interaction of chloroplasts with inhibitors - plant physiology
TRANSCRIPT

Plant Physiol. (1977) 59, 724-732
Interaction of Chloroplasts with Inhibitors
INDUCTION OF CHLOROSIS BY DIURON DURING PROLONGED ILLUMINATION IN VITRO
Received for publication June 3, 1976 and in revised form December 10, 1976
STUART M. RIDLEYBiochemistry Section, Jealott's Hill Research Station, Bracknell, Berkshire RG12 6EY, England
ABSTRACT
A primary symptom of diuron (DCMU) phytotoxicity in plants is thedestruction of chlorophyll. To study this process in vitro, chloroplstsfrom pea leaves (Pisum sativum L.) have been incubated in the lghtwith DCMU for periods of up to 34 hours. The sequence of photode-struction of chlorophylls and carotenoids has been followed to try andestablish the nature of the chloroplast protection mechanisms that aredestroyed by DCMU. 13-Carotene decays most rapidly, followed bychlorophyll a and xanthophylls which are destroyed in a constant ratio,followed filly by chlorophyll b. Bypassing the DCMU block in theelectron transport system with an artificia electron donor providescomplete protection against chlorophyll and carotenoid photodestruc-tion. The same protection by this electron donor system is afforded tostroma-free lamellae from which soluble reductants have been removedso that NADPH formation, which has been proposed as an essential partof a protective xanthophyll cyde, is not possible. Both this and thesimltaneous loss of chlorophyll a and xanthophylls tend to predude thebreakdown of a xanthophyll cyde from the possible protective mecha-nisms inhibited or destroyed by DCMU.
Cofactors of cylic electron transport also protect against DCMU-induced photodestruction of pigments. Their concentration dependencefor this protection appears to reflect their various abilities to catalyzecyclic photophosphorylation. The extent to which the chlorophyUls aredestroyed in the major pigment-protein complexes from chloroplastsEuminated with and without DCMU has been measured. In the absenceof DCMU, the light-harvestig chlorophyll a/b protein complex is de-stroyed most rapidly. In the presence of DCMU, the losses of chloro-phyll a from the photosystem I P700-chlorophyll a protein and thechlorophyll a/b complex are about the same. Chlorophyll losses arematched by simultaneous losses of the protein moieties; spectral analysesshow that the remahing chlorophyll a is held in a loose association withthe protein. Phenazine methosuate protects the chlorophyll of the lght-harvesting complex in DCMU-treated chloroplasts more than it protectsthat in photosystem I. Data published on DCMU-induced fluorescenceand its quenching are used to interpret the longer term DCMU-inducedchlorosis and its protection. By blocking electron transport, conforma-tional changes in the membrane that alow spUlover of excitation energyfrom photosystem II to photosystem I (and quenching of fluorescence bythis means) are prevented. The mechanism that normally protects thechloroplast against excessive illumination is then overloaded which im-pairs the harmless dissipation of absorbed light energy; consequently,the pigments are destroyed. When photosystem I is allowed to functionagain through cyclic electron flow, a necessary conformational change isbelieved to be reintroduced that once again allows the harmless dissipa-tion of excitation energy through spiElover. A functional electron trans-port system associated with photosystem I will protect against DCMU-induced chlorosis when the thylakoid membranes are intact, but whenthe P700-chlorophyll a protein complex is in isolation, there is only alimited degree of protection.
Diuron has been widely used as a research tool for studyingelectron transport, fluorescence, and energy transfer in chloro-plasts. The broad application of DCMU in research should notobscure the fact that this inhibitor, together with many othersacting in a similar manner, is among the most widely usedherbicides throughout the world. It is well established that withsuch herbicides whose mechanism of action is to block electrontransport in chloroplasts, the extent of phytotoxicity is a functionof light intensity (4, 20), and Chl is the principal absorbingpigment (4). A primary symptom of this type of inhibition ischlorosis brought about by the inability of Chl to dissipate itsabsorbed light energy which leads to irreversible photodestruc-tion (12, 13).The loss of pigments in treated plants points to a breakdown
or overloading of the mechanism that normally protects thechloroplast against excessive illumination. It has long been rec-ognized that the carotenoids play a major role in this protection(2, 8), and two mechanisms for the dissipation of light energyabsorbed by Chl in excess of that used for photosynthesis mayoperate in plants: (a) Witt (41) has provided evidence for triplet-triplet excitation energy transfer from Chl a to a special carote-noid which is then rapidly deactivated; (b) Krinsky (18) haspostulated that xanthophylls may remove excess excitation en-ergy from Chl through an epoxide cycle.The epoxide cycle hypothesis has been used by Stanger and
Appleby (32) in proposing a mechanism for DCMU-inducedphytotoxicity. By stopping the generation of reduced pyridinenucleotide, DCMU prevents the regeneration of oxidized carot-enoid, which in turn leads to destruction of Chl. These authorsdemonstrated that in DCMU-treated chloroplasts illuminatedfor 14 hr, carotenoids began to be destroyed before Chl, andthat protection was afforded to the pigments when the block inelectron transport was bypassed with an artificial electron donor(ascorbate plus DCIP'). Both of these findings appear to supportKrinsky's hypothesis of protection, and illustrate one way inwhich DCMU might cause a breakdown of a protection mecha-nism. However, there are a number of objections to the role ofan epoxide cycle in protection (19). For instance, the de-epoxi-dase is situated on the inner (loculus) side of the thylakoidmembrane (10), a site that is unlikely to be readily accessible tothe pyridine nucleotide reduced in the stroma and needed tofulfil Stanger and Appleby's hypothesis (32). Recent data sug-gest that the cycle is linked to PSI rather than PSII (31); further-more, NADPH is required for the epoxidation rather than thede-epoxidation reactions (30). It is therefore necessary to seekanother interpretation of the way in which DCMU might destroy
l Abbreviations: DCIP: 2,6-dichlorophenol indophenol; FMN: flav-ine mononucleotide; PMS: phenazine methosulfate; TMPD: N,N,N',N'-tetramethvl-p-nhenvlenediamine.
724
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

DCMU-INDUCED CHLOROSIS IN VITRO
the Chl protection mechanisms in plants and so induce chlorosis.The use of DCMU as a research tool in vitro can provide a
valuable insight of the consequences of its action in vivo. At-tempts are made in this paper to explain the mechanisms forDCMU-induced chlorosis by drawing on information reportingconditions that affect fluorescence yield. The site of action ofDCMU is known to be located close to the primary acceptor (Q)of PSII, which allows the reduction of Q but not its oxidation. Qquenches the fluorescence produced by PSII but not when in thereduced form (6) as it is when chloroplasts are poisoned withDCMU. Photosystem I reactions such as those involving non-physiological cofactors of cyclic electron transport do quenchPSII fluorescence when DCMU is present (see review 23), andalso, as shown in this paper, protect against chlorosis. Fluores-cence in DCMU-treated chloroplasts can then be used as anindicator of the spillover of excitation energy from PSII to PSI(23).An examination of chloroplasts in isolation under prolonged
periods of illumination with inhibitors provides a useful tool forstudying selective destruction of assembled plastid components.Particular reference is made in this paper to the fate of thepigments, which during prolonged illumination, act as photosen-sitizers for the destruction of the chloroplasts, and ultimately aredestroyed themselves. The availability of techniques for isolatingChl-containing fractions of chloroplasts enables one perhaps topinpoint more accurately the selective destruction of the pig-ments within these complexes. The evidence suggests that anepoxide cycle for the protection of Chl is unlikely to be relevantto the mechanism of action of DCMU, and that conformationalchanges in the membrane which allow optimum dissipation ofexcessive excitation energy are an essential part of the protectivemechanism that DCMU destroys.
MATERIALS AND METHODS
Plant Material. Chloroplasts were isolated from pea leaves(Pisum sativum L. var. Onwards). The plants were grown for 2to 3 weeks in a glasshouse under 18,000 lux illumination from400 w Atlas 'Kolorlux' lamps and the regime was 16 hr lightfollowed by 8 hr dark.
Isolation of Chloroplasts. Fifty g of leaves were homogenizedin an Atomix for 8 sec in an isolation medium containing 330mM sorbitol, 50 mm Tricine, 10 mm NaHCO3, 2 mm KNO3, 2mM EDTA (Na2 salt), 1 mM MgCl2, 1 mm MnCl2, 0.5 mMK2HPO4 (pH 7.4). Leaf debris and whole cells were removed byfiltering the slurry through four layers of butter muslin into a 85-mm Hartley-Buchner funnel in which were clamped six 12-cmdiscs of 25-,um nylon bolting cloth (Marine Biological Supplies,Plymouth, England). The filtrate was centrifuged at 2,000g for 1min and the chloroplast pellet resuspended in a medium contain-ing 330 mm sorbitol, 50 mm HEPES, 10 mm NaHCO3, 2 mMKNO3, 2 mm EDTA (Na2 salt), 1 mM MgCI2, 1 mm MnCl2, 0.5mM K2HPO4 (pH 7.4). All operations were carried out at 0 to 2C. This yielded a chloroplast preparation that was 80 to 90%intact as judged by phase contrast microscopy of the numbers ofclass I and intermediate class chloroplasts present (25).
For the preparation of stroma-free lamellae, the chloroplastswere sedimented at 2,000g for 10 min and resuspended in 10mM HEPES-NaOH buffer (pH 7.5), homogenized in a Potter-Elvehjem homogenizer, and resedimented at 48,000g for 20min. The buffer wash was repeated twice more to remove asmuch of the soluble proteins as possible without causing unduedamage to the lamellar system. The lamellae were finally resus-pended in the same medium used for the whole chloroplasts.
Incubation of Chloroplasts. Chloroplast or lamellar prepara-tions were diluted with the resuspension medium to roughly 250,UM Chl. Samples (4 ml) were incubated in 10-ml flasks attached
to a 10-cm disc which was rotated at 1 rpm at a slight angle toallow mixing. The flasks were immersed in a glass water bathmaintained at 20 C and illuminated from below by six 150 wincandescent lamps giving a light intensity of 14,000 lux at thewater surface.Aqueous solutions of DCMU, and cofactors of cyclic and
noncyclic electron flow were added to the chloroplast suspensionbefore illumination. In some experiments, DCMU in an etha-nolic solution was added but never in a greater volume than 201lI. On these occasions, the same volume of ethanol was addedto all of the other flasks. All treatments were carried out induplicate. Samples of 100 to 200 ,ul were removed from thechloroplast suspensions at intervals for pigment analyses.
Solubilization of Lameflae with Detergent. Stroma-free chlo-roplast lamellae were solubilized in 1% SDS in 50 mm tris-HClbuffer containing 2 mM MgCl2, at pH 8 when the extract wasneeded for electrophoresis, or 1% Triton X-100 in 50 mM tris-HCI containing 20 mm sodium isoascorbate at pH 7.4 when theextract was needed for the preparation of the P700-Chl a pro-tein. The final ratios of detergent to Chl were 10:1 (w/w) for theSDS extracts, and 75:1 (w/w) for the Triton extracts (1). In atimed experiment when there had been a loss of Chl, the ratio ofdetergent to Chl was based on the Chl in the unilluminatedcontrol samples; then the same quantity of detergent was addedto the other samples irrespective of the Chl concentration so thatthe detergent to chloroplast ratio was kept the same for allsamples. The SDS extracts were centrifuged at 40,000g for 50min, and the Triton X-100 extracts at 40,000g for 10 min; thepellets were discarded.
Analytical Polyacrylamide Gel Electrophoresis. Thirty ,ul ofSDS-solubilized lamellae were loaded onto polyacrylamide gelsprepared as described by Thornber (36) except that 8% gelscontaining 0.25% SDS in 50 mm tris-HCl buffer containing 2mM MgCl2 at pH 8 were used in glass tubes of 7 mm (i.d.) x 85mm. The lamellae components were resolved by electrophoresisat 4 mamp/tube (about 75 v) into three major pigmented zonesas reported by Thornber et al. (38).
For qualitative spectral analysis, the pigment bands weresliced from the gels, placed between two glass sheets (13 x 25mm; 0.5 mm thick) hinged at one end, and scanned in anAminco DW-2 recording spectrophotometer.
For the location of proteins after electrophoresis, the gelswere fixed and stained in 0.05% (w/v) Coomassie brilliant bluein 25% isopropyl alcohol-10% acetic acid. Excess dye wasleached from the gels with 10% acetic acid in which the gels werealso stored. Gels were scanned in a Joyce Loebl Chromoscandensitometer using a red filter (No. 620).
Preparative Polyacrylamide Gel Electrophoresis. For the di-rect quantitative estimation of Chl in the pigmented zones, 300,ul of the SDS-solubilized lamellae were loaded onto 8% poly-acrylamide gels in glass tubes of 16 mm (i.d.) x 80 mm. Thecomponents were resolved by electrophoresis at 20 mamp/tube(about 80 v) into the three major pigmented zones. The zoneswere sliced from the gel as soon as there was a gap of about 2mm between the light-harvesting Chl a/b protein band and thefree pigment zone; as much pigment-free gel as possible wasretained in the slice on one side of the band. The bands werethen removed from the gel slice by electrophoresis into a smallcolumn of Sephadex G-200. This was accomplished in a tube (16x 80 mm) one end of which was sealed with 24/322 Viskingdialysis membrane held in position by two 0-rings. A slurry ofSephadex G-200 in 0.05% SDS, 10 mm tris-HCl containing 1mM-MgCl2 at pH 8 was poured onto the membrane and allowedto settle to a height of 50 mm and held in place with a glass fiberdisc. The gel slice was pushed into position on the glass fiber discand air allowed to escape through a fine 75-mm long microsy-ringe needle. A current of 20 mamp/tube was applied until all of
Plant Physiol. Vol. 59, 1977 725
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 59, 1977
the pigment had moved into the Sephadex column; a 50-mmcolumn was sufficient to allow complete migration from the gelinto the Sephadex without the pigment migrating as far as thedialysis membrane. The Chl-protein was eluted from the Sepha-dex with acetone for Chl determination.
Isolation of the P700-Chlorophyl a Protein. The techiqueusing a hydroxylapatite column (60 mm high x 27 mm diam)was essentially that reported by Shiozawa et al. (29).
Determination of P700 to Chlorophyll Ratios. The Chl con-centration in a Triton X-100 extract of chloroplast lamellae, andin the isolated P700-Chl protein was determined from the ab-sorption spectrum using e = 60 mm-' cm-' at 677 nm (35).P700 concentration was estimated on an Aminco DW-2 dual
wavelength, double beam recording spectrophotometer from thereversible light-induced absorbance change at 697 nm, using anisosbestic point of 725 nm as a reference (29). An extinctioncoefficient for P700 at 697 nm of 64 mm-' cm-' was used (14).
Incubation of the P700-Chl a Complex. The conditions werethe same as those for incubation of whole chloroplasts exceptthat 3-ml samples of the P700-Chl a complex in duplicate wereincubated.
Chlorophyll Estimation. The concentration of Chl a and b in80% acetone extracts of chloroplast lamellae or fractions thereofwas estimated by the method of Arnon (3).
Carotenoid Estimation. Bulk carotenoids were estimated in80% acetone extracts of chloroplasts from the absorption at 480nm after correction for Chl interference (17); an extinctioncoefficient of 100 mm-' cm-' at 480 nm (corrected) was thenemployed. This coefficient was established by direct estimationof xanthophylls and carotenes from five separate acetone ex-tracts.
Carotenoids were fractionated into carotenes and xantho-phylls by the method of Davies (5). An 80% (v/v) acetoneextract of chloroplasts or P700-Chl a protein was combined withan equal volume of hexane in a separating funnel. The loweracetone phase was re-extracted with hexane, and the hexaneextracts combined and washed four times with an equal volumeof water. Xanthophylls were removed from the hexane fractionin four extractions with an equal volume of 90% methanol,which were combined, evporated to dryness under reduced pres-sure, and taken up in peroxide-free diethyl ether. Both thehexane and the ether extracts were saponified with 30% metha-nolic KOH and then washed with water; water was removedfrom the solvent extracts by partition into a 30% solution ofNaCl. The concentration of total xanthophylls in ether wasdetermined from the spectra using an absorption coefficient of142 mm-' cm-' at 443 nm (based on lutein [19]). The concentra-tion of 8-carotene in hexane was determined using an absorptioncoefficient of 137 mm-' cm-' at 451 nm.
Photosystem II Activity. Oxygen evolution by uncoupledchloroplasts was recorded in a water-jacketed Rank electrodefitted with a polyethylene membrane No. 5937 (Yellow SpringsInstrument Co.). Illumination was from a 100 w halogen projec-tor lamp focused onto the electrode cell. The light beam waspassed through a Calflex C interference heat filter, and two 3-mm red Perspex filters (ICI No. 400), giving a spectral cut-off at592 nm. The light energy at the surface of the electrode cell was1400 ILeinsteins/m2/sec. The cell was attached to a ChurchillChiller Thermo Circulator and maintained at 20 C, and 02evolution recorded on a W + W 1100 recorder (Kontron Tech-nik Ltd.) fitted with a zero suppressor.The reaction mixture (3 ml) contained 330 mm sorbitol, 50
mM Tricine, 40 mM KHCO3 (pH 7.5), 5 mm potassium ferricya-nide, 3 mm ammonium chloride, and chloroplasts containingapproximately 100 ,ug Chl.
RESULTSLoss of Chlorophyll in Intact Chloroplasts. When chloroplasts
are incubated with 5 ,UM DCMU, there is a light-dependent lossof Chl showing linear kinetics over 34 hr after a lag period of 2 to
4 hr (Fig. 1). Illuminated samples without DCMU (light con-trols) also show a loss of Chl over the same period but to a lesserextent and only after a lag of about 6 hr. PSII activity fallsrapidly in the first 2 hr of illumination but the subsequent loss ofactivity is gradual. Variations in the rate of photodestructioncould be observed in chloroplast suspensions with Chl concen-trations that were greater (slower rate of destruction) or less(faster rate of destruction) than the 250 .tM normally used. Achange in chloroplast density alters the effective intensity of thelight for destruction.When electrons are fed artificially to PSI using isoascorbate/
TMPD, and so bypassing the block, the Chl loss is the same asthat in the dark control.
Loss of Chlorophyls in Stroma-free Lamellae. The experi-ment in Figure 1 was repeated with stroma-free lamellae fromwhich all NADP and soluble reductants had been removed bythorough washing with buffer. Similar kinetic data of Chl loss arefound with DCMU-treated lamellae as found with intact chloro-plasts, except that .the rate of loss is slightly greater (Fig. 2). Thesame protection is afforded to the DCMU-treated lamellae byisoascorbate/TMPD. This clearly discounts the requirement ofNADPH in any protective mechanism involving an epoxide cyclebut does point to the involvement of a functional PSI for protec-tion.
Loss of Carotenoid Pigments. The loss of total carotenoidsrecorded over 26 hr is somewhat greater than that of the Chl inDCMU-treated chloroplasts but the essential difference is thelack of any lag period before photodestruction begins (Fig. 3).
Lighcontrol
80|1 <-<\4T M P~TMD
-602401\lV
20 Photosystem 11 °2 evolution(%) cmU
0 5 10 1 5 20 25 30 35Hours illumination
FIG. 1. Photodestruction of Chl in isolated chloroplasts in the pres-ence of 5 ,UM DCMU, and the absence (light control) of DCMU, andwith DCMU plus 500 jAM TMPD reduced with 10 mm Na isoascorbate,or with DCMU in the absence of light (dark control), and the loss of PSIIactivity in the illuminated control. Points represent the percentage of Chlor 02 evolution capacity compared with that in freshly prepared chloro-plasts.
0 5 10 15 20 25 30 35Hours illumination
FIG. 2. Photodestruction of Chl in stroma-free lamellae in the pres-ence and absence of DCMU, and with DCMU plus reduced TMPD.Details as in Figure 1.
726 RIDLEY
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

DCMU-INDUCED CHLOROSIS IN VITRO
Similar kinetic data of carotenoid loss and protection are foundwith illuminated stroma-free lamellae.
Fractionation of carotenoids into carotenes and xanthophyllsafter illumination of DCMU-treated chloroplasts shows that thegreatest percentage loss is in the carotene fraction, followed byChl a and xanthophylls and finally by Chl b (Table I). The ratioof Chl a to xanthophylls remains constant at 5:1 indicatingconcomitant loss of these pigments. The Chl a to carotene ratioincreases from 10:1 in the dark control samples to 34:1 in theDCMU-treated plastids, thereby showing preferential destruc-tion of these carotenoids over those of the xanthophylls.
Protection of Pigments against DCMU-induced Photode-struction. Not only does isoascorbate/TMPD protect Chl andcarotenoids against photodestruction (Figs. 1 and 3), but cofac-tors of cyclic electron transport (e.g. PMS and FMN) also giveprotection to the pigments in DCMU-treated chloroplasts duringillumination (Table II). DCIP and TMPD protect the chloroplastpigments from photodestruction whether reduced by isoascor-bate or not; in the absence of isoascorbate, 500 ,UM TMPD ismore effective than 500 ,UM DCIP in this protection (Table II).Sodium isoascorbate at 10 mm gives some protection (Table II),but as ascorbate does not readily donate electrons to P700 (16)this is more likely to be a direct antioxidant property.The concentration dependence of PMS, TMPD, and DCIP in
protecting against chlorosis in DCMU-treated chloroplasts isshown in Figure 4. PMS acts as an artificial proton carrier whilecatalyzing cyclic electron flow but it may also interact directlywith excited Chl molecules (23). The plot of PMS concentrationversus percentage protection against chlorosis is hyperbolic andwill give a linear reciprocal plot. It can be calculated from thisthat the PMS concentration for 50% protection is 6.5 ,UM. Insimilar experiments with stroma-free lamellae, the PMS concen-tration required is found to be less than 3 ,UM. Neither DCIP norTMPD yields linear reciprocal plots (from Fig. 4) suggestingdifferent or more complex protection mechanisms.
Distribution of Chlorophylls in Pigment Complexes of FreshlyPrepared Chloroplasts. Before studying the photodestruction of
15 20Hours illumination
FIG. 3. Photodestruction of total carotenoids in isolated chloroplastsin the presence and absence of DCMU, and with DCMU plus reducedTMPD. Details as in Figure 1.
Chl in detail, it is necessary to establish the Chl distribution inpigment complexes of fresh chloroplasts prior to DCMU treat-ment.
Chloroplasts were extracted with SDS and the pigment com-plexes separated by gel electrophoresis. Chl a and Chl b concen-
trations were determined and the percentage distribution calcu-lated in the three major pigment zones. Of the total (from fiveseparate experiments), complex I (the SDS-altered form of theP700-Chl a protein) contained 10 to 12% Chl a, the light-harvesting Chl a/b protein contained 28 to 30% Chl a and 22 to23% Chl b, and the free pigment zone contained 28 to 30% Chla and 10 to 11% Chl b. This distribution of Chl in the pigmentcomplexes is close to that published by others using differentspecies, and different methods of isolation and estimation (7,29). The free pigment zone contains about one-third of the totalChl b, most of which is assumed to come from the Chl a/bprotein complex because the true Chl a/b ratio in this complex is1.0 (37).DCMU-induced Photodestruction of Chlorophyll-Protein
Complexes. Table III shows that upon illumination of controlchloroplast suspensions, the greatest absolute losses of Chl occurin the Chl a/b protein and there is a rather small loss in thecomplex I band. In DCMU-treated chloroplasts, the increasedloss of Chl a over that in the illuminated control, from complex Iand the Chl a/b protein is similar, indicating an approximatelyequal DCMU-induced photodestruction of the two pigmentcomplexes in absolute terms; the percentage loss of Chl fromcomplex I is greater because this represents a smaller percentageof the total Chl compared with that in the Chl a/b protein.The addition of 500 ,UM PMS to DCMU-treated chloroplasts
increases the efficiency of energy dissipation and prevents the
TABLE II. Photodestruction of chlorophylls and total carotenoidsin DC.U-treated pea chloroplasts following 26 hours
illunination, and the protection conferred by, electrondonors and acceptors.
Chloroplasts were illuninated with the following final concentrationsof additions: DC4U, 5 uM; PYS, 500 M; FEN, I mM; DCIP, 500 uM;TMPD, 500 pM; sodiun isoascorbate, 10 mM.
* FMN interfered with the caretenoid estimation
TABLE I. Losses of individual chlorophylls and carotenoids in chloroplast suspensions illuminatedfor 20 hours with and without DCMU
Chl a Chl b Total carotenoidAdditions
um % loss uM % loss uM % loss
Before illumination 200 88 78
After 26 hrillumination:
Dark control 196 2 88 0 76 3
Light control 134 33 63 28 50 36
PMS 188 6 91 0 71 9
DCMU 64 68 47 47 20 74
DCKU + PMS 182 9 85 3 68 13
DCOU + FMN 189 5 88 0 --
DO!U + TMPD 188 6 88 0 78 0
DC1U. DCIP 155 22 84 4 57 27
DCMU + isoascorbate 135 32 73 17 40 49
OiLOROPHYLLS CARorENOIDS
Samnple i~ /OTotal Chl Chl a Chl b Carotenes Carotee Xanthophylls Xanth /llsuM % loss UM % loss uM % loss uM % loss uM % loss
Dartc control 2 38 .9 172 .2 66 .7 2 3.2 10: 1 36 .8 5: 1
Light control 209.1 12 140.7 18 68.4 0 14.5 37 14:1 31.1 16 5:1
+5LM DCMU 132.0 45 80.2 53 51.8 22 3.9 83 34:1 17.1 54 5:1
Plant Physiol. Vol. 59, 1977 727
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 59, 1977
light-induced Chl loss; the small losses found in the dark controlsuspensions were unaffected by this concentration of PMS. The10 ,UM PMS treatment affords a greater protection of the Chl inthe Chl a/b protein (35% protection of Chl a and 56% protec-tion of Chl b) than in complex I (12% protection of Chl a). Inother preliminary experiments with PMS in DCMU-poisonedplastids illuminated for shorter periods (data not shown here), ithas been possible to retain complete protection of the Chl a/bprotein while still inducing as much as 45% loss in the complex IChl a protein.The proteins associated with the Chl in the pigment complexes
are also subject to destruction. This is revealed in densitometerscans of polyacrylamide gels after electrophoresis of SDS ex-tracts from control and DCMU-treated chloroplasts (Fig. 5).The protein-stained gels show the greatest losses in those bandsthat also contain Chl; the major nonpigmented protein is unaf-fected by the DCMU-induced photodestruction. The scans alsoclearly demonstrate the marked relative DCMU-induced loss ofthe photosystem I band.A comparison of spectra of the pigment-containing zones in
situ in the gels from chloroplasts illuminated with and withoutDCMU (Fig. 6) shows shifts in the absorption maxima in the twomajor Chl-protein complexes but not in the free pigment zones.In the DCMU-treated chloroplasts, the red maximum of Chl a incomplex I is shifted from 675 to 669 nm which is closer to themaximum found in the free pigment zone. Changes in the Soretband at 437 nm relative to the maximum for pheophytin at 422
60 80
liM cofoctor
FIG. 4. Protection against DCMU-induced loss of Chl in isolatedchloroplasts by different concentrations of cofactors of cyclic electrontransport after 22-hr illumination. Protection is plotted as a percentageof the Chl in the dark control after subtracting the Chl remaining in theDCMU-treated chloroplasts to which no cofactors had been added.
TABLE III. Loss of chlorophylls from the pigmentcompared with freshly prepared plastid
nm indicate Mg2+ loss from the Chl molecule itself. The light-harvesting Chl protein shows that the red maximum for Chl b isunchanged at 652 nm in spite of a 43% loss of this Chl (TableIII); a small red shift from 671 to 668 nm might be accounted forentirely by the relatively greater contribution by Chl b, althougha looser association with the protein may also be involved. Thesmall downward shift in the Soret band of Chl b from 471 to 469nm is due to the loss of carotenoids absorbing around 480 nm;these are the most readily destroyed pigments (Table I).
Photodestruction and Protection of the Isolated P700-Chl aComplex. The heart of PSI (the P700-Chl a protein) remainsphotochemically active in isolation and we can therefore exam-
FIG. 5. Densitometer tracings of polyacrylamide gels of the electro-phoretically separated pigment complexes extracted with SDS fromchloroplasts after they had been illuminated with or without DCMU for21 hr. Electrophoresis was from left to right. Gels stained for proteintend to swell slightly so that the horizontal scale in comparison with theunstained gels is slightly extended. LH-Chl: light-harvesting Chl a/bprotein.
complexes of chloroplasts illuninated for 21 hrs
Picaent bands were separated by preparative gel electrophoresis of SDS extracts of lamellae,(each extract had the same chloroplast to SDS ratio - see Methods). A loading of fresh chloroplastscontained 261 nmol of Chl a and 123 nmol of Chl b. Negative figures represent gains.
Total chl Complex I Chl a/b protein Free Pigmentloaded
Additions
Chl a Chl b Chl a Loss l Loss Chl b Loss l Loss l Loss- lost ~ lost lost lost lost
nmol nmol % nmol % nmol % nmol % nmol %
Dark control 243 106 0 0 6 6 9 11 14 13 8 13
Light control 230 96 5 11 21 20 19 23 5 5 8 21
D36JU (SuM) 167 90 31 67 55 52 36 43 8 7 -3 -8
DCMU + Pt'S (500aM) 257 115 0 0 10 9 5 6 -5 -5 3 8
DC JU+ PM3 (O1 M) 179 102 27 58 38 36 21 25 17 16 0 0
728 RIDLEY
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

DCMU-INDUCED CHLOROSIS IN VITRO
ILLUMINATION -DCMU ILLUMINATION + DCMU
u
z
0tn
0400 45 50 600
L600 65350 400 450 S00 S50 6500 650 700 350 400 450 500 550 600 650O 700
Wavelength (nm)
FIG. 6. Absorption spectra of the pigmented zones following separation by gel electrophoresis of SDS extracts of chloroplasts which had beenilluminated with or without DCMU for 21 hr. The zones were sliced from the gels and scanned directly. PSI Chl protein refers to the SDS-alteredP700-Chl a protein (complex I).
ine photodestruction in this chloroplast fraction in more detail.In preparations of the P700-Chl a protein complex, the Chl to
P700 ratio ranged from 40:1 to 48:1. (-Carotene was presentand in a ratio with P700 of 8 to 9: 1, amounting to approximately20% of the total chloroplast carotene; this ratio is higher thanthat reported in similar preparations from a number of differentspecies (29). Preparations of the complex were illuminated forperiods of up to 18 hr with direct analyses of P700 and Chl atintervals. Photodestruction of both P700 activity and antennaeChl begins without any lag (data not shown). Prior treatment ofthe complex with 5 ,UM DCMU produces no significant effectover that of the illuminated control (Table IV). There is a
greater percentage loss of P700 than of the antennae Chl but theP700 to (-carotene ratio remains the same in all experimentsboth before and after illumination, suggesting a close associationof the two (Table IV).
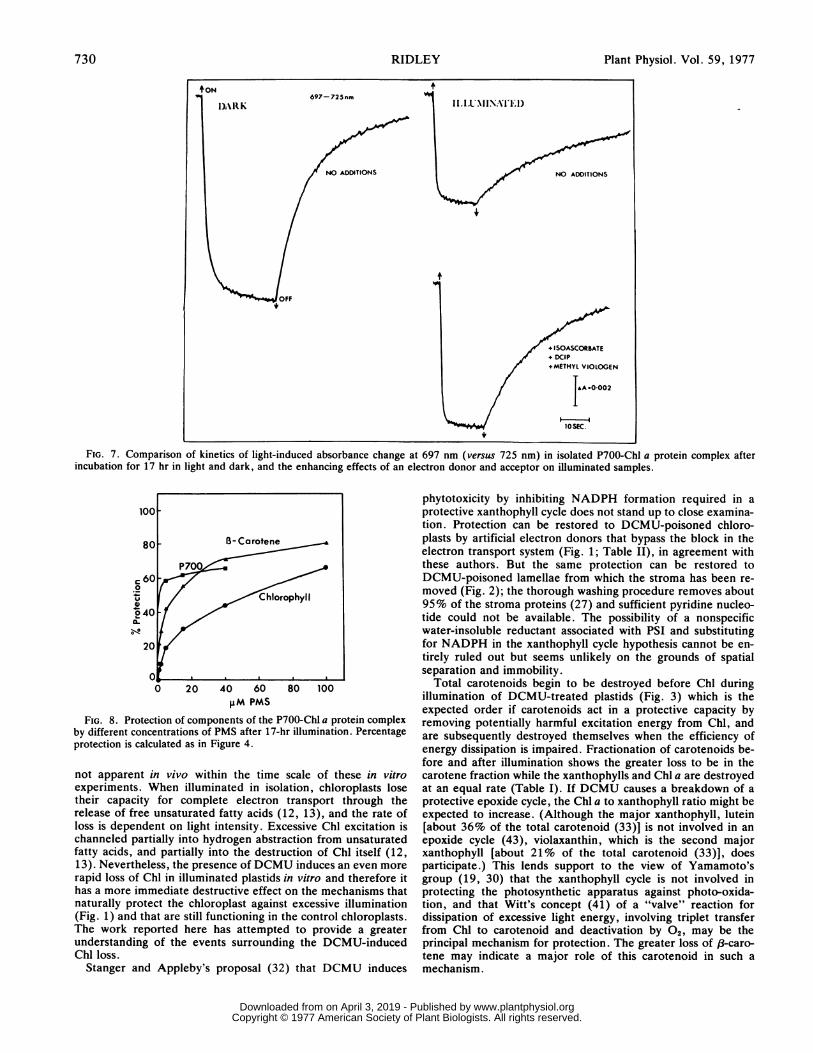
In the assay for P700 activity, the dark reduction followinglight-induced oxidation in the absence of added electron donorsand acceptors is markedly slower after prolonged illumination ofthe P700-Chl a complex (Fig. 7) indicating that the ability of theendogenous reductant to cycle electrons back to P+700 has beenimpaired. The addition of isoascorbate, DCIP, and methyl violo-gen to a control sample of the complex after prolonged illumina-tion increases the P700 activity by about 50% over that in theabsence of electron donors and acceptors; no such increase isfound in the dark control samples after prolonged incubation.These differences may reflect a partial light-induced dislocationof the P700 reaction center from the other electron transportcomponents within the P700-Chl a protein complex.Isoascorbate/TMPD added to the P700-Chl a protein in the
same concentrations used with whole chloroplasts (Fig. 1) failsto protect the Chl in the complex during illumination; P700activity cannot be measured using light activation in the presenceof these additives. PMS provides partial protection to the com-ponents of the complex (Fig. 8) with a preference for ,-carotene
TABLE IV. Photodestruction of chlorophyll and B-carotene in the isolatedP700-Ol a protein complex.
Duplicate 3 ml samrles of the complex were incubated and illuminatedfor 18 hr uneer the same conditions used for isolated chloroplasts (seeMethods).
and P700. The percentage protection of Chl versus PMS concen-tration does not yield a linear reciprocal plot as it did with wholechloroplasts. P700 activity cannot be assayed using light activa-tion in samples containing more than 40 AM PMS because it isnot possible to poise the system in the oxidized state. Theaddition of different concentrations of PMS to the illuminatedcontrol after the 17-hr illumination did not raise the measurableP700 activity to the levels in samples to which the same concen-trations of PMS had been added before the 17-hr illumination.Protection of P700 by PMS is therefore genuine and not just anartifact of the assay.
DISCUSSION
In prolonged experiments with chloroplasts in vitro, theDCMU-induced effects have to be superimposed on the photo-inhibition effects of control samples lacking DCMU. Isolatedchloroplasts exhibit light-induced decay characteristics that are
p700 Chl a hl/ 0-carotene O-carotene/SamplesP0070iuM % loss M % loss
P700
UN % lossP700
Zero time 0.2226 10.68 48:1 2.06 9:1
18 hr
Ililuminated:Dark control 0.2226 0 9.51 11 43:1 2.05 0 9:1
Light control 0.0804 64 6.12 42 77:1 0.77 63 10:1
*5uM DCMU 0.0831 63 6.13 43 74:1 0.69 66 8:1
729Plant Physiol. Vol. 59, 1977
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 59, 1977
FIG. 7. Comparison of kinetics of light-induced absorbance change at 697 nm (versus 725 nm) in isolated P700-Chl a protein complex afterincubation for 17 hr in light and dark, and the enhancing effects of an electron donor and acceptor on illuminated samples.
C ou
Chlorophyll1040
240
20
0 20 40 60 80 100PM PMS
FIG. 8. Protection of components of the P700-Chl a protein complexby different concentrations of PMS after 17-hr illumination. Percentageprotection is calculated as in Figure 4.
not apparent in vivo within the time scale of these in vitroexperiments. When illuminated in isolation, chloroplasts losetheir capacity for complete electron transport through therelease of free unsaturated fatty acids (12, 13), and the rate ofloss is dependent on light intensity. Excessive Chl excitation ischanneled partially into hydrogen abstraction from unsaturatedfatty acids, and partially into the destruction of Chl itself (12,13). Nevertheless, the presence of DCMU induces an even morerapid loss of Chl in illuminated plastids in vitro and therefore ithas a more immediate destructive effect on the mechanisms thatnaturally protect the chloroplast against excessive illumination(Fig. 1) and that are still functioning in the control chloroplasts.The work reported here has attempted to provide a greaterunderstanding of the events surrounding the DCMU-inducedChl loss.
Stanger and Appleby's proposal (32) that DCMU induces
phytotoxicity by inhibiting NADPH formation required in aprotective xanthophyll cycle does not stand up to close examina-tion. Protection can be restored to DCMU-poisoned chloro-plasts by artificial electron donors that bypass the block in theelectron transport system (Fig. 1; Table II), in agreement withthese authors. But the same protection can be restored toDCMU-poisoned lamellae from which the stroma has been re-moved (Fig. 2); the thorough washing procedure removes about95% of the stroma proteins (27) and sufficient pyridine nucleo-tide could not be available. The possibility of a nonspecificwater-insoluble reductant associated with PSI and substitutingfor NADPH in the xanthophyll cycle hypothesis cannot be en-tirely ruled out but seems unlikely on the grounds of spatialseparation and immobility.
Total carotenoids begin to be destroyed before Chl duringillumination of DCMU-treated plastids (Fig. 3) which is theexpected order if carotenoids act in a protective capacity byremoving potentially harmful excitation energy from Chl, andare subsequently destroyed themselves when the efficiency ofenergy dissipation is impaired. Fractionation of carotenoids be-fore and after illumination shows the greater loss to be in thecarotene fraction while the xanthophylls and Chl a are destroyedat an equal rate (Table I). If DCMU causes a breakdown of aprotective epoxide cycle, the Chl a to xanthophyll ratio might beexpected to increase. (Although the major xanthophyll, lutein[about 36% of the total carotenoid (33)] is not involved in anepoxide cycle (43), violaxanthin, which is the second majorxanthophyll [about 21% of the total carotenoid (33)], doesparticipate.) This lends support to the view of Yamamoto'sgroup (19, 30) that the xanthophyll cycle is not involved inprotecting the photosynthetic apparatus against photo-oxida-tion, and that Witt's concept (41) of a "valve" reaction fordissipation of excessive light energy, involving triplet transferfrom Chl to carotenoid and deactivation by 02, may be theprincipal mechanism for protection. The greater loss of /3-caro-tene may indicate a major role of this carotenoid in such amechanism.
730 RIDLEY
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

DCMU-INDUCED CHLOROSIS IN VITRO
The protection afforded to DCMU-treated chloroplasts bycofactors reduced with isoascorbate is through the promotion ofelectron transport associated with PSI, but not in the generationof NADPH; a direct involvement of these reduced cofactors as
substitutes for NADPH in a de-epoxidation reaction is notlikely. In the absence of isoascorbate, the concentration depend-ence of PMS, TMPD, and DCIP in protection (Fig. 4) reflectsthe ability of these cofactors to catalyze cyclic photophosphory-lation. DCIP supports cyclic electron flow but involves moreelection transport intermediates than PMS (40) and so is lessefficient at dissipating excitation energy. On the other hand,only at high concentrations does TMPD catalyze any degree ofphotophosphorylation (28), which agrees with the concentrationeffect of TMPD on Chl protection (Fig. 4).The dissipation of excitation energy may be promoted either
through cyclic electron transport (native or artificial) by activat-ing an energy-conserving site (11, 28), or (with reduced cofac-tors) through noncyclic electron flow to an endogenous compo-nent that consumes 02 in a Mehler reaction; this is particularlytrue of chloroplasts that have become disrupted (26). In theformer alternative (and possibly to some degree in the latter as
well), the mechanism for the nonradiative energy dissipationmay be analogous to fluorescence-quenching through conforma-tional changes in the membrane, which occur when PSI-associ-ated electron transport is functioning; this allows a greater access
of 02 to quench the excitation energy (22, 24, 42). Theseconformational changes have been assigned to the generation ofa proton gradient, and also to structural changes in the couplingfactor (15, 21-23). Gross and Prasher (9) have demonstratedusing fluorescence and electron microscopy that conformationalchanges in the membrane result in increased spillover of excita-tion energy. The protection of Chl in DCMU-treated chloro-plasts by various redox compounds, whether in the reduced formor not, implies that there must be a considerable spillover ofexcitation energy from PSII to PSI in order to circumvent theblock in the electron transport system. Comparative work iscurrently being pursued to see to what extent an analogy be-tween data on fluorescence yield and the mechanisms that in-duce the much longer term effects of chlorosis may be taken. It isnot yet clear whether cofactors of cyclic electron flow re-estab-lish in the chloroplasts conditions that allow the natural protec-tive mechanism to work, or whether a different artificial mecha-nism is introduced.The sequence of energy transfer between pigments in the
highly ordered lamellae membrane structure is likely to be re-
flected in the order and extent to which individual pigments are
photodestroyed during prolonged illumination of chloroplasts,either protected or unprotected by cofactors of cyclic electrontransport, and in the presence or absence of DCMU. Thissequence has been revealed by fractionating the major pigment-protein complexes and analyzing the Chl. In the illuminatedcontrol, the Chl a/b protein is principally affected (Table III).The capacity for 02 evolution falls to 13% of its original level in3 hr (Fig. 1) so that the Chl loss is likely to be caused by theaccumulation of a PSII oxidant as found by Yamashita andButler (44) in tris-washed chloroplasts. The 6-fold increase inChl loss from complex I when DCMU is present (Table III)suggests that a different mechanism of photodestruction associ-ated with PSI is introduced. DCMU induces similar losses (overthe illuminated control) of Chl a in complex I and in the Chl a/bprotein (Table III), which points to a lack of any gross energyspillover.The protective action of PMS (especially as found under
conditions of partial protection at low concentration) favors theChl of the Chl a/b complex, with less protection of Chl a incomplex I. The sequence of Chl photodestruction reflects thepromotion of excitation energy spillover from PSII to PSI andagrees with the effect of PMS in quenching PSII fluorescence by
this means (23). However, the interpretation of selective loss ofChl in these complexes is complicated by movement of Chl to thefree pigment zone as well as actual photodestruction (Table III).Chl b is located principally (if not entirely) in the light-harvestingChl a/b protein (39) but is subject to less photodestruction thanChl a because of efficient energy transfer from Chl b to Chl a.That the sequence of pigment photodestruction in DCMU-poi-soned chloroplasts reflects the sequence of energy migrationbetween the pigments seems to be generally upheld for theduration of these experiments, although it is fully realized thatthe mechanism of this selective destruction is complex and mustdepend on the accessibility of 02 to the various pigments in thesequence, and the rate at which each excited pigment is deacti-vated (reviewed by Witt [41]).The proteins to which the pigments are attached are also
destroyed in illuminated chloroplasts treated with DCMU (Fig.5). The Chl (or carotenoids) may act as photosensitizers for thedisruption of the protein moieties into subunits (this could bedirectly or indirectly through peroxidation of lipids associatedwith the complexes). Alternatively, the pigments may be essen-tial for the structural integrity of the proteins so that photodes-truction of Chl and carotenoid molecules results in structuraldisintegration of the complexes. The red maxima of Chl a in bothcomplex I and the Chl a/b protein are shifted to become morelike that in the free pigment zone indicating that the Chl associa-tion with the protein in these complexes has loosened (Fig. 6).The simultaneous losses of Chl and protein, seen in densitometerscans (Fig. 5), must leave the residual complexes in a disorgan-ized state but still with Chl attached to the protein. This isfurther demonstrated in the assay for P700 activity because ofthe reduced ability of endogenous reductants to cycle electronsback to P+700 without the aid of added electron donors andacceptors (Fig. 7).The involvement of a functional PSI in the protection of Chl in
DCMU-treated chloroplasts, particularly when cyclic electronflow is "short circuited" through PMS, has led to studies with theisolated P700-Chl a protein (37). This complex lacks electrontransport and light energy spillover from PSII, and contains littleor no lipid so that a membrane conformational change, believedto be an essential part of a protective mechanism, is not possible.The complex contains no xanthophylls that can participate in aprotective epoxide cycle.
Illumination of the isolated complex causes an increase in theChi to P700 ratio (Table IV) showing that the reaction center ispreferentially photodestroyed. On the other hand, the ,8-caro-tene to P700 ratio remains constant. If ,8-carotene were to play aprotective role for P700 by removing excessive excitation energyfrom this Chl, the carotene should decay before the Chl and theratio decrease. While the P700-Chl a protein in isolation maydiffer from that assembled in the thylakoid membrane, theconcomitant loss of P700 and /8-carotene suggests their interde-pendence. The structural integrity of those protein subunits inthe complex that contain P700 (only one-third of them do [37])could depend directly or indirectly on /-carotene, so that photo-destruction of this carotenoid leads to a dissociation of the P700-containing subunits in such a way that the measurable activity ofP700 is affected. This would account for the constant ratio ofP700 to /3-carotene. Support for this structural concept alsocomes from the densitometer tracings (Fig. 5) in which both theChl and the protein in the P700-Chl a complex are seen to bediminished after illumination. DCMU did not enhance photo-destruction in the complex which confirms that chlorosis inducedby 5 ,UM DCMU in whole chloroplasts results entirely from itsaction on PSII and not through any additional action on PSI.Some protection is afforded to the P700-Chl a complex by
artificial electron donors. Isoascorbate/TMPD does not protectthe Chl in the complex as it does in the whole chloroplast,probably because of the inability to catalyze noncyclic electron
731Plant Physiol. Vol. 59, 1977
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 59, 1977
flow. PMS, on the other hand, can facilitate significant protec-tion of P700 and ,8-carotene during an 18-hr illumination (Fig.8) but with less effect on the bulk Chl, which suggests that thoseparticles that contain P700 are protected through cyclic electrontransport when excitation energy is assumed to be dissipated asheat. The protein subunits containing only antennae Chl will notbe protected by PMS, except perhaps through direct quenchingof the excited Chl, and are therefore preferentially destroyed. Itis clear that the protection in the isolated P700-Chl a protein byPMS is not nearly as effective as that in whole chloroplasts(compare Fig. 4 with Fig. 8), which emphasizes the requirementof an intact membrane system for the full expression of a protec-tive mechanism. The ratio of PMS to P700 in preparations of theisolated complex and in whole chloroplasts is similar if calculatedon the basis of complex I accounting for 10 to 12% of the totalChl.The expression of herbicidal activity by DCMU may be sum-
marized as follows: (a) the inhibition of the electron transportsystem prevents the dissipation of light energy by this means andalso prevents conformational changes in the chloroplast mem-
brane; (b) the spillover of excitation energy from the light-harvesting Chl to PSI (dependent on membrane conformationalchange) does not occur; (c) the absorbed energy is now excessiveto requirement and must be dissipated through the protectivemechanism involving the carotenoids; (d) energy is transferredfrom Chl to carotenoid but the input is greater than can beharmlessly dissipated; (e) photo-oxidative destruction of thepigment complexes ensues (in waves starting from the two reac-tion centers) and, eventually, superimposed upon this is thedestruction of lipids which causes a further disarray or discon-nection of the pigment protein complexes.
PSI may initially promote a conformational change throughendogenous cyclic electron flow, which allows some energy spill-over (and fluorescence-quenching [34]). The gradual decay ofthe ,8-carotene associated with P700 would eventually ensurethat this dissipation route was also blocked. A more rapid herbi-cidal action of DCMU might therefore be achieved under condi-tions in which P700 activity was simultaneously destroyed. Moredefinitive proof of the inhibition of conformational changes inthe chloroplast membrane by DCMU, and the nature of suchchanges in these long term experiments is currently beingsought.
LITERATURE CITED
1. ALBERTE RS, JP THORNBER, AW NAYLOR 1972 Time of appearance of photosystems I andII in chloroplasts of greening jack bean leaves. I Exp Bot 23: 1060-1069
2. ANDERSON IC, DS ROBERTSON 1960 Role of carotenoids in protecting chlorophyll from
photodestruction. Plant Physiol 35: 531-534
3. ARNON DI 1949 Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta
vulgaris. Plant Physiol 24: 1-154. ASHTON FM 1965 Relationship between light and toxicity symptoms caused by atrazine and
monuron. Weeds 13: 164-168
5. DAVIES BH 1965 Analysis of carotenoid pigments. In TW Goodwin, ed, Chemistry and
Biochemistry of Plant Pigments. Academic Press, New York pp 489-5316. DUYSENS LNM, HE SWEERS 1963 Mechanism of two photochemical reactions in algae as
studied by means of fluorescence. In Studies on Microalgae and Photosynthetic Bacteria.
Japan. Soc Plant Physiol, University of Tokyo Press, Tokyo pp 353-3727. GENGE S, D PILGER, RG HILLER 1974 The relationship between chlorophyll b and pigment
protein complex II. Biochim Biophys Acta 347: 22-308. GRIFFITHS M, WR SISTROM, G COHEN-BAZIRE, RV STANIER 1955 The function of carote-
noids in photosynthesis. Nature 176: 1211-12159. GROSS EL, SH PRASHER 1974 Correlation between monovalent cation-induced decreases in
chlorophyll a fluorescence and chloroplast structural changes. Arch Biochem Biophys 164:460-468
10. HAGER A 1969 Lichtbedingte pH-Erniedrigung in einem Chloroplastenkompartiment alsUrsache der enzymatischen Violaxanthin-Zeaxanthin-Umwardlung; Beziehungen zur
Photophosphorylierung. Planta 89: 224-243
11. HAUSKA G, S REIMER, A TREBST 1974 Native and artificial energy-conserving sites in cyclicphotophosphorylation systems. Biochim Biophys Acta 357: 1-13
12. HEATH RL, L PACKER 1968 Photoperoxidation in isolated chloroplasts. I. Kinetics andstoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125: 189-198
13. HEATH RL, L PACKER 1968 Photoperoxidation in isolated chloroplasts. II. Role of electrontransfer. Arch Biochem Biophys 125: 850-857
14. HIYAMA T, B KE 1972 Difference spectra and extinction coefficients of P700. BiochimBiophys Acta 267: 160-171
15. JENNINGS RC, FM GARLASCHI, G FORTI 1976 Studies on the slow fluorescence decline inisolated chloroplasts. Biochim Biophys Acta 423: 264-274
16. KATOH S, A SAN PIETRO 1967 Ascorbate-supported NADP photoreduction by heatedEuglena chloroplasts. Arch Biochem Biophys 122: 144-152
17. KIRK JTO, RL ALLEN 1965 Dependence of chloroplast pigment synthesis on proteinsynthesis: effect of actidione. Biochem Biophys Res Commun 21: 523-530
18. KRINSKY NI 1966 The role of carotenoid pigments as protective agents against photosensi-tized oxidations in chloroplasts. In TW Goodwin, ed, Biochemistry of the Chloroplast VolI. Academic Press, London pp 423-430
19. LEE KH, HY YAMAMOTO 1968 Action spectra for light-induced de-epoxidation and epoxi-dation of xanthophylls in spinach leaf. Photochem Photobiol 7: 101-107
20. MINSHALL WH 1957 Influence of light on the effect of 3-p-(chlorophenyl)-1,1-dimethylureaon plants. Weeds 5: 29-33
21. MURAKAMI S, J TORES-PEREIRA, L PACKER 1975 Structure of the chloroplast membrane -relation to energy coupling and ion transport. In Govindjee, ed, Bioenergetics of Photo-synthesis. Academic Press, New York pp 555-618
22. MURATA N, K SUGAHARA 1969 Control of excitation transfer in photosynthesis. III. Light-induced decrease of chlorophyll a fluorescence related to photophosphorylation system inspinach chloroplasts. Biochim Biophys Acta 189: 182-192
23. PAPAGEORGIou G 1975 Chlorophyll fluorescence: an intrinsic probe of photosynthesis. InGovindjee, ed., Bioenergetics of Photosynthesis. Academic Press, New York pp 319-371
24. PAPAGEORGIOU G, J ISAKIDOU, C ARGOUDELIS 1972 Structure dependent control ofchlorophyll a excitation density: the role of oxygen. FEBS Lett 25: 139-142
25. RIDLEY SM, RM LEECH 1968 The survival of chloroplasts in vitro: particle volume distribu-tion patterns as a criterion for assessing the degree of integrity of isolated chloroplasts.Planta 83: 20-34
26. RIDLEY SM, RM LEECH 1970 Light-dependent consumption of oxygen by Vicia fabachloroplasts. Arch Biochem Biophys 139: 351-360
27. RIDLEY SM, JP THORNBER, JL BAILEY 1967 A study of the water-soluble proteins ofspinach beet chloroplasts with particular reference to fraction I protein. Biochim BiophysActa 140: 62-79
28. SCHWARTZ M 1966 N-Tetramethyl-p-phenylenediamine as a catalyst of photophosphoryla-tion. Biochim Biophys Acta 112: 204-212
29. SHIOZAWA JA, RS ALBERTE, JP THORNBER 1974 The P700-chlorophyll a-protein. Isolationand some characteristics of the complex in higher plants. Arch Biochem Biophys 165:388-397
30. SIEFERMANN D, HY YAMAMoTO 1975 Properties of NADPH and oxygen-dependentzeaxanthin epoxidation in isolated chloroplasts. A transmembrane model for the violaxan-thin cycle. Arch Biochem Biophys 171: 70-77
31. SIEFERMANN D, HY YAMAMOTO 1976 Light-induced de-epoxidation in lettuce chloroplasts.VI. De-epoxidation in grana and in stroma lamellae. Plant Physiol 57: 939-940
32. STANGER CE, AP APPLEBY 1972 A proposed mechanism for diuron-induced phytotoxicity.Weed Sci 20: 357-363
33. TAKEGUCHI CA, HY YAMAMOTO 1968 Light-induced 1802 uptake by epoxy xanthophylls inNew Zealand spinach leaves (Tetragonia expansa). Biochim Biophys Acta 153: 459-465
34. TELFER A, J BARBER, J NICOLSON 1975 Energy-dependent quenching of chlorophyll a
fluorescence. Evidence for coupled cyclic electron flow in isolated intact chloroplasts.Plant Sci Lett 5: 171-176
35. THORNBER JP 1969 Comparison of a chlorophyll a-protein complex isolated from a blue-green alga with chlorophyll-protein complexes obtained from green bacteria and higherplants. Biochim Biophys Acta 172: 230-241
36. THORNBER JP 1970 Photochemical reactions of purple bacteria as revealed by studies ofthree spectrally different carotenobacteriochlorophyll-protein complexes isolated fromChromatium, strain D. Biochemistry 9: 2688-2698
37. THORNBER JP 1975 Chlorophyll-proteins: light-harvesting and reaction center componentsof plants. Annu Rev Plant Physiol 26: 127-158
38. THORNBER JP, RPF GREGORY, CA SMITH, JL BAILEY 1967 Studies on the nature of thechloroplast lamella. I. Preparation and some properties of two chlorophyll-protein com-
plexes. Biochemistry 6: 391-39639. THORNBER JP, HR HIGHKIN 1974 Composition of the photosynthetic apparatus of normal
barley leaves and a mutant lacking chlorophyll b. Eur J Biochem 41: 109-11640. TREBST A 1974 Energy conservation in photosynthetic electron transport of chloroplasts.
Annu Rev Plant Physiol 25: 423-45841. Wrrr HT 1971 Coupling of quanta, electrons, fields, ions and phosphorylation in the
functional membrane of photosynthesis. Results by pulse spectroscopic methods. Q RevBiophys 4: 365-477
42. WRAIGHT CA, AR CROFrs 1970 Energy-dependent quenching of chlorophyll a fluorescencein isolated chloroplasts. Eur J Biochem 17: 319-327
43. YAMAMOTO HY, L KAMITE, Y-Y WANG 1972 An ascorbate-induced absorbance change inchloroplasts from violaxanthin de-epoxidation. Plant Physiol 49: 224-228
44. YAMASHITA T, WL BUTLER 1969 Photooxidation by photosystem II of tris-washed chloro-plasts. Plant Physiol 44: 1342-1346
732 RIDLEY
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.