inhibition of myogenic differentiation in myoblasts - development
TRANSCRIPT

INTRODUCTION
The decision of a cell to proliferate, differentiate and migrateduring development is intimately connected to its environment.Transplantation and ablation studies have shown the impor-tance of cell-cell interactions and the extracellular milieu in thedetermination of cell fate (Greenwald and Rubin, 1992;Gurdon, 1992). Several peptide growth factors have beenshown to play a role in inductive processes that give rise to dif-ferentiated cell types (Jessell and Melton, 1992). Transform-ing growth factor-
β (TGF-β), one such factor, affects thegrowth and differentiation of many cell types in vitro, espe-cially those of mesenchymal origin (Ignotz and Massagué,1985; Rosen et al., 1988; Torti et al., 1989). Its expressionpattern during mouse development also suggests an importantfunction in specific morphogenetic and differentiation eventsin vivo (Heine et al., 1987; Millan et al., 1991; Pelton et al.,1991). In particular, TGF-β is highly expressed during periodsof morphogenesis or remodeling of mesenchyme and at sitesof mesenchymal-epithelial interactions (Heine et al., 1987).
TGF-β may play an important role in myogenic differen-tiation (Olson et al., 1986). TGF-β is thought to be involved incardiomyogenesis (Potts et al., 1989, 1991) and its expressionis induced during experimental myocardial infarction(Thompson et al., 1988; Lefer et al., 1990). However, the roleof TGF-β in skeletal myoblast differentiation is unclear, sinceexogenous TGF-β has both positive and negative effects onmuscle cell development in vitro. On the one hand, treatmentof skeletal myoblast cell lines or primary muscle cells withTGF-β in low serum inhibits terminal differentiation(Massagué et al., 1986). In addition, TGF-β blocks theexpression and function of two muscle-specific transcriptionfactors, MyoD (Vaidya et al., 1989) and myogenin (Brennanet al., 1991), thereby preventing expression of downstreammuscle transcripts. On the other hand, TGF-β in normal serumcan induce differentiation of myoblasts (Zentella andMassagué, 1992) and treatment of embryonic stem cells withTGF-β results in preferential differentiation of cells intomuscle (Slager et al., 1993).
TGF-β acts by binding to a set of specific receptors
1085Development 120, 1085-1095 (1994)Printed in Great Britain © The Company of Biologists Limited 1994
Transforming growth factor-β (TGF-β) is thought to playa role in mesenchymal cell development and, specifically,in muscle differentiation, yet its precise role in the latterprocess remains unclear. TGF-β has been shown to bothinhibit and induce myoblast maturation in vitro, dependingon the culture conditions. Whether the type I or type IITGF-β receptor mediates the various TGF-β effects onmyogenesis is not known. In the present study, C2C12myoblasts were transfected with an expression vector for atruncated type II TGF-β receptor, which has been shownto act as a dominant negative inhibitor of type II receptorsignaling. In contrast to the parental cells, the transfectedclones did not efficiently form myotubes or induceexpression of MyoD, myogenin and several other differen-tiation markers following incubation in low serum media.However, some muscle differentiation markers continuedto be expressed in the transfected cells suggesting that atleast two pathways are involved in muscle cell differen-tiation. These cells could still growth arrest in low serummedia, showing that decreased proliferation can be disso-
ciated from differentiation. Unlike several oncogenesknown to block myogenic differentiation, expression of thetruncated TGF-β receptor did not result in myoblast trans-formation. Injection of the parental or the transfectedC2C12 cells into the limb muscle of nude mice revealedquantitative and qualitative differences in their behavior,and suggested that myoblasts expressing the truncatedTGF-β receptor cannot fuse in vivo. Finally, retrovirus-mediated expression of MyoD in the transfected cellsrestored their ability to form myotubes in vitro, indicatingthat inhibition of myoblast differentiation by the truncatedTGF-β receptor may depend on decreased MyoDexpression. We propose that TGF-β signaling through thetype II receptor is required for several distinct aspects ofmyogenic differentiation and that TGF-β acts as a compe-tence factor in this multistep process.
Key words: TGF-β, differentiation, muscle, myoblasts, signaling,growth factor
SUMMARY
Inhibition of myogenic differentiation in myoblasts expressing a truncated
type II TGF-β receptor
Ellen H. Filvaroff, Reinhard Ebner and Rik Derynck*
Departments of Growth and Development, and Anatomy, Programs in Cell Biology and Developmental Biology, University ofCalifornia at San Francisco, San Francisco, CA 94143-0640, USA
*Author for correspondence

1086
(Massagué, 1992). Of these, the type I and type II TGF-βreceptors mediate most of its biological effects (Laiho et al.,1990a, 1991; Geiser et al., 1992). The cytoplasmic domains ofthe cloned type I and type II TGF-β receptors have sequencescharacteristic of serine-threonine kinases (Lin et al., 1992;Ebner et al., 1993a, Franzén et al., 1993), and the type IIreceptor has been shown to be a functional kinase (Wrana etal., 1992). A physical association between the type I and typeII receptors has been proposed (Wrana et al., 1992), and thetype I receptor may require the type II receptor for its activity(Ebner et al., 1993a,b, Franzén et al., 1993, Bassing et al.,1994). Functional inactivation of the type II receptor in anepithelial cell line has indicated that the type II receptor isrequired for the antiproliferative effect of TGF-β and that itseffect on extracellular matrix protein synthesis may bemediated through the type I receptor (Chen et al., 1993). Whichof these two receptors mediates the effects of TGF-β on myo-genesis is not yet known. Whereas most cells in culture haveboth receptors, the type II receptor is expressed in vivo at highlevels in undifferentiated mesenchyme and in differentiatedmuscle tissue (Lawler et al., 1994). The co-localization ofTGF-β expression suggests a developmental role for TGF-βduring muscle cell differentiation (Heine et al., 1987; Pelton etal., 1991; Lawler et al., 1994).
To address the seemingly paradoxical observations on theeffects of TGF-β on myoblasts and to gain insight into its roleduring myogenic differentiation, we transfected C2C12myoblasts (Yaffe and Saxel, 1977; Blau et al., 1983) with atruncated form of the type II TGF-β receptor. This truncatedreceptor inhibits signaling by the type II, but not the type I,TGF-β receptor in a dominant negative fashion (Brand et al.,1993; Chen et al., 1993). We show that myoblasts expressingthe truncated type II TGF-β receptor do not undergo morpho-logical or biochemical differentiation. This inhibition of differ-entiation is associated with decreased expression of themyogenic determining genes, MyoD and myogenin, and canbe rescued following infection with a MyoD-expressing retro-virus. These studies suggest that some, but not all, changesassociated with myotube formation require signaling throughthe TGF-β receptor, and that TGF-β functions in an autocrineor paracrine fashion as a competence factor for myogenicdifferentiation.
MATERIALS AND METHODS
Cell culture and transfectionsC2C12 cells which have been clonally purified for reproduciblemyogenic differentiation (Blau et al., 1983) from C2 cells (Yaffe andSaxel, 1977), were obtained from Dr H. Blau and grown in 20% fetalcalf serum (FCS) in DMEM. To induce differentiation intomyotubes, the cells were switched into DMEM containing 2% horseserum for 1 to 3 days. C2C12 cells were transfected withpcDNA1Neo (InVitrogen) expressing the truncated type II TGF-βreceptor and encoding neomycin resistance (Chen et al., 1993) usingthe calcium phosphate precipitation method (Sambrook, 1989). Thetransfected cells were then cultured in medium containing G418 (400µg/ml). After 14 to 21 days, G418 resistant clones were isolated,expanded and screened by northern hybridization for expression ofmRNA for the truncated receptor. The three clones selected for thisstudy were chosen on the basis of cell surface expression of thetruncated receptor, as determined by chemical cross-linking using
125I-TGF-β. Five randomly selected neomycin-resistant, control-transfected clones were also analyzed and found to undergo normalmyogenic differentiation.
Northern hybridization analysis3 days after plating equal numbers of cells (determined using aCoulter counter), cells were switched to differentiation media (2%horse serum) or were kept in growth media (20% fetal calf serum) forthree more days. RNA was isolated (Chomczynski and Sacchi, 1987)and analyzed by northern analysis (Sambrook, 1989) using cDNAprobes radiolabelled by random priming using a commercial kit(Boehringer Mannheim). Following hybridization at 42˚C for 24hours, the nitrocellulose blots were washed in 0.5× SSC, 0.5% SDSfor 20 minutes at 42˚C and 0.1× SSC, 0.5% SDS for 30 minutes at55˚C. cDNAs for myosin light chain 2, MyoD (Davis et al., 1987) andmyosin light chain 1/3 (Periasamy et al., 1984, modified by Dr N.Rosenthal), the acetylcholine receptor α subunit (Isenberg et al.,1986) and myogenin (Wright et al., 1989) were used as hybridizationprobes to assess the differentiation state.
Receptor cross-linking analysisRecombinant human TGF-β1 was 125I-labelled using a slightlymodified chloramine T method (Frolik et al., 1984). Cross-linking ofparental and transfected C2C12 cells was carried out as described(Gazit et al., 1993).
Western analysisEqual quantities of protein (determined by the Biorad colorimetricassay) were electrophoresed in 12% (for troponin T or desmin) or7.5% (for pRB or myosin heavy chain) denaturing polyacrylamidegels and transferred to nitrocellulose membranes at 40 V for 2 hours.The blots were washed for 10 minutes in TBST (25 mM Tris pH 8.0,125 mM NaCl, 0.025% Tween 20), incubated in blocking buffer (25mM Tris pH 8.0, 125 mM NaCl, 0.1% Tween 20, 1% BSA, 0.1%NaN3) for 2 hours at room temperature or 24 hours at 4˚C, and thenincubated with the antibody for 2 hours at room temperature. Theywere then washed three times with blocking buffer (15 minutes perwash) and incubated with a secondary, alkaline phosphatase-conju-gated antibody (Promega) for 1 hour. After three 15 minute washeswith blocking buffer and three 5 minute washes with TBST, the blotswere incubated in phosphatase substrate (BCIP/NBT, Kirkegaard andPerry Laboratories) for 5-30 minutes and, once developed, washedwith distilled water. Antibodies for troponin T, myosin heavy chain(i.e. MF20 antibody, Sigma) and desmin were provided by Dr CharlesOrdahl (UCSF). The antibodies to pRB (DeCaprio et al., 1988) werepurchased from Pharmingen.
ImmunofluorescenceCells on coverslips were washed three times with phosphate-bufferedsaline (PBS) and fixed in methanol (for myosin heavy chain antibod-ies) or in 4% formaldehyde in PBS (for troponin T or desmin anti-bodies) for 10 minutes at room temperature. After washing with PBS,cells were incubated in 0.1% Triton in PBS for 10 minutes and rinsedagain with PBS. The samples were then treated with 3% bovine serumalbumin (BSA) in PBS for 15-30 minutes and incubated with primaryantibody for 45 minutes at 37˚C. After three washes in PBS, the cellswere incubated with a rhodamine-conjugated secondary antibody for45 minutes at 37˚C. The cells were then washed three times with PBS,hydrated in 70% then 100% ethanol (3 minute washes each), air dried,mounted with Fluoromount-G (Fisher Scientific) and a coverslip, andphotographed with a Zeiss Axioplan microscope..
Growth curvesOn day 0, the cells were counted and plated such that by the day oftreatment, all samples would have approximately the same cellnumber. On day 4, one set of plates was changed to differentiationmedia (2% horse serum), while the other set remained in growth
E. H. Filvaroff, R. Ebner and R. Derynck

1087Myogenesis and type II TGF-β receptor
media (20% serum). All cells were changed and counted every otherday using a Coulter counter.
Retrovirus infectionsTo construct the retrovirus encoding hygromycin resistance and β-galactosidase, the β
-galactosidase gene was cut from plasmid pML62(constructed by Drs M. Landowski and G. Martin) with SalI andBamHI and ligated into the SalI and BamHI sites of a derivative ofpLXSH (Miller et al., 1993), obtained from Drs M. Lochric and H.Varmus. The packaging cell line, PA317 (Miller et al., 1993), wastransfected with this plasmid and the retrovirus-containing condi-tioned media, harvested 48 hours later, was used to infect anotherpackaging cell line, PE501. These cells were grown for 2 weeks inDMEM/10% serum with hygromycin, and individual colonies wereisolated and grown. Viruses produced from these clones were titeredusing NIH-3T3 fibroblasts, and the highest titer virus stock was usedto infect C2C12 cells. 48 hours after infection, C2C12 cells wereswitched to growth media with hygromycin (500 µg/ml) and selectedfor two weeks prior to X-gal staining.
The high titer MyoD retrovirus was kindly supplied by Dr DustyMiller (Weintraub et al., 1989). Infections were performed by incu-bating growing cells overnight with viral stock plus 4 µg/ml ofpolybrene.
In vivo injections and analysisFor intramuscular injections into mice, the cells were trypsinized,washed twice and resuspended in PBS at 4˚C. Approximately 4×105
myoblasts (in 20 µl) were delivered via 4 injections (5 µl each) intothe hind limbs of anesthetized 3- to 4-week-old nude mice. 2 weekslater, these limbs were removed from euthanized mice and frozen inisopentane, serially sectioned, and fixed and stained for β-galactosi-dase activity as described (Dhawan et al., 1991). Muscle fibers wereexamined across multiple serial sections throughout the length of thelimb. Sections were mounted in Gel/Mount (Biomeda Corporation)and photographed with a Zeiss Axioplan microscope.
RESULTS
Myoblast fusion is inhibited in cells expressing atruncated type II TGF-β receptor The mouse C2C12 myoblast cell line (Yaffe and Saxel, 1977;Blau et al., 1983) is frequently used as a model to study theprocess of myogenic differentiation in vitro. This clonal cellpopulation has highly reproducible differentiation properties(Blau et al., 1983). These cells proliferate as mononuclearmyoblasts in high (20%) serum media (growth media), butbecome growth arrested and undergo morphological and bio-chemical differentiation after a switch to low (2%) serummedia (differentiation media) (Blau et al., 1983). To examinethe role of TGF-β and its signaling through the type II receptorduring myogenesis, C2C12 cells were transfected with atruncated type II TGF-β receptor expression vector contain-ing a neomycin-resistance marker. This truncated receptor,which lacks its cytoplasmic domain, has been shown to inhibitsignaling specifically through the type II TGF-β receptor in adominant negative fashion (Chen et al., 1993). After 2 weeks,G418-resistant colonies were isolated, propagated as stablecell lines, and tested for expression of the truncated receptormRNA by northern blotting (data not shown). To verifyexpression of the truncated TGF-β receptor protein, the trans-fected cells were incubated with 125I-labelled TGF-β, and thecross-linked receptors were analyzed by polyacrylamide gelelectrophoresis. As shown in Fig. 1A, the truncated receptor
was expressed at the cell surface of transfected clones andbound TGF-β. The 125I-TGF-β cross-linked receptor could beimmunoprecipitated with antibodies against an epitope tagengineered at its carboxy terminus (Fig. 1B). Of the fiveclones that expressed the truncated receptor mRNA, threeshowed expression of the corresponding protein at the cellsurface. These three clones were used for further analysis andcompared with parental and neomycin-resistant controlC2C12 cells.
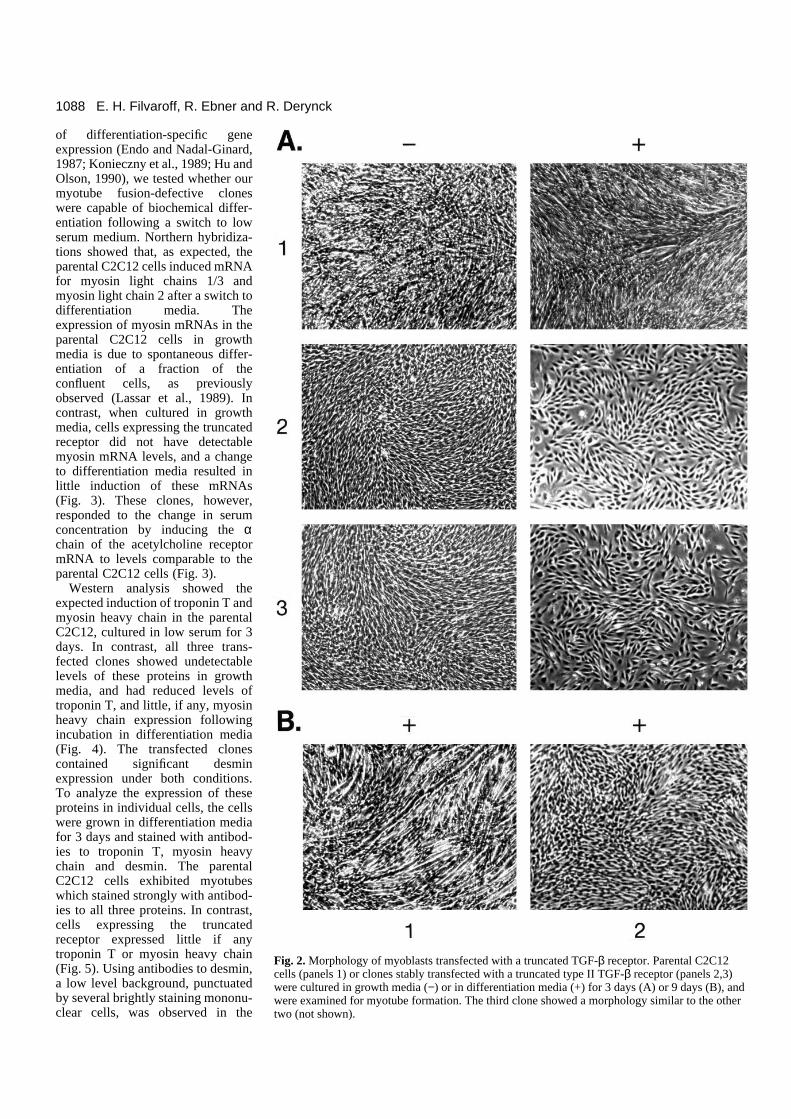
Under proliferative conditions, cells transfected with thetruncated receptor were morphologically similar to the parentalC2C12 cells (Fig. 2A−). To induce differentiation, cells wereswitched to low serum (2%) media. Within 3 days, the parentalC2C12 cells ceased proliferating and fused to form multinu-cleated fibers. In contrast, in the three C2C12 clones express-ing the truncated type II TGF-β receptor, very few (<5%) ofthe cells formed myotubes (Fig. 2A+). Their lack of differen-tiation was not due to delayed onset of differentiation, sincetransfectants maintained for up to 9 days in low serum did notshow significant myotube fusion (Fig. 2B).
These clones are unlikely to be spontaneous, nondifferenti-ating mutants, since the probability of picking a single natural,non-differentiating clone of these cells is less than 10−3 (Lassaret al., 1989). Therefore, the probability that all three clones arenatural, non-differentiating mutants is less than 10−9. Further-more, neither we nor others (Lassar et al., 1989) have detecteda non-differentiating phenotype in control, neomycin-resistantcolonies. Our results therefore suggest that expression of atruncated type II TGF-β receptor inhibits morphological differ-entiation of C2C12 myoblasts.
Induction of muscle cell differentiation markers isinhibited in myoblasts expressing the truncatedTGF-β receptorSince myoblast fusion is not required for the induction
Fig. 1. Binding of 125I-TGF-β to the truncated type II TGF-βreceptor at the cell surface in transfected myoblasts. Parental C2C12cells (lane 1) or clones stably transfected with a truncated type IITGF-β receptor (lanes 2,3) were incubated with 125I-TGF-β, andcross-linked proteins were (A) analyzed by SDS-PAGE or (B)immunoprecipitated with antibodies to an epitope tag in the truncatedtype II TGF-β receptor expression vector, and then separated onSDS-polyacrylamide gels. The third clone gave similar results (notshown). The type I, type II and type III TGF-β receptors areindicated by Roman numerals. DN and arrow indicate the bandcorresponding to the truncated receptor.
1088
of differentiation-specific geneexpression (Endo and Nadal-Ginard,1987; Konieczny et al., 1989; Hu andOlson, 1990), we tested whether ourmyotube fusion-defective cloneswere capable of biochemical differ-entiation following a switch to lowserum medium. Northern hybridiza-tions showed that, as expected, theparental C2C12 cells induced mRNAfor myosin light chains 1/3 andmyosin light chain 2 after a switch todifferentiation media. Theexpression of myosin mRNAs in theparental C2C12 cells in growthmedia is due to spontaneous differ-entiation of a fraction of theconfluent cells, as previouslyobserved (Lassar et al., 1989). Incontrast, when cultured in growthmedia, cells expressing the truncatedreceptor did not have detectablemyosin mRNA levels, and a changeto differentiation media resulted inlittle induction of these mRNAs(Fig. 3). These clones, however,responded to the change in serumconcentration by inducing the αchain of the acetylcholine receptormRNA to levels comparable to theparental C2C12 cells (Fig. 3).
Western analysis showed theexpected induction of troponin T andmyosin heavy chain in the parentalC2C12, cultured in low serum for 3days. In contrast, all three trans-fected clones showed undetectablelevels of these proteins in growthmedia, and had reduced levels oftroponin T, and little, if any, myosinheavy chain expression followingincubation in differentiation media(Fig. 4). The transfected clonescontained significant desminexpression under both conditions.To analyze the expression of theseproteins in individual cells, the cellswere grown in differentiation mediafor 3 days and stained with antibod-ies to troponin T, myosin heavychain and desmin. The parentalC2C12 cells exhibited myotubeswhich stained strongly with antibod-ies to all three proteins. In contrast,cells expressing the truncatedreceptor expressed little if anytroponin T or myosin heavy chain(Fig. 5). Using antibodies to desmin,a low level background, punctuatedby several brightly staining mononu-clear cells, was observed in the
E. H. Filvaroff, R. Ebner and R. Derynck
Fig. 2. Morphology of myoblasts transfected with a truncated TGF-β receptor. Parental C2C12cells (panels 1) or clones stably transfected with a truncated type II TGF-β receptor (panels 2,3)were cultured in growth media (−) or in differentiation media (+) for 3 days (A) or 9 days (B), andwere examined for myotube formation. The third clone showed a morphology similar to the othertwo (not shown).

1089Myogenesis and type II TGF-β receptor
transfected clones (Fig. 5). Thus, C2C12 myoblasts express-ing the truncated receptor do not exhibit normal morphologi-cal or biochemical differentiation. However, not all differen-tiation markers are equally affected by expression of thetruncated receptor.
Regulated expression of MyoD and myogenin, butnot Id, is blocked in cells expressing the dominantnegative receptor mutantProliferating myoblasts express the protein Id, an inhibitor ofdifferentiation (Benezra et al., 1990). In high serum media, Idprevents MyoD activation of downstream genes by competingwith MyoD for binding to the E12 protein (Murre et al.,1989a,b). Upon a switch to low serum media, Id expression isgreatly decreased in cultured myoblasts (Benezra et al., 1990).Since down-regulation of Id expression is required for normalmyoblast differentation (Weintraub et al., 1991), we comparedthe level of Id mRNA in the parental and transfected C2C12cells. As expected, exposure of C2C12 cells to low serumdecreased Id mRNA levels (Fig. 3). Surprisingly, cells express-ing the truncated receptor had a similar downregulation of IdmRNA levels when switched to low serum, suggesting that theregulation of this gene is not affected by expression of thetruncated TGF-β receptor. This result agrees with studiesshowing that TGF-β does not affect Id expression (Brennan etal., 1991). Furthermore, the inability of the transfected clonesto differentiate cannot be explained by a failure of Id to bedownregulated in low serum.
MyoD and myogenin are expressed only in skeletal musclecells, and their expression is upregulated during myogenicdifferentiation in vitro (Davis et al., 1987; Edmondson andOlson, 1989; Wright et al., 1989). In addition, expression oftransfected cDNAs for these transcription factors convertsvarious cell types to a muscle phenotype (Davis et al., 1987;Weintraub et al., 1989). Thus, MyoD and myogenin arebelieved to be involved in the commitment of cells to themyogenic lineage and in the establishment and maintenance ofthe differentiated phenotype (Weintraub et al., 1991). Toascertain whether the truncated TGF-β receptor affected thedetermination of cell identity, we examined the expression ofMyoD and myogenin. As expected, C2C12 cells induced tran-scripts for both MyoD and myogenin after a switch to differ-entiation media. Expression of myogenin in these cells ingrowth media is most likely due to spontaneous differentiationwhich occurs at high cell density (Lassar et al., 1989). Unlikethe parental C2C12, the cells expressing the truncated receptorshowed no MyoD or myogenin expression under growing con-ditions, and exhibited little induction of these transcripts indifferentiation media (Fig. 3). The failure of these cells to dif-ferentiate may be due to inadequate levels of MyoD andmyogenin, and raises the possibility that these clones are nolonger committed to the myogenic lineage.
Expression of the truncated TGF-β receptor doesnot prevent growth arrest or cause transformationThe inability of the transfected cells to differentiate could, intheory, be due to an increase in cell growth, since proliferationand differentiation are assumed to be mutually exclusive inmyoblasts (Olson, 1992). Furthermore, the differentation-resistant phenotype of myoblasts, created by introduction andexpression of some oncogenes, such as v-myc (Falcone et al.,1985), v-ras (Olson et al., 1987; Konieczny et al., 1989; Lassaret al., 1989) and v-fos (Lassar et al., 1989), or the Hox 7.1homeobox gene (Song et al., 1992), is associated with cellulartransformation. Thus, we evaluated the proliferation rate andtransformation state of the transfected clones.
Fig. 3. Expression of muscle-specific mRNAs in myoblastsexpressing the truncated TGF-β receptor. RNA was extracted fromparental C2C12 cells (lanes 1) or cells transfected with a truncatedtype II TGF-β receptor (lanes 2,3,4) cultured in growth media (−) orin differentiation media (+) for 3 days, and analyzed by northernblotting using probes for myosin light chain 2 (MLC2), myosin lightchain 1/3 (MLC 1/3), the α chain of the acetylcholine receptor (Ach-R), myogenin, MyoD or Id. Equal loading was ascertained byethidium bromide staining of the 28S and 18S rRNA bands and by acontrol hybridization using the cDNA for the constitutivelyexpressed glyceraldehyde-phosphate dehydrogenase (not shown).
Fig. 4. Expression ofmuscle-specific proteinsin myoblasts transfectedwith the truncated TGF-β receptor. Equalquantities of proteinextracts, prepared fromparental C2C12 cells(lanes 1) or the stablytransfected clones (lanes2,3,4) cultured ingrowth (−) ordifferentiation (+)
media for 3 days, were analyzed by western blotting using antibodiesto troponin T (TnT), myosin heavy chain (MHC), or desmin.

1090
Neither our transfected cells nor the parental cells displayedanchorage-independent growth in a soft agar colony formationassay unlike our positive control, Cos-1 cells (data not shown).Therefore, inhibition of TGF-β responsiveness through thetype II receptor does not induce transformation. Furthermore,the proliferation rate of the cells expressing the truncatedreceptor was significantly slower than of the parental C2C12cells (Fig. 6). Thus, the inability of the transfected cells to dif-ferentiate into myotubes can not be explained by increased oruncontrolled growth. In fact, a decreased proliferation ratemight be considered an advantage for differentiation.
Another possible way in which expression of the truncatedTGF-β receptor could block differentiation would be to preventthe growth arrest that normally occurs in myoblasts in lowserum media (Olson, 1992). The proliferation rate before andafter a switch to low serum media indicated that the transfectedcells ceased proliferation similarly to the parental C2C12 cells(Fig. 6). Thus, expression of the truncated type II TGF-βreceptor does not prevent growth arrest of the cells in differ-entiation media.
Phosphorylation of the growth suppressor gene product pRBhas been used as a cell cycle marker, since this protein becomesphosphorylated at the G1/S boundary and dephosphorylatedduring G2/M phase (reviewed in Marshall, 1991; Weinberg,
E. H. Filvaroff, R. Ebner and R. Derynck
Fig
. 5.I
mm
unofl
uore
scen
ce a
naly
sis
of e
xpre
ssio
n of
mus
cle
mar
kers
in th
e tr
ansf
ecte
dcl
ones
. Par
enta
l C2C
12 c
ells
(to
p pa
nels
) an
d on
e of
the
clon
es tr
ansf
ecte
d w
ith th
etr
unca
ted
TG
F-β
rece
ptor
(bo
ttom
pan
els)
wer
e in
cuba
ted
with
no
prim
ary
antib
ody
(−)
or
antib
odie
s to
des
min
, tro
poni
n T
(T
nT),
or
myo
sin
heav
y ch
ain
(MH
C)
follo
wed
by
rhod
amin
e-co
njug
ated
sec
onda
ry a
ntib
ody
and
phot
ogra
phed
by
phas
e-co
ntra
st a
ndep
ifluo
resc
ence
mic
rosc
opy.
All
thre
e cl
ones
beh
aved
sim
ilarl
y (n
ot s
how
n).
MH
CT
nT
Des
min
−
1 2
Fig. 6. Proliferation and growth arrest of myoblasts expressing thetruncated TGF-β receptor. (A) C2C12 cells (closed circles) or thetransfected clones (open symbols) were plated on day 0, and counted3 days later, when they were either maintained in growth media(Normal Serum) or were switched to differentiation media (LowSerum). Cells were counted every other day, up to a week after theinitial plating. (B) Protein extracts, prepared from parental C2C12cells (lanes 1) or the transfected clones (lanes 2,3,4) cultured ingrowth (−) or differentiation (+) media for 3 days, were analyzed bywestern blotting using antibodies to the retinoblastoma protein (Rb).

1091Myogenesis and type II TGF-β receptor
1991). Thus, in growth-arrested myoblasts, only the unphos-phorylated form of pRB can be detected (Gu et al., 1993). Inaddition, pRB has been implicated in the antiproliferativeeffect of TGF-β (Laiho et al., 1990b; Moses et al., 1990) andplays a key role in myogenic differentiation by interacting with
MyoD (Gu et al., 1993). We therefore examined the phospho-rylation state of pRB under growing or differentiating con-ditions. Withdrawal of high serum shifted pRB to its fastermigrating, dephosphorylated form in the parental C2C12 cells,as previously observed (Gu et al., 1993). A similar shift in pRB
Fig. 7. Behavior of transfected myoblasts in vivo. Cross sections of mouse hind limbs 14 days after injection with β-galactosidase-expressinguntransfected C2C12 myoblasts (panel 1) or myoblasts expressing the truncated TGF-β type II receptor (panels 2,3) were stained for β-galactosidase activity (blue), mounted and photographed. The third clone gave results similar to those shown in panel 2.
1 2 3
Fig. 8. Phenotype oftransfected myoblastsexpressing thetruncated type IIreceptor which havebeen infected with aMyoD-expressingretrovirus. The threeclones of myoblaststransfected with thetruncated TGF-βreceptor (panels 1,2,3)were infected with ahigh titer, MyoD-expressing retrovirus.These cells weretransferred todifferentiation mediumfor 3 days andanalyzed for (a)myotube formationand (b) expression ofmyosin heavy chain byimmunofluorescence.
b
a
1 2 3

1092
migration was observed in the transfected clones (Fig. 6).Thus, expression of the truncated TGF-β receptor does notinhibit the change in pRB phosphorylation induced by lowserum in C2C12 cells, nor, by extrapolation, their withdrawalfrom the cell cycle and subsequent growth arrest.
Myoblasts expressing the truncated type II receptordo not form myofibers in vivoWhen injected into skeletal muscle, C2C12 myoblasts incor-porate into and fuse to form myofibers (Barr and Leiden,1991; Dhawan et al., 1991). To evaluate the effect ofexpression of the truncated receptor on myoblast differen-tiation in vivo, we injected the transfected and controlmyoblasts into the skeletal muscle of hind limbs of nude mice.Prior to injection, the cells were infected with a retrovirusexpressing β-galactosidase and hygromycin-resistance.Following selection in hygromycin, more than 90% of thecells expressed β-galactosidase, as assessed by histochemicalstaining. Furthermore, retroviral infection and β-galactosidaseexpression did not affect myotube formation of the parentalC2C12 cells in vitro, nor did myotube formation decrease β-galactosidase expression (data not shown). 2 weeks afterinjection, mouse limbs were serially sectioned and analyzedfor β-galactosidase expression by histochemical staining withX-gal. Sections from muscle tissue injected with the parentalC2C12 cells showed many β-galactosidase-expressing musclefibers, which resembled in morphology and size the sur-rounding muscle tissue (Fig. 7). Thus, as previously shown(Barr and Leiden, 1991; Dhawan et al., 1991), the parentalC2C12 cells fused and incorporated into the myofibers. Incontrast, tissue injected with two of the transfected cell linescontained few β-galactosidase-positive cells, and those cellsappeared primarily between the endogenous muscle fibers.Similarly, fibroblasts injected into muscle do not incorporateinto myofibers and gradually lose their ability to proliferate(G. Pavlath and H. Blau, personal communication). Tissueinjected with the third clone showed a significant number ofβ-galactosidase-positive cells adjacent to, but generally notwithin, muscle fibers. The vast majority of these cells wereunicellular and were part of a disorganized tissue (Fig. 7).Thus in contrast to the parental cells, cells expressing thetruncated receptor did not fuse into endogenous muscle tissue.
The viability of the cells in vivo was verified by trypan bluestaining, showing that the transfected and parental cells werenot differentially sensitive to the preinjection conditions. Fur-thermore, injected limbs sectioned one day after injection ofthe clones showed many β-galactosidase-positive cells, sug-gesting similar early survival and β-galactosidase expressionof the parental and transfected cells in vivo (data not shown).Thus, our results suggest that the in vivo environment is unableto overcome the block in differentiation observed in the trans-fected cells in vitro. Furthermore, the inability of the clones tomature in vivo suggests that their in vitro differences from theparental C2C12 cells have physiological relevance.
Retroviral expression of MyoD rescues the ability ofcells expressing the truncated receptor todifferentiateTo determine whether downregulation of MyoD was requiredfor the inhibition of myoblast differentiation, the transfectedcells expressing the truncated TGF-β receptor were infected
with a high titer MyoD-expressing retrovirus and tested intransient assays for their ability to differentiate. Northernanalysis showed that exogenous MyoD was expressed in theinfected cell population (data not shown). Based on the ratioof G418-resistant to G418-sensitive C2C12 cells, we estimatean infection efficiency of 50-70%. Of those cells expressingMyoD, we would expect approximately 50% to undergomyogenic differentiation (Davis et al., 1987; Koniezcny et al.,1989). Accordingly, 10-50% of the transfected cells infectedwith the MyoD retrovirus formed myotubes when cultured inhigh or low serum media (Fig. 8A). Furthermore, cells infectedwith the MyoD virus showed a similar frequency of myosinheavy chain-expressing cells, as visualized by immunofluores-cence (Fig. 8B). Thus, forced expression of MyoD allows cellsexpressing the truncated receptor to differentiate, suggestingthat downregulation of MyoD is essential for inhibition ofmyogenic differentiation by the truncated TGF-β receptor.
DISCUSSION
We have examined the function of TGF-β and its type IIreceptor-mediated signaling system during the growth anddifferentiation of myoblasts using C2C12 cells as a modelsystem. Our approach was to generate cells expressing atruncated type II receptor which inhibits signaling in adominant negative fashion (Brand et al., 1993; Chen et al.,1993). The resulting changes in the ability of myoblasts to dif-ferentiate in vitro and in vivo led to several conclusions.
Responsiveness to TGF-β, endogenously synthesized by thecells or in the media, has positive effects on the growth anddifferentiation of myoblasts. TGF-β may assist in myoblastproliferation as suggested by the stimulatory effect of TGF-βon C2C12 proliferation (data not shown) and the considerablyslower growth rate of cells expressing the truncated type IITGF-β receptor. These transfected cells do not differentiatewhen cultured in low serum, as evidenced by their lack offusion and inability to induce expression of several myoblastdifferentiation markers. In contrast, control transfected C2C12cells differentiated normally. The C2C12 cells expressing thetruncated receptor were still differentiation competent, since,as the parental cells, they formed myotubes followingtreatment with dexamethasone (data not shown). Thus, TGF-βresponsiveness appears to be required for muscle-specific genetranscription and fusion during low serum-induced myogenicdifferentiation. Interestingly, attempts to create myoblastsoverexpressing the full-length type II TGF-β receptor wereunsuccessful. This would be expected if type II TGF-β receptoroverexpression favored myotube formation, just as functionalinactivation of the endogenous receptors blocked differen-tiation.
The growth arrest and change in phosphorylation of pRB,which occurred in the transfected cells after a switch to lowserum, indicate that the inhibition of differentiation cannot beexplained by an inability to exit the cell cycle. Transfection ofmyoblasts with v-myc (Falcone et al., 1985), v-ras (Olson et al.,1987; Konieczny et al., 1989; Lassar et al., 1989), v-fos (Lassaret al., 1989) or Hox 7.1 (Song et al., 1992), which block lowserum-induced differentiation, also results in cellular transfor-mation. In contrast, cells expressing the truncated receptor werenot transformed, as determined by a soft agar assay.
E. H. Filvaroff, R. Ebner and R. Derynck

1093Myogenesis and type II TGF-β receptor
The parental C2C12 cells express both MyoD andmyogenin, and these proteins stimulate expression of them-selves as well as of each other (Thayer et al., 1989; Edmondsonet al., 1991). In contrast, C2C12 cells transfected with thetruncated type II receptor no longer express MyoD andmyogenin suggesting an involvement of TGF-β in the mainte-nance of their expression, perhaps by affecting their positiveautoregulatory loops. Since MyoD and myogenin induce aprogram leading to myogenic differentiation (Weintraub et al.,1991), their suppression in the transfected cells may be thebasis of the block in differentiation. This hypothesis issupported by our finding that infection of our transfectedC2C12 cells with a MyoD retrovirus permits expression ofmyosin heavy chain and myotube formation, as observed in v-ras or v-fos transformed C2C12 cells (Lassar et al., 1989). Thissimilarity is intriguing since c-ras (Mulder and Morris, 1992)and c-fos (Kim et al., 1990; Matrisian et al., 1992) may be partof the signaling pathways induced by TGF-β.
The effect of TGF-β on differentiation could depend on thepresence of other growth factors (Allen and Boxhorn, 1989;Zentella and Massagué, 1992) and the immediate cellular envi-ronment. However, the inhibition of myogenic differentiationalso occurred in vivo, in the context of the muscle tissue fromwhich C2C12 cells were derived (Yaffe and Saxel, 1977).Thus, the in vivo environment could not overcome the blockin differentiation, and the inhibitory effect of the truncatedreceptor appears to predominate. Furthermore, these in vivoresults indicate that the inhibition of differentiation observedin vitro may have physiological relevance.
The dominant negative mutant of the TGF-β receptor doesnot block all aspects of differentiation, suggesting that at leasttwo pathways are involved in muscle cell differentiation invitro. One pathway regulates the induction of MyoD,myogenin, myosin heavy chain, myosin light chain andtroponin T. This pathway is inhibited in the transfected clonesand, thus, requires signaling through the type II TGF-βreceptor. The other pathway(s) involves basal desminexpression and other low-serum-induced changes — such asinduction of the α chain of the acetylcholine receptor, down-regulation of Id, hypophosphorylation of pRB and growtharrest — and is not affected by inhibition of type II receptorsignaling. Induction of the latter pathway in the transfectedcells is not sufficient to cause full myogenic differentiation.Similarly, v-ras transformed myoblasts induce expression ofthe acetylcholine receptor and maintain desmin expression butare blocked in expression of other markers including MyoD(Lassar et al., 1989). Thus, one pathway may be independentof MyoD expression, as cells expressing desmin and otherspecific contractile proteins, but not MyoD, have beendescribed (Kaufman et al., 1991; Peterson et al., 1990; Cusella-De Angelis et al., 1992). Our results also imply that myoblastdifferentiation is a multistep process. Our transfected cells,which undergo growth arrest but do not differentiate, mayallow dissection of the pathways involved in myoblast differ-entiation.
The seemingly conflicting reports on the inhibitory and stim-ulatory effects of TGF-β in muscle formation could beexplained in several ways. One possibility is that the effect ofTGF-β depends on the intrinsic, developmental state of thecell. In this scenario, TGF-β could help maintain MyoDexpression in immature myoblasts, thus providing a state of
competence for differentiation. Further differentiation wouldthen be prevented by the inhibitory effect of TGF-β until anappropriate signal to differentiate occurred, or until cells havemigrated away from areas of TGF-β production. Thus, TGF-βcould maintain a proliferating population of committed cells,poised to become myotubes, while also preventing prematuremyoblast differentiation until sufficient muscle mass exists(Olson et al., 1986; Massagué et al., 1986). Interestingly, theexpression pattern of TGF-β during mouse muscle develop-ment shows early expression in somites at day 11 (Heine et al.,1987), which is consistent with the above model.
Another possibility is that the two effects of TGF-β, stimu-lation versus inhibition of myoblast differentiation, aremediated by two different receptors. We have used a truncatedtype II receptor, which specifically inhibits signaling by thetype II, but not the type I, TGF-β receptor in a dominantnegative fashion (Chen et al., 1993). Based on that study inepithelial cells, the type II receptor is believed to be requiredfor the growth inhibition induced by TGF-β, and the type Ireceptor may be involved in TGF-β induction of the extracel-lular matrix (Chen et al., 1993). Since collagen (Heino andMassagué, 1990), and c-jun (Bengal et al., 1992) inhibit myo-genesis, signaling through the type I receptor could indirectlyprevent myoblast differentiation by inducing expression ofthese inhibitory factors. In contrast, since unphosphorylatedpRB may play a critical role in myogenesis by interacting withMyoD (Gu et al., 1993), signaling through the type II receptorcould have a positive effect on skeletal muscle cell differen-tiation. Thus, the pleiotropic effects of TGF-β on myoblastsmight be mediated by at least two pathways involving twoTGF-β receptor types. Under normal circumstances, a balancemay exist between the negative and positive signaling throughthe type I and type II receptors. Introduction of a truncated typeII receptor into C2C12 cells may upset this balance, such thatthe negative effect of signaling through the type I receptor pre-dominates. C2C12 cells in culture produce all three TGF-βisoforms (Lafyatis et al., 1991). According to this model, theinability of cells expressing the truncated receptor to differen-tiate could be due to autocrine stimulation of the type I TGF-β receptor.
The possibility that TGF-β can have mutually distinct effectsthat result in the synchronization of cells in a specific devel-opmental stage is an intriguing one. Furthermore, the effect ofTGF-β on a cell may depend as much on the balance ofexpression of specific receptors within the cell, as on the extra-cellular concentration of the ligand.
We thank Dr Charles Ordahl and the members of his lab forvaluable reagents and advice, Drs Helen Blau and Andrew Lassar forcritical reading of the manuscript, Drs Nadia Rosenthal, HaroldWeintraub, Harold Varmus and Woody Wright for plasmids, DrDusty Miller for the high titer MyoD retrovirus, and Dr Grace Pavlathfor teaching us the in vivo experiments. We are also grateful to AdrianErlebacher and Dr Ruey-Hwa Chen for helpful suggestions andsupport. This work was supported by an NIH postdoctoral traininggrant to E. H. F., by a grant from the NIH to R. D. and by the MarkeyCharitable Trust to the Programs in Biological Sciences.
REFERENCES
Allen, R. E. and Boxhorn, L. K. (1989). Regulation of skeletal muscle satellite

1094
cell proliferation and differentiation by transforming growth factor-beta,insulin-like growth factor-I, and fibroblast growth factor. J. Cell. Physiol.138, 311-15.
Bassing, C. H., Yingling, J. M., Howe, D. J., Wang, T., He, W. W.,Gustafson, M. L., Shah, P., Donahoe, P. K. and Wang, X.-F. (1994). Atransforming growth factor-β type I receptor that signals to activate geneexpression. Science 263, 87-89.
Barr, E. and Leiden, J. M. (1991). Systemic delivery of recombinant proteinsby genetically modified myoblasts. Science 254, 1507-1509.
Benezra, R., Davis, R. L., Lockshon, D., Turner, D. L. and Weintraub, H.(1990). The protein Id: a negative regulator of helix-loop-helix DNA bindingproteins. Cell 61, 49-59.
Bengal, E., Ransone, L., Scharfmann, R., Dwarki, V. J., Tapscott, S. J.,Weintraub, H. and Verma, I. M. (1992). Functional antagonism between c-jun and MyoD proteins: a direct physical association. Cell 68, 507-519.
Blau, H. M., Chiu, C-P. and Webster, C. (1983). Cytoplasmic activation ofhuman nuclear genes in stable heterocaryons. Cell 32, 1171-1180.
Brand, T., MacLellan, R. and Schneider, M. D. (1993). A dominant-negativereceptor for type β transforming growth factors created by deletion of thekinase domain. J. Biol. Chem. 268, 11500-11503.
Brennan, T. J., Edmondson, D. G., Li, L. and Olson, E. N. (1991).Transforming growth factor β represses the actions of myogenin through amechanism independent of DNA binding. Proc. Natl. Acad. Sci. USA 88,3822-3826.
Chen, R-H., Ebner, R. and Derynck, R. (1993). Inactivation of the type IIreceptor reveals two receptor pathways for the diverse TGF-β activities.Science 260, 1335-1338.
Chomczynski, P. and Sacchi, N. (1987). Single-step method of RNA isolationby acid guanidium thiocyanate-phenol-chloroform extraction. Anal.Biochem. 162, 156-159.
Cusella-De Angelis, M. G., Lyons, G., De Angelis, L., Vivarelli, E., Farmer,K., Wright, W. E., Molinaro, M., Bouche, M., Buckingham, M. andCossu, G. (1992). MyoD, myogenin independent differentiation ofprimordial myoblasts in mouse somites. J. Cell Biol. 116, 1243-1255.
Davis, R. L., Weintraub, H. and Lassar, A. (1987). Expression of a singletransfected cDNA converts fibroblasts to myoblasts. Cell 51, 987-1000.
DeCaprio, J. A., Ludlow, J. W., Figge, J., Shew, J-Y., Huang, C-M., Lee,W-H., Marsilio, E., Paucha, E. and Linvingston, D. M. (1988). SV40 largetumor antigen forms a specific complex with the product of theretinoblastoma susceptibility gene. Cell 54, 275-283.
Dhawan, J., Pan, L. C., Pavlath, G. K., Travis, M. A., Lanctot, A. M. andBlau, H. M. (1991). Systemic delivery of human growth hormone byinjection of genetically engineered myoblasts. Science 254, 1509-1512.
Ebner, R., Chen, R-H., Shum, L., Lawler, S., Zioncheck, T. F., Lee, A.,Lopez, A. R. and Derynck, R. (1993a). Cloning of a type I TGF-β receptorand its effect on TGF-β binding to the type II receptor. Science 260, 1344-1348.
Ebner, R., Chen, R-H., Lawler, S., Zioncheck, T. and Derynck, R. (1993b).The type II receptors for TGF-β or activin determine the ligand specificity ofa single type I receptor. Science 262, 900-902.
Edmondson, D. G. and Olson, E. N. (1989). A gene with homology to the mycsimilarity region of MyoD1 is expressed during myogenesis and is sufficientto activate the muscle differentiation program. Genes Dev. 3, 628-640.
Edmondson, D. G., Brennan, T. J. and Olson, E. N. (1991). Mitogenicrepression of myogenin autoregulation. J. Biol. Chem. 266, 21343-21346.
Endo, T. and Nadal-Ginard, B. (1987). Three types of muscle-specific geneexpression in fusion-blocked rat skeletal muscle cells: translational control inEGTA-treated cells. Cell 49, 515-526.
Falcone, F., Tato, F. and Alema, S. (1985). Distinctive effects of the viraloncogenes myc, erb, fps, and src on the differentiation program of quailmyogenic cells. Proc. Natl. Acad. Sci. USA 82, 426-430.
Franzén, P., ten Dijke, P., Ichijo, H., Yamashita, H., Schulz, P., Heldin, C.-H. and Miyazono, K. (1993). Cloning of a TGF-β type I receptor that formsa heteromeric complex with the TGF-β type II receptor. Cell 75, 681-692.
Frolik, C. A., Wakefield, L. M., Smith, D. M. and Sporn, M. B. (1984).Characterziation of a membrane receptor for transforming growth factor-betain normal kidney fibroblasts. J. Biol. Chem. 259, 10995-11000.
Gazit, D., Ebner, R., Kahn, A. J. and Derynck, R. (1993). Modulation ofexpression and cell surface binding of members of the transforming growthfactor-β superfamily during retinoic acid-induced osteoblasticdifferentiation of multipotential mesenchymal cells. Mol. Endocrinol. 7,189-198.
Geiser, A. G., Burmester, J. K., Webbink, R., Roberts, A. B. and Sporn, M.B. (1992). Inhibition of growth by transforming growth factor-β following
fusion of two nonresponsive human carcinoma cell lines. J. Biol. Chem. 267,2588-2593.
Greenwald, I. and Rubin, G. M. (1992). Making a difference: the role of cell-cell interactions in establishing separate identities for equivalent cells. Cell58, 271-281.
Gu, W., Schneider, J. W., Condorelli, G., Kaushal, S., Mahdavi, V. andNadal-Ginard, B. (1993). Interaction of myogenic factors and theretinoblastoma protein mediates muscle cell commitment anddifferentiation. Cell 72, 309-324.
Gurdon, J. B. (1992). The generation of diversity and pattern in animaldevelopment. Cell 68, 185-199.
Heine, U. I., Munoz, E. F., Flanders, K. C., Ellingsworth, L. R., Lam, H-Y.P., Thompson, N. L., Roberts, A. B. and Sporn, M. B. (1987). Role oftransforming growth factor-β in the development of the mouse embryo. J.Cell Biol. 105, 2861-2876.
Heino, J. and Massagué, J. (1990). Cell adhesion to collagen and decreasedmyogenic gene expression implicated in the control of myogenesis bytransforming growth factor β. J. Biol. Chem. 265, 10181-10184,
Hu, J. S. and Olson, E. N. (1990). Functional receptors for transforminggrowth factor-beta are retained by biochemically differentiated C2 myocytesin growth factor-deficient medium containing EGTA but down-regulatedduring terminal differentiation. J. Biol. Chem. 265, 7914-9.
Ignotz, R. A. and Massagué, J. (1985). Type β transforming growth factorcontrols the adipogenic differentiation of 3T3 fibroblasts. Proc. Natl. Acad.Sci. USA 82, 8530-8534.
Isenberg, K. E., Mudd, J., Shah, V. and Merlie, J. P. (1986). Nucleotidesequence of the mouse acetylcholine receptor α subunit. Nucleic Acids Res.14, 5111.
Jessell, T. M. and Melton, D. A. (1992). Diffusible factors in vertebrateembryonic induction. Cell 68, 257-270.
Kaufman, S. J., George-Weinstein, M. and Foster, R. F. (1991). In vitrodevelopment of precursor cells in the myogenic lineage. Dev. Biol. 146, 228-238.
Kim, S-J., Angel, P., Lafyatis, R., Hattori, K., Kim, K. Y., Sporn, M. B.,Karin, M. and Roberts, A. B. (1990). Autoinduction of TGF-β1 is mediatedby the AP-1 complex. Mol. Cell. Biol. 10, 1492-97.
Konieczny, S. F., Drobes, B. L., Menke, S. L. and Taparowsky, E. J. (1989).Inhibition of myogenic differentiation by the H-ras oncogene is associatedwith the down regulation of the MyoD1 gene. Oncogene 4, 4773-481.
Lafyatis, R., Lechleider, R., Roberts, A. B. and Sporn, M. B. (1991).Secretion and transcriptional regulation of transforming growth factor-β3during myogenesis. Mol. Cell. Biol. 11, 3795-3803.
Laiho, M., Weis, F. M. B. and Massagué, J. (1990a). Concomitant loss oftransforming growth factor receptor types I and II in TGF-β resistant cellmutants implicates both receptor types in signal transduction. J. Biol. Chem.265, 18518-18524.
Laiho, M., DeCaprio, J. A., Ludlow, J. W., Livingston, D. M. andMassagué, J. (1990b). Growth inhibition by TGF-β linked to suppression ofretinoblastoma protein phosphorylation. Cell 62, 175-185.
Laiho, M., Weis, F. M. B., Boyd, F., Ignotz, R. A. and Massagué, J. (1991).Responsiveness to transforming growth factor-β restored by geneticcomplementation between cells defective in TGF-β receptors I and II. J. Biol.Chem. 266, 9108-9112.
Lassar, A. B., Thayer, M. J., Overell, R. W. and Weintraub, H. (1989).Transformation by activated ras or fos prevents myogenesis by inhibitingexpression of MyoD1. Cell 58, 659-667.
Lawler, S., Candia, A. F., Ebner, R., Lopez, A. R., Moses, H. L., Wright, C.V. E. and Derynck, R. (1994). The murine type II TGF-β receptor has acoincident embryonic expression and binding preference for TGF-β1.Development, in press.
Lefer, A. M., Tsao, P., Aoki, N. and Palladino, M. A., Jr. (1990). Mediationof cardioprotection by transforming growth factor-beta. Science 249, 61-64.
Lin, H. Y., Wang, X-F., Ng-Eaton, E., Weinberg, R. A. and Lodish, H. F.(1992). Expression cloning of the TGF-β type II receptor, a functionaltransmembrane serine/threonine kinase. Cell 68, 775-785.
Marshall, C. J. (1991). Tumor suppressor genes. Cell 64, 313-326. Massagué, J., Cheifetz, S., Endo, T. and Nadal-Ginard, B. (1986). Type β
transforming growth factor is an inhibitor of myogenic differentiation. Proc.Natl. Acad. Sci. USA 83, 8206-8210.
Massagué, J. (1992). Receptors for the TGF-β family. Cell 69, 1067-1070. Matrisian, L. M., Ganser, G. L., Kerr, L. D., Pelton, R. W. and Wood, L. D.
(1992). Negative regulation of gene expression by TGF-β. Mol. Reprod.Develop. 32, 111-120.
Millan, F. A., Denhez, F., Kondaiah, P. and Akhurst, R. J. (1991).
E. H. Filvaroff, R. Ebner and R. Derynck

1095Myogenesis and type II TGF-β receptor
Embryonic gene expression of TGF β1, β2, and β3 suggest differentdevelopmental functions in vivo. Development 111, 131-144.
Miller, A. D., Miller, D. G., Garcia, J. V. and Lynch, C. M. (1993). Use ofretroviral vectors for gene transfer and expression. Methods in Enzym. 217,581-99.
Moses, H. L., Yang, E. Y. and Pietenpol, J. A. (1990). TGF-β stimulation andinhibition of cell proliferation: new mechanistic insights. Cell 63, 245-247.
Mulder, K. M. and Morris, S. L. (1992). Activation of p21ras by transforminggrowth factor β in epithelial cells. J. Biol Chem. 267, 5029-5031.
Murre, C., McCaw, P. S. and Baltimore, D. (1989a). A new DNA bindingand dimerization motif in immunoglobulin enhancer binding, daughterless,MyoD, and myc proteins. Cell 56, 777-783.
Murre, C., McCaw, P. S., Vaessin, H., Caudy, M., Jan, L. Y., Jan, Y. N.,Cabrera, C. V., Buskin, J. N., Hauschka, S. D., Lassar, A. B., Weintraub,H. and Baltimore, D. (1989b). Interactions between heterologous helix-loop-helix proteins generate complexes that bind specifically to a commonDNA sequence. Cell 58, 537-544.
Olson, E. N., Sternberg, E., Hu, J. S., Spizz, G. and Wilcox, C. (1986).Regulation of myogenic differentiation by type β transforming growthfactor. J. Cell. Biol. 103, 1799-1805.
Olson, E. N., Spizz, G. and Tainsky, M. A. (1987). The oncogenic forms of N-ras or H-ras prevent skeletal myoblast differentiation. J. Cell. Biol. 7, 2104-2111.
Olson, E. N. (1992). Interplay between proliferation and differentiation withinthe myogenic lineage. Dev. Biol. 154, 261-272.
Pelton, R. W., Saxena, B., Jones, M., Moses, H. L. and Gold, L. I. (1991).Immunohistochemical localization of TGFβ1, TGFβ2, and TGF-β3 in themouse embryo: expression of patterns suggest multiple roles duringembryonic development. J. Cell. Biol. 115, 1091-1105.
Periasamy, M., Strehler, E. E., Garfinkel, L. I., Gubits, R. M., Ruiz-Opazo,N. and Nadal-Ginard, B. (1984). Fast skeletal muscle myosin light chains 1and 3 are produced from a single gene by a combined process of differentialRNA transcription and splicing. J. Biol. Chem. 259, 11395-13604.
Peterson, C. A., Gordon, H., Hall, Z. W., Paterson, B. M. and Blau, H. M.(1990). Negative control of the helix-loop-helix family of myogenicregulators in the NFB mutant. Cell 62, 493-502.
Potts, J. D. and Runyan, R. B. (1989). Epithelial-mesenchymal celltransformation in the embryonic heart can be mediated, in part, bytransforming growth factor β. Dev. Biol. 134, 392-401.
Potts, J. D., Dagle, J. M., Walder, J. A., Weeks, D. L. and Runyan, R.(1991). Epithelial-mesenchymal transformation of embryonic cardiacendothelial cells is inhibited by a modified antisense oligodeoxynucleotide totransforming growth factor b3. Proc. Natl. Acad. Sci. USA 88, 1516-1520.
Rosen, D., Stempien, S. A., Thompson, A. Y. and Seyedin, S. M. (1988).Transforming growth factor β modulates the expression of osetoblast andchondroblast phenotypes in vitro. J. Cell. Physiol. 134, 337-346.
Sambrook, J. (1989). Molecular cloning; a laboratory manual. (ed. J.
Sambrook, E. F. Fritsch, and T. Maniatis). 2nd ed. Cold Spring Harbor, N. Y.: Cold Spring Harbor Laboratory.
Slager, H. G., van Inzen, W., Freund, E., van den Eijnden-van Raaij, A. J.M. and Mummery, C. L. (1993). Transforming growth factor-β in the earlymouse embryo: implications for the regulation of muscle formation andimplantation. Dev. Gen. 14, 212-224.
Song, K., Wang, Y. and Sassoon, D. (1992). Expression of Hox-7. 1 inmyoblasts inhibits terminal differentiation and induces cell transformation.Nature 360, 477-481.
Thayer, M. J., Tapscott, S. J., Davis, R. L., Wright, W. E., Lassar, A. B. andWeintraub, H. (1989). Positive autoregulation of the myogenicdetermination gene MyoD1. Cell 58, 241-248.
Thompson, N. L., Bazoberry, F., Speir, E. H., Casscells, W., Ferrans, V. J.,Flanders, K. C., Kondaiah, P., Geiser, A. G. and Sporn, M. B. (1988).Transforming growth factor beta-1 in acute myocardial infarction in rats.Growth Factors 1, 91-99.
Torti, F. M., Torti, S. V., Larrick, J. W. and Ringold, G. M. (1989).Modulation of adipocyte differentiation by tumor necrosis factor andtransforming growth factor beta. J. Cell Biol. 108, 1105-1113.
Vaidya, T. B., Rhodes, S. J., Taparowsky, E. J. and Konieczny, S. F. (1989).Fibroblast growth factor and transforming growth factor β represstranscription of the myogenic regulatory gene MyoD1. Mol. Cell. Biol. 9,3576-3579.
Weinberg, R. A. (1991). Tumor suppressor genes. Science 254, 1138-1146. Weintraub, H., Tapscott, S. J., Davis, R. L., Thayer, M. J., Adam, M. A.,
Lassar, A. B. and Miller, A. D. (1989). Activation of muscle-specific genesin pigment, nerve, fat, liver, and fibroblast cell lines by forced expression ofMyoD. Proc. Natl. Acad. Sci. USA 86, 5434-5438.
Weintraub, H., Davis, R., Tapscott, S., Thayer, M., Krause, M., Benezra,R., Blackwell, T. K., Turner, D., Rupp, R., Hollenberg, S., Zhuang, Y.and Lassar, A. (1991). The myoD gene family: nodal point duringspecification of the muscle cell lineage. Science 251, 761-766.
Wrana, J. L., Attisano, L., Carcamo, J., Zentella, A., Doody, J., Laiho, M.,Wang, X-F. and Massagué, J. (1992). TGF-β signals through a heteromericprotein kinase receptor complex. Cell 71, 1003-1014.
Wright, W. E., Sassoon, D. A. and Lin, V. K. (1989). Myogenin, a factorregulating myogenesis, has a domain homologous to MyoD. Cell 56, 607-617.
Yaffe, D. and Saxel, O. (1977). Serial passaging and differentiation ofmyogenic cells isolated from dystrophic mouse muscle. Nature 270, 725-727.
Zentella, A. and Massagué, J. (1992). Transforming growth factor β inducesmyoblast differentiation in the presence of mitogens. Proc. Natl. Acad. Sci.USA 89, 5176-5180.
(Accepted 31 January 1994)