interaction of cholera toxin and membrane gm1 ganglioside of small
TRANSCRIPT

Proc. Nat. Acad. Sci. USAVol. 72, No. 7, pp. 2520-2524, July 1975Biochemistry
Interaction of cholera toxin and membrane GM1 ganglioside of smallintestine
(mucosal receptors/[3HJGMl ganglioside/ Vibrio cholerae sialidase/diarrhea)
JAN HOLMGREN*, IVAR LONNROTH*, JAN-ERIC MANSSONt, AND LARS SVENNERHOLMt* Institute of Medical Microbiology and t Department of Neurochemistry, University of G~teborg, Goteborg, Sweden
Communicated by Saul Roseman, March 31, 1975
ABSTRACT Ganglioside GM, was isolated from thesmall intestinal mucosa of man, pig, and beef and amountedto 0.1, 2.0, and 43 nmol per g fresh weight, respectively.These differences in GMI content were associated with aquantitatively differing ability of the mucosal cells to bindcholera toxin. Human cells bound about 15,000 toxin mole-cules when saturated with the toxin, porcine cells 120,000,and bovine cells 2,600,000 molecules. The association con-stant (KA) of the cholera toxin binding was, for cells of allthree species, about 109 liters/mol. Exogenously added GM1ganglioside was incorporated in intestinal mucosal cells aswell as in intact rabbit small bowel. The increment in GM1was associated with a correspondingly increased number ofbinding sites for cholera toxin, whereas KA was unchanged.GM! incorporation increased the sensitivity of the rabbitsmall bowel to the diarrheogenic action of cholera toxin.
Vibrio cholerae sialidase hydrolyzed isolated intestinal di-and trisialogangliosides to GM,. However, the enzyme didnot change the ganglioside pattern of intestinal mucosa, hadvery little influence on the number of toxin binding sites onintestinal cells, and. did not alter the sensitivity of the smallbowel to the diarrheogenic action of the toxin.These results demonstrate a relationship in the intestinal
mucosa between the GM, ganglioside concentration, thenumber of binding sites for cholera toxin, and the sensitivityto the biologic action of the toxin. Thus, the study stronglysupports the concept that the GM, ganglioside is the intesti-nal binding receptor for cholera toxin.
Diarrhea in cholera is due to the action on the small intestineof a protein exotoxin produced by the infecting Vibrio chol-erae (2). The isolated cholera toxin, molecular weight 84,000(3), consists of two types of noncovalently linked subunits,one heavy (H) and several light (L) ones (4-6). The complexof L subunits gives a rapid, tight binding of toxin to theouter membrane of mammalian cells, which is followed by aslower biologic action probably mediated by the H subunit(4-7). The key action of toxin on cellular metabolism seemsto be activation of adenylate cyclase, and in the intestine theresulting cyclic AMP (cAMP) accumulation causes body-to-lumen secretion of chloride, which leads to diarrhea (8).The monosialosylganglioside GM1 was recently found to
bind and inactivate cholera toxin with an affinity and speci-ficity which suggested that this ganglioside, believed to be anormal constituent of the plasma membrane of most mam-malian cells, might be the membrane binding receptor fortoxin (9-11). The observation that aninactive toxin deriva-tive, choleragenoid toxoid, which binds as efficiently as toxinto target cells (7, 12), also had toxin-like binding propertiesto the GMi ganglioside in vitro (10) supports this view. Fur-Abbreviations: The nomenclature of Svennerholm (ref. 1) is used todesignate the gangliosides: GM3, monosialylgalactosylglucosylcera-mide; GD3, disialylgalactosylglucosylceramide; GM1, galactosyl-N-acetylgalactosaminyl-(sialyl)-galactosylglucosylceramide; GD1, disi-alylgalactosyl-N-acetylgalactosaminylgalactosylglucosylceramide;GTI, trisialylgalactosyl-N-acetylgalactosaminylgalactosylglucosyl-ceramide; GA1, ganglio-N-tetraose, galactosyl-N-acetylgalactosam-inylgalactosylglucosylceramide.
ther evidence for the receptor function of GM, is indicatedby the finding that incubation of fat cells with GM, in-creased the toxin binding ability of the cells and also the li-polytic response to the toxin, which suggested that the exog-enous ganglioside was incorporated as functional receptor inthe cell membrane (13).
In the present study, we investigate the binding propertiesof cholera toxin to intestinal mucosa cells of various species,including man, and examine how these properties relate tothe ganglioside pattern of the mucosa, in particular to thecontent of GM1 ganglioside. We further directly demonstrateincorporation of GM, ganglioside in intestinal cells and ana-lyze the effects of such incorporation on both binding oftoxin to the cells and on the diarrheogenic action of choleratoxin on the intact small bowel. It is well known that GM1,which by itself is resistant to sialidase, can be generated invitro from more complex gangliosides by the action of V.cholerae sialidase (1). The possibility that such hydrolysis ofmembrane-located di- and trisialogangliosides could occuras a contributory pathogenic event in cholera infection hasbeen recognized (9, 10). Consequently, we tried to evaluatethe ganglioside-converting effect of V. cholerae sialidase onintestinal ganglioside preparations, cells, and intact smallbowel mucosa as well as the possible influence of enzyme ac-tion for binding and diarrheogenicity of cholera toxin.
MATERIALS AND METHODSCholera Toxin and Toxoid. Culture filtrate of V. cholerae
(lot 4493 G) and highly purified toxin (lot 0572) were re-ceived from Dr. Carl Miller, Cholera Research Program,NIAID, Bethesda, Md. Purified choleragenoid toxoid was agift from Dr. R. A. Finkelstein, Dallas, Texas. The purifiedtoxin and toxoid had been isolated as described (14). Neitherthe purified nor crude toxin had demonstrable sialidase ac-tivity, as tested with GD1a ganglioside as substrate. 125I-La-beled toxin was prepared with Na 125I and the ChloramineT coupling method as described (7), and the fraction of pro-tein similar in size to native toxin was isolated by column fil-tration through Sephadex G-100. More than 90% of the ra-dioactivity of this fraction showed specific binding to cells(mouse thymocytes) or to the ganglioside GM, (coupled topolystyrene tubes or to agarose beads), and the bindingproperties of the labeled toxin were indistinguishable fromthose of unlabeled toxin. The biologic activity was also unaf-fected by the labeling (permeability increase of rabbit skinand thymocyte cAMP generation).
Glycosphingolipids. Pure GM1 ganglioside, the sialicacid-free derivative GA1, and GD1a ganglioside were pre-pared from human brain and characterized as described(15). [3H]GMl ganglioside was prepared by sequential treat-ment with galactose oxidase and sodium [3H]borohydride(16). The radioactive ganglioside was purified by column
2520

Proc. Nat. Acad. Sci. USA 72 (1975) 2521
chromatography on Sephadex G-25 and on silicic acid. Theganglioside was finally purified by preparative thin-layerchromatography on Silica Gel G in a solvent system of chlo-roform/methanol/water 60:32:7 (by volume). The specificactivity was 2.0 X 106 dpm/nmol.
Sephadex G-25 and DEAE-Sephadex A-25 were pur-chased from Pharmacia Ltd., Uppsala, Sweden; Silica Gel Gand H from Fluka AG, Buchs, Switzerland. V. cholerae sial-idase (500 U/ml) was purchased from Behringswerke AG,Marburg-Lahn, West Germany.
Intestinal Preparations for Binding and Activity Stud-ies. Human cells were prepared from surgically removedsegments of jejunum, 15-20 inches (38-50 cm) from the py-lorus, of three patients. Porcine and bovine cells were pre-pared from 1.5-m segments taken immediately after slaugh-ter from the middle of the small intestines of adult animals.The gut segments were placed on ice, cut open longitudinal-ly, and with Ringer's solution gently washed free from visi-ble mucus. After incubation with shaking at 370 for 30 minin Ringer's solution, the cell-rich fluid was poured through anylon mesh. The passed cells were sedimented at 200 X gfor 10 min and washed once with Ringer's solution. About80-90% of the human cells were viable and more than 90%of the animal cells, as judged by trypan blue exclusion andphase contrast microscopy. They were suspended in Eagle'sminimal essential medium supplemented with 1 mg/ml ofCaCl2 to give cell densities of 5 X 106 per ml for the humancells and 2 X 106 per ml for the porcine and bovine cells. Al-iquots of the cell suspensions were incubated at 370 for 25min with GM1 or [3H]GM1 ganglioside in concentrationsranging from 0.07-2 ,gM (0.1-3.0 ,g/ml), V. cholerae sial-idase (1-50 U/ml), or with no additive. The cells were thenwashed four times in minimal medium by centrifugation at200 X g for 10 min and resuspended to the same densities inminimal medium supplemented with 0.2 mg/ml of freshlydissolved bovine serum albumin.
Small bowel segments were prepared in live, anesthetized8- to 12-week-old rabbits, that had been starved for 30-48hr. Starting 50 cm from the pylorus, about 100 cm of the gutwas ligated, and via connecting tubes to each end of this seg-ment it was rinsed with borate/gelatin buffer, pH 7.5 (7),supplied under mild hydrostatic pressure until the outflowfluid was clear. The rinsed gut was then tied off into 25- or50-cm segments, which were incubated for 20 min with 0.5ml/cm of gut of GM1 ganglioside (0.07 or 0.7 ,M in borate/gelatin buffer), V. cholerae sialidase (1, 10, or 20 U/ml inborate/gelatin buffer with 1 mg/ml of CaC12), or, as control,with the buffer alone or, in a few instances, with gangliote-traose GA1 (0.7 ,M in borate/gelatin buffer). Thereafter thesegments were rinsed as above with 200-300 ml of borate/gelatin buffer, and further ligated into approximately 4-cmlong test loops.Binding of Cholera Toxin. Binding of toxin to the dis-
persed cells was tested by means of 125I-labeled toxin mixedwith multiple concentrations of unlabeled toxin. The testprocedure and the Scatchard plot calculations of the associa-tion constant, KA, and number, n, of bound toxin moleculeswhen the cells were saturated with toxin have been de-scribed for thymus cells (7). Modifications were the use of agreater number of toxin concentrations, and testing aliquotsof 4 X 105 porcine and bovine cells as well as 1 X 106 and 5X 106 human cells.
Binding of toxin to rabbit small bowel mucosa was testedby removing the described entire loop-divided intestinalsegment, placing it on ice, and incubating the loops for 20min with 1 ml each of 125I-labeled toxin mixed in minimal
medium/bovine serum albumin with different concentra-tions of unlabeled toxin. Each loop was then cut open anddip-washed several times in minimal medium; its radioactiv-ity was determined.
Biological Action of Cholera Toxin and Inhibition withToxoid. The ability of crude toxin (V. cholerae culture fil-trate) to cause fluid secretion in the small bowel of live rab-bits was quantitated in the prepared intestinal loops (17).Quantitative inhibition of the diarrheogenic action of thecrude toxin by choleragenoid (18, 19) was studied by inject-ing different amounts of choleragenoid in a volume of 0.5ml into the intestinal loops 15 min before injection of anequal volume containing 0.1 or 1.0 mg of V. cholerae cul-ture filtrate. The assay was the same as that used for the de-termination of toxin action.
Isolation of Gangliosides from Human, Pig, and BovineIntestine. The same bowel material was used as for the iso-lation of intestinal cells. The jejunum and ileum were evert-ed over a cold rod, and the mucosa was carefully rinsedfrom mucus with physiological saline and then scraped offwith a surgical knife. Great care was taken not to obtain anyof the muscular layer, particularly at the preparation of thehuman jejunum. Porcine mucosa (555 g), porcine muscularlayer (2.0 kg), bovine mucosa (185 g), bovine muscular layer(650 g), human mucosa (8-25 g; 4 samples), and humanmuscular layer (54 g) were available for ganglioside isola-tion. To two of the samples of human mucosa 0.1 nmol of[3H]GM1 was added. The mucosa scrapings and the samplesof muscular layer, after homogenization in a Waring blen-dor, were extracted with 6 volumes of chloroform/methanol1:2 (by volume). The gangliosides, except for gangliosideGM3, were isolated by partition and purified by columnchromatography on Sephadex G-25 and Silica Gel G (Sven-nerholm, in preparation). All the ganglioside preparations,particularly those of human and pig mucosa, were still veryimpure, and the gangliosides of all the six sources were fur-ther purified by mild alkaline hydrolysis, chromatographyon Sephadex G-25, DEAE-Sephadex A-25 (20), SephadexG-25, and finally on Silica Gel H (21). The quantitative de-terminations of gangliosides were performed on the final el-uate of the Silica Gel H column by the resorcinol method(22). The ganglioside pattern was determined as for brain(23). In the human mucosa, the ganglioside concentrationwas extremely low and total gangliosides were only sepa-rated into three fractions: ganglioside GM1, gangliosides GD1+ GT1, and ganglioside GD3. Ganglioside GM, and ganglios-ides GD1 and GT1 were hydrolyzed with V. cholerae sial-idase, and the hydrolyzed gangliosides were semiquantita-tively estimated on thin-layer plates sprayed with resorcinol(22).The neutral carbohydrate moiety of the gangliosides of
bovine and porcine intestine was characterized in the fol-lowing manner. The apparent ganglioside GM1 and the gan-gliosides GD1 and GT1 were isolated by thin-layer chroma-tography. These fractions were hydrolyzed with V. choleraesialidase at pH 6.8, and the gangliosides that migrated asganglioside GM1 were subjected to acid hydrolysis with 1 Mformic acid at 1000 for 1 hr (24). The neutral tetraglycosyl-ceramide formed was analyzed with thin-layer chromatog-raphy in chloroform/methanol/water 65:25:4 and 60:32:7(by volume) with neutral glycosylceramides of known chem-ical structure as reference substances.
Sialidase Hydrolysis of Pig Mucosa. Porcine mucosa(100 g) was hydrolyzed for 30 min at 370 with 200 units ofV. cholerae sialidase in a total volume of 200 ml of 0.05 Macetate buffer pH S.0 or 0.05 M Tris-maleate buffer pH 6.8.
Biochemistry: Ho1mgren et al.
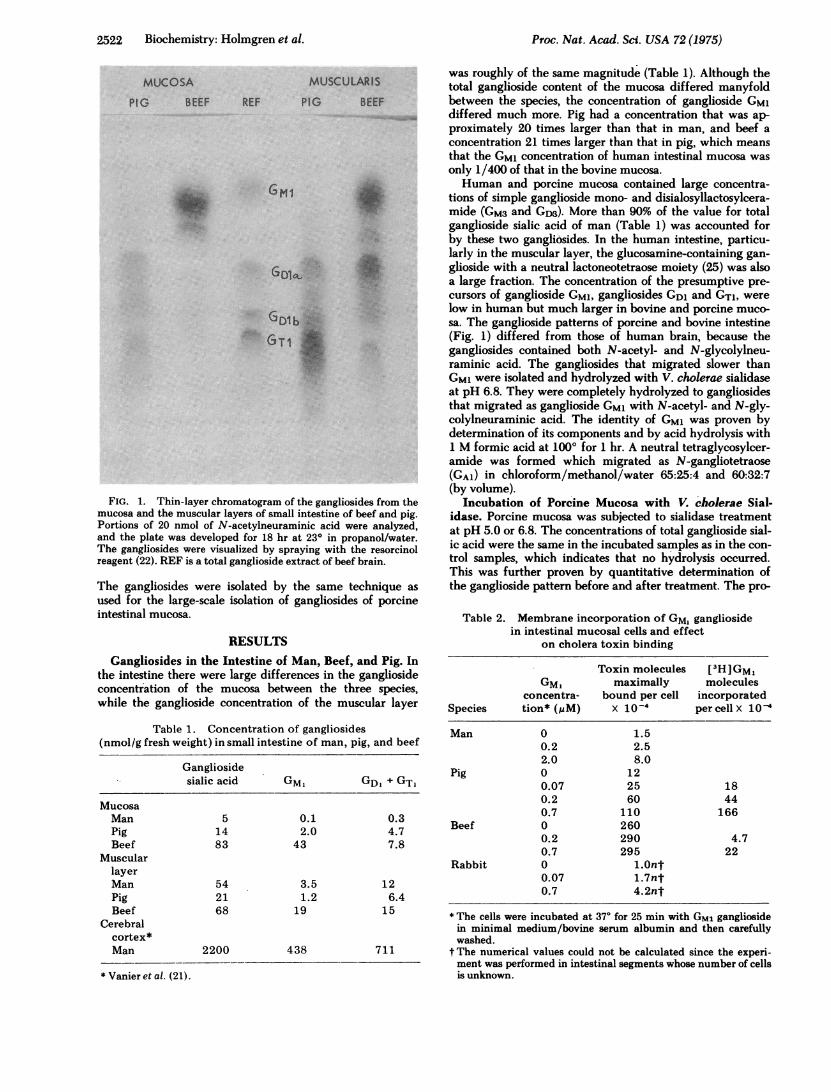
2522 Biochemistry: Holmgren et al.
MUCOSA MUSCULARIS
PIG BEEF REF PIG BEEF
. ~~GM1
G
FIG. 1. Thin-layer chromatogram of the gangliosides from the
mucosa and the muscular layers of small intestine of beef and pig.Portions of 20 nmol of N-acetylneuraminic acid were analyzed,and the plate was developed for 18 hr at 230 in propanol/water.The gangliosides were visualized by spraying with the resorcinol
reagent (22). REF is a total ganglioside extract of beef brain.
The gangliosides were isolated by the same technique as
used for the large-scale isolation of gangliosides of porcineintestinal mucosa.
RESULTS
Gangliosides in the Intestine of Man, Beef, and Pig. In
the intestine there were large differences in the gangliosideconcentration of the mucosa between the three species,
while the ganglioside concentration of the muscular layer
Table 1. Concentration of gangliosides
(nmol/g fresh weight) in small intestine of man, pig, and beef
Gangliosidesialic acid GM, GD, + GT,
Mucosa
Man 5 0.1 0.3
Pig 14 2.0 4.7
Beef 83 43 7.8
Muscularlayer
Man 54 3.5 12
Pig 21 1.2 6.4
Beef 68 19 15
Cerebral
cortex*
Man 2200 438 711
* Vanier et al. (21).
was roughly of the same magnitude (Table 1). Although thetotal ganglioside content of the mucosa differed manyfoldbetween the species, the concentration of ganglioside GM1differed much more. Pig had a concentration that was ap-proximately 20 times larger than that in man, and beef aconcentration 21 times larger than that in pig, which meansthat the GM1 concentration of human intestinal mucosa wasonly 1/400 of that in the bovine mucosa.Human and porcine mucosa contained large concentra-
tions of simple ganglioside mono- and disialosyllactosylcera-mide (GM3 and GDM). More than 90% of the value for totalganglioside sialic acid of man (Table 1) was accounted forby these two gangli6sides. In the human intestine, particu-larly in the muscular layer, the glucosamine-containing gan-glioside with a neutral lactoneotetraose moiety (25) was alsoa large fraction. The concentration of the presumptive pre-cursors of ganglioside GM1, gangliosides GD1 and GT1, werelow in human but much larger in bovine and porcine muco-sa. The ganglioside patterns of porcine and bovine intestine(Fig. 1) differed from those of human brain, because thegangliosides contained both N-acetyl- and N-glycolylneu-raminic acid. The gangliosides that migrated slower thanGM1 were isolated and hydrolyzed with V. cholerae sialidaseat pH 6.8. They were completely hydrolyzed to gangliosidesthat migrated as ganglioside GM1 with N-acetyl- and N-gly-colylneuraminic acid. The identity of GM1 was proven bydetermination of its components and by acid hydrolysis with1 M formic acid at 1000 for 1 hr. A neutral tetraglycosylcer-amide was formed which migrated as N-gangliotetraose(GA1) in chloroform/methanol/water 65:25:4 and 60:32:7(by volume).
Incubation of Porcine Mucosa with V. cholerae Sial-idase. Porcine mucosa was subjected to sialidase treatmentat pH 5.0 or 6.8. The concentrations of total ganglioside sial-ic acid were the same in the incubated samples as in the con-trol samples, which indicates that no hydrolysis occurred.This was further proven by quantitative determination ofthe ganglioside pattern before and after treatment. The pro-
Table 2. Membrane incorporation of GM, gangliosidein intestinal mucosal cells and effect
on cholera toxin binding
Toxin molecules [3H]GM1GM, maximally molecules
concentra- bound per cell incorporatedSpecies tion* (,M) X 10-4 per cell X 10
Man 0 1.50.2 2.52.0 8.0
Pig 0 120.07 25 180.2 60 440.7 110 166
Beef 0 2600.2 290 4.70.7 295 22
Rabbit 0 L.Ont0.07 1.7nt0.7 4.2nt
* The cells were incubated at 370 for 25 min with GM, gangliosidein minimal medium/bovine serum albumin and then carefullywashed.
t The numerical values could not be calculated since the experi-ment was performed in intestinal segments whose number of cellsis unknown.
Proc. Nat. Acad. Sci. USA 72 (1975)
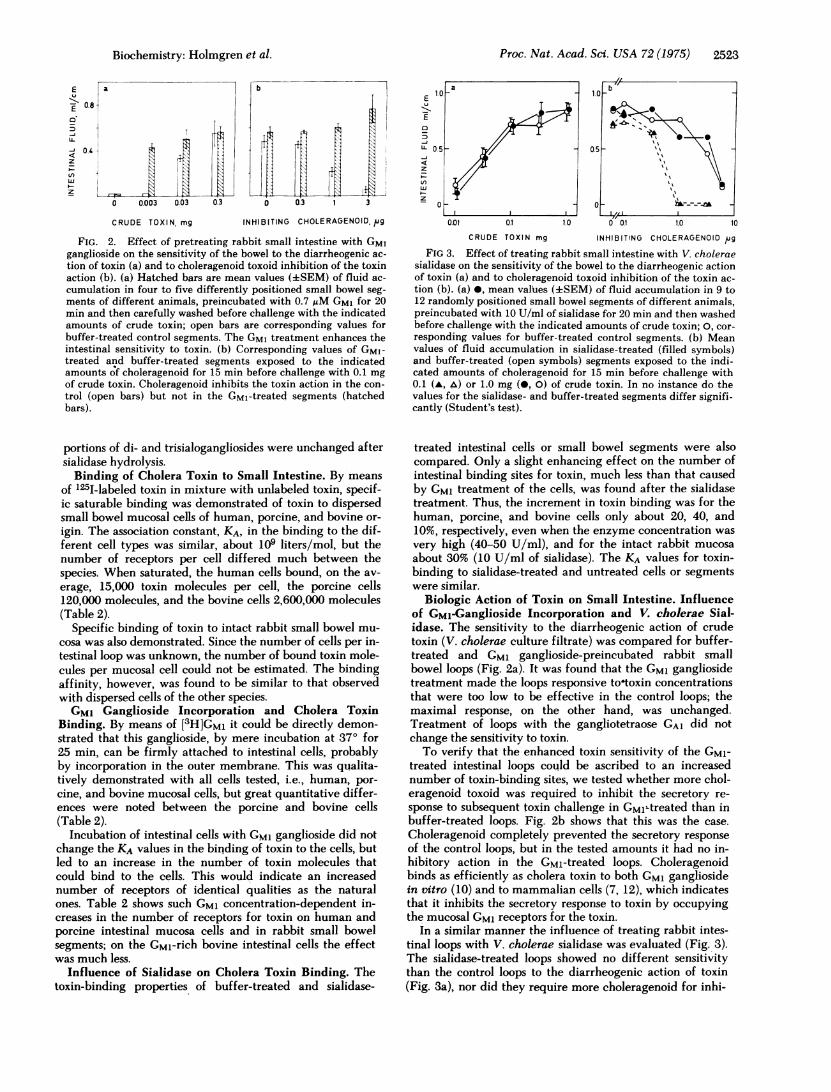
Proc. Nat. Acad. Sci. USA 72 (1975) 2523
Ia
0 0.003 0.03 0.3
CRUDE TOXI N. mg
b
~1-XA I0 0.3 1 3
INHIBITING CHOLERAGENOID. ug
FIG. 2. Effect of pretreating rabbit small intestine with GM1ganglioside on the sensitivity of the bowel to the diarrheogenic ac-
tion of toxin (a) and to choleragenoid toxoid inhibition of the toxinaction (b). (a) Hatched bars are mean values (+SEM) of fluid ac-
cumulation in four to five differently positioned small bowel seg-ments of different animals, preincubated with 0.7 gM GM1 for 20min and then carefully washed before challenge with the indicatedamounts of crude toxin; open bars are corresponding values forbuffer-treated control segments. The GM1 treatment enhances theintestinal sensitivity to toxin. (b) Corresponding values of GM1-treated and buffer-treated segments exposed to the indicatedamounts of choleragenoid for 15 min before challenge with 0.1 mgof crude toxin. Choleragenoid inhibits the toxin action in the con-
trol (open bars) but not in the GMl-treated segments (hatchedbars).
portions of di- and trisialogangliosides were unchanged aftersialidase hydrolysis.Binding of Cholera Toxin to Small Intestine. By means
of '25I-labeled toxin in mixture with unlabeled toxin, specif-ic saturable binding was demonstrated of toxin to dispersedsmall bowel mucosal cells of human, porcine, and bovine or-
igin. The association constant, KA, in the binding to the dif-ferent cell types was similar, about 109 liters/mol, but thenumber of receptors per cell differed much between thespecies. When saturated, the human cells bound, on the av-
erage, 15,000 toxin molecules per cell, the porcine cells120,000 molecules, and the bovine cells 2,600,000 molecules(Table 2).
Specific binding of toxin to intact rabbit small bowel mu-
cosa was also demonstrated. Since the number of cells per in-testinal loop was unknown, the number of bound toxin mole-cules per mucosal cell could not be estimated. The bindingaffinity, however, was found to be similar to that observedwith dispersed cells of the other species.GM1 Ganglioside Incorporation and Cholera Toxin
Binding. By means of [3H]GM1 it could be directly demon-strated that this ganglioside, by mere incubation at 370 for25 min, can be firmly attached to intestinal cells, probablyby incorporation in the outer membrane. This was qualita-tively demonstrated with all cells tested, i.e., human, por-
cine, and bovine mucosal cells, but great quantitative differ-ences were noted between the porcine and bovine cells(Table 2).
Incubation of intestinal cells with GM1 ganglioside did notchange the KA values in the binding of toxin to the cells, butled to an increase in the number of toxin molecules thatcould bind to the cells. This would indicate an increasednumber of receptors of identical qualities as the naturalones. Table 2 shows such GM1 concentration-dependent in-creases in the number of receptors for toxin on human andporcine intestinal mucosa cells and in rabbit small bowelsegments; on the GMl-rich bovine intestinal cells the effectwas much less.
Influence of Sialidase on Cholera Toxin Binding. Thetoxin-binding properties of buffer-treated and sialidase-
CRUDE TOXIN mg INHIBITING CHOLERAGENOID ug
FIG 3. Effect of treating rabbit small intestine with V. choleraesialidase on the sensitivity of the bowel to the diarrheogenic actionof toxin (a) and to choleragenoid toxoid inhibition of the toxin ac-
tion (b). (a) mean values (+SEM) of fluid accumulation in 9 to12 randomly positioned small bowel segments of different animals,preincubated with 10 U/ml of sialidase for 20 min and then washedbefore challenge with the indicated amounts of crude toxin; 0, cor-
responding values for buffer-treated control segments. (b) Meanvalues of fluid accumulation in sialidase-treated (filled symbols)and buffer-treated (open symbols) segments exposed to the indi-cated amounts of choleragenoid for 15 min before challenge with0.1 (A, A) or 1.0 mg (-, 0) of crude toxin. In no instance do thevalues for the sialidase- and buffer-treated segments differ signifi-cantly (Student's test).
treated intestinal cells or small bowel segments were alsocompared. Only a slight enhancing effect on the number ofintestinal binding sites for toxin, much less than that causedby GM1 treatment of the cells, was found after the sialidasetreatment. Thus, the increment in toxin binding was for thehuman, porcine, and bovine cells only about 20, 40, and10%, respectively, even when the enzyme concentration was
very high (40-50 U/ml), and for the intact rabbit mucosa
about 30% (10 U/ml of sialidase). The KA values for toxin-binding to sialidase-treated and untreated cells or segmentswere similar.
Biologic Action of Toxin on Small Intestine. Influenceof GMl-Ganglioside Incorporation and V. cholerae Sial-idase. The sensitivity to the diarrheogenic action of crudetoxin (V. cholerae culture filtrate) was compared for buffer-treated and GM1 ganglioside-preincubated rabbit smallbowel loops (Fig. 2a). It was found that the GM1 gangliosidetreatment made the loops responsive tootoxin concentrationsthat were too low to be effective in the control loops; themaximal response, on the other hand, was unchanged.Treatment of loops with the gangliotetraose GA1 did notchange the sensitivity to toxin.To verify that the enhanced toxin sensitivity of the GMI-
treated intestinal loops could be ascribed to an increasednumber of toxin-binding sites, we tested whether more chol-eragenoid toxoid was required to inhibit the secretory re-
sponse to subsequent toxin challenge in GMlLtreated than inbuffer-treated loops. Fig. 2b shows that this was the case.
Choleragenoid completely prevented the secretory response
of the control loops, but in the tested amounts it had no in-hibitory action in the GM1-treated loops. Choleragenoidbinds as efficiently as cholera toxin to both GM1 gangliosidein vitro (10) and to mammalian cells (7, 12), which indicatesthat it inhibits the secretory response to toxin by occupyingthe mucosal GM1 receptors for the toxin.
In a similar manner the influence of treating rabbit intes-tinal loops with V. cholerae sialidase was evaluated (Fig. 3).The sialidase-treated loops showed no different sensitivitythan the control loops to the diarrheogenic action of toxin(Fig. 3a), nor did they require more choleragenoid for inhi-
E
E 0.8
0
IL
z
z
Biochemistry: Holmgren et al.

2524 Biochemistry: Holmgren et al.
bition of fluid production to toxin challenge (Fig. Sb). Thequantitative inhibition by choleragenoid of two differentdoses of challenge toxin gives further support for the con-cept of receptor competition with similar affinity betweenthe toxoid and toxin.To study a possible delayed influence of sialidase on the
intestinal sensitivity to toxin, the incubation time with theenzyme in the intestine was extended to 200 min in threeanimals. This, however, did not change the sensitivity fortoxin of the enzyme-treated segments from that of the con-trol segments incubated with borate/gelatin buffer.
Finally, it was found that incubation of the intestine inmvo with 10 U/ml of sialidase for 20 min did not result inliberation of more free sialic than incubation with buffer;the enzyme was fully active after the incubation, however,as tested with GDla ganglioside as substrate.
DISCUSSIONThe concept, derived from the observations described in theintroduction, that the GM1 ganglioside is the binding recep-tor for cholera toxin in the small bowel has gained muchsupport from the present study. The occurrence of GM1 isdemonstrated in the small bowel mucosa of various mamma-lian species, including man, and the marked differences inthe content of this ganglioside among the species are foundto correspond to different numbers of toxin molecules thatcan bind per intestinal cell. Further, it is shown by means of[3H]GMS that the exogenously added GM1 can be incorporat-ed in intestinal mucosal cells or intact small bowel, and thatsuch incorporation increases the number of toxin moleculesthat can bind per cell as well as the sensitivity of the intes-tine to the diarrheogenic action of toxin.
Large differences were found in the concentration of GM1ganglioside in the intestinal mucosa of man, pig, and beef. Itis striking that the 21-fold higher GM1 concentration in beefas compared with pig mucosa was associated with a 22-foldhigher average number of binding sites to toxin in the bo-vine than in the porcine mucosal cell. It is also notable thatthe naturally GM1-rich bovine cells incorporated manyfewer [3H]GM1 molecules than the porcine cells, and paral-lelly did not increase the number of toxin binding sites percell as much as the porcine cells. In consideration of thecomplicated experimental procedures, the rather closeagreement in the porcine cells between the number of[3H]GM1 molecules incorporated per cell and the increase innumber of toxin molecules that were bound by a toxin-satu-rated cell, the mean ratio being 1.3, suggests that almostevery GM1 molecule incorporated is in a position to attachone toxin molecule. Moreover, the association constant inbinding is similar, around 109 liters/mol, in the GMl-treatedcells as in the untreated ones, further indicating that the in-corporated GM1 molecules have identical toxin-bindingproperties as the natural receptors.
V. cholerae sialidase can convert isolated di- and trisialo-gangliosides to the sialidase-resistant GM1 ganglioside, asshown previously (1) and in the present study. However,when permitted to act in vitro on intestinal cells or mucosaor in ivo in the small bowel, the enzyme did not change theganglioside pattern of the mucosa, had little influence on thenumber of toxin-binding sites per intestinal cell, and alteredneither the sensitivity of the small bowel to the diarrheogen-ic action of toxin nor the amount of choleragenoid toxoid re-
quired to inhibit the toxin action on the gut. These findingscontradict a major pathogenetic role of V. cholerae sialidaseproduction as a toxin-receptor-creating enzyme in the intes-tine. In contrast to our findings, King and van Heyningen(9) found with sialidase treatment an 8-fold increase of thetoxin-inactivating capacity of rabbit and bovine intestinalmucosa scrapings; they also claimed that the ganglioside pat-terns of bovine brain and intestinal mucosal scrapings didnot show any remarkable differences. Our Fig. 1 illustratesthat the ganglioside pattern of the intestinal muscular layeris similar to that of brain, which is not unexpected since thegangliosides of the muscular layer are partly derived fromthe nerve cells in this tissue. Contamination of the mucosalscrapings tested by King and van Heyningen with muscularlayer tissue might possibly explain the divergent findings ascompared with those of the present study.
We thank Dr. Tore Schersten for the specimens of human smallintestine and Dr. Harald Almgren, Scan, Uddevalla, for the facili-ties he put at our disposal for the work with beef and pig intestine.This work was supported by grants from the Swedish Medical Re-search Council (no. 6X-3383 and 3X-627), the W.H.O., and theHesselman Foundation.
1. Svennerholm, L. (1963) J. Neurochem. 10, 613-623.2. Pierce, N. F., Greenough, W. B., III & Carpenter, C. C. (1971)
Bacteriol. Rev. 35, 1-13.3. LoSpalluto, J. J. & Finkelstein, R. A. (1972) Biochim. Biophys.
Acta 257, 158-166.4. Lonnroth, I. & Holmgren, J. (1973) J. Gen. Microbiol. 76,
417-427.5. Cuatrecasas, P., Parikh, I. & Hollenberg, D. (1973) Biochemis-
try 12, 4253-4264.6. Holmgren, J. & Lonnroth, I. (1975) J. Gen. Microbiol 86, 49-
66.7. Holmgren, J., Lindholm, L. & Lonnroth, I. (1974) J. Exp.
Med. 139, 801-819.8. Field, M. (1971) N. Engl. J. Med. 284, 1137-1144.9. King, C. A. & van Heyningen, W. E. (1973) J. Infect. Dis.
127,639-647.10. Holmgren, J., Lonnroth, I. & Svennerholm, L. (1973) Infect.
Immun. 8,208-214.11. Cuatrecasas, P. (1973) Biochemistry 12,3547-3558.12. Cuatrecasas, P. (1973) Biochemistry 12,3577-3581.13. Cuatrecasas, P. (1973) Biochemistry 12, 3558-3566.14. Finkelstein, R. A. & LoSpalluto, J. J. (1970) J. Infect. Dis. 121,
S 63-72.15. Svennerholm, L. (1972) in Methods in Carbohydrate Chemis-
try, eds. Whistler, R. L. & BeMiller, J. N. (Academic PressInc., New York), Vol. 6, pp. 464-474.
16. Radin, N. S., Hof, L., Bradley, R. M. & Brady, R. 0. (1969)Brain Res. 14,497-505.
17. Kasai, G. J. & Burrows, W. (1966) J. Infect. Dis. 116, 606-614.
18. Pierce, N. F. (1973) J. Exp. Med. 137, 1009-1023.19. Holmgren, J. (1973) Infect. Immun. 8, 851-859.20. Yu, R. K. & Ledeen, R. W. (1972) J. Lipid Res. 13,680-686.21. Vanier, M.-T., Holm, M., MAnsson, J.-E. & Svennerholm, L.
(1973) J. Neurochem. 21, 1375-1384.22. Svennerholm, L. (1957) Biochim. Biophys. Acta 24,604-611.23. Vanier, M.-T., Holm, M., Ohman, R. & Svennerholm, L.
(1970) J. Neurochem. 18,581-592.24. Holm, M., Mansson, J.-E., Vanier, M.-T. & Svennerholm, L.
(1972) Biochim. Biophys. Acta 280,356-364.25. Li, Y.-T., MAnsson, J.-E., Vanier, M.-T. & Svennerholm, L.
(1973) J. Biol. Chem. 248, 2634-2636.
Proc. Nat. Acad. Sci. USA 72 (1975)