
En el estudio realizado para O. mimus en las costas de Chile, se
analizan tanto contenidos estomacales como acumulaciones de restos
en los refugios, mostrando una dieta compuesta principalmente de
cangrejos braquiuros (Canchriidae y Grapsidae) y anomuros (según
método numérico y frecuencia de aparición), también de peces
blénnidos (según el método gravimétrico). Describe que la dieta es
afectada en gran medida por el sexo y la maduración, en donde los
individuos de mayor edad consumen menos presas y más pequeñas
8

(Cortez et al., 1995). Esto está relacionado a la disminución en la
ingestión de alimento durante el periodo reproductivo de los pulpos
(Boucher-Rodoni et al., 1987).
Otra metodología utilizada en el estudio de la ecología trófica de
pulpos es la observación directa en campo, dado el carácter
territorialista de los pulpos, la cual puede complementar el análisis de
estómagos y de restos de presas en los refugios. Esta metodología tiene
las ventajas de poder identificar y medir directamente las presas, y de
causar un mínimo de disturbios en el hábitat de los pulpos y su
comunidad (Mather, 1991; Smith, 2003). Por otro lado, este tipo de
estudio permite observar directamente cómo se lleva a cabo la captura
de las presas, factores de elección y de consumo, o bien estrategias de
caza y forrajeo (Hanlon y Messenger, 2002), aspectos no considerados
en las metodologías tradicionales de hábitos alimenticios. También es
factible hacer observaciones sobre la clasificación de las presas en tipo
de recursos según su hábitat (excavadores del fondo, larvas que viven
entre las rocas, etc) como recomienda Gerking (1994) para estudios de
alimentación.
Existe un estudio que utiliza las tres metodologías mencionadas
(análisis de tractos digestivos, de acumulaciones de restos y
observaciones in situ), (Smith, 2003). Este autor reporta 39 presas
para O. vulgaris en las costas de Sudáfrica, 30 se presentaron en las
acumulaciones, 14 en los estómagos y 8 en las observaciones. Las
presas más importantes fueron el cangrejo Plagusia chabrus (64.6%
de IIR) y el abulón Haliotis midae (21.6%). También reportó otros
moluscos y crustáceos, así como peces, poliquetos y un
equinodermo. El autor observa una diversificación de la dieta con el
incremento de la talla, lo cual coincide con lo observado para otros
cefalópodos (Boucher-Rodoni et al., 1987).
En general se observa un mayor número de presas y taxa con el
empleo de métodos combinados, lo cual ofrece una mejor estimación
del espectro trófico.
9

Estudios sobre pulpos en México.
Existen pocos estudios sobre las especies de Octopus que habitan
en las costas de México. La mayoría están enfocados a aspectos de su
pesquería (Aguilar-Chávez, 1995; Solís-Ramírez et al., 1998; Alejo-Plata
y Cerdenares-Ladrón-De-Guevara, 2002; Sánchez-Cruz, 2006). Otros
tratan aspectos biológicos generales, como el realizado para O. maya
(Solís-Ramírez, 1967; Aguilar-Chávez, 1995). Otro estudio que incluye
una parte de alimentación de O. hubbsorum en las costas de Oaxaca, el
cual menciona que su principal alimento es el cangrejo Petrolisthes sp.
(Sánchez-Cruz, 2006). Leyva-Villarreal et al., (1987) reporta que O.
veligero se alimenta de almejas del género Anomalocardia sp., langostas
(Panulirus sp.) y cangrejos no identificados. Este autor describe además
aspectos de tipo de hábitat y estrategias para evitar la depredación.
JUSTIFICACIÓN
En comparación a otros grupos marinos como peces, mamíferos,
y crustáceos, los estudios ecológicos en cefalópodos son escasos.
Actualmente y debido a este desconocimiento, los esfuerzos de
investigación se están concentrando en el estudio de la abundancia de
los stocks y su importancia en las cadenas tróficas, ya sea en su rol de
presas o de consumidores (Piatkowski et al., 2001).
La información existente sobre O. bimaculatus es muy escasa, ya
que solamente hay estudios realizados en California, E.U.A. en el
periodo 1976-1979 (Ambrose 1982; 1984; 1986; 1988) y ninguno para
el Golfo de California. Específicamente el estudio de la ecología trófica
como consumidor proporciona información de las relaciones
interespecíficas con miembros de su comunidad, con posibles
organismos de importancia comercial en esta misma zona, y detecta
posibles cambios temporales en las mismas. Asimismo, la información
generada ampliará el conocimiento sobre el ecosistema de Bahía de Los
Ángeles, necesario para el marco teórico de la propuesta que se esta
realizando para que sea considerada como Área Natural Protegida.
10

Por otro lado, la pesquería de pulpos en el noroeste de México va
en aumento y es necesario tomar medidas para proteger el recurso. El
desarrollo del cultivo del pulpo es una alternativa viable, la cual se esta
implementando en otros países donde el recurso ha sido
sobreexplotado, y sirve para repoblar las áreas naturales y para su
producción en cultivos integrados (Solís y Chávez, 1985). En este
sentido, se considera que el estudio de la ecología trófica puede ser el
primer paso para la elaboración de dietas naturales o preparadas en
cultivos, como alternativa a posibles casos de sobreexplotación o
cuando la producción no sea suficiente.
HIPÓTESIS
Según los cambios estacionales y ontogenéticos en la dieta ya
mencionados para pulpos del género Octopus (Hernándes-López, 2000;
Smith, 2003), así como estos mismos cambios observados para otros
cefalópodos (Boucher-Rodoni et al., 1987; Rodhouse y Nigmatulin,
1996). Se esperan los siguientes resultados:
Habrá una diversificación de la dieta de acuerdo al incremento en
talla.
Habrá cambios en la composición de la dieta de acuerdo a los
cambios de estación.
Habrá una disminución en la ingestión de alimento durante los
meses de temporada reproductiva.
OBJETIVO
Conocer la ecología trófica de Octopus bimaculatus frente a las
costas de Bahía de Los Ángeles, B.C., México.
OBJETIVOS ESPECÍFICOS
Conocer la composición específica de la dieta de O. bimaculatus
en Bahía de Los Ángeles, B.C.
Analizar la variación del espectro trófico, en función a la talla y
sexo.
11

Determinar la amplitud del espectro trófico de esta especie en
Bahía de los Ángeles.
Determinar traslapamiento entre dietas en función a la talla y el
sexo.
Describir aspectos de comportamiento alimentario.
ÁREA DE ESTUDIO
El golfo de California es una cuenca de evaporación larga y
estrecha (1000 km por 150 km) que tiene comunicación directa con el
Océano Pacífico (Roden y Groves, 1959). Se caracteriza por ser un área
con tasas excepcionalmente altas de productividad primaria, la cual
parece aumentar de sur a norte y ser mayor en la primavera y el
comienzo del verano (Álvarez-Borrego, 1983).
Bahía de los Ángeles está situada en la costa oriental de Baja
California, en la zona de las grandes islas. Situada frente a numerosas
islas pequeñas cercanas (Smith, Ventana, Cabeza de Caballo, Pata
Bota, Flecha, Jorobado, Llave, Cerraja y Los Gemelos) y a una mayor
distancia frente a Isla Ángel de la Guarda (Fig. 3). Presenta 3 canales
profundos en su interior. Está protegida de los vientos del este, aunque
los vientos crean grandes olas que pasan entre los canales de las islas;
la costa oeste está protegida por cerros de granito de la Sierra de San
Borjas. El suelo de la bahía tiene forma de cuenca que va desde los 20
m de profundidad al oeste, hasta los 170-180 al este y al norte (Barnard
y Grady, 1968).
12

Figura 1. Ubicación del área de estudio en el Golfo de California.
MATERIAL Y MÉTODOS.
Análisis del contenido estomacal y de los restos de presas
próximos a los refugios.
Se realizaron muestreos mensuales de O. bimaculatus obtenidos
de la pesca artesanal que se lleva a cabo en la zona submareal de
agosto de 2006 a julio de 2007. Se obtuvieron de 20 a 30 individuos de
ambos sexos de diferentes tallas. De la misma manera, se extrajeron los
restos de presas en zonas contiguas a los refugios donde se les pesca
(conchas de bivalvos, gasterópodos y exoesqueletos de crustáceos con
las posibles marcas características a su depredación). A cada individuo
le acompañó en todos los casos posibles su correspondiente
acumulación de restos de presas así como presas vivas presentes en los
refugios al momento de la captura. Durante los muestreos del invierno
(febrero y marzo) y primavera (abril y junio) de 2007 se hicieron
recolectas de invertebrados de la zona intermareal y submareal con el
fin de conocer cualitativamente algunas de las posibles presas en estos
meses.
Los pulpos se transportaron en frío al laboratorio de
Invertebrados Marinos del Centro Interdisciplinario de Ciencias
Marinas, en La Paz, B.C.S. A cada organismo se le registró el sexo, la
13

longitud total y del manto, así como el peso total, se les extrajo el
estómago y el buche, los cuales se guardaron en un congelador a –20°C.
Se pesó el contenido de los órganos digestivos, y se les asignó un
valor arbitrario de llenado. Con ello se calculó el Índice de Llenado (FI)
(Breiby y Jobling, 1985) en el que 0 es vacío, 1 es 25% de llenado, 2 el
50%, 3 el 75% y 4 lleno. También se calculó el Índice de Peso-Llenado
(FWI) (Quetglas et al., 1999) como sigue:
FWI= peso del estómago /(peso del organismo - peso del estómago)*100
Se asignó un valor arbitrario de estado de digestión de acuerdo a
la descripción de las presas propuesta por Karpov y Cailliet (1978), cuya
escala va de 1 a 4: (1:recién ingerido; 2, poco digerido; 3, digestión
avanzada; 4, muy digerido).
Los contenidos del tracto digestivo se preservaron en alcohol al
70% para la posterior identificación y cuantificación de las presas. La
identificación de las presas, tanto las encontradas en los refugios como
en los tractos digestivos, en la observación directa y de las recolectas de
invertebrados durante invierno-primavera, se realizó con la ayuda de
literatura especializada según el grupo taxonómico: para crustáceos, la
guía FAO para plantas e invertebrados (Hendrickx, 1995), Kertisch y
Bertsch (2007) y el apoyo del personal del Museo de Historia Natural de
la UABCS y del laboratorio de Invertebrados del CICIMAR; para
bivalvos y gasterópodos la guía FAO (Poutiers, 1995b) y los trabajos de
Keen (1971) y Brusca (1985); para gusanos equiuros y sipuncúlidos el
apoyo del personal del Museo de Historia Natural de la UABCS así como
de los trabajos de Fisher (1946), Brusca (1980) y Brusca y Brusca
(2005); para restos de peces se contó con la referencia y apoyo de la
Colección de Peces del CICIMAR.
Se midieron las conchas de bivalvos y gasterópodos (longitud de
la concha, LC) presas con la finalidad de encontrar posibles relaciones
entre tallas del pulpo y su presa, así como relación entre el porcentaje
de perforación por talla y especie presa.
14

Índices cuantitativos para determinación porcentual de las
presas.
Para este fin se utilizaron los siguientes índices: el numérico, el
volumétrico, el de frecuencia de aparición (Cailliet et al., 1996) y
el índice de importancia relativa (Pinkas et al., 1971).
o Método Numérico (% N): es el porcentaje del número de
individuos de una determinada presa con respecto al total de
presas y se obtiene mediante la siguiente relación:
% N = (n/NT)*100
Donde: n es el número total de presas de una determinada
especie o categoría de presa y NT es el número total de presas de todas
las categorías.
o Método de Frecuencia de Aparición (% FA). Con este método se
registra el número de estómagos en los cuales aparece un
determinado tipo de presa. El resultado se expresa como el
porcentaje del valor anterior con respecto al número total de
estómagos con alimento:
% FA= (n/NE)*100
Donde: n es el número de estómagos que presentan un
determinado tipo de alimento y NE es el número total de estómagos con
alimento.
o Índice de Importancia Relativa (% IIR). Este método integra los
índices anteriores de la siguiente forma:
% IIR= (%N)* (%FA)
Índices Ecológicos.
o Índice estandarizado de Levin (Hurlbert, 1978). Se calculó la
amplitud de la dieta, aplicando este índice a los valores absolutos
del Índice Numérico. Los valores van de 0 a 1; un valor menor a
0.6 indica un depredador especialista y mayor a 0.6, indica que
es generalista.
15

Bi= 1/ n-1 (1/ΣPij²)-1)
Bi= Índice de Levin para del depredador i.
Pij= Proporción de la dieta del depredador i que utiliza la presa j.
n= Número de categorías de las presas.
Índice de Morisita Horn (Smith y Zaret, 1982). Se evaluó el traslapo de
dietas entre tallas y sexos. Los valores comprenden de 0 a 1. Valores
mayores a 0.60 reflejan un alto traslapo. De 0.30 a 0.59 se considera
un bajo traslapo.
Cλ= -----------------------------
n
2Σ(Pxi * Pyi) i=1
n n (Σ P xi² + Σ Pyi²) i=1 i=1
Cλ= Índice de Morisita Horn de sobreposición entre depredador x y
depredador y.
Pxi = Proporción de la presa i del total de presas usadas por el
depredador x.
Pyi = Proporción de la presa i del total de las presas usadas por el
depredador y.
n = Número total de presas.
16

RESULTADOS
Se analizaron un total de 288 individuos de ambos sexos de
agosto de 2006 a julio de 2007. De los cuales 73 fueron del verano de
2006, 70 del otoño de 2006, 74 del invierno de 2007 y 71 de la
primavera de 2007. 121 fueron hembras, 132 machos y 31 indefinidos
(Fig. 2).
Se registró el porcentaje de individuos con restos de
alimento y sin restos del mismo, considerando tractos digestivos,
acumulaciones alrededor de sus refugios y presas vivas. 222
individuos se presentaron con alimento, encontrando un menor
porcentaje de estos durante los meses del verano (Fig.3).
05
1015202530354045
Verano 06 Otoño 06 Invierno 07 Prim 07
Nú
mer
o d
e in
div
idu
os
HembrasMachosSin Definir
N=73 N= 70 N=74 N=71
Figura 2. Distribución de la muestra por estación
0% 20% 40% 60% 80% 100%
Verano 06
Otoño 06
Invierno 07
Primavera 07
Número de individuos
Con Alimento Sin Alimento
Figura 3. Porcentaje de individuos con o sin alimento por estación.
17

Según el Índice de Peso-Llenado, se registraron valores más bajos
también para los meses del verano (Fig. 4).
0
0,2
0,4
0,6
0,8
Verano 06 Otoño 06 Invierno 07 Primav 07
Índ
ice
de
Pes
o-L
len
ado
Figura 4. Índice de Peso-Llenado promedio según la estación.
Se identificaron un total de 85 item o categorías de presa a
diferentes niveles de clasificación, de las cuales 44 fueron a especie, 12
a género, 12 a familia, 1 a superfamilia, 1 a infraorden, 2 a orden, 1 a
infraclase, 2 a suclase, 7 a clase, 1 a subphylum y 2 a phylum (Fig. 5).
Los 85 item pertenecieron a uno de 8 phyla (Arthropoda,
Mollusca, Echiura, Sipuncula, Annelida, Echinodermata, Ectoprocta,
Chordata) registrados en total, de los cuales, todos fueron detectados
por los tractos digestivos, 4 por las acumulaciones y 3 por observación
directa (Tabla 1).
18
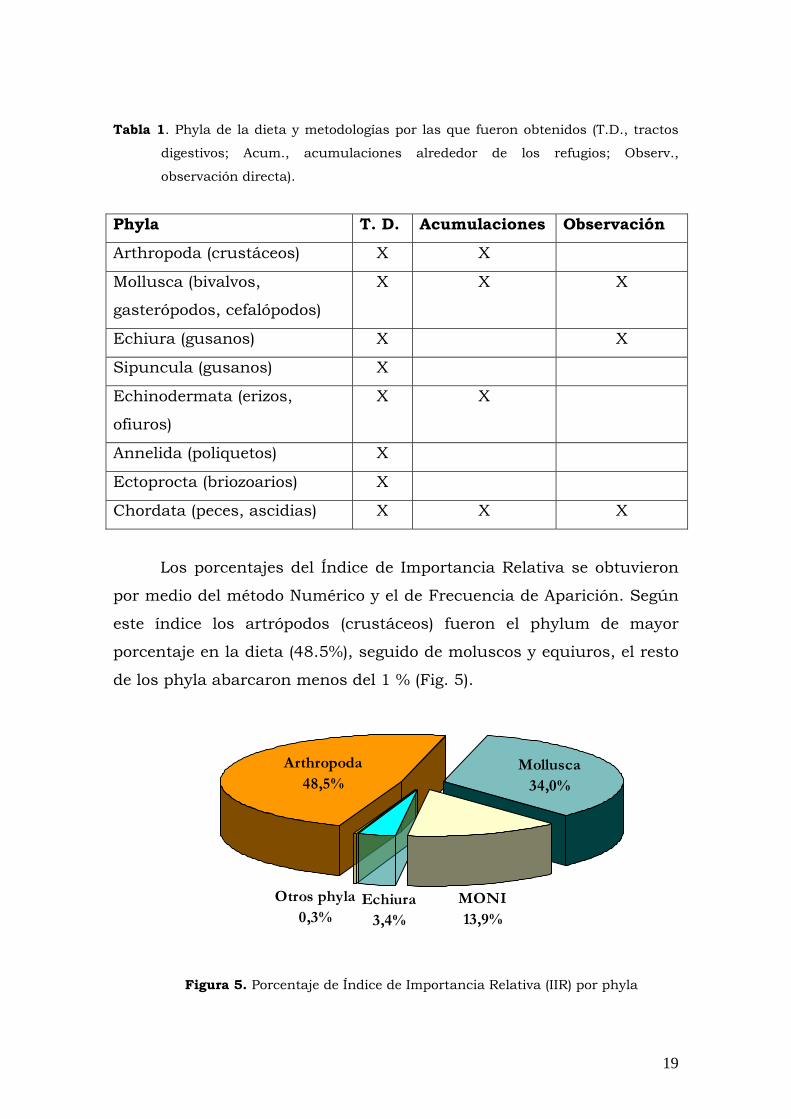
Tabla 1. Phyla de la dieta y metodologías por las que fueron obtenidos (T.D., tractos
digestivos; Acum., acumulaciones alrededor de los refugios; Observ.,
observación directa).
Phyla T. D. Acumulaciones Observación
Arthropoda (crustáceos) X X
Mollusca (bivalvos,
gasterópodos, cefalópodos)
X X X
Echiura (gusanos) X X
Sipuncula (gusanos) X
Echinodermata (erizos,
ofiuros)
X X
Annelida (poliquetos) X
Ectoprocta (briozoarios) X
Chordata (peces, ascidias) X X X
Los porcentajes del Índice de Importancia Relativa se obtuvieron
por medio del método Numérico y el de Frecuencia de Aparición. Según
este índice los artrópodos (crustáceos) fueron el phylum de mayor
porcentaje en la dieta (48.5%), seguido de moluscos y equiuros, el resto
de los phyla abarcaron menos del 1 % (Fig. 5).
Arthropoda48,5%
Mollusca34,0%
Echiura3,4%
Otros phyla0,3%
MONI13,9%
Figura 5. Porcentaje de Índice de Importancia Relativa (IIR) por phyla
19

Según el IIR, el item de mayor importancia en la dieta son los
cangrejos xanthoideos (46%), seguido de materia orgánica no
identificada (MONI; 14%), Megapitaria squalida (12%), Trigoniocardia
biangulata (6%), Echiurus spp. (3%), Glycimeris gigantea (3%), Anadara
multicostata (3%), Pecten vogdesi (2%) y otras presas (11%) (Fig. 6; Tabla
2).
Según las recolectas de invertebrados en la zona intermareal y
submareal que se hicieron durante los muestreos de invierno y
primavera de 2007, se registraron las siguientes taxa: Glyptoxanths
meandricus, Eurypanopeus spp., Speleolophorus schmitti, Petrochirus
californiensis, de los cuales el primero estuvo presente en los 4 meses
de muestreo.
Echiurus spp.Trigoniocardia
biangulata
Glycimeris gigantea
Anadara multicostata
Pecten vogdesi
Otras presas
Megapitaria squalida
MONIXanthoidea
Figura 6. Presas más importantes según el porcentaje del
Índice de Importancia Relativa (IIR).
20

Tabla 2. Item con sus respectivos porcentajes de Frecuencia de Aparición (FA),
Numérico (N) e Índice de Importancia Relativa (IIR).
% FA % N % IIR ARTHOPODA Crustacea n.i. 0,5 0,1 0,00Copepoda Harpacticoidea 0,5 0,1 0,00Ostracoda Ostracoda 0,9 0,1 0,01Decapoda Decapoda (camarón) 0,5 0,4 0,01 Alpheidae 6,8 0,1 0,03 Decapoda (cangrejo) 0,5 1,0 0,03 Diogenidae 1,8 0,1 0,01 Lithodidae 4,5 1,9 0,50 Brachyura 1,8 0,3 0,03 Majidae 0,5 0,1 0,00 Portunidae 9,5 2,7 1,52 Portunus xantusii 0,5 0,1 0,00 Speleophorus schmitti 0,5 0,1 0,00 Xanthoidea 28,4 27,6 46,33MOLLUSCA Bivalvia 0,9 0,1 0,01Bivalvia Arcidae 0,5 0,1 0,00 Anadara multicostata 13,5 3,5 2,78 Myrakeena angelica 0,9 0,2 0,01 Argopecten circularis 6,8 1,2 0,46 Barbatia reeveana 0,5 0,1 0,00 Cardita affinis 1,4 0,4 0,03 Veneridae 1,4 0,3 0,02 Chione californiensis 8,1 1,5 0,74 Chione pulicaria 0,9 0,1 0,01 Chione tumens 0,9 0,1 0,01 Chione undatella 4,5 0,7 0,19 Dosinia ponderosa 8,6 1,9 0,95 Eucrassatella digueti 1,4 0,3 0,02 Gari regularis 2,7 0,5 0,07 Glycimeridae 0,9 0,1 0,01 Glycimeris gigantea 13,1 3,9 2,99 Glycimeris multicostata 8,6 2,8 1,40 Hiatellidae 0,5 0,1 0,00 Macoma spp. 0,9 0,1 0,01 Megapitaria auriantiaca 2,3 0,3 0,04 Megapitaria squalida 23,4 8,6 11,97 Mytella guayanensis 2,7 0,4 0,06 Nudipecten subnodosus 0,9 0,2 0,01 Papyridea aspersa 8,1 1,4 0,65 Pecten vogdesi 11,7 2,7 1,88 Pinna rugosa 3,2 0,5 0,08 Pitar spp. 0,9 0,1 0,01 Placuanomia cumingii 0,9 0,1 0,01
21
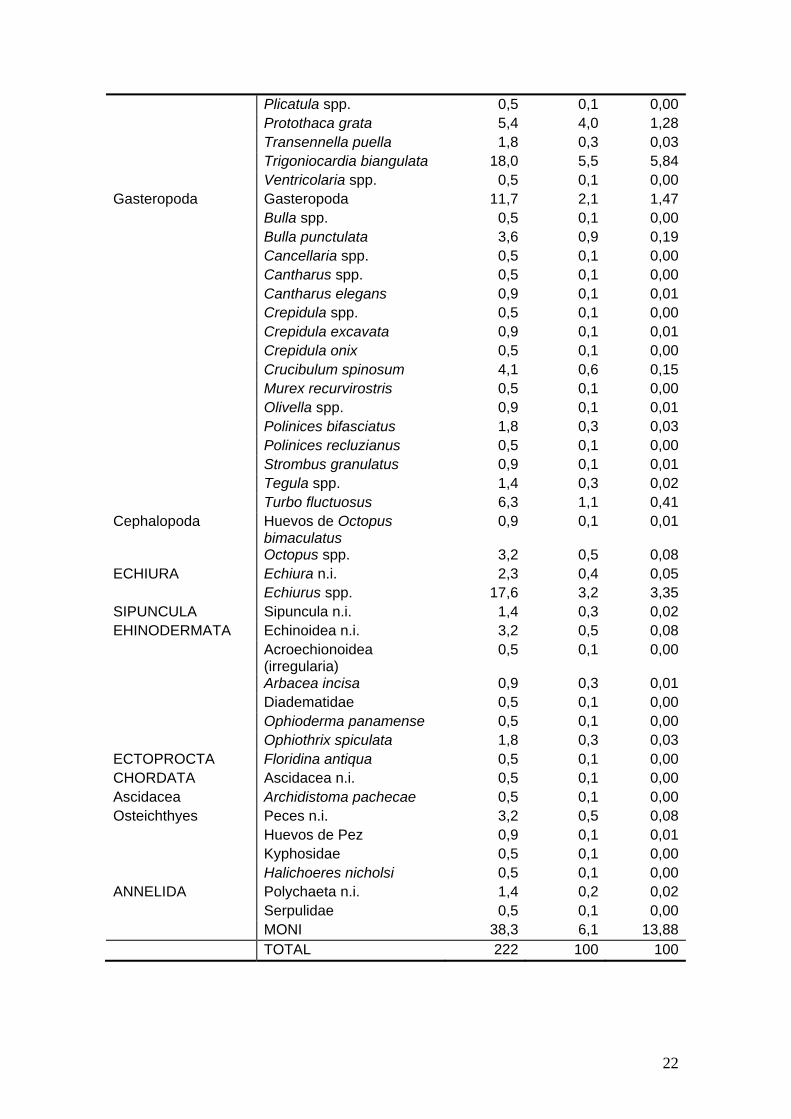
Plicatula spp. 0,5 0,1 0,00 Protothaca grata 5,4 4,0 1,28 Transennella puella 1,8 0,3 0,03 Trigoniocardia biangulata 18,0 5,5 5,84 Ventricolaria spp. 0,5 0,1 0,00Gasteropoda Gasteropoda 11,7 2,1 1,47 Bulla spp. 0,5 0,1 0,00 Bulla punctulata 3,6 0,9 0,19 Cancellaria spp. 0,5 0,1 0,00 Cantharus spp. 0,5 0,1 0,00 Cantharus elegans 0,9 0,1 0,01 Crepidula spp. 0,5 0,1 0,00 Crepidula excavata 0,9 0,1 0,01 Crepidula onix 0,5 0,1 0,00 Crucibulum spinosum 4,1 0,6 0,15 Murex recurvirostris 0,5 0,1 0,00 Olivella spp. 0,9 0,1 0,01 Polinices bifasciatus 1,8 0,3 0,03 Polinices recluzianus 0,5 0,1 0,00 Strombus granulatus 0,9 0,1 0,01 Tegula spp. 1,4 0,3 0,02 Turbo fluctuosus 6,3 1,1 0,41Cephalopoda Huevos de Octopus
bimaculatus 0,9 0,1 0,01
Octopus spp. 3,2 0,5 0,08ECHIURA Echiura n.i. 2,3 0,4 0,05 Echiurus spp. 17,6 3,2 3,35SIPUNCULA Sipuncula n.i. 1,4 0,3 0,02EHINODERMATA Echinoidea n.i. 3,2 0,5 0,08 Acroechionoidea
(irregularia) 0,5 0,1 0,00
Arbacea incisa 0,9 0,3 0,01 Diadematidae 0,5 0,1 0,00 Ophioderma panamense 0,5 0,1 0,00 Ophiothrix spiculata 1,8 0,3 0,03ECTOPROCTA Floridina antiqua 0,5 0,1 0,00CHORDATA Ascidacea n.i. 0,5 0,1 0,00Ascidacea Archidistoma pachecae 0,5 0,1 0,00Osteichthyes Peces n.i. 3,2 0,5 0,08 Huevos de Pez 0,9 0,1 0,01 Kyphosidae 0,5 0,1 0,00 Halichoeres nicholsi 0,5 0,1 0,00ANNELIDA Polychaeta n.i. 1,4 0,2 0,02 Serpulidae 0,5 0,1 0,00 MONI 38,3 6,1 13,88 TOTAL 222 100 100
22

Aunque se presentaron cambios en la composición de la dieta
estacional, durante todas las estaciones se encontraron cangrejos
xanthoideos. Durante el verano se alimentaron de gusanos equiuros,
cangrejos portúnidos, almeja voladora y peces; durante otoño
incrementaron los bivalvos y los xanthoideos; en el invierno
incrementaron los bivalvos en especies y abundancia, también
consumieron más gasterópodos; y finalmente durante la primavera
tuvieron un notable incremento en el consumo de cangrejos
xanthoideos, los cuales abarcaron más del 80% de la dieta según el IIR
(Fig. 7).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Ver 06 Oto 06 Inv 07 Pri 07
IIR
Otros items
Osteichthyes
Gasteropoda n.i.
Prothotaca grata
Pecten vogdesi
Glycimeris multicostata
Glycimeris gigantea
Dosinia ponderosa
Anadara multicostata
Trigoniocardia biangulata
Megapitaria squalida
Xanthoidea
Portunidae
Lithodidae
Decapoda Figura 7. Porcentaje del Índice de Importancia Relativa comparativo
por estaciones (verano de 2006: Ver 06; Otoño de 2006: Oto 06;
invierno de 2007: Inv 07; primavera de 2007: Pri 07).
Se calculó el IIR de las presas en la dieta por sexos, a pesar de
que presentan una dieta similar en la composición, es notable el amplio
23

consumo de xanthoideos de los machos y en hembras una dieta más
diversificada en proporción y composición de las presas (Fig. 8).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
MACHOS HEMBRAS
IIR
Otros items
Echiurus spp.
Gasteropoda
Anadara multicostata
Trigoniocardia biangulata
Megapitaria squalida
Glycimeris gigantea
Portunidae
Xanthoidea
Figura 8. Presas más importantes según el Índice
de Importancia Relativa por sexos.
Se calculó el IIR para dos intervalos de talla según la talla de
primera madurez (124 mm LM para machos y 147 mm LM para
hembras), quedando como inmaduros todos los menores a 124 mm, que
es la talla mínima para ambos.
Los cangrejos xantoideos fueron la presa de mayor importancia
para ambos intervalos de talla. Los pulpos inmaduros se alimentaron de
una mayor cantidad de presas, destacando bivalvos de menor talla
como Trigoniocardia biangulata y gusanos equiuros. Los maduros
incrementaron el consumo de xanthoideos y consumieron bivalvos de
mayor talla como Megapitaria squalida (Fig. 9).
24

0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Inmaduros Maduros
IIR
Otros items
Echiurus spp.
Gasteropoda
Trigoniocardia biangulata
Protothaca grata
Pecten vogdesi
Glycimeris gigantea
Megapitaria squalida
Portunidae
Xanthoidea
Figura 9. Presas más importantes según el Índice de
Importancia Relativa por intervalos de madurez gonádica.
Indices ecológicos
Los valores del índice de Morisita-Horn señala un alto traslapo
entre dietas de hembras y machos (0.8), así como de inmaduros y
maduros (0.74).
El índice estandarizado de Levin dio valores de dieta especialista
para el total, para machos, inmaduros y maduros (0.003), así como
para hembras (0.004) Lo anterior indica que a pesar de que se
alimentan de 85 categorías de presa diferentes, principalmente lo hacen
de unas pocas.
Análisis de tallas y perforación de las conchas de bivalvos y
gasterópodos.
Se registró un total de 643 conchas recientes de bivalvos y
gasterópodos obtenidas de las acumulaciones de restos duros de los
refugios. 591 de bivalvos de 29 taxa y 52 de gasterópodos de 11 taxa.
Para este análisis los pulpos se agruparon en tres intervalos de talla
arbitrarios: chicos (10-105 mm LM), medianos (106-140 mm LM) y
grandes (141-190 cm LM).
25

Considerando el espectro completo de tallas de las conchas, se
observó que pulpos de todas las tallas se alimentaron casi de todos las
tamaños de moluscos con concha aunque se observó cierta
especialización por algunas tallas; todos se alimentaron principalmente
de conchas de 3 a 5 cm (LC) pero los pulpos diversificaron su dieta
hasta conchas de 9 cm ó más, por otro lado los pulpos pequeños se
concentraron las tallas antes mencionadas (3-5 cm LC). Los pulpos
medianos mostraron un comportamiento intermedio entre el de chicos y
el de grandes (Fig. 10).
0
10
20
30
40
50
60
1 2 3 4 5Longitud de la Concha (cm)
Por
cen
taje
(N
) d
e p
ulp
os
dep
red
ador
es
Chicos
Medianos
Grandes
Figura 10. Distribución de tallas de conchas encontradas en refugios de
pulpos chicos (10-105 mm LM), medianos (106-140 mm LM) y grandes
(141-190 cm LM).
No se encontró un porcentaje de perforación diferencial entre
pulpos chicos, medianos y grandes, ya que todos perforaron alrededor
del 40 % de las conchas (Fig. 11).
Se realizaron pruebas de correlación entre las tallas de las
conchas de las 8 especies más abundantes en la dieta (todos bivalvos) y
las tallas de los pulpos depredadores, y no se encontró una correlación
significativa entre estas (coeficientes de correlación: 0.06 a 0.1) (Fig. 12
y 13). Tampoco se encontró una distribución distintiva de las conchas
perforadas (puntos anaranjados) y las no perforadas (puntos azules) en
el espectro de tallas de las conchas en relación al depredador.
26

0%
20%
40%
60%
80%
100%
Chicos Medianos Grandes
Núm
ero
de c
onch
asNo perforadasPerforadas
Figura 11. Porcentaje de perforación por intervalos de talla
de los pulpos . (Chicos 10-105 mm LM, medianos
106-140 mm LM y grandes 141-190 cm LM).
Anadara multicostata
R2 = 0,01260
40
80
120
160
200
0 20 40 60 80 100
Glycimeris gigantea
R2 = 0,15970
40
80
120
160
200
0 20 40 60 80 100
Trigoniocardia biangulata
R2 = 0,05410
40
80
120
160
200
0 20 40 60 80 100
Megapitaria squalida
R2 = 0,01270
40
80
120
160
200
0 20 40 60 80 100
Lon
gitu
d d
e m
anto
(m
m)
Figura 12. Correlación de tallas concha/depredador de cuatro de
los 8 bivalvos más consumidos de la dieta.
27

Pecten vogdesi
R2 = 0,05360
40
80
120
160
200
0 20 40 60 80 100
Protothaca grata
R2 = 0,00680
40
80
120
160
200
0 20 40 60 80 100
Dosinia ponderosa
R2 = 0,0410
40
80
120
160
200
0 20 40 60 80 100
Glicymeris multicostata
R2 = 0,04290
40
80
120
160
200
0 20 40 60 80 100
Lon
gitu
d d
el m
anto
(m
m)
Figura 13. Correlación de tallas concha/depredador de
Longitud de la Concha (mm)
cuatro de los 8 bivalvos más consumidos de la dieta.
Por último se obtuvo el porcentaje de perforación para las mismas
8 especies de bivalvos y se encontró que las conchas de Dosinia
ponderosa son las más perforadas (95%) y Prothotaca grata también fue
una de las más perforadas (58%), Megapitaria squalida y Trigoniocardia
biangulata (que son las más abundantes en la dieta) se encontraron
alrededor del 40% de perforación y el resto con menos del 20% de
perforación para cada especie (Fig. 14).
28

0%
20%
40%
60%
80%
100%
Dosinia p
onde
rosa
Prototha
ca gra
ta
Trigon
iocard
ia bian
gulata
Megapit
aria s
qualid
a
Glycim
eris g
igantea
Pecten
vogd
esi
Anadar
a mult
icosta
ta
Glycim
eris m
ultico
stata
N
No perforadas
Perforadas
Figura 14. Porcentaje de perforación por especie de los
8 bivalvos más abundantes de la dieta.
CONCLUSIONES PARCIALES.
• O.bimaculatus se alimenta principalmente de cangrejos
xanthoideos y bivalvos.
• Se presentó mayor llenado de tractos digestivos durante
primavera y menor en el verano.
• Hay un incremento en el consumo de cangrejos en la
Primavera, durante la época de maduración.
• Las hembras se alimentan de mayor variedad de presas y los
machos principalmente de xanthoideos.
• Presenta una dieta especialista y con alto traslapamiento entre
sexos y tallas según madurez.
• No existe una relación significativa entre la longitud de manto
de los pulpos y la longitud de la concha de los moluscos de la
dieta.
• Hay diferencias en el porcentaje de perforación según la
especie presa.
29

Impacto
Los datos obtenidos en el presente proyecto amplían el
conocimiento de los hábitos alimenticios de esta especie de
pulpo. Los datos obtenidos servirán para mejorar los intentos de
cultivo e este recurso tan importante en México y en el mundo.
BIBLIOGRAFÍA
Aguilar-Chávez, S.G. 1995. Estudio biológico del Pulpo Octopus sp.
(Cephalopoda: Octopoda) en la Costa Sur de Jalisco. Tesis de
Licenciatura. Universidad de Guadalajara, México. 60 pp.
Álvarez-Borrego, S. 1983. Golfo of California, México. En: B.H. Ketchum
(ed.). pp. 427-449. Ecosystems of the World, vol. 26, Estuaries
an Enclosed Seas. Elsevier Science Publications. Netherlands.
Alejo-Plata, M.C. y G. Cerdenares-Ladrón de Guevara. 2002.
Caracterización de la pesca de Pulpo (Cephalopoda: Octopoda)
en las Costas de Oaxaca, México. En: Mem. I Foro Científico de
Pesca Ribereña. Instituto Nacional de la Pesca Centro Regional
de Investigaciones Pesqueras Guaymas, Son.
Ambrose, R.F. 1982. Shelter Utilization by the Molluscan Cephalopod
Octopus bimaculatus. Marine Ecology- Progress Series. 7:67-73.
Ambrose, R.F. 1984. Food preferences, prey availability and the diet of
Octopus bimaculatus, Verril. Journal of Experimental Marine
Biology and Ecology. 77: 29-44.
Ambrose, R.F. 1986. Effects of octopus predation on motile
invertebrates in a rocky subtidal community. Marine Ecology-
Progress Series. 30: 261-273.
Ambrose, R.F. 1988. Population Dynamics of Octopus bimaculatus:
influence of life history patterns, synchronous reproduction and
recruitment. Malacology. 29 (1): 23-39.
Bachok, Z., M.I. Mansor and R.M. Noordin. 2004. Diet composition and
food habits of demersal and pelagic marine fishes from
30

Terengganu waters, east coast of Peninsular Malaysia. NAGA,
World Fish Center Quarterly. 27: Jul-Dec.
Boucher-Rodoni, R., E. Boucaud-Camou and K. Mangold. 1987.
Feeding and digestion. pp. 85-108. En: P.R. Boyle (ed.).
Cephalopod Life Cycle. Vol. II. Comparative Reviews. Academic
Press. England.
Breiby, A. and M. Jobling. 1985. Predatory role of the flying squid
(Todarodes sagittatus) in Norwegian waters. North Atlantic
Fisheries Organization (NAFO) Scientific Council Studies. 9:
125-132.
Brusca, R. C. Common Intertidal Invertebrates of the Gulf of California.
University of Arizona Press. Arizona, E.U.A.1582 pp.
Brusca R.C. y G.J. Brusca. 2005. Invertebrados. McGraw Hill-
Interamericana. España. 1005 pp.
Cailliet, M.G., M.S. Love and A.W. Ebeling. 1996. Fishes. A field and
laboratory manual on their structure identification and natural
history. Wadsworth Publishing Company. 194 pp.
Cardoso, F., P. Villegas y C. Estrella. 2004. Observaciones sobre la
biología de Octopus mimus (Cephalopoda: Octopoda) en la costa
peruana. Revista Peruana de Biología. 11(1): 45 – 50.
Castro, J.J. 1993. Feeding ecology of chub mackerel (Scomber
japonicus) in Canary Islands area. South African Journal of
Marine Science. 13:323-328.
Castro, J.J. and V. Hernández-García. 1995. Ontogenic changes in
mouth structures, foraging behavior and habitat use of Scomber
japonicus and Illex coindetti. Scientia Marina. 59 (3-4): 347-355.
Clarke, M.R. 1987. Cephalopod biomass estimation from predation. pp.
221-238. En: P.R. Boyle (ed.). Cephalpod Life Cycles Vol 2.
Academic Press. England.
Clarke, M.R. 1996. Cephalopod as prey. III. Cetaceans. pp 1053-1065.
En: M. R. Clarke (ed.) The Role of Cephalopods in the world´s
oceans. Philosophical Transactions of the Royal Society of
London B Biological Sciences Science.
31

CONABIO (Comisión Nacional para el Conocimiento y Uso de la
Biodiversidad). 1996. Regiones prioritarias para la conservación
en México. Resultados del taller de consulta realizado los días
26, 27 y 28 de febrero de 1996. México, D.F. 45 pp.
Cortez, T., B.G. Castro and A. Guerra. 1995. Feeding dynamics of
Octopus mimus (Mollusca: Cephalopoda) in northern Chile
waters. Marine Biology. 123: 497-503
Croxall, J.P. and P.A. Prince, 1996. Cephalopod as prey. I. Seabirds. pp
1023-1044. En: M. R. Clarke (ed.). The Role of Cephalopods in
the world´s oceans. Philosophical Transactions of the Royal
Society of London B Biological Sciences Science. 351 pp.
Daneman, G. y C. Peynador. 2004. Estudio Técnico Justificativo para la
creación del Parque Nacional “Bahía de Los Ángeles”, Baja
California. Versión preliminar para revisión (séptima edición).
Comunidad de Bahía de los Ángeles; PRONATURA Noroeste;
Centro de Investigación Científica y Educación Superior de
Ensenada; Comunidad y Biodiversidad A.C.; Conservación
Internacional de México A.C.; Departamento de Fauna Silvestre,
Pesquerías y Biología de la Conservación de la Universidad de
California en Davis; Ejido Ganadero y Turístico “Tierra y
Libertad”, Grupo de Ecología y Conservación de Islas A.C.;
Investigación y Conservación de Mamíferos Marinos de
Ensenada A.C.; ProEsteros, Lagunas y Marismas de las
Californias S.C.; Sociedad de Historia Natural Niparajá A.C.;
Wild Coast; WWF-Programa México. 156 pp y siete anexos.
Dodge, R. and Scheel, D. 1999. Remains of the prey, recognizing the
midden piles of Octopus dofleini (Wulker). Veliger. 42: 260-266.
Doyle, P., D. T. Donovan and M. Nixon. 1994. Phylogeny and
systematics of the Coleoidea. Univ. Kansas Paleont. Contrib. No.
5: 1-15.
Enríquez-Andrade, R. y G. Danemann. 1998. Identificación y
establecimiento de prioridades para las acciones de
conservación y oportunidades de uso sustentable de los
32

recursos marinos de la Península de Baja California. Reporte
técnico de proyecto. PRONATURA Península de Baja California.
77 pp.
FAO. 2000. Total production 1950-1998. FAO Homepage (Former
Yearbook fisheries statistics “Catches and Landings”).
Ferron, A. and W.C. Leggend. 1994. An appraisal of conditions
measures for marine fish larvae. Advances in Marine Biology.
30: 217-303.
Fitch, J.E. 1969. Fossil records of certain shooling fishes of the
California Current system. California Cooperative Oceanic
Fisheries Investigations (CalCOFI) Reports. 13: 71-80.
Fisher, W.K. 1946. Equiuroid worms of the North Pacific Ocean. Pp
281-310. En: Proceedings of the United States National
Museum. Smithsonian Museum. Washington, E.U.A.
Forsythe, J. W. and W.F. Van Heukelem. 1987. Growth. pp 135-156.
En: P.R. Boyle (ed.). Cephalopod Life Cycles, Vol. II: Comparative
Reviews. Academic Press. London, U.K. 135-156.
Gerking, Sh.D. 1994. Feeding Ecology of Fish. Academic Press. U.S.A.
416 pp.
Ghiretti, F. 1959. Cephalothoxin: the crab-paralyizing posterior salivary
glands in the cephalopods. Nature. 182: 1192-1193.
Guerra, A. 1992. Fauna Ibérica. Mollusca: Cephalopoda. Museo
Nacional de ciencias Naturales. Consejo Superior de
Investigaciones Científicas. Madrid. 627 pp.
Guerra-Sierra, A y J.L. Sánchez-Lizaso. 1998. Fundamentos de
explotación de recursos vivos marinos. Editorial Acribia, S.A.
Zaragoza, España. 249 pp.
Guerra, A., Nixon, M., 1987. Crab and mollusc shell drilling by Octopus
vulgaris (Mollusca: Cephalopoda) in the Ria de Vigo (north-west
Spain). Journal of Zoology 211, 515–523
Hanlon, R. and J. B. Messenger. 2002. Cephalopod Behavior.
Cambridge University Press. U.S.A. 232 pp.
33

Harvey, J., T. Loughlin, M. Perez y D. Oxman. 2000. Relationship
between Fish Size and Otolith Length for 63 species of Fishes
from the Eastern North Pacific Ocean. NOAA Technical Report.
NMFS 150. E.U.A. 38 pp.
Hernández-Flores, A., M. Solís-Ramírez, J.C. Espinoza-Méndez, R.
Mena-Aguilar y F. Ramírez-Gil. 2000. Pesquerías del Golfo de
México y Mar Caribe: Pulpo. pp 615-630. En: Cisneros Mata
M.A., L.F. Meléndez-Moreno, E. Zárate-Becerra, T.Gaspar-
Dillanes, L.C. López-González, C. Saucedo-Ruíz, Javier Tovar-
Avila (eds.): Sustentabilidad y pesca responsable en México,
Evaluación y Manejo, 1999-2000. Instituto Nacional de la Pesca.
Hernández-López, J.L. 2000. Biología, ecología y pesca del pulpo común
(Octopus vulgaris, Cuvier 1797) en aguas de Gran Canaria. Tesis
Doctoral. Universidad de Las Palmas de Gran Canaria. 210 pp.
Hendrickx, M.E.. 1995. Cangrejos. pp.565-636. En: Fischer, W, F.
Krupp, W. Schneider, C. Sommer, K.E. Carpenter y V.H. Niem.
(eds.). Guía FAO para la identificación de especies para los fines
de la pesca. Pacífico Centro-Oriental. Vol. I. Roma, FAO. 1813
pp.
Villalobos-Hirriart, J.L., J.C. Nates-Rodríguez, A. Cantú-Díaz-Borrego,
M.D. Valles-Martínez, P. Florez-Hernández, E. Lira-Fernández,
P. Smidtsdorf-Valencia. 1989. Listados faunísticos de México I.
Crustáceos estomatópodos y decápodos intermareales de las
Islas del Golfo de California, México. UNAM. México. 114 pp.
Hochberg, F.G. and W.G. Fields. 1980. Cephalopods: the squids and
octopuses. Chapter 17. pp 429-444. En : Morris R., Abbot, D.
and Haderlie, E. (eds.). Intertidal invertebrates of California.
Stanford Un. Press. U.S.A.
Hochberg, F.G. 1980. Class Cephalopoda. pp 201-204. En: R.C. Brusca
(ed.). Common Invertebrates of the Gulf of California. The Univ.
of Arizona Press. U.S.A.
Hurlbert, S.H. 1978. The measurement of the niche overlap and some
relatives. Ecology. 59: 67-77.
34

Karpov, K.A. and G.M. Cailliet. 1978. Feeding Dynamics of Loligo
opalescens. Pages 45-65. In Recksiek, C.W. and H.W. Frey, ed.
Biological, Oceanographic, and Acoustic Aspects of the Market
Squid, Loligo opalescens Berry. State of California The
Resources Agency Department of Fish and Game Fish Bulletin
169.
Keen, M. 1971. Sea Shells of Tropical West America. Marine Mollusks
from California to Peru. Stanford Univ. Press. U.S.A. 1064 pp.
Kertish, A. y H. Bertsch. 2007. Sea of Cortez. Marine Invertebrates. A
Guide for the Pacific Coast, México to Perú. Sea challengers.
California, E.U.A. 124.
Leyva-Villarreal, M., S. Osuna-Marroquín, A. Ley-Montoya, F.
Cervantes-Galáviz and J. Quiñónez-Cruz. 1987. Contribución al
conocimiento biológico del Pulpo Octopus sp. en la Bahía de
Mazatlán, Sinaloa, México. Memoria de Servicio Social de
Licenciatura. Universidad Autónoma de Sinaloa. 73 pp.
Markaida, U. 2005. Cephalopods in the Diet of Swordfish (Xiphias
gladius) Caught off the West Coast of Baja California, Mexico.
Pacific Science. 59 (1): 25-41.
Mather, J.A. 1991. Foraging, feeding and prey remains in middens of
Octopus vulgaris. Journal of Zoology. London. 224: 27-39.
Mather, J.A., Nixon, M., 1990. Octopus vulgaris drills Chiton. Journal of
Cephalopod Biology 1 (2), 113–116.
Millen, S. V. and H. Bertsch. 2000. Three new species of dorid
nudibranchs from southern California, USA, and the Baja
California Peninsula, Mexico. The Veliger 43(4):354-366.
Morales-Bojórquez, E., A. Hernández-Herrera, M.O. Nevárez-Martínez,
G.I. Rivera-Parra, A. Balmori-Ramírez, C. Cervantes-Valle and E.
Miranda-Mier. 2000. Pesquerías del Pacífico: Calamar. pp 97-
117. En: Cisneros Mata, M.A., L.F. Meléndez-Moreno, E. Zárate-
Becerra, T.Gaspar-Dillanes, L.C. López-González, C. Saucedo-
Ruíz, Javier Tovar-Avila (eds.). Sustentabilidad y pesca
35

responsable en México, Evaluación y Manejo, 1999-2000.
Instituto Nacional de la Pesca.
Nixon, M. 1987. Cephalopod diets. pp 201-217. En: P.R. Boyle (ed.)
Cephalopod Life Cycle. Vol. II. Comparative Reviews. Academic
Press. England.
Nixon, M., Boyle, P.R., 1982. Hole-drilling in crustaceans by Eledone
cirrhosa (Mollusca: Cephalopoda). Journal of Zoology 196, 439–
444.
Nixon, M., Maconnachie, E., 1988. Drilling by Octopus vulgaris
(Mollusca: Cephalopoda) in the Mediterranean. Journal of Zoology
216, 687–716.
Pecl, G.T., N. A. Moltschaniwskyj . S. R. Tracey and A. R. Jordan. 2004.
Inter-annual plasticity of squid life history and population
structure: ecological and management implications. Oecologia.
139: 515–524
Piatkowski, U., G.J. Pierce and M. Morais de Cunha. 2001. Impact of
cephalopods in the food chain and the interaction with the
environment and fisheries: an overview. Fisheries Research. 52:
5-10.
Pinkas, L., M.S. Oliphant and I.L.K Iverson. 1971. Food habits of
albacore, bluefin tuna and bonito in California waters. Fisheries
Bulletin. California, U.S.A. 152: 105.
Poutiers, J.M. 1995a. Bivalves. pp. 99-222. En: Fischer, W, F. Krupp,
W. Schneider, C. Sommer, K.E. Carpenter y V.H. Niem. (eds.).
Guía FAO para la identificación de especies para los fines de la
pesca. Pacífico Centro-Oriental. Vol. I. Roma, FAO. 1813 pp.
Poutiers, J.M. 1995b. Gasterópodos. pp. 223-297. En: Fischer, W, F.
Krupp, W. Schneider, C. Sommer, K.E. Carpenter y V.H. Niem.
(eds.). Guía FAO para la identificación de especies para los fines
de la pesca. Pacífico Centro-Oriental. Vol. I. Roma, FAO. 1813
pp.
Quetglas, A., F. Alemany, A. Carbonell, P. Merilla and P. Sánchez. 1998.
Biology and shery of Octopus vulgaris Cuvier, 1797, caught by
36

trawlers in Mallorca (Balearic Sea, Western Mediterranean).
Fisheries Research. 36 : 237-249.
Quetglas, A. F Alemany, A. Carbonell, P. Merella, P. Sánchez. 1998.
Biology and Fishery of Octopus vulgaris Cuvier, 1797, caught by
trawlers in Mallorca (Balearic Sea, Western Mediterranean).
Fisheries Research 36. 237-249.
Quetglas, A., F. Alemany, A. Carbonell, P. Merella and P. Sánchez.
1999. Diet of the European flying squid Todarodes sagittatus
(Cephalopoda: Ommastrephidae) in the Balearic Sea (western
Mediterranean). Journal Marine Biology Assessment. United
Kingdom. 79:479-486.
Quetglas, A., M. González and I. Franco. 2005. Biology of the upper-
slope cephalopod Octopus salutii from the western
Mediterranean Sea. Marine Biology. 146: 1131–1138.
Rathjen, W.F. 1991. Cephalopod capture methods: an overview. Bulletin
of Marine Science. 49 (1-2): 494-505.
Roden, G.I. and G.W. Groves. 1959. Recent oceanographic investigation
in the Gulf of California. Journal of Marine Research. 18: 177-
183.
Rodhouse, P.G. y Ch.M. Nigmatullin. 1996. Role as Consumers.
Philosophical Transactions: Biological Sciences. 351. Issue
1343. The Role of Cephalopods in the world´s oceans. 1003-
1022.
Roper, C.F. E., M.J. Sweeney and F.G. Hocchberg. 1995. Cephalopoda.
pp. 306-535. En: Fischer, W, F. Krupp, W. Schneider, C.
Sommer, K.E. Carpenter and V.H. Niem (eds.). Guía FAO para la
identificación de especies para los fines de la pesca. Pacífico
Centro-Oriental. Vol. I, II y III. Roma, FAO.
Ridley, M. 1995. Animal Behavior. Blackwell Scientific Publications.
Boston, U.S.A. 288 pp.
Sánchez-Cruz, Y. 2006. Alimentación y Reproducción del pulpo Octopus
hubbsorum (Cefalopoda: Octopoda) en Puerto Ángel, Oaxaca,
37

durante el periodo 2002-2003. Tesis Licenciatura. Universidad
del Mar. Oaxaca.
Segura-Zarzosa, J. C., L. A. Abitia-Cárdenas and F. Galván-Magaña.
1997. Observations on the feeding habits of the shark
Heterodontus francisci Girard 1854 (Chondrichthyes:
Heterodontidae), in San Ignacio Lagoon, Baja California Sur,
México. Ciencias Marinas. 23:111–128.
Serrano, G. S.J. 1992. Dispersión de larvas de bivalvos y seston por
corrientes costeras, dentro del sistema Bahía de los Angeles, B.
C., México. Tesis de Maestría. Centro de Investigaciones
Estudios Superiores de Ensenada. Ensenada, B.C., México.
Smale, M.J. 1996. Cephalopod as prey. IV. Fishes. pp 1067-1082. En:
M. R. Clarke (ed.). The Role of Cephalopods in the world´s
oceans. Philosophical Transactions of the Royal Society of
London B Biological Sciences Science.
Smith, C. D. 2003. Diet of Octopus vulgaris in False Bay, South Africa.
Marine Biology. 143: 1127-1133.
Smith P.E. y M.P Zaret. 1982. Bias in estimating niche overlap. Ecology.
63: 1248-1253.
Saunders, W.B., Knight, R.L., Bond, P.N., 1991. Octopus predation on
Nautilus: Evidence from Papua New Guinea. Bulletin of Marine
Science 49 (1-2), 280–287.
Solís Ramírez, M. 1967. Aspectos biológicos del Octopus maya. En: Voss
y Solís (eds.) Publicaciones del Instituto Nacional de
Investigaciones Biológicas Pesqueras. México. 18:1-90.
Solís Ramírez M., J. I. Fernández M. y F. Márquez F. 2000. Pulpo.
Océano Pacífico. En: Cisneros-Mata M., L. Meléndez-Moreno, E.
Zárate-Becerra, T. Gaspar-Dillanes, L. López-González, C.
Saucedo-Ruiz and J. Tovar-Ávila (eds.). Sustentabilidad y pesca
responsable en México. Evaluación y Manejo, 1999-2000.
Instituto Nacional de la Pesca. México.
38

Tucker, J.K., Mapes, R.H., 1978. Possible predation on Nautilus
pompilius. Veliger 21, 95–98.
Valdez, O. V. M, E. Torreblanca R. y L. Avendaño C. 2006. Estrategias
de manejo para la pesquerías del pulpo (Octopus spp.) en Bahía
de los Ángeles, Baja California. Informe Técnico para: Área de
Protección de Flora y Fauna Islas del Golfo de California,
Comisión Nacional de Áreas Naturales Protegidas y Asociación de
Buzos de Bahía de los Ángeles, A.C. 21 pp.
Vázquez-Solórzano E., M.L. Salgado-Rogel, F. Uribe O. y J. Molina M.
2006. Propuesta para mejorar la pesquería de pulpo en Bahía de
los Ángeles, B.C. I.N.P. Centro Regional de Investigaciones
Pesqueras. En: Esteban Torreblanca R. (ed.). Memorias del Taller:
Bases para la conservación y el manejo de la pesquería del pulpo
(Octopus spp.) en Bahía de los Ángeles, Baja California. Agosto de
2006, Ensenada. Baja California, México. PRONATURA Noroeste.
Ensenada, B.C., México.
Villanueva, R. 1993. Diet and mandibular growth of Octopus magnificus
(Cephalopoda). South Africa. Journal of Marine Science, 13: 121-
126.
Villanueva, R., N.Koueta, J. Riba and B. Camou. 2000. Growth and
proteolytic activity of Octopus vulgaris paralarvae with different
food rations during first-feeding, using Artemia nauplii and
compound diets. Aquaculture. Netherlands. 205: 269-286.
http://www.sagarpa.gob.mx/conapesca/planeacion/anuario/anuario2
003.pdf
http://www.fao.org/figis/servlet/SQServlet?file=/usr/local/tomcat/FI/
5.5.9/fi5/webapps/figis/temp/hqp_75682.xmlandouttype=html
andxp_check=1:32001:1425,1:32001:1426,1:32001:1427,1:320
01:1428,1:32001:1429,1:32001:1430,1:32001:1431,1:32001:14
32.
39

40