inhibition of efflux pumps as a novel approach to combat ... pump inhibition 225 inhibition of...
TRANSCRIPT

Efflux Pump Inhibition 225
Inhibition of Efflux Pumps as a Novel Approachto Combat Drug Resistance in Bacteria
*For correspondence. Email [email protected]; Tel. (650) 428-3548;Fax. (650) 428-3550.
J. Mol. Microbiol. Biotechnol. (2001) 3(2): 225-236.
© 2001 Horizon Scientific Press
JMMB Symposium
Olga Lomovskaya* and Will Watkins
Microcide Pharmaceuticals, Inc., 850 Maude Avenue,Mountain View, CA 94043, USA
Abstract
Efflux mechanisms have become broadly recognizedas major components of resistance to many classesof antibiotics. Some efflux pumps selectively extrudespecific antibiotics, while others, referred to asmultidrug resistance (MDR) pumps, expel a variety ofstructurally diverse compounds with differingantibacterial modes of action. There are numerouspotentially beneficial consequences of the inhibitionof efflux pumps in improving the clinical performanceof various antibiotics, and several companies andresearch laboratories have initiated programs todiscover and develop efflux pump inhibitors. Thisreview will summarize recent achievements in this new,very exciting and equally challenging field.
Introduction
Efflux mechanisms have become broadly recognized asmajor components of resistance to many classes ofantibiotics. Some efflux pumps selectively extrude specificantibiotics, while others, referred to as multidrug resistance(MDR) pumps, expel a variety of structurally diversecompounds with differing antibacterial modes of action(Lawrence and Barrett, 1998; Levy, 1992; Lewis, 1994;Nikaido, 1998a; Nikaido, 1998b; Van Bembeke et al., 2000).Antibiotic-specific efflux pumps are usually encoded ontransmissible plasmids and transposons, while genesencoding many MDR pumps are normal constituents ofbacterial chromosomes. Thus, bacteria have the potentialto develop multi-drug resistance without acquisition ofmultiple specific resistance determinants.
Genes encoding some MDR pumps are expressedconstitutively in wild-type cells. As a result these cells havebasal levels of efflux activity, contributing to decreasedantibiotic susceptibility. This intrinsic resistance may be lowenough for the bacteria still to be susceptible to therapy.However, they would be even more susceptible if effluxpumps were rendered non-functional, allowing lower dosesof antibiotics to be used in therapy. This could be especiallyimportant for antibiotics with narrow therapeutic indices.
If it were not for efflux pumps, the spectrum of activityof many so-called “gram-positive” antibiotics could beexpanded to previously non-susceptible gram-negative
species. This has been demonstrated for β-lactams,macrolides, lincosamides, streptogramines, rifamycins,fusidic acid and oxazolidinones, all of which have potentantibacterial effects against engineered mutants ofEscherichia coli and Pseudomonas aeruginosa lackingefflux pumps (Nikaido, 1998a; Nikaido, 1998b; Poole,2000).
These facts taken together have led to the recognitionof numerous potential beneficial consequences of theinhibition of efflux pumps in improving the clinicalperformance of various antibiotics, and prompted severalcompanies and research laboratories to initiate programsto discover and develop efflux pump inhibitors. This reviewwill summarize recent achievements in this new, veryexciting and equally challenging field.
Identification of Target Antibiotics and Bacteria
While inhibition of efflux pumps appears to be an attractiveapproach for improving the clinical efficacy of antibioticsthat are substrates of such pumps, it is important to identifyantibiotics and target bacteria for which this approach wouldbe the most productive. Several factors should beconsidered, such as 1) the prevalence of efflux-mediatedresistance; 2) the multiplicity of efflux pumps; 3) othermechanisms besides efflux that contribute to resistance toa particular antibiotic; and 4) interactions between differentmechanisms of resistance.
MacrolidesMacrolides account for 10-15% of the worldwide oralantibiotic market (Kirst, 1991). Inhibition of the transposon-encoded macrolide-specific efflux pumps MefA/E (Clancyet al., 1996; Luna et al., 1999; Tait-Kamradt et al., 1997) ingram-positive bacteria would enhance the efficacy ofmacrolides, but only against a sub-population of resistantorganisms (Arpin et al., 1999; Giovanetti et al., 1999;Nishijima et al., 1999; Poutanen et al., 1999; Roberts etal., 1999). Another rather large sub-population withresistance due to target modification (dimethylation of 23Sribosomal RNA within the macrolide binding site by theproducts of erm genes) would remain unaffected. On theother hand, in Haemophilus influenzae (a primary causativeagent of respiratory infections), inhibition of a single MDRpump (HIAcrAB, encoded by a constitutively expressedhousekeeping operon (Sanchez et al., 1997) andapparently the major factor affecting susceptibility tomacrolides in this organism) should make the entirepopulation of bacteria more susceptible to macrolides suchas azithromycin or clarithromycin. The consequentimprovement in potency would significantly enhance theutility of the antibacterial class for the treatment of thiscommon clinical syndrome.

• MALDI-TOF Mass Spectrometry in Microbiology
Edited by: M Kostrzewa, S Schubert (2016) www.caister.com/malditof
• Aspergillus and Penicillium in the Post-genomic Era
Edited by: RP Vries, IB Gelber, MR Andersen (2016) www.caister.com/aspergillus2
• The Bacteriocins: Current Knowledge and Future Prospects
Edited by: RL Dorit, SM Roy, MA Riley (2016) www.caister.com/bacteriocins
• Omics in Plant Disease Resistance
Edited by: V Bhadauria (2016) www.caister.com/opdr
• Acidophiles: Life in Extremely Acidic Environments
Edited by: R Quatrini, DB Johnson (2016) www.caister.com/acidophiles
• Climate Change and Microbial Ecology: Current Research and Future Trends
Edited by: J Marxsen (2016) www.caister.com/climate
• Biofilms in Bioremediation: Current Research and Emerging Technologies
Edited by: G Lear (2016) www.caister.com/biorem
• Microalgae: Current Research and Applications
Edited by: MN Tsaloglou (2016) www.caister.com/microalgae
• Gas Plasma Sterilization in Microbiology: Theory, Applications, Pitfalls and New Perspectives
Edited by: H Shintani, A Sakudo (2016) www.caister.com/gasplasma
• Virus Evolution: Current Research and Future Directions
Edited by: SC Weaver, M Denison, M Roossinck, et al. (2016) www.caister.com/virusevol
• Arboviruses: Molecular Biology, Evolution and Control
Edited by: N Vasilakis, DJ Gubler (2016) www.caister.com/arbo
• Shigella: Molecular and Cellular Biology
Edited by: WD Picking, WL Picking (2016) www.caister.com/shigella
• Aquatic Biofilms: Ecology, Water Quality and Wastewater Treatment
Edited by: AM Romaní, H Guasch, MD Balaguer (2016) www.caister.com/aquaticbiofilms
• Alphaviruses: Current Biology
Edited by: S Mahalingam, L Herrero, B Herring (2016) www.caister.com/alpha
• Thermophilic Microorganisms
Edited by: F Li (2015) www.caister.com/thermophile
• Flow Cytometry in Microbiology: Technology and Applications
Edited by: MG Wilkinson (2015) www.caister.com/flow
• Probiotics and Prebiotics: Current Research and Future Trends
Edited by: K Venema, AP Carmo (2015) www.caister.com/probiotics
• Epigenetics: Current Research and Emerging Trends
Edited by: BP Chadwick (2015) www.caister.com/epigenetics2015
• Corynebacterium glutamicum: From Systems Biology to Biotechnological Applications
Edited by: A Burkovski (2015) www.caister.com/cory2
• Advanced Vaccine Research Methods for the Decade of Vaccines
Edited by: F Bagnoli, R Rappuoli (2015) www.caister.com/vaccines
• Antifungals: From Genomics to Resistance and the Development of Novel Agents
Edited by: AT Coste, P Vandeputte (2015) www.caister.com/antifungals
• Bacteria-Plant Interactions: Advanced Research and Future Trends
Edited by: J Murillo, BA Vinatzer, RW Jackson, et al. (2015) www.caister.com/bacteria-plant
• Aeromonas
Edited by: J Graf (2015) www.caister.com/aeromonas
• Antibiotics: Current Innovations and Future Trends
Edited by: S Sánchez, AL Demain (2015) www.caister.com/antibiotics
• Leishmania: Current Biology and Control
Edited by: S Adak, R Datta (2015) www.caister.com/leish2
• Acanthamoeba: Biology and Pathogenesis (2nd edition)
Author: NA Khan (2015) www.caister.com/acanthamoeba2
• Microarrays: Current Technology, Innovations and Applications
Edited by: Z He (2014) www.caister.com/microarrays2
• Metagenomics of the Microbial Nitrogen Cycle: Theory, Methods and Applications
Edited by: D Marco (2014) www.caister.com/n2
Caister Academic Press is a leading academic publisher of advanced texts in microbiology, molecular biology and medical research. Full details of all our publications at caister.com
Further Reading
Order from caister.com/order

226 Lomovskaya and Watkins
TetracyclinesTetracyclines have been used extensively against gram-positive and gram-negative bacteria, mycoplasma,rickettsiae and protozoan parasites (Chopra et al., 1992).Besides their therapeutic use in man and animals, theyhave also been used indiscriminately for growth promotionin animal husbandry. As a result, tetracycline resistancehas become widespread in both gram-positive and gram-negative species. The two major mechanisms of resistanceare active efflux by tetracycline-specific pumps (Tet-pumps)and ribosomal protection (Schnappinger and Hillen, 1996).Tet-pumps are widely spread among both gram-negativeand gram-positive bacteria (Roberts, 1994) and belong totwo subfamilies of the MF superfamily (see below),classified as 12- and 14-TMS export proteins, respectively.Within each sub-family, there is ca. 80% homology in aminoacids. However, the Tet-proteins of gram-negative andgram-positive bacteria are less similar to each other thanto other exporters from the corresponding families(Sheridan and Chopra, 1991). Inhibitors of Tet-transportersshould clearly improve activity of tetracyclines against Tet-containing bacteria, but the discovery of a single agentwith activity against both gram-negative and gram-positivepumps is a daunting prospect.
β -LactamsThis class includes expanded-spectrum penicillins,cephalosporins, monobactams and carbapenems, andbenefits from bactericidal activity combined with low toxicity.The main resistance mechanism in the majority of bacterialspecies is degradation of the antibiotic by variouschromosomal- or plasmid-encoded β-lactamases (Bush etal., 1995). In addition, resistance to carbapenems(imipenem, meropenem) can be due to mutational loss ofthe porin OprD, which is their main route of uptake (Triasand Nikaido, 1990). In P. aeruginosa, resistance to mostβ-lactams (with exception of the carbapenem imipenem)is also conferred by the overproduction of the MexAB-OprMRND MDR efflux pump (Li et al., 1994; Morshed et al.,1995; Yoneyama et al., 1997). Moreover, several surveyshave found a high proportion of MexAB-OprM-overexpressing strains among β-lactam-resistant clinicalisolates of P. aeruginosa (Bonfiglio et al., 1998; Chen etal., 1995; Ziha-Zarifi et al., 1999). It is noteworthy that allthese various resistance mechanisms can coexist in asingle bacterial cell and that their interactions have distincteffects on drug resistance. In the case of the carbapenemmeropenem, increased MexAB-OprM-mediated efflux anddecreased permeability due to the loss of OprD has amultiplicative effect on drug resistance (i.e. the fold increasein MIC due to their combined presence is close to theproduct of the individual fold increases). Consequently, thedeletion of MexAB-OprM decreases resistance tomeropenem even in strains lacking OprD (Kohler et al.,1999). The same would be expected for an inhibitor of theMexAB-OprM efflux pump.
A quite different picture emerges from a study of theinterplay between MexAB-OprM and chromosomal β-lactamase with respect to cephalosporins such asceftazidime and cefepime, and the monobactamaztreonam. Here, over-expression of MexAB-OprMsignificantly increased (16- to 64-fold) resistance towards
each of these antibiotics in strains lacking β-lactamase.Derepression of β-lactamase in a strain lacking MexAB-OprM conferred comparable or even higher increases.However, over-expression or deletion of efflux pumps inthe strain with de-repressed enzyme had only a weak effectadditive effect (Masuda et al., 1999; Nakae et al., 1999).An important conclusion from these experiments is thatinhibition of the MexAB-OprM pump in strains over-expressing β-lactamases is not expected to have asignificant effect on β-lactam resistance in P. aeruginosa.
AminoglycosidesThese are rapidly bactericidal agents that impair proteinsynthesis by binding to bacterial ribosomes. Developmentof new analogs has been driven both by the necessity toidentify compounds that resist structural modification bymultiple aminoglycoside modifying enzymes, and by theneed for better therapeutic indices. Two main mechanismsof aminoglycoside resistance have been identified: specificenzymatic inactivation by modifying enzymes, and a moregeneral mechanism often referred to as impermeabilityresistance. The prevalence of these resistancemechanisms is different for various bacterial species. Forexample, in Enterobacteriaceae and Staphylococcus, atendency to acquire multiple aminoglycoside modifyingenzymes, resulting in multiple genotypes and complexphenotypes, has been noted. In contrast, in P. aeruginosa,the main changes were towards an increase in thefrequency of the impermeability type of resistance (Milleret al., 1997).
Until recently, aminoglycosides were among the fewexamples of antibiotic classes for which no active extrusiondue to multidrug resistance pumps (MDR) had beendemonstrated. This situation changed with the discoveryof the MDR pump AmrAB-OprA in Burkholderiapseudomalei (Moore et al., 1999). More recently, otheraminoglycoside efflux pumps have been discovered:MexXY-OprM in P. aeruginosa (Aires et al., 1999;Westbrock-Wadman et al., 1999) and AcrD in E. coli(Rosenberg et al., 2000). Importantly, the clinical relevanceof the MexXY-OprM-mediated efflux of aminoglycosideshas been established: elevated expression of mexXYgenes has been detected in clinical isolates of P. aeruginosaexhibiting impermeability-type resistance (Westbrock-Wadman et al., 1999). Although it appears thatimpermeability-type resistance is a multifactorialphenomenon, the MexXY-OprM efflux pump may prove tobe its essential component, suggesting the applicability ofthe efflux pump inhibitor approach. Another potential benefitrelates to the phenomenon of induced adaptive resistanceto the bactericidal effect of aminoglycosides after initialexposure to these antibiotics (Barclay et al., 1996; Daikoset al., 1990; Daikos et al., 1991; Karlovsky et al., 1996;Xiong et al., 1997): mutants lacking MexXY-OprM showedalmost no adaptation, unlike the MexXY-OprM-containingstrain (Mao et al., 2000). This is consistent with the recentfinding that expression of the mexXY operon is induced byaminoglycosides (Masuda et al., 2000). Yet anotherpotential benefit of an inhibitor of MexXY-OprM relates tothe well-documented phenomenon of antagonism ofaminoglycosides by divalent cations (Medeiros et al., 1971;Zimelis and Jackson, 1973). It is generally understood that

Efflux Pump Inhibition 227
of this pump renders strains significantly (8- to 16-fold) moresusceptible to fluoroquinolones as compared to the wild-type. While no other Mex pumps have been implicated influoroquinolone resistance in the clinical setting, recent dataindicate that more of them may be involved in intrinsicresistance (Masuda et al., 2000; Srikumar et al., 2000).
Known MDR pumps from gram-positive bacteriaimplicated in resistance to fluoroquinolones belong to theMF family (Saier et al., 1999a) (see below). Examplesinclude NorA from Staphylococcus aureus (Neyfakh et al.,1993; Ng et al., 1994; Yoshida et al., 1990b) and PmrAfrom Streptococcus pneumoniae (Gill et al., 1999). WhileNorA, like MexAB-OprM, is produced by wild-type cells(contributing ca. 4-fold to resistance to ciprofloxacin, Hsiehet al., 1998; Yamada et al., 1997), expression of the genepmrA appears to occur only after the acquisition ofregulatory mutations (Gill et al., 1999). The possibility ofthe presence of additional efflux pumps in these organismshas recently been demonstrated (Brenwald et al., 2000;Piddock et al., 1997; Piddock et al., 1998). Active efflux offluoroquinolones has also been seen in wild-type strainsof Enterococcus faecalis and Enterococcus faecium (Lynchet al., 1997). Among gram-positive bacteria, theseorganisms are the most resistant to fluoroquinolones. Therelevant efflux pumps still await identification.
In P. aeruginosa single-step gyrase and efflux mutantsconferred similar 4- to 8-fold increases in fluoroquinoloneresistance (Lomovskaya et al., 1999a). Of note was theobservation that the frequencies of mutations resulting inpump over-expression were ca. 10-fold higher thanfrequencies of target-based mutations (Kohler et al., 1997b;Lomovskaya et al., 1999a). Again, both types of resistancemechanisms may coexist in a single bacterial cell, and suchstrains have been identified among highly fluoroquinolone-resistant clinical isolates of many gram-negative and gram-positive bacteria (Cho et al., 1999; Everett et al., 1996;Guerin et al., 2000; Jalal and Wretlind, 1998). In fact, recentstudies indicated that high-level fluoroquinolone resistanceoccurs due to the interplay of the two mechanisms.
This interplay has been studied in detail in P.aeruginosa (Lomovskaya et al., 1999a), E. coli (Oethingeret al., 2000) and S. aureus (Kaatz and Seo, 1997) usinglaboratory mutants over-expressing or lacking efflux pumpsand containing various combinations of target-based andefflux-mediated mutations. When both mechanisms werecombined in a single cell, their relative contributions toresistance were independent of each other; the combinedresistance was much higher than that conferred by eachof the mechanisms when present singly. In fact, the foldincrease in drug resistance was close to the product of thefold increases due to the individual mechanisms. In P.aeruginosa (Lomovskaya et al., 1999a), abolishment of thebasal level of mexAB-oprM expression by deletion of theoperon resulted in ca. 8-fold increased susceptibility tolevofloxacin in the wild-type strain (whose MIC was 0.125µg/ml). The same 8-fold increase in susceptibility uponpump deletion was seen irrespective of the number of targetmutations. For this particular antibiotic, where a singletarget-based mutation in the presence of an efflux pumpresults in clinically relevant resistance (MIC 1-2 µg/ml), theabsence of functioning efflux pump(s) will therefore resultin an 8-fold reduction in the MIC, rendering the organism
divalent cations interfere with uptake of aminoglycosides,mainly at the level the outer membrane (Abdel-Sayed etal., 1982; Hancock et al., 1981). Interaction ofaminoglycosides with negatively chargedlipopolisaccharides (LPS) is believed to result inpermeabilization of the outer membrane, which facilitatesuptake of aminoglycosides into the periplasmic space(Hancock et al., 1981). Divalent cations apparently competewith aminoglycosides for binding to LPS, and consequentlyprevent the so-called self-promoted uptake of antibiotics.We found, unexpectedly, that while both Mg2+ and Ca2+
antagonized aminoglycosides, the antagonism was onlyseen in strains of P. aeruginosa that contained a functioningMexXY-OprM efflux pump (Mao et al., 2000). This indicatesthat decreased uptake contributes to increased resistanceto aminoglycosides only in the presence of active efflux.Thus inhibition of the MexXY-OprM efflux pump maysignificantly increase the (notoriously narrow) therapeuticindex of aminoglycosides and improve the clinical utility ofthis important class of antibiotics.
FluoroquinolonesFluoroquinolones are oral, broad-spectrum bactericidalagents widely used in both community and hospital settings.Older compounds such as norfloxacin, ciprofloxacin andenofloxacin perform better against gram-negative thangram-positive pathogens such as S. pneumoniae or S.aureus (Gootz and Brighty, 1996). Newer fluoroquinolonessuch as levofloxacin, sparfloxacin and trovafloxacin haveimproved activity against gram-positive organisms (Brightyand Gootz, 1997). The antibacterial activity offluoroquinolones is based on inhibition of type IItopoisomerases, DNA gyrase (encoded by the gyrA andgyrB genes) and DNA topoisomerase IV (encoded by theparC and parE genes). Only two mechanisms, targetmodification and efflux by MDR pumps, contribute toresistance to this important class of antibiotics. Most ofthe target-based mutations in both gram-negative andgram-positive bacteria have been mapped to the so-calledquinolone resistance-determining regions (Piddock, 1999;Yoshida et al., 1990a) of the gyrase or topoisomerase IVgenes. MDR efflux pumps from clinically relevant gram-negative and gram-positive bacteria belong to twoevolutionally distinct families of transporters. In gram-negative bacteria, fluoroquinolones are extruded out of cellsby tripartite MDR pumps of the RND family (Tseng et al.,1999) (see below), such as AcrAB-TolC in E. coli andseveral Mex pumps in P. aeruginosa (Nikaido, 1998a;Poole, 2000). Similar RND pumps involved in efflux offluoroquinolones have been also discovered in other gram-negative bacteria (Poole, 2000). Notably, a single bacterialstrain may contain several fluoroquinolone pumps. Themost well studied case is that of P. aeruginosa, whichharbors at least three such pumps, MexAB-OprM (Pooleet al., 1993), MexCD-OprJ (Poole et al., 1996a) andMexEF-OprN (Kohler et al., 1997a) that contribute to fluoro-quinolone resistance in clinical settings (Cho et al., 1999;Fukuda et al., 1990; Fukuda et al., 1995; Hirai et al., 1987;Jakics et al., 1992; Jalal and Wretlind, 1998; Ziha-Zarifi etal., 1999). Of these, the MexAB-OprM is the only one thatis produced constitutively (Poole et al., 1996b) andconsequently contributes to intrinsic resistance. Deletion

228 Lomovskaya and Watkins
clinically susceptible. Deletion of mexAB-oprM from strainsoverproducing the corresponding pump resulted in a 64-fold increase in susceptibility, also regardless of thepresence of target mutations, indicating that the foldreduction in MIC due to inhibition of the pump(s) isdependent only on the levels of pump expression.
A major beneficial consequence of the inhibition ofefflux pumps apparent from these results is that thefrequency of emergence of strains with clinically relevantlevels of resistance to fluoroquinolones will be significantlydecreased. Obviously, inhibition of efflux pumps shouldprevent the appearance of efflux-mediated mutants, but areduced frequency of emergence of strains with otherresistance phenotypes is also to be expected, since in orderfor bacteria without efflux pumps to attain clinically relevantresistance, they need to acquire more than one target-based mutation. Such simultaneous acquisition of multiplemutations is a rare event. Indeed, a dramatic decrease inthe frequency of emergence of resistant strains has beenobserved in P. aeruginosa using mutants lacking effluxpumps (Lomovskaya et al., 1999a). The sameconsiderations can also be applied to gram-positivebacteria and can explain results obtained earlier with S.aureus and S. pneumonia: addition of reserpine (an inhibitorof MDR pumps) to the selecting media suppressedemergence of mutants with resistance to fluoroquinolones(Markham and Neyfakh, 1996; Markham, 1999).
In conclusion, it is expected that inhibition of MDRpumps should decrease intrinsic resistance tofluoroquinolones, should significantly reverse both target-based and efflux-mediated acquired resistance, and(perhaps most important) should reduce the frequency ofemergence of highly resistant mutants. Undoubtedly,fluoroquinolones are one of the antibiotic classes for whichthe efflux pump inhibitor approach appears to be mostcompelling.
Interplay Between Different Efflux PumpsAs discussed above, multiple mechanisms of resistanceto a particular antibiotic may coexist in a single bacterialcell. A single cell may also contain more than one effluxpump capable of effluxing the same antibiotic. Cells of E.coli or P. aeruginosa may acquire plasmid-encodedtransporters such as TetA or CmlA, despite the presencein their genomes of endogenous MDR pumps (AcrAB-TolCand MexAB-OprM, respectively) that can also effluxtetracycline and chloramphenicol. P. aeruginosa containsmultiple MDR pumps that can confer resistance tofluoroquinolones.
Structurally, efflux pumps occur either as single-component or multi-component systems. In gram-negativebacteria, single-component efflux pumps, such as TetA orCmlA, extrude their substrates into the periplasmic space(Thanassi et al., 1997) and maintain a concentrationgradient of antibiotics across the inner membrane. Multi-component efflux pumps (which are found exclusively ingram-negative bacteria), by contrast, traverse both innerand outer membranes; each contains a transporter locatedin the cytoplasmic membrane (as exemplified by AcrB orMexB), an outer membrane channel (TolC or OprM) and aperiplasmic ‘linker’ protein (AcrA or MexA), which is thoughtto bring the other two components into contact (Zgurskaya
and Nikaido, 1999a; Zgurskaya and Nikaido, 1999b). Thisstructural organization allows extrusion of substratesdirectly into the external medium (Zgurskaya and Nikaido,2000). The relative impermeability of the outer membraneof gram-negative bacteria to both hydrophobic andhydrophilic antibiotics (Nikaido, 1994; Nikaido, 1998c;Nikaido and Vaara, 1985) is thereby exploited as a meansof lowering the intracellular accumulation of antibioticsthrough the function of these pumps. Consequently, directefflux into the medium leads to a decreased amount ofdrug in the periplasmic space; a concentration gradient ismaintained across the outer membrane.
We sought to investigate the effects of simultaneousexpression of two different efflux pumps upon susceptibilityto antibiotics that are the substrates of both pumps (Lee etal., 2000a). Several types of pump combinations have beenstudied: 1) simultaneous expression of a single-componentefflux pump (exporting antibiotics into the periplasm) incombination with a multi-component efflux pump(accomplishing efflux directly into the external medium);2) simultaneous expression of two single-componentpumps; and 3) simultaneous expression of two multi-component pumps. When efflux pumps of differentstructural types were combined in the same cell, theobserved antibiotic resistance was much higher than thatconferred by either of the pumps expressed singly, andthe fold increase in drug resistance was close to the productof the fold increases due to the individual pumps. Forexample, when singly expressed, both AcrAB-TolC andCmlA conferred similar levels of resistance tochloramphenicol (MICs ranged from 16-32 µg/ml) in P.aeruginosa; since the MIC for the pumpless strain was 1µg/ml, each pump afforded a 16- to 32-fold increase inresistance. The strain expressing both pumps had an MICof >256 µg/ml. By contrast, simultaneous expression ofeither two single-component or two multi-component effluxpumps did not produce large increases in antibioticresistance; the effects were only additive. For instance,MexAB-OprM and MexCD-OprJ when individually over-expressed conferred a comparable level of resistance tolevofloxacin (1 µg/ml), which corresponded to a 64-foldincrease in resistance when compared to the pumplessstrain. The strain over-expressing both pumps had an MICof 2 µg/ml (128-fold increase).
These effects are explicable according to the notionthat pumps of the same structural type are organized inparallel, while pumps of different structural types areorganized in series. Parallel pumps extrude antibiotics intothe same compartment (either the periplasm or the externalmedium). For pumps in series, the single component pumptransfers antibiotics from the cytoplasm to the periplasm,and the multi-component pump extrudes them into externalmedium (Figure 1). We have developed a very simplemathematical model explaining our results (Lee et al.,2000a).
The arrangement of pumps both in series and inparallel appears to be relevant in clinical settings. In somestrains of E. coli that are highly resistant (MIC >128 µg/ml)to chloramphenicol, the resistance observed is theconsequence of simultaneous over-expression of both themulti-component AcrAB-TolC pump and the single-component “phenicol-specific” ef flux pump, Flo (A. Lee

Efflux Pump Inhibition 229
and O. Lomovskaya, unpublished data). The latter was firstdiscovered in Pasteurella piscicida (Kim and Aoki, 1996)but recently has also been detected in clinical strains ofSalmonella typhimurium (Arcangioli et al., 1999; Bolton etal., 1999) and E. coli (Keyes et al., 2000). In this case,although inhibition of either pump is expected to decreasedrug resistance to some extent, complete reversal ofresistance will require simultaneous inhibition of both.Strains over-expressing pairs of Mex pumps have beenreported among clinical isolates of P. aeruginosa (Cho etal., 1999), and as predicted, the MICs of these strains wereno higher than those overexpressing single Mex pumps.In this case, inhibition of both pumps was required toachieve any antibiotic potentiating effect at all.
Inhibitors of Efflux Pumps as Novel Agents forCombination Therapy
Two approaches to combat efflux-mediated resistance thatare currently being advanced are: 1) modification of existingantibiotics to identify derivatives minimally affected by effluxpumps; and 2) discovery and development of therapeutic
agents which inhibit the transport activity of efflux pumpsand could be used in combination with existing antibiotics.Several successful examples of the first approach include:1) a new class of semisynthetic tetracyclines, theglycylcyclines (Sum et al., 1998), that overcome efflux-mediated resistance because they are not recognized bythe transporter protein (Someya et al., 1995); 2) newmacrolides and ketolides that appear to be much lessaffected by the Mef-pumps (Biedenbach et al., 1998;Brennan et al., 1998; Jamjian et al., 1997; Davies et al.,2000); and 3) new fluoroquinolones which overcome NorA-or PmrA-mediated efflux in gram-positive bacteria (Fukudaet al., 1998; Gill et al., 1999; Gootz et al., 1999; Pestova etal., 1999; Piddock et al., 1998). In the latter case, directtransport experiments are still needed to clarify whetherthis is a consequence of lack of binding to the pumps, oran increased rate of diffusion into the cytoplasm.
It is perhaps significant, however, that these examplesall relate to pumps that span a single membrane. It appearsmuch more difficult to outsmart MDR pumps from gram-negative bacteria by this approach. Part of the problem isthat in these bacteria particular restrictions are imposedon the structure of successful drugs, in that they must beamphiphilic to be able to cross both membranes.Apparently, it is this very amphiphilicity that makesantibiotics good substrates of MDR pumps in theseorganisms.
The second approach based on inhibition of effluxpumps, while very compelling scientifically and of clearpotential clinical benefit, brings with it many challenges.Besides the obvious issues common to all infectiousdisease drug development programs (appropriate potencyand spectrum of activity, bioavailability, clearance,avoidance of mechanism-based and/or non-mechanism-based toxicity etc.), several are specific to this strategy. Inthe discovery phase, the pharmacokinetics of the partnerantibiotic must be taken into account; viablepharmacokinetic profiles of the potentiator may besignificantly restricted as a result. In development, clinicaltrials must be designed so as to demonstrate improvementof a combination product over existing therapies. Despitethese inherent challenges, several programs of discoveryand development of efflux pump inhibitors have beeninitiated and their achievements are reviewed below.
Inhibitors of Tetracycline TransportersSince tetracycline is effluxed by specific transporters it isreasonable to expect that inhibitors might be found amongtetracycline analogs; for example, inhibition might beachieved by higher binding affinity and a lower dissociationconstant. Indeed, certain semisynthetic tetracyclineanalogs inhibited tetracycline efflux in everted vesiclesprepared from E. coli cells containing the TetB protein. Themost potent analog, 13-CPTC, interfered with tetracyclinetransport by competitive binding to TetB, but wastransported less efficiently than tetracycline. Whencombined with doxycycline, 13-CPTC exhibited synergyagainst E. coli strains expressing either TetA or TetBproteins by lowering the MIC by a factor of 2 or more(Nelson and Levy, 1999; Nelson et al., 1993; Nelson et al.,1994).
A second class of Tet pump inhibitors, unrelated to
A
B
C
D
... .
. .
.
.
. . .. ..
. . .. ....
.. . .. ...... . .. ....
.. . .. .... ...
. .. ... .
....
. . .. . .. ... .. . .. .
...
..
Medium Periplasm Cytoplasm
... ..
. . .. .. . .. ... .
OM IM
R1
R1
R2
R2
Figure 1. The models show the fluxes of antibiotics at steady state. Externalconcentrations of antibiotics are equal to MICs and the density of dotscorresponds approximately to the concentrations of antibiotics in eachcompartment. In each case, the concentration of antibiotics in the cytoplasmis the same and sufficient to inhibit the target for antibiotics. Arrows indicatedirections and thickness indicates rate of flux. The thick line separating theperiplasm and the external medium indicates the low permeability of theouter membrane as compared to the inner membrane. In cells withoutefflux pumps (A) all compartments are essentially in equilibrium. In cellsexpressing a single-component pump (B), which extrudes substrates intothe periplasm thus balancing the rapid influx across the inner membrane,the external medium and the periplasm are in equilibrium, and aconcentration gradient (R1) exists at the inner membrane. In cells expressingmulti-component efflux pump (C), which extrudes antibiotics in the externalmedium bypassing the outer membrane barrier, the cytoplasm and theperiplasm are in equilibrium and the concentration gradient (R2) is seenacross the outer membrane. In the case of simultaneous expression ofboth efflux pumps (D), concentration gradients are maintained at both theinner and outer membranes. This results in a multiplicative, or synergistic,effect on drug resistance.

230 Lomovskaya and Watkins
the tetracyclines, has also been reported (Silvia et al.,1999). By high throughput screening using TetC as a target,UK-57,562 was discovered. This agent lacks conventionalantibacterial activity (MIC >50 µg/ml), but reduced the MICfor tetracycline 8- to 32-fold against resistant field isolatesof P. haemolytica and P. multocida.
One other compound, Ro 07-3149, has been reportedto inhibit the efflux of tetracyclines from S. aureus 743(Hirata et al., 1998). This compound was shown to increasethe intracellular concentration of tritiated tetracycline atconcentrations at or above 25 µg/ml. Although intrinsicallyantibacterial (MIC 32 µg/ml), the compound was shown toreduce the MIC to tetracycline two-fold at 8 µg/ml, whereasthe MIC of chloramphenicol was unchanged.
Inhibitors of Multidrug Resistance PumpsSelectivity of Inhibitors of Bacterial MDR PumpsBased on sequence similarity, bacterial efflux pumps areclassified into five superfamilies known as 1) ATP BindingCassette transporters (ABC), 2) Major Facilitators (MF),3) Resistance-Nodulation-Division (RND), 4) Small Multi-drug Resistance (SMR), and 5) Multi-drug And Toxiccompound Extrusion (MATE). Representatives of all ofthese families are also present in mammalian cells (Paulsenet al., 1998; Saier et al., 1999a; Saier, 1998; Saier et al.,1999b; Saier et al., 1998; Tseng et al., 1999). Moreover,the precedent of a compound that can inhibit bothprokaryotic and eukaryotic MDR pumps is well established:reserpine, long since identified as an inhibitor of amammalian MDR ABC-transporter P-glycoprotein (Becket al., 1988), also inhibits many bacterial pumps such asthe MF-transporters NorA (Neyfakh et al., 1993) and PmrA(Gill et al., 1999) from gram-positive bacteria, and MdfA(Edgar and Bibi, 1999) from E. coli and the E. coli SMRtransporter EmrE (Yerushalmi et al., 1995). Interestingly,although reserpine does not inhibit RND pumps, it still hasaffinity for at least one: in a strain of E. coli over-expressingAcrAB-TolC it did not inhibit the MdfA transporter, whereasexcellent inhibition was observed in the ∆acrAB mutant(Angela Lee and Olga Lomovskaya, unpublished data).Thus, although none of these data show that selectivitycannot be achieved, they highlight the need to ascertainthe selectivity of bacterial inhibitors at a very early stage ina discovery program.
Spectrum of Activity of Inhibitors of Bacterial MDR PumpsWhile most (if not all) currently identified MDR pumps fromgram-positive bacteria that confer clinically relevant drugresistance belong to the MF family, the vast majority ofrelevant pumps in gram-negative bacteria are RNDtransporters. This implies a low probability for identificationof a broad-spectrum non-competitive inhibitor. However,many very different pumps still extrude the same substrates(for example, fluoroquinolones), implying some similarityin their substrate binding sites. Therefore, it may be possibleto identify a competitive inhibitor with activity against severalrelevant transporters.
Screening for Efflux Pump Inhibitors and Criteria for EffluxPump Inhibitory Mode of Action of the Screening HitsUnlike drug-specific pumps, MDR-transporters havemultiple, structurally dissimilar substrates, so the starting
point for design of a potential inhibitor is not obvious. Inthis case, random screening of synthetic compounds andnatural product libraries appears to be a more compellingapproach. However, this situation might change if naturalsubstrates of relevant MDR pumps with more specific andtighter binding were to be discovered.
While several screens for the identification of effluxpump inhibitors may be envisioned (Trias et al., November,1999), one of the simplest employs whole-cell growthinhibition assays in which strains expressing efflux pumpsare grown in the presence of sub-inhibitory concentrationsof antibiotics that are substrates of those pumps. Syntheticcompounds or natural product extracts that inhibit growthin the presence but not in the absence of the antibiotic willscore as hits. Various follow-up assays are then requiredto discriminate between false positives and true efflux pumpinhibitors. To qualify as efflux pump inhibitors (EPIs), hitcompounds have to satisfy several criteria:1) they must enhance the activity of multiple substrates
of the pump;2) they must not potentiate antibiotics that are not effluxed;3) they must not have activity in strains lacking efflux
pumps;4) they must increase accumulation and decrease
extrusion of efflux pump substrates;5) they must not affect the proton gradient across the inner
membrane.This approach has been used successfully to identifyinhibitors of the NorA pump from S. aureus and Mex pumpsfrom P. aeruginosa.
NorA Pump InhibitorsInhibitors of the NorA pump that potentiate ciprofloxacinagainst both wild-type and NorA-overexpressing strainsof S. aureus have been reported by authors at Influx Inc.(Markham et al., 1999). A library of 9,600 syntheticcompounds was screened for potentiation of ethidiumbromide in an engineered strain of B. subtilis lacking twomajor MDR transporters and containing the plasmid with anorA gene. As many as 4% of the compounds screenedwere active, most of which could be divided into severalunrelated chemical groups. While surprising at first sight,the large number of hits may reflect the very nature of thetarget protein, which has evolved so as to bind to manystructural types. Representatives of the most potentchemically diverse series are shown in Figure 2.
All of these compounds decreased MICs tociprofloxacin up to 4-fold (at concentrations ranging from0.2 µg/ml to 1.5 µg/ml) in strains of S. aureus over-expressing the NorA pump. A 4-fold decrease in MIC isclose to the maximum theoretical effect (8-fold), based ondata from deletion of norA (Hsieh et al., 1998; Yamada etal., 1997). Importantly, all compounds dramaticallysuppressed the frequency of emergence of ciprofloxacin-resistant strains when selection was performed at 1 µg/mlof ciprofloxacin using the wild-type strain of S. aureus. Inthis strain, the maximal effect of pump inhibition shouldresult in an MIC of 0.125 µg/ml; apparently single-steptarget mutations could not afford the requisite 8-foldincrease in resistance. No data on the efficacy of thesenovel NorA inhibitors in animal models are currentlyavailable.

Efflux Pump Inhibition 231
A very important fundamental question is whetherinhibitors of NorA will have actual clinical utility. Studies onrecent representative clinical isolates (including MRSAstrains) with high-level ciprofloxacin resistance arenecessary to evaluate whether a 4- to 8-fold extent ofpotentiation (regardless of the potency of the potentiator)would shift the MIC50 and MIC90 of the bacterial populationbelow the susceptibility barrier.
INF271 and INF55 also inhibited efflux-mediatedresistance in S. pneumoniae. Recent results from severallaboratories indicate that MDR pumps other than PmrAmay be involved in fluoroquinolone resistance in thisorganism. Furthermore, these pump(s) confer resistanceto newer compounds, such as levofloxacin andtrovafloxacin, that are only minimally affected by PmrA(Brenwald et al., 2000).
RND MDR InhibitorsA collaboration between Microcide Pharmaceuticals andDaiichi Pharmaceutical Co. resulted in the identification ofthe first inhibitor of multiple RND transporters in gram-negative bacteria, MC-207,110 (Renau et al., 1999;Lomovskaya et al., 2001) (Figure 3).
This compound was among the few hits identified in alevofloxacin potentiation assay after the screening of alibrary containing over 200,000 synthetic compounds andnatural product extracts using strains of P. aeruginosa over-expressing Mex pumps, which contribute to fluoroquinoloneresistance in this organism. The striking difference inamount of hits found in efflux screens against gram-positivevs. gram-negative bacteria may be explained by theeffective permeability barrier for compounds, includingpotential efflux pump inhibitors, provided by the outer
membrane in the latter case.The efflux pump inhibitory mode of action of MC-
207,110 was confirmed in a series of microbiological,accumulation and efflux assays (Lomovskaya et al., 1999b;Lomovskaya et al., 2001). While MC-207,110 potentiateda variety of antibiotics that are substrates of Mex pumps,the magnitude of the potentiating effect was stronglydependent on the nature of the particular substrate,perhaps indicating that different antibiotics may havedifferent binding sites on the pump and that inhibition byMC-207,110 is binding site-specific. The possibility ofmultiple substrate binding sites was recently demonstratedfor several MDR pumps (Mitchell et al., 1999; Putman etal., 1999). According to this interpretation, the pump, whenexposed to MC-207,110, can still extrude some (but notall) substrates. Furthermore, MC-207,110 can itself beeffluxed by RND pumps, providing direct evidence of aninteraction between them (Warren et al., 2000).
Remarkably, several antibiotics, all substrates ofMexAB-OprM, did not compete with MC-207,110 inaccumulation assays even when tested at very highconcentrations (up to 500 µg/ml). It is quite possible thatMC-207,110 appears as an inhibitor since it is a much bettersubstrate of efflux pumps as compared to the antibiotics.It is noteworthy that many compounds that enhance activityof anti-cancer agents against mammalian cellsoverexpressing the MDR-pump, P-glycoprotein, have alsobeen shown to be efficiently extruded by this pump (Stein,1997).
Numerous potential benefits of using MC-207,110 incombination with levofloxacin against P. aeruginosa havebeen evident. MC-207,110 decreased the intrinsicresistance to levofloxacin ca. 8-fold in a wild-type strain ofP. aeruginosa, while in strains over-expressing efflux pumpsthe susceptibility was increased up to 64-fold. As expected,MC-207,110 potentiated levofloxacin irrespective of thepresence of target based mutations (Lomovskaya et al.,2001).
Recent clinical isolates of P. aeruginosa with a widerange of resistant phenotypes also showed increasedsusceptibility to levofloxacin in the presence of MC-207,110.Remarkably, both MIC50 and MIC90 were decreased to thesame extent (16-fold) by MC-207,110, providing additionalevidence that the potentiating effect is not dependent on
Figure 2. NorA inhibitors.
Figure 3. MC-207,110.

232 Lomovskaya and Watkins
the absolute level of resistance, but solely on the level ofefflux pump expression. In the presence of the inhibitor,both MIC50 and MIC90 were below the susceptibility barrier(Figure 4).
A major beneficial consequence of the inhibition ofmultiple efflux pumps was a dramatic decrease in thefrequency of emergence of P. aeruginosa strains withclinically relevant levels of resistance to fluoroquinolones:resistant mutants with an MIC of 1 µg/ml for levofloxacinwere isolated with a frequency less than 10-11 (versus 10-7
in the absence of efflux pump inhibitors). These data areparticularly important in view of results indicating that under
stress conditions, such as those in acute clinical infections,there is an increased frequency of selection ofhypermutable strains of P. aeruginosa (Oliver et al., 2000).
MC-207,110 (and/or its analogs) also has activityagainst RND pumps in many representatives ofPseudomonadaceae, Enterobacteriaceae and H.influenzae (Blais et al., 1999). The potentiation ofmacrolides in gram-negative bacteria may prove to be animportant application of efflux pump inhibitors (Figure 5).
Thus, a single compound, when combined withdifferent antibiotics, may have multiple clinical applications.This inhibitor does not, however, show activity against MF
0
20
40
60
80
100
0.00
8
0.01
6
0.03
1
0.06
3
0.12
5
0.25 0.5 1 2 4 8
16 32 64
MIC (µg/ml)
Cum
ulat
ive %
Sus
cept
ibili
tyLVX LVX+EPI
MIC50
(µg/ml)MIC90
(µg/ml)
LVFX 0.5 8
LVFX +MC-207,110 0.03 0.5
Figure 4. Cumulativesusceptibility to levofloxacinwith or without MC207,110for 50 clinical isolates of P.aeruginosa.
Haemophillus influenzaen=34
0
2 0
4 0
6 0
8 0
1 0 0
0.00
8
0.01
6
0.03
1
0.06
3
0.12
5
0.25 0.5 1 2 4 8
16 32
Cu
mu
lati
ve %
Su
scep
tib
le
Escherichia colin=24
0
20
40
60
80
100
0.01
6
0.03
1
0.06
3
0.12
5
0.25 0.5 1 2 4 8
>8 16 32 64
128
>12
8
Cu
mu
lati
ve %
Su
scep
tib
le
Klebsiella pneumoniaen=20
0
20
40
60
80
100
0.01
6
0.03
1
0.06
3
0.12
5
0.25 0.
5 1 2 4 8
>8 16 32 64
128
>12
8
MIC (µg/ml)
Cu
mu
lati
ve %
Su
scep
tib
le
AZM AZM + MC-04,124 CLR CLR + MC-04,124
Acinetobacter baumanniin=11
0
20
40
60
80
100
0.03
1
0.06
3
0.12
5
0.25 0.
5 1 2 4 8
16 32 64
128
MIC (µg/ml)
Cu
mu
lati
ve %
Su
scep
tib
le
AZM AZM + MC-04,124 CLR CLR + MC-04,124
Figure 5. Cumulative susceptibility to azithromycin and clarythromycin alone or in combination with MC-04,124 (analog of MC-207,110) for gram-negativeclinical isolates.
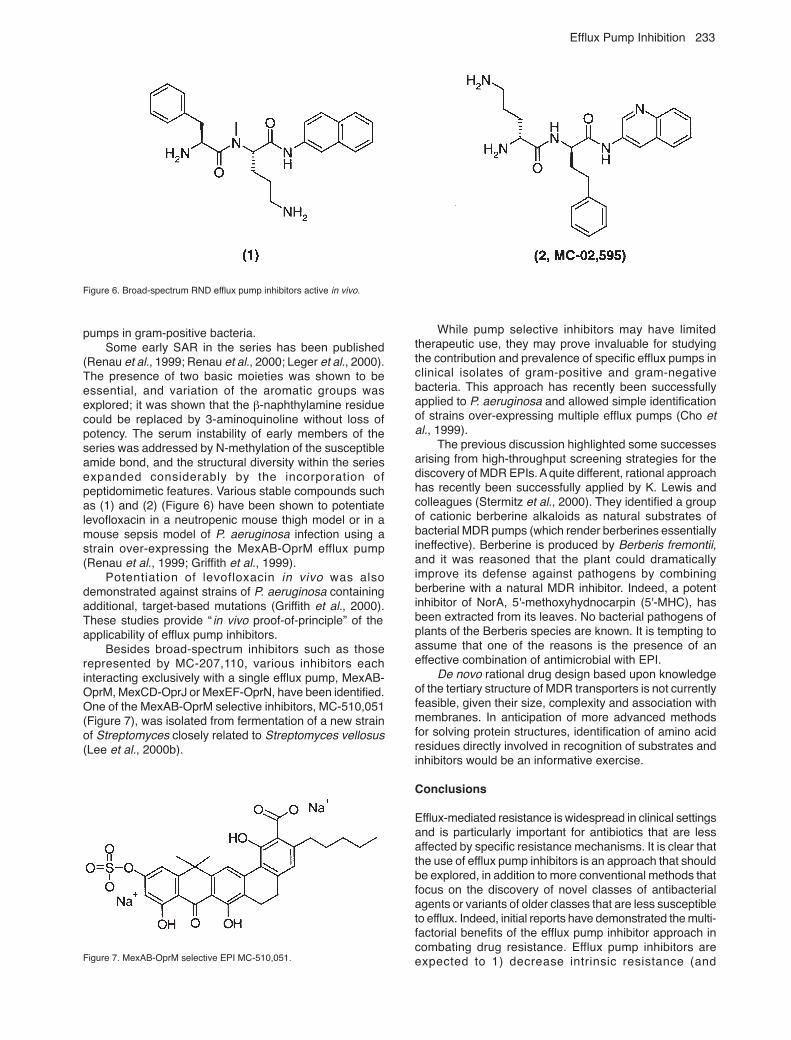
Efflux Pump Inhibition 233
pumps in gram-positive bacteria.Some early SAR in the series has been published
(Renau et al., 1999; Renau et al., 2000; Leger et al., 2000).The presence of two basic moieties was shown to beessential, and variation of the aromatic groups wasexplored; it was shown that the β-naphthylamine residuecould be replaced by 3-aminoquinoline without loss ofpotency. The serum instability of early members of theseries was addressed by N-methylation of the susceptibleamide bond, and the structural diversity within the seriesexpanded considerably by the incorporation ofpeptidomimetic features. Various stable compounds suchas (1) and (2) (Figure 6) have been shown to potentiatelevofloxacin in a neutropenic mouse thigh model or in amouse sepsis model of P. aeruginosa infection using astrain over-expressing the MexAB-OprM efflux pump(Renau et al., 1999; Griffith et al., 1999).
Potentiation of levofloxacin in vivo was alsodemonstrated against strains of P. aeruginosa containingadditional, target-based mutations (Griffith et al., 2000).These studies provide “ in vivo proof-of-principle” of theapplicability of efflux pump inhibitors.
Besides broad-spectrum inhibitors such as thoserepresented by MC-207,110, various inhibitors eachinteracting exclusively with a single efflux pump, MexAB-OprM, MexCD-OprJ or MexEF-OprN, have been identified.One of the MexAB-OprM selective inhibitors, MC-510,051(Figure 7), was isolated from fermentation of a new strainof Streptomyces closely related to Streptomyces vellosus(Lee et al., 2000b).
While pump selective inhibitors may have limitedtherapeutic use, they may prove invaluable for studyingthe contribution and prevalence of specific efflux pumps inclinical isolates of gram-positive and gram-negativebacteria. This approach has recently been successfullyapplied to P. aeruginosa and allowed simple identificationof strains over-expressing multiple efflux pumps (Cho etal., 1999).
The previous discussion highlighted some successesarising from high-throughput screening strategies for thediscovery of MDR EPIs. A quite different, rational approachhas recently been successfully applied by K. Lewis andcolleagues (Stermitz et al., 2000). They identified a groupof cationic berberine alkaloids as natural substrates ofbacterial MDR pumps (which render berberines essentiallyineffective). Berberine is produced by Berberis fremontii,and it was reasoned that the plant could dramaticallyimprove its defense against pathogens by combiningberberine with a natural MDR inhibitor. Indeed, a potentinhibitor of NorA, 5'-methoxyhydnocarpin (5'-MHC), hasbeen extracted from its leaves. No bacterial pathogens ofplants of the Berberis species are known. It is tempting toassume that one of the reasons is the presence of aneffective combination of antimicrobial with EPI.
De novo rational drug design based upon knowledgeof the tertiary structure of MDR transporters is not currentlyfeasible, given their size, complexity and association withmembranes. In anticipation of more advanced methodsfor solving protein structures, identification of amino acidresidues directly involved in recognition of substrates andinhibitors would be an informative exercise.
Conclusions
Efflux-mediated resistance is widespread in clinical settingsand is particularly important for antibiotics that are lessaffected by specific resistance mechanisms. It is clear thatthe use of efflux pump inhibitors is an approach that shouldbe explored, in addition to more conventional methods thatfocus on the discovery of novel classes of antibacterialagents or variants of older classes that are less susceptibleto efflux. Indeed, initial reports have demonstrated the multi-factorial benefits of the efflux pump inhibitor approach incombating drug resistance. Efflux pump inhibitors areexpected to 1) decrease intrinsic resistance (and
Figure 6. Broad-spectrum RND efflux pump inhibitors active in vivo.
Figure 7. MexAB-OprM selective EPI MC-510,051.

234 Lomovskaya and Watkins
consequently expand the spectrum of activity of someantibiotics to previously non-susceptible species); 2)reverse acquired resistance; and very importantly 3)decrease the frequency of emergence of resistant strains.
While this review was dedicated exclusively to theinhibitors of bacterial pumps, the same efflux pumpinhibitory approach can be applied to pathogenic fungi witha goal of improving the clinical performance of antifungaltherapy. Indeed, recently several inhibitors of MDR pumpsfrom various Candida species have been identified in ourscreening efforts. These compounds are currently beingextensively evaluated and will be the subject of the futurecommunications. It is clear that substantial efforts areneeded before inhibitors of efflux pumps can be usedclinically and be fully accepted by the medical community.
References
Abdel-Sayed S., Gonzalez M., and Eagon R. 1982. The role of the outermembrane of Pseudomonas aeruginosa in the uptake of aminoglycosideantibiotics. J. Antimicrob. Chemother. 10: 173-183.
Aires J. R., Kohler T., Nikaido H., and Plesiat P. 1999. Involvement of anactive efflux system in the natural resistance of Pseudomonas aeruginosato aminoglycosides. Antimicrob. Agents. Chemother. 43: 2624-2628.
Arcangioli M. A., Leroy-Setrin S., Martel J. L., and Chaslus-Dancla E. 1999.A new chloramphenicol and florfenicol resistance gene flanked by twointegron structures in Salmonella typhimurium DT104. FEMS Microbiol.Lett. 174: 327-32.
Arpin C., Canron M. H., Noury P., and Quentin C. 1999. Emergence ofmefA and mefE genes in beta-haemolytic streptococci and pneumococciin France. J. Antimicrob. Chemother. 44: 133-134.
Barclay M., Begg E., Chambers S., Thornley P., Pattemore P., and GrimwoodK. 1996. Adaptive resistance to tobramycin in Pseudomonas aeruginosalung infection in cystic finrosis. J. Antimicrob. Chemother. 37: 1155-1164.
Beck W., Cirtain M., Glover C., Felsted R., and Safa A. 1988. Effects ofindole alkaloids on multidrug resistance and labeling of P-glycoproteinby a photoaffinity analog of vinblastine. Biochem. Biophys. Res. Com.153: 959-966.
Biedenbach D. J., Barrett M. S., and Jones R. N. 1998. Comparativeantimicrobial activity and kill-curve investigations of novel ketolideantimicrobial agents (HMR 3004 and HMR 3647. tested againstHaemophilus influenzae and Moraxella catarrhalis strains. Diag. Microbiol.Infect. Dis. 31: 349-353.
Blais J., Cho D., Tangen K., Ford C., Lee A., Lomovskaya O., andChamberland S. 1999. Efflux pump inhibitors enhance the activity ofantimicrobial agents against a broad selection of bacteria. 39thInterscience Conference for Antimicrobial Agents and Chemotherapy, SanFrancisco.
Bolton L. F., Kelley L. C., Lee M. D., Fedorka-Cray P. J., and Maurer J. J.1999. Detection of multidrug-resistant Salmonella enterica serotypetyphimurium DT104 based on a gene which confers cross-resistance toflorfenicol and chloramphenicol. J. Clin. Microbiol. 37: 1348-1351.
Bonfiglio G., Laksai Y., Franchino L., Amicosante G., and Nicoletti G. 1998.Mechanisms of beta-lactam resistance amongst Pseudomonasaeruginosa isolated in an Italian survey. J. Antimicrob. Chemother. 42:697-702.
Brennan L., Duignan J., Petitras J., Anderson M., Fu W., Retsema J., RaiJ., Smyth K., Su W., and Sutcliffe J. 1998. CP-544372: MIC90 studiesand killing kinetics against key espiratory tract pathogens. 38thInterscience Conference for Antimicrobial Agents and Chemotherapy, SanDiego.
Brenwald N., Gill M., Davies T., and Appelbaum P. 2000. Efflux-mediatedresistance in mutants of Streptococcus pneumonia selected with newerfluoroquinolones. 40th Interscience Conference for Antimicrobial Agentsand Chemotherapy, Toronto.
Brighty K. E., and Gootz T. D. 1997. The chemistry and biological profile oftrovafloxacin. J. Antimicrob. Chemother. 39 Suppl B: 1-14.
Bush L. M., Calmon J., and Johnson C. C. 1995. Newer penicillins andbeta-lactamase inhibitors. Infect. Dis. Clin. North. Am. 9: 653-686.
Chen H. Y., Yuan M., and Livermore D. M. 1995. Mechanisms of resistanceto beta-lactam antibiotics amongst Pseudomonas aeruginosa isolatescollected in the UK in 1993. J. Med. Microbiol. 43: 300-309.
Cho D., Blais J., Tangen K., Ford K., Lee A., Lomovskaya O., ChamberlandS., and Miller G. 1999. Prevalence of efflux mechanisms among clinicalisolates of fluoroquinolone resistant Pseudomonas aeruginosa. 39thInterscience Conference for Antimicrobial Agents and Chemotherapy, San
Francisco.Chopra I., Hawkey P. M., and Hinton M. 1992. Tetracyclines, molecular and
clinical aspects. J. Antimicrob. Chemother. 29: 245-277.Clancy J., Petitpas J., Dib-Hajj F., Yuan W., Cronan M., Kamath A., Bergeron
J., and Retsema J. 1996. Molecular cloning and functional analysis of anovel macrolide resistance determinant mefA from Streptomycespyogenes. Mol. Microbiol. 22: 867-879.
Daikos G., Jackson G., Lolans V., and Livermore D. 1990. Adaptiveresistance to aminoglycoside antibiotics from first-exposure down-regulation. J. Infect. Dis. 162: 414-420.
Daikos G., Lolans V., and Jackson G. 1991. First-exposure adaptiveresistance to aminoglycoside antibiotics in vivo with meaning for optimalclinical use. Antimicrob. Agents Chemother. 35: 117-123.
Davies T., Ednie L., D. H., Pankuch G., Jacobs M., and Appelbaum P. 2000.Antipneumococcla activity of ABT-773 compared to those of 10 otheragents. Antimicrob. Agents Chemother. 44: 1894-1899.
Edgar R., and Bibi E. 1999. A single membrane-embedded negative chargeis critical for recognizing positively charged drugs by the Escherichia colimultidrug resistance protein MdfA. Embo J. 18: 822-832.
Everett M. J., Jin Y. F., Ricci V., and Piddock L. J. 1996. Contributions ofindividual mechanisms to fluoroquinolone resistance in 36 Escherichiacoli strains isolated from humans and animals. Antimicrob. AgentsChemother. 40: 2380-2386.
Fukuda H., Hori S., and Hiramatsu K. 1998. Antibacterial activity ofgatifloxacin (AM-1155, CG5501, BMS-206584), a newly developedfluoroquinolone, against sequentially acquired quinolone-resistant mutantsand the norA transformant of Staphylococcus aureus. Antimicrob. AgentsChemother. 42: 1917-1922.
Fukuda H., Hosaka M., Hirai K., and Iyobe S. 1990. New norfloxacinresistance gene in Pseudomonas aeruginosa PAO. Antimicrob. AgentsChemother. 34: 1757-1761.
Fukuda H., Hosaka M., Iyobe S., Gotoh N., Nishino T., and Hirai K. 1995.nfxC-type quinolone resistance in a clinical isolate of Pseudomonasaeruginosa. Antimicrob. Agents Chemother. 39: 790-792.
Gill M. J., Brenwald N. P., and Wise R. 1999. Identification of an effluxpump gene, pmrA, associated with fluoroquinolone resistance inStreptococcus pneumoniae. Antimicrob. Agents Chemother. 43: 187-189.
Giovanetti E., Montanari M. P., Mingoia M., and Varaldo P. E. 1999.Phenotypes and genotypes of erythromycin-resistant Streptococcuspyogenes strains in Italy and heterogeneity of inducibly resistant strains.Antimicrob. Agents Chemother. 43: 1935-1940.
Gootz T. D., and Brighty K. E. 1996. Fluoroquinolone antibacterials: SARmechanism of action, resistance, and clinical aspects. Med. Res. Rev.16: 433-86.
Gootz T. D., Zaniewski R. P., Haskell S. L., Kaczmarek F. S., and MauriceA. E. 1999. Activities of trovafloxacin compared with those of otherfluoroquinolones against purified topoisomerases and gyrA and grlAmutants of Staphylococcus aureus. Antimicrob. Agents Chemother. 43:1845-1855.
Griffith D., Lomovskaya O., Lee V., and Dudley M. 1999. Potentiation oflevofloxacin by a broad-spectrum efflux pump inhibitor in mouse modelsof infection caused by Pseudomonas aeruginosa. 39th InterscienceConference for Antimicrobial Agents and Chemotherapy, San Francisco.
Griffith D., Lomovskaya O., Lee V., and Dudley M. 2000. Potentiation oflevoflocaxin by MC-02,595, a broad-spectrum efflux pump inhibitor inmouse models of infection due to Pseudomonas aeruginosa withcombinations of different Mex pumps expression and target mutations.40th Interscience Conference for Antimicrobial Agents and Chemotherapy,Toronto.
Guerin F., Varon E., Hoi A., Gutmann L., and Podglajen I. 2000.Fluoroquinolone resistance associated with target mutations and activeefflux in oropharyngeal colonizing isolates of viridans group streptococci.Antimicrob. Agents Chemother. 44: 2197-2200.
Hancock R., Raffle V., and Nicas T. 1981. Involvement of the outermembrane in gentamicin and streptomycin uptake and killling inPseudomonas aeruginosa. Antimicrob. Agents Chemother. 19: 777-785.
Hirai K., Suzue S., Irikura T., Iyobe S., and Mitsuhashi S. 1987. Mutationsproducing resistance to norfloxacin in Pseudomonas aeruginosa.Antimicrob. Agents Chemother. 31: 582-586.
Hirata T., Nielsen J., Sato T., Nirira S., and A. Y. 1998. Screening of aninhibitor of the tetracycline efflux pump in a tetracycline-resistant clinical-isolate of Staphylococcus aureus 743. A Biol. Pharm. Bull. 21: 678.
Hsieh P. C., Siegel S. A., Rogers B., Davis D., and Lewis K. 1998. Bacterialacking a multidrug pump: a sensitive tool for drug discovery. Proc. Natl.Acad. Sci. USA. 95: 6602-6606.
Jakics E. B., Iyobe S., Hirai K., Fukuda H., and Hashimoto H. 1992.Occurrence of the nfxB type mutation in clinical isolates of Pseudomonasaeruginosa. Antimicrob. Agents Chemother. 36: 2562-2565.
Jalal S., and Wretlind B. 1998. Mechanisms of quinolone resistance in clinicalstrains of Pseudomonas aeruginosa. Microb. Drug Resist. 4: 257-261.

Efflux Pump Inhibition 235
Jamjian C., Biedenbach D. J., and Jones R. N. 1997. In vitro evaluation ofa novel ketolide antimicrobial agent, RU-64004. Antimicrob. AgentsChemother. 41: 454-459.
Kaatz G. W., and Seo S. M. 1997. Mechanisms of fluoroquinolone resistancein genetically related strains of Staphylococcus aureus. Antimicrob. AgentsChemother. 41: 2733-2737.
Karlovsky J., Saunders M., Harding G., Hoban D., and Zhanel G. 1996. Invitro characterization of aminoglycoside adaptive resistance inPseudomonas aeruginosa. Antimicrob. Agents Chemother. 40: 1387-1393.
Keyes K., Hudson C., Maurer J. J., Thayer S., White D. G., and Lee M. D.2000. Detection of florfenicol resistance genes in Escherichia coli isolatedfrom sick chickens. Antimicrob. Agents Chemother. 44: 421-424.
Kim E., and Aoki T. 1996. Sequence analysis of the florfenicol resistancegene encoded in the transferable R-plasmid of a fish pathogen, Pasteurellapiscicida. Microbiol. Immunol. 40: 665-9.
Kirst H. A. 1991. New macrolides: expanded horizons for an old class ofantibiotics. J. Antimicrob. Chemother. 28: 787-90.
Kohler T., Michea-Hamzehpour M., Epp S. F., and Pechere J. C. 1999.Carbapenem activities against Pseudomonas aeruginosa: respectivecontributions of OprD and efflux systems. Antimicrob. Agents Chemother.43: 424-7.
Kohler T., Michea-Hamzehpour M., Henze U., Gotoh N., Curty L. K., andPechere J. C. 1997a. Characterization of MexE-MexF-OprN, a positivelyregulated multidrug efflux system of Pseudomonas aeruginosa. Mol.Microbiol. 23: 345-54.
Kohler T., Michea-Hamzehpour M., Plesiat P., Kahr A. L., and Pechere J.C. 1997b. Differential selection of multidrug efflux systems by quinolonesin Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 41: 2540-3.
Lawrence L. E., and Barrett J. F. 1998. Efflux pumps in bacteria: overview,clinical relevance, and potential pharmaceutical target. Exp. Opin. Invest.Drugs 7: 199-217.
Lee A., Mao W., Warren M., Mistry A., Hoshino K., Okumura R., Ishida H.,and Lomovskaya O. 2000a. Interplay between efflux pumps may provideeither additive or multiplicative effects on drug resistance. J. Bacteriol.182: 3142-3150.
Lee J., Galazzo J., Ianiro T., Warren M., Cano R., Chamberland S.,Lomovskaya O., and Lee M. 2000b. MF-EA-371a and MF-EA-371b, novelbacterial efflux pump inhibitors from microbial phermentations. 40thInterscience Conference for Antimicrobial Agents and Chemotherapy,Toronto.
Leger R., Renau T. E., Zhang J. Z., Belej P., Flamme E. M., McMillen W.,Sangalang J., She M. W., Yen R., Chamberland S., Gannon C. L.,Lomovskaya O., and Lee V. J. 2000. Peptidomimetics of dipeptide amideefflux pump inhibitors potentiate the activity of levofloxacin inPseudomonas aeruginosa. 40th Interscience Conference for AntimicrobialAgents and Chemotherapy, Toronto.
Levy S. B. 1992. Active efflux mechanisms for antimicrobial resistance.Antimicrob. Agents Chemother. 36: 695-703.
Lewis K. 1994. Multidrug resistance pumps in bacteria: variations on atheme. Trends Biochem. Sci. 19: 119-123.
Li X. Z., Ma D., Livermore D. M., and Nikaido H. 1994. Role of efflux pump(s)in intrinsic resistance of Pseudomonas aeruginosa: active efflux as acontributing factor to beta-lactam resistance. Antimicrob. AgentsChemother. 38: 1742-1752.
Lomovskaya O., Lee A., Hoshino K., Ishida H., Mistry A., Warren M. S.,Boyer E., Chamberland S., and Lee V. J. 1999a. Use of a genetic approachto evaluate the consequences of inhibition of efflux pumps inPseudomonas aeruginosa. Antimicrob. Agents Chemother. 43: 1340-6.
Lomovskaya O., Lee A., Warren M., Galazzo J., Fronko R., Lee M.,Chamberland S., Hecker S., Lee V., Ishida H., and Hoshino K. 1999b.Targeting efflux pumps in Pseudomonas aeruginosa. 39th InterscienceConference for Antimicrobial Agents and Chemotherapy, San Francisco.
Lomovskaya O., Warren M., Lee A., Galazzo J., Fronko R., Lee M., Blais,J., Cho, D., Chamberland S., Renau, T., Leger, R., Hecker, S., Watkins,W., Hoshino, K., Ishida H., and Lee, V.J. 2001. Identification andcharacterization of inhibitors of multidrug resistance efflux pumps inPseudomonas aeruginosa: Novel agents for combination therapy.Antimicrob. Agents Chemother. 45: 105-116.
Luna V., Jernigan D., Tice A., Kellner J., and Roberts M. 1999. A variety ofgram-positive bacteria carry mobile mef genes. J. Antimicrob. Chemother.44: 19-25.
Lynch C., Courvalin P., and Nikaido H. 1997. Active efflux of antimicrobialagents in wild-type strains of enterococci. Antimicrob. Agents Chemother.41: 869-871.
Mao W., Warren M., Lee A., and Lomovskaya O. 2000. MexXY-OprM effluxpump is required for antagonism of aminoglycosides by divalent cationsin Pseudomonas aeruginosa. 40th Interscience Conference forAntimicrobial Agents and Chemotherapy, Toronto.
Markham P. N. 1999. Inhibition of the emergence of ciprofloxacin resistance
in Streptococcus pneumoniae by the multidrug efflux inhibitor reserpine.Antimicrob. Agents Chemother. 43: 988-989.
Markham P. N., and Neyfakh A. A. 1996. Inhibition of the multidrugtransporter NorA prevents emergence of norfloxacin resistance inStaphylococcus aureus. Antimicrob. Agents Chemother. 40: 2673-2674.
Markham P. N., Westhaus E., Klyachko K., Johnson M. E., and Neyfakh A.A. 1999. Multiple novel inhibitors of the NorA multidrug transporter ofStaphylococcus aureus. Antimicrob. Agents Chemother. 43: 2404-2408.
Masuda N., Gotoh N., Ishii C., Sakagawa E., Ohya S., and Nishino T. 1999.Interplay between chromosomal beta-lactamase and the MexAB-OprMefflux system in intrinsic resistance to beta-lactams in Pseudomonasaeruginosa. Antimicrob. Agents Chemother. 43: 400-402.
Masuda N., Sakagawa E., Ohya S., Gotoh N., Tsujimoto H., and Nishino T.2000. Contribution of the MexXY-OprM efflux system to intrinsic resistancein Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 44: 2242-2246.
Medeiros A., O’Brien T., Wacker W., and Yulug N. 1971. Effect on saltconcentration on the apparent in vitro susceptibility of Pseudomonas andother gram-negative bacilli to gentamicin. J. Infect. Dis. 124: 59-64.
Miller G. H., Sabatelli F. J., Hare R. S., Glupczynski Y., Mackey P., ShlaesD., Shimizu K., and Shaw K. J. 1997. The most frequent aminoglycosideresistance mechanisms—changes with time and geographic area: areflection of aminoglycoside usage patterns? Aminoglycoside ResistanceStudy Groups. Clin. Infect. Dis. 24 Suppl 1: S46-62.
Mitchell B. A., Paulsen I. T., Brown M. H., and Skurray R. A. 1999.Bioenergetics of the staphylococcal multidrug export protein QacA.Identification of distinct binding sites for monovalent and divalent cations.J. Biol. Chem. 274: 3541-3548.
Moore R. A., DeShazer D., Reckseidler S., Weissman A., and Woods D. E.1999. Efflux-mediated aminoglycoside and macrolide resistance inBurkholderia pseudomallei. Antimicrob. Agents Chemother. 43: 465-470.
Morshed S. R., Lei Y., Yoneyama H., and Nakae T. 1995. Expression ofgenes associated with antibiotic extrusion in Pseudomonas aeruginosa.Biochem. Biophys. Res. Com. 210: 356-362.
Nakae T., Nakajima A., Ono T., Saito K., and Yoneyama H. 1999. Resistanceto beta-lactam antibiotics in Pseudomonas aeruginosa due to interplaybetween the MexAB-OprM efflux pump and beta-lactamase. Antimicrob.Agents Chemother. 43: 1301-1303.
Nelson M. L., and Levy S. B. 1999. Reversal of tetracycline resistancemediated by different bacterial tetracycline resistance determinants byan inhibitor of the Tet(B) antiport protein. Antimicrob. Agents Chemother.43: 1719-1724.
Nelson M. L., Park B. H., Andrews J. S., Georgian V. A., Thomas R. C., andLevy S. B. 1993. Inhibition of the tetracycline efflux antiport protein by 13-thio- substituted 5-hydroxy-6-deoxytetracyclines. J. Med. Chem. 36: 370-377.
Nelson M. L., Park B. H., and Levy S. B. 1994. Molecular requirements forthe inhibition of the tetracycline antiport protein and the effect of potentinhibitors on the growth of tetracycline-resistant bacteria. J. Med. Chem.37: 1355-1361.
Neyfakh A. A., Borsch C. M., and Kaatz G. W. 1993. Fluoroquinoloneresistance protein NorA of Staphylococcus aureus is a multidrug effluxtransporter. Antimicrob. Agents Chemother. 37: 128-129.
Ng E. Y., Trucksis M., and Hooper D. C. 1994. Quinolone resistancemediated by norA: physiologic characterization and relationship to flqB,a quinolone resistance locus on the Staphylococcus aureus chromosome.Antimicrob. Agents Chemother. 38: 1345-1355.
Nikaido H. 1994. Prevention of drug access to bacterial targets: permeabilitybarriers and active efflux. Science. 264: 382-388.
Nikaido H. 1998a. Antibiotic resistance caused by gram-negative multidrugefflux pumps. Clin. Infect. Dis. 27 Suppl 1: S32-41.
Nikaido H. 1998b. Multiple antibiotic resistance and efflux. Curr. Opin.Microbiol. 1: 516-523.
Nikaido H. 1998c. The role of outer membrane and efflux pumps in theresistance of gram-negative bacteria. can we improve drug access? DrugRes. Updates. 1: 93-98.
Nikaido H., and Vaara M. 1985. Molecular basis of bacterial outer membranepermeability. Microbiol. Rev. 49: 1-32.
Nishijima T., Saito Y., Aoki A., Toriya M., Toyonaga Y., and Fujii R. 1999.Distribution of mefE and ermB genes in macrolide-resistant strains ofStreptococcus pneumoniae and their variable susceptibility to variousantibiotics. J. Antimicrob. Chemother. 43: 637-643.
Oethinger M., Kern W. V., Jellen-Ritter A. S., McMurry L. M., and Levy S. B.2000. Ineffectiveness of topoisomerase mutations in mediating clinicallysignificant fluoroquinolone resistance in Escherichia coli in the absenceof the AcrAB efflux pump. Antimicrob. Agents Chemother. 44: 10-13.
Oliver A., Canton R., Campo P., bacuero F., and Blazquez J. 2000. Highfrequency of hypermutable Pseudomonas aeruginosa in cystic fibrosislung infection. Science. 288: 1251-1253.
Paulsen I. T., Sliwinski M. K., and Saier M. H., Jr. 1998. Microbial genome

236 Lomovskaya and Watkins
analyses: global comparisons of transport capabilities based onphylogenies, bioenergetics and substrate specificities. J. Mol. Biol. 277:573-592.
Pestova E., Beyer R., Cianciotto N. P., Noskin G. A., and Peterson L. R.1999. Contribution of topoisomerase IV and DNA gyrase mutations inStreptococcus pneumoniae to resistance to novel fluoroquinolones.Antimicrob. Agents Chemother. 43: 2000-2004.
Piddock L. 1999. Mechanism of fluoroquinolone resistance: an update 1994-1998. Drugs. 58: 11-18.
Piddock L. J., Jin Y. F., and Everett M. J. 1997. Non-gyrA-mediatedciprofloxacin resistance in laboratory mutants of Streptococcuspneumoniae. J. Antimicrob. Chemother. 39: 609-615.
Piddock L. J., Johnson M., Ricci V., and Hill S. L. 1998. Activities of newfluoroquinolones against fluoroquinolone-resistant pathogens of the lowerrespiratory tract. Antimicrob. Agents Chemother. 42: 2956-2960.
Poole K. 2000. Efflux-mediated resistance to fluoroquinolones in gram-negative bacteria. Antimicrob. Agents Chemother. 44: 2233-2241.
Poole K., Gotoh N., Tsujimoto H., Zhao Q., Wada A., Yamasaki T., NeshatS., Yamagishi J., Li X. Z., and Nishino T. 1996a. Overexpression of themexC-mexD-oprJ efflux operon in nfxB-type multidrug-resistant strainsof Pseudomonas aeruginosa. Mol. Microbiol. 21: 713-724.
Poole K., Heinrichs D. E., and Neshat S. 1993. Cloning and sequenceanalysis of an EnvCD homologue in Pseudomonas aeruginosa: regulationby iron and possible involvement in the secretion of the siderophorepyoverdine. Mol. Microbiol. 10: 529-544.
Poole K., Tetro K., Zhao Q., Neshat S., Heinrichs D. E., and Bianco N.1996b. Expression of the multidrug resistance operon mexA-mexB-oprMin Pseudomonas aeruginosa: mexR encodes a regulator of operonexpression. Antimicrob. Agents Chemother. 40: 2021-2028.
Poutanen S. M., de Azavedo J., Willey B. M., Low D. E., and MacDonald K.S. 1999. Molecular characterization of multidrug resistance inStreptococcus mitis. Antimicrob. Agents Chemother. 43: 1505-1507.
Putman M., Koole L., van Veen H., and Konings W. 1999. The secondarymultidrug transporter LmrP contains multiple drug interaction sites.Biochemistry. 38: 13900-13905.
Renau T., Leger R., Gannon K., Mathias K., Lomovskaya O., ChamberlandS., Lee V., Ohta T., nakayama K., and Ishida R. 2000. Addressing thestability of c-capped dipeptide efflux pump inhibotors that potentiate theactivity of levofloxacin in Pseudomonas asruginosa. 40th InterscienceConference for Antimicrobial Agents and Chemotherapy, Toronto.
Renau T. E., Leger R., Flamme E. M., Sangalang J., She M. W., Yen R.,Gannon C. L., Griffith D., Chamberland S., Lomovskaya O., Hecker S. J.,Lee V. J., Ohta T., and Nakayama K. 1999. Inhibitors of efflux pumps inPseudomonas aeruginosa potentiate the activity of the fluoroquinoloneantibacterial levofloxacin. J. Med. Chem. 42: 4928-4931.
Roberts M., Sutcliffe J., Courvalin P., Jensen L., Rood J., and Seppala H.1999. Nomenclature for macrolide and macrolide-lincosamide-streptogramine B resistance determinants. Antimicrob. Agents Chemother.43: 2823-2830.
Roberts M. C. 1994. Epidemiology of tetracycline-resistance determinants.Trends Microbiol. 2: 353-7.
Rosenberg E., Ma D., and Nikaido H. 2000. AcrD of Eschericia coli is anaminoglycoside efflux pump. J. Bacteriol. 182: 1754-1756.
Saier M., Beatty J., Goffeau A., Harley K., Heijne W., Huang S., Jack D.,Jahn P., Lew K., Liu J., Pao S., Paulsen I., T. T., and Virk P. 1999a. Themajor facilitator superfamily. J. Mol. Microbiol. Biothechnol. 1: 257-279.
Saier M. H., Jr. 1998. Molecular phylogeny as a basis for the classificationof transport proteins from bacteria, archaea and eukarya. Adv. Microb.Physiol. 40: 81-136.
Saier M. H., Jr., Eng B. H., Fard S., Garg J., Haggerty D. A., Hutchinson W.J., Jack D. L., Lai E. C., Liu H. J., Nusinew D. P., Omar A. M., Pao S. S.,Paulsen I. T., Quan J. A., Sliwinski M., Tseng T. T., Wachi S., and YoungG. B. 1999b. Phylogenetic characterization of novel transport proteinfamilies revealed by genome analyses. Biochim. Biophys. Acta. 1422: 1-56.
Saier M. H., Jr., Paulsen I. T., Sliwinski M. K., Pao S. S., Skurray R. A., andNikaido H. 1998. Evolutionary origins of multidrug and drug-specific effluxpumps in bacteria. Faseb J. 12: 265-274.
Sanchez L., Leranoz S., Puig M., Loren J. G., Nikaido H., and Vinas M.1997. Molecular basis of antimicrobial resistance in non-typableHaemophilus influenzae. Microbiologia. 13: 309-314.
Schnappinger D., and Hillen W. 1996. Tetracyclines: antibiotic action, uptake,and resistance mechanisms. Arch. Microbiol. 165: 359-369.
Sheridan R. P., and Chopra I. 1991. Origin of tetracycline efflux proteins:conclusions from nucleotide sequence analysis. Mol. Microbiol. 5: 895-900.
Silvia A., Gavitt N., Pere W., and Hayashi S. 1999. Identification of ateracycline efflux resistance reverser UK-57,562. 39th InterscienceConference for Antimicrobial Agents and Chemotherapy, San Francisco.
Someya Y., Yamaguchi A., and Sawai T. 1995. A novel glycylcycline, 9-
(N,N-dimethylglycylamido)-6-demethyl-6- deoxytetracycline, is neithertransported nor recognized by the transposon Tn10-encoded metal-tetracycline/H+ antiporter. Antimicrob. Agents Chemother. 39: 247-249.
Srikumar R., Yan H., and hancock R. P., K. 2000. Role of OprM in theintrinsic drug resistance and pathogenicity of Pseudomonas aeruginosa.40th Interscience Conference for Antimicrobial Agents and Chemotherapy,Toronto.
Stein W. 1997. Kinetics of the multidrug transporter (P-glycoprotein) andits reversal. Physiol. Rev. 77: 545-590.
Stermitz F., Lorenz P., Tawara J., Zenewicz L., and Lewis. K. 2000. Synergyin a medicinal plant: antimicrobial action of berberine potentiated by 5'-methoxyhydnocarpine, a multidrug pump inhibitor. Proc. Natl. Acad. Sci.USA. 97: 1433-1437.
Sum P. E., Sum F. W., and Projan S. J. 1998. Recent developments intetracycline antibiotics. Curr. Pharm. Des. 4: 119-132.
Tait-Kamradt A., Clancy J., Cronan M., Dib-Hajj F., Wondrack L., Yuan W.,and Sutcliffe J. 1997. mefE is necessary for the erythromycin-resistant Mphenotype in Streptococcus pneumoniae. Antimicrob. Agents Chemother.41: 2251-2255.
Thanassi D. G., Cheng L. W., and Nikaido H. 1997. Active efflux of bilesalts by Escherichia coli. J. Bacteriol. 179: 2512-2518.
Trias J., Chamberland C., Hecker S., and Lee V. J. 1999. Efflux PumpInhibitors. U. S. patent 5,989,832.
Trias J., and Nikaido H. 1990. Outer membrane protein D2 catalyzesfacilitated diffusion of carbapenems and penems through the outermembrane of Pseudomonas aeruginosa. Antimicrob. Agents Chemother.34: 52-57.
Tseng T., Gratwick K., Kollman J., Park D., Nies D., Goffeau A., and SaierM. 1999. The RND permease surefamily: an ancient, ubiquitous anddiverse family that includes human disease and development proteins. J.Mol. Microbiol. Biotechnol. 1: 107-125.
Van Bembeke F., Balzi E., and Tulkens P. 2000. Antibiotic efflux pumps.Biochem. Pharmacol. 60: 457-470.
Warren M., Lee J., Lee A., Hoshino K., Ishida H., and lomovskaya O. 2000.An inhibitor of Mex pumps from Pseudomonas aeruginosa is also asubstrate of these pumps. 40th Interscience Conference for AntimicrobialAgents and Chemotherapy, Toronto.
Westbrock-Wadman S., Sherman D. R., Hickey M. J., Coulter S. N., Zhu Y.Q., Warrener P., Nguyen L. Y., Shawar R. M., Folger K. R., and Stover C.K. 1999. Characterization of a Pseudomonas aeruginosa Efflux PumpContributing to Aminoglycoside Impermeability. Antimicrob. AgentsChemother. 43: 2975-2983.
Xiong Y., Caillon J., Kergueris M., Drugeon H., Baron D., Potel G., andBayer A. 1997. Adaptive resistance of Pseudomonas aeruginosa inducedby aminoglycosides and killing kinetics in a rabit endocarditis model.Antimicrob. Agents Chemother. 41: 823-826.
Yamada H., Kurose-Hamada S., Fukuda Y., Mitsuyama J., Takahata M.,Minami S., Watanabe Y., and Narita H. 1997. Quinolone susceptibility ofnorA-disrupted Staphylococcus aureus. Antimicrob. Agents Chemother.41: 2308-2309.
Yerushalmi H., Lebendiker M., and Schuldiner S. 1995. EmrE, anEscherichia coli 12-kDa multidrug transporter, exchanges toxic cationsand H+ and is soluble in organic solvents. J. Biol. Chem. 270: 6856-6863.
Yoneyama H., Ocaktan A., Tsuda M., and Nakae T. 1997. The role of mex-gene products in antibiotic extrusion in Pseudomonas aeruginosa.Biochem. Biophys. Res. Com. 233: 611-618.
Yoshida H., Bogaki M., Nakamura M., and Nakamura S. 1990a. Quinoloneresistance-determining region in the DNA gyrase gyrA gene of Escherichiacoli. Antimicrob. Agents Chemother. 34: 1271-1272.
Yoshida H., Bogaki M., Nakamura S., Ubukata K., and Konno M. 1990b.Nucleotide sequence and characterization of the Staphylococcus aureusnorA gene, which confers resistance to quinolones. J. Bacteriol. 172: 6942-6949.
Zgurskaya H., and Nikaido H. 2000. Multidrug resistance mechanisms: drugefflux across two membranes. Mol. Microbiol. 37: 219-225.
Zgurskaya H. I., and Nikaido H. 1999a. AcrA is a highly asymmetric proteincapable of spanning the periplasm. J. Mol. Biol. 285: 409-420.
Zgurskaya H. I., and Nikaido H. 1999b. Bypassing the periplasm:reconstitution of the AcrAB multidrug efflux pump of Escherichia coli. Proc.Natl. Acad. Sci. USA. 96: 7190-7195.
Ziha-Zarifi I., Llanes C., Kohler T., Pechere J. C., and Plesiat P. 1999. Invivo emergence of multidrug-resistant mutants of Pseudomonasaeruginosa overexpressing the active efflux system MexA-MexB-OprM.Antimicrob. Agents Chemother. 43: 287-291.
Zimelis V., and Jackson G. 1973. Activity of aminoglycoside antibioticsagainst Pseudomonas aeruginosa: specificity and site of calcium andmagnesium antagonism. J. Infect. Dis. 127: 663-669.